Embed Size (px)
Citation preview

R
To
NRa
b
c
3
a
ARRAA
KACCLOS
1
b2tta12
h1
Perspectives in Plant Ecology, Evolution and Systematics 22 (2016) 23–35
Contents lists available at ScienceDirect
Perspectives in Plant Ecology, Evolution and Systematics
journa l homepage: www.e lsev ier .com/ locate /ppees
esearch article
richome-like emergences in Croton of Brazilian highland rockutcrops: Evidences for atmospheric water uptake
arah Costa Vitarelli a,c, Ricarda Riina b, Mariana Franco Cassino a,enata Maria Strozi Alves Meira a,∗
Departamento de Biologia Vegetal, Pós-Graduac ão em Botânica, Universidade Federal de Vic osa, Vic osa 36570-900, BrazilReal Jardín Botánico, RJB-CSIC, Plaza de Murillo 2, ES-28014 Madrid, SpainDepartamento de Educac ão e Ciência, Instituto Federal de Educac ão, Ciência e Tecnologia do Sudeste de Minas Gerais (IFSUDESTEMG), Juiz de Fora,6080-001, Brazil
r t i c l e i n f o
rticle history:eceived 28 September 2015eceived in revised form 1 July 2016ccepted 29 July 2016vailable online 3 August 2016
ewwords:tmospheric water absorptionampos de altitudeampos rupestreseaf anatomyntogenyclereids
a b s t r a c t
Background and aims: Although trichomes are typical features of the species-rich genus Croton, little isknown about their development, anatomical structure and function. This study aims to characterize theanatomy of leaf trichomes of three Croton species restricted to Brazilian rock outcrops, and investigatetheir functional role in an environment with limited soil water availability.Methods: Samples of leaves at different stages of maturity were submitted to standard anatomicallight and scanning electron microscopy techniques to perform a structural and developmental study.Atmospheric water uptake was investigated through the absorption of aqueous solutions of berberinehemisulfate and Indian ink.Key results: The so-called trichomes actually correspond to emergences because they originate fromprotodermal and ground meristem activity rather than from epidermal tissue. These emergences showa complex anatomy, are closely associated with the mesophyll sclereids and might be involved in leafatmospheric water uptake. The outermost cells of the emergences are arranged radially and presentnon-lignified thick walls. We demonstrate that an aqueous solution can effectively penetrate throughthe leaf emergences, continuing through sclereids and reaching the vascular bundle or flowing across the
mesophyll maximizing water distribution in the leaf tissue.Conclusions: The complex structure and hypothesized function of Croton trichome-like emergences rep-resent a novel finding in Euphorbiaceae. In the three species studied, these leaf emergences might playa role in atmospheric water absorption and they may be one of the clues for the occurrence of Croton inhabitats with limited soil water supply where this genus is usually species-rich and abundant.© 2016 Elsevier GmbH. All rights reserved.
. Introduction
Trichomes of diverse types characterize Croton L. (Euphor-iaceae), one of the largest genera of flowering plants (Berry et al.,005) with over 1200 species (Govaerts et al., 2000). Because ofheir great diversity in morphology, density, and distribution pat-erns on the plant, trichomes have been used by Croton systematists
s a rich source of characters at different taxonomic levels (Webster,993; Lima and Pirani, 2003, 2008; Riina et al., 2010; Caruzo et al.,011; van Ee and Berry, 2011; Vitarelli et al., 2015).∗ Corresponding author.E-mail addresses: [email protected], renata meira @hotmail.com (R.M.S.A. Meira).
ttp://dx.doi.org/10.1016/j.ppees.2016.07.002433-8319/© 2016 Elsevier GmbH. All rights reserved.
Plant trichomes are specialized epidermal structures originat-ing from protodermal cells. The existence of a connection betweentrichomes and structures in the mesophyll has been reported in sev-eral species of Croton (Solereder, 1908; Metcalfe and Chalk, 1950;Sá-Haiad, 1987; Starling, 1993; Louro et al., 2003; Appezzato-da-Glória and Carmello-Guerreiro, 2006; Lucena and Sales, 2006; Silva,2006; Sá-Haiad et al., 2009; Barros and Soares, 2013), but withoutany experimental demonstration about the trichomes functionalrole.
In Croton, seven types of trichomes (stellate, fasciculate,multiradiate-rosulate, dendritic, lepidote, papillate, and glandu-
lar) have been described (Webster et al., 1996). Trichomes areimportant features in systematics, physiology and ecology, how-ever studies about their anatomy and development are scarce. We
2 logy,
srterslC
mc2eowwl2TomMAMrudevaOiaiu
wedp1ls1fa1tctRsietfl(i
gCaia
4 N.C. Vitarelli et al. / Perspectives in Plant Eco
et to conduct a detailed study of leaf trichomes of three closelyelated species of Croton section Lamprocroton, namely Croton ery-hroxyloides Baill., C. pygmaeus L.R. Lima, and C. splendidus Mart.x Colla. In a recent taxonomic study, lepidote trichomes wereeported for the abaxial side of the leaves of C. pygmaeus and C.plendidus and on both leaf surfaces in C. erythroxyloides, and stel-ate trichomes on the adaxial side of the leaves of C. pygmaeus and. splendidus (Lima and Pirani, 2008).
Croton erythroxyloides, C. pygmaeus and C. splendidus are com-on in high elevation rocky habitats in southeastern Brazil; the so
alled campos rupestres and campos de altitude (Lima and Pirani,008). Plants living in these ecosystems are exposed to similarxtreme environmental conditions, such as high daily variationf temperature, risk of frost, intensive sun irradiation, stronginds, and the nutrient-impoverished shallow soils with a lowater-holding capacity that leads to long periods of severe water
imitation for plant productivity (e.g., Safford, 1999; Benites et al.,003; Lüttge et al., 1998; Morales et al., 2015; Silveira et al., 2015).hese conditions exert continuous selective pressure on plants col-nizing these environments, which could result in the evolution oforphoanatomical and/or physiological adaptations (Eames andcDaniels, 1947; Metcalfe and Chalk, 1983; Hamba et al., 2002;
lcantara et al., 2015), especially those involved in water economy.any species have superficial root systems that would allow for
apid use of rain pulses (Nobel et al., 1990). Whilst leaves of res-rrection plants are physiologically inactive during the dry season,iffering from deciduous species in that the former retain leaf nutri-nts during the dry season (Griffiths et al., 2014). Aerial roots withelamen or pseudostems are common in Velloziaceae (Porembskind Barthlott, 1995), Bromeliaceae (Pita and Menezes, 2002) andrchidaceae (Silva et al., 2010). Velamen may play a role in access-
ng moisture from dew and rain in campos rupestres and campos deltitude (Moreira et al., 2009; Silva et al., 2010) or could be moremportant in preventing water loss than in moisture or nutrientptake (Dycus and Knudson, 1957).
Among the anatomical traits, trichomes stand out by theiridespread presence in phylogenetically unrelated species (Araújo
t al., 2010; Lusa et al., 2014; Francino et al., 2015). They might haveifferent functions, such as protection against herbivores either byhysical (Levin, 1973; Wagner, 1991) or chemical defense (Levin,973), reduction of the fraction of radiation that is absorbed by
eaves (Fahn, 1986; Karabourniotis and Esseas, 1996), protection oftomata and reduction of water loss by evapotranspiration (Fahn,986). Trichomes may play a role in atmospheric water uptake, per-orming as specialized epidermal structures that could speed thetmospheric water absorption process (Stuart, 1968; Benzing et al.,976; Martin and von Willert, 2000; Fernández et al., 2014). Besidesrichomes, other epidermal structures, such as hydathodes anduticule structure, could be related to atmospheric water absorp-ion (Grundell, 1933; Meidner, 1954; Leyton and Armitage, 1968;iederer 2006; Eller et al., 2013). All of these epidermal structureseem to provide an important alternative water supply, especiallyn environments where soil water availability is deficient. Severalxperiments could be conducted to evaluate the water pathwayhrough epidermal structures, for example, using berberine sulfateuorochrome (Heide-Jørgensen, 1990), aqueous safranin solutionMilanez and Machado, 2008), and measuring the hydric potentialn wettable leaves (Fernández et al., 2014).
This study aims to perform anatomical, histochemical and onto-enetic characterization of leaf trichomes of three closely relatedroton species occurring in similar habitats, i.e., campos rupestresnd campos de altitude, as well as to evaluate their functional role
n plants growing in such extreme and harsh environments char-cterized by limited soil water availability.Evolution and Systematics 22 (2016) 23–35
2. Materials and methods
2.1. Tissue sampling and study site
Croton erythroxyloides, C. pygmaeus and C. splendidus were cho-sen because they present different types of trichomes (lepidote,stellate, and fasciculate) and occur in extreme habitats, allowing usto compare trichome structure, development, and function undersimilar environmental conditions. Leaf samples were obtained fromnatural populations or from herbarium material (Table 1). Voucherspecimens were deposited in the herbarium of the UniversidadeFederal de Vic osa (VIC).
Croton erythroxyloides and C. splendidus were collected from nat-ural populations (Fig. 1A–E) in campos de altitude above 1700 melevation on ‘Serra das cabec as’, ‘Parque Estadual da Serra doBrigadeiro’ (PESB), Minas Gerais State, Brazil (Fig. 1A–C). Camposde altitude (altitudinal grasslands) are rupestrian ecosystems ofigneous formation that occur mostly on the ‘Serra da Mantiqueira’and ‘Serra do Mar’ ranges in southeastern Brazil. Campos rupestresand campos de altitude share several plant genera and are knownto harbor considerable numbers of endemic species (Safford, 2007;Alves and Kolbek, 2010). In spite of their floristic and physionomicsimilarities, they are geologically distinct (Benites et al., 2007; Alvesand Kolbek, 2010). For example, campos de altitude have more floris-tic affinities with the páramo vegetation of the Andes than camposrupestres (Alves and Kolbek, 2010). On the other hand, both ecosys-tems have soils developed on ironstones that are shallow and acidicwith a low water-holding capacity.
The PESB is located at the summit of ‘Serra da Mantiqueira’between 42◦20′ and 42◦40′S and 20◦20′ and 21◦00′W (Engevix,1995). The regional climate is classified as mesothermal medium– CWb (Koeppen, 1948), the average annual precipitation is1300 mm. Although, the annual precipitation does not imply a xericenvironment, the high sun light exposition, shallow depth and highpermeability of soils and intense wind, produce an environmentwith extreme low water supply (Benites, 1998). The rainy sea-son is from November to March and the dry season from Mayto September. Leaf samples were collected in both the dry andrainy season. The average annual temperature is 18 ◦C (64.4 ◦F)(Engevix, 1995), with June, July and August as the coldest months.The recorded minimum temperature was below 0 ◦C. The presenceof clouds covering the peaks of the PESB is observed daily, especiallyin early morning hours (Fig. 1B and C). The individuals sampledgrow in dense grass tussocks directly on bare rock outcrops withlow water-holding capacity (Fig. 1C).
Samples of Croton pygmaeus were obtained from herbariummaterial (Table 1). We were unable to obtain fresh material fromthis species because of the difficult and expensive logistics dueto its disjunct location from the other two species. Accordingto the herbarium collection labels the individuals sampled areshrubs 1–2 m tall, growing directly on rocky substrates in the stateof Rio Grande do Sul, southern Brazil (approximate location at30◦32′6.92′′S, 53◦33′2.14W).
Leaves at different developmental stages (leaf primordia, youngand mature leaves) from each specimen were used in this study.Four specimens of Croton splendidus, four of C. erythroxyloides andtwo of C. pygmaeus were used as replicates (Table 1), using a min-imum of three leaves per specimen. Sections of leaf samples weretaken from the apical, median and basal portions of the blade(including midrib, margin, and region between them).
2.2. Light microscopy
Leaf tissue sampled from herbarium specimens was boiled indistilled water for 10 min, treated with 2% potassium hydroxidefor 2 h at room temperature, rinsed in tap water five times, dehy-

N.C. Vitarelli et al. / Perspectives in Plant Ecology, Evolution and Systematics 22 (2016) 23–35 25
Table 1List of species, emergence type, and voucher information.
Species Emergence type Voucher (herbarium)
Croton erythroxyloides Baill. Lepidote (adaxial and abaxial) Vitarelli 002 (VIC), Vitarelli 003 (VIC), Vitarelli005 (VIC), Vitarelli s.n. (VIC)
Croton pygmaeus L.R. Lima Stellate (adaxial) Lepidote (abaxial) Falkenberg 6483 (BHCB), Marchett 549 (BHCB)
Croton splendidus Mart. ex. Colla Fasciculate (adaxial) Lepidote (abaxial) Alvim s.n. (HUEMG), Alvim 40 (HUEMG),Vitarelli s.n. (VIC), Vitarelli s.n. (VIC)
Fig. 1. Campos de altitude of ‘Serra das Cabec as’ (‘Parque Estadual da Serra do Brigadeiro’, Minas Gerais, Brazil). (A) View from the summit with a clear sky at 12:00 noon. (B)V on onb e to tho
d(evgAtiHisLtM
iew from the summit with sky covered by fog, early morning. (C) View of vegetatiare rock. (D) Croton splendidus, detail of leaf surfaces showing a discolor aspect duf lepidote emergences, visible to the naked eye, as brown pits.
rated in an ethanol series (30–70%) and stored in 70% ethanolSmith and Smith, 1942). Fresh leaf material was fixed in the fieldither in FAA (glacial acetic acid, formalin, 50% ethanol, 1:1:18 byolume) (Johansen, 1940) or NBF (neutral buffered formalin) foreneral characterization and histochemical testing (Clark, 1973).fter fixation, the samples were also stored in 70% ethanol. Part of
he samples was dehydrated in an ethanol series and embeddedn methacrylate (Historesin, Leica Microsystems Nussloch GmbH,eidelberg, Germany, prepared according to the manufacturer
nstructions). The embedded samples were cross and longitudinallyectioned (5 �m thick) in an automatic rotary microtome (RM2155,
eica Microsystems Inc., Deerfield, USA) using glass knives, andhe sections were stained with toluidine blue pH 4.0 (O’Brien andcCully, 1981).
the summit. Croton populations grow, along with dense grass tussocks, directly one high density of lepidote emergences. (E) C. erythroxyloides, showing high density
Hand-cut sections were obtained from both fresh and fixedsamples for performing the following histochemical tests: SudanIV (Pearse, 1980) and neutral red under fluorescence (Kirk, 1970)for detection of total lipids; phloroglucinol for lignin (Johansen,1940); Rutenium red (Johansen, 1940) for pectins. Some sectionsof methacrylate embedded samples were tested for total polysac-charides using periodic acid Schiff (PAS) (Mcmanus, 1948).
For the description of leaf surface characters, estimation of den-sity of trichome-like emergences and measurements of stalk lengthof the lepidote emergences, whole leaves were cleared using asolution of 10% sodium hydroxide and 20% sodium hypochlorite,
interspersed with successive washes in distilled water (Johansen,1940, modified). The leaf material was stained with safranin (1%alcoholic solution) and mounted in glycerinated gelatin.
2 logy,
mbs(rvo
mwGvm
c
2
tpsTcmso
atawtno
hwrpsp
2
ieTTuIC
3
ato
cbsu
6 N.C. Vitarelli et al. / Perspectives in Plant Eco
Density of trichome-like emergences and stalk length wereeasured on the following specimens: Croton pygmaeus (Falken-
erg 6483; Marchett 549), Croton splendidus (Alvim s.n.; Vitarelli.n.) and Croton erythroxyloides (Vitarelli 002; Vitarelli 003)Table 1). Trichome-like emergences were counted at the middleegion of both adaxial and abaxial leaf surfaces, excluding the mid-ein area. The stalk length of lepidote emergences was measuredn leaf cross sections from embedded samples.
Observations and photographs were obtained using a lighticroscope (AX70TRF, Olympus Optical, Tokyo, Japan) equippedith a U-photo system and digital camera (AxioCam HRc; Zeiss,öttingen, Germany), and an epifluorescence HBO 50W mercuryapor lamp and filter block A (exciter filter BP 340–380, dichroicirror 450, barrier filter LP-430).
The denomination of trichome-like emergence types follows thelassification by Webster et al. (1996).
.3. Leaf absorption experiment
The capacity of the trichomes to absorb an aqueous solution wasested in fully expanded leaves from the thirth or fourth node of livelants (Fig. 1D). Fresh branch samples of Croton splendidus wereprayed with a 1% solution of fluorochrome berberine hemisulfate.hese samples were kept in darkness overnight or longer. Hand-ut cross sections were made and the slides were immediatelyounted in glycerol and visualized, to prevent secondary diffu-
ion, under a fluorescent microscope. This methodology is basedn experiments conducted by Heide-Jørgensen (1990).
Whole leaves were also dipped in an India ink aqueous solution,nd kept in that solution during 4 h. Leaves were vertically posi-ioned inside a recipient containing a solution of India ink in such
way that only the apical and median portions of the leaf bladeere in contact with the solution. This was achieved by attaching
he petiole to the upper part of the walls of the container. Leavesot subjected to the above treatments with berberine hemisulfater India ink solutions were also analyzed as control samples.
The experiments using solution of fluorochrome berberineemisulfate and India Ink solution were chosen to evaluate the leafater absorption ability because they are simple and show direct
esults. This is important since it is very difficult to cultivate Crotonlants out of their natural environment and there are many con-traints to conduct experimental practices in the field due to theroblematic access to the areas where the studied species occur.
.4. Scanning electron microscopy
For analysis with a scanning electron microscope, samples fixedn Karnovsky solution (Karnovsky, 1965) were dehydrated in anthanol series and submitted to critical-point drying (CPD 020; Bal-ec, Balzers, Liechtenstein) using CO2 (Bozzola and Russel, 1992).he samples were then mounted onto stubs and coated with goldsing a FDU 010 sputter coater (Bal-Tec, Balzers, Liechtenstein).
mage captures were conducted using a Leo 1430VP SEM (Zeiss,ambridge, United Kingdom).
. Results
The epidermal structures observed in the three species studiedre formed by the activity of protoderm and ground meristem. Forhis reason, from now on we refer to them as emergences insteadf trichomes.
We observed three types of emergences: lepidote (Fig. 2A), fasci-
ulate (Fig. 2B) and stellate (Fig. 2C, D). The lepidote type occurs onoth leaf surfaces of Croton erythroxyloides and only on the abaxialide on the leaves of both C. pygmaeus and C. splendidus. The fascic-late emergences are present in the adaxial leaf side of C. splendidusEvolution and Systematics 22 (2016) 23–35
and the stellate emergences are present on the adaxial side of theleaves of C. pygmaeus.
The lepidote emergences cover all the abaxial leaf sur-face and are visible to the naked eye (Fig. 1D, E) as browndots. The density of lepidote emergences on the abaxial leafsurface was 3 emergences/�m2 in Croton erythroxyloides, 5emergences/�m2 in C. pygmaeus, and 4 emergences/�m2 inC. splendidus. On the adaxial leaf surface, the emergence den-sity was 4 lepidote emergences/�m2 in C. erythroxyloides, 1–2stellate emergences/�m2 in C. pygmaeus, and 4–5 fasciculateemergences/�m2 in C. splendidus.
3.1. Ontogenetic development of emergences
Emergences in early and mature stages could be observed onthe same leaf primordia due to their asynchrony and early devel-opment (Figs. 3 and 4). The beginning of the development of thethree types of emergences is recognized by the presence of a pro-todermal cell that stands out because of its larger volume, evidentnucleus and dense cytoplasm (Figs. 3A; 4A). The adjacent protoder-mal cells undergo a series of anticlinal divisions (Figs. 3B; 4B), giveorigin to a protuberance (Fig. 3C), in which the central cell showsan increasing in volume while the adjacent cells (lateral cells) dis-play successive anticlinal divisions (Figs. 3C–F; 4B–D). The lateralcells are radially positioned to the central cell and will develop intolateral rays by stretching (Figs. 3G, H; 4D, E). The ray cells haveperipheral nuclei (Fig. 3E, F), indicating their growing direction.
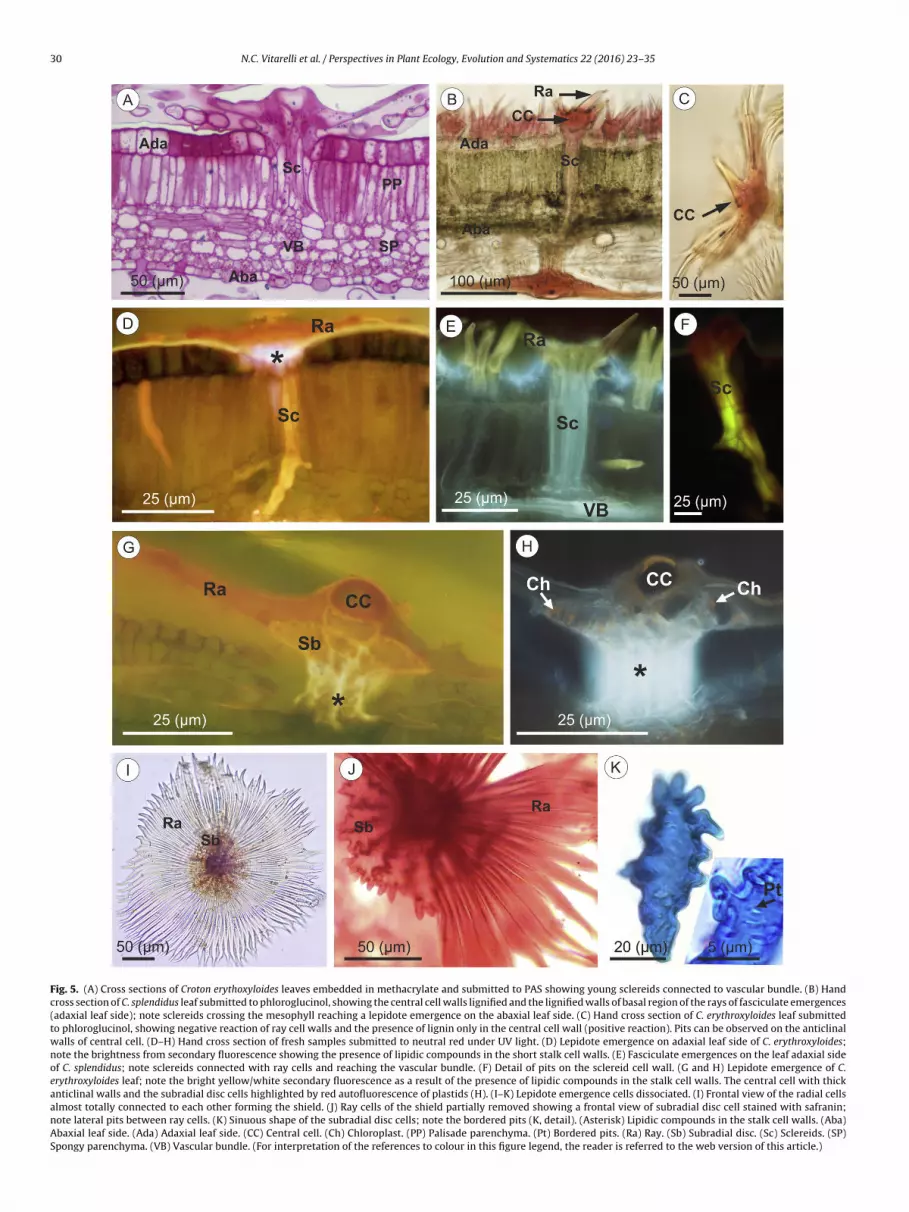
In the lepidote emergences, the lateral rays remain united bywebbing and form a shield-like structure. Simultaneously with theray cells stretching process protodermal cells, localized below therays, become bulky and form a subradial disc (Fig. 3E–H). Underthe subradial disc, protodermal and ground meristem cells forma multiseriate stalk through periclinal divisions (Fig. 3C–G). Atthe central part of the stalk, cells arising from the ground meris-tem stretch out and become precursors of sclereids (Fig. 3F, H).The sclereids allow connection between the outer region of theemergences and the interior of the leaf (Fig. 5A, B, D–F) reach-ing the vascular bundles (Fig. 5A, E) and crossing the mesophyll(Fig. 5B). Ray cells are dead in mature emergences. The ray cellwalls become thick, but non-lignified, as demonstrated by the neg-ative reaction of phloroglucinol (Table 2; Fig. 5C). Additionally, thepositive reaction to Rutenium red, along with the negative reactionto phloroglucinol, confirm that the lateral walls of the ray cells arerich in pectins and remain non-lignified even in mature structures(Table 2; Fig. 5C). In contrast, the central cell walls become thickerand undergo lignification (Table 2; Fig. 5C).
In completely mature lepidote emergences, the central cell isdead and shows a circular region at its centre (Fig. 5C, G, H), whichprobably is a more permeable zone since the anticlinal walls arethicker than the periclinal external wall. The anticlinal walls arelignified and pitted (Fig. 5C).
The central cell of the lepidote emergence is surrounded by arounded shield formed by the union of the lateral ray cells (Fig. 5I),whose anticlinal walls bear pits (Fig. 5J). In the subradial region(or subshield region) there is a disc of cells with markedly sinuousoutline and thick walls (Fig. 5J, K), that show bordered pits (Fig. 5K,detail). These sinuous cells remain metabolically active at maturityand contain numerous chloroplasts (Fig. 5H).
The above described structure (shield and subshield regions)is supported by a stalk of variable length: 80 �m on the abaxialleaf surface of C. erythroxyloides, 44.18–58.3 �m on the abaxial leafsurface of C. pygmaeus, and 80.72–101.09 �m on the abaxial leaf
surface of C. splendidus. The stalk of lepidote emergences presenton the adaxial leaf surface of C. erythroxyloides is almost sessilewith approximately 40 �m of length (Figs. 3H; 5A, D), and stel-late and fasciculate emergences are sessile (Fig. 5B). At the end
N.C. Vitarelli et al. / Perspectives in Plant Ecology, Evolution and Systematics 22 (2016) 23–35 27
Fig. 2. Types of emergences observed in the three species studied. (A and B) Scanning electron microscopy images from Croton splendidus leaf. (A) Lepidote emergenceson the leaf abaxial side. (B) Fasciculate emergences on the leaf adaxial side. (C and D) Frontal view of cleared leaves of C. pygmaeus from the adaxial side showing stellateemergences.
Table 2Chemical nature of substances found in the cellular walls of different parts of leaf emergences.
Region Chemical nature of thesubstance
Reaction results/emergence type
Ray cells Lipids (−) lepidote; (−) stellate; (−) fasciculate
Lignin (−) lepidote; (+) stellate, in the rays basal region; (+) fasciculate, in therays basal region
Pectin (+) lepidote; (+) stellate; (+) fasciculate
Central cell Lipids (−) lepidote, stellate and fasciculate
Lignin (+) lepidote, stellate and fasciculate
Pectin (−) lepidote, stellate and fasciculate
Subradial disc cells Lipids (−) lepidote
Lignin Inconclusive due the brown natural color of these cells
Pectin (+) lepidote
Stalk cell walls Lipids (+) lepidote
Lignin (−) lepidote
Pectin (+) lepidote
Sclereids Lipids (−) lepidote, stellate and fasciculate
Lignin (−) lepidote, stellate and fasciculate; or weak reaction in the threetypes
odwanc
bcrr(
eH
Pectin
f the emergence development the stalk cells appear dead and aense lipid layer is deposited over their anticlinal and periclinalalls constituting the external region of the stalk (Fig. 5D, G, H),
s displayed by the positive reaction to Sudan IV (visible light) andeutral red (under fluorescent light, showing a bright white/blueolor as shown in Fig. 5D, G, H).
Completely developed lepidote emergences are characterizedy three different regions: the outermost part comprised by theentral cell and the lateral rays or shield (Fig. 5C, I); the medianegion, formed by the subradial disc cells (Fig. 5J); and the basalegion or stalk (Fig. 5G, H) internally constituted by sclereids
Fig. 5D).The ontogenetic development of the stellate and fasciculatemergences is similar to the development of the lepidote ones.owever, stellate and fasciculate emergences are always sessile or
(+) lepidote, stellate and fasciculate
short-stalked, while in lepidote emergences the stalk can be longor short. In the sessile and short-stalked emergences the rays aredirectly connected with the sclereids of the mesophyll (Fig. 5D–F).Additionally, the central cell of the stellate and fasciculate emer-gences becomes more stretched, forming a central porrect ray(Fig. 4C–E). The lateral cells stretch and form the rays in a singlewhorl (Fig. 4E). The same process takes place in lepidote, stellateand fasciculate emergences. The difference between them is thatin the lepidote ones the ray cells become totally or partially unitedby the lateral walls forming a rounded shield (Figs. 2A; 6 A). Inthe stellate emergences, the ray cells remain free in the same plane
(Fig. 2C, D). The fasciculate emergences differ from the stellate onesin having ascending rays rather than positioned in a horizontalplane (Figs. 2B; 4E).
28 N.C. Vitarelli et al. / Perspectives in Plant Ecology, Evolution and Systematics 22 (2016) 23–35
Fig. 3. (A–H) Ontogenetic development of lepitote emergences on the leaves of Croton erythroxyloides. Cross sections of leaves embedded in methacrylate and stained withtoluidine blue. (A) A typical initial cell of an emergence in the protoderm. (B) Dome-shaped epidermal protuberance produced by protoderm and ground meristem activity.Note the anticlinal divisions of protodermal cells adjacent to central cell. (C and D) Dome in radial section (C), showing the lateral cells (LC1 and LC2) arranged radially tothe central cell (*); dome in tangential section (D), showing the lateral cells (or radial cells) originated by anticlinal divisions of protoderm. In C and D, note the periclinaldivisions of ground meristem cells, starting the development of the stalk (arrow). (E and F) Anticlinal divisions of radial cells of emergence in tangential section (E) and radialsection (F); note nucleus located peripherally in the radial cells; note the start of development of the subradial disc (arrow head) and the elongation of the stalk (arrow). (G)W e subs . (Arroc mal c
3
ghelot
all thickening of central cell, stretching of the radial cells and development of thhowing live central cell, radial cells, subradial cells, short stalk and young sclereidsell or radial cell that will become a ray. (GM) Ground meristem cells. (Pd) Protoder
.2. Foliar absorption through emergences
The absorption capacity of the lepidote and fasciculate emer-ences was demonstrated by the experiments with berberineemisulfate and India ink solutions. The water pathways werelucidated by these experiments and were observed by the yel-
ow/blue fluorescence under UV light (Fig. 6B–E) after applicationf berberine hemisulfate spray. No fluorescence was produced byhe control samples.radial disc (arrow head) and stalk (arrow). (H) Almost fully developed emergencew) Stalk cells. (Arrow head) Subradial disc cells. (Asterisk) Central cell. (LC) Lateral
ells. (Sc) Sclereids.
The lepidote, stellate and fasciculate emergences of the threeCroton species studied are structurally complex. The rays of theemergences increase the contact area with the exterior (Fig. 5A–J),thus increasing the surface area for absorption of atmosphericmoisture. The lepidote emergences are even more specialized inwater capture than the other two (stellate and fasciculate) because
they possess subradial disc cells (Fig. 5J) and many more rays form-ing their shield-like structure (Fig. 5C, I, J). These two featuresprovide a larger surface of contact for the lepidote emergence.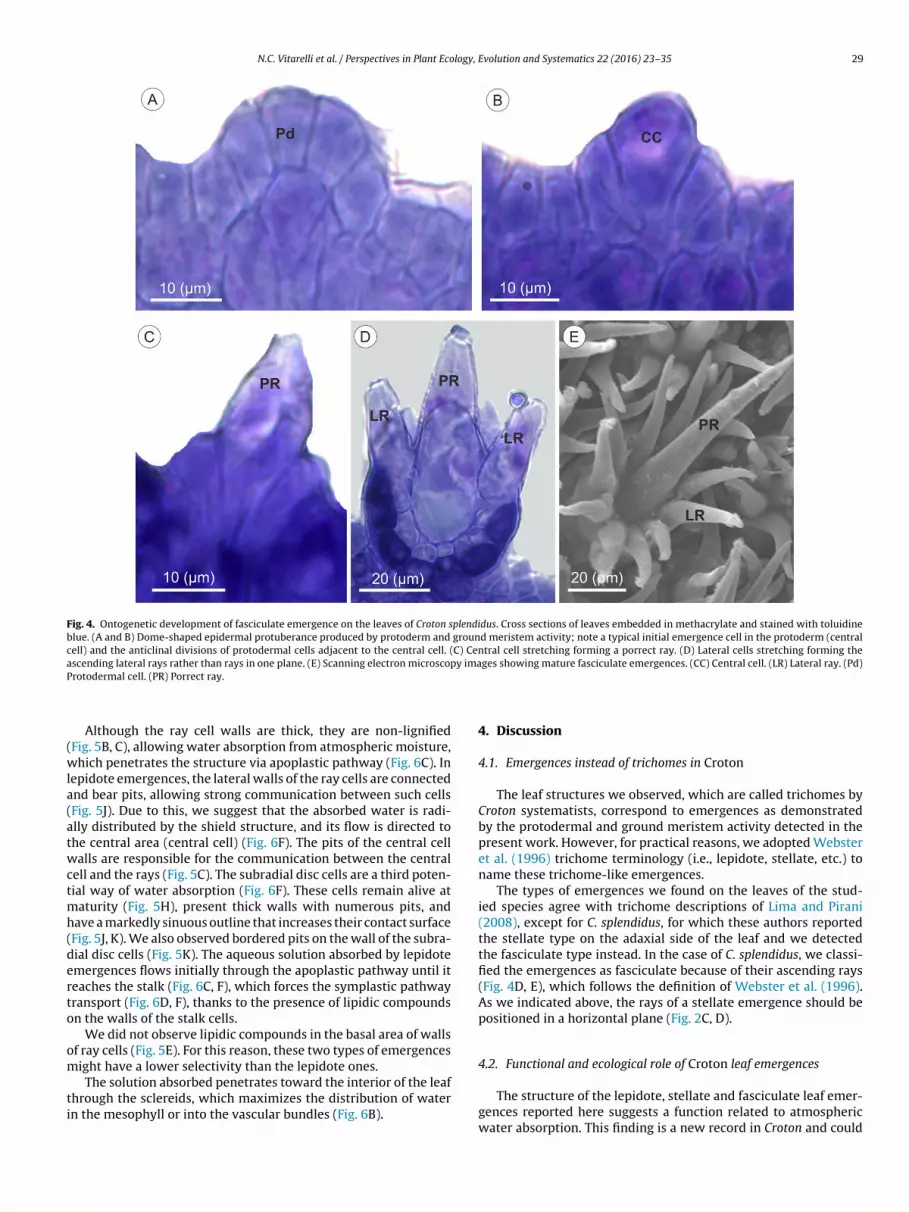
N.C. Vitarelli et al. / Perspectives in Plant Ecology, Evolution and Systematics 22 (2016) 23–35 29
Fig. 4. Ontogenetic development of fasciculate emergence on the leaves of Croton splendidus. Cross sections of leaves embedded in methacrylate and stained with toluidineblue. (A and B) Dome-shaped epidermal protuberance produced by protoderm and ground meristem activity; note a typical initial emergence cell in the protoderm (centralcell) and the anticlinal divisions of protodermal cells adjacent to the central cell. (C) Central cell stretching forming a porrect ray. (D) Lateral cells stretching forming thea y imaP
(wla(atwctmh(derto
om
ti
scending lateral rays rather than rays in one plane. (E) Scanning electron microscoprotodermal cell. (PR) Porrect ray.
Although the ray cell walls are thick, they are non-lignifiedFig. 5B, C), allowing water absorption from atmospheric moisture,hich penetrates the structure via apoplastic pathway (Fig. 6C). In
epidote emergences, the lateral walls of the ray cells are connectednd bear pits, allowing strong communication between such cellsFig. 5J). Due to this, we suggest that the absorbed water is radi-lly distributed by the shield structure, and its flow is directed tohe central area (central cell) (Fig. 6F). The pits of the central cellalls are responsible for the communication between the central
ell and the rays (Fig. 5C). The subradial disc cells are a third poten-ial way of water absorption (Fig. 6F). These cells remain alive at
aturity (Fig. 5H), present thick walls with numerous pits, andave a markedly sinuous outline that increases their contact surfaceFig. 5J, K). We also observed bordered pits on the wall of the subra-ial disc cells (Fig. 5K). The aqueous solution absorbed by lepidotemergences flows initially through the apoplastic pathway until iteaches the stalk (Fig. 6C, F), which forces the symplastic pathwayransport (Fig. 6D, F), thanks to the presence of lipidic compoundsn the walls of the stalk cells.
We did not observe lipidic compounds in the basal area of wallsf ray cells (Fig. 5E). For this reason, these two types of emergencesight have a lower selectivity than the lepidote ones.
The solution absorbed penetrates toward the interior of the leafhrough the sclereids, which maximizes the distribution of watern the mesophyll or into the vascular bundles (Fig. 6B).
ges showing mature fasciculate emergences. (CC) Central cell. (LR) Lateral ray. (Pd)
4. Discussion
4.1. Emergences instead of trichomes in Croton
The leaf structures we observed, which are called trichomes byCroton systematists, correspond to emergences as demonstratedby the protodermal and ground meristem activity detected in thepresent work. However, for practical reasons, we adopted Websteret al. (1996) trichome terminology (i.e., lepidote, stellate, etc.) toname these trichome-like emergences.
The types of emergences we found on the leaves of the stud-ied species agree with trichome descriptions of Lima and Pirani(2008), except for C. splendidus, for which these authors reportedthe stellate type on the adaxial side of the leaf and we detectedthe fasciculate type instead. In the case of C. splendidus, we classi-fied the emergences as fasciculate because of their ascending rays(Fig. 4D, E), which follows the definition of Webster et al. (1996).As we indicated above, the rays of a stellate emergence should bepositioned in a horizontal plane (Fig. 2C, D).
4.2. Functional and ecological role of Croton leaf emergences
The structure of the lepidote, stellate and fasciculate leaf emer-gences reported here suggests a function related to atmosphericwater absorption. This finding is a new record in Croton and could
30 N.C. Vitarelli et al. / Perspectives in Plant Ecology, Evolution and Systematics 22 (2016) 23–35
Fig. 5. (A) Cross sections of Croton erythoxyloides leaves embedded in methacrylate and submitted to PAS showing young sclereids connected to vascular bundle. (B) Handcross section of C. splendidus leaf submitted to phloroglucinol, showing the central cell walls lignified and the lignified walls of basal region of the rays of fasciculate emergences(adaxial leaf side); note sclereids crossing the mesophyll reaching a lepidote emergence on the abaxial leaf side. (C) Hand cross section of C. erythroxyloides leaf submittedto phloroglucinol, showing negative reaction of ray cell walls and the presence of lignin only in the central cell wall (positive reaction). Pits can be observed on the anticlinalwalls of central cell. (D–H) Hand cross section of fresh samples submitted to neutral red under UV light. (D) Lepidote emergence on adaxial leaf side of C. erythroxyloides;note the brightness from secondary fluorescence showing the presence of lipidic compounds in the short stalk cell walls. (E) Fasciculate emergences on the leaf adaxial sideof C. splendidus; note sclereids connected with ray cells and reaching the vascular bundle. (F) Detail of pits on the sclereid cell wall. (G and H) Lepidote emergence of C.erythroxyloides leaf; note the bright yellow/white secondary fluorescence as a result of the presence of lipidic compounds in the stalk cell walls. The central cell with thickanticlinal walls and the subradial disc cells highlighted by red autofluorescence of plastids (H). (I–K) Lepidote emergence cells dissociated. (I) Frontal view of the radial cellsalmost totally connected to each other forming the shield. (J) Ray cells of the shield partially removed showing a frontal view of subradial disc cell stained with safranin;note lateral pits between ray cells. (K) Sinuous shape of the subradial disc cells; note the bordered pits (K, detail). (Asterisk) Lipidic compounds in the stalk cell walls. (Aba)Abaxial leaf side. (Ada) Adaxial leaf side. (CC) Central cell. (Ch) Chloroplast. (PP) Palisade parenchyma. (Pt) Bordered pits. (Ra) Ray. (Sb) Subradial disc. (Sc) Sclereids. (SP)Spongy parenchyma. (VB) Vascular bundle. (For interpretation of the references to colour in this figure legend, the reader is referred to the web version of this article.)

N.C. Vitarelli et al. / Perspectives in Plant Ecology, Evolution and Systematics 22 (2016) 23–35 31
Fig. 6. (A) Scanning electron microscopy showing a general view of a Croton splendidus leaf in cross section. (B–E) Hand-cut cross section of fresh C. splendidus leavesafter sprayed with berberine hemisulfate showing the water uptake pathways by secondary fluorescence under UV light. (B) General view of the water pathways. Observethe absorption of the solution by emergences and their distribution to the vascular bundles by sclereids. (C and D) Detail of the solution pathway in lepidote emergenceabsorption. The solution is absorbed by rays, central cell and subradial disc, following the apoplastic pathway. The transfer of water from apoplastic (C) to symplastic pathway(D) occurs in the stalk cells. (E) The absorbed solution reaching the leaf main vasculature. (F) Schematic drawing of lepidote emergence showing the water pathway. Thewater, represented by blue balls, is absorbed by the thick and non-lignified walls (gray color) of the ray cells, central cell and subradial disc cells. The water flows initiallythrough the apoplastic pathway until it reaches the stalk. The lipidic walls (beige color) of the cells of the stalk force the symplastic pathway transport. (Arrow head) Solutionp ) FasciS the ret
reirteaor
athway. (Asterisk) Central Cell. (Aba) Abaxial leaf side. (Ada) Adaxial leaf side. (FEubradial disc. (Sc) Sclereids. (St) Stalk. (VB) Vascular bundle. (For interpretation ofhis article.)
epresent a survival strategy for populations of this genus living innvironments with evident water restrictions such as the Brazil-an rock outcrops (campos rupestres and campos de altitude). Waterestriction in these habitats is caused by the low soil water reten-ion due to the rocky surface and rough relief (Safford, 1999; Benites
t al., 2003; Caiafa and Silva, 2007). Most of the literature on waterbsorption through leaves and stems in vascular plants has focusedn the significance of this pathway for plants living in arid envi-onments (Rundel, 1982). Even if the Brazilian rock outcrops areculate emergence. (LE) Lepidote emergence. (MV) Main vasculature. (Ra) Ray. (Sb)ferences to colour in this figure legend, the reader is referred to the web version of
characterized by water soil restriction, there is a significant hydricand nutrition supplement available from the daily fogs (Gundelet al., 1994; Burgess and Dawson, 2004; Eller et al., 2013), which arepresent all year long. According to Eller et al. (2013), the occurrenceof frequent fog events is a defining climatic attribute of tropical
montane environments. Cavelier et al. (1996) and Bruijnzeel et al.(2011) report that the direct contact of fog water droplets withthe surface of stems or leaves causes water to drip to the soil, andthis additional precipitation is considered to be a major hydrologi-
3 logy,
cattnacfsaa
r2SmtBascVrf(
hsa1C
4
ttdglFfpascs2lin
sof1o2mopwMart
2 N.C. Vitarelli et al. / Perspectives in Plant Eco
al input in these tropical montane environments. Species that areble to use this atmospheric source of water and nutrients, as thehree species studied here, might be more successful in colonizinghese types of environment. It is important to highlight that fog isot pure water but a heterogeneous substance composed of waternd dissolved ions (Houghton, 1955; Gundel et al., 1994), whichould be also important for plant nutrition. Questions requiringurther investigation are: how relevant is this atmospheric waterupply to the hydric balance and nutrition of the studied species?nd how significant is it for the species success in habitats with lowvailability of soil water?
The ability to capture water through leaves has been indirectlyeported in species occurring in deserts (Martin and von Willert,000) but also in cloud-immersed mountain habitats (Berry andmith, 2012, 2013; Berry et al., 2014; Eller et al., 2013) and coastalountain regions where fog is a significant climatic contribu-
or of moisture (Burgess and Dawson, 2004; Oliveira et al., 2005;reshears et al., 2008; Limm et al., 2009; Simonin et al., 2009; Limmnd Dawson, 2010; Berry et al., 2014; Gotsch et al., 2014). Crotonpecies distributed in forest ecosystems also have sclereids asso-iated with epidermal trichome-like emergences (Soares, 2013;itarelli, 2013), and these structures may play the same absorption
ole reported here, similarly to what has been described for speciesrom tropical montane cloud forest such as Drimys brasiliensis MiersEller et al., 2013).
We believe that many other Croton species, if not all of them,ave emergences that are similar to the ones described here,ince several authors reported the presence of Croton trichomesssociated with sclereids in the mesophyll (Metcalfe and Chalk,950; Starling, 1993; Louro et al., 2003; Appezzato-da-Glória andarmello-Guerreiro, 2006; Sá-Haiad et al., 2009).
.3. Atmospheric water uptake in Croton and other plant groups
The results of this study show the capacity of leaves fromhree species of Croton to absorb atmospheric water throughhe leaves. According to Rundel (1982), laboratory experimentsemonstrated the ability of leaves to absorb water in differentroup of plants. However there are still controversies on the eco-ogical and physiological significance of this way of water uptake.or water absorption and transfer to occur, a source of water and aavorable water potential gradient are necessary. In addition, mor-hological or anatomical structures must be present to speed thebsorption process (Rundel, 1982). These structures may includepecialized trichomes (Stuart, 1968; Benzing et al., 1976), spe-ialized epidermal cells (Grundell, 1933; Meidner, 1954) and/orpecialized cuticle features (Leyton and Armitage, 1968; Eller et al.,013). There are only a few reports in the literature demonstrating
eaf water absorbtion in flowering plants (see below), which mightndicate that either leaf water absorbtion is a rare strategy or it hasot been systematically studied across angiosperms.
The experiments with berberine hemisulfate and Indian inkolutions confirmed the absorption ability of the leaf emergencesf the studied Croton species. Similar experiments have been per-ormed in Hakea suaveolens R. Br. (Proteaceae) (Heide-Jørgensen,990) using berberine sulfate fluorochrome, and in Microlepisleaefolia (DC.) Triana (Melastomataceae) (Milanez and Machado,008) using aqueous safranin solution. In both cases, the per-eability of the epidermis was demonstrated by the penetration
f the solution through trichomes and sclereids to the meso-hyll and vascular bundles. Fernández et al. (2014) studied theater absorption of holm oak leaves (Quercus ilex L.), an evergreen
editerranean species, and found that the upper side of youngnd mature leaves was wettable and absorbed water, and theyelated this uptake mechanism to the presence of trichomes andrichome scars. Burrows et al. (2013) detected stellate trichomes
Evolution and Systematics 22 (2016) 23–35
with intrusive bases on the leaves of Solanum elaeagnifolium Cav.They reported that the trichome basal cells penetrate into the mes-ophyll and sometimes reach the bundle sheath of the vein, butwithout a direct contact with the xylem cells. The stellate trichomesof Solanum elaeagnifolium are very similar to the stellate emer-gences found in Croton. However, in S. elaeagnifolium the transportbetween trichomes and veins was not confirmed by symplastic(carboxyfluorescein diacetate) or apoplastic (lucifer yellow) tracerdyes (Burrows et al., 2013). Trichomes associated with sclereidswere also reported in Jovetia humilis Guédès (Rubiaceae), a Mala-gasy xerophytic shrub, and their structure suggests a water uptakefunction (Guédès, 1975).
The overall structure of the emergences of Croton erythroxy-loides, C. pygmaeus and C. splendidus is similar to the xerophytictrichomes described by Fahn (1986) in Correa backhouseana Hook.(Rutaceae) and Olea europaea L. (Oleaceae), which have lipidic com-pounds covering the walls of their basal cell and are also associatedwith atmospheric water absorption. However, C. backhouseana andO. europaea lack the complex structure of the stalk-sclereid andsubradial disc cells described in the Croton emergences. Accordingto Fahn (1986), the layer of lipidic compounds observed in basalor stalk cells of the trichomes of 12 xeromorphic species he stud-ied plays a function similar to that of the endodermis, inducing aswitch from the apoplastic to the symplastic pathway. A similarstructure was observed in absorbent trichomes in species of sub-tribe Pleurothallidinae in the family Orchidaceae (Pridgeon, 1981).The selectivity function of absorbent trichomes (or other leaf epi-dermal structures) is relevant, since fog water is a solution of waterand ions (Gundel et al., 1994; Houghton, 1955).
In the lepidote emergences of the Croton studied here, the stalkis the region where the selectivity of the absorbing solution occurs.This function is also assigned to the subradial disc cells, since thosecells are apparently the only ones that remain alive and metabol-ically active in mature emergences, as attested by the presence ofplastids. Nevertheless, there is no physical obstruction to preventthe apoplastic flow through the subradial disc cells. This regionof the lepidote emergence could also be associated with move-ments that change the orientation of the lateral rays in responseto changes in atmospheric humidity, resembling the movementsof the Tillandsioideae scales in Bromeliaceae (Benzing et al., 1976).However, complementary ultrastructural and morphophysiolog-ical studies are required to confirm this hypothesis. Our resultsshow that the subradial disc cells are highly specialized structuresimplied in absorption and transport of water due to their markedlysinuous outline and the presence of bordered pits on their walls. Thesubradial disc cells are similar to the modified tracheids present inthe transfusion tissue of gymnosperms (von Mohl, 1871), which areresponsible for short distance water transport.
Anatomically, the lepidote emergences on the leaves of Crotonerythroxyloides, C. pygmaeus and C. splendidus are someway simi-lar to the leaf scales of Tillandsioideae species (Tomlinson, 1969;Benzing et al., 1976). However in Tillandsioideae, the stalk cells(named dome cells) are alive and present a complex structure withinvaginations of the plasmatic membrane and numerous mito-chondria, being specialized in the absorption function (Benzinget al., 1976), whereas in Croton the stalk cells are specialized in theselectivity function. Besides this, the lepidote emergences in Crotonshow a greater structural complexity than the scales of Tilland-sioideae, due to the presence of the subradial disc cells and thesclereids.
4.4. The emergence-sclereid complex in Croton
Sclereids are the most polymorphic cells in the plant body andcan be found in clusters or as isolated cells (named idioblasts)(Evert, 2006). Several studies have focused on their morphologi-

logy, E
cRssrEoeA
EMseCWtCSss
iT(((pAnfigtoicumsaba
5
ybtosstbtow
F
mat
N.C. Vitarelli et al. / Perspectives in Plant Eco
al characterization and taxonomic importance (e.g., Foster, 1956;ao and Das, 1979; Rao, 1991), however, the functional role of theclereids is still incompletely understood. Sclereids are described asclerenchyma cells that present secondary thick and lignified wallselated to the support tissue of plant organs (Haberlandt, 1882;vert, 2006). However, some studies demonstrate the active rolef sclereids in the water pathway in leaves of species adapted tonvironments with low water supply (Tucker, 1964; Fahn, 1986;lvin, 1987; Heide-Jørgensen, 1990; Milanez and Machado, 2008).
The presence of sclereids connected with leaf trichomes inuphorbiaceae species has already been reported (Solereder, 1908;etcalfe and Chalk, 1950; Rudall, 1994), including species of Croton
uch as C. alchorneicarpus Croizat, C. caldensis Müll. Arg. (Sá-Haiadt al., 2009), C. compressus Lam. (Sá-Haiad, 1987; Louro et al., 2003),. fuscescens Spreng. (Metcalfe and Chalk, 1950), C. rhamnifoliusilld. (Appezzato-da-Glória and Carmello-Guerreiro, 2006) and C.
riqueter Lam. (Starling, 1993). This emergence-sclereid complex inroton leaves has been interpreted as tissue support (Barros andoares, 2013), although the alternative role of absorption has beenuggested by Louro et al. (2003), but without experimental demon-tration.
The sclereids from lepidote, stellate and fasciculate emergencesn Croton have thick and non-lignified (or weakly lignified) walls.his unusual pattern in sclereids was also observed in Olea europaeaArzee, 1953) and in populations of Androstachys johnsonii PrainPicrodendraceae) growing in environments with water restrictionAlvin, 1987). The connection between leaf trichomes and meso-hyll sclereids has been directly related with leaf water uptake in. johnsonii (Alvin, 1987). This author suggested the name “gelati-ous sclereids” due to the structural similarity with gelatinousbers, which possess a G-layer (so-called gelatinous layer) thatives them the hygroscopic capacity to catch and store water. Inhe leaves of Hakea suaveolens, the surface of the branches of mes-phyll osteosclereids constitutes a pseudohypodermis and their
nternal arms are in contact with the modified tracheids in the vas-ular ends. The sclereids remain alive and take part in the waterptake function, allowing the water to flow from the leaf epider-is to the photosynthetic tissues (Heide-Jørgensen, 1990). In the
tudied Croton species, the sclereids carry water into the xylemnd/or distribute it into the mesophyll, resembling the role of theundle-sheath extension described by Plymale and Wylie (1944)nd reported in xerophytic leaves (Fahn, 1986).
. Conclusions
The so called trichomes present on the leaves of Croton erythrox-loides, C. pygmaeus and C. splendidus correspond to emergencesecause they originate from the protodermis and ground meris-em. The anatomical origin, structure, as well as the functional rolef these emergences are described in detail for the first time. Theirtructural complexity along with our experimental results providetrong evidence that they may play an important role in the absorp-ion of atmospheric water by these Croton species and potentiallyy other species in the genus. This ability to capture water throughhe leaves may be one of the clues for the evolutionary successf this speciose group common in environments with limited soilater supply in tropics and subtropics worldwide.
unding
This work was supported by Conselho Nacional de Desenvolvi-ento Científico e Tecnológico [477867/2013-8]; Fundac ão de
mparo à Pesquisa do Estado de Minas Gerais [CRA-APQ]; and Cen-ro de Ciências Ambientais Floresta Escola [FUNARBE-6991].
volution and Systematics 22 (2016) 23–35 33
Acknowledgements
We are grateful to the IEF (Instituto Estadual de Floresta) for thepermits to collect samples in Parque Estadual da Serra do Brigadeiro(Minas Gerais, Brazil), the Centro de Microscopia e Microanálisesat the Universidade Federal de Vic osa where the scanning electronmicroscopy analysis was conducted, and the curators of herbariaBHCB, HUEMG, and VIC. Inês Cordeiro confirmed the identificationof specimens. Katiuscia Freire de Souza conducted measurementson emergence density and stalk length. This work was supported byCNPq, FAPEMIG and Centro de Ciências Ambientais Floresta Escola(FUNARBE-6991). NCV received support from CAPES (Cordenac ãode Aperfeic oamento de Pessoal de Nível Superior), and RMSAM wassupported by a scholarship from CNPq. RR was supported by a vis-iting scholar grant to Brazil in 2015 (FAPEMIG CRA-BPV-00043-14).Four anonymous reviewers provided helpful comments and usefulsuggestions on earlier versions of this paper.
References
Alcantara, S., Mello-Silva, R., Teodoro, G., Drequeceler, K., Ackerly, D.D., Oliveira,R.S., 2015. Carbon assimilation and habitat segregation in resurrection plants:a comparison between desiccation- and non-desiccation-tolerant species ofNeotropical Velloziaceae (Pandanales). Funct. Ecol., http://dx.doi.org/10.1111/1365-2435.12462.
Alves, R.J.V., Kolbek, J., 2010. Can campos rupestres vegetation be floristicallydelimited based on vascular plant genera? Plant Ecol. 207, 67–79.
Alvin, K.L., 1987. Leaf anatomy of Androstachys johnsonii Prain and its functionalsignificance. Ann. Bot. 59, 579–591.
Appezzato-da-Glória, B., Carmello-Guerreiro, S.M., 2006. Anatomia Vegetal, 2ndedn. UFV, Vic osa.
Araújo, J.S., Azevedo, A.A., Silva, L.C., Meira, R.M.S.A., 2010. Leaf anatomy as anadditional taxonomy tool for 16 species of Malpighiaceae found in the Cerradoarea. Plant Syst. Evol. 286, 117–131 (Brazil).
Arzee, T., 1953. Morphology and ontogeny of foliar sclereids in Olea europaea I.Distribution and structure. Am. J. Bot. 40, 680–687.
Barros, I.O., Soares, A.A., 2013. Adaptac ões anatômicas em folhas de marmeleiro evelame da caatinga brasileira. Rev. Cienc. Agron. 44, 192–198.
Benites, V.M., Caiafa, A.N., Mendonc a, E.S., Schaefer, C.E., Ker, J.C., 2003. Solos evegetac ão nos complexos rupestres de altitude da Mantiqueira e do Espinhac o.Floresta e Ambiente 10, 76–85.
Benites, V.M., Schaefer, C.E.R., Simas, F.N.B., Santos, H.G., 2007. Soil associated withrock outcrops in the Brazilian mountain ranges Mantiqueira and Espinhac o.Rev. Bras. Bot. 30, 569–577.
Benites, V.M., 1998. Caracterizac ão química e espectroscópica da matéria orgânicae suas relac ões com a gênese dos solos da Serra do Brigadeiro, Zona da MataMineira. MSc Thesis. Universidade Federal de Vic osa, Brazil.
Benzing, D.H., Henderson, K., Kessel, B., Sulak, J., 1976. The absorptive capacities ofbromeliad trichomes. Am. J. Bot. 63, 1009–1014.
Berry, P.E., Hipp, A.L., Wurdack, K.J., van Ee, B.W., Riina, R., 2005. Molecularphylogenetics of the giant genus Croton and tribe Crotoneae (Euphorbiaceaesensu stricto) using ITS and trnL-trnF DNA sequence data. Am. J. Bot. 92,1520–1534.
Berry, Z.C., Smith, W.K., 2012. Cloud pattern and water relations in Picea rubensand Abies fraseri southern Appalachian Mountains, USA. Agric. For. Meteorol.162, 27–34.
Berry, Z.C., Smith, W.K., 2013. Ecophysiological importance of cloud immersion in arelic spruce-fir forest at elevational limits, southern Appalachian Mountains,USA. Oecologia 173, 637–648.
Berry, Z.C., Hughes, N.M., Smith, W.K., 2014. Cloud immersion: an important watersource for spruce and fir saplings in the southern Appalachian Mountains.Oecologia 174, 319–326.
Bozzola, J.J., Russel, L.D., 1992. Electron Microscopy. Jones and Bartlett Publishers,Boston.
Breshears, D.D., McDowell, N.G., Goddard, K.L., et al., 2008. Foliar absorption ofintercepted rainfall improves woody plant water status most during drought.Ecology 89, 41–47.
Bruijnzeel, L.A., Mulligan, M., Scatena, F.N., 2011. Hydrometeorology of tropicalmontane cloud forests: emerging patterns. Hydrol. Process. 25, 465–498.
Burgess, S.S.O., Dawson, T.E., 2004. The contribution of fog to the water relations ofSequoia sempervirens (D. Don): foliar uptake and prevention of dehydration.Plant Cell Environ. 27, 1023–1034.
Burrows, G.E., White, R.G., Harper, J.D.I., et al., 2013. Intrusive trichome bases in theleaves of silverleaf nightshade (Solanum elaeagnifolium; Solanaceae) do notfacilitate fluorescent tracer uptake. Am. J. Bot. 100, 2307–2317.
Caiafa, A.N., Silva, A.F., 2007. Structural analysis of the vegetation on a highlandgranitic rock outcrop in southeast Brazil. Rev. Bras. Bot. 30, 657–664.
Caruzo, M.B.R., van Ee, B.W., Cordeiro, I., Berry, P.E., Riina, R., 2011. Molecularphylogenetics and character evolution of the Sacaca clade: novel relationshipsof Croton section Cleodora. Mol. Phylogenet. Evol. 60, 193–206 (Euphorbiaceae).

3 logy,
C
CD
E
E
E
E
F
F
F
F
G
G
G
G
G
G
H
H
H
H
JK
K
K
KL
LL
L
L
L
L
L
L
L
M
4 N.C. Vitarelli et al. / Perspectives in Plant Eco
avelier, J., Solis, D., Jaramillo, M.A., 1996. Fog interception in montane forestsacross the central Cordillera of Panama. J. Trop. Ecol. 12, 357–369.
lark, G., 1973. Staining Procedures. The Williams & Wilkins Co., Baltimore.ycus, A.M., Knudson, L., 1957. The role of the velamen of the aerial roots of
orchids. Bot. Gaz. 119, 78–87.ames, A.J., McDaniels, L.H., 1947. An Introduction to Plant Anatomy, 2nd edn.
McGraw-Hill, New York USA.ller, C.B., Lima, A.L., Oliveira, R.S., 2013. Foliar uptake of fog water and transport
belowground alleviates drought effects in the cloud forest tree species, Drimysbrasiliensis. New Phytol. 199, 151–162 (Winteraceae).
ngevix, 1995. Caracterizac ão do meio físico da área autorizada para a criac ão doParque Estadual da Serra do Brigadeiro. Relatório técnico final dos estudos.Instituto Estadual de Floresta, BIRD/PRÓ-FLORESTA/SEPLAN, Brazil.
vert, R.F., 2006. Esau’s Plant Anatomy. In: Meristems, Cells, and Tissues of thePlant Body–Their Structure, Function, and Development, 3rd edn. Wiley, NewJersey.
ahn, A., 1986. Structural and functional properties of trichomes of xeromorphicleaves. Ann. Bot. 57, 631–637.
ernández, V., Sancho-Knapik, D., Guzmán, P., et al., 2014. Wettability, polarity,and water absorption of holm oak leaves: effect of leaf side and age. PlantPhysiol. 166, 168–180.
oster, A.S., 1956. Plant idioblasts: remarkable examples of cell specialization.Protoplasma 46, 184–193.
rancino, D.M.T., Coutinho, I.A.C., Dalvi, V.C., Azevedo, A.A., Conceic ão, A.S., Meira,R.M.S.A., 2015. Anatomical interpretations of the taxonomy of Chamaecrista(L.) Moench sect. Absus (Leguminosae–Caesalpinioideae). Plant Syst. Evol. 301,2087–2103.
otsch, S.G., Asbjornsen, H., Holwerda, F., Goldsmith, G.R., Weintraub, A.E., Dawson,T.E., 2014. Foggy days and dry nights determine crown-level water balance in aseasonal tropical montane cloud forest. Plant Cell Environ. 37, 261–272.
ovaerts, R., Frodin, D.G., Radcliffe-Smith, A., 2000. World Checklist andBibliography of Euphorbiaceae (and Pandaceae), vol. 2. Royal Botanic Gardens,Kew.
riffiths, C.A., Gaff, D.F., Neale, A.D., 2014. Drying without senescence inresurrection plants. Front. Plant Sci. 5, 36.
rundell, R., 1933. Zur anatomie von Myrothamnus flabellifolia Welw. Symb. Bot.Upsal. 2, 1–17.
uédès, M., 1975. Intrusive hair sclereids in Jovetia. Bot. J. Linn. Soc. 71, 141–144(Rubiaceae).
undel, L.A., Benner, W.H., Hansen, A.D.A., 1994. Chemical composition of fogwater and interstitial aerosol in Berkeley, California. Atmos. Environ. 28,2715–2725.
aberlandt, G., 1882. Vergleichend anatomie des assimilatorischengewebe-systems der pflanzen. Jahrb. f. Wiss. Bot. 13, 74–188.
amba, Y.T., Kogami, H., Terashima, I.H., 2002. The effect of growth irradiance onleaf anatomy and photosynthesis in Acer species differing in light demand.Plant Cell Environ. 25, 1021–1030.
eide-Jørgensen, H.S., 1990. Xeromorphic leaves of Hakea suaveolens R. Br.Ontogeny, structure and function of sclereids. Aust. J. Bot. 38, 25–43.
oughton, H.G., 1955. On the chemical composition of fog and cloud water. J.Meteorol. 12, 355–357.
ohansen, D.A., 1940. Plant Microtechnique. McGraw-Hill, New York.oeppen, W., 1948. Climatologia. Fondo de Cultura Económica, México – Buenos
Aires.arabourniotis, G., Esseas, C., 1996. The dense indumentum with its polyphenol
content may replace the protective role of the epidermis in some youngxeromorphic leaves. Can. J. Bot. 74, 347–351.
arnovsky, M.J., 1965. A formaldehyde glutaraldehyde fixative of high osmolalityfor use in electron microscopy. J. Cell Biol. 27, 137–138.
irk Jr., P.W., 1970. Neutral red as a lipid fluorochrome. Stain Technol. 45, 1–4.üttge, U., Haridasan, M., Fernandes, G.W., Mattos, E.A., Trimborn, P., Franco, A.C.,
et al., 1998. Photosynthesis of mistletoes in relation to their host at varioussites of tropical Brazil. Trees 12, 167–174.
evin, D.A., 1973. The role of trichomes in plant defense. Q. Rev. Biol. 48, 3–15.eyton, L., Armitage, I.P., 1968. Cuticle structure and water relations of the needles
of Pinus radiata (D. Don). New Phytol. 67, 31–38.ima, L.R., Pirani, J.R., 2003. O gênero Croton L. (Euphorbiaceae) na Cadeia do
Espinhac o, Minas Gerais, Brasil. Bol. Bot. Univ. São Paulo 21, 299–344.ima, L.R., Pirani, J.R., 2008. Revisão taxonômica de Croton sect. Lamprocroton
(Müll.Arg.) Pax (Euphorbiaceae s.s.). Biota Neotrop. 8, 21–75.imm, E.B., Dawson, T.E., 2010. Polystichum munitum (Dryopteridaceae) varies
geographically in its capacity to absorb fog water by foliar uptake within theredwood forest ecosystem. Am. J. Bot. 97, 1121–1128.
imm, E.B., Simonin, K.A., Bothman, A.G., Dawson, T.E., 2009. Foliar water uptake: acommon water acquisition strategy for plants of the redwood forest. Oecologia161, 449–459.
ouro, R.P., Ortiz, B., Santiago, L.J.M., 2003. Leaf ultrastructure of Croton compressusLam. (Euphorbiaceae). Acta Microsc. 12, 61–62.
ucena, M.F.A., Sales, M.F., 2006. Tricomas foliares em espécies de Croton L.(Crotonoideae-Euphorbiaceae). Rodriguesia 57, 11–25.
usa, M.G., Appezzato-da-Glória, B., Loeuille, B., Bártoli, G., Ciccarelli, D., 2014.
Functional groups in Lychnophorinae (Asteraceae: Vernonieae) based onmorphological and anatomical traits. Aust. J. Bot. 62, 150–163.artin, C.E., von Willert, D.J., 2000. Leaf epidermal hydathodes and theecophysiological consequences of foliar water uptake in species of Crassulafrom the Namib Desert in Southern Africa. Plant Biol. 2, 229–242.
Evolution and Systematics 22 (2016) 23–35
Mcmanus, J.F.A., 1948. Histological and histochemical uses of periodic acid. StainTechnol. 23, 99–108.
Meidner, H., 1954. Measurements of water intake from the atmosphere by leaves.New Phytol. 53, 423–426.
Metcalfe, C.R., Chalk, L., 1950. Anatomy of the Dicotyledons: Leaves Stem andWood in Relation to Taxonomy with Notes on Economic Uses, 1. ClarendonPress, Oxford.
Metcalfe, C.R., Chalk, L., 1983. Anatomy of the Dicotyledons, 2., 2nd edn. ClarendonPress, Oxford.
Milanez, C.R.D., Machado, S.R., 2008. Leaf emergences in Microlepis oleaefolia (DC.)Triana (Melastomataceae) and their probable function: an anatomical andultrastructural study. Micron 39, 884–890.
Morales, M., Garcia, Q.S., Munné-Bosch, S., 2015. Ecophysiological response toseasonal variations in water availability in the arborescent, endemic plantVellozia gigantea. Tree Physiol. 35, 253–265.
Moreira, A.S.F.P., Lemos-Filho, J.P., Zotz, G., Isaias, R.M.S., 2009. Anatomy andphotosynthetic parameters of roots and leaves of two shade-adapted orchids,Dichaea cogniauxiana Shltr. and Epidendrum secundum Jacq. Flora 204, 604–611.
Nobel, P.S., Schulte, P.J., North, G.B., 1990. Water influx characteristics andhydraulic conductivity for roots of Agave deserti Engelm. J. Exp. Bot. 41,409–415.
O’Brien, T.P., McCully, M.E., 1981. The Study of Plant Structure Principles andSelected Methods. Termarcarphi Pty Ltd., Melbourne.
Oliveira, R.S., Dawson, T.E., Burgess, S.O., 2005. Evidence for direct waterabsorption by the shoot of the desiccation-tolerant plant Vellozia flavicans inthe savannas of central Brazil. J. Trop. Ecol. 21, 585–588.
Pearse, A.G.E., 1980. Histochemistry Theoretical and Applied, 2., 4th edn. ChurchillLivingston, Edinburgh, UK.
Pita, P.B., Menezes, N.L., 2002. Anatomia da raiz de espécies de Dyckia Schult. eEncholirium Mart. ex Schult. & Schult. f. (Bromeliaceae Pitcairnioideae) da Serrado Cipó (Minas Gerais, Brasil), com especial referência ao velame. Rev. Bras.Bot. 25, 25–34.
Plymale, E.L., Wylie, R.B., 1944. The major veins of mesomorphic leaves. Am. J. Bot.31, 99–106.
Porembski, S., Barthlott, W., 1995. On the occurrence of a velamen radicum inCyperaceae and Velloziaceae. Nord. J. Bot. 15, 625–629.
Pridgeon, A.M., 1981. Absorbing trichomes in the Pleurothallidinae. Am. J. Bot. 68,64–71 (Orchidaceae).
Rao, T.A., Das, S., 1979. Leaf sclereids – occurrence and distribution in theAngiosperms. Bot Not 132, 319–324.
Rao, T.A., 1991. Compendium of Foliar Sclereids in Angiosperms: Morphology andTaxonomy. Wiley, New Delhi.
Riederer, M., 2006. Thermodynamics of the water permeability of plant cuticles:characterization of the polar pathway. J. Exp. Bot. 57, 2937–2942.
Riina, R., van Ee, B.W., Wiedenhoeft, A.C., Cardozo, A., Berry, P.E., 2010. Sectionalrearrangement of arborescent clades of Croton (Euphorbiaceae) in SouthAmerica: evolution of arillate seeds and a new species, Croton domatifer. Taxon59, 1147–1160.
Rudall, P., 1994. Laticifers in Crotonoideae (Euphorbiaceae): homology andevolution. Ann. Missouri Bot. Gard. 81, 270–282.
Rundel, P.W., 1982. Water uptake by organs other than roots. In: Lange, O.L., Nobel,P.S., Osmond, C.B., Ziegler, H. (Eds.), Physiological Plant Ecology II: WaterRelations and Carbon Assimilation. Springer-Verlag, Berlin, Heidelberg NewYork, pp. 111–134.
Sá-Haiad, B., Serpa-Ribeiro, A.C.C., Barbosa, N.C., et al., 2009. Leaf structure ofspecies from three closely related genera from tribe Crotoneae Dumort.(Euphorbiaceae s.s., Malpighiales). Plant Syst. Evol. 283, 179–202.
Sá-Haiad, B., 1987. Taxonomia e morfologia das espécies do gênero Croton L. sec ãoCroton, subsec ão Lasiogyne (Klotzsch) Müll. Arg., série Gonocladium (Baill.)Müll. Arg. ocorrentes em restingas do estado do Rio de Janeiro. MSc Thesis.Universidade Federal do Rio de Janeiro, Brazil.
Safford, H.D., 1999. Brazilian Páramos II: Macro- and mesoclimate of the camposde altitude and affinities with high mountain climates of the tropical Andesand Costa Rica. J. Biogeogr. 26, 713–737.
Safford, H.D., 2007. Brazilian Páramos IV: Phytogeography of the campos dealtitude. J. Biogeogr. 17, 1–22.
Silva, I.V., Meira, R.M.S.A., Azevedo, A.A., 2010. Anatomia de raízes de espécies deOrchidaceae do Parque Estadual da Serra do Brigadeiro, Minas Gerais. Hoehnia37, 147–161.
Silva, B.O., 2006. Análise estrutural e histoquímica de Croton macrobothrys Baillon eCroton floribundus Spreng (Euphorbiaceae). MSc Thesis. Universidade Federaldo Rio de Janeiro, Brazil.
Silveira, F.A.O., Negreiros, D., Barbosa, N.P.U., Buisson, E., Carmo, F.F., Cartensen,D.W., et al., 2015. Ecology and evolution of plant diversity in the endangeredcampo rupestre: a neglected conservation priority. Plant Soil, http://dx.doi.org/10.1007/s11104-015-2637-8.
Simonin, K.A., Santiago, L.S., Dawson, T.E., 2009. Fog interception by Sequoiasempervirens (D. Don) crowns decouples physiology from soil water deficit.Plant Cell Environ. 32, 882–892.
Smith, F.H., Smith, E.C., 1942. Anatomy of the inferior ovary of Darbya. Am. J. Bot.29, 464–471.
Soares, D.A., 2013. Morfoanatomia foliar de espécies de Croton sect. Luntia(Euphorbiaceae) como contribuic ão para a taxonomia do gênero. MSc Thesis.Universidade Federal de Vic osa, Brazil.
Solereder, H., 1908. Systematic Anatomy of the Dicotyledons, 2. Clarendon Press,Oxford.

logy, E
S
S
T
T
V
V
von Mohl, H., 1871. In: von Mohl, H., Bary, A. (Eds.), Morphologische BetrachtungDer Blätter Von Sciadopitys. Verlag von Arthur Felix, Botanische zeitung,Leipzig, pp. 17–32.
N.C. Vitarelli et al. / Perspectives in Plant Eco
tarling, M.F.V., 1993. Aspectos taxonômicos do Julocroton triqueter (Baill.) Müll.Arg. (Euphorbiaceae) do Campus da UFES, Vitória/ES, BIOS, Cadernos do.Departamento de Ciências Biológicas da PUC-MG 1, pp. 7–13.
tuart, T.S., 1968. Revival of respiration and photosynthesis in dried leaves ofPolypodium polypodioides. Planta 83, 185–206.
omlinson, P.B., 1969. Anatomy of the Monocothyledons: III.Commelinales-Zingiberales. Oxford University Press, London.
ucker, S.C., 1964. The terminal idioblasts in Magnoliaceous leaves. Am. J. Bot. 51,1051–1062.
itarelli, N.C., Riina, R., Caruzo, M.B.R., Cordeiro, I., Fuertes-Aguilar, J., Meira,
R.M.S.A., 2015. Foliar secretory structures in Crotoneae (Euphorbiaceae):diversity, anatomy and evolutionary significance. Am. J. Bot. 102, 1–15.itarelli, N.C., 2013. Anatomia foliar como subsídio para a sistemática de Croton L.(secões Cleodora e Lamprocroton), Euphorbiaceae. Doctoral Thesis.Universidade Federal de Vic osa, Brazil.
volution and Systematics 22 (2016) 23–35 35
Wagner, G.J., 1991. Secreting glandular trichomes: more than just hairs. PlantPhysiol. 96, 675–679.
Webster, G.L., 1993. A provisional synopsis of the sections of the genus Croton(Euphorbiaceae). Taxon 42, 793–823.
van Ee, B.W., Berry, P.E., 2011. Croton section Pedicellati (Euphorbiaceae), a novelnew world group, and a new subsectional classification of Croton section lLamprocroton. Syst. Bot. 36, 88–98.
Webster, G.L., Del-Arco-Aguilar, M.J., Smith, B.A., 1996. Systematic distribution offoliar trichome types in Croton. Bot. J. Linn. Soc. 121, 41–57.





























