Embed Size (px)
Citation preview

1 23
Plant Systematics andEvolution ISSN 0378-2697Volume 292Combined 1-2 Plant Syst Evol (2010) 292:1-14DOI 10.1007/s00606-010-0388-9
Morphology, development and homologiesof the perianth and floral nectaries inCroton and Astraea (Euphorbiaceae-Malpighiales)

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

ORIGINAL ARTICLE
Morphology, development and homologies of the perianthand floral nectaries in Croton and Astraea(Euphorbiaceae-Malpighiales)
Orlando Cavalari De-Paula • Maria das Gracas Sajo •
Gerhard Prenner • Ines Cordeiro • Paula J. Rudall
Received: 6 June 2010 / Accepted: 31 October 2010 / Published online: 26 November 2010
� Springer-Verlag 2010
Abstract New observations are presented on the ontogeny,
vasculature and morphology of both staminate and pistillate
flowers of Croton and Astraea. These data support earlier
hypotheses that the filamentous structures in pistillate flowers
represent reduced and transformed petals. Staminate flowers
of both genera possess five free nectaries, which are vascu-
larised by divergences of the sepal traces in Croton and
unvascularised in Astraea. In pistillate flowers, there are five
separate non-vascularised nectaries in Astraea, but in Croton
there is a single nectariferous disk that is vascularised by
divergences of the sepal traces. The nectaries are initiated late
in floral development, but their location indicates that they
could represent the outer stamen whorl transformed into
secretory staminodes. Other glandular structures occur in
pistillate flowers of most Croton species, resulting in flowers
with two secretory organ whorls. In these cases, the inner
whorl is formed by modified staminodes. Our observations
support the recent segregation of Astraea species from the
larger genus Croton. Despite strong similarities between the
two genera, there are clear structural differences, including
the presence of colleters in Astraea (absent in Croton),
moniliform trichomes on petals (rather than simple trichomes
in Croton), non-vascularised nectaries (vascularised in Cro-
ton) and reduced, non-secretory filamentous structures (well
developed and secretory in Croton).
Keywords Crotoneae � Euphorbiaceae � Flowers �Inaperturate crotonoids � Nectary evolution �Obdiplostemony � Petals � Staminodes
Introduction
As currently circumscribed, the family Euphorbiaceae
sensu stricto (order Malpighiales) is subdivided into four
subfamilies based on morphological and molecular studies:
Acalyphoideae, Cheilosoideae, Crotonoideae and Euphor-
bioideae (Wurdack et al. 2005; Tokuoka and Tobe 2006;
Tokuoka 2007; Davis et al. 2007; Wurdack and Davis
2009). The subfamily Crotonoideae is polyphyletic, and
some of its putative synapomorphies (such as crotonoid
pollen) are unsupported (Wurdack et al. 2005). Crotonoi-
deae includes 12 tribes and 74 genera from tropical and
temperate regions, distributed in 4 informal groups: Ade-
noclineae s.l., Gelonieae, the articulated crotonoids and the
inaperturate crotonoids (Webster 1994; Radcliffe-Smith
2001; Wurdack et al. 2005; Tokuoka and Tobe 2006;
Tokuoka 2007).
Croton and Astraea belong to the inaperturate croto-
noids, characterised by inaperturate pollen, ovules with
vascularised inner integument and petals present in at least
one of the floral types (i.e. pistillate or staminate flowers)
(Wurdack et al. 2005). Species of the large genus Croton
(ca. 1,200 tropical and subtropical species: Webster 1993;
Govaerts et al. 2000) are easily recognised by their star-
shaped or scaly trichomes, long or condensed monoclinous
inflorescences, frequent petiolar glands, and senescent
leaves that turn orange before abscising (Berry et al. 2005).
Croton was considered monophyletic with the exclusion of
section Astraea (elevated to the category of genus) and the
inclusion of Crotonopsis, Cubacroton, Eremocarpus,
O. C. De-Paula (&) � M. das Gracas Sajo
Universidade Estadual Paulista (UNESP), Rio Claro,
Sao Paulo 13506-900, Brazil
e-mail: [email protected]
G. Prenner � P. J. Rudall
Royal Botanic Gardens, Kew,
Richmond, Surrey TW9 3DS, UK
I. Cordeiro
Instituto de Botanica, Sao Paulo 04301-012, Brazil
123
Plant Syst Evol (2011) 292:1–14
DOI 10.1007/s00606-010-0388-9
Author's personal copy

Julocroton and Moacroton, which were previously
assigned to a tribe Crotoneae (Berry et al. 2005; van Ee
et al. 2008; Riina et al. 2009). The American genus Astraea
(10 species) is sister to Croton and shares many morpho-
logical features (Berry et al. 2005). Astraea is characterised
by the presence of deeply lobate leaves, glabrous floral
receptacle, staminate flowers with imbricate perianth mem-
bers, petals densely pilose at the base, stamen filaments
narrow and reddish, and stigma densely ramified. Its seeds
are quadrangular, rugose and carunculate (Miller and Web-
ster 1966; Berry et al. 2005; Caruzo and Cordeiro 2007).
Flowers of both Croton and Astraea are unisexual and
grouped in terminal inflorescences, usually with the pistil-
late flowers at the base and staminate flowers at the apex
(Caruzo and Cordeiro 2007). Male flowers possess a com-
plete perianth with calyx and corolla (Baillon 1858; Nair and
Abraham 1962; Venkata-Rao and Ramalakshmi 1968),
whereas in pistillate flowers only filamentous structures are
found in the positions of petals. These filamentous structures
have been interpreted as reduced petals, even though they
are non-vascularised (Baillon 1858; Nair and Abraham
1962; Venkata-Rao and Ramalakshmi 1968; Webster 1993;
Radcliffe-Smith 2001; Caruzo and Cordeiro 2007). Flowers
of both Croton and Astraea possess five-lobed nectaries
located opposite the sepals that are in staminate flowers and
range from entire to variably lobed in pistillate flowers
(Caruzo and Cordeiro 2007). Although the precise homol-
ogies of these nectaries are unknown, they have been
interpreted either as staminodes (Michaelis 1924) or nectary
disks (Caruzo and Cordeiro 2007). However, since nectary
disks develop from the floral receptacle (Weberling 1989)
and ontogenetic studies of Croton flowers are not yet
available, this interpretation remains to be tested.
In this paper, we investigate the morphology and floral
development of the pistillate and staminate flowers of
Croton and Astraea in order to clarify the homologies of
the different floral parts. Floral homologies among other
Euphorbiaceae are notoriously controversial (Prenner and
Rudall 2007; Prenner et al. 2008). Some Euphorbiaceae
possess perfect flowers (e.g. Jatropha), whereas in the
cyathium of Euphorbia and its allies, each flower is widely
interpreted as a single stalked stamen or a single naked
stalked pistil. Few previous studies have focused on the
flowers of non-cyathial euphorbiaceous taxa, which possess
less specialised inflorescences (Nair and Abraham 1962;
Venkata-Rao and Ramalakshmi 1968).
Materials and methods
Plants were collected in their natural environment, and
specimens were prepared and incorporated into the
Rioclarense Herbarium (HRCB) and the Herbario do
Estado ‘‘Maria Eneyda P. Kaufmann Fidalgo’’ (SP), as
follows: Astraea lobata (L.) Klotzsch, De-Paula 10
(HRCB)—Botucatu, Sao Paulo, Brazil; A. praetervisa
(Mull. Arg.) P.E. Berry, Cordeiro 3016 (SP)—Salvador,
Bahia, Brazil; Croton floribundus Spreng., De-Paula 25
(HRCB)—Botucatu, Sao Paulo, Brazil; C. fuscescens
Spreng., De-Paula 29 (HRCB)—Sao Paulo, Sao Paulo,
Brazil; C. glandulosus Mull. Arg., De-Paula 31
(HRCB)—Botucatu, Sao Paulo, Brazil; C. lundianus
(Didr.) Mull. Arg., De-Paula 26 (HRCB)—Jundiaı, Sao
Paulo, Brazil; C. piptocalyx Mull. Arg., De-Paula 28
(HRCB)—Campinas, Sao Paulo, Brazil; C. urucurana
Baill., De-Paula 30 (HRCB)—Botucatu, Sao Paulo,
Brazil; C. triqueter Lam., 3016 (SP)—Feira de Santana,
Bahia, Brazil. The specimens to be microscopically
examined were fixed in FAA 50 (Johansen 1940) and
subsequently stored in 50% ethanol.
For anatomical studies, entire flowers were dehydrated
through an ethanol series and embedded in Leica methac-
rylate. The material was sectioned using a rotary microtome
at 8–10 lm thickness. Sections were stained with 0.05%
toluidine blue at pH 4.7 (O’Brien et al. 1964) and mounted
with Entellan. Relevant features were recorded using a DFC
290 digital camera attached to a Leica DMLB microscope.
For scanning electron microscopy (SEM), flowers of
different developmental stages were carefully dissected,
dehydrated through an alcohol series and processed in an
Autosamdri-815B CPD CO2 critical point drier. The sam-
ples were mounted on aluminium stubs, coated with plat-
inum using an Emitech K550 sputter coater and examined
using a cold-field emission Hitachi SEM S-4700-II at the
Jodrell Laboratory, Royal Botanic Gardens, Kew.
Photographs were processed using Adobe Photoshop
software (Redwood City, CA). Illustrations were prepared
from digital photographs using Adobe Illustrator software
(Redwood City, CA).
Results
Ontogeny (Figs. 1, 2)
Flower primordia of both Astraea and Croton are sub-
tended by a single abaxial bract. Flowers are preceded by
two lateral bracteoles, which arise in a rapid sequence
(Fig. 1a–c). The sepal primordia develop in a spiral
sequence (Fig. 1c), except in C. fuscescens and C. triquet-
er, which have zygomorphic corollas and unidirectional
sepal initiation (Fig. 1d, e). Alternating with the sepals of
pistillate flowers, five primordia give rise to filamentous
structures (Fig. 1f–i). In C. triqueter, only one primor-
dium develops in an adaxial position (Fig. 1k, l), and in
C. fuscescens no such structure is formed (Fig. 1j).
2 O. C. De-Paula et al.
123
Author's personal copy
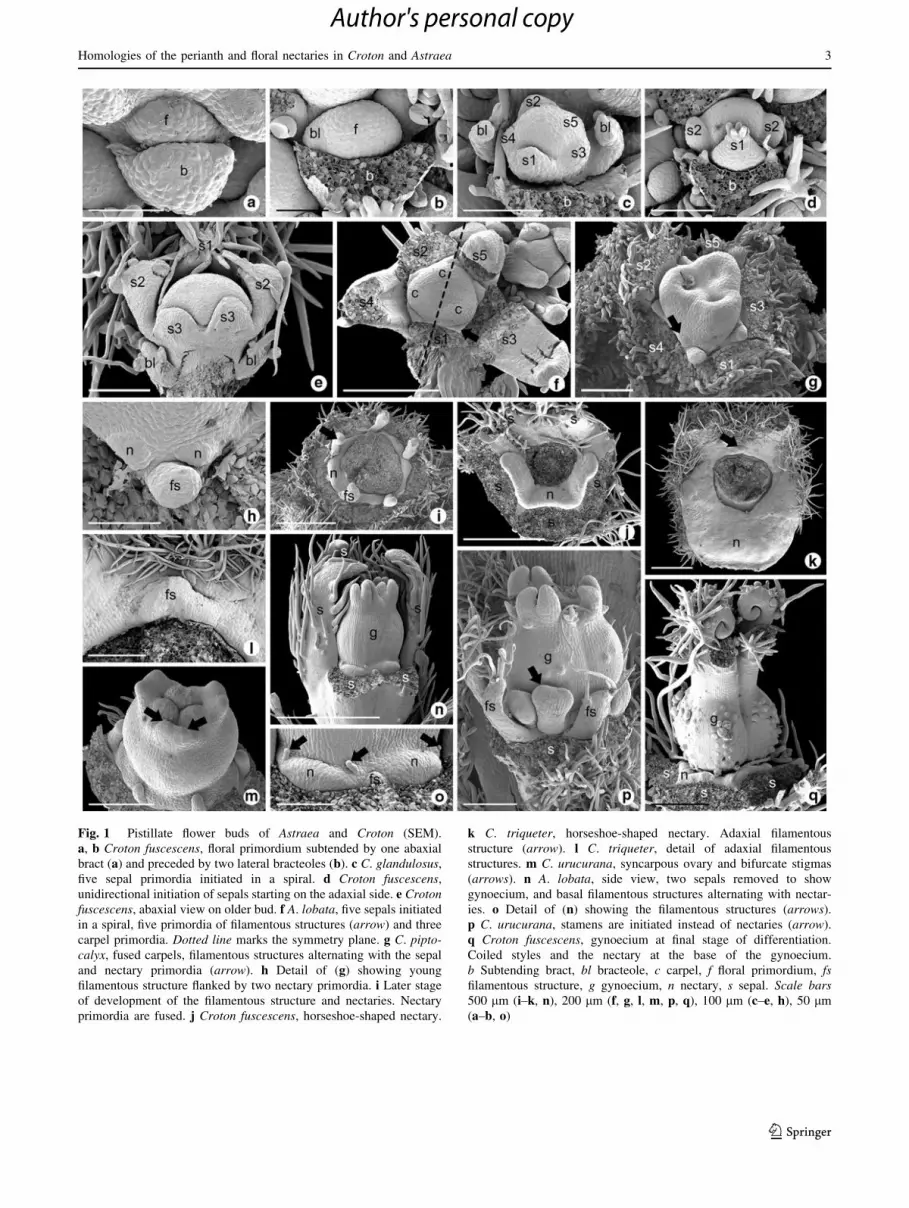
Fig. 1 Pistillate flower buds of Astraea and Croton (SEM).
a, b Croton fuscescens, floral primordium subtended by one abaxial
bract (a) and preceded by two lateral bracteoles (b). c C. glandulosus,
five sepal primordia initiated in a spiral. d Croton fuscescens,
unidirectional initiation of sepals starting on the adaxial side. e Crotonfuscescens, abaxial view on older bud. f A. lobata, five sepals initiated
in a spiral, five primordia of filamentous structures (arrow) and three
carpel primordia. Dotted line marks the symmetry plane. g C. pipto-calyx, fused carpels, filamentous structures alternating with the sepal
and nectary primordia (arrow). h Detail of (g) showing young
filamentous structure flanked by two nectary primordia. i Later stage
of development of the filamentous structure and nectaries. Nectary
primordia are fused. j Croton fuscescens, horseshoe-shaped nectary.
k C. triqueter, horseshoe-shaped nectary. Adaxial filamentous
structure (arrow). l C. triqueter, detail of adaxial filamentous
structures. m C. urucurana, syncarpous ovary and bifurcate stigmas
(arrows). n A. lobata, side view, two sepals removed to show
gynoecium, and basal filamentous structures alternating with nectar-
ies. o Detail of (n) showing the filamentous structures (arrows).
p C. urucurana, stamens are initiated instead of nectaries (arrow).
q Croton fuscescens, gynoecium at final stage of differentiation.
Coiled styles and the nectary at the base of the gynoecium.
b Subtending bract, bl bracteole, c carpel, f floral primordium, fsfilamentous structure, g gynoecium, n nectary, s sepal. Scale bars500 lm (i–k, n), 200 lm (f, g, l, m, p, q), 100 lm (c–e, h), 50 lm
(a–b, o)
Homologies of the perianth and floral nectaries in Croton and Astraea 3
123
Author's personal copy
Alternating with the filamentous structures and opposite
the sepals, five pairs of primordia give rise to five nectaries
(Fig. 1g–i) that fuse at later stages to form a disk. In
C. fuscescens and C. triqueter, the nectary forms an arc
(Fig. 1j, k). In the pistillate flowers of Astraea, in addition
to the filamentous structures that alternate with the sepals,
there are other filamentous structures alternating with the
nectaries (Fig. 1n, o). In some pistillate flowers, the pri-
mordia that would normally be expected to give rise to
nectaries actually form stamens (Fig. 1p). The gynoecium
arises from three carpel primordia (Fig. 1f) that fuse into a
syncarpous ovary (Fig. 1g, m, p). The stigma is bifid
(Fig. 1m, n, p, q).
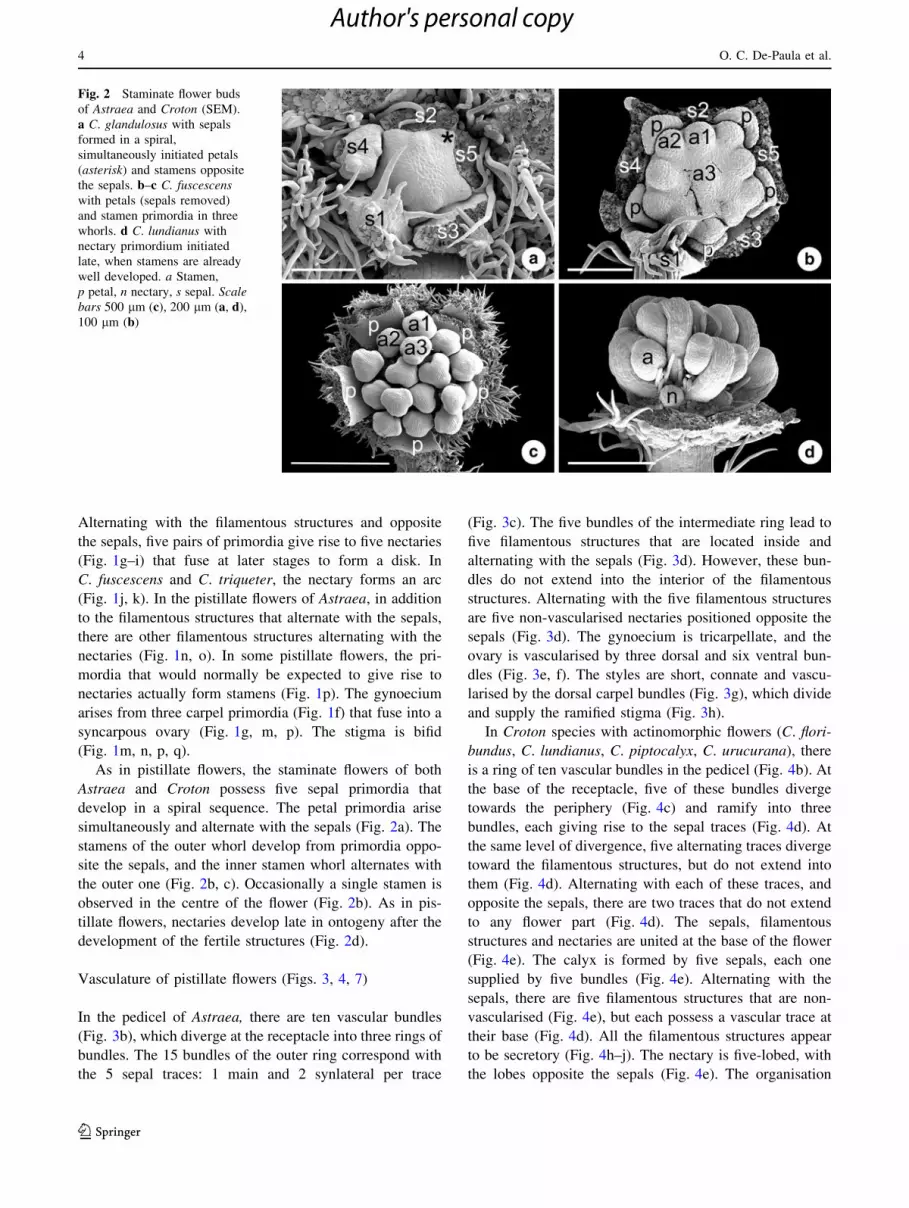
As in pistillate flowers, the staminate flowers of both
Astraea and Croton possess five sepal primordia that
develop in a spiral sequence. The petal primordia arise
simultaneously and alternate with the sepals (Fig. 2a). The
stamens of the outer whorl develop from primordia oppo-
site the sepals, and the inner stamen whorl alternates with
the outer one (Fig. 2b, c). Occasionally a single stamen is
observed in the centre of the flower (Fig. 2b). As in pis-
tillate flowers, nectaries develop late in ontogeny after the
development of the fertile structures (Fig. 2d).
Vasculature of pistillate flowers (Figs. 3, 4, 7)
In the pedicel of Astraea, there are ten vascular bundles
(Fig. 3b), which diverge at the receptacle into three rings of
bundles. The 15 bundles of the outer ring correspond with
the 5 sepal traces: 1 main and 2 synlateral per trace
(Fig. 3c). The five bundles of the intermediate ring lead to
five filamentous structures that are located inside and
alternating with the sepals (Fig. 3d). However, these bun-
dles do not extend into the interior of the filamentous
structures. Alternating with the five filamentous structures
are five non-vascularised nectaries positioned opposite the
sepals (Fig. 3d). The gynoecium is tricarpellate, and the
ovary is vascularised by three dorsal and six ventral bun-
dles (Fig. 3e, f). The styles are short, connate and vascu-
larised by the dorsal carpel bundles (Fig. 3g), which divide
and supply the ramified stigma (Fig. 3h).
In Croton species with actinomorphic flowers (C. flori-
bundus, C. lundianus, C. piptocalyx, C. urucurana), there
is a ring of ten vascular bundles in the pedicel (Fig. 4b). At
the base of the receptacle, five of these bundles diverge
towards the periphery (Fig. 4c) and ramify into three
bundles, each giving rise to the sepal traces (Fig. 4d). At
the same level of divergence, five alternating traces diverge
toward the filamentous structures, but do not extend into
them (Fig. 4d). Alternating with each of these traces, and
opposite the sepals, there are two traces that do not extend
to any flower part (Fig. 4d). The sepals, filamentous
structures and nectaries are united at the base of the flower
(Fig. 4e). The calyx is formed by five sepals, each one
supplied by five bundles (Fig. 4e). Alternating with the
sepals, there are five filamentous structures that are non-
vascularised (Fig. 4e), but each possess a vascular trace at
their base (Fig. 4d). All the filamentous structures appear
to be secretory (Fig. 4h–j). The nectary is five-lobed, with
the lobes opposite the sepals (Fig. 4e). The organisation
Fig. 2 Staminate flower buds
of Astraea and Croton (SEM).
a C. glandulosus with sepals
formed in a spiral,
simultaneously initiated petals
(asterisk) and stamens opposite
the sepals. b–c C. fuscescenswith petals (sepals removed)
and stamen primordia in three
whorls. d C. lundianus with
nectary primordium initiated
late, when stamens are already
well developed. a Stamen,
p petal, n nectary, s sepal. Scalebars 500 lm (c), 200 lm (a, d),
100 lm (b)
4 O. C. De-Paula et al.
123
Author's personal copy

Fig. 3 Vasculature of pistillate flower of Astraea lobata. a Flower in
longitudinal section indicating regions where transverse sections were
made. b Pedicel with ten vascular bundles. c Receptacle. Divergence
of sepal, petal and stamen traces. d Base of sepals with filamentous
structures (arrow) and nectaries (grey). e Median region of ovary.
f Uppermost part of ovary where ovule traces originate. g Style.
h Stigma. arrow Filamentous structures, at stamen traces, db dorsal
bundles of the carpel, dt dorsal traces of the carpel, ft traces of the
filamentous structures, n nectary, ov ovule, ot ovule trace, s sepal, sgstigma, sgb stigma bundle, sgt stigma trace, sl style, st sepal trace, vbventral bundles of the carpel, vt ventral carpel traces. Scale bars200 lm
Homologies of the perianth and floral nectaries in Croton and Astraea 5
123
Author's personal copy

Fig. 4 Vasculature of pistillate flower of Croton. a–e Crotonfloribundus. a Flower in longitudinal section indicating regions where
transverse sections were made. b Pedicel with ten vascular bundles.
c Receptacle. Divergence of sepal traces. d Receptacle at base of
nectaries (grey). Divergence of petal and stamen traces. e Base of
sepals with filamentous structures (arrows) and nectaries (grey).
Divergence of the dorsal carpel traces. f–i Croton glandulosus.
f Flower in longitudinal section, indicating the regions where the
transverse sections were made. g Base of receptacle. Divergence of
filament traces and dorsal and ventral carpel traces. h Basal region of
flower. i Median region of flower. The nectaries are fused to the sepals
and are vascularised filamentous structures (arrow). j–m Croton
triqueter. j Flower in longitudinal section, indicating regions where
transverse sections were made. k Base of receptacle. The nectariferous
tissues (grey) opposite the groups of sepal traces. l Base of sepals with
stamen traces in abaxial region, carpellary bundles in central region
and a single non-glandular filamentous structure in the adaxial region
(arrow). m Basal regions of the sepals and gynoecium, and median
region of the nectary (grey). Floral zygomorphy. arrow Filamentous
structures, at stamen trace, db dorsal bundle, dt dorsal trace, fb bundle
of the filamentous structures, ft trace of the filamentous structures,
g gynoecium, lo locule, n nectary, ov ovule, s sepal, sb sepal bundle, sgstigma, sl style, st sepal trace, vb ventral bundle, vt ventral carpel trace.
Scale bars 400 lm (a–e, g–h, j–m), 200 lm (f, i)
6 O. C. De-Paula et al.
123
Author's personal copy

and vasculature of the gynoecium resembles that of
Astraea.
In C. glandulosus, there are ten vascular bundles in the
pedicel, as in Astraea. At the base of the receptacle, five of
these bundles diverge to the sepals (Fig. 4g, h), and five
alternating traces diverge to supply the five filamentous
structures (Fig. 4g–i). The calyx, the filamentous struc-
tures, the nectaries and the gynoecium are fused at the
flower base (Fig. 4h). The calyx is formed by five sepals,
each with a single vascular bundle (Fig. 4i). The filamen-
tous structures alternate with the sepals; they are vascu-
larised (Fig. 4h, i) and apparently secretory (Fig. 4h–j).
The nectaries are fused to the sepals and are vascularised
by branches of the sepal traces (Fig. 4h, i). The tricarpel-
late ovary is vascularised by three dorsal and three ventral
vascular bundles (Fig. 4h, i). The style is short and vas-
cularised by dorsal carpellary bundles, which ramify and
supply the stigmas (Fig. 4f).
The filamentous structures of C. floribundus, C. lundi-
anus, C. piptocalyx, C. urucurana and C. glandulosus
appear secretory (Fig. 4h–j), so in these species the pistil-
late flowers possess two whorls of secretory structures.
In C. fuscescens and C. triqueter with zygomorphic
flowers, there are ten vascular bundles in the pedicel. At the
base of the receptacle, five groups of traces diverge to
supply each of the five sepals (Fig. 4k). In the same region,
four areas of nectariferous tissue alternate with the sepal
traces (Fig. 4k). The zygomorphic calyx is formed by two
free adaxial sepals and three larger abaxial ones that are
connate at their bases (Fig. 4l). Each of the larger sepals
possesses 5–7 vascular bundles and each of the smaller
sepals only one (Fig. 4m). In C. triqueter, a single non-
vascularised filamentous structure alternates with the
smaller sepals (Fig. 4l). In C. fuscescens there are no fila-
mentous structures. In these two species, the nectary is
abaxial and semicircular, and supplied by branches of the
sepal traces (Fig. 4l). Gynoecium vasculature resembles
that of C. glandulosus, though the ovary is zygomorphic in
both C. triqueter and C. fuscescens (Fig. 7i).
Vasculature of staminate flowers (Fig. 5)
The staminate flowers of Astraea are actinomorphic and
possess a horseshoe-shaped vascular bundle in the pedicel
(Fig. 5b), which divides at the base of the receptacle into
ten bundles: the five external bundles extend to the sepals,
and the five alternating inner bundles extend to the petals
(Fig. 5c–f). The calyx consists of five sepals united at their
bases, each with a single vascular bundle (Fig. 5e). The
corolla consists of five free petals alternating with the
sepals (Fig. 5e, f). Alternating with the petals there are five
free nectaries, adnate to the androphore (Fig. 5d, e). In the
centre of the flower there is an androphore with three
alternating whorls of five stamens (Fig. 5f). The outer-
whorl stamens are located opposite the petals. The stamens
are curved inwards in the flower bud, so that the filaments
appear external in transverse sections (Fig. 5f). Each anther
is supplied by a single bundle that extends to one-third of
its length (Fig. 5a).
The staminate flowers of Croton show the same pat-
tern of vasculature as those of Astraea, though the
pedicel in Croton possesses a ring of bundles and not an
arc (Fig. 5h). The nectaries are vascularised (Fig. 5k), in
contrast to Astraea, which has non-vascularised nectaries
(Fig. 5e).
Floral morphology (Figs. 6, 7)
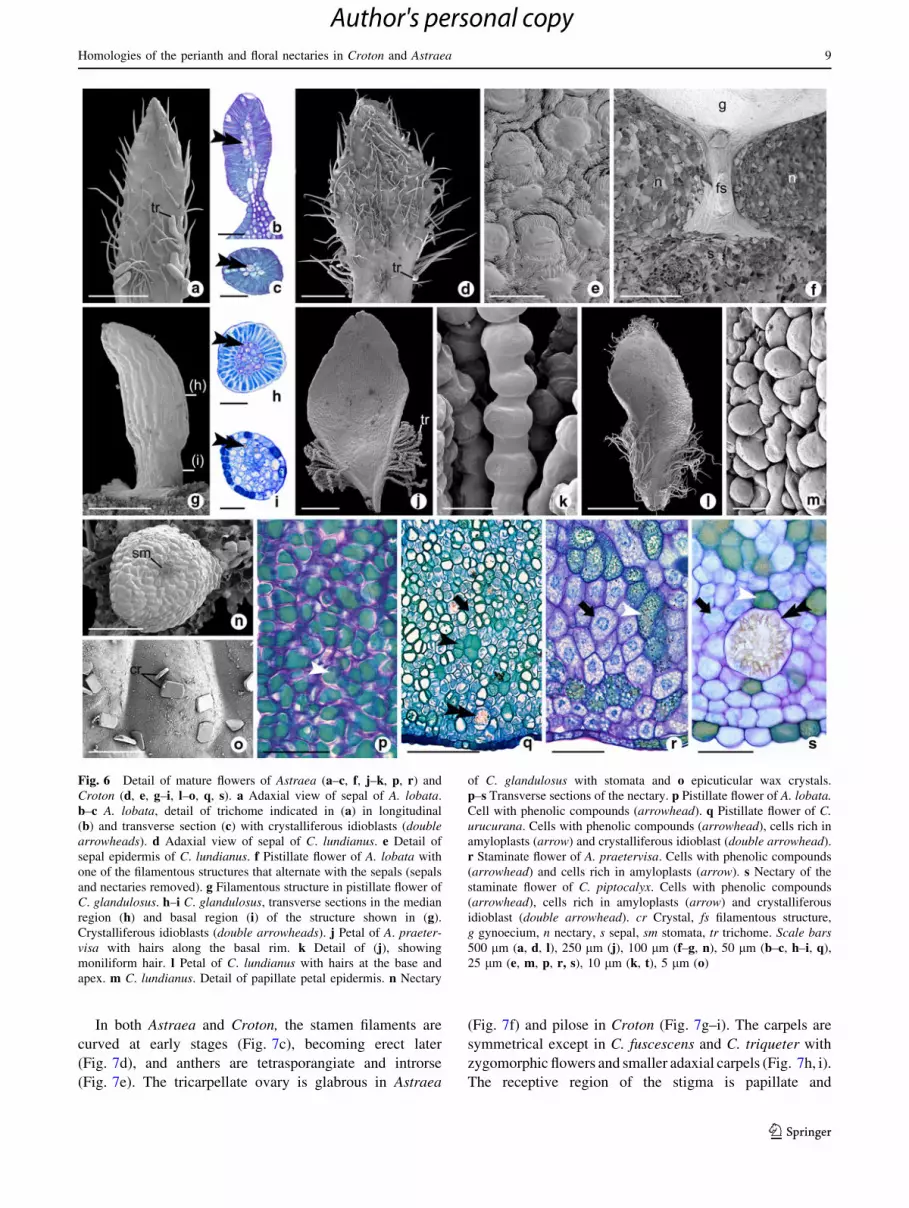
On the sepals of Astraea and Croton, there are both
secretory and non-secretory trichomes (Fig. 6a–d). Non-
secretory trichomes occur over the entire sepal surface
(Fig. 6a, d), whereas secretory trichomes are restricted to
the sepal margins (Fig. 6a–d). Secretory trichomes are
elongated in Astraea (Fig. 6a) and spherical in Croton
(Fig. 6d). They are multicellular, covered by elongated
epidermal cells and contain crystalliferous idioblasts in
Astraea (Fig. 6b, c) and resemble colleters. In C. lundi-
anus, the sepals are covered by an ornamented epidermis
(Fig. 6e).
In staminate flowers of Astraea and Croton, the petals
have only non-secretory trichomes (Fig. 6j, l). In Astraea,
the trichomes are moniliform and occur at the petal base
(Fig. 6j, k); in Croton they are simple and found at both the
base and apex of the petals (Fig. 6l). In C. lundianus the
entire petal surface is papillate (Fig. 6m).
In the pistillate flowers of Astraea, the filamentous
structures are reduced and alternate with the sepals (com-
pare Fig. 6g). The filamentous structures of Croton are
larger than the Astraea trichomes and supplied by a single
vascular bundle (Fig. 6i).
The nectaries, here interpreted as secretory staminodes,
possess a uniseriate epidermis (Fig. 6q–s) with occasional
stomata (Fig. 6n); the epidermis is covered by epicuticular
wax in the shape of bars (Fig. 6o). Internally, Astraea
nectaries consist of thick-walled parenchymatous cells of
dense cytoplasm in pistillate flowers (Fig. 6p) and cells
with amyloplasts in staminate flowers (Fig. 6r). In Croton,
the nectaries of both pistillate (Fig. 6q) and staminate
(Fig. 6s) flowers are formed by three cell types: cells rich
in amyloplasts, cells containing phenolic compounds and
idioblasts containing druses.
Both flower types in Astraea also contain other fila-
mentous structures (Fig. 7a–c), which alternate with the
nectaries in the pistillate flowers (Fig. 7a, b), but lack any
specific pattern of distribution in the staminate flowers
(Fig. 7c).
Homologies of the perianth and floral nectaries in Croton and Astraea 7
123
Author's personal copy

Fig. 5 Vasculature of staminate flowers of Astraea and Croton.
a–f A. praetervisa. a Flower in longitudinal section, indicating
regions where transverse sections were made. b Pedicel with
U-shaped vascular bundle. c Base of receptacle from which the sepal
traces diverge. d Basal region of flower. Divergence of the petal
bundles. e Median region of nectaries and basal region of androphore.
f Median region of flower. g–l Croton piptocalyx. g Flower in
longitudinal section, indicating regions where transverse sections
were made. h Pedicel with eight vascular traces. i Base of receptacle.
j Median region of receptacle. k Detail of base of nectary. l Median
region of nectaries and basal region of androphore. Divergence of the
stamen traces, with the outer whorl opposite the petals. a Stamen, afandrophore, at1 stamen traces of the external whorl, at2 stamen traces
of the second whorl, at3 stamen traces of the internal whorl, n nectary,
nb nectary bundle, p petal, pb petal bundle, pt petal trace, s sepal, sbsepal bundle, st sepal trace. Scale bars 400 lm (g–l), 200 lm (a–f)
8 O. C. De-Paula et al.
123
Author's personal copy
In both Astraea and Croton, the stamen filaments are
curved at early stages (Fig. 7c), becoming erect later
(Fig. 7d), and anthers are tetrasporangiate and introrse
(Fig. 7e). The tricarpellate ovary is glabrous in Astraea
(Fig. 7f) and pilose in Croton (Fig. 7g–i). The carpels are
symmetrical except in C. fuscescens and C. triqueter with
zygomorphic flowers and smaller adaxial carpels (Fig. 7h, i).
The receptive region of the stigma is papillate and
Fig. 6 Detail of mature flowers of Astraea (a–c, f, j–k, p, r) and
Croton (d, e, g–i, l–o, q, s). a Adaxial view of sepal of A. lobata.
b–c A. lobata, detail of trichome indicated in (a) in longitudinal
(b) and transverse section (c) with crystalliferous idioblasts (doublearrowheads). d Adaxial view of sepal of C. lundianus. e Detail of
sepal epidermis of C. lundianus. f Pistillate flower of A. lobata with
one of the filamentous structures that alternate with the sepals (sepals
and nectaries removed). g Filamentous structure in pistillate flower of
C. glandulosus. h–i C. glandulosus, transverse sections in the median
region (h) and basal region (i) of the structure shown in (g).
Crystalliferous idioblasts (double arrowheads). j Petal of A. praeter-visa with hairs along the basal rim. k Detail of (j), showing
moniliform hair. l Petal of C. lundianus with hairs at the base and
apex. m C. lundianus. Detail of papillate petal epidermis. n Nectary
of C. glandulosus with stomata and o epicuticular wax crystals.
p–s Transverse sections of the nectary. p Pistillate flower of A. lobata.Cell with phenolic compounds (arrowhead). q Pistillate flower of C.urucurana. Cells with phenolic compounds (arrowhead), cells rich in
amyloplasts (arrow) and crystalliferous idioblast (double arrowhead).
r Staminate flower of A. praetervisa. Cells with phenolic compounds
(arrowhead) and cells rich in amyloplasts (arrow). s Nectary of the
staminate flower of C. piptocalyx. Cells with phenolic compounds
(arrowhead), cells rich in amyloplasts (arrow) and crystalliferous
idioblast (double arrowhead). cr Crystal, fs filamentous structure,
g gynoecium, n nectary, s sepal, sm stomata, tr trichome. Scale bars500 lm (a, d, l), 250 lm (j), 100 lm (f–g, n), 50 lm (b–c, h–i, q),
25 lm (e, m, p, r, s), 10 lm (k, t), 5 lm (o)
Homologies of the perianth and floral nectaries in Croton and Astraea 9
123
Author's personal copy

Fig. 7 Details of mature flowers of Astraea (a–c, f, j–l) and Croton(d–e, g–i, m–o). A. lobata (a–c, f, j–l), C. glandulosus (d),
C. piptocalyx (e), C. urucurana (g), C. triqueter (h–i), C. lundianus(m–o). a Pistillate flower of A. lobata with two sepals and nectaries
removed. Filamentous structures (arrow) alternating with the nectar-
ies. b Detail of (a) showing filamentous structures. c–d A. lobata,
aspect of young staminate flower (c) and a single mature stamen (d).
Stamen with filamentous structures at the base. e Transverse section
of anther of C. piptocalyx. f–o Pistillate flower. f A. lobata: transverse
section of the ovary. Glabrous surface. g C. urucurana: transverse
section of the ovary. Hirsute surface. h–i C. triqueter: transverse
sections of the ovary in the median region (h) and basal region (i).Arrow pointing to the adaxial smaller carpel. j–l Stigma of A. lobata:
general aspect (j) and its receptive region in frontal view (k) and in
transverse section (l). m–o Stigma of C. lundianus: general aspect
(m) and its receptive region in frontal view with pollen grain and
papillate surface (arrow) (n) and in transverse section (o). g Gynoe-
cium, n nectary, ov ovule, po pollen grain, s sepal. Scale bars 500 lm
(a, c, j), 400 lm (g), 250 lm (d, m), 200 lm (b, e–f, h–i), 100 lm
(k–l), 50 lm (n), 25 lm (o)
10 O. C. De-Paula et al.
123
Author's personal copy

secretory in Croton (Fig. 7m–o), and formed by elongated
non-secretory cells in Astraea (Fig. 7j–l).
Discussion
Homologies of filamentous structures in flowers
of Croton and Astraea (Fig. 8)
Our observations of floral ontogeny and vasculature lead us
to propose new interpretations for the filamentous struc-
tures of the pistillate flowers of Croton and Astraea. Distal
to the sepals and alternating with them, five primordia give
rise to five petals in staminate flowers and to five fila-
mentous structures in pistillate flowers (Fig. 8). In both
Croton and Astraea, these filamentous structures are
unvascularised; traces of associated vascular tissue at their
bases extend into the filamentous structures only in
C. glandulosus. Based on Puri’s (1951) floral vascularisa-
tion model, in which vascular traces that alternate with
sepals are interpreted as petal traces, the filamentous
structures of the pistillate flowers of Croton and Astraea
represent reduced petals. This supports the view of Nair
and Abraham (1962) and Venkata-Rao and Ramalakshmi
(1968), who described the presence of a corolla in pistillate
flowers of Croton. The reduction of the vascular bundles
Fig. 8 Floral diagrams and floral formulae of pistillate and staminate flowers of Croton and Astraea. Floral formulae follow the style proposed
by Prenner et al. (2010)
Homologies of the perianth and floral nectaries in Croton and Astraea 11
123
Author's personal copy

observed here (filamentous structures vascularised in
C. glandulosus but possessing only basal traces in other
species) indicates a reduction of the internal perianth
whorl, which consists of five elements in some Crotonoi-
deae and is entirely absent from others. The inner perianth
whorl is represented by a single petal in C. fuscescens.
Filamentous structures similar to those of Astraea and
Croton also occur in the pistillate flowers of Hevea and
Codiaeum (Crotonoideae) and in Micrococca, Acalypha
and Mercurialis (Acalyphoideae). These structures were
interpreted as staminodes in Hevea (Nair and Abraham
1962), reduced petals in Codiaeum (Nair and Abraham
1962; Venkata-Rao and Ramalakshmi 1968) and nectaries
in Micrococca (Venkata-Rao and Ramalakshmi 1968) and
Mercurialis (Baillon 1858).
Sepals and petals are not consistently present in
Euphorbiaceae flowers. In most Crotonoideae there are two
perianth whorls, but staminate flowers of Hevea, Manihot
and Baliaspermum and pistillate flowers of Hevea and
Manihot possess only a calyx whorl (Nair and Abraham
1962; Venkata-Rao and Ramalakshmi 1968), Garcia pos-
sesses a calyx whorl and two-whorled corolla in both
flower types (Baillon 1858), and Croton and Astraea pos-
sess petals transformed into filamentous structures, as
described here. Flowers of subfamilies Cheilosoideae and
(most) Acalyphoideae possess only a calyx (Radcliffe-
Smith 2001). In Euphorbioideae, a perianth is either
entirely absent, or perianth-like structures are present, or
they possess a calyx of 1–8 sepals (in staminate flowers) or
1–6 sepals (in pistillate flowers) (Radcliffe-Smith 2001;
Prenner and Rudall 2007; Prenner et al. 2008). Wurdack
et al. (2005) hypothesised that in Euphorbiaceae the peri-
anth has an important role in floral specialisation. They
highlighted the occurrence of petals in at least one of the
floral types of inaperturate crotonoids as a synapomorphy
for this group. This is corroborated with the evidence
presented here that the filamentous structures of the pis-
tillate flowers of Croton and Astraea represent reduced and
transformed petals.
Homologies of nectaries
Our observations also allow new interpretations for the
nectaries of both pistillate and staminate flowers of Croton
and Astraea. Within the perianth, flowers of both Croton
and Astraea possess structures described either as a ‘‘nec-
tariferous disk’’, based on their morphology (Caruzo and
Cordeiro 2007), or as ‘‘extrastaminal nectaries’’, based on
their position (Webster 1993; Bernardello 2007). Our study
shows that staminate flowers of both Croton and Astraea
possess five free nectaries, which are vascularised by
divergences of the sepal traces in Croton and unvascular-
ised in Astraea. In pistillate flowers of Astraea the nectaries
are free and non-vascularised, but in Croton they are fused
into a nectariferous disk that is vascularised by divergences
of the sepal traces. The nectaries originate from five sep-
arate primordia that become evident only after the other
elements of the flower are formed (Figs. 1g, h, 2d). They
are located opposite the sepals and alternate with the petals
(in staminate flowers) or alternate with the filamentous
structures (in pistillate flowers).
Despite the late initiation of the nectaries, their posi-
tion indicates that they could represent the outer stamen
whorl transformed into secretory staminodes (Fig. 8). Our
observations that in most species the nectaries are vas-
cularised by divergences of the sepal traces and the out-
ermost stamens are located opposite the petals reinforce
this interpretation. Furthermore, occasional pistillate
flowers of C. urucurana develop stamens in positions
opposite the sepals that are normally occupied by nec-
taries (Fig. 1p).
Floral nectaries are relatively common in other Croto-
noideae, occurring in both flower types in Jatropha, Man-
ihot, Baliospermum, Codiaeum and Aleurites (Baillon 1858;
Nair and Abraham 1962; Venkata-Rao and Ramalakshmi
1968) and in the staminate flowers of Ricinocarpus (Baillon
1858). As in Croton and Astraea, the nectaries of the sta-
minate flowers are located opposite the sepals, and the outer
stamens lie in the same sector as the petals. These obser-
vations suggest that in all these genera the nectaries are
homologous with the outer stamen whorl and that secretory
staminodes are a synapomorphy for Crotonoideae.
Floral nectaries of unknown nature occur in other
Euphorbiaceae, such as Acalyphoideae (Clutia, Micrococca,
Chrozophora, Argythamnia and Ditaxis) and some
Cheilosoideae (Wurdack et al. 2005). Within Euphorbioi-
deae, the nectaries related to reproduction (nuptial nec-
taries) are apparently not of floral origin, but are strictly
extrafloral because they are associated with the cyathium,
which is normally interpreted as an inflorescence (Prenner
and Rudall 2007). Staminodial nectaries are also common
in other families of the order Malphigiales, such as Bon-
netiaceae (Dickinson and Weitzman 1998), Caryocaraceae
(Prance and Freitas da Silva 1973; Dickinson 1990),
Clusiaceae (Robson 1961; Ronse Decraene and Smets
1991), Chrysobalanaceae (Matthews and Endress 2008),
Linaceae (Brown 1938; Cronquist 1981), Passifloraceae
(Cronquist 1981; Bernhard 1999) and Violaceae (Smets
1986; Vogel 1998; Freitas and Sazima 2003). However,
staminodial nectaries are absent from Rafflesiaceae and
Peraceae, the putative sister families of Euphorbiaceae
(Davis et al. 2007; Wurdack and Davis 2009).
In addition to staminodial nectaries, nectariferous
structures occur in pistillate flowers of most Croton spe-
cies, resulting in flowers with two secretory whorls, as
Freitas et al. (2001) reported for Croton sarcopetalus. In
12 O. C. De-Paula et al.
123
Author's personal copy

these cases, the outer secretory whorl is filamentous and
formed by transformed petals, and the inner one is necta-
riferous and formed by modified staminodes. C. triqueter
possesses a single filamentous structure between the two
adaxial sepals, but this structure is lacking in C. fuscescens,
in which a continuous horseshoe-shaped nectary is formed
internally and in an abaxial position. In these cases, the
early developing nectary lobes alternate with the sepal
traces (Fig. 4k) so the nectaries are probably petal-derived
structures, though this interpretation requires further
investigation.
The function of the floral nectaries in Croton is not yet
fully understood. In C. suberosus, a supposed anemophi-
lous species (Domınguez and Bullock 1989), the nectaries
do not appear to be essential for pollination. However, in
the entomophilous C. sarcopetalus, both the outer floral
nectaries (here interpreted as petals) and the inner nectaries
(here interpreted as staminodes) secrete fructose and glu-
cose at anthesis. Furthermore, the external nectaries remain
secretory in the fruit when they secrete glucose only
(Freitas et al. 2001).
Segregation of Astraea from Croton is supported
by floral structure
Although Astraea and Croton are sister genera and could
be placed together without loss of monophyly, our results
support the recent segregation of Astraea species from the
larger genus Croton because there are considerable mor-
phological and anatomical differences between the two
genera. These differences include the presence of colleters
in Astraea (absent in Croton), moniliform trichomes on
petals in Astraea (simple in Croton) and non-vascularised
nectaries (vascularised in Croton). Additionally, the fila-
mentous structures of Astraea, which we interpret as
petals, are highly reduced and non-secretory, in contrast
to Croton with well-developed, secretory filamentous
structures.
In Astraea, besides the filamentous petals, other fila-
mentous structures are initiated after the staminodial nec-
taries (Fig. 1n, o) and alternate with them (Fig. 7a–c).
These filamentous structures could represent an additional
whorl of staminodes, but the absence of vasculature does
not fully support this hypothesis. Endress (2008) noted that
organs that have decreased in size during evolution and lost
their original function frequently become labile in number.
This could be the case in some instances in the current
study (e.g. in Astraea lobata), where the number of fila-
mentous structures does not correspond with the original
petal number.
Acknowledgments The authors thank J. Marzinek, C.J. Campos,
J. Lombardi and L.C. Bernacci for their help with field work. O.C.
De-Paula acknowledges financial support (2006/61641-0) from the
Fundacao de Amparo a Pesquisa do Estado de Sao Paulo (FAPESP)
and the Kew Latin American Research Fellowship scheme. We thank
Peter Endress and an anonymous reviewer for their critical and
helpful reviews.
References
Baillon H (1858) Etude generale du groupe des Euphorbiacees.
V. Masson, Paris
Bernardello G (2007) A systematic survey of floral nectaries. In:
Nicolson SW, Nepi M, Pacini E (eds) Nectaries and nectar.
Springer, Dordrecht, pp 19–128
Bernhard A (1999) Flower structure, development, and systematics in
Passifloraceae and in Abatia (Flacourtiaceae). Int J Pl Sci
160:135–150
Berry PE, Hipp AL, Wurdack KJ, van Ee BW, Riina R (2005)
Molecular phylogenetics of the giant genus Croton and tribe
Crotoneae (Euphorbiaceae sensu stricto) using ITS and trnL-
trnF DNA sequence data. Amer J Bot 92:1520–1534
Brown W (1938) The bearing of nectaries on the phylogeny of
flowering plants. Proc Amer Phil Soc 79:549–595
Caruzo MBR, Cordeiro I (2007) Sinopse da tribo Crotoneae Dumort.
(Euphorbiaceae s.s.) no Estado de Sao Paulo, Brasil. Hoehnea
34:571–585
Cronquist A (1981) An integrated system of classification of
flowering plants. Columbia University Press, New York
Davis CC, Latvis M, Nickrent DL, Wurdack KJ, Baum DA (2007)
Floral gigantism in Rafflesiaceae. Science 315:1812
Dickinson WC (1990) A study of the floral morphology and anatomy
of the Caryocaraceae. Bull Torrey Bot Club 117:123–137
Dickinson WC, Weitzman AL (1998) Floral morphology and
anatomy of Bonnetiaceae. J Torrey Bot Soc 125:268–286
Domınguez CA, Bullock SH (1989) La reproduccion de Crotonsuberosus (Euphorbiaceae) en luz y sombra. Rev Biol Tropical
37:1–10
Endress PK (2008) The whole and the parts: relationships between
floral architecture and floral organ shape and their repercussions
on the interpretation of fragmentary floral fossils. Ann Mo Bot
Gard 95:101–120
Freitas L, Sazima M (2003) Floral biology and pollination mecha-
nisms in two Viola species—from nectar to pollen flowers? Ann
Bot 91:311–317
Freitas L, Bernardello G, Galetto L, Paoli AAS (2001) Nectaries and
reproductive biology of Croton sarcopetalus (Euphorbiaceae).
Bot J Linn Soc 136:267–277
Govaerts R, Frodin DG, Radcliffe-Smith A (2000) World checklist of
Euphorbiaceae (and Pandaceae). Royal Botanic Gardens, Kew
Johansen DA (1940) Plant microtechnique. McGraw-Hill Book, New
York
Matthews ML, Endress PK (2008) Comparative floral structure and
systematics in Chrysobalanaceae s.l. (Chrysobalanaceae,
Dichapetalaceae, Euphroniaceae, and Trigoniaceae; Malpighi-
ales). Bot J Linn Soc 157:249–309
Michaelis P (1924) Blutenmorphologische Untersuchungen an den
Euphorbiaceen unter besonderer Berucksichtigung der Phylog-
enie der Angiospermenblute. Bot Abh 3:1–150
Miller KI, Webster GL (1966) Chromosome numbers in the
Euphorbiaceae. Brittonia 18:372–379
Nair NC, Abraham V (1962) Floral morphology of a few species of
Euphorbiaceae. Proc Ind Acad Sci B 56:1–12
O’Brien TP, Feder N, McCully ME (1964) Polychromatic staining of
plant cell walls by toluidine blue O. Protoplasma 59:368–373
Homologies of the perianth and floral nectaries in Croton and Astraea 13
123
Author's personal copy

Prance GT, Freitas da Silva MF (1973) Caryocaraceae. Flora
Neotropica Monograph 12. The New York Botanical Garden,
New York
Prenner G, Rudall PJ (2007) Comparative ontogeny of the cyathium in
Euphorbia (Euphorbiaceae) and its allies: exploring the organ–
flower–inflorescence boundary. Amer J Bot 94:1612–1629
Prenner G, Hopper S, Rudall PJ (2008) Pseudanthium formation in
Calycopeplus paucifolius with particular reference to the evo-
lution of the cyathium in Euphorbieae (Euphorbiaceae–Mal-
pighiales). Aust Syst Bot 21:153–161
Prenner G, Bateman RM, Rudall PJ (2010) Floral formulae updated
for routine inclusion in formal taxonomic descriptions. Taxon
59:241–250
Puri V (1951) The role of floral anatomy in the solution of
morphological problems. Bot Rev 17:471–553
Radcliffe-Smith A (2001) Genera Euphorbiacearum. Royal Botanic
Gardens, Kew
Riina R, Berry PE, van Ee BW (2009) Molecular phylogenetics of the
Dragon’s Blood Croton section Cyclostigma (Euphorbiaceae): a
polyphyletic assemblage unraveled. Syst Bot 42:360–374
Robson NKB (1961) Guttiferae. In: Exel AW, Will H (eds) Flora
Zambesiaca 1. Crown Agents for Overseas Governments and
Administrations, London, pp 371–375
Ronse Decraene LP, Smets EF (1991) Androecium and floral
nectaries of Harungana madagascariensis (Clusiaceae). Plant
Syst Evol 178:179–194
Smets EF (1986) Localization and systematic importance of the floral
nectaries in the Magnoliatae (Dicotyledons). Bull Jard Bot Natl
Belg 56:51–76
Tokuoka T (2007) Molecular phylogenetic analysis of Euphorbiaceae
sensu stricto based on plastid and nuclear DNA sequences and
ovule and seed character evolution. J Pl Res 120:511–522
Tokuoka T, Tobe H (2006) Phylogenetic analyses of Malpighiales
using plastid and nuclear DNA sequences, with particular
reference to the embryology of Euphorbiaceae sens. str. J Pl
Res 119:599–616
van Ee BW, Berry PE, Riina R, Amaro JEG (2008) Molecular
phylogenetics and biogeography of the Caribbean-Centered
Croton subgenus Moacroton (Euphorbiaceae s.s.). Bot Rev
74:132–165
Venkata-Rao C, Ramalakshmi T (1968) Floral anatomy of the
Euphorbiaceae–I. Some non-cyathium taxa. J Ind Bot
47:278–300
Vogel S (1998) Remarkable nectaries: structure, ecology, organo-
phyletic perspectives: III. Nectar ducts. Flora 193:113–131
Weberling F (1989) Morphology of flowers and inflorescences.
Cambridge University Press, Cambridge
Webster GL (1993) A provisional synopsis of the sections of the
genus Croton (Euphorbiaceae). Taxon 42:793–823
Webster GL (1994) Synopsis of the genera and suprageneric taxa of
Euphorbiaceae. Ann Mo Bot Gard 81:33–144
Wurdack KJ, Davis CC (2009) Malpighiales phylogenetics: gaining
ground on one of the most recalcitrant clades in the angiosperm
tree of life. Amer J Bot 96:1551–1570
Wurdack KJ, Hoffmann P, Chase MW (2005) Molecular phylogenetic
analysis of uniovulate Euphorbiaceae (Euphorbiaceae sensu
stricto) using plastid rbcL and trnL-trnF sequences. Amer J Bot
92:1397–1420
14 O. C. De-Paula et al.
123
Author's personal copy





















