Embed Size (px)
Citation preview

Research article
Received: 29 November 2013 Revised: 30 April 2014 Accepted: 9 May 2014 Published online in Wiley Online Library: 30 May 2014
(wileyonlinelibrary.com) DOI 10.1002/jrs.4517
1180
Identification of endolithic traces on stonemonuments and natural outcrops: preliminaryevidences†
A. Casanova Municchia,* G. Caneva, M. A. Ricci and A. Sodo
Four different lithotypes of rock gathered from stone monuments and natural outcrops in Temperate and Mediterraneanbioclimates, with endolithic microorganism colonization (cyanobacteria, fungi, and lichens), have been analysed by Ramanspectroscopy. Endolithic microorganisms can penetrate actively into the rocks giving rise to biodeterioration phenomena.Often the biological origin of these deterioration phenomena is not recognized and confused with abiotic damages. Theaim of this work is to identify by Raman spectroscopy the endolithic traces on stones, due to current or past biologicalcolonization. The Raman spectra show the presence of bio-markers characteristic of the strategy adopted by microorganismsagainst strong UV-radiation and dry conditions, such as scytonemin, anthraquinone compounds, and carotenoids. Moreover,we have identified different geo-markers such as goethite and lepidocrocite because of the mobilization of iron by lichenmetabolic activity. Importantly, we have identified for the first time by Raman spectroscopy, traces of endolithicmicroorganisms on stone monuments located in Temperate and Mediterranean bioclimate, where extreme climate conditionsare not foreseen. Copyright © 2014 John Wiley & Sons, Ltd.
Keywords: endolithic; stone monuments; biodeterioration; Raman spectroscopy
* Correspondence to: Annalaura Casanova Municchia, Università Roma Tre,Dipartimento di Scienze, Viale G. Marconi 446, 00146 Rome, Italy.E-mail: [email protected]
† This article is part of the special issue of the Journal of Raman Spectroscopyentitled “Raman in Art and Archaeology 2013” edited by Polonca Ropret andJuan Manuel Madariaga.
Dipartimento di Scienze, Università Roma Tre, Viale G. Marconi 446, 00146Rome, Italy
IntroductionEndolithic growth forms can be present in different microorgan-isms groups including cyanobacteria, green and red algae, fungi,and lichens. This kind of growth inside the rocks derives from theneed to protect against environmental factors inhibiting epilithicgrowth, such as intense solar radiations, adverse temperature anddesiccation conditions.[1–3] Their depth of penetration dependson their species, rock composition and ecologic factors.[4–12]
The observation of their presence dates back to the 1960s oflast century, when the pioneering work of Friedman andcoworkers,[3] on the desert environment, appeared. Since then,traces of endolithic life have been detected in several environ-ments, including the Antarctic desert, and have captured theinterest of several scientific communities, ranging from astro-biology to geo-biology.[13–16] More recently the presence ofendolithic colonies has been proved also in Mediterranean,Temperate and Tropical regions,[6,12,17–19] in particular onmonuments, on statues and historic buildings, damaged by thegrowth and survival strategy of these microrganisms.[2,20–23]
The survival strategy of endolithic microorganisms encompassesproduction of photoprotective or light harvesting pigments againstUV-radiation and darkness, respectively, or calcium oxalates againstdrought.[24–26] Moreover, as they migrate deep in the stone, theymay modify the substratum by selective mobilization of rockconstituents and crystallization into new mineral phases.[26,27]
Along with an aesthetic alteration of surfaces, they can cause adetachment of stone as a scale (chasmoendolithic) or they candissolve the substrate giving rise to the formation of circularholes on the surfaces (euendolithic).[17] Their impact on thecultural heritage is usually underestimated, since the endolithicdeterioration marks can be confused with weathering ormechanic damages.[2,28]
J. Raman Spectrosc. 2014, 45, 1180–1185
Raman spectroscopy is an efficient tool in order to identifytraces of the organic and inorganic compounds. In particular,since endolithic bio- and geo-markers can be preserved in thegeological record, this technique can be used to identifycompounds sedimented by endolithic microorganisms as protectivemechanisms against adverse environmental conditions. As a matterof fact this technique has been extensively employed in astro-biology, in order to identify key spectral markers of extremophilesmicroorganisms, and a rich spectra database of bio-markersand geo-markers related to life activity in hostile environments isnow available.[24,25,29–32]
For instance, Edwards et al.,[33,34] in a study about the survivalstrategies of lichens in stressed habitats, report the Raman bio-marker bands in the spectra of metabolic lichen products(atranorin, gyrophoric acid, fumarprotocetraric acid rhizocarpicacid, calycin, pulvinic dilactone, usnic acid, parietin and emodin).Other studies identify key bio- and geo-markers, produced bymicroorganisms as UV-protective pigments (carotenoids,scytonemin), antidesiccants (calcium oxalate), and pigments usefulin light-deficient conditions (phycocyanin or phycoerythrin).[26]
Edwards et al.,[35] establish the important role of scytonemin asUV-screening pigment found in cyanobacterial sheaths, and assign
Copyright © 2014 John Wiley & Sons, Ltd.

Identification of endolithic traces on stone monuments and natural outcrops: preliminary evidences
four vibrational Raman bands to the scytonemin compound.Another strategy of endolithic microorganisms against UV-radiation is the mobilization of some iron oxides.[27]
In this paper we perform a preliminary Raman spectroscopicanalysis on endolithic samples from Temperate andMediterranean bioclimatic regions, in order to detect traces ofsuch colonization on stones.
Experimental
Samples
Four different rock types, colonized by endolithic microorgan-isms (Fig. 1), namely two fragments belonging to stone monu-ments and two from natural outcrops set in Temperate andMediterranean bioclimate regions (Fig. 2) were investigated byRaman spectroscopy. The monument fragments were blackschist and carbonatic limestone; one fragment from naturaloutcrops was Carrara marble, the other was a dolomite rock. Thisselection has been done in order to have a wide choice of rocks,including the most common stone used in monuments and mostaffected by endolithic attack[6] (Carrara Marble) and the lessstudied so far (dolomitic limestone). In detail, the samples were:
E1: Marble rock showing colonization of cyanobacteria andendolithic fungi (Fig. 1), from quarries in Carrara Area,characterized by a Temperate bioclimate
Figure 1. Optical microscope images of the cross sections of samples E1, E2, ansample E1 is a marble rock from quarries in Carrara Area; sample E2 is a dolomHebrew’s cemetery tombstone in Venice; image E4 is the rear side of the fine-gr
Figure 2. Bioclimatic map of Europe, showing the four sites, located in Tsamples come from. (Worldwide Bioclimatic Classification System, 1996 e 20
J. Raman Spectrosc. 2014, 45, 1180–1185 Copyright © 2014 Joh
E2: Dolomitic limestone showing colonization of endolithic
d E3itic limained
empe09).[3
n Wi
cyanobacteria (Fig. 1), from cliff of the Amalfi Coast in the Capod’Orso area, characterized by a Mediterranean bioclimate
E3: Black schist fromHebrew’s cemetery tombstone in Venice, which is
characterized by a Temperate bioclimate (Fig. 1). This sample showsendolithic lichens, indeed its surface is covered by chalky whitesmooth thallus with perithecia completely sunken in the rockE4: Fine-grained limestone from the Church of the Virgin in
Martvili inWestern Georgia, which is characterized by a Temper-ate bioclimate (Fig. 1). The back side of the sample shows orangetraces ascribable to the presence of cyanobacteria endolithiccommunities.Sample preparation and optical microscopy
Raman measurements were performed on E4 sample withoutany preparation, because a biological colonization was alreadyevident on the rear side of the fragment (Fig. 1), implyingcolonization in depth. On the contrary for samples E1, E2 andE3, polished cross-sections were prepared, in order to investigatethe fragment in depth. These were obtained after inclusion in apolyester resin according to UNI 10922,[37] for the sake of identi-fying endolithic microorganisms inside the stone.
All samples were photographed by a Wild M3 stereomicroscope,then the cross-sections were observed with a Leitz Orthoplanmicroscope and digital photographs were taken with a LeicaDC300 digital camera and the IM100 software.
, along with the stereomicroscope image of sample E4. We remind thatestone from cliff of the Amalfi Coast; sample E3 is a black schist fromlimestone from the Church of the Virgin in Martvili in Western Georgia.
rate and Mediterranean macrobioclimates, where the investigated6]
ley & Sons, Ltd. wileyonlinelibrary.com/journal/jrs
1181

A. Casanova Municchia et al.
1182
The characterization of endolithic microorganisms wascarried out through observation by microscope with animmersion objective at 100x magnification (Olympus BX41)as described by standard method UNI 10923, based on thepreparation of biological specimens for observation underthe optical microscope.[38]
Figure 3. Microphotographs of the sample E1 (from Carrara area) inpolished cross section, showing the points corresponding to the reportedRaman spectra.
Raman microscopy
The substrates and the endolithic metabolic products of the rockfragments were investigated by a Renishaw In-Via Reflex Ramanmicroscope equipped with two different sources, at 785.5 andat 514.5 nm respectively.All the samples were investigated by using a Renishaw diode
laser at 785 nm (nominal output power 300mW) and onlywhen we could not obtain any Raman information above theintense fluorescence band (samples E2 and E4), a 514.5 nm laser(Ar+ ions, nominal output power 25mW) was used. Usually,near-infrared laser excitation minimizes the interferences causedby fluorescence background in organic compounds and it isrecommended by several studies[25,26,39] for this kind of samples.In particular, Villar et al.,[26] in their work reporting a comparativeevaluation of Raman spectroscopy at different wavelength forendolithic investigations, conclude that the 785nm laserwavelength is the most suitable for the analysis of both geo- andbiomarkers for a wide range of environmental conditions experi-enced by these microorganisms. The 514.5 nm excitation is insteadmore suitable in identification of carotenoid compounds.In order to avoid degradation of bio-markers, neutral filters,
when necessary, were used. No absolute a priori power ‘at thesample’ could be chosen, depending on the investigated sample.For each, trial measurements were attempted, starting fromabout 0.1mW at the surface, and gradually increasing the intensityuntil an acceptable signal-to-noise ratio was obtained. The finalexperimental conditions varied between 1 and 25mW. Thebackscattered light was dispersed by a 1200 line/mm grating forthe 785.5 nm laser and by a 1800 line/mm grating for the514.5 nm source. The Raman signal was detected by a Peltiercooled (–70 °C) deep depletion charge- coupled device (CCDRD-VIU, 578 x384 pixel, with spectral response in the range200–1025nm). Nominal spectral resolution was about 3 cm-1. Thesystem, equipped with a Leica DMLMmicroscope to focus the laseron the sample and a color video camera, allows for positioning ofthe sample and selection of a specific area for investigation. Spec-tral acquisitions (three accumulations, 50 s each, in the range200–3200 cm-1) were performed with a 50x objective (N.A. = 0.75).
Results and discussion
Raman spectra obtained from the endolithic specimens havebeen recorded on the surface and in the inner part of thesamples, in order to identify a possible difference of the bio- andgeotraces detected at different depths. Raman spectra have beenregistered using both the 785nm and 514nm excitation lines.Several Raman measurements on different fragments of the
same type of rocks were performed, and in this work only themost representative ones are reported.In good agreement with earlier studies,[26,30] in all samples
investigated bio-traces ascribable to endolithic microorganismsoccurrence were detected.The spectra of the individual samples are described below.
wileyonlinelibrary.com/journal/jrs Copyright © 2014 John
E1 (Marble quarries of Carrara)
We report five spectra among those collected on this sampleby using the laser exciting line in the near-infrared region:the points probed are labeled from 1 to 5 in Fig. 3 and spanfrom the stone surface, where was evident a greenish coloni-zation, down to the inner part of the sample, not apparentlycolonized.
Figure 4A reports the Raman spectra collected from this sample.All the spectra clearly show the calcite [CaCO3] most intense bandat 1086 cm�1 while the calcite bands at 712 and 282 cm�1 arevisible only in the spectra 2, 3 and 4 e 5. Calcite is indeed expectedto be the main component of this marble. Moreover, the spectracollected from points 3 and 4 show the four corroborative Ramanbands of scytonemin at 1590, 1554, 1323 and 1176 cm�1[35]
(labeled with arrows in Fig. 4B, where a detail in the range1100–1800 cm�1 is reported). The absence of the Raman bands at1627 and 1502 cm-1, and the presence of the band at 1177 cm-1
could confirm the presence of oxidized scytonemin.[40] Scytoneminis exclusively synthesized by cyanobacteria as extracellular sheathpigment, and it plays an essential role as photoprotective compoundagainst UV radiation.[30,41] This is a significant finding, indicative ofthe presence of endolithic cyanobateria in conditions of environ-mental stress. Raman spectrum from point 2 shows, in addition toscytonemin fingerprints, a very broad band, centered at 1334 cm�1
(labeled with an asterisk in Fig. 4B), ascribable to the presence ofchlorophyll. Spectrum 1 shows the scytonemin contribution, plusbands at 1650, 1553, 1387,1276 cm�1 and weaker signals at 1181and 463 cm-1: these signatures have been assigned to anthraquinonecompounds[25,33,39] and are labeled with circles in Fig. 4A. Finally, inspectrum 5 Raman bands at 1279, 1244,1173 cm�1 (labeled withcircles in Fig. 4b), ascribable to anthraquinone compounds, wereobserved. Usually anthraquinones are reported as lichenproducts which protect the photobiont against intense radia-tion, but, interestingly, Villar et al.,[26] have found an anthraqui-none, namely parietin, among the products of a halophiliccyanobacteria community.
Some of these spectra are affected by the presence of the resinbands at 1733,1602,1584, 1453, 1290, 1198, 1165, 1039, 1002 cm-1,as a residue of the sample preparation (Fig. 4B).
Wiley & Sons, Ltd. J. Raman Spectrosc. 2014, 45, 1180–1185
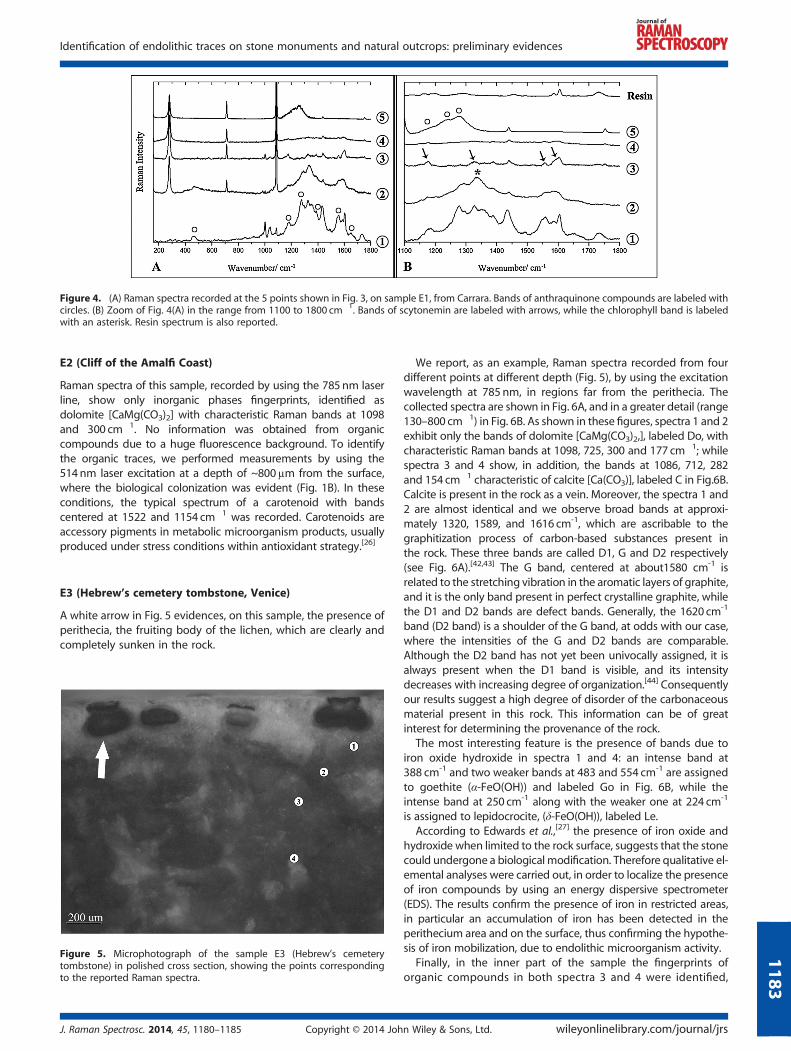
Figure 4. (A) Raman spectra recorded at the 5 points shown in Fig. 3, on sample E1, from Carrara. Bands of anthraquinone compounds are labeled withcircles. (B) Zoom of Fig. 4(A) in the range from 1100 to 1800 cm�1. Bands of scytonemin are labeled with arrows, while the chlorophyll band is labeledwith an asterisk. Resin spectrum is also reported.
Identification of endolithic traces on stone monuments and natural outcrops: preliminary evidences
E2 (Cliff of the Amalfi Coast)
Raman spectra of this sample, recorded by using the 785nm laserline, show only inorganic phases fingerprints, identified asdolomite [CaMg(CO3)2] with characteristic Raman bands at 1098and 300 cm�1. No information was obtained from organiccompounds due to a huge fluorescence background. To identifythe organic traces, we performed measurements by using the514nm laser excitation at a depth of ~800μm from the surface,where the biological colonization was evident (Fig. 1B). In theseconditions, the typical spectrum of a carotenoid with bandscentered at 1522 and 1154 cm�1 was recorded. Carotenoids areaccessory pigments in metabolic microorganism products, usuallyproduced under stress conditions within antioxidant strategy.[26]
E3 (Hebrew’s cemetery tombstone, Venice)
A white arrow in Fig. 5 evidences, on this sample, the presence ofperithecia, the fruiting body of the lichen, which are clearly andcompletely sunken in the rock.
Figure 5. Microphotograph of the sample E3 (Hebrew’s cemeterytombstone) in polished cross section, showing the points correspondingto the reported Raman spectra.
J. Raman Spectrosc. 2014, 45, 1180–1185 Copyright © 2014 Joh
118
We report, as an example, Raman spectra recorded from fourdifferent points at different depth (Fig. 5), by using the excitationwavelength at 785 nm, in regions far from the perithecia. Thecollected spectra are shown in Fig. 6A, and in a greater detail (range130–800 cm�1) in Fig. 6B. As shown in these figures, spectra 1 and 2exhibit only the bands of dolomite [CaMg(CO3)2,], labeled Do, withcharacteristic Raman bands at 1098, 725, 300 and 177 cm�1; whilespectra 3 and 4 show, in addition, the bands at 1086, 712, 282and 154 cm�1 characteristic of calcite [Ca(CO3)], labeled C in Fig.6B.Calcite is present in the rock as a vein. Moreover, the spectra 1 and2 are almost identical and we observe broad bands at approxi-mately 1320, 1589, and 1616 cm-1, which are ascribable to thegraphitization process of carbon-based substances present inthe rock. These three bands are called D1, G and D2 respectively(see Fig. 6A).[42,43] The G band, centered at about1580 cm-1 isrelated to the stretching vibration in the aromatic layers of graphite,and it is the only band present in perfect crystalline graphite, whilethe D1 and D2 bands are defect bands. Generally, the 1620 cm-1
band (D2 band) is a shoulder of the G band, at odds with our case,where the intensities of the G and D2 bands are comparable.Although the D2 band has not yet been univocally assigned, it isalways present when the D1 band is visible, and its intensitydecreases with increasing degree of organization.[44] Consequentlyour results suggest a high degree of disorder of the carbonaceousmaterial present in this rock. This information can be of greatinterest for determining the provenance of the rock.
The most interesting feature is the presence of bands due toiron oxide hydroxide in spectra 1 and 4: an intense band at388 cm-1 and two weaker bands at 483 and 554 cm-1 are assignedto goethite (α-FeO(OH)) and labeled Go in Fig. 6B, while theintense band at 250 cm-1 along with the weaker one at 224 cm-1
is assigned to lepidocrocite, (δ-FeO(OH)), labeled Le.According to Edwards et al.,[27] the presence of iron oxide and
hydroxide when limited to the rock surface, suggests that the stonecould undergone a biological modification. Therefore qualitative el-emental analyses were carried out, in order to localize the presenceof iron compounds by using an energy dispersive spectrometer(EDS). The results confirm the presence of iron in restricted areas,in particular an accumulation of iron has been detected in theperithecium area and on the surface, thus confirming the hypothe-sis of iron mobilization, due to endolithic microorganism activity.
Finally, in the inner part of the sample the fingerprints oforganic compounds in both spectra 3 and 4 were identified,
n Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jrs
3

Figure 6. (A) Raman spectra recorded at the 4 points shown in Fig. 5, on sample E3, from Hebrew’s cemetery tombstone in Venice. Anthraquinonecompounds are labeled with circles, G D1 and D2 label the finger prints of carbonaceous materials. (B) Zoom of the spectra in the range from 130 to800 cm�1. (Do, dolomite; C, calcite; Go, goethite; Le, Lepidocrocite).
Table 1. Summary of the Raman bands of substrate and of the geo- and biotraces detected in the four samples
Characteristic Raman signatures (cm�1)
Compounds E1 E2 E3 E4
Bio-traces
Chlorophyll 1334
Carotenoid 1522, 1154 1507, 1148
Scytonemin 1590,1554,1323,1176
Anthraquinone 1650,1553,1387,1276, 1181, 463 1279, 1171
Geo-traces
Lepidocrocite 250, 224
Ghoetite 554, 483, 388
Substrate
Calcite 1086, 712, 282 1086, 712, 282, 154 1086, 712, 282
Dolomite 1098, 300 1143, 1098, 725, 300, 177
Carbon-based substances 1616, 1589, 1329
The corroborative bands of each compound are reported in bold.
A. Casanova Municchia et al.
1184
where an intense Raman band at 1279 cm-1 and a weakerband at 1171 cm�1 (labeled with circles in Fig. 6A) wereobserved that could be assigned to anthraquinone com-pounds. This organic compound has been only detected inthe spectra where the calcite was present. Probably due tothe higher solubility of calcite compared to dolomite andconsequent higher sensitivity to acid attack by endolithicmicroorganisms.
E4 (Church of the Virgin in Martvili, Western Georgia)
The substrate of this sample is mainly calcite identified by thetypical bands at 1086, 712 and 282 cm�1. As for sample E2, nouseful information on bio and geo-markers was obtained byusing the 785.5 nm laser line. On the contrary, by using the laserline at 514 nm, the presence of carotenoid bands at 1507 and1148 cm�1 was revealed.A summary of all the results obtained for the four samples, in
terms of substrate and bio- and geotraces, and their characteris-tic bands assignment is reported in Table 1.
wileyonlinelibrary.com/journal/jrs Copyright © 2014 John
Conclusions
Ourwork is the first study showing Raman spectroscopic applicationsto stone monuments and natural outcrops affected by endolithiccolonizations in Temperate and Mediterranean bioclimate regions.
Traces of both biological and geological compounds used byendolithic colonies within their survival strategy have beendetected not only just below the sample surface but also indepth, where the sample does not show evident traces ofcolonization. This demonstrates that endolithic microorganismscan penetrate down to millimeters inside the rock.
We observe no strict connection between detected bio and geo-markers and the bioclimates provenance of our samples. Indeed, inboth E2 (Cliff of the Amalfi Coast, Mediterranean bioclimatic region)and E4 (Church of the Virgin in Martvili, Temperate bioclimaticregion) samples we have identified carotenoid compounds (antiox-idant). The presence, in either bioclimates, of these biological tracesuseful against the high UV-radiation could be explained accordingto the work of Lombardozzi et al.,.[6] These authors report that theoccurrence of the endolithic microorganisms on stones depends
Wiley & Sons, Ltd. J. Raman Spectrosc. 2014, 45, 1180–1185

Identification of endolithic traces on stone monuments and natural outcrops: preliminary evidences
on several different factors. Not only bioclimate, but also specificmicroclimatic parameters (exposure to high solar radiation, low wa-ter input, high ventilation), orientation of the surface (as vertical orsubvertical surfaces) and stone characteristic (preferably low poros-ity), could favor the growth of endolithic microorganisms. This im-plies that similar extreme conditions on a stone surface may begenerated in quite different climatic regions, and this couldexplain the same survival strategy adopted by endoliths in oursamples from Mediterranean and Temperate bioclimates.
Both E1 (Marble of Carrara, Temperate bioclimatic regions) andE3 (Tombstone in Venice, Temperate bioclimatic regions)samples show traces of compounds known to be effectiveagainst UV-radiation damage. In particular, in sample E1 we havefound scytonemin and anthraquinone compounds and in sampleE3 anthraquinone compounds and iron localization at the surfaceof the stone, where the lichens structure is visible.
Finally we have shown that endolithic microorganism colonizestones also at non-extreme bioclimatic conditions, if microclimaticfactors give rise to local extreme-conditions. A systematic study onsamples of different mineralogical composition and provenance isneeded to assess the relevance on endolithic colonization of chemi-cal-physical parameters versus climatic or topographic factors.
Our paper confirms that Raman spectroscopy is a useful tool toinvestigate samples colonized by endolithic microorganisms. Theuse of this technique, in the field of biodeterioration of stone mon-uments, can be useful for a clear identification of the presence ofendolithic microorganisms as opposite to abiotic alterations. Westress however that such studies require micro-sampling whichmay be not always feasible on cultural heritage.
Acknowledgements
The authors wish to thank Dr. M. Bicchieri, Responsible for thelaboratory of Chemistry (ICRCPAL); Dr. C. Romano, Head of theLaboratory for Experimental Vulcanology and Petrology (Univer-sity of Roma Tre), and Dr. E. Bemporad, Responsible for theInterdepartmental Laboratory of Electron Microscopy (LIME), forthe technical support.
References
[1] J. J. Walker, N. R. Pace, Annu. Rev. Microbiol. 2007, 61, 331.[2] G. Caneva, V. Lombardozzi, S. Ceschin, A. Casanova Municchia,
O. Salvadori, J. Cult. Herit. 2013, http://dx.doi.org/10.1016/j.culher.2013.10.003
[3] E. Imre Friedmann, Science 1982, 215, 1045.[4] G. Caneva, S. Ceschin, in Plant Biology for Cultural Heritage. Biodeteri-
oration and Conservation (Eds: G. Caneva, M. P. Nugari, O. Salvadori),The Getty Conservation Institute, Los Angeles, 2009, pp. 35–58.
[5] U. Matthes, S. J. Turner, D. W. Larson, Int. J. Plant Sci. 2012, 162, 263.[6] V. Lombardozzi, T. Castrignanò, M. D’Antonio, A. Casanova
Municchia, G. Caneva, Int. Biodeterior. Biodegrad. 2012, 73, 8.[7] A. Danin, Isr. J. Earth Sci. 1992, 41, 201.[8] A. Gorbushina, Environ. Microbial. 2007, 9, 1613.[9] S. Onofri, E. Imre Friedmann, in Newsletter of the Italian Biological
Research in Antarctica (Eds: M. Tamburini, R. d’Avino), UniversityPress, Camerino, 1999, 2, pp. 45–51.
J. Raman Spectrosc. 2014, 45, 1180–1185 Copyright © 2014 Joh
[10] K. Sterflinger, Geomicrobiol. J. 2000, 17, 97.[11] A. De Los Ríos, L. G. Sancho, M. Grube, J. Wierzchos, C. Ascaso,
New Phytol. 2005; 165, 181.[12] D. Pinna, O. Salvadori, M. Tetriach, Plant Biosyst. 1998, 132, 183.[13] H. G. M. Edwards, Phil. Trans. R. Soc. A 2010, 368, 3059–3065[14] H. G. M. Edwards, E. M. Newton, D. L. Dickensheets, D. D. Wynn-Williams.
Spectrochim. Acta Part A 2003, 59, 2277.[15] D. D.Wynn-Williams, H. G. M. Edwards, Planet. Space Sci. 2000, 48, 1065.[16] H. G. M. Edwards, M. A. Mohsin, F. N. Sadooni, N. F. N. Hassan, T.
Munshi, Anal. Bioanal. Chem. 2006, 385, 46.[17] S. Golubic, R. D. Perkins, K. J. Lukas, in Study of Trace Fossils (Ed: R.
Frey), Springler-Verlag, New York, 1975, pp. 229–259.[18] C. Saiz-Jimenez, J. Garcia-Rowe, M. A. Garcia Del Cura, J. J. Ortega-
Calvo, E. Roekens, R. Van Grieken, Sci.Total Environ. 1990, 94, 209.[19] C. J. McNamara, T. D. Perry, K. A. Bearce, G. Hernandez-Duque,
R. Mitchell, Microb. Ecol. 2006, 51, 51.[20] C. Ascaso, J. Wierzchos, J. Delgado Rodrigues, L. Aires-Barros, F. M. A.
Henriques, A. E. Charola, Intern. Z. Bauinstandsetzen 1998, 4, 627.[21] G. Caneva, A. Danin, S. Ricci, C. Conti, Rend. accad. Lincei 1914, 88, 77.[22] G. Caneva, M. P. Nugari, O. Salvadori, in Proceedings of the Seventh In-
ternational Congress on Deterioration and Conservation of Stone (Eds:J. D. Rodriguez, F. Henriques, F. T. Jeremias), LNNA, Lisbon, 1992, p. 521.
[23] O. Salvadori, in Of Microbes and Art. The Role of Microbial Commu-nities in the Degradation and Protection of Cultural Heritage(Eds: O. Ciferri, P. Tiano, G. Mastromei), Plenum, London, 2000,pp. 89–101.
[24] S. E. Jorge Villar, H. G. M. Edwards, C. S. Cockell, Analyst, 2005, 130, 156.[25] S. E. Jorge Villar, H. G. M. Edwards, Anal. Bioanal. Chem. 2006, 384, 100.[26] S. E. Jorge Villar, H. G. M. Edwards, L. Benning, Icarus 2006, 184, 158.[27] H. G. M. Edwards, D. D. Wynn-Williams, S. E. Jorge Villar, J. Raman
Spectrosc. 2004, 35, 470.[28] D. Pinna, O. Salvadori, in Proceedings of the Ninth International Con-
gress on Deterioration and Conservation of Stone (Ed: V. Fassina),Elsevier, Venice, 2000, p. 513.
[29] S. E. Jorge Villar, L. G. Benning, H. G. M. Edwards, Geochem. Trans.2007, 8, 8.
[30] S. E. Jorge Villar, H. G. M. Edwards, L. G. Benning, Anal. Bioanal. Chem.2011, 401, 2927.
[31] D. L. Dickensheets, D. D. Wynn-Williams, H. G. M. Edwards, C. Schoen,C. Crowder, E. M. Newton, J. Raman Spectrosc. 2000, 31, 633.
[32] S. E. Jorge Villar, H. G. M. Edwards, Life 2013, 3, 276.[33] H. G. M. Edwards, E. M. Newton, D. D. Wynn-Williams, S. R. Coombes,
J. Mol. Struct. 2003, 648, 49.[34] H. G. M. Edwards, E. M. Newton, D. D. Wynn-Williams, J. Mol. Struct.
2003, 651, 27.[35] H. G. M. Edwards, F. Garcia-pichel, E. M. Newton, D. D. Wynn-williams,
Spectrochim. Acta Part A 1999, 56, 193.[36] S. Rivas-Martinez, S. Rivas-Saenz, Worldwide bioclimatic classification
system. Phytosociological Research Center, Spain, 1996-2009,http://www.globalbioclimatics.org.
[37] UNI 10922, Beni Culturali. Materiali lapidei naturali ed artificiali.Allestimento di sezioni sottili e sezioni lucide di materiali lapideicolonizzati da biodeteriogeni. UNI, Milano, 2001.
[38] UNI 10923, Beni Culturali. Materiali lapidei naturali ed artificiali.Allestimento di preparati biologici per l’osservazione al microscopioottico. UNI, Milano, 2000.
[39] S. E. Jorge Villar, H. G. M. Edwards, M. R. Worland, Orig. Life Evol.Biosphere 2005, 35, 489.
[40] T. Varnali, H. G. M. Edwards, Spectrochim. Acta Part A 2014, 117, 72.[41] J. G. Dillon, R. W. Castenholz, J. Phycol. 1999, 35, 673.[42] J. M. Rahl, K. M. Anderson, M. T. Brandon, C. Fassoulas, Earth Planet.
Sci. Lett. 2005, 240, 339.[43] O. Beyssac, J. N. Rouzaud, B. Goffé, F. Brunet, C. Chopin, Contrib.
Mineral. Petrol. 2002, 143, 19.[44] O. Beyssac, B. Goffé, J. Petitet, E. Froigneux, M. Moreau, J. Rouzaud,
Spectrochim. Acta Part A 2003, 59, 2267.
n Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jrs
1185





























