Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Biomaterials 33 (2012) 4975e4987
Contents lists available
Biomaterials
journal homepage: www.elsevier .com/locate/biomater ia ls
tedLeading opinion
Theoretical considerations on mechanisms of harvesting cells cultured onthermoresponsive polymer brushesq
Avraham Halperin a,*, Martin Kröger b,*aUniversity of Grenoble 1/CNRS, LIPhy UMR 5588, BP 87, 38041 Grenoble, Franceb Polymer Physics, Department of Materials, ETH Zurich, CHe8093 Zurich, Switzerland
a r t i c l e i n f o
Article history:Received 10 February 2012Accepted 17 March 2012Available online 12 April 2012
Keywords:Cell cultureFibronectinIntegrinMembraneModelingProtein adsorption
Rq Editor’s Note: This paper is one of a newly institut
that provide evidence-based scientific opinions on tobiomaterials science. They have some features of an inon scientific facts, and some features of a review papcomprehensive. These papers have been commissionreviewed for factual, scientific content by referees.* Corresponding authors.
E-mail addresses: [email protected] (M. Kröger).
0142-9612/$ e see front matter � 2012 Elsevier Ltd.doi:10.1016/j.biomaterials.2012.03.060
vided
copy
n not
perm
ita b s t r a c t
Poly (N-isopropylacrylamide) (PNIPAM) brushes and hydrogels serve as temperature-responsive cellculture substrates. The cells adhere at 37 �C and are detached by cooling to below the lower critical solutiontemperature TLCST z 32 �C, an effect hitherto attributed to change in PNIPAM hydration. The articleproposes a mechanism coupling the change of hydration to integrin mediated environmental sensing forcell culture on brushes and hydrogels in serum containing medium. Hydration is associated with swellingand higher osmotic pressure leading to two effects: (i) The lower osmotic pressure in the collapsed brush/hydrogel favors the adsorption of serum borne extracellular matrix (ECM) proteins enabling cell adhesion;(ii) Brush/hydrogel swelling at T < TLCST gives rise to a disjoining force fcell due to confinement by theventral membrane of a cell adhering via integrin-ECM bonds. fcell places the integrineECM bonds undertension thus accelerating their dissociation and promoting desorption of ECMproteins. Self consistentfieldtheory of PNIPAM brushes quantifies the effect of the polymerization degree N, the area per chain S, andthe temperature, T on ECM adsorption, fcell and the dissociation rate of integrineECM bonds. It suggestsguidelines for tuning S and N to optimize adhesion at 37 �C and detachment at T < TLCST. The mechanismrationalizes existing experimental results on the influence of the dry thickness and the RGD fraction onadhesion and detachment.
� 2012 Elsevier Ltd. All rights reserved.
ro o iAuthor
p
edist
ribut1. Introduction
Cultured anchorage dependent cells are often harvested usingproteolitic enzymes [1] thus damaging the extracellular matrix(ECM), cellecell junctions and cell surface receptors. Temperatureresponsive cell culture substrates enable “non invasive” detach-ment sparing these components and thus conducive to tissueengineering applications [2,3]. Among these substrates, surfacelayers of poly (N-isopropylacrylamide) (PNIPAM) received mostattention leading to corresponding technological developments[4e7]. These layers allow cell adhesion and proliferation at 37 �C,above the lower critical transition temperature (LCST) of PNIPAM at
ed series of scientific articlespical and important issues invited editorial but are baseder, without attempting to beed by the Editor-in-Chief and
le.fr (A. Halperin), mk@
All rights reserved.
TLCST z 32 �C. The cells spontaneously detach upon lowering thetemperature T to below the LCST and efficient harvesting is typi-cally achieved at T ¼ 20 �C. The current understanding of theseeffects involves two postulates [4,7e9]: (i) Both cell spreading at37 �C and cell retraction at T < TLCST are metabolically activeprocesses involving integrin mediated environmental sensing. (ii)The thermal switch between cell adhesive and cell detaching statesis related to change in PNIPAM hydration. It is often discussed interms of hydrophobicity/hydrophilicity as characterized by contactangle measurement. The mechanism allowing the cells to detectthe hydration state of PNIPAM and react to it is currently unspec-ified. In the following we present theoretical considerations sug-gesting such mechanism focusing on the swelling degree asa measure for hydration. We further explore the mechanism’sdependence on the structural parameters of the PNIPAM layers andsuggest guidelines for optimizing brush performance allowing forcell adhesion/proliferation as well as efficient detachment.
Studies of PNIPAM based temperature-responsive cell culturesubstrates utilized diverse structures including PNIPAM co-adsorbed with collagen [10], surface hydrogel layers cross linkedby electron beam (e-b) irradiation [6,11], plasma polymerizedlayers [12e15], brushes of terminally anchored chains [16e20] and

d cop
y
o no
t perm
itted
Fn chain
cell
5 μm
50 n
m
PNIPAM brush
50 μm
150 nm
25 n
m5
μm
Integrin
cell membraneFA
a
b
c
I
Fig. 1. A schematic view of a cell adhering via integrin bonds to ECM proteins adsorbedat the grafting surface of a PNIPAM brush. It depicts views at different length scales (a)-(c) at 37 �C for brushes with hdry < HFA z 25 nm such that there is no brushconfinement by the ventral membrane even at the focal adhesion (FA) regions.
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e49874976
Author
provid
e
Redist
ributi
microgels [21]. For concreteness, our discussion focuses on two ofthese system: e-b hydrogels and brushes. In particular we aim torationalize the following observations: (1) e-b hydrogels andbrushes with dry thickness hdry T 30 nm do not support celladhesion in serum containingmedium [22,23] while thin hydrogelswith hdry ( 15 nm do not promote cell detachment [16]. (2) Celladhesion to e-b PNIPAM hydrogels in serum free medium improveswith increasing the amount of randomly attached RGD peptideswhereas the corresponding detachment efficiency decreases[24,25]. (3) Cell adhesion to PNIPAM brushes in serum containingmedium decreases with increasing hdry while the detachmentefficiency grows with hdry [16]. Similarly, cell adhesion increases asthe polymerization degree N decreases and as the area per chain S
increases whereas the efficiency of cell detachment exhibits theopposite trend [17] (Appendix A).
Our theoretical analysis is implemented in detail to the case ofcell culture on PNIPAM brushes in serum containing medium. Thefocus on brushes is warranted by the singular balance of physialand cell culture studies concerning this system. The emergingphysical understanding of PNIPAM brushes [26] benefits froma backdrop of extensive research concerning brushes in general[27,28]. Their structure [29e31], physical properties [32e35] andtheory [36,37] were already explored. These studies indicate thatthe brush properties are determined by three parameters: thepolymerization degree N, the substrate area per chain, S, and thetemperature, T. In turn, detailed experimental results on the effectof N, S, and T on cell culture [17,20] are now available thus settingthe ground for theory efforts concerning this system. Our analysis isqualitatively applicable to e-b hydrogels to the extent they may beconsidered as weakly crosslinked brushes.
The starting point of our discussion is the current view that twoprocesses occur when cells in serum containing medium adhere toa solid substrate: First, serum borne ECM proteins, Fibronectin (FN)and Vitronectin (VN), adsorb onto the surface and subsequentlycells bind to the adsorbed ECM proteins [38]. Our analysis is basedon the hypothesis that ECM proteins undergo “primary” adsorptionat the grafting surface thus providing ligands to integrin receptorsanchored at the cell ventral membrane. While other adsorptionmodes are conceivable [37] we focus on the primary adsorptionhypothesis because it allows to rationalize the experimentalobservations noted earlier. Within this picture, the brush modifiesthe two-stage cell adhesion process in two respects: (i) It intro-duces a free energy penalty opposing the adsorption of ECMproteins. This reduces the total amount adsorbed [37,39] as well asalters the composition of adsorbed ECM proteins i.e., the ratio of FNand VN adsorbed. This free energy penalty increases as S decreasesand is proportional to the inserted, P(z)eweighted volume of theprotein. Consequently, high grafting density decreases the overalladsorption of ECM proteins while favoring the smaller VN over thelarger FN. (ii) Formation of integrin ECM bonds imposes a distancebetween the ventral cell membrane and the grafting surface (Fig.1).The measured separation between a FN coated surface and theventral membrane in focal adhesions is z25 nm [40] reflecting thelength of the extracellular integrin tail and the thickness of theadsorbed ECM layer (see section 4 for a detailed discussion). Sincethe membrane is impenetrable to the polymer chains, cell adhesioncan confine the underlying brush thus giving rise to a disjoiningforce fcell (Fig. 2). At 37 �C when the brush is collapsed and onlyweakly compressible, this fcell prevents cell adhesion forhdry T 25 nm.When cells do adhere to thinner layers, lowering T tobelow TLCST causes brush swelling while initially maintaining thefocal adhesions formed at 37 �C. The resulting brush confinementgives rise to fcell placing the integrineECM bonds under tensionthus shortening their life time according to Bell’s law [41,42]. fcellmay also accelerate the desorption of adsorbed ECM proteins
nbound to the cell (Fig. 2). Both effects contribute to the kinetics ofcell detachment. They also provide a coupling mechanism to theenvironmental sensing mechanism of the cells [43e45] anda possible signal to active detachment. Importantly, the brushconfinement is weakened when hdry and the correspondingswollen brush height are low. This reduces fcell and thus weakensthe driving force for detachment. Consequently, a thin brush maysupport cell adhesion while performing poorly in cell detachment.These qualitative considerations apply to both sparsely seeded cellsand to confluent cell sheets. For harvesting cells sheets theyrationalize the preservation of cellecell junctions because fcell actsalong the normal to the substrate and does not generate tangentialstress. As we shall elaborate in the Discussion, somewhat modifiedconsiderations allow to qualitatively rationalize observations con-cerning e-b hydrogels functionalized with pendant RGD peptides.
A complete theoretical analysis of this system, allowing forsolvent transport and cell activity is currently unfeasible. We thusfocus on two aspects susceptible to numerical estimation: (a) Theeffect of the brush on the adsorption of VN and FN. (b) The initial fcellacting on isolated cells and the corresponding effect on the integrin-FN bond dissociation rate. In both cases we explore the dependenceon N, S and T. To this end we utilize three ingredients: (i) The selfconsistent field (SCF) theory [27,46] of PNIPAM brushes [37]incorporating the empirical mixing free energy of PNIPAM solu-tions proposed by Afroze, Nies and Berghmans (ANB) [47]. The ANBfree energy reproduces the phase boundaries of PNIPAM and itscombination with SCF theory yields brush concentration profiles insemi-quantitative agreement with experimental data [26,29e31].(ii) The height distribution of the cell ventral membrane as

ded c
opy
ot pe
rmitte
d
Fig. 2. A schematic view of the cell detachment mechanism upon lowering T. At T ¼ 37 �C the cell adheres with no brush confinement (a). Brush confinement arises upon lowering Tto below TLCST because of brush swelling underneath the adhering cell and gives rise to a disjoining force, fcell. fcell reflects the contributions of the local contributions of disjoiningpressures whose amplitude increases with the local confinement (b). fcell drives dissociation of the integrin-ECM protein bonds (c) and/or desorption of the bound ECM proteins (d).
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e4987 4977
Author
provi
Redist
ributi
oobtained from the height maps reported by Iwanaga, Braun andFromherz (IBF) [40]. The IBF data was obtained for fibroblastsadhering to FN coated silica. While the height profiles may wellvary with the cell type, its state and the substrate we assume thatthe IBF data is representative of adhering cells (see detaileddiscussion in section 4). (iii) The force dependent integrineFNdissociation rate as measured and modeled by Li, Redick, Ericksonand Moy (LREM) [48]. While the LERM expression may requirerefinement in view of recent reports of catch bond behavior at lowtension [44], it summarizes the leading features of the experi-mentally observed behavior. For concreteness, our analysis focuseson the integrineFN bonds because there is no corresponding dataon the detachment of adsorbed ECM proteins.
The ABM, IBF and LERM results provide a minimal inputnecessary to obtain fcell and its effect on cell detachment. Ouranalysis utilized these results in order to obtain concrete numericaloutcome. We should emphasize that the analysis is not specific tothis choice and it can be generalized to allow for different inputs.However, using this minimal set, for lack of alternatives, imposeslimitations on our analysis. For example, the numerical results donot allow for desorption of FN and VN nor for the compliance of thecell membrane. Despite these limitations our approach allows torationalize the observed trends of cell adhesion and detachment interms of the three parameters determining brush structure, S, Nand T. In contrast, alternative approaches invoking contact anglemeasurements do not fully account for the observed phenomena[17]. Furthermore, our results suggest experiments to confront thetheory as well as tentative guidelines for the design of cell culturesubstrates based on PNIPAM brushes.
The different dimensions of cells and proteins give rise to theirdistinctive interaction modes with brushes. The relevant polymer
n ntheory of these interactions is briefly discussed in section 2 whilethe numerical implementation is summarized in Appendix B. Thebrush effects on the “primary” adsorption of ECM proteins at thegrafting surface are analyzed in section 3. In particular, we imple-ment earlier results [37] to the case of serum borne FN and VN andaddress the consequences of their different size. In section 4 weanalyze brushecell interactions on the basis of the IBF ventralmembrane height histogram. In particular, we consider the brushconfinement at T¼ 37 �C and at T< TLCST, the corresponding fcell andthe effect on the integrin-FN dissociation rate. In the Discussion wecombine the results of sections 3 and 4 to formulate designguidelines for PNIPAM brushes specifying the range of N and S thatoptimizes adhesion/proliferation at T ¼ 37 �C and detachment atT < TLCST. We also outline qualitative extension of the theory toaddress e-b hydrogels and the effect of pendant RGD peptides. Toconclude we comment on the possible role of “ternary” adsorptionwithin the brush itself and on the limitations of the theory as wellas propose experiments to confront the model and facilitate futuretheory efforts.
2. Brush-particle interactions: ecm proteins vs. cells
Chains in brushes comprising crowded terminally anchoredpolymers are stretched along the normal to the grafting surface.Their height and structure reflect a local balance between theosmotic pressure of the monomers and the average chain tensionper unit area [27,28]. The brush structure is specified by themonomer concentration c(z) at height z or the correspondingvolume fraction f(z) ¼ c(z)a3 of monomers of size a. Brushes ofneutral water soluble polymers (NWSP) affect both the amount ofadsorbed ECM proteins and their accessibility to adhering cells. The

o
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e49874978
hor p
rovide
ributi
underlying mechanisms involve two different scenarios ofparticleebrush interactions whose applicability is determined bythe particle size relative to the equilibrium height of the unconfinedbrush, h(T) (Fig. 3) [39,49,50]. Large particles can only approach thegrafting surface by compressing the brush. On the other hand, theinsertion of small particles into the brush causes only localperturbation in the monomer concentration profile. In physicalterms the second, “insertive”mode is realized when the trajectoriesof the chains can circumvent the particle. The “compressive” modefor spherical particles of radius R corresponds to R[ h(T) while theinsertive one is realized for R � h(T). For cylinders of radius r andlength L the two limits are specified by r� h(T) and r[ h(T). Cells,with spans in the range of 10 mm�60 mm, can only approach thesurface via the compressive mode. In contrast, proteinebrushinteractions may be usefully discussed in terms of the insertivemode.
The amount of adsorbed proteins is reduced because of the freeenergy penalty, Fins, incurred upon inserting the protein into thebrush. Fins reflects the work expended against the osmotic pressureof the unperturbed brush. In particular, bringing a protein intograzing contact with the grafting surface of an extended brush isassociated with
FinszP0Vp: (1)
Here P0 ¼ P(f(0)) is the osmotic pressure at the graftingsurface, at altitude z ¼ 0 where the monomer volume fraction isf(0), and the Vp is the volume of protein p: Vp ¼ 4pR3/3 fora globular protein and Vp ¼ pr2L for a cylindrical protein. While eq(1) is an approximation it is essentially exact for proteins fullyinserted into thick brushes [37].
The accessibility of the adsorbed ECM proteins to the cell isaffected by the brush because the cell ventral membrane isimpenetrable to chains. Since the formation of focal adhesionimposes a local membrane-substrate separation of z25 nm, celladhesion may lead to brush confinement with an associated dis-joining force. This effect is determined by the height map of theventral membrane H(x, y) at lateral position x, y. It identifies theconfined regions where H(x, y) < h(T) and specifies the localconfinement degree H(x, y)/h(T). The disjoining pressure P(H) dueto brush confinement by an infinite planar piston placed at H< h(T)is [50,51]
PðHÞ ¼ PðfðHÞÞ (2)
P(H) is thus identical to the osmotic pressure at the outer edge ofthe compressed brush, at z ¼ H < h(T) where the monomer volume
Aut
Redist
insertive mode
h
compressive mode
H
h
a
b
Fig. 3. A schematic view comparing the (a) compressive mode characteristic of cellsand the (b) insertive mode often realized by proteins.
d cop
y
n not
perm
itted
fraction is f(H). In turn, P(f(H)) is obtainable via the SCF theorythat yields the complete monomer volume fraction profile f(z) asa function of altitude z for confines as well as free brushes(Appendix B). So long as H(x, y) varies slowly on the scale of h(T),the disjoining force experienced by the cell, fcell, may be estimatedby the Derjaguin approximation i.e., by summing up the contri-butions due to area elements of certain H assuming that each onelocally imposes P(H) of a planar piston. Altogether, fcell for a cellwith a projected area Aproj is
fcell ¼ Aproj
ZN
0
pventralðHÞPðHÞdH (3)
where pventralðHÞ is the height probability distribution density ofthe ventral membrane. Note that this expression accounts only forcompressed regions with H < h(T) because the osmotic pressurevanishes at the edge of a unconfined brush, P(f(h)) ¼ 0, andbeyond.
The discussion as presented above applies to NWSP brushes ingeneral. The distinctive features of PNIPAM brushes are associatedwith their behavior in the vicinity of TLCST. They are evident fromthe S and T dependence of the reduced height h(T)/Na and P0 asobtained from the SCF theory of brushes combined with the ANBfree energy (Figs. 4,5). In common with all brushes, h(T) and P0
increase with the grafting density, as S decreases. In distinction tothe “classical” scenarios, h(T) and P0 of PNIPAM brushes decreasewith increasing T. Furthermore, the T dependence is steep witha sharp fall around T ¼ 32 �C followed by a plateau at higher T. Inour analysis h(T) andP0 quantify the swelling of the brush and thusthe hydration of the chains. The numerical implementation of theSCF theory and the ANB free energy utilized are summarized inAppendix B. The effects on ECM adsorption and cell adhesion/detachment are discussed in the following two sections.
3. ECM Protein adsorption
Protein adsorption onto brush displaying surfaces may involvedifferent modes distinguished by the adsorption altitude and thenature of the driving attraction [37,39]. In particular, it is useful todistinguish between (Fig. 6): (i) Primary adsorption at the graftingsurface due to surface-protein attraction. (ii) Secondary adsorption
Fig. 4. The reduced equilibrium brush height h(T)/N as a function of temperature T andarea per chain S as obtained from SCF theory of PNIPAM brushes utilizing the ANB fint(Appendix B).

d
Fig. 5. The osmotic pressure at the grafting surface of PNIPAM brushes, P0, as functionof the area per chain S and the temperature T as obtained from SCF theory of PNIPAMbrushes.
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e4987 4979
Author
provid
e
edist
ributi
o
at the outer edge of the brush thus avoiding the insertion penalty. Itmay arise because of van der Waals attraction to the graftingsurface. For collapsed brushes secondary adsorption may resultfrom contact attraction between the proteins and the outerboundary of the dense brush. (iii) Ternary adsorption within thebrush itself due tomonomer-protein attraction. In the following wefocus on the primary adsorption scenario because it allows torationalize hdry effects on the cell adhesion and detachment. Wewill comment on the role of ternary adsorption of ECM proteins inthe Discussion.
Among the different scenarios noted above, primary adsorptionis the counterpart of adsorption onto a bare surface. Both are drivenby surface-protein attraction. The two differ because primaryadsorption also incurs an extra free energy penalty Fins due to thebrush. Fins affects both the equilibrium adsorption isotherm and therate of adsorption. The brush effect is clearest when consideringadsorption out of a single component protein solution. Theadsorption isotherm specifies the adsorbed amount per unit area Gfor a surface at equilibrium with a single component bulk proteinsolution of molar concentration cbp. In the dilute surface limit, whenthe adsorbed proteins do not interact with each other
G ¼ cbpK0pexpð � Fins=kBTÞhcbpKp (4)
Here K0p characterizes the adsorption of protein p on a “brush-
less” surface of identical structure and composition. The rate ofadsorption is reduced because the proteins must traverse an acti-vation barrier [39] due to the brush leading to
adsorption rate ¼ cbpk0pexpð � Fins=kBTÞ (5)
RA
B
C
Fig. 6. A schematic view of three protein adsorption modes: (a) primary adsorption atthe grafting surface, (b) ternary adsorption within the brush and (c) secondaryadsorption at the brush outer edge.
copy
n not
perm
itted
where k0p is the adsorption rate constant for the bare surface. Thus,while the ECM adsorption may be controlled either thermody-namically or kinetically, it is reduced by a factor of exp(�Fins/kBT)due to the brush for both cases. A simple approximation for Fins ispossible for extended brushes such that the protein is fully insertedand the monomer concentration profile experienced by the proteinis f(z) z f(0) z const’. In this limit Fins z P0Vp is essentially exactwhile for thinner brushes this is an upper bound allowing torecover leading trends. The value of Fins z P0Vp depends on thebrush viaP0 and on the protein via Vp. As we discussed earlier,P0 istuned via S and T and P0 ¼ P0(S,T) for PNIPAM brushes modeledusing the ANB free energy is depicted in Fig. 5.
The quantitative results, reflecting the exp(�P0Vp/kBT) factor,depend on the volume of the specific protein, Vp. Two ECM proteinsmay play a role in cell adhesion in serum containing culturemedium: FN (molecular weight MW z 500 kDa) and VN(MW z 75 kDa) whose molar concentrations in plasma arerespectively cFN z 1.2 mM [52] and cVN z 2.66e5.33 mM [53].Electron microscopy suggests that VN in its monomeric form isglobular with a diameter of dVN ¼ 6e8 nm [54] while FN in itsextended state is long and thin strand 160 nm in length havingan average diameter of dFN ¼ 2 nm. In turn, these valuessuggest the estimates VVN z 180 nm3 and VFN z 500 nm3.KVN=K0
VN ¼ expð�P0VVN=kBTÞ and KFN=K0FN ¼ expð�P0VFN=kBTÞ
as function of S and T are depicted in Fig. 7. In both cases adsorptionat 37 �C is much higher than at 20 �C and is strongly repressed forlower S. However the effect on FN is stronger because of its largervolume. Accordingly, primary adsorption onto brush coatedsurfaces tends to favor VN in comparison to FN. In any case, topromote cell adhesion/proliferation at 37 �C it is preferable toutilize brushes with ST 30 nm2 when Finsz kBT and GVN as well asGFN are only weakly reduced. Note also that the equilibrium GVNand GFN are strongly reduced upon lowering T below TLCST. Theresulting desorption of VN and FN may contribute to cell detach-ment. However, this last effect depends on the desorption rate, yetto be explored. We should add that the effect of the adsorbed ECMproteins on cell adhesion depends on additional effects, beyond thesimple theory considered above: (i) unknown K0
p traceable to theinteractions of VN and FN with the bare surface. (ii) The confor-mation of the adsorbed ECM protein and the accessibility of theRGD domains. (iii) Competitive adsorptionwith “non-ECM” plasmaproteins. All three effects depend on the composition and structureof the grafting surface.
4. Cell Adhesion and detachment
Within our model, cells sense the hydration state of the brushvia the disjoining force fcell arising because of brush confinement bythe impenetrable ventral membrane of adhering cells anchored viaintegrins binding to adsorbed ECM proteins. In turn, the instanta-neous fcell is determined by the height map of the ventralmembrane, H(x, y), which specifies the local height H as it varieswith the in-plane position specified by x, y. The local confinementand the resulting disjoining pressure are determined by the H(x, y)/h(T) ratio. To judge themerits of this picture it is necessary to utilizerealistic numerical values of H(x, y) in obtaining the associatedtension dependent dissociation of the integrin bonds.
4.1. Topography of the ventral membrane and brush confinement
The key input required to this end are topographical mapsspecifying the absolute height of ventral membrane so as to enablecomparison with h(T). In the absence of data regarding timedependence we base our analysis on two complementary sets ofdata describing instantaneous snapshots H(x, y). Detailed H(x, y) of

o
itted
Fig. 7. The effect of the osmotic insertion penalty Fins on the adsorption of (a) Fibronectin (FN) and (b) Vitronectin (VN) as characterized by plots of log10(Kp /K0p ) ¼ �P0Vp/kBT vs. the
area per chain S and T. In the range depicted FN adsorption is reduced for all T while VN adsorption is not affected for sparse brushes at T ¼ 37�C.
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e49874980
Author
provid
e
Redist
ributi
fibroblasts adhering to hydrophobized silica coated with FN wereobtained using fluorescense interference contrast microscopy [40]of cellular membranes labeled with a fluorescent dye. These IBFH(x, y) maps describe five typical 5 mm � 5 mm regions within threedifferent cell with lateral resolution z 400 nm and height resolu-tion of z 1 nm. The corresponding distribution of vinculin wasmonitored by using cells expressing GFP-vinculin. A second study,utilizing transmission electron microscopy (TEM), provides onedimensional cross sections H(x) of human embryonic kidneyHEK293 cells cultured on SiO2 coated by various proteins includingFN [55]. Each cross section corresponds to a different cell, andcombining these data yields an average height histogram withlateral and vertical resolution ofz 2 nm. The two sets of data differin two respects: (i) The TEM study indicates that regions of0 nm�H� 10 nm account forz 20% of the histogramwhile the IBFdata reports a minimal Hmin z 15 nm. This discrepancy may beattributed to the TEM sample preparation that requires detachmentof the SiO2 substrate with a possible effect on the protein layer. (ii)The maximal height, Hmax, in the TEM study reachedHmax z 750 nmwhile the IBF data Hmax z 100 nm is much smaller.This second difference is due to contributions from the cellperiphery that are not included in the IBF data. With these caveats,the average height as determined by TEM for FN coated surface ishHiz 55 nm and comparable to the IBF data discussed below.Importantly, the superior lateral resolution of the TEM dataconfirms that H(x) varies smoothly thus justifying the use of theDerjaguin approximation.With these observations inmindwe baseour subsequent discussion on the IBF data because the topo-graphical H(x, y) map, combined with the corresponding vinculindistribution, facilitates its interpretation.
The following six features characterize the IBF data:(i) An average hHiz 50 nm, (ii) Upward bulges extending toHmax z 100 nm, (iii) No correlation between the vinculin and theheight distribution, (iv) A prominent vinculin spot associated withHvin z 25 nm, (v) Small regions of Hmin z 15 nm not correlatedwith vinculin expression, (vi) H(x, y) is slowly varying and isessentially constant over length scales of z500 nm. Among thesefeatures Hvin z 25 nm is of special significance because of threeadditional observations. First, stringent identification of focaladhesions ideally requires co-staining for integrins to evidenceclustering, noting that different integrins may be involved. Whilevinculin staining is considered as a necessary sign for focal adhe-sions it can occur without co-staining of integrins. Second, recent
d cop
y
n not
permneutron reflectometry studies demonstrated that FN adsorbed onto
silica forms a highly hydrated 4e5 nm thick layer [56]. Similarresults obtained by imaging ellipsometry suggest a hydrated layerof adsorbed FN 4.6 � 1.9 nm in thickness [55]. Finally, the length ofthe integrin extracellular domain is lint z 20 nm [57,58]. Takentogether, these observations suggest that focal adhesions (FA)correspond toH reflecting the sum of lintz 20 nm and the height ofthe adsorbed ECM protein layer. Allowing for certain upwardsbuckling of the adsorbed FN chains this argument implies thatHFA z 28�5 nm is an appropriate topographical diagnostic of a FA.
Two observations place the IBF H(x, y) picture in perspective inthe context of PNIPAM based tissue culture substrates. First, hdry THFA correlates to repression of cell adhesion. Second, high N PNI-PAM brushes with hdry z 10 nm retain h(37 �C) z hdry in theircollapsed state while swelling to h(20 �C)[ h(37 �C) when cooledto T¼ 20 �C [34]. We emphasize that the swelling/collapse behaviorof PNIPAM brushes can depend on N, an issue considered in theDiscussion. With this caveat, comparison of h(T) and H(x, y)suggests the occurrence of three broad regimes: (i)HFA < h(37 �C) < h(T < TLCST) and the adhering cell always confinesthe brush irrespective of T and cell adhesion is repressed altogether.(ii) h(37 �C) < HFA ( h(T < TLCST) and the cell confines the swollenbrush below TLCST but not in its collapsed state, above the TLCST. Inthis range cells will adhere at 37 �C and detach upon cooling toT < TLCST. (iii) h(37 �C) < h(T < TLCST) < HFA and there is no brushconfinement due to adhering cells irrespective of T, a situationfavoring cell adherence while repressing cell detachment.
4.2. The disjoining pressure and fcell
The occurrence of the brush confinement regimes, as discussedabove, is a necessary condition for the viability of our mechanism. Itis next important to estimate the magnitude of the associated fcelland the tension per bound integrin to establish that they caninfluence cell adhesion. To this end we assume that the IBF H(x, y)data are representative and extract from it the probability densityof ventral membrane height, pventral(H) (Fig. 8). In doing so weignore negligible contribution due to H(x, y) � 22 nm. Noting theslow variation ofH(x, y) we are in a position to invoke the Derjaguinapproximation and obtain the histograms of forces acting on thecells, Aproj pventral(H)P(H)ΔH, and the corresponding fcell by using eqs(2) and (3). Note that in invoking the Derjaguin approximation inthis form we introduce an assumption that the brush relaxation is

d
0 20 40 60 80 1000
0.01
0.02
0.03
0.04
0.05
0.06
0.07
H [nm]
p ven
tral(H
) [n
m−1]
H(x,y)
x
y
100nm
75nm
50nm
25nm
Fig. 8. The height histogram pventral(H) of the cell’s ventral membrane as obtainedfrom the height maps H(x, y) of Iwanaga, Braun and Fromherz (IBF) [40], shown asinsets. The empty bars were omitted from the calculations concerning cells adhering toPNIPAM brushes. Insets reproduced with permission.
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e4987 4981
Author
provid
e
Redist
ributi
o
faster than that of the cell membrane i.e., P(H) attains its equilib-rium value subject to a fixed H(x, y) and pventral(H). This roughapproximation is tenable at a very short interval following a quasiinstantaneous change of T. It is somewhat justified by reports of fastrelaxation in the thickness of brushes, with time scales in the rangeof millisecond to seconds [59,60]. However, this approach over-estimates fcell since the cell will deform when subjected to dis-joining pressure thus lowering fcell. With these reservations inmind, the plots of fcell vs 10 nm2� S� 50 nm2 and 10 �C� T� 40 �Cas obtained for a typical projected cell area Aproj z 2000 mm2
(Fig. 9) reveal three relevant features: (i) fcell z 0 at T ¼ 37 �C whenhdry ( HFA, (ii) fcell increases as T is lowered below TLCST, (iii) Theincrease in fcell is steeper for higher N. The maximal fcell isz5 � 105 pN for N ¼ 200 and reaches 109 pN for N ¼ 1000. Thuswhile cell compliance will lower fcell, it is sufficient to drive celldetachment. For comparison,measured cell pulling strength vary inthe range 1 pN < fcell < 1.5 � 106 pN [44].
4.3. The Effect on the dissociation rate of integrineFN bonds
The immediate effect of fcell on cell detachment may result fromdesorption of adsorbed ECM proteins or accelerated dissociation ofintegrin-ECM protein bonds. The effect of the tension on thedesorption of adsorbed ECM proteins is difficult to estimate in theabsence of force spectroscopy data on the subject. This is also thecase for integrin-VN dissociation. We thus consider the effect of fcellon the dissociation of integrin-FN bonds assuming that there is nodesorption of ECM proteins. In particular, we analyze the initialbond dissociation rate for an adhering cell anchored via m0integrin-FN bonds at the instant fcell increases upon lowering Tfrom 37�C to T < TLCST. To this end we argue that fcell is equallypartitioned among m0 bonds loaded in parallel [42]. In this situa-tion the effective initial rate of dissociation for a single barrierenergy landscape is given by keff(m0) ¼ m0k0 exp(fcellxb/m0kBT)where xb is the position of the barrier as measured from theminimum of the well and k0 ¼ s�1 exp(�Eb/kBT) is the dissociationrate constant of the unloaded bond. k0 is defined in terms of theheight of the activation barrier, Eb, and a characteristic attempt times. The exp(fcellxb/m0kBT) factor, reflecting Bell’s law [41], accountsfor the effect of the tension on the barrier height. The m0 prefactorallows for the number of possible ways of breaking a bond. The
copy
n not
perm
itted
room temperature force spectroscopy measurement of LREM [48]suggest however that the integrin-FN bond energy landscapeinvolves two barriers at xb(1) z 0.09 nm and xb(2) z 0.42 nmwithbarrier heights of Eb(1)z 11.7 kBT and Eb(2)z 17.3 kBT as measuredwith respect to the fundamental minimum of the potential well.The corresponding dissociation energy is 18.8 kBT. Their results forsingle bond dissociation are described by keff
�1 ¼ P2j¼1 k0
�1(j) exp[�fcellxb(j)/kBT ] with room T values of k0(1) ¼ s�1 exp(�Eb(1)/kBT) ¼ 33.5sec�1 and k0(2) ¼ s�1 exp(�Eb(2)/kBT) ¼ 0.13sec�1. TheLERM experiment involved varying the applied force at a fixed T sothat k0(1) and k0(2) are constants. In contrast, in our situation fcell istuned by lowering T thus affecting both k0 and the exp(fcellxb/m0kBT) factors. Altogether, the dissociation rate upon lowering T tobelow TLCST is specified by keff(m0) given by
1keff ðm0Þ
¼ sm0
exp�Ebð1ÞkBT
� fcellxbð1Þm0kBT
�
þ sm0
exp�Ebð2ÞkBT
� fcellxbð2Þm0kBT
� (6)
keff(m0) thus strongly depends on the number of bound integrins,m0. In turn m0 is determined by the density of bound integrins ina FA, sINT, times the total area occupied by FA. For sINT we utilizesINT ¼ 300/mm2 as proposed by Moore et al. [44] on the basis ofexperimental data of Arnold et al. [61]. The IBF data indicates thatFAs as defined by HFA z 28�5 nm account for z 10% of the pro-jected cell area, Aproj. For a cell with Aproj z 2000 mm2 the IBF datasuggests z60 FA having an average area of 3 mm2 and thus leadingto m0 z 5 � 104. The average area per FA as specified by thisargument is comparable to the experimentally measured value of5.5 � 2.9 mm2 reported by Arnold et al. [61].
Because of the exp(fcellxb/m0kBT) factor the keff(m0) landscape(Fig. 10) accentuates the features of the fcell plots leading to steepervariation. We distinguish again between three qualitative regimes:(i) No amplification above TLCST, (ii) Onset of amplification atTz TLCST, (iii) For T< TLCST the amplification grows with decreasingS and T and with increasing N. In the 10 nm2 � S � 50 nm2 and10 �C� T� 40 �C range the maximal amplification is of order of 102
for N ¼ 200 and reaches z 106 for N ¼ 1000. While the approxi-mations invoked may lead to overestimate of keff, the trends are inagreement with experimental results and suggest that fcellcontributes to cell detachment.
5. Discussion
Brush effects on cell adhesion/detachment vary with the densityand altitude of integrin ligands within the brush. Integrin ligandssuch as RGD peptides, can be chemically linked to the chains. In thiscase their density and height distribution are determined by thesynthesis. Alternatively, integrin ligands can be introduced viaadsorption of ECM proteins. In this second case it is helpful todistinguish between primary adsorption at the grafting surface andternary adsorption within the brush itself. The relative importanceof the two adsorption modes depends on protein-surface interac-tions specific to the substrate utilized. For brevity, our analysisfocused on cell adhesion from serum containing medium assumingthat the ECM proteins undergo primary adsorption at the graftingsurface. This scenario is of interest since the majority of experi-ments utilize serum containing culture medium and because itallows to rationalize hdry effects on adhesion and detachment. Inthis situation the brush tunes the cell adhesion/detachmentbehavior via two effects: (i) It affects the concentration of adsorbedECM proteins and thus the maximum density of integrin ligandsavailable to the cells. Proteins can only contact the surface followinginsertion into the brush thus incurring a free energy penalty,

Author
provid
ed co
py
Redist
ributi
on no
t perm
itted
Fig. 9. The disjoining force, fcell, experienced by cell having a projected areaAproj ¼ 2000 mm2 as calculated using the SCF theory of PNIPAM brushes and the IBFheight histogram depicting its variation with the area per chain
Pand T for N ¼ 200,
300, 500, and 1000.
Fig. 10. The amplification of the integrineFN dissociation by the disjoining force log10keff(fcell) calculated using equation (6) as function of the area per chain
Pand T for
N ¼ 200, 300, 500, and 1000.
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e49874982
d
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e4987 4983
r prov
ide
utio
Fins z P0Vp, reflecting the work done against its osmotic pressure.P0 increases as S and T decrease. Adsorption at T ¼ 37 �C is thushigher than at T < TLCST but can be repressed altogether for small S.For fully inserted proteins Fins w Vp and since VFN >VVN, FN incursa higher Fins thus favoring adsorption of VN. The FN/VN ratio isultimately determined by Fins together with the surface-proteincontact energy and thus substrate dependent. (ii) Once the celladheres to the surface via integrin bonds to the adsorbed ECMproteins, it presents an impenetrable ventral membrane to theunderlying brush. Within this view, cells adhere at 37 �C when thecollapsed brush lies below the ventral membrane and there is nobrush compression. Upon lowering T the brush swells and isconfined by the anchored ventral membrane giving rise to a dis-joining force fcell. In turn fcell places the integrin-ECM bonds undertension thus shortening their life time according to Bell’s law and,in addition, accelerating the desorption of the adsorbed ECMproteins. These two processes directly contribute to the kinetics ofcell detachment by lowering the numbers of tethers anchoring thecell. They also provide a possible signal for triggering active celldetachment via the integrin mediated environmental sensing ofthe cells. Our numerical results concerned the effect of fcell on thedissociation rate of integrin-FN bonds. However, as noted earlier,fcell can also lead to accelerated desorption of adsorbed ECMproteins. In this context it is useful to note that FN adsorbed on e-bhydrogels at 37 �C does not detach at room T even after prolongedincubation in 0.5 M SDS or 0.5 M SDSþ 8M urea [62]. This suggestsstrong adsorption such that a change inP0 and Fins does not lead todesorption. On the other hand there is evidence suggesting cellmediated detachment of FN. In particular, the appearance of FN freeregions, similar in size and shape to adhering cells, followingdetachment by low T treatment [62]. These observations wereinterpreted as evidence for the involvement of metabolically activeprocess but may reflect FN desorption driven by the fcell asproposed in our model. We should emphasize that our discussionconcerns only dimeric FN and monomeric VN as present in theserum. It does not concern endogenous ECM proteins whichpresumably reside outside the brush and detach with the cells.
5.1. Design guidelines
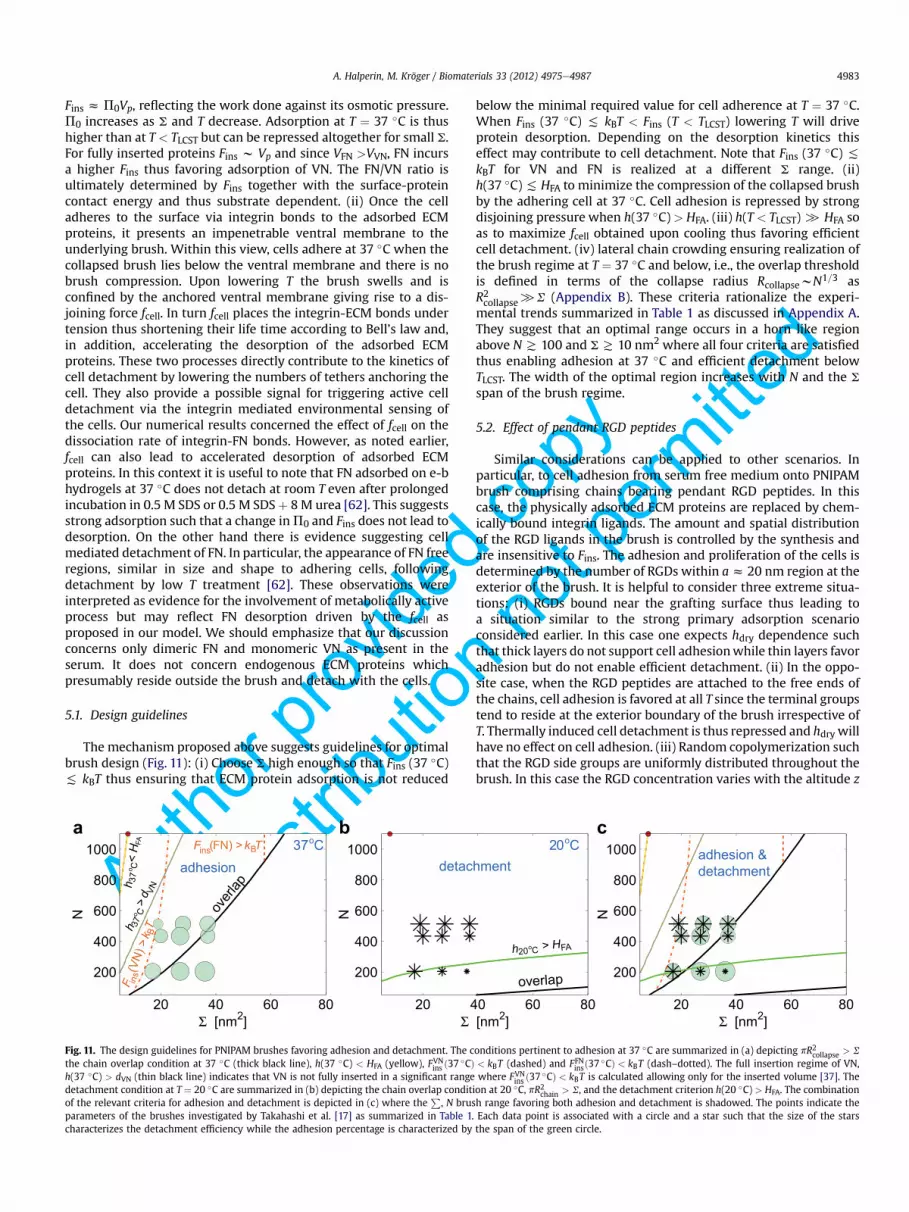
The mechanism proposed above suggests guidelines for optimalbrush design (Fig. 11): (i) Choose S high enough so that Fins (37 �C)( kBT thus ensuring that ECM protein adsorption is not reduced
Autho
Redist
rib
Σ
N
20 4
200
400
600
800
1000
Σ [nm2]
N
20 40 60 80
200
400
600
800
1000
h37
o C <
H
FAh37
o C > d
VN
Fins(FN) > kBT
detacadhesion
overl
ap
37oC
Fin
s(VN
) > k BT
ba
TT
Fig. 11. The design guidelines for PNIPAM brushes favoring adhesion and detachment. The cthe chain overlap condition at 37 �C (thick black line), h(37 �C) < HFA (yellow), FVNins ð37 �CÞh(37 �C) > dVN (thin black line) indicates that VN is not fully inserted in a significant rangedetachment condition at T ¼ 20 �C are summarized in (b) depicting the chain overlap conditof the relevant criteria for adhesion and detachment is depicted in (c) where the
P, N brus
parameters of the brushes investigated by Takahashi et al. [17] as summarized in Table 1.characterizes the detachment efficiency while the adhesion percentage is characterized by
copy
n not
perm
itted
below the minimal required value for cell adherence at T ¼ 37 �C.When Fins (37 �C) ( kBT < Fins (T < TLCST) lowering T will driveprotein desorption. Depending on the desorption kinetics thiseffect may contribute to cell detachment. Note that Fins (37 �C) (kBT for VN and FN is realized at a different S range. (ii)h(37 �C)( HFA to minimize the compression of the collapsed brushby the adhering cell at 37 �C. Cell adhesion is repressed by strongdisjoining pressure when h(37 �C) > HFA. (iii) h(T < TLCST)[ HFA soas to maximize fcell obtained upon cooling thus favoring efficientcell detachment. (iv) lateral chain crowding ensuring realization ofthe brush regime at T ¼ 37 �C and below, i.e., the overlap thresholdis defined in terms of the collapse radius RcollapsewN1=3 asR2collapse[S (Appendix B). These criteria rationalize the experi-mental trends summarized in Table 1 as discussed in Appendix A.They suggest that an optimal range occurs in a horn like regionabove N T 100 and ST 10 nm2 where all four criteria are satisfiedthus enabling adhesion at 37 �C and efficient detachment belowTLCST. The width of the optimal region increases with N and the S
span of the brush regime.
5.2. Effect of pendant RGD peptides
Similar considerations can be applied to other scenarios. Inparticular, to cell adhesion from serum free medium onto PNIPAMbrush comprising chains bearing pendant RGD peptides. In thiscase, the physically adsorbed ECM proteins are replaced by chem-ically bound integrin ligands. The amount and spatial distributionof the RGD ligands in the brush is controlled by the synthesis andare insensitive to Fins. The adhesion and proliferation of the cells isdetermined by the number of RGDs within az 20 nm region at theexterior of the brush. It is helpful to consider three extreme situa-tions: (i) RGDs bound near the grafting surface thus leading toa situation similar to the strong primary adsorption scenarioconsidered earlier. In this case one expects hdry dependence suchthat thick layers do not support cell adhesionwhile thin layers favoradhesion but do not enable efficient detachment. (ii) In the oppo-site case, when the RGD peptides are attached to the free ends ofthe chains, cell adhesion is favored at all T since the terminal groupstend to reside at the exterior boundary of the brush irrespective ofT. Thermally induced cell detachment is thus repressed and hdry willhave no effect on cell adhesion. (iii) Random copolymerization suchthat the RGD side groups are uniformly distributed throughout thebrush. In this case the RGD concentration varies with the altitude z
Σ [nm2]
N
20 40 60 80
200
400
600
800
1000
[nm2]0 60 80
h20oC > HFA
hment
overlap
02 oCadhesion &detachment
c
onditions pertinent to adhesion at 37 �C are summarized in (a) depicting pR2collapse > S
< kBT (dashed) and FFNinsð37 �CÞ < kBT (dash–dotted). The full insertion regime of VN,where FVNins ð37 �CÞ < kBT is calculated allowing only for the inserted volume [37]. The
ion at 20 �C, pR2chain > S, and the detachment criterion h(20 �C) > HFA. The combinationh range favoring both adhesion and detachment is shadowed. The points indicate theEach data point is associated with a circle and a star such that the size of the starsthe span of the green circle.

o
d
Table 1The grafted amount G, and N as obtained from tables 2 and 3 of Takahashi et al. [17]while the fraction of adhering cells (AC) was extracted from their Fig. 4, whichspecifies error bars. AC37
�C24h is specified with respect to the number of adhering cells
on the initiator surface after 24 h incubation at 37 �C in an identical culture medium.AC20
�C2h characterizes the fraction of adhering cells after 2 h incubation at 20 �C. The
remaining quantities were calculated using the PNIPAM parameters a ¼ 0.5 nm,p ¼ 1, r ¼ 1.1 g/cm3, M0 ¼ 113 g/mol and the relationships hdry ¼ G/r, R20 ¼ Npa2,2pR3collapsed=3 ¼M0N=r, i.e., Rcollapsedz0:43� N1=3 nm, S ¼ MW/G, s ¼ a2/S. Thedetachment efficiency is defined by DE¼ 1� AC20
�C2h =AC37
�C24h . The G values marked by
# and ## are below the detection limit. Following Takahashi et al. they were replacedby entries * and ** assuming that S is identical to that of the N ¼ 434 samples withidentical initiator densities. For * and **G and hdry are calculated using S and N.Fig. 11 depicts the * and ** values.
MW G N hdry R0 Rcollapsed S AC24h37�C AC2h20�C DE
[g/mol] [mg/cm2] [nm] [nm] [nm] [nm2] [%] [%] [%]
125000 2.47 1106 22.5 16.6 4.5 8 0 0 e
54000 0.93 478 8.5 10.9 3.4 10 yes na yes58000 0.50 513 4.5 11.3 3.5 19 50 0 10058000 0.35 513 3.2 11.3 3.5 28 80 10 8858000 0.26 513 2.4 11.3 3.5 37 80 10 8849000 0.41 434 3.7 10.4 3.3 20 70 10 8649000 0.30 434 2.7 10.4 3.3 27 90 20 7849000 0.22 434 2.0 10.4 3.3 37 75 30 6023000 0.22 204 2.0 7.1 2.6 17 82 15 8223000# 0.17 204 1.5 7.1 2.6 22 92 45 5123000## 0.17 204 1.5 7.1 2.6 22 100 70 3023000* 0.14 204 1.3 7.1 2.6 27 92 45 5123000** 0.11 204 1.0 7.1 2.6 37 100 70 30
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e49874984
Author
provid
e
Redist
ributi
as xRGDc(z) where xRGD is the mole fraction of monomers with RGDside groups and c(z) is the monomer concentration profile of thebrush. For a given xRGD the average RGD concentration is higher incollapsed brushes as is the number of RGD within lint z 20 nm ofthe brush boundary. Both factors favor cell adhesion at T ¼ 37 �C.However, at high xRGD the number of accessible RGD will be suffi-cient at all T thus reducing the efficiency of thermal cell detach-ment. The performance of PNIPAM brushes with RGD pendantgroups remains to be explored. However, e-b hydrogels incorpo-rating RGD moieties were studied [24,25,63] and in this case highxRGD was indeed associatedwith poor detachment [25]. This picturesuggests that cell adhesion to RGD functionalized e-b gels at highxRGD will not depend on hdry, a prediction that remains to be tested.
5.3. The role of ternary adsorption
Yet another cell adhesion mode involves ternary adsorption ofECM proteins within the brush due to attractive interactionsbetween the proteins and the PNIPAM chains. Recent experimentsby Xue et al. [20] suggest cell adhesion mediated by ternaryadsorption of ECM proteins. The study concerned 3T3 fibroblastscultured on PNIPAM brushes anchored to a grafting surfacepassivated by non-fouling oligoethylene oxide coating thusrepressing primary adsorption. The ternary adsorption mode ofproteins and its role in mediating cell adhesion are evidenced bythe S dependence of the amount of adsorbed protein per unit area,G. In particular: (i) G(S,T) at given T exhibits a maximum at high S
for both FN and bovine serum albumin and is higher than the barepassivated surface G(S ¼ N). (ii) G(S,T) decreases strongly as T islowered below TLCST. (iii) The S and T dependence of the number ofadhering cells and their area is similar to the behavior of G(S,T). Anumber of issues merit attention in this context. For ternaryadsorption G is expected to increase with N [64]. The realization ofthis signature for ECM proteins is yet to be observed. Moreover,within the ternary adsorption scenario the proteins adsorbthroughout the brush. This suggests that cell adhesion is inde-pendent of hdry provided that S is sufficiently high to ensure lowFins and significant adsorption within lint z 20 nm from the brush
edge. This regimewas not explored by Xue et al. but such trend wasobserved by Xu et al. [18] for culture of 3T3 fibroblasts on PNIPAMbrushes of up to hdry z 31 nmwhose S and N were not specified. Itis of interest to note two additional points. First, the resultssuggestive of ternary adsorption involved immortal cell lines whilethe reports of hdry dependence concerned primary cells. Second,ECM adsorption onto a substrate such as tissue culture polystyreneserves a double role: It localized the ECM protein at the surface andalso induces exposure of cryptic integrin binding sites in VN and FN[65,66]. G on its own does not specify the biological activity of theadsorbed ECM proteins. The effect of ternary adsorption withinPNIPAM brushes on the configurations of adsorbed ECM proteinsremains to be established. Finally, the mechanism of cell detach-ment within this scenario depends on the kinetics of proteindesorption, a subject that remains to be investigated. fcell maycontribute if the desorption is slow.
d cop
y
n not
perm
itte5.4. Confrontation with experiment
In confronting experiments it is important to note a number ofissues. One concerns brush characterization. Experimental studiesof cell culture on PNIPAM brushes often utilize surface initiatedatom transfer radical polymerization (ATRP) “grafting from”
synthesis. This approach allows to produce dense and thick brushesat the price of difficulties in determining S, N and their poly-dispersity. Unambiguous determination of N requires gel perme-ation chromatography of chains detached from the substrate [32],a challenging procedure requiring samples with large surface areas.On the other hand, detailed confrontation with theory requiresknowledge of both S and N and characterization in terms of hdryalone is insufficient since identical hdry ¼ Na3=S values can berealized by different combinations of N and S.
A second group of problems concerns the modeling of PNIPAMbrushes. These were discussed in detail earlier [37]. Here we brieflyoutline the underlying assumptions and comment on two issuesrelevant to the numerical calculation presented above. Our theoryconcerns a planar, uniform grafting surface having no attraction toPNIPAM and displaying a laterally uniform and monodispersebrush with constant N and S. It does not allow for polydispersity inN and S or for chemical heterogeneity of the surface. Within thebrush the PNIPAM chains are flexible with a T independentpersistence length. The analytical SCF theory captures the leadingfeatures of brush structure. It does not account for a narrowdepletion layer at the grafting surface and for a weaker decay of themonomer concentration profile at the outer edge of the brush. Inaddition, phase boundaries within this description appear asdiscontinuities in the concentration profile. To account for thedistinctive features of PNIPAM, we utilized the ANB free energy[47]. This reproduces the phase diagram of PNIPAM as observed byANB and later confirmed by Zhou et al. [67]. Its combination withSCF theory yields concentration profiles consistent with the profilesobtained by neutron reflectivity [29,30] and SANS [31]. In partic-ular, it predicts collapse upon heating and the occurrence of verticalphase separationwithin the brush, in semi-quantitative agreementwith experimental results for high N. Note however that theapplicability regime of the ANB free energy remains to be estab-lished. It would be especially important to confront the bulkosmotic pressure, P(f,T) as obtained from the ANB free energy(Appendix B) with experimentally measured values. It is also usefulto explore alternatives to the ANB free energy. Of particular interestis the free energy obtained from molecular theory of PNIPAMsolutions proposed by Okada and Tanaka [68]. This howeverrequires additional analysis since the phenomenological parame-ters of this model were determined from a different phase diagram.
d
y itted
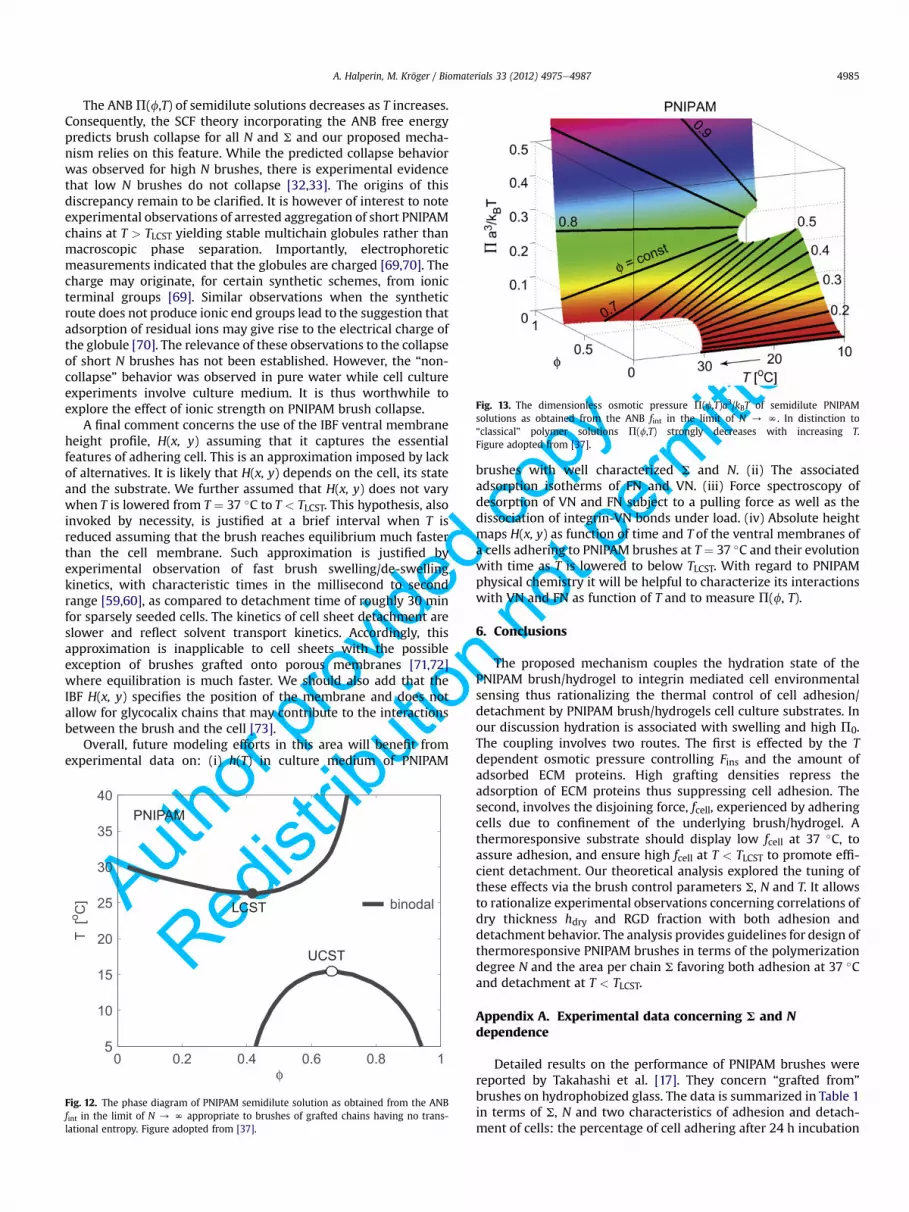
Fig. 13. The dimensionless osmotic pressure P(f,T)a3/kBT of semidilute PNIPAMsolutions as obtained from the ANB fint in the limit of N / N. In distinction to“classical” polymer solutions P(f,T) strongly decreases with increasing T.Figure adopted from [37].
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e4987 4985
prov
ide
utio
The ANBP(f,T) of semidilute solutions decreases as T increases.Consequently, the SCF theory incorporating the ANB free energypredicts brush collapse for all N and S and our proposed mecha-nism relies on this feature. While the predicted collapse behaviorwas observed for high N brushes, there is experimental evidencethat low N brushes do not collapse [32,33]. The origins of thisdiscrepancy remain to be clarified. It is however of interest to noteexperimental observations of arrested aggregation of short PNIPAMchains at T > TLCST yielding stable multichain globules rather thanmacroscopic phase separation. Importantly, electrophoreticmeasurements indicated that the globules are charged [69,70]. Thecharge may originate, for certain synthetic schemes, from ionicterminal groups [69]. Similar observations when the syntheticroute does not produce ionic end groups lead to the suggestion thatadsorption of residual ions may give rise to the electrical charge ofthe globule [70]. The relevance of these observations to the collapseof short N brushes has not been established. However, the “non-collapse” behavior was observed in pure water while cell cultureexperiments involve culture medium. It is thus worthwhile toexplore the effect of ionic strength on PNIPAM brush collapse.
A final comment concerns the use of the IBF ventral membraneheight profile, H(x, y) assuming that it captures the essentialfeatures of adhering cell. This is an approximation imposed by lackof alternatives. It is likely that H(x, y) depends on the cell, its stateand the substrate. We further assumed that H(x, y) does not varywhen T is lowered from T ¼ 37 �C to T < TLCST. This hypothesis, alsoinvoked by necessity, is justified at a brief interval when T isreduced assuming that the brush reaches equilibrium much fasterthan the cell membrane. Such approximation is justified byexperimental observation of fast brush swelling/de-swellingkinetics, with characteristic times in the millisecond to secondrange [59,60], as compared to detachment time of roughly 30 minfor sparsely seeded cells. The kinetics of cell sheet detachment areslower and reflect solvent transport kinetics. Accordingly, thisapproximation is inapplicable to cell sheets with the possibleexception of brushes grafted onto porous membranes [71,72]where equilibration is much faster. We should also add that theIBF H(x, y) specifies the position of the membrane and does notallow for glycocalix chains that may contribute to the interactionsbetween the brush and the cell [73].
Overall, future modeling efforts in this area will benefit fromexperimental data on: (i) h(T) in culture medium of PNIPAM
Author
Redist
rib
0 0.2 0.4 0.6 0.8 15
10
15
20
25
30
35
40
φ
T [o C
] binodal
UCST
LCST
PNIPAM
Fig. 12. The phase diagram of PNIPAM semidilute solution as obtained from the ANBfint in the limit of N / N appropriate to brushes of grafted chains having no trans-lational entropy. Figure adopted from [37].
cop
n not
permbrushes with well characterized S and N. (ii) The associated
adsorption isotherms of FN and VN. (iii) Force spectroscopy ofdesorption of VN and FN subject to a pulling force as well as thedissociation of integrin-VN bonds under load. (iv) Absolute heightmaps H(x, y) as function of time and T of the ventral membranes ofa cells adhering to PNIPAM brushes at T ¼ 37 �C and their evolutionwith time as T is lowered to below TLCST. With regard to PNIPAMphysical chemistry it will be helpful to characterize its interactionswith VN and FN as function of T and to measure P(f, T).
6. Conclusions
The proposed mechanism couples the hydration state of thePNIPAM brush/hydrogel to integrin mediated cell environmentalsensing thus rationalizing the thermal control of cell adhesion/detachment by PNIPAM brush/hydrogels cell culture substrates. Inour discussion hydration is associated with swelling and high P0.The coupling involves two routes. The first is effected by the Tdependent osmotic pressure controlling Fins and the amount ofadsorbed ECM proteins. High grafting densities repress theadsorption of ECM proteins thus suppressing cell adhesion. Thesecond, involves the disjoining force, fcell, experienced by adheringcells due to confinement of the underlying brush/hydrogel. Athermoresponsive substrate should display low fcell at 37 �C, toassure adhesion, and ensure high fcell at T < TLCST to promote effi-cient detachment. Our theoretical analysis explored the tuning ofthese effects via the brush control parameters S, N and T. It allowsto rationalize experimental observations concerning correlations ofdry thickness hdry and RGD fraction with both adhesion anddetachment behavior. The analysis provides guidelines for design ofthermoresponsive PNIPAM brushes in terms of the polymerizationdegree N and the area per chain S favoring both adhesion at 37 �Cand detachment at T < TLCST.
Appendix A. Experimental data concerning S and Ndependence
Detailed results on the performance of PNIPAM brushes werereported by Takahashi et al. [17]. They concern “grafted from”
brushes on hydrophobized glass. The data is summarized in Table 1in terms of S, N and two characteristics of adhesion and detach-ment of cells: the percentage of cell adhering after 24 h incubation

A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e49874986
at 37 �C, AC37�C24h , and the percentage of detached cell after 2 h
incubation at 20 �C, AC20�C2h . The percentage is taken with respect to
the number of adhering cells on the initiator surface after 24 hincubation. N was determined for the free chains produced duringthe brush synthesis. It should be emphasized that the maximal hdryin the samples listed, hdry ¼ 22.4 nm, is below the “e-b hydrogel”threshold yet does not support cell adhesion. As noted by theauthors [17] the trends cannot be explained in terms of contactangle measurements.
Comparing the data of Table 1 with the design criteria (Fig. 11)brings up a number of observations: (i) Cell adhesion on the brushdisplaying surface is lower than on the corresponding initiatorsurface. This is consistent with the assumption that primaryadsorption of ECM proteins provides integrin ligands to theadhering cells. (ii) The hdry < HFA condition is satisfied for allreportedN and S. (iii) Two of the N¼ 204 samples do not satisfy thebrush condition at 37 �C. Their behavior thus reflects the baresurface characteristics of superior cell adhesion and poor detach-ment. (iii) Overall, AC37
�C24h increases with S reflecting the corre-
sponding decrease in P0 and the enhancement of ECM proteinadsorption. The non adherence of cells to the N¼ 1106, S¼ 8 nm2 isattributed to strong repression of the adsorption of both VN and FN.(iv) Generally, DE grows with increasing N and with decreasing S.This is consistent with expectation that brush confinement by anadhering cell and the associated fcell grow with h(20 �C).
oo
Author
provid
e
Redist
ributi
Appendix B. Outline of brush structure
The brush effects on primary protein adsorption and celladhesion are mediated by Fins and fcell determined respectively byP0 and byP(f(H)) i.e., the osmotic pressure at the grafting surface,z ¼ 0, in an unperturbed brush and at the brush boundary,z ¼ H < h, in a confined brush. To calculate P0 and P(f(H)) weutilize a self consistent field theory of brushes [27]. It specifies themonomer volume fraction f(z) at altitude z and all related prop-erties including the osmotic pressure profile P(f(z)).Within theanalytical SCF theory we utilize [46] the chains are Gaussian andstrongly stretched with the free ends distributed throughout thebrush. The theory applies to laterally crowded chains when the areaper chain S is smaller than the cross sectional area of the free chain,pR2chainðTÞ as determined by its T dependent radius RchainðTÞ.
At each z the brush is considered as a semidilute polymersolution characterized by an interaction free energy density kBTfintspecific to the polymer and solvent under consideration. Irre-spective of the precise form of fint, minimization of the total freeenergy per chain subject to the constraint of a polymerizationdegree N leads to
mðfðzÞÞkBT
¼ L� 3p2
8pa2N2z2 (B1)
where p is the number of monomers in a persistent segment anda is the monomer size. In other words, the monomer exchangechemical potential within the brush m/kBT ¼ a3vfint(f(z))/vf variesparabolically with the altitude z. This, together with the normali-zation condition
S
a3
Zh
0
fðzÞdz ¼ N (B2)
specifies f(z) and h and the corresponding osmotic pressure P/kBT ¼ fvfint/vf � fint of a free brush upon imposingP(f(h)) ¼ 0. Fora confined brush, on the other hand, the upper limit of the integral
d cop
y
t perm
itted
in eq (B2) is H rather than h and the pressure P(H) h P(f(H)) isobtained rather than imposed. The results noted above are general.To describe the behavior of PNIPAM brushes in water we utilize theempirical ANB fint [47] in the limit of N / N applicable to termi-nally anchored chains having no translational entropy:
a3fintðfÞ ¼ ð1� fÞlnð1� fÞ þ ceff ðf; TÞfð1� fÞ (B3)
with ceff (f, T)¼�12.947þ 0.044959 T/Kþ 17.920f� 0.056944 fT/K þ 14.814 f2 � 0.051419 f2T/K, where T is the temperature inKelvin. The resulting f(z) and P(z) profiles of free, unconfinedbrushes are described in detail in an earlier article [37].
The ANB fint was obtained by fitting the phase boundary ofaqueous PNIPAM solutions in the 26 �C ( T ( 35 �C range asobtained from their experimental data later confirmed by Zhouet al. [67]. The applicability of this fint outside this T range remainsto be established.With this caveat in mind, the ANB fint leads to twooutcomes playing a role in our discussion: (i) A phase diagramwithboth a LCST and UCST with critical points at fLCST
c ¼ 0:413,TLCST ¼ 26.360 �C, and fUCST
c ¼ 0:661, TUCST ¼ 15.396 �C (Fig. 12).The fLCST
c ¼ 0:413 indicates a “second type of phase transition”leading a vertical phase separation within the brush associatedwith a discontinuity in f(z) in semi-quantitative agreement withexperiments. (ii) The osmotic pressure of semidilute PNIPAMsolutions, P(f,t)∕kBT ¼ fvfint/vf � fint predicted by the ANB fintdecreases as T increases, in contrast to “classical” polymer solutions(Fig. 13). Accordingly, within this picture individual chains as wellas brushes collapse upon increasing T above TLCST. Since our analysisis based on the ANB fint, it assumes that collapse occurs irrespectiveof N and the overlap threshold is thus defined in terms of the radiusof a collapsed chain, i.e., S < pR2collapsewN2=3a2.
n nReferences
[1] Freshney RI. Culture of animal cells. A manual of basic technique. 3rd ed. NewYork, USA: Wiley & Sons; 1994.
[2] da Silva RMP, Mano JF, Reis RL. Smart thermoresponsive coatings and surfacesfor tissue engineering: switching cell-material boundaries. Trends Biotechnol2007;25:577e83.
[3] Cole MA, Voelcker NH, Thissen H, Griesser HJ. Stimuli-responsive interfacesand systems for the control of protein-surface and cell-surface interactions.Biomaterials 2009;30:1827e50.
[4] Yang J, Yamato M, Okano T. Cell-sheet engineering using intelligent surfaces.MRS Bull 2005;30:189e93.
[5] Yamato M, Akiyama Y, Kobayashi J, Yang J, Kikuchi A, Okano T. Temperature-responsive cell culture surfaces for regenerative medicine with cell sheetengineering. Progr Polym Sci 2007;32:1123e33.
[6] Nagase K, Kobayashi J, Okano T. Temperature-responsive intelligent interfacesfor biomolecular separation and cell sheet engineering. J Royal Soc Interface2009;6:S293e309.
[7] Cooperstein MA, Canavan HE. Biological cell detachment from poly(N-isopropylacrylamide) and its applications. Langmuir 2010;26:7695e707.
[8] Yamato M, Okuhara M, Karikusa F, Kikuchi A, Sakurai Y, Okano T. Signaltransduction and cytoskeletal reorganization are required for cell detachmentfrom cell culture surfaces grafted with a temperature-responsive polymer.J Biomed Mater Res 1999;44:44e52.
[9] Okano T, Yamada N, Okuhara M, Sakai H, Sakurai Y. Mechanism of celldetachment from temperature-modulated, hydrophilic-hydrophobic polymersurfaces. Biomaterials 1995;16:297e303.
[10] Takezawa T, Mori Y, Yoshizato K. Cell-culture on a thermoresponsive polymersurface. Nat Biotechn 1990;8:854e6.
[11] Yamada N, Okano T, Sakai H, Karikusa F, Sawasaki Y, Sakurai Y. Thermores-ponsive polymeric surfaces - control of attachment and detachment ofcultured cells. Makromol Chem Rapid Commun 1990;11:571e6.
[12] Canavan HE, Cheng XH, Graham DJ, Ratner BD, Castner DG. Cell sheetdetachment affects the extracellular matrix: a surface science studycomparing thermal liftoff, enzymatic, and mechanical methods. J BiomedMater Res 2005;75A:1e13.
[13] Cheng XH, Canavan HE, Stein MJ, Hull JJR, Kweskin SJ, Wagner MS, et al.Surface chemical and mechanical properties of plasma-polymerized N-isopropylacrylamide. Langmuir 2005;21:7833e41.
[14] Canavan HE, Cheng XH, Graham DJ, Ratner BD, Castner DG. Comparison ofnative extracellular matrix with adsorbed protein films using secondary ionmass spectrometry. Langmuir 2007;23:50e6.

d
A. Halperin, M. Kröger / Biomaterials 33 (2012) 4975e4987 4987
Author
provid
e
Redist
ributi
o
[15] Reed JA, Love SA, Lucero AE, Haynes CL, Canavan HE. Effect of polymerdeposition method on thermoresponsive polymer films and resulting cellularbehavior. Langmuir 2012;28:2281e7.
[16] Mizutani A, Kikuchi A, Yamato M, Kanazawa H, Okano T. Preparation ofthermoresponsive polymer brush surfaces and their interaction with cells.Biomaterials 2008;29:2073e81.
[17] Takahashi H, Nakayama M, Yamato M, Okano T. Controlled chain length andgraft density of thermoresponsive polymer brushes for optimizing cell sheetharvest. Biomacromolecules 2010;11:1991e9.
[18] Xu FJ, Zhong SP, Yung LYL, Kang ET, Neoh KG. Surface-active and stimuli-responsive polymer Si(100) hybrids from surface-initiated atom transferradical polymerization for control of cell adhesion. Biomacromolecules 2004;5:2392e403.
[19] Nagase K, Watanabe M, Kikuchi A, Yamato M, Okano T. Thermo-responsivepolymer brushes as intelligent biointerfaces: preparation via ATRP andcharacterization. Macromol Biosci 2011;11:400e9.
[20] Xue C, Choi BC, Braun PV, Kong Q, Leckband DE. Protein adsorption modesdetermine reversible cell attachment on poly(N-isopropylacrylamide)brushes. Adv Funct Mater; in press.
[21] Schmidt S, Zeiser M, Hellweg T, Duschl C, Fery A, Möhwald H. Adhesion andmechanical properties of PNIPAM microgel films and their potential use asswitchable cell culture substrates. Adv Funct Mater 2010;20:3235e43.
[22] Yamato M, Konno C, Koike S, Isoi Y, Shimizu T, Kikuchi A, et al. Nano-fabrication for micropatterned cell arrays by combining electron beam-irradiated polymer grafting and localized laser ablation. J Biomed Mater Res2003;67A:1065e71.
[23] Akiyama Y, Kikuchi A, Yamato M, Okano T. Ultrathin poly(N-isopropylacrylamide) grafted layer on polystyrene surfaces for cell adhe-sion/detachment control. Langmuir 2004;20:5506e11.
[24] Ebara M, Yamato M, Aoyagi T, Kikuchi A, Sakai K, Okano T. Immobilization ofcell-adhesive peptides to temperature-responsive surfaces facilitates bothserum-free cell adhesion and noninvasive cell harvest. Tissue Eng 2004;10:1125e35.
[25] Ebara M, Yamato M, Aoyagi T, Kikuchi A, Sakai K, Okano T. Temperature-responsive cell culture surfaces enable “on-off” affinity control between cellintegrins and RGDS ligands. Biomacromolecules 2004;5:505e10.
[26] Toomey R, Tirrell M. Functional polymer brushes in aqueous media from self-assembled and surface-initiated polymers. Annu Rev Phys Chem 2008;59:493e517.
[27] Milner ST. Polymer brushes. Science 1991;251:905e14.[28] Halperin A, Tirrell M, Lodge TP. Tethered chains in polymer microstructures.
Adv Polym Sci 1992;100:31e71.[29] Yim H, Kent MS, Satija S, Mendez S, Balamurugan SS, Balamurugan S, et al.
Evidence for vertical phase separation in densely grafted, high-molecular-weight poly(N-isopropylacrylamide) brushes in water. Phys Rev E 2005;72:051801.
[30] Yim H, Kent MS, Mendez S, Lopez GP, Satija S, Seo Y. Effects of grafting densityand molecular weight on the temperature-dependent conformational changeof poly(N-isopropylacrylamide) grafted chains in water. Macromolecules2006;39:3420e6.
[31] Koga T, Tanaka F, Motokawa R, Koizumi S, Winnik FM. Theoretical modeling ofassociated structures in aqueous solutions of hydrophobically modified tele-chelic PNIPAM based on a neutron scattering study. Macromolecules 2008;41:9413e22.
[32] Plunkett KN, Zhu X, Moore JS, Leckband DE. PNIPAM chain collapse dependson the molecular weight and grafting density. Langmuir 2006;22:4259e66.
[33] Zhu X, Yan C, Winnik FM, Leckband DE. End-grafted low-molecular-weightPNIPAM does not collapse above the LCST. Langmuir 2007;23:162e9.
[34] Malham IB, Bureau L. Density effects on collapse, compression, and adhesionof thermoresponsive polymer brushes. Langmuir 2010;26:4762e8.
[35] Balamurugan S, Mendez SS, Balamurugan MJ, O’Brien II, Lopez GP. Thermalresponse of poly(N isopropylacrylamide) brushes probed by surface plasmonresonance. Langmuir 2003;19:2545e9.
[36] Baulin VA, Zhulina EB, Halperin A. Self-consistent field theory of brushes ofneutral water-soluble polymers. J Chem Phys 2003;119:10977e88.
[37] Halperin A, Kröger M. Collapse of thermoresponsive brushes and the tuning ofprotein adsorption. Macromolecules 2011;44:6986e7005.
[38] Wilson CJ, Clegg RE, Leavesley DI, Pearcy MJ. Mediation of biomaterial-cellinteractions by adsorbed proteins: a review. Tissue Eng 2005;11:1e18.
[39] Halperin A. Polymer brushes that resist adsorption of model proteins: designparameters. Langmuir 1999;15:2525e33.
[40] Iwanaga Y, Braun D, Fromherz P. No correlation of focal contacts and closeadhesion by comparing GFP-vinculin and fluorescence interference of dil. EurBiophys J 2001;30:17e26.
[41] Bell GI. Models for specific adhesion of cells to cells. Science 1978;200:618e27.
[42] Evans E, Williams P. Dynamic force spectroscopy. In: Jülicher F, Ormos P,David F, Flyvbjerg H, editors. Physics of bio-molecules and cells. Berlin:Springer; 2002. p. 145e204.
copy
n not
perm
itted
[43] Discher DE, Janmey P, Wang YL. Tissue cells feel and respond to the stiffness oftheir substrate. Science 2005;310:1139e43.
[44] Moore SW, Roca-Cusachs P, Sheetz MP. Stretchy proteins on stretchysubstrates: the important elements of integrin-mediated rigidity sensing. DevCell. 2010;19:194e206.
[45] Vogel V, Sheetz M. Local force and geometry sensing regulate cell functions.Nat Rev Molec Cell Biol 2006;7:265e75.
[46] Zhulina EB, Borisov OV, Priamitsyn VA. Theory of the steric stabilization ofcolloid dispersions by grafted polymers. J Coll Interf Sci 1990;137:495e511.
[47] Afroze F, Nies E, Berghmans H. Phase transitions in the system poly(N-isopropylacrylamide)/water and swelling behavior of the correspondingnetworks. J Mol Struct 2000;554:55e68.
[48] Li F, Redick SD, Erickson HP, Moy VT. Force measurements of the a5b1integrin-fibronectin interaction. Biophys J 2003;84:1252e62.
[49] Ermilov V, Lazutin A, Halperin A. Colloids in brushes: the insertion free energyvia Monte Carlo simulation with umbrella sampling. Macromolecules 2010;43:3511e20.
[50] Halperin A, Zhulina EB. Atomic force microscopy of polymer brushes: colloidalversus sharp tips. Langmuir 2010;26:8933e40.
[51] Halperin A. Compression induced phase transitions in PEO brushes: the n-cluster model. Eur Phys J B 1998;3:359e64.
[52] Pankov R, Yamada KM. Fibronectin at a glance. J Cell Sci 2002;115:3861e3.[53] Schvartz I, Seger D, Shaltiel S. Vitronectin. Int J Biochem Cell Biol 1999;31:
539e44.[54] Stockmann A, Hess S, Declerck P, Timpl R, Preissner KT. Multimeric vitronectin
- identification and characterization of conformation-dependent self-associ-ation of the adhesive protein. J Biol Chem 1993;30:22874e82.
[55] Wrobel G, Höller M, Ingebrandt S, Dieluweit S, Sommerhage F, Bochem HP,et al. Transmission electron microscopy study of the cell-sensor interface. J RSoc Interf 2008;5:213e22.
[56] Al-Jawad M, Fragneto G, Liu J, Chang SR, Clarkson B. Fibronectin adsorptionstudied using neutron reflectometry and complementary techniques. EurPhys J E 2009;30:175e9.
[57] Xiong JP, Stehle T, Diefenbach B, Zhang RG, Dunker R, Scott DL, et al. Crystalstructure of the extracellular segment of integrin aVb3. Science 2001;294:339e45.
[58] Xiong JP, Stehle T, Zhang RG, Joachimiak A, Frech M, Goodman SL, et al. Crystalstructure of the extracellular segment of integrin aVb3in complex with anArg-Gly-Asp ligand. Science 2002;296:151e5.
[59] Popa AM, Angeloni S, Bürgi T, Hubbell JA, Heinzelmann H, Pugin R. Dynamicperspective on the function of thermoresponsive nanopores from in situ AFMand ATIZ-IR investigations. Langmuir 2010;26:15356e65.
[60] Naini CA, Franzka S, Frost S, Ulbricht M, Hartmann N. Probing the intrinsicswitching kinetics of ultrathin thermoresponsive polymer brushes. AngewChem Int Ed 2011;50:4513e6.
[61] Arnold M, Cavalcanti-Adam EA, Glass R, Eck W, Kantlehner M, Kessler H, et al.Activation of integrin function by nanopatterned adhesive interfaces. Chem-PhysChem. 2004;5:383e8.
[62] Yamato M, Konno C, Kushida A, Hirose M, Utsumi M, Kikuchi A, et al. Releaseof adsorbed fibronectin from temperature-responsive culture surfacesrequires cellular activity. Biomaterials 2000;21:981e6.
[63] Hatakeyama H, Kikuchi A, Yamato M, Okano T. Bio-functionalized thermor-esponsive interfaces facilitating cell adhesion and proliferation. Biomaterials2006;27:5069e78.
[64] Halperin A, Kröger M. Ternary protein adsorption onto brushes: strong versusweak. Langmuir 2009;25:11621e34.
[65] Ugarawa TP, Zamarron C, Veklich Y, Bowditch RD, Ginsberg MH, Weisel JW,et al. Conformational transitions in the cellebinding domain of fibronectin.Biochemistry 1995;34:4457e66.
[66] Seiffert D, Smith JW. The cell adhesion domain in plasma vitronectin is cryptic.Biol Chem 1991;21:13705e10.
[67] Zhou X, Li J, Wu C, Zheng B. Constructing the phase diagram of an aqueoussolution of poly(Nisopropylacrylamide) by controlled microevaporation ina nanoliter microchamber. Macromol Rapid Commun 2008;29:1363.
[68] Okada Y, Tanaka F. Cooperative hydration, chain collapse, and flat LCSTbehavior in aqueous poly(Nisopropylacrylamide) solutions. Macromolecules2005;38:4465e71.
[69] Chan K, Pelton R, Zhang J. On the formation of colloidally dispersed phase-separated poly(Nisopropylacrylamide). Langmuir 1999;15:4018e20.
[70] Balu C, Delsanti M, Guenoun P, Monti F, Cloitre M. Colloidal phase separationof concentrated PNIPAm solutions. Langmuir 2007;23:2404e7.
[71] Kwon OH, Kikuchi A, Yamato M, Sakurai Y, Okano T. Rapid cell sheetdetachment from poly(N isopropylacrylamide)-grafted porous cell culturemembranes. J Biomed Mater Res 2000;50:82e9.
[72] Kwon OH, Kikuchi A, Yamato M, Okano T. Accelerated cell sheet recovery byco-grafting of PEG with PIPAAm onto porous cell culture membranes.Biomaterials 2003;24:1223e32.
[73] Sackmann E, Goennenwein S. Cell adhesion as dynamic interplay of lock-and-key, generic and elastic forces. Progr Theor Phys Suppl 2006;165:78e99.





























