Embed Size (px)
Citation preview

lable at ScienceDirect
Biomaterials 32 (2011) 4830e4839
Contents lists avai
Biomaterials
journal homepage: www.elsevier .com/locate/biomateria ls
The promotion of axon extension in vitro using polymer-templated fibrin scaffolds
John B. Scott a,b, Mehdi Afshari c,d, Richard Kotek c, Justin M. Saul a,b,*aVirginia Tech eWake Forest University School for Biomedical Engineering and Sciences, Medical Center Boulevard, 2nd Floor, MRI Building, Winston-Salem, NC 27157, United StatesbWake Forest Institute for Regenerative Medicine, Medical Center Boulevard, Winston-Salem, NC 27157, United StatescNorth Carolina State University, College of Textiles, 2401 Research Drive, Raleigh, NC 27695, United Statesd Fiberweb, Inc., 70 Old Hickory Boulevard, Old Hickory, TN 37138, United States
a r t i c l e i n f o
Article history:Received 11 January 2011Accepted 18 March 2011Available online 13 April 2011
Keywords:FibrinMicropatterningNerve guideNerve tissue engineeringScaffold
* Corresponding author. Virginia Tech e WakeBiomedical Engineering and Sciences and Wake ForeMedicine, Medical Center Boulevard, 2nd Floor, MNC 27157, United States. Tel.: þ1 336 713 1326; fax: þ
E-mail address: [email protected] (J.M. Saul).
0142-9612/$ e see front matter � 2011 Elsevier Ltd.doi:10.1016/j.biomaterials.2011.03.037
a b s t r a c t
Biomaterial nerve cuffs are a clinical alternative to autografts and allografts as ameans to repair segmentalperipheral nerve defects. However, existing clinical biomaterial constructs lack true incorporation ofphysical guidance cues into their design. In both two- and three-dimensional systems, it is known thatsubstrate geometry directly affects rates of axon migration. However, the ability to incorporate these cuesinto biomaterial scaffolds of sufficient porosity to promote robust nerve regeneration in three-dimensionalsystems is a challenge.We have developed fibrin constructs fabricated by a sacrificial templating approach,yielding scaffolds withmultiple 10e250 mmdiameter conduits depending on the diameter of the templatefibers. The resulting scaffolds contained numerous, highly aligned conduits, had porosity of w 80%, andshowed mechanical properties comparable to native nerve (150e300 kPa Young’s modulus). We studiedthe effects of the conduit diameters on the rate of axon migration through the scaffold to investigate ifmanipulation of this geometry could be used to ultimately promotemore rapid bridging of the scaffold. Alldiameters studied led to axon migration, but in contrast to effects of fiber diameters in other systems, therate of axon migration was independent of conduit diameter in these templated scaffolds. However,aligned conduits did support more rapid axon migration than non-aligned, tortuous controls.
� 2011 Elsevier Ltd. All rights reserved.
1. Introduction
There are over 50,000 surgical procedures for peripheral nerveinjuries reported annually [1] in both civilian and military settingsdue tomotor vehicle accidents, knife injuries, gunshot wounds, andother sources of trauma [2,3]. Though small segmental injuries canbe repaired via surgical coaptation of nerve ends, the implantationof a graft is required if direct suture would result in tension on thenerve [1].
The current clinical gold standard of autografting haswell-knowndetriments including multiple surgical sites and donor sitemorbidity [4,5]. Allografts have been employed [1,6], but concernspersist regarding the potential for disease transmission [7] andethical concerns [8]. Biomaterial nerve cuffs based on collagen havealso been employed at the clinical level. While these nerve cuffmaterials avoid drawbacks of human tissue grafting, it is not clearthat their performance matches autografts and allografts [4]. In
Forest University School forst Institute for RegenerativeRI Building, Winston-Salem,1 336 713 7290.
All rights reserved.
addition to the absence of neurotrophic factors present in autograftsand allografts, nerve cuffs possess only one lumen, and thereforehave far less surface area per volume than, for example, decellular-ized constructs.
At the pre-clinical level, a number of filler materials, oftenhydrogels [9e15], have been incorporated into nerve cuff materials.These filler materials are believed to provide a physical matrix ontowhich infiltrating cells can attach. However, achieving robustregeneration with these filler materials is generally believed torequire the presentation of soluble or immobilized chemical cuessuch as growth factors or cell stimulating peptide sequences [16,17].
Besides chemical cues, there are multiple lines of evidencesuggesting that physical cues play an important role in promotingaxonmigration. The efficacy of regeneration through autografts andallografts may be due to the presence of aligned conduits inconjunction with neurotrophins [18]. Further, decellularization ofnerve tissue results in hollow, aligned conduits that support nerveregeneration [19,20]. It is also known that axons will track alongfibers formed by extrusion or electrospinning in a fashion depen-dent on the diameter of the fibers [21,22]. Importantly, neuralscaffolds consisting only of electrospun fibers are known to affectregenerative processes in vitro [22] and have led to axonal bridgingof large neural defects [23].

J.B. Scott et al. / Biomaterials 32 (2011) 4830e4839 4831
From a materials perspective, one drawback to bundled fiberapproaches is that the overall porosity of nerve guidance channelspacked with these fibers is expected to be quite low. In fact,approaches to provide physical cues with fibers while providingsufficient porosity are being actively investigated [24]. Here, wedescribe a different approach to the development of biomaterialscaffolds that can be fabricated de novo, provide physical guidancecues, and provide sufficient porosity to allow robust axonal growth.
The goals of the present study were twofold. The first goal was tofabricate highly porous scaffolds containing large numbers ofaxially-aligned luminal conduitswithin a biomaterial scaffold havingmechanical properties suitable for neural repair. To accomplish this,we utilized a sacrificial templating approach within the context ofa degradable fibrin hydrogel (see Fig. 1). Numerous natural andsynthetic polymer scaffolds are compatible with this type ofapproach [25e29]. However, we focused on the use of fibrin in thesestudies in part because fibrin is a component of native peripheralnerve repair, being laid down in a cable ahead of the growing axonbundle as a template for growth [30]. Moreover, the sacrificialtemplating approach has the advantage of generating alignedcylindrical conduits similar in morphology to denervated endoneu-rial tubes left after Wallerian degeneration is complete [31], whichwe reasonedwould provide a conducive substrate to axonmigration.As a final design consideration, we aimed to incorporate scaffoldswith mechanical integrity similar to that of native nerve so theywould handle well and could ultimately integrate properly withtarget tissue.
The second goal was to assess the rate of axonal migrationthrough the conduits of these scaffolds in order to determine theeffect of the conduit diameter on the rate of axonal extension. Weemployed an embryonic chicken dorsal root ganglia (DRG)model toassess rates of axon migration [32]. Speed of axon migration isa critical metric for nerve regeneration as more positive functionaloutcomes are expected with more rapid neural regeneration.
Fig. 1. Scaffold fabrication process. (A) Sacrificial poly(methyl methacrylate) (pMMA)template fibers, shown in gray, were bundled within a Teflon� mold. (B) A solution offibrinogen in PBS was back-filled around the aligned fiber bundle via centrifugationand polymerized to fibrin, shown in brown, using thrombin. (C) pMMA was dissolvedusing acetone, leaving an array of aligned empty conduits, shown in white, within thefibrin matrix. (For interpretation of the references to color in this figure legend, thereader is referred to the web version of this article.)
Employing sacrificial template fibers of well-known diameterallowed for the fabrication of scaffolds with conduits that appearedto span the full length of the scaffold in a wide range of diameters.This allowed us to assess the effects of the conduit diameters on therate of axon migration within the context of a three-dimensionalconstruct of mechanical properties comparable to native nerve.This report describes the fabrication and characterization of thescaffolds as well as the axonal response as a function of conduitdiameter in these scaffolds.
2. Methods
2.1. Template fiber fabrication and characterization
Monofilament strands of poly(methyl-methacrylate) (pMMA) were formed bymelt extrusion at the North Carolina State University College of Textiles. 100 kDapMMA (Polysciences, Warrington, PA) was melt extruded on two different extrudersystems to achieve a range of fiber diameters by variation of system parametersincluding type of extruder, temperature, rate of extrusion, and collection godetspeed (see Table 1). One set of extrusions was performed on an Alex James andAssociates, Incorporated melt extruder with a spinneret containing 64 holes. Eachorifice had a diameter of 0.6 mm and a length:diameter ratio of 2.3:1. The processtemperatures were controlled at five different zones in the extruder. The tempera-tures from the feed to the die zone were 210, 220, 240, 245, and 250 �C. Undrawnfilaments were taken up at 200 or 800 m/min (Table 1). Melt spinning was alsoperformed on the Bradford piston system with a one hole spinneret 0.5 mm indiameter and length:diameter ratio of 2. The piston temperature was maintainedbetween 240 and 250 �C (Table 1). The undrawnmonofilament was taken up at 27.5,80 or 96.5 m/min (Table 1).
Cellulose acetate (CA) fibers (generous donation of Eastman Chemical Co.,Kingsport, TN) were also used. Brightfield imaging of both CA and pMMA fibers wasperformed on a Zeiss Axiovert II microscope, and micrographs were captured withAxioVision software (both Carl Zeiss, Inc. North America, Thornwood, NY). Imageswere imported into ImageJ software (National Institutes of Health, Bethesda, MD)and fiber diameter was measured for at least N ¼ 5 samples. Average and standarddeviation of fiber diameters were recorded for use in studying the relationshipbetween template fiber size and fibrin scaffold microarchitecture.
2.2. Scaffold fabrication
Fibrin scaffolds with highly aligned conduits were fabricated by using a poly-mer-templating approach, summarized graphically in Fig. 1, in which polymericfibers were used as sacrificial templates. To begin, aligned pMMA fibers or celluloseacetate (CA) fibers were bundled tightly and inserted into a Teflon� tube (McMaster-Carr, Los Angeles, CA). Scaffolds were fabricated in Teflon� tubing with either 3 mmor 8 mm inside diameter, depending on the nature of the experiment (see below).Care was taken to fill as much free space within the mold as possible in order tomaximize porosity of the final scaffold. CA fibers averaged 11.7 mm in diameter, whilefive different diameters of pMMA fiber were used, averaging 18.5 mm, 40.5 mm,105.4 mm, 161.3 mm, and 247.7 mm. Each scaffold was templated with fibers ofa uniform diameter, resulting in a range of six scaffold types for use in furtherexperiments. A 200 mg/mL solution of bovine fibrinogen (SigmaeAldrich, St. Louis,MO) in PBS was then back-filled around the fiber bundle by centrifugation ina Sorvall Legend RT benchtop centrifuge (Kendro Laboratory Products, Newtown,CT) at 50 rcf. The fibrinogen surrounding the pMMA templates was then polymer-ized for at least 16 h in a thrombin working solution consisting of 26 parts PBS, twoparts 5.5 mg/mL CaCl2 (Thermo Fisher Scientific, Waltham, MA), and two parts 250U/mL thrombin (Sigma). After polymerization, excess fibrin was removed from eachend of the fiber bundle with a razor blade. The resulting scaffold material was then
Table 1Template fiber fabrication parameters and resulting diameter.
ExtrusionMethod
Speed of AlexJames Motor(RPM) or Piston(mm/min)
ExtrusionTemperature(�C)
GodetSpeed(m/min)
FiberDiameter (mm)Average � SD
CelluloseAcetate
a a a 11.7 � 2.5
Alex James 15 250 800 18.5 � 3.7Alex James 15 250 200 40.5 � 3.3Piston 2.5 250 96.5 105.4 � 6.8Piston 3.0 250 80 161.3 � 24.0Piston 3.6 240 27.5 247.7 � 3.3
a Donated cellulose acetate fibers were manufactured using a proprietary tech-nique and are therefore only characterized by diameter.

J.B. Scott et al. / Biomaterials 32 (2011) 4830e48394832
placed in acetone (Fisher) to selectively dissolve the sacrificial CA or pMMA fibers.Three acetone washes were used lasting two hours, four hours, and 16 hours,respectively. This acetone step also proved sufficient to sterilize scaffolds for use intissue culture experiments.
Random porous and non-porogen control scaffolds were fabricated similarly. Tocreate scaffolds incorporated with a random porous network, a template wasformed by filling the Teflon� mold with 180e212 mm diameter pMMA beads (Pol-ysciences, Warrington, PA). This size range of beads was obtained by sieving. Thebead-filled molds were vigorously tapped to settle beads into a close-packedarrangement and heated at 140 �C for 22 h to sinter the beads together at points ofcontact [25,33]. Fibrinogen back-filling, polymerization into fibrin, and templateremoval with acetone were performed as above for the sacrificial fibers. Non-porogen scaffolds were created by omitting the inclusion of a polymer template.Instead, fibrinogen was pipetted into the Teflon� mold, a wedge was removed fromthe wall of the mold with a razor blade, and the entire assembly was immersed inthrombin working solution as described above to polymerize the fibrin block. Theremaining Teflon�was then removed, and acetonewashes were performed as aboveto sterilize the scaffolds. These resulting non-porogen scaffolds were essentiallya fibrin hydrogel plug.
2.3. Characterization of scaffold morphology
Scaffold morphology was evaluated by using scanning electron microscopy(SEM) of scaffolds consisting of each different sacrificial template diameter. Scaffoldsfabricated as described abovewere cut in half longitudinally with a scalpel, hydratedin sterile PBS, dried in a model EMS850X critical point dryer (Electron MicroscopySciences, Hatfield, PA), mounted on SEM chucks for longitudinal or cross-sectionalimaging, and sputter coated with an Au-Pd mixture using a Hummer 6.2 sputtercoater (Anatech Ltd., Battle Creek, MI). Scaffolds were imaged with an S-2600 Nenvironmental scanning electron microscope (Hitachi High Technologies America,Inc., Schaumburg, IL) in either longitudinal- or cross-section. Images of the longi-tudinally-sectioned scaffolds were imported into ImageJ software to quantify thedegree of conduit alignment within the scaffold. Five regions within the micrographwere randomly selected and, within each region, the angular deviation from thescaffold longitudinal axis was recorded for all observed conduits. Similarly, thediameters of the conduits were determined by ImageJ analysis of the cross-sectionalSEM images by measuring five separate regions of a single scaffold.
2.4. Scaffold porosity determined by mercury porosimetry
Porosity was measured for scaffolds made with each fiber template by mercuryporosimetry. 8 mm diameter scaffolds of different luminal conduit dimensions wereprepared as described above. Samples were then trimmed to 1 cm length, weighed,and evaluated using an AutoPore IV mercury porosimeter (Micromeritics, Norcross,GA). Mercury intrusion was performed over a range of 0.10e33000 psia, and totalintrusion volume was measured over this range. Total pore volume was calculatedautomatically by the bundled software package (AutoPore IV 9500 v1.09, Micro-meritics) by subtracting intrusion volume at maximum pressure from intrusionvolume at zero pressure [34]. Sample bulk volume was assumed to equal totalpenetrometer volume minus intrusion volume at zero pressure. Sample percentporosity was then calculated from bulk volume and pore volume by software usingthe equation:
P% ¼ (Vpore/Vbulk) * 100% [34].N ¼ 3 were evaluated for each scaffold type, and analysis of variance (ANOVA)
was performed on resulting averages using Prism 5 (GraphPad Software, La Jolla, CA)to determine if any significant differences existed between scaffold types usinga value of P < 0.05 to determine statistical significance.
2.5. Mechanical properties determined by tensile testing
To determine stiffness and strength, scaffolds were fabricated as describedabove, re-hydrated in PBS for at least six hours, and trimmed to 3 cm length. Scaf-folds of each type (n¼ 3) weremounted into the grips of aModel 5544 tension tester(Instron, Canton, MA) equipped with a 100 N load cell, and scaffold diameter andlength were measured. Samples were subjected to 10 cycles of 0.3 mm maximumextension at 0.1667 mm/s as a preconditioning step prior to tensile testing to failure.Raw data were exported to MS Excel 2007 (Microsoft, Redmond, WA) and plotted astensile load vs. extension and tensile stress vs. strain. Maximum load was directlylocated in tabular raw data and reported. The linear region of the stress vs. strain plotwas selected manually, and linear regression was performed on this data set. Theslope of the resulting line was reported as the scaffold Young’s Modulus (E). ANOVAand post-hoc Tukey’s test (where indicated by positive ANOVA) of calculated E andmaximum load were performed using Prism 5. N ¼ 3 samples were evaluated foreach scaffold and P < 0.05 was taken as statistically significant.
2.6. DRG seeding in vitro
To determine the rate of axon infiltration through the scaffolds in response toscaffold architecture, we employed an embryonic chicken dorsal root ganglia (DRG)
model [32]. DRG culture medium was composed of HyClone high-glucose Dulbec-co’s modified Eagle’s medium (Fisher),10% v/v fetal bovine serum (GIBCO Invitrogen,Carlsbad, CA), 1% v/v 5-fluoro-20-deoxyuridine (FDU, Sigma), 1% v/v Uridine (Sigma),1% v/v penicillin-streptomycin (Invitrogen), 20 mg/mL bovine aprotinin (Sigma), and50 ng/mL nerve growth factor (NGF, BD Biosciences). Aprotinin was included toreduce fibrinolytic scaffold degradation, and NGF was used to stimulate axonalextension [32]. In particular, we note that FDU and uridine were added to minimizeproliferation of non-neuronal cells, especially Schwann cells. The purpose ofinhibiting Schwann cell proliferationwas to allow us to focus on axonal extension inresponse to the conduit diameters alone rather than in response to Schwann cellsand other cell-based trophic cues.
3 mm diameter aligned conduit scaffolds and control scaffolds, fabricated asdescribed above, were trimmed to 5 mm in length and washed three times in sterilePBS for at least two hours, four hours, and 16 hours respectively to remove acetone.Small amounts of 100 mg/mL fibrinogen and thrombin working solution were usedas a glue to attach scaffolds to the wall of individual wells in 24-well Falcon poly-styrene culture dishes (BD Biosciences, Franklin Lakes, NJ) under aseptic conditions.Scaffolds were kept wetted with PBS to prevent contraction and retain normalmorphology of the fibrin matrix until DRG seeding.
DRGwere dissected from E7-E8 chicken embryos (Tyson Chicken, Hays, NC) andstored on ice in sterile 1% penicillin/streptomycin in PBS for a time not exceeding onehour. Ten DRG were seeded on the top surface of each scaffold to increase thelikelihood of obtaining a longitudinal section from the scaffold that had a DRG andassociated axons (see below). DRG culture mediumwas added to a height just belowthe seeded scaffold top to allow DRG attachment to the scaffolds. This cell clusterwas allowed to attach to the scaffold for 24 h, after which additional medium wasadded to cover the top of the scaffold and DRG. Samples were removed after 1, 2, or 3days, with n ¼ 3 for each combination of scaffold type and time point. Removedscaffolds were briefly washed once in PBS, oriented in a plastic tray for longitudinalsectioning, and then snap frozen in OCT compound (Sakura Finetek USA, Torrance,CA) with liquid nitrogen.
2.7. Immunohistochemistry
Axon infiltration into fibrin scaffolds was determined by immunohistochemi-cally staining longitudinal sections for neurofilament. 50 mm-thick sections were cutfrom OCT-embedded scaffolds along longitudinal planes using a CM 1950 cryostat(Leica Microsystems, Bannockburn, IL) and immobilized on silane-coated glassslides (LabScientific, Livingston, NJ). Sections containing DRGwere fixed for 3 min in10% neutral-buffered formalin (Leica) and then washed to remove excess fixativeand OCT compound. Sections were blocked using Dual Endogenous Enzyme Block(Dako North America, Carpinteria, CA) and Serum-Free Protein Block (Dako).Sections were then treated with mouse monoclonal antibody to 160 kDa and200 kDa neurofilament (AbCam, Cambridge, MA) or to mouse IgG (isotype) controlat 1:100 dilution in Antibody Diluent Solution (Dako) for 30min. Serum-Free ProteinBlock was applied a second time before then treating sections with biotinylated goatanti-mouse IgG or horse anti-mouse IgG (Vector Laboratories, Burlingame, CA) at1:200 dilution in Antibody Diluent Solution. Sections were then treated with ABC-AP reagent (Vector) before staining with Vector Red Alkaline Phosphatase Substrate(Vector). Coverslips were mounted to slides using Fluoromount-G (SouthernBiotech, Birmingham, Alabama) and dried overnight at 4 �C.
2.8. Image analysis
Staining was imaged via brightfield microscopy using a DM4000 B microscope(Leica) with attached Retiga 2000RV camera (QImaging, Surrey, BC, Canada),coupled with ImagePro 6.2 software (Media Cybernetics Inc., Bethesda, MD).Maximum axon length was measured using ImageJ software. After the longest axonwas identified, a line parallel to the scaffold longitudinal axis was drawn betweenthe axon tip and scaffold surface and the length of the linewasmeasured. One imagefrom each seeded scaffold was analyzed and average maximum axon lengthcalculated for each combination of time point and scaffold type, resulting in N ¼ 3for each reported data point. This method of quantification of maximum length ofaxon extension is consistent with those reported by others [21]. Statistical analysis ofthese averages was performed in Prism 5 using ANOVA and post-hoc pairwiseBonferroni comparisons where appropriate. P < 0.05 was used as the significancethreshold.
3. Results
3.1. Template fiber characterization
To fabricate scaffolds and assess the effects of conduit diameteron the rate of axon extension, it was necessary to acquire polymericfiber templates of a range of diameters. Acetone-soluble polymerfibers were either custom extruded or donated to serve as thesetemplates. 12 mm templates were obtained by donation of cellulose

J.B. Scott et al. / Biomaterials 32 (2011) 4830e4839 4833
acetate fibers, but to obtain template fibers of other diameters,processing parameters for extrusion were modulated to obtainfibers diameters from approximately 19e250 mm as determined bylight microscopy and summarized in Table 1. We anticipated thatthis wide micro-scale range would provide a good basis for evalu-ating the effects of conduit diameter on axon growth in threedimensions.
3.2. Scaffold morphology
One goal of these studies was to fabricate three-dimensionalscaffolds with well-defined conduit dimensions to assess the effectof conduit diameters on axon infiltration rate. We employedscanning electron microscopy (SEM) to qualitatively and quantita-tively assess the physical architecture of the scaffolds. Fig. 2 showsSEM micrographs of scaffolds in both the longitudinal and trans-verse planes at magnifications of 25e50X. All scaffolds had thesame general architecture with densely-packed hollow conduitsthat were axially-aligned. It can be clearly observed that with smalldiameter templates, the conduit diameters are much smaller (e.g.,12 mm template fibers) compared to larger diameter template fibers(e.g., 250 mm template fibers). Higher magnification images areprovided in Fig. 2 for smaller diameter templates to demonstratethe presence of conduits difficult to see at lower magnification.Based on the longitudinal sections of all scaffolds, the conduitsappear to run the full length of the scaffolds, indicating the ability ofaxons tomigrate through the length of thematerial (see below).Wenoted artifacts in the SEM images of 19 and 41 mm templates thatwe believe may be a slight contamination of the pMMA templateswith higher molecular weight polymers. This contamination didnot lead to any observable effects in other studies and appears to bea small fraction of the total polymer template used.
To quantitate the axial alignment of the scaffolds, we measuredthe deviation of these conduits from the main axis of the scaffold.Fig. 3 shows the alignment of the conduits within the scaffoldsmeasured by this technique. Quantification of conduit orientationrevealed that all patterned channels fell within 16� of the scaffoldaxis and assumed a roughly Gaussian distribution centered near 0�.The results of this characterization indicate that the scaffolds havehighly aligned and densely-packed conduits.
As described above, we had measured the diameters of thetemplating fibers, which should provide an indication of theresulting conduits. However, we also estimated the diameters ofthe conduits by measurement from the SEM images. Fig. 4 showsthe results of these SEM measurements compared to the templatediameters. We note that in the larger diameter scaffolds the SEMmeasurements are less than the templating diameters. However,we observed significant contraction of the scaffolds during criticalpoint drying for these larger scaffolds (e.g., see Fig. 2F), which is thereason for the difference. Error bars denote standard deviations ofabout 5e10% of the average, indicating that the pores were quiteuniform in their diameter for a given template. We also noted thatthe scaffolds did have “memory” in that after contraction due todehydration, scaffolds immersed in aqueous media did return tonormal size through swelling of the hydrogel.
3.3. Scaffold porosity
To further characterize the observation of the packing density ofthe conduits from the SEM studies, we performed mercury poros-imetry. Table 2 summarizes the scaffold porosity. These resultsindicate that the average porosity for all templates fell near 80%with little variability. This is important for studies below with axonmigration. Because all scaffolds had nearly the same porosity,differences in axon migration could be attributed to the conduit
diameters or pattern (aligned vs. random porous) but not todifferences in scaffold porosity.
3.4. Mechanical testing
A second design parameter for these studies was to developscaffolds that would be suitable in mimicking the mechanicalcharacteristics of native tissue and to ensure that they would havesufficient mechanical integrity. To determine the mechanicalproperties of the fabricated scaffolds, we performed tensile tests onscaffolds to determine the elastic modulus and maximal load asa function of the conduit dimensions. Fig. 5 summarizes the tensiletest data from these studies. We observed that the maximal loadfell between 0.5 N and 1.2 N for all scaffolds, with significantdifference observed between only one pair. The elastic moduli ofscaffolds featuring smaller conduits were shown to be significantlyhigher than that of scaffolds fabricated with larger conduits. Wenote that elastic moduli in the range of 100e330 kPa are compa-rable in magnitude to the elastic modulus of native nerve [35].
3.5. Axon infiltration in vitro
Others have previously shown in fiber-based systems that therate of axon migration is dependent upon the dimensions of thefiber on which axons are tracking [21,22]. The fabrication of scaf-folds with varying conduit diameters but similar porositiesallowed us to explore the effect of conduit dimensions on axoninfiltration within the context of a three-dimensional, porousscaffold. We employed an embryonic chicken DRG model, as thisprovided a system in which a full neural explant could be culturedwith the scaffolds. We utilized antimitotics (uridine and FDU) inorder to isolate effects of the scaffold on axons alone rather thanadding a confounding factor related to the issue of Schwann cellmigration. Moreover, preliminary studies revealed that scaffolddegradation by DRG occurred on a time scale similar to that of axoninfiltration, potentially complicating axon length measurements.For this reason, we included aprotinin to the culture media to haltfibrinolytic degradation of the scaffold and allow for reliablemeasurements of axon infiltration.
To assess axonal response to the scaffolds, we used immunohis-tochemical staining with anti-neurofilament antibodies. A colori-metric stain (Vector Red Alkaline Phosphatase) was used instead offluorometric stainingmethods due to the levels of autofluorescencefrom the high fibrinogen (and ultimately fibrin) concentration usedto fabricate the scaffolds. A representative image from each scaffoldtype after one day in culture is shown in Fig. 6. Similar results wereobtained with anti-betaIII-tubulin antibodies (images not shown),though staining was more intense with neurofilament. As can beseen in Fig. 6, DRG attached to scaffolds after 24 h in culture andaxons began to migrate both across the scaffold edge and into theconduits of the scaffold.
We employed two control scaffolds to contrast the behaviorwith the axially-aligned conduits. Fibrin plugs (non-porogencontrols) were used to demonstrate the need for porous patterningwithin dense fibrin hydrogels created using 200 mg/mL fibrinogen.As shown in Fig. 6, no axon infiltration was observed into the non-porogen fibrin hydrogel control. A fibrin scaffold with randomlyoriented spherical pores (rather than axially-aligned conduits) wasalso used to determine the effect of the axial alignment. As can beseen in Fig. 6, we did not observe the same degree of axon infil-tration into these spherically-templated scaffolds. Dotted lineswere added to Fig. 6 using Photoshop Elements 6.0 (Adobe SystemsIncorporated, San Jose, CA) to highlight the length of axonalextension into the scaffolds.
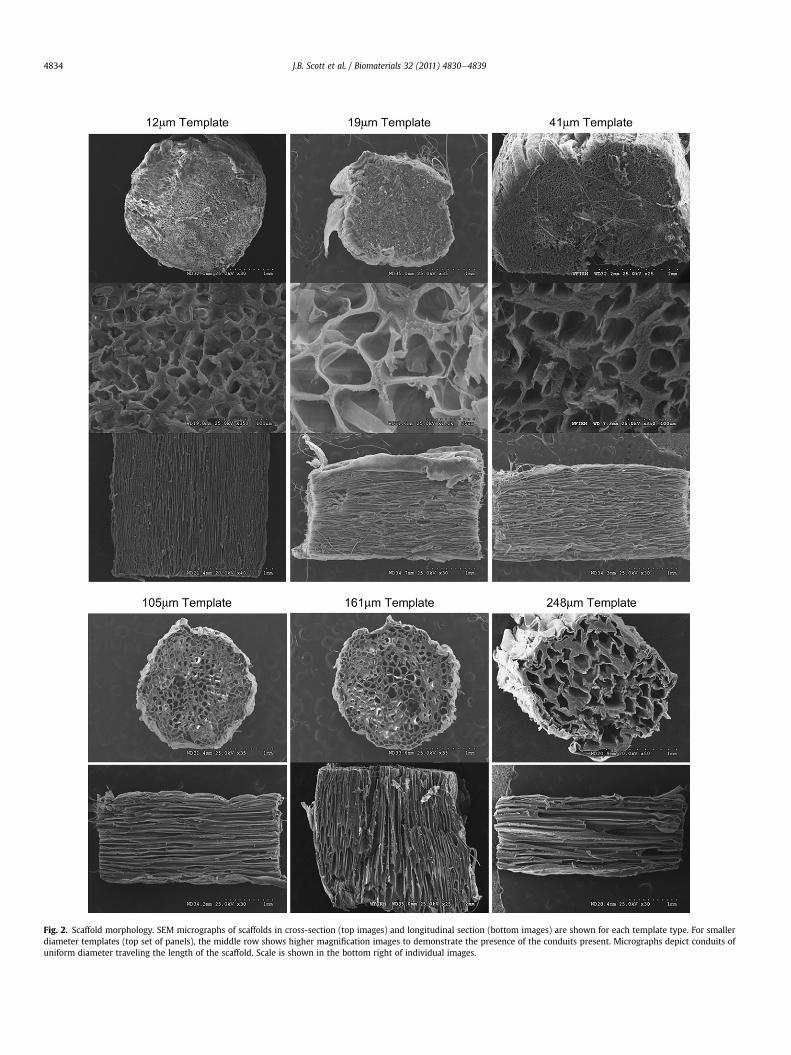
Fig. 2. Scaffold morphology. SEM micrographs of scaffolds in cross-section (top images) and longitudinal section (bottom images) are shown for each template type. For smallerdiameter templates (top set of panels), the middle row shows higher magnification images to demonstrate the presence of the conduits present. Micrographs depict conduits ofuniform diameter traveling the length of the scaffold. Scale is shown in the bottom right of individual images.
J.B. Scott et al. / Biomaterials 32 (2011) 4830e48394834
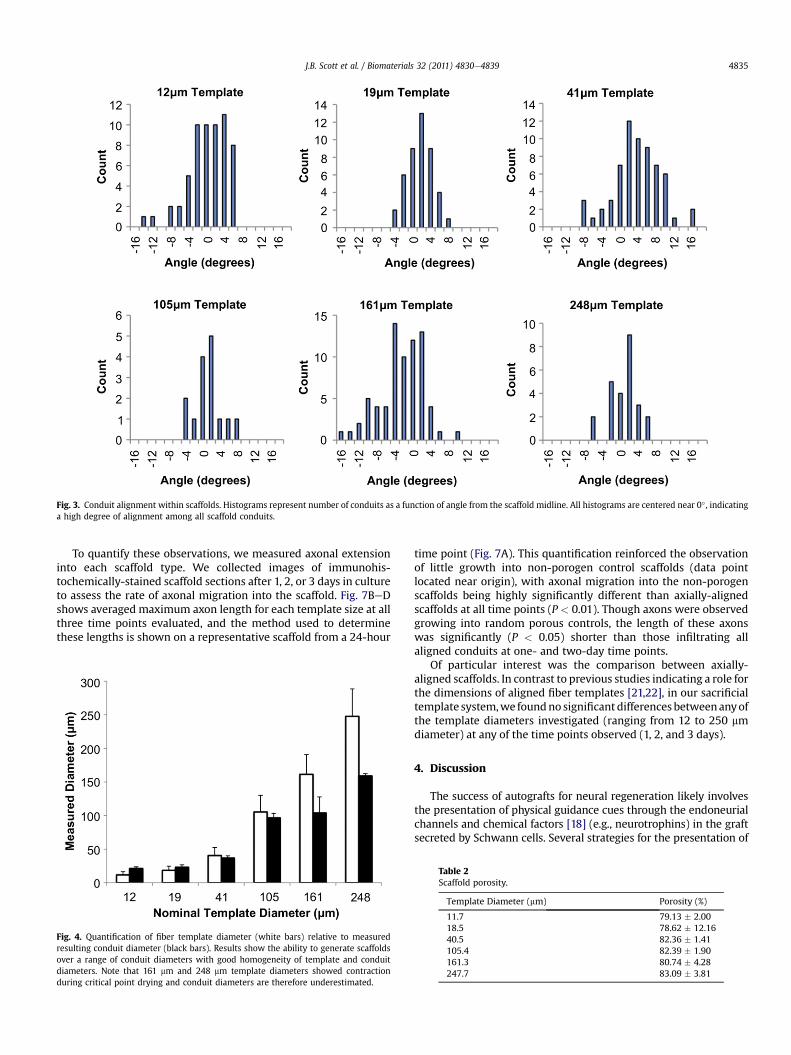
Fig. 3. Conduit alignment within scaffolds. Histograms represent number of conduits as a function of angle from the scaffold midline. All histograms are centered near 0� , indicatinga high degree of alignment among all scaffold conduits.
J.B. Scott et al. / Biomaterials 32 (2011) 4830e4839 4835
To quantify these observations, we measured axonal extensioninto each scaffold type. We collected images of immunohis-tochemically-stained scaffold sections after 1, 2, or 3 days in cultureto assess the rate of axonal migration into the scaffold. Fig. 7BeDshows averaged maximum axon length for each template size at allthree time points evaluated, and the method used to determinethese lengths is shown on a representative scaffold from a 24-hour
Fig. 4. Quantification of fiber template diameter (white bars) relative to measuredresulting conduit diameter (black bars). Results show the ability to generate scaffoldsover a range of conduit diameters with good homogeneity of template and conduitdiameters. Note that 161 mm and 248 mm template diameters showed contractionduring critical point drying and conduit diameters are therefore underestimated.
time point (Fig. 7A). This quantification reinforced the observationof little growth into non-porogen control scaffolds (data pointlocated near origin), with axonal migration into the non-porogenscaffolds being highly significantly different than axially-alignedscaffolds at all time points (P < 0.01). Though axons were observedgrowing into random porous controls, the length of these axonswas significantly (P < 0.05) shorter than those infiltrating allaligned conduits at one- and two-day time points.
Of particular interest was the comparison between axially-aligned scaffolds. In contrast to previous studies indicating a role forthe dimensions of aligned fiber templates [21,22], in our sacrificialtemplate system,we foundno significant differences betweenanyofthe template diameters investigated (ranging from 12 to 250 mmdiameter) at any of the time points observed (1, 2, and 3 days).
4. Discussion
The success of autografts for neural regeneration likely involvesthe presentation of physical guidance cues through the endoneurialchannels and chemical factors [18] (e.g., neurotrophins) in the graftsecreted by Schwann cells. Several strategies for the presentation of
Table 2Scaffold porosity.
Template Diameter (mm) Porosity (%)
11.7 79.13 � 2.0018.5 78.62 � 12.1640.5 82.36 � 1.41105.4 82.39 � 1.90161.3 80.74 � 4.28247.7 83.09 � 3.81
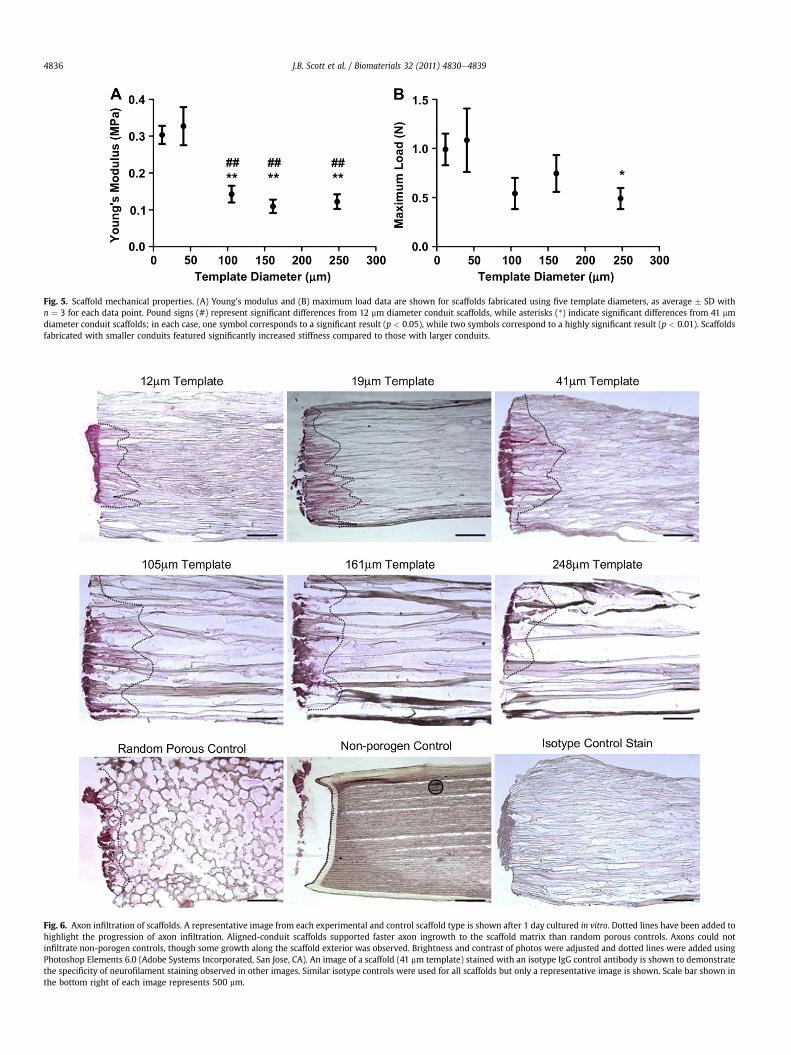
Fig. 5. Scaffold mechanical properties. (A) Young’s modulus and (B) maximum load data are shown for scaffolds fabricated using five template diameters, as average � SD withn ¼ 3 for each data point. Pound signs (#) represent significant differences from 12 mm diameter conduit scaffolds, while asterisks (*) indicate significant differences from 41 mmdiameter conduit scaffolds; in each case, one symbol corresponds to a significant result (p < 0.05), while two symbols correspond to a highly significant result (p < 0.01). Scaffoldsfabricated with smaller conduits featured significantly increased stiffness compared to those with larger conduits.
Fig. 6. Axon infiltration of scaffolds. A representative image from each experimental and control scaffold type is shown after 1 day cultured in vitro. Dotted lines have been added tohighlight the progression of axon infiltration. Aligned-conduit scaffolds supported faster axon ingrowth to the scaffold matrix than random porous controls. Axons could notinfiltrate non-porogen controls, though some growth along the scaffold exterior was observed. Brightness and contrast of photos were adjusted and dotted lines were added usingPhotoshop Elements 6.0 (Adobe Systems Incorporated, San Jose, CA). An image of a scaffold (41 mm template) stained with an isotype IgG control antibody is shown to demonstratethe specificity of neurofilament staining observed in other images. Similar isotype controls were used for all scaffolds but only a representative image is shown. Scale bar shown inthe bottom right of each image represents 500 mm.
J.B. Scott et al. / Biomaterials 32 (2011) 4830e48394836
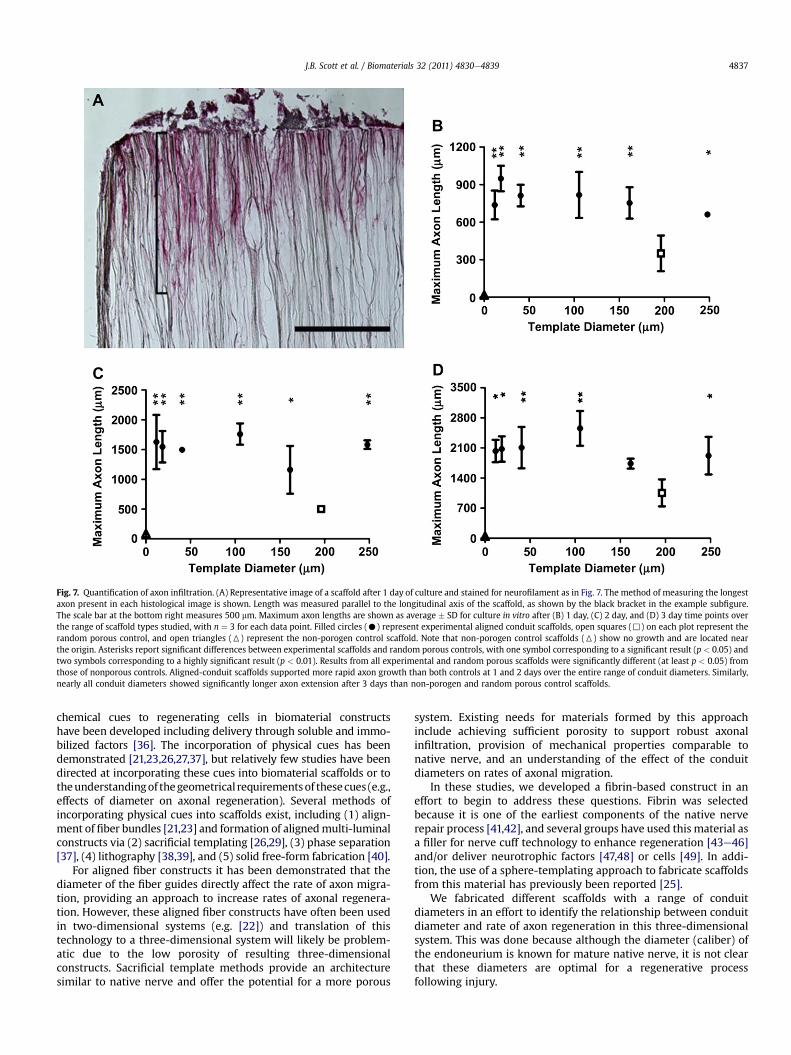
Fig. 7. Quantification of axon infiltration. (A) Representative image of a scaffold after 1 day of culture and stained for neurofilament as in Fig. 7. The method of measuring the longestaxon present in each histological image is shown. Length was measured parallel to the longitudinal axis of the scaffold, as shown by the black bracket in the example subfigure.The scale bar at the bottom right measures 500 mm. Maximum axon lengths are shown as average � SD for culture in vitro after (B) 1 day, (C) 2 day, and (D) 3 day time points overthe range of scaffold types studied, with n ¼ 3 for each data point. Filled circles (C) represent experimental aligned conduit scaffolds, open squares (,) on each plot represent therandom porous control, and open triangles (6) represent the non-porogen control scaffold. Note that non-porogen control scaffolds (6) show no growth and are located nearthe origin. Asterisks report significant differences between experimental scaffolds and random porous controls, with one symbol corresponding to a significant result (p < 0.05) andtwo symbols corresponding to a highly significant result (p < 0.01). Results from all experimental and random porous scaffolds were significantly different (at least p < 0.05) fromthose of nonporous controls. Aligned-conduit scaffolds supported more rapid axon growth than both controls at 1 and 2 days over the entire range of conduit diameters. Similarly,nearly all conduit diameters showed significantly longer axon extension after 3 days than non-porogen and random porous control scaffolds.
J.B. Scott et al. / Biomaterials 32 (2011) 4830e4839 4837
chemical cues to regenerating cells in biomaterial constructshave been developed including delivery through soluble and immo-bilized factors [36]. The incorporation of physical cues has beendemonstrated [21,23,26,27,37], but relatively few studies have beendirected at incorporating these cues into biomaterial scaffolds or totheunderstandingof thegeometrical requirementsof thesecues (e.g.,effects of diameter on axonal regeneration). Several methods ofincorporating physical cues into scaffolds exist, including (1) align-ment of fiber bundles [21,23] and formation of alignedmulti-luminalconstructs via (2) sacrificial templating [26,29], (3) phase separation[37], (4) lithography [38,39], and (5) solid free-form fabrication [40].
For aligned fiber constructs it has been demonstrated that thediameter of the fiber guides directly affect the rate of axon migra-tion, providing an approach to increase rates of axonal regenera-tion. However, these aligned fiber constructs have often been usedin two-dimensional systems (e.g. [22]) and translation of thistechnology to a three-dimensional system will likely be problem-atic due to the low porosity of resulting three-dimensionalconstructs. Sacrificial template methods provide an architecturesimilar to native nerve and offer the potential for a more porous
system. Existing needs for materials formed by this approachinclude achieving sufficient porosity to support robust axonalinfiltration, provision of mechanical properties comparable tonative nerve, and an understanding of the effect of the conduitdiameters on rates of axonal migration.
In these studies, we developed a fibrin-based construct in aneffort to begin to address these questions. Fibrin was selectedbecause it is one of the earliest components of the native nerverepair process [41,42], and several groups have used this material asa filler for nerve cuff technology to enhance regeneration [43e46]and/or deliver neurotrophic factors [47,48] or cells [49]. In addi-tion, the use of a sphere-templating approach to fabricate scaffoldsfrom this material has previously been reported [25].
We fabricated different scaffolds with a range of conduitdiameters in an effort to identify the relationship between conduitdiameter and rate of axon regeneration in this three-dimensionalsystem. This was done because although the diameter (caliber) ofthe endoneurium is known for mature native nerve, it is not clearthat these diameters are optimal for a regenerative processfollowing injury.

J.B. Scott et al. / Biomaterials 32 (2011) 4830e48394838
The scaffolds fabricated by the sacrificial fiber approach in thecurrent study had a physical resemblance similar to native nervetissue following decellularization or post-injury Wallerian degen-eration (compare Fig. 2 with [31]). This approach therefore offersthe ability to achieve important structureefunction relationships ofnative tissue while optimizing the system for regeneration.Although contraction during processing for SEM was observed forlarger diameter fiber templates (see Fig. 2), it is clear that conduitdiameter was directly related to the fiber template diameter.Further, the conduits were highly aligned and SEM images suggestthat the conduits spanned the length of the scaffolds. We also notethat there was no observed impediment to axon infiltration and wewere able to visually observe, for the large diameters, that conduitsran the full length of the scaffold.
As a control for the role of alignment, we also incorporatedporosity into the scaffolds by the use of spherical templates [25]that created a more tortuous path than the aligned fibertemplates. The more tortuous spherical templates resulted in axoninfiltration into the scaffolds, but with a rate of extension signifi-cantly less than the less tortuous aligned scaffold conduits (300 mmper day compared to 600e900 mm per day). These resultsdemonstrate the importance of non-tortuous paths for axonalextension in three dimensional scaffolding systems and areconsistent with previous results indicating that oriented patternsare superior in directing rapid axon extension in two-dimensionalsystems [50].
Importantly, the scaffolds were also highly porous, with porosityvalues of approximately 80% regardless of the template fiberdiameter. In addition, the mechanical properties of these scaffolds(Fig. 5) were similar to native nerve, with an elastic modulus ofapproximately 100e330 kPa compared to native nerve with anelastic modulus of 580 kPa [35]. This combination of a highlyporous scaffold material with mechanical properties similar tonative tissue is of considerable importance for several reasons.Although the fibrin hydrogel alone has goodmechanical properties,the porosity provided by the conduits is necessary to allow themigration of axons through the material. In addition, the materialhas sufficient mechanical strength to prevent collapse of theconduits during handling, indicating the ability to handle thesescaffolds within a surgical setting. Lastly, the similarity of themechanical properties of these scaffolds relative to native nervewould likely minimize scarring that could occur due to mechanicalmismatch at site of implantation. Thus, the fibrin scaffolds fabri-cated by this approach have material properties suitable andfavorable for nerve grafting applications.
We evaluated the rate of axon migration into the scaffolds asa function of conduit diameter by immunohistochemistry withhistomorphometry. For these in vitro studies we employed anembryonic chicken DRG model. This model was well-suited tothese studies as the size of the DRG allowed them to be placed onthe top of the scaffold (similar to the proximal end of a nerve injury)for all conduit diameters tested. Dissociation of DRG to isolateneuronal cells was not feasible due to the small size of the neuronsand the large diameter of some conduits. We therefore elected touse whole DRG explants for these studies. To focus on the axonextension through the conduits in response to conduit diameters,we included uridine and FDU in the culture medium to inhibitmitotic growth of Schwann cells. We observed during pilot studiesthat the smaller diameter conduits (e.g., 12 and 19 mm) showedlower numbers of Schwann cells because these cells were excludedby the diameter of the conduits. To achieve similar trophic condi-tions for all scaffold types, we therefore inhibited Schwann cellmitosis with uridine and FDU. While Schwann cells play a key rolein the functional nerve recovery in vivo, the hydrogel nature ofthese scaffolds offers the potential to provide exogenous trophic
support through controlled release until Schwann cells can ulti-mately associate with the regenerated axons. We reason that ifmore rapid axon extension can be achieved in vivo this will providemore favorable long-term functional recovery. Aprotinin was alsoadded to the culture to prevent degradation of the scaffold bymatrix metalloproteinases (MMPs) [49]. We also added exogenousNGF to the media to stimulate axonal extension from the DRG. Werecognize that future strategies utilizing this approach may requiremeans to prevent proteolytic degradation of fibrin and to providecontrolled release of growth factors to promote axonal extension.We have found that the hydrogel nature of these scaffolds iscompatiblewith the slow release of small molecule proteins such asaprotinin and growth factors (data not shown).
We evaluated themaximum distance of axonmigration throughthe scaffolds as reported by others [21]. Other approaches toquantify number of axons were not used because of variabilityobserved in DRG spreading on the scaffold surfaces (e.g., see Fig. 2).
A key finding from these studies was that no significant differ-ences in axon length with time were observed over the wide rangeof conduit diameters studied. We had hypothesized that smallerdiameter conduits would lead to increased rates of axonal exten-sion based on previous studies that indicated changes in length ofaxon extension at a given time point (indicating differences in therate of extension [21,22]). Our results may indicate that physicalshape of the curvature observed by axonal growth cones (i.e.,convex for a fiber compared to concave for a conduit) affects axonalmigration. The results could also indicate that the porosity of thematerial, such as the conduit scaffolds used in these studies, isa more important parameter than the geometrical dimensions ofthe physical guidance cues. We speculate that one advantage to thesmaller diameter conduits is that they may prevent infiltration ofinflammatory cells (e.g., neutrophils andmonocytes/macrophages),allowing unimpeded infiltration of axons.
We note that other studies looking at effects of geometricalparameters on axonmigration have utilized different time points (5or 7 days) rather than the 3 time points in our studies [21,22]. It isconceivable that differences could occur at longer time points inour system. However, because we observedmultiple time points (1,2, and 3 days), we can see the linear rate of axon migration, indi-cating that similar results would be expected at 5 or 7 days.Unfortunately a direct comparison to these other studies was notpossible. Although the fibrin scaffolds can be fabricated in anylength, the mechanical properties were not sufficient to allow theiruse in the culture system used in our study; that is, the scaffolds didnot have sufficient rigidity to remain upright when glued to theculture plate. Wen and Tresco were able to evaluate at longer timepoints, presumably because the more rigid nature of their scaffoldsallowed them to employ longer scaffolds in a similar culture system[21]. It is also noteworthy that in the three-dimensional fibrinsystem in our study the axon extension length was approximately1500 mm at 3 days (w500 mm/day), compared to Wang et al, whosaw extension of 1400 mm over 5 days (w280 mm/day) in the samechick DRG system. However, differences in culture conditions andDRG model make direct comparisons to other studies challenging[21,22].
5. Conclusions
In these studies, we have described an approach to the fabri-cation of fibrin scaffolds suitable for peripheral nerve repairapplications. Although the sacrificial templating approach shouldbe compatible with other materials, the mechanical propertiesachieved with fibrin scaffolds are comparable to native nerve andare expected to support axonal migration due to the role of fibrin inthe natural repair process following nerve injury. An important

J.B. Scott et al. / Biomaterials 32 (2011) 4830e4839 4839
finding from these studies was that the use of highly alignedconduits supports more rapid axon migration through the scaffoldsthan tortuous, non-aligned pores. However, in contrast to two-dimensional fiber-based systems, the diameters in these three-dimensional scaffolds were not found to significantly affect the rateof axon migration through the materials. The approach used for thefabrication of these scaffolds represent a useful means by which toincorporate physical guidance cues and mechanical integrity intobiomaterials useful for peripheral nerve repair.
Acknowledgments
This work was funded by the Wake Forest University HealthSciences Venture Fund, the Department of Biomedical Engineering,and the Wake Forest Institute for Regenerative Medicine. Theauthors gratefully acknowledge the laboratory of Dr. CarolanneMilligan, Wake Forest University Health Sciences Department ofNeurobiology and Anatomy, for assistance with isolation andculture of embryonic chicken DRG.
References
[1] Evans GR. Peripheral nerve injury: a review and approach to tissue engineeredconstructs. Anat Rec 2001;263:396e404.
[2] Nichols CM, Brenner MJ, Fox IK, Tung TH, Hunter DA, Rickman SR, et al. Effectsof motor versus sensory nerve grafts on peripheral nerve regeneration. ExpNeurol 2004;190:347e55.
[3] Noble J, Munro CA, Prasad VS, Midha R. Analysis of upper and lower extremityperipheral nerve injuries in a population of patients with multiple injuries.J Trauma 1998;45:116e22.
[4] Taras JS, Nanavati V, Steelman P. Nerve conduits. J Hand Ther 2005;18:191e7.[5] Siemionow M, Bozkurt M, Zor F. Regeneration and repair of peripheral nerves
with different biomaterials: review. Microsurgery 2010;30:574e88.[6] Whitlock EL, Tuffaha SH, Luciano JP, Yan Y, Hunter DA, Magill CK, et al. Pro-
cessed allografts and type I collagen conduits for repair of peripheral nervegaps. Muscle Nerve 2009;39:787e99.
[7] Larsen M, Habermann TM, Bishop AT, Shin AY, Spinner RJ. Epstein-Barr virusinfection as a complication of transplantation of a nerve allograft from a livingrelated donor. Case report. J Neurosurg 2007;106:924e8.
[8] Champney TH. A proposal for a policy on the ethical care and use of cadaversand their tissues. Anat Sci Educ 2011;4:49e52.
[9] Madison RD, da Silva C, Dikkes P, Sidman RL, Chiu TH. Peripheral nerveregeneration with entubulation repair: comparison of biodegradeable nerveguides versus polyethylene tubes and the effects of a laminin-containing gel.Exp Neurol 1987;95:378e90.
[10] Midha R, Munro CA, Dalton PD, Tator CH, Shoichet MS. Growth factorenhancement of peripheral nerve regeneration through a novel synthetichydrogel tube. J Neurosurg 2003;99:555e65.
[11] Labrador RO, Buti M, Navarro X. Influence of collagen and laminin gelsconcentration on nerve regeneration after resection and tube repair. ExpNeurol 1998;149:243e52.
[12] Sierpinski P, Garrett J, Ma J, Apel P, Klorig D, Smith T, et al. The use of keratinbiomaterials derived from human hair for the promotion of rapid regenera-tion of peripheral nerves. Biomaterials 2008;29:118e28.
[13] Yu TT, Shoichet MS. Guided cell adhesion and outgrowth in peptide-modifiedchannels for neural tissue engineering. Biomaterials 2005;26:1507e14.
[14] Deister C, Aljabari S, Schmidt CE. Effects of collagen 1, fibronectin, laminin andhyaluronic acid concentration in multi-component gels on neurite extension.J Biomater Sci Polym Ed 2007;18:983e97.
[15] Suri S, Schmidt CE. Cell-laden hydrogel constructs of hyaluronic acid, collagen,and laminin for neural tissue engineering. Tissue Eng Part A 2010;16:1703e16.
[16] Dodla MC, Bellamkonda RV. Differences between the effect of anisotropic andisotropic laminin and nerve growth factor presenting scaffolds on nerveregeneration across long peripheral nerve gaps. Biomaterials 2008;29:33e46.
[17] Yu X, Bellamkonda RV. Tissue-engineered scaffolds are effective alternativesto autografts for bridging peripheral nerve gaps. Tissue Eng 2003;9:421e30.
[18] Nguyen QT, Sanes JR, Lichtman JW. Pre-existing pathways promote preciseprojection patterns. Nat Neurosci 2002;5:861e7.
[19] Hudson TW, Liu SY, Schmidt CE. Engineering an improved acellular nervegraft via optimized chemical processing. Tissue Eng 2004;10:1346e58.
[20] Hudson TW, Zawko S, Deister C, Lundy S, Hu CY, Lee K, et al. Optimizedacellular nerve graft is immunologically tolerated and supports regeneration.Tissue Eng 2004;10:1641e51.
[21] Wen X, Tresco PA. Effect of filament diameter and extracellular matrixmolecule precoating on neurite outgrowth and Schwann cell behavior onmultifilament entubulation bridging device in vitro. J Biomed Mater Res A2006;76:626e37.
[22] Wang HB, Mullins ME, Cregg JM, McCarthy CW, Gilbert RJ. Varying thediameter of aligned electrospun fibers alters neurite outgrowth and Schwanncell migration. Acta Biomater 2010;6:2970e8.
[23] Kim YT, Haftel VK, Kumar S, Bellamkonda RV. The role of aligned polymerfiber-based constructs in the bridging of long peripheral nerve gaps. Bioma-terials; 2008.
[24] Clements IP, Kim YT, English AW, Lu X, Chung A, Bellamkonda RV. Thin-filmenhanced nerve guidance channels for peripheral nerve repair. Biomaterials2009;30:3834e46.
[25] Linnes MP, Ratner BD, Giachelli CM. A fibrinogen-based precision micropo-rous scaffold for tissue engineering. Biomaterials 2007;28:5298e306.
[26] Flynn L, Dalton PD, Shoichet MS. Fiber templating of poly(2-hydroxyethylmethacrylate) for neural tissue engineering. Biomaterials 2003;24:4265e72.
[27] Gros T, Sakamoto JS, Blesch A, Havton LA, Tuszynski MH. Regeneration oflong-tract axons through sites of spinal cord injury using templated agarosescaffolds. Biomaterials 2010;31:6719e29.
[28] Wei G, Ma PX. Macroporous and nanofibrous polymer scaffolds and polymer/bone-like apatite composite scaffolds generated by sugar spheres. J BiomedMater Res A 2006;78:306e15.
[29] Stokols S, Sakamoto J, Breckon C, Holt T, Weiss J, Tuszynski MH. Templatedagarose scaffolds support linear axonal regeneration. Tissue Eng 2006;12:2777e87.
[30] Bellamkonda RV. Peripheral nerve regeneration: an opinion on channels,scaffolds and anisotropy. Biomaterials 2006;27:3515e8.
[31] Lee SK, Wolfe SW. Peripheral nerve injury and repair. J Am Acad Orthop Surg2000;8:243e52.
[32] Leon A, Benvegnu D, Dal Toso R, Presti D, Facci L, Giorgi O, et al. Dorsal rootganglia and nerve growth factor: a model for understanding the mechanismof GM1 effects on neuronal repair. J Neurosci Res 1984;12:277e87.
[33] Osathanon T, Linnes ML, Rajachar RM, Ratner BD, Somerman MJ, Giachelli CM.Microporous nanofibrous fibrin-based scaffolds for bone tissue engineering.Biomaterials 2008;29:4091e9.
[34] Webb PA. An introduction to the physical characterization of materials bymercury intrusion porosimetry with emphasis on reduction and presentationof experimental data. Micrometrics Instrument Corp; 2001:1e9.
[35] Borschel GH, Kia KF, Kuzon Jr WM, Dennis RG. Mechanical properties ofacellular peripheral nerve. J Surg Res 2003;114:133e9.
[36] Mieszawska AJ, Kaplan DL. Smart biomaterials - regulating cell behaviorthrough signaling molecules. BMC Biol 2010;8:59.
[37] Bozkurt A, Brook GA, Moellers S, Lassner F, Sellhaus B, Weis J, et al. In vitroassessment of axonal growth using dorsal root ganglia explants in a novelthree-dimensional collagen matrix. Tissue Eng 2007;13:2971e9.
[38] Wang DY, Huang YY. Fabricate coaxial stacked nerve conduits through softlithography and molding processes. J Biomed Mater Res A 2008;85:434e8.
[39] Lacour SP, Fitzgerald JJ, Lago N, Tarte E, McMahon S, Fawcett J. Long micro-channel electrode arrays: a novel type of regenerative peripheral nerveinterface. IEEE Trans Neural Syst Rehabil Eng 2009;17:454e60.
[40] Lee W, Lee V, Polio S, Keegan P, Lee JH, Fischer K, et al. On-demand three-dimensional freeform fabrication of multi-layered hydrogel scaffold withfluidic channels. Biotechnol Bioeng 2010;105:1178e86.
[41] Williams LR. Exogenous fibrin matrix precursors stimulate the temporalprogress of nerve regeneration within a silicone chamber. Neurochem Res1987;12:851e60.
[42] Williams LR, Danielsen N, Muller H, Varon S. Exogenous matrix precursorspromote functional nerve regeneration across a 15-mm gap within a siliconechamber in the rat. J Comp Neurol 1987;264:284e90.
[43] Kalbermatten DF, Pettersson J, Kingham PJ, Pierer G, Wiberg M, Terenghi G.New fibrin conduit for peripheral nerve repair. J Reconstr Microsurg 2009;25:27e33.
[44] Kalbermatten DF, Kingham PJ, Mahay D, Mantovani C, Pettersson J, Raffoul W,et al. Fibrin matrix for suspension of regenerative cells in an artificial nerveconduit. J Plast Reconstr Aesthet Surg 2008;61:669e75.
[45] Nakayama K, Takakuda K, Koyama Y, Itoh S, Wang W, Mukai T, et al.Enhancement of peripheral nerve regeneration using bioabsorbable polymertubes packed with fibrin gel. Artif Organs 2007;31:500e8.
[46] Nakayama K, Takakuda K, Koyama Y, Itoh S, Wang W, Shirahama N, et al.Regeneration of peripheral nerves by bioabsorbable polymer tubes with fibringel. J Nanosci Nanotechnol 2007;7:730e3.
[47] Wood MD, Moore AM, Hunter DA, Tuffaha S, Borschel GH, Mackinnon SE,et al. Affinity-based release of glial-derived neurotrophic factor from fibrinmatrices enhances sciatic nerve regeneration. Acta Biomater 2009;5:959e68.
[48] Johnson PJ, Parker SR, Sakiyama-Elbert SE. Controlled release of neurotrophin-3 from fibrin-based tissue engineering scaffolds enhances neural fibersprouting following subacute spinal cord injury. Biotechnol Bioeng 2009;104:1207e14.
[49] Hurtado A, Moon LD, Maquet V, Blits B, Jerome R, Oudega M. Poly (D, L-lacticacid) macroporous guidance scaffolds seeded with Schwann cells geneticallymodified to secrete a bi-functional neurotrophin implanted in the completelytransected adult rat thoracic spinal cord. Biomaterials 2006;27:430e42.
[50] Cao H, Liu T, Chew SY. The application of nanofibrous scaffolds in neural tissueengineering. Adv Drug Deliv Rev 2009;61:1055e64.





























