Embed Size (px)
Citation preview

Behaviour (2013) DOI:10.1163/1568539X-00003096 brill.com/beh
The evolution of behavioural systems:a study of grooming in rodents
J. Malange a,b,∗, C.C. Alberts c, E. Spinelli Oliveira a,d and H.F. Japyassú a,b
a Programa de Pós-Graduação em Neurociências e Comportamento, NeC,Instituto de Psicologia, Universidade de São Paulo, São Paulo, Brazil
b Núcleo de Etologia e Evolução, Instituto de Biologia,Universidade Federal da Bahia, Bahia, Brazil
c Universidade Estadual Paulista, São Paulo, Brazild Departamento de Biologia, Faculdade de Filosofia, Universidade de São Paulo,
Ciências e Letras de Ribeirão Preto, São Paulo, Brazil*Corresponding author’s e-mail address: [email protected]
Accepted 7 May 2013
AbstractIn the present paper we focus on the study of complex behavioural systems, within an explicit phy-logenetic framework. We reconstruct the phylogeny of rodents using grooming sequences from 12terminals. Using a method derived from graph theory, we decompose complex behavioural systemsinto strings of behavioural units (behavioural routines) which are then used as behavioural charac-ters to compose the phylogenetic matrix in addition to three mitochondrial markers as molecularcharacters (the cytochrome b gene (cytb), the 16S ribosomal RNA gene and the 12S ribosomalRNA gene). Our results point to a highly structured behavioural morphospace: only a few char-acters have been selected for, within the total space of possibilities. The optimization of hundredsof non-homoplastic routines onto three distinct phylogenies (behavioural, combined data and themolecular supertree of Fabre et al., 2012) reveals the same evolutionary trend from simple tocomplex: while simple behavioural routines (zero- or first-order sequences) are synapomorphiesat basal levels of the phylogeny, progressively more complex behaviours evolve later, appearingcloser to the tips of the phylogeny. Also, the optimization shows that the organisation of unitsinto modules of coordinated action patterns first evolved around large body parts, namely the headand the trunk, modules that were later fused into one single organising module among rodents.We support the use of complex behavioural systems as a promising tool in the study of evolution-ary scenarios and discuss the role of routines length and microstructure to provide phylogeneticinformation and elucidate evolutionary processes.
Keywords
Grooming, evolution, behavioural systems, rodents.
© 2013 Koninklijke Brill NV, Leiden DOI:10.1163/1568539X-00003096

2 Behaviour (2013) DOI:10.1163/1568539X-00003096
1. Introduction
Complex behavioural systems are not as easy to study in a comparativeperspective as are simple, reflex-like units of behaviour. When we add com-plexity to a system, it usually shows more context sensitivity, a property thatis not much appreciated in phylogenetic studies. This is probably at the rootof the diminishing numbers of papers dealing with phylogenetic informationin behavioural journals (Price et al., 2011).
Extreme context sensitivity is probably the reason behind the long stand-ing controversy regarding the use of behavioural data to estimate phylo-genies, a controversy revolving mainly around the alleged inherent plas-ticity of behaviour and the inapplicability of the homology criteria to thisevanescent database (Lorenz, 1941; Atz, 1970; Baroni Urbani, 1989). Therehave been many successful empirical studies deploying phylogenetical rela-tionships from behaviour (McLennan, 1993; Paterson et al., 1995; Alberts,1996; Slikas, 1998; Stuart & Hunter, 1998; Bucheli et al., 2002; Noll, 2002;Desutter-Grandcolas & Robillard, 2003; Robillard et al., 2006; Cap et al.,2008), but these studies are mostly concerned with simple behavioural pat-terns, simple units of species typical behaviour.
The success of the use of simple, reflex-like behavioural units within anexplicitly phylogenetic context has encouraged the use of other, more com-plex aspects of the performance, such as the sequential organisation of theseunits (Robillard et al., 2006; Legendre et al., 2008), but this is still a quite un-usual approach. Even more unusual is the simultaneous use of simple units,short and long sequences of behaviour in a phylogenetic context; such anapproach could possibly give a picture of the evolution of complexity withinbehavioural systems. It would allow one to answer questions regarding possi-ble evolutionary trends from simple to complex systems, or vice versa. Also,it would help to settle the question of the suitability of behaviour as a phy-logenetic character to its highest level: if even complex behavioural systems(as for example long sequences of behaviour) prove to be phylogeneticallyinformative, then the relationship between behavioural plasticity and evolu-tionary processes (West-Eberhard, 2003, pp. 337–352) must be readdressed.In the present paper we deal with these questions tracing the evolution ofvarying levels of behavioural complexity in the phylogeny of rodents.
Rodents have a quite long history of phylogenetic reconstructions. Somemolecular studies have considered them as a polyphyletic group (Graur etal., 1991; Li et al., 1992; D’Erchia et al., 1996; Reyes et al., 2000; Adkins,

J. Malange et al. / Behaviour (2013) 3
2001, 2003; Arnason et al., 2002; Lin et al., 2002), while other studies havefavoured their monophyly (Martignetti & Brosius, 1993; Quintana, 1998;da Silva Neto, 2000; Reyes et al., 2000; Adkins, 2001, 2003; Huchon &Douzery, 2001; Arnason et al., 2002; Lin et al., 2002; Rowe & Honeycutt,2002; Reyes et al., 2004; Narita et al., 2006; Poux et al., 2006). Nevertheless,recent papers have converged on the monophyly hypothesis (Blanga-Kanfi etal., 2009; Fabre et al., 2012). In this paper, we present grooming as a new,potentially rich database for rodent phylogeny, and explore this database inunder sampled rodent taxa.
Grooming is a well-defined and described behaviour (Eisenberg, 1963;Roth & Rosenblatt, 1966, 1967) that has been studied with regard to itsfunction (Cromwell & Berridge, 1996; Shanas & Terkel, 1996; Ferkin etal., 2001; Greer & Capecchi, 2002; Wolff et al., 2002; Kalueff & Tuohimaa,2004; Berridge et al., 2005; Hawlena et al., 2007), hierarchical (Fentress,1972, 1973, 1987; Dawkins & Dawkins, 1976) and sequential organisa-tion (Berridge, 1990; Colonnese et al., 1996; Bursten et al., 2000), andalso with regard to its evolutionary pattern (Farish, 1972; Sachs, 1988;Berridge, 1990). Grooming occurs among vertebrates and invertebrates,and seems to be an old and evolutionarily conservative behavioural pattern(Berridge, 1990; Colonnese et al., 1996). Despite this evolutionary conser-vatism, grooming units and their sequential organisation show enough vari-ability as to be potentially informative for phylogenetic studies (Dawkins &Dawkins, 1976; Berridge, 1990; Bursten et al., 2000).
When dealing with behavioural sequences, it is desirable to take intoconsideration the probabilistic nature of this kind of data, a condition thathas been recently treated by the direct optimization of small behaviouralsequences (Japyassú & Machado, 2010). The use of longer behavioural se-quences would be welcome, since it would result in an exponentially largernumbers of phylogenetic characters (Japyassú & Viera, 2002), but the ex-tension of this direct optimization method to longer behavioural sequencesis computationally intensive, and has not been developed up to now. In thispaper we follow a different analytical procedure to circumvent the compu-tational difficulties of the direct optimization approach. We use a methodderived from graph theory to extract behavioural sequences that will besubsequently used as characters in a phylogenetic analysis (Alberts, 1996;Japyassú et al., 2006). We focus on sequences of an easily observable be-haviour (self-grooming) in order to understand the evolution of this complex

4 Behaviour (2013) DOI:10.1163/1568539X-00003096
behavioural system, mapping the behavioural characters in three distinct treetopologies.
2. Methods
2.1. Behavioural data
Grooming is a well-known behaviour, and its main units, as described forRattus norvegicus (Silverman, 1978), are ‘wash’, ‘groom’, ‘scratch’ and ‘lickpenis’ (urinary papilla). We complemented this behavioural catalogue withother units obtained from a review of the literature and from preliminaryobservations of the animals under study, so that the new catalogue comprisesa total of 27 behavioural units (Appendix A). We did not consider variationsin the form of each behavioural unit because we focus not on the evolutionof the units themselves, but rather on the evolution of the routines, that is, ofthe probabilistic sequences of the units.
A total of 42 specimens comprising 12 terminals were videotaped.The terminals analyzed were Calomys callosus Rengger, 1830 (2�, 2�),Mesocricetus auratus Waterhouse, 1839 (2�, 1�) and Meriones unguicu-latus Milne-Edwards, 1867 (1�, 2�), of the superfamily Muroidea; Caviaaperea Erxleben, 1777 (3�, 2�), C. intermedia Cherem, Olimpio e Ximenez,1999 (3�, 4�) and C. porcellus Linnaeus, 1758 (1�, 2�), of the family Cavi-idae; Trinomys yonenagae Rocha, 1995 (1�, 1�), Thrichomys pachyurusWagner, 1845 (1�, 1�) and T. laurentius Thomas, 1904 (2�, 2�), of thefamily Echimyidae. Specimens of the species T. laurentius were further sub-divided in two distinct terminals, because they exhibit different behaviouraland morphometrical features (Reis et al., 2002; Basile, 2003; Mendes et al.,2004), and also because they come from isolated populations of differentbiomes and distant geographic areas, one from São Raimundo Nonato (Pi-auí; 1� and 1�; 09°00′55′′S, 42°41′58′′W) and another from Caetité (Bahia;1� and 1�; 14°04′10′′S, 42°28′30′′W). Felis silvestres f. catus Linnaeus,1758 (4�, 3�) (order Carnivora) and Oryctolagus cuniculus Linnaeus, 1758(1�, 2�) (order Lagomorpha) constituted outgroups in the analysis.
C. callosus, M. auratus, M. unguiculatus, C. aperea, C. porcellus, T. yone-nagae, T. pachyurus, T. laurentius and O. cuniculus were kept under standardlaboratory conditions with controlled climate parameters (21–24°C, 45–55%humidity and 12/12 h light/dark cycle). All animals were provided with foodand water ad libitum. The rodents were placed in acrylic cages (41 × 34 ×

J. Malange et al. / Behaviour (2013) 5
20 cm). These cages were kept at the laboratory where the animals belonged(Adolfo Lutz Institute at São Paulo; Psychology Institute and FFCLRP at theUniversity of São Paulo; Oswaldo Cruz Institute at Rio de Janeiro). C. in-termedia were filmed at their natural habitat (Moleques do Sul Island, SantaCatarina State, Brazil, 27°51′S, 48°26′W). F. catus were also filmed at theirnatural, domestic habitat (in São Paulo city, Brazil).
Grooming was videotaped in focal animal method (Lehner, 1996). Wesampled a total of approximately 75 h, resulting in nearly 2000 events foreach terminal. All terminals reached a plateau when plotted on accumulationcurves, revealing a sufficient and comparable sampling effort (Appendix B).The sessions were transcribed with the aid of the program EthoLog (Ottoni,2000).
2.2. Obtaining behavioural routines
First-order transition matrices were analysed using the software EthoSeq,available online at http://www2.assis.unesp.br/cats/ethoseq.htm. The detailsof the software algorithm are described elsewhere (Japyassú et al., 2006),but below we provide a brief description of the main steps in the graphanalysis. This graph analysis results in probabilistic behavioural sequences(routines). In order to obtain these routines, EthoSeq builds a first-order tran-sition matrix from the raw sequence data, for each species. Next, it searches,in each matrix, for the most probable directed hierarchical graph, given thedyadic association between the behavioural units in the transition matrix.Behavioural units are the vertices (V), and transition probabilities the arrows(A), in each of the possible directed hierarchical graphs G(V, A) present inthe transition matrix. Each behavioural unit appears only once in a G(V, A),which connects all units in one single hierarchy. EthoSeq simply chooses themost probable hierarchical graph for a given matrix and a given initiating be-haviour (the root, r), and it does so by maximizing the transition probabilitiesbetween behavioural units, with the function:
P(r, . . . , l) = P(r)∏
(i,j)∈(r,...,l)
C(i, j),
where P(r) is the unconditional probability of the behavioural unit r , at theroot of the hierarchical graph; C(i, j) is the conditional probability of thebehavioural unit i given the unit j ; P(r, . . . , l) is the probability of the linearbranch from the behavioural units r through l (from the root to the tip of thehierarchy). The routines then are the set of these linear branches within themost probable graphs found for each species.

6 Behaviour (2013) DOI:10.1163/1568539X-00003096
2.3. Phylogenetic analyses
The behavioural phylogenetic matrix with informative routines (available assupplementary material in the online edition of this journal, which can be ac-cessed via http://booksandjournals.brillonline.com/content/1568539x) wasanalysed using the software TNT (v. 1.1, Goloboff, Farris Nixon, 2008), us-ing heuristic search (RAS + TBR) parameters, 10 replications, to search themost parsimonious tree. Bootstrap (Felsenstein, 1985) and jackknife (Farriset al., 1996) were used as measures of clade support (100 replications).
In order to investigate the association between the size of the routine andits position (from the root to the tips of the tree) as a synapomorphy on thephylogeny, we performed a Spearman’s correlation test (Spearman, 1904).
For the molecular analysis we used three mitochondrial markers, the cy-tochrome b gene (cytb) (approximately 1140 bp), the 16S ribosomal RNAgene (approximately 1560 bp) and the 12S ribosomal RNA gene (approxi-mately 1020 bp). The molecular data were obtained from GenBank (Table 1).While for cytb most species presented the complete sequence, for 16S and12S only partial sequences were obtained, or no data at all.
To align molecular sequences and to reconstruct the phylogeny using mi-tochondrial data via dynamic optimization we employed the POY software,version 3.0.11 (Wheeler, 2003). The molecular data undergone a completesearch and for the jackknife analysis the command jackboot was used withthe 100 replicates parameter.
Table 1.GenBank accession numbers for the molecular data according to each taxon and to eachmitochondrial gene used on this study.
Species cytb 16S 12S
Felis catus AB194812.1 DQ334823.1 AY012149.1Oryctolagus cuniculus AY292717.1 DQ334838.1 AY292691.1Thrichomys apereoides laurentius AY083332.1 n/a n/aThrichomys pachyurus AY083329.1 n/a n/aTrinomys yonenagae AF194295.1 AF422899.1 AF422865.1Meriones unguiculatus AB381901.1 EU078984.1 AJ851249.1Calomys callosus DQ447282.1 n/a n/aMesocricetus auratus AJ973379.1 DQ334843.1 X84390.1Cavia aperea GU136743.1 n/a AF433908.1Cavia porcellus DQ017040.1 DQ334847.1 AF433909.1
n/a, not available.

J. Malange et al. / Behaviour (2013) 7
We also used POY to reconstruct a phylogeny based on the concomitantuse of both molecular and behavioural data employing the program’s Com-bined Analysis.
3. Results
3.1. Phylogeny of rodents
From the 6083 grooming routines, 2832 were phylogenetically informative.The analysis of the character matrix with these informative routines (seesupplementary material in the online edition of this journal, which can beaccessed via http://booksandjournals.brillonline.com/content/1568539x) re-sulted in one single most parsimonious tree (Figure 1).
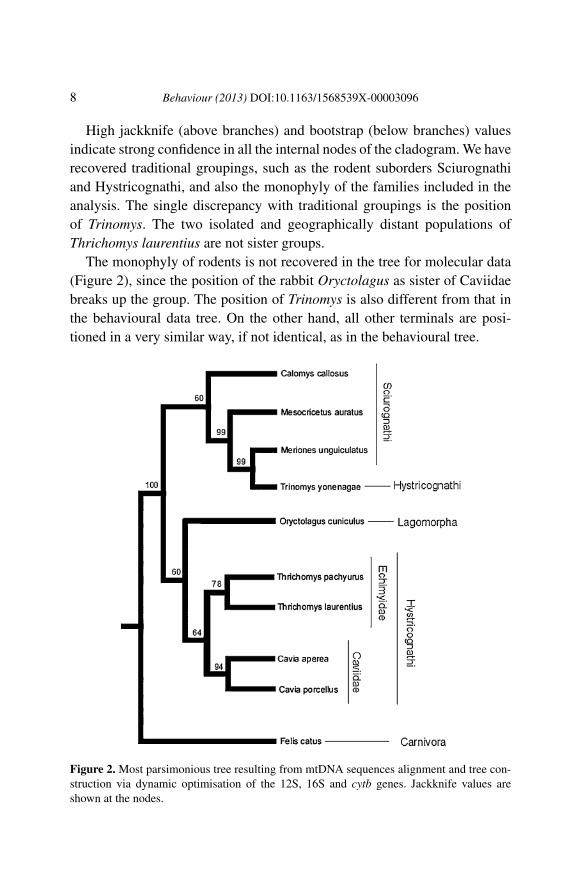
Figure 1. Single most parsimonious tree resulting from the analysis of 2832 informativegrooming sequences (6970 steps; CI = 40; RI = 31). Jackknife and bootstrap values areshown above and below the branches, respectively.
8 Behaviour (2013) DOI:10.1163/1568539X-00003096
High jackknife (above branches) and bootstrap (below branches) valuesindicate strong confidence in all the internal nodes of the cladogram. We haverecovered traditional groupings, such as the rodent suborders Sciurognathiand Hystricognathi, and also the monophyly of the families included in theanalysis. The single discrepancy with traditional groupings is the positionof Trinomys. The two isolated and geographically distant populations ofThrichomys laurentius are not sister groups.
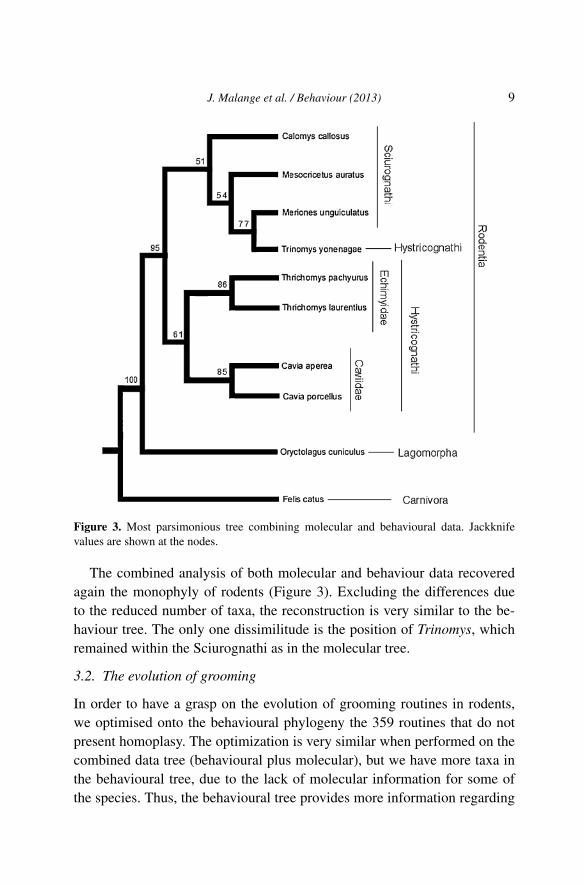
The monophyly of rodents is not recovered in the tree for molecular data(Figure 2), since the position of the rabbit Oryctolagus as sister of Caviidaebreaks up the group. The position of Trinomys is also different from that inthe behavioural data tree. On the other hand, all other terminals are posi-tioned in a very similar way, if not identical, as in the behavioural tree.
Figure 2. Most parsimonious tree resulting from mtDNA sequences alignment and tree con-struction via dynamic optimisation of the 12S, 16S and cytb genes. Jackknife values areshown at the nodes.
J. Malange et al. / Behaviour (2013) 9
Figure 3. Most parsimonious tree combining molecular and behavioural data. Jackknifevalues are shown at the nodes.
The combined analysis of both molecular and behaviour data recoveredagain the monophyly of rodents (Figure 3). Excluding the differences dueto the reduced number of taxa, the reconstruction is very similar to the be-haviour tree. The only one dissimilitude is the position of Trinomys, whichremained within the Sciurognathi as in the molecular tree.
3.2. The evolution of grooming
In order to have a grasp on the evolution of grooming routines in rodents,we optimised onto the behavioural phylogeny the 359 routines that do notpresent homoplasy. The optimization is very similar when performed on thecombined data tree (behavioural plus molecular), but we have more taxa inthe behavioural tree, due to the lack of molecular information for some ofthe species. Thus, the behavioural tree provides more information regarding

10 Behaviour (2013) DOI:10.1163/1568539X-00003096
the evolution of grooming sequences, including the interesting non-sisterrelationship between the two populations of Thrichomys laurentius.
Overall, this analysis shows a striking difference between the groomingbehaviour of rodents and that of the outgroup taxa, and this result holds irre-spective of which of the outgroup taxa we choose to root the cladogram. BothF. catus and O. cuniculus (possible outgroup species) usually perform facialand body grooming in separate behavioural bouts, while rodents mostly per-form complete grooming sequences, involving both the head and the restof the body. For example, in one single bout cats and rabbits groom onlybody parts alternating between licking and biting the flank, and licking andbiting the belly. Rodents do not present this routine, and instead they makeregular use of complete sequences such as rub ear/rub eye/rub vibrissae/lickforelimb/lick back/lick flank/bite flank/lick belly; they perform these rou-tines quickly, especially in the species of the suborder Sciurognathi. Catsand rabbits frequently show long pauses between grooming sequences, whilerodents mostly perform uninterrupted sequences. Thus, it seems that rodentevolution resulted in the unification of two previously distinct behaviouraldomains: the head and the body grooming modules. Notwithstanding thisevolutionary unification of two modules into a single structure, at a largerscale there is still a similarity between the plesiomorphic and the apomorphicorganisation, in the sense that both groups perform grooming sequences in ahead-tail direction.
In the rodent clade, we remark that histricognathids frequently freeze inthe middle of their grooming sequences. For example, histricognathids fre-quently display the routine scratch head/bite hind foot/lick tail/freeze/washanogenital area.
Hystricognathid rodents are also characterised by grooming sequencesintermingled with non-grooming behaviour, as for example, in the routinerub vibrissae/lick forelimb/exploration/walking/exploration/bite belly. Also,hystricognathids have lost many routines that sciurognathids share with theoutgroups. These are usually small routines such as, for example, the routinebite/lick/bite (the belly).
Within sciurognathids, cricetids differ markedly from the other rodents(they present 56 synapomorphies). One major difference is the intense use ofthe hind legs to scratch other body parts, composing routines such as scratchhead/bite hind legs/scratch belly/bite hind legs/scratch flank. Also, besidesusing the usual head-tail overall direction of grooming activities, cricetidsadditionally make use of a dorsoventral flow of behaviours.

J. Malange et al. / Behaviour (2013) 11
3.3. Evolution of complexity in grooming behaviour
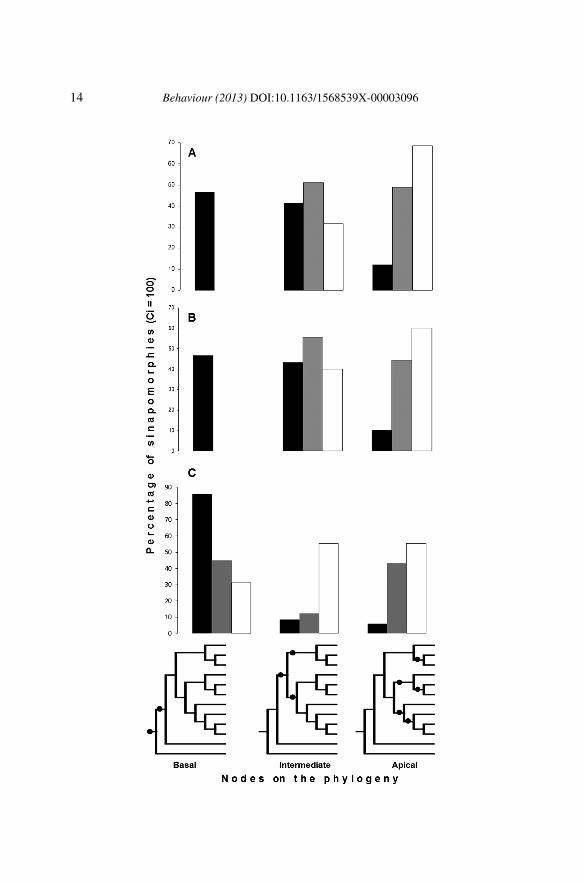
Our main question relates to the evolution of complexity in this behaviouralsystem, which can be assessed by a correlation between the size of theroutine and the level in the phylogeny where this routine appears as a synapo-morphy. In order to evaluate this correlation we selected from the availableroutines the ones without homoplasy (CI = 100, N = 359 for the behaviouralphylogeny, N = 263 for the combined data phylogeny, and N = 388 for thetopology of Fabre et al., 2012), and inspected the level of the three phyloge-nies, from the root to the tip of the tree, where they stuck as synapomorphies(to see synapomorphies for the major clades, check Table 2). In all analy-ses, the distribution of the behavioural characters follow a common, verysimilar pattern: short routines are more informative at the basal nodes of thephylogeny, medium-sized routines inform the intermediate nodes, and largeroutines are synapomorphies mainly at the more distal nodes (near the tips)of the phylogeny (Figure 4). There is a significant positive correlation be-tween the complexity (size) of the routine and its position on the phylogeny(N = 359; p < 0.01; r = 0.244).
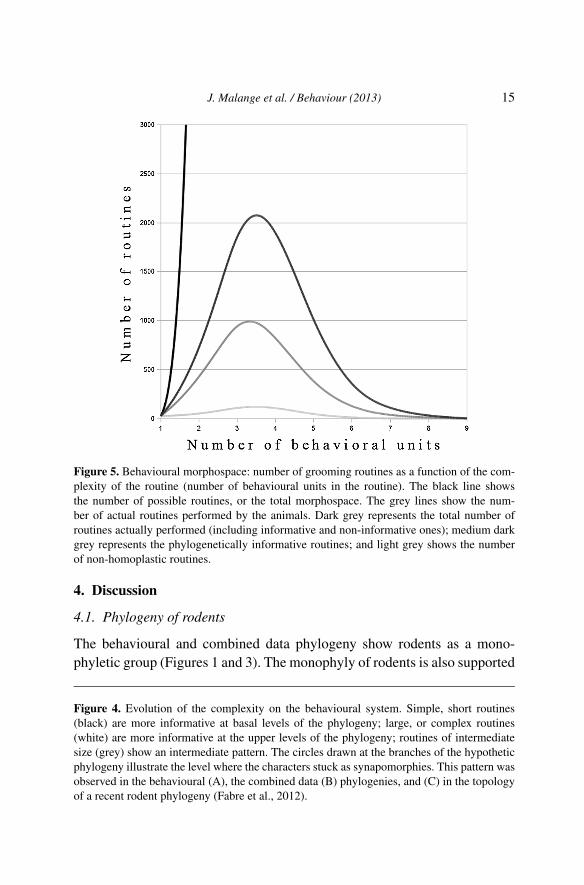
There are routines of all kinds, from sequences of up to nine behaviouralunits connected in a string, to simple dyads (a sequence of only two units)or even monads (zero order sequences, or isolated units). The number ofpossible sequences increases exponentially according to the function R =k(k−1)(n−1), where k is the number of behavioural units present in the wholerepertoire, and n is the size of the sequence (number of behavioural units inthe sequence). There is a strong disconnection between the number of pos-sible sequences and the number of actual routines uncovered by our datamining analyses (Figure 5): from a total of 5.9 × 1012 possible grooming se-quences of up to 9 units (up to 8th-order sequences), EthoSeq has uncoveredonly 6083 grooming routines actually exhibited by rodents.
This result points to a highly structured behavioural morphospace. Thereare actually very few long routines in the repertoire of rodents, what amountsto saying that there are very few long behavioural sequences that are biologi-cally structured. One could wonder if the low number of long routines couldbe due to insufficient sample size. If the reason for the presence or absenceof long routines was low sample size, the resulting sequences mined by thegraph algorithm method would be a collection of randomly assorted routines,with no phylogenetic signal. Instead, our results show that the long routines

12 Behaviour (2013) DOI:10.1163/1568539X-00003096
Table 2.Synaptomorphic characters with consistency index equal to 100% for the major groups in thestudy: Order Rodentia, Suborder Sciurognathi and Suborder Hystricognathi.
(Sub)Order State
Rodentiabit_hindf, bit_forelimb stch_head, bit_hindf, stch_head +bit_forelimb, exploration lick_forelimb, lick_belly, lick_flanc +stch_flanc, exploration lick_forelimb, bit_forelimb, exploration +drinking, walking, exploration exploration, bit_hindf, shake +walking, lick_flanc stch_flanc, exploration, walking, exploration +drinking, wash_anoge bit_forelimb, exploration, walking, exploration +stch_belly, lick_hindf exploration, walking, exploration, bit_forelimb +lick_belly, lick_forelimb, bit_forelimb +pause, lick_belly drinking, lick_forelimb –pause, lick_flanc lick_flanc, exploration –pause, lick_back walking, eating, shake –lick_hindf, pause pause, lick_forelimb, walking –lick_flanc, stopped lick_hindf, walking, exploration –lick_back, pause bit_forelimb, lick_forelimb, shake –lick_flanc, pause shake, eating, shake –lick_back, shake shake, eating, walking –lick_flanc, wash_anoge bit_back, lick_back, lick_hindf –stch_belly, pause lick_back, lick_flanc, bit_flanc –bit_tail, exploration bit_hindf, lick_hindf, walking –bit_forelimb, lick_hindf shake, walking, eating –pause, stch_belly lick_hindf, bit_hindf, exploration, walking –mord_flanc, stch_flanc eating, walking, exploration, shake –bit_tail, lick_tail shake, walking, eating, walking –stopped, lick_forelimb walking, exploration, walking, bit_flanc –bit_hindf, exploration shake, eating, walking, exploration –lick_hindf, lick_belly –
Sciurognathistch_flanc, bit_hindf, bit_forelimb +walking, stch_flanc, exploration +walking, stch_flanc, exploration, walking +lick_hindf, lick_forelimb, rub_eye, rub_vibss +rub_eye, rub_vibss, lick_forelimb, bit_forelimb, lick_forelimb +rub_eye, rub_vibss, lick_forelimb, exploration, walking +rub_vibss, lick_forelimb, bit_forelimb, exploration, walking +lick_forelimb, rub_eye, rub_vibss, lick_forelimb, wash_anoge +rub_eye, rub_vibss, lick_forelimb, exploration, walking, exploration +rub_vibss, lick_forelimb, rub_eye, rub_vibss, lick_forelimb, lick_belly +rub_eye, rub_vibss, lick_forelimb, pause, lick_forelimb, rub_eye +

J. Malange et al. / Behaviour (2013) 13
Table 2.(Continued.)
(Sub)Order State
walking, exploration, lick_forelimb, pause, lick_forelimb, rub_vibss +eating, shake –shake, stch_head –stopped, shake –shake, stopped –bit_forelimb, lick_forelimb, exploration –exploration, eating, shake –rub_eye, rub_vibss, lick_forelimb, shake –
Hystricognathistch_head, stopped +bit_forelimb, exploration, stopped +stopped, bit_forelimb, stopped +stopped, stch_head, stopped +stch_head, bit_hindf, stopped +bit_forelimb, wash_anoge, pause +wash_anoge, exploration, shake +bit_hindf, exploration, stopped +lick_forelimb, exploration, stch_flanc +exploration, bit_forelimb, shake +pause, bit_forelimb, pause +bit_forelimb, stopped, rub_vibss +pause, bit_forelimb, lick_forelimb +bit_hindf, rub_vibss, lick_forelimb, bit_forelimb +exploration, rub_vibss, lick_forelimb, bit_forelimb +stch_head, bit_hindf, shake, walking +wash_anoge, exploration, shake, exploration +bit_hindf, stch_head, bit_hindf, stopped +stopped, exploration, lick_forelimb, exploration +bit_belly, lick_belly –lick_belly, bit_belly –lick_flanc, lick_hindf, lick_forelimb –
The behavioural characters where optimised onto the topology of Fabre et al. (2012). Thestates of the characters mean presence (+) or absence (–).
found are the very synapomorphies that shape our behavioural reconstruc-tion (Figure 1), which is almost identical to the topology that can be seen inphylogenies reconstructed using completely different characters (e.g., Fabreet al., 2012). Our few long routines do structure the phylogeny and, thus,cannot be the result of random processes.
14 Behaviour (2013) DOI:10.1163/1568539X-00003096
J. Malange et al. / Behaviour (2013) 15
Figure 5. Behavioural morphospace: number of grooming routines as a function of the com-plexity of the routine (number of behavioural units in the routine). The black line showsthe number of possible routines, or the total morphospace. The grey lines show the num-ber of actual routines performed by the animals. Dark grey represents the total number ofroutines actually performed (including informative and non-informative ones); medium darkgrey represents the phylogenetically informative routines; and light grey shows the numberof non-homoplastic routines.
4. Discussion
4.1. Phylogeny of rodents
The behavioural and combined data phylogeny show rodents as a mono-phyletic group (Figures 1 and 3). The monophyly of rodents is also supported
Figure 4. Evolution of the complexity on the behavioural system. Simple, short routines(black) are more informative at basal levels of the phylogeny; large, or complex routines(white) are more informative at the upper levels of the phylogeny; routines of intermediatesize (grey) show an intermediate pattern. The circles drawn at the branches of the hypotheticphylogeny illustrate the level where the characters stuck as synapomorphies. This pattern wasobserved in the behavioural (A), the combined data (B) phylogenies, and (C) in the topologyof a recent rodent phylogeny (Fabre et al., 2012).

16 Behaviour (2013) DOI:10.1163/1568539X-00003096
by morphological (Carleton & Musser, 2005) and many other molecular phy-logenies (Nedbal et al., 1996; DeBry & Sagel, 2001; Reyes et al., 2004;Blanga-Kanfi et al., 2009), and is also recovered in the recent comprehen-sive meta-analysis of Fabre et al. (2012).
Within rodents, our behavioural data support the monophyly of Sciurog-nathi, a result that is mostly uncontroversial, since many authors agree on asister group relationship for the families Cricetidae and Muridae (McKenna& Bell, 1997; De Brye et al., 2001; Carleton & Musser, 2005), both includedin the present study. Also, many authors agree that the families Echimyi-dae and Caviidae are embedded within Histricognathi (Carleton & Musser,2005), a result that was recovered only on our behavioural tree (Figure 1).The only topological difference between behavioural and recent reconstruc-tions (Fabre et al., 2012) is the position of T. yonenagae, a poorly known,and recently described rodent species. So it seems that our behavioural char-acters (routines), recovered by the graph algorithm (Japyassú et al., 2006),do indeed provide accurate phylogenetic information.
The two populations of T. laurentius are possibly two different species.There is clearly need for more data in order to confirm this result, but weare confident of it because we have observed further morphological and be-havioural differences between them. For example, specimens from the Piauípopulation are smaller, more silent, and less aggressive towards humans, thanare the specimens from Bahia. Also, these populations differ in food and wa-ter intake, urine concentration (Mendes et al., 2004) and anatomy (Reis etal., 2002; Basile, 2003).
4.2. The evolution of grooming
Units of grooming behaviour are uninformative at the level of the order Ro-dentia, implying that they are possibly older than the whole group. Manystudies show that grooming units are present in many different rodent species(Steiner, 1973; Borchelt et al., 1976; Lacher Jr., 1981; Randall, 1981; Geyer& Kornet, 1982; Chou et al., 1985; Messias, 1995; Shanas & Terkel, 1996;Bursten et al., 2000; Manaf & Spinelli Oliveira, 2000; Sabatini & Costa,2001; Stopka & Graciasová, 2001), and it seems that these units have beenused for more than 50 million years, before the separation between the altri-cial and precocial rodents (Colonnese et al., 1996). Grooming units can beshared even among tetrapods (Sachs, 1988), a possibility that extends theirorigin to an even deeper evolutionary point.

J. Malange et al. / Behaviour (2013) 17
Present in such a wide range of taxa, grooming probably exerts importantadaptive functions. Within mammals it is related to obvious functions suchas ectoparasite or dirt removal (Geyer & Kornet, 1982; Hart, 1990; Mooring& Hart, 1997; Hawlena et al., 2007) and bacterial control by saliva (Kohari etal., 2009), but also to presumably derived functions such as thermoregulation(Shanas & Terkel, 1996), and even communication (Ferkin et al., 1996, 2001;Wolff et al., 2002) or ritualised territory defence (Bursten et al., 2000).
Among these functions, we suggest that the reason behind the evolution-ary merging of head and trunk grooming modules in rodents is an increasein the efficiency of ectoparasite removal. It is well known that ectoparasiteshave a stronger effect on smaller mammals, a condition known as the ‘bodysize principle’ (Hart et al., 1992; Mooring et al., 2000). The reason behindthis principle is that smaller mammals are more affected by ectoparasites dueto their higher surface/volume relationship, a condition that should selectfor more effective ectoparasite removal. Enhanced parasite infestations, dueto habitat selection or social vigilance stress, for example, select for highergrooming rates in ungulates (Mooring et al., 2004).
Enhanced parasite load, due to the reduced rodent size, could also se-lect for more efficient grooming patterns. The all-body bouts of groomingbehaviour performed by rodents (starting on the head and ending in thetrunk) could indeed be more effective by reducing the chances of crossedinfestations between the head and the trunk. The idea of cross-infestation re-lates to the possibility that, right after a head cleaning bout, trunk parasitescould, for example, re-infect the head. This would be especially common insmaller mammals, because ectoparasites like ticks could more rapidly findtheir way to uninfected, recently cleaned areas. In this situation, there wouldbe selective pressure for whole body (head plus trunk), complete cleaningbouts. Thus, our point is that, besides the evolutionary enhancement of thefrequency of grooming in smaller mammals (Hart et al., 1992; Mooring etal., 2004), there would also be selective pressure for a more efficient patternof grooming in these smaller animals, and the head-trunk, fused groomingpattern, would be more efficient in preventing possible crossed-infestationsbetween distinct body parts.
In line with this reasoning, we see that within rodents, the smaller ones,like the sciurognathids, perform all-body grooming sequences in a presum-ably more efficient way: they perform very quick all-body grooming se-quences, and they make use of an additional overall direction of grooming

18 Behaviour (2013) DOI:10.1163/1568539X-00003096
activities (they make use of a dorsoventral flow of behaviours in addition tothe usual head-tail flow); these results are also accounted for by the body sizehypothesis, and accordingly we suggest that body size is an important factoralso in the evolution of the pattern of grooming in rodents.
4.3. Evolution of complexity in grooming behaviour
Our results point to an increase of behavioural complexity through time.As a general trend, rodent behaviour evolves from short to long groomingroutines, adding each time more connectivity to the emerging behaviouralsystem. This is a very robust result, since the pattern remains when weoptimise the routines in three distinct tree topologies (Figure 4), includinga recent comprehensive molecular meta-analysis (Fabre et al., 2012).
This result has implications for the delimitation of behavioural characters:short sequences should be preferred if one is looking for deep phylogeneticstructure, that is, if one is interested in the phylogeny of very ancient taxa.The other way around, long behavioural routines should be preferred if one iswilling to uncover historical relationships between recent taxa. This is analo-gous to what happens with molecular data, where some sequences are used toreveal the topology of ancient taxa, while others reveal relationships betweenpopulations of a single species (Michot et al., 1990; Nee et al., 1998). Thisresult points to the possibility of using hyper-variable behavioural sequencesfor inferring rapid evolutionary events among very recent taxa.
The exponentially growing function of the number of possible behaviouralroutines contrasts markedly with the bell shaped function of the number ofactual behavioural routines (Figure 5). The exponentially growing functionrefers to the total space of possibilities, that is, the complete behaviouralmorphospace. The bell shaped function represents the routines that are bi-ologically structured, the ones that have been shaped by evolutionary pro-cesses that acted upon rodent grooming behaviour. The strong differencebetween these curves means that evolutionary processes have selected fora very small portion of the complete behavioural morphospace. Selection orbehavioural constraints are particularly harsh for long behavioural routines,since the number of actual routines declines pronouncedly for sequenceswith more than four behavioural units, implying that there is a restrictionfor the increase of complexity in this group. So it seems that, besides thegeneral trend for the progressive increase in behavioural complexity, thereare upper biological limits to complexity.

J. Malange et al. / Behaviour (2013) 19
4.4. Upper bounds to complexity
Although there are numerous papers detecting significant evolutionary trendsin the number of behavioural units (Ord & Blumstein, 2002; McComb &Semple, 2005; Japyassú & Caires, 2008; delBarco-Trillo et al., 2012; Free-berg et al., 2012; Gustison et al., 2012; Ord & Garcia-Porta, 2012; Pollard &Blumstein, 2012), only a few of them look for the evolution of syntax, tryingto detect patterns in the organisation of the behavioural units. This is usuallythe case for communication sequences repertoire size, for example, in birds(Ord & Garcia-Porta, 2012), where some groups have even been shown tohave open ended song repertoires (Hailman et al., 1985; Freeberg & Lucas,2012). Traditionally, song and repertoire size in songbirds have been shownto be sexually selected markers of male quality, based on the notions thatlearning more song elements is cognitively difficult (see review in Catchpole& Slater, 2008), a task that gets additionally expensive when consistency inthe repetition of songs is relevant (Sakata & Vehrencamp, 2012). Thus, theevidence we have so far about syntax is that the evolution of open endedrepertoires is rare, and that the use of long song streams is energeticallycostly, not only due to motor performance per se, but more generally be-cause it requires more neural tissue to be maintained even in quiet moments(Brenowitz, 2004; Van der Linden et al., 2009). Longer routines, that areusually connected to larger behavioural repertoires (Hailman et al., 1985),thus, have additional costs that could help explain the clear absence of verylong grooming routines (Figure 5).
Besides the physiological costs, another possible selective pressureagainst very long routines could be related to a trade-off with vigilance.There is evidence that grooming frequencies are smaller among territoryholding male cervids, than among females or bachelor males (Hart et al.,1992). This is because territory holding males allocate more time to vigilance(either against possible contesting males, or in the search for reproductivefemales) and, thus, reduce grooming frequency and efficiency (Mooring etal., 2004). This suggests an indirect cost for enhancing grooming rates: thenegative payoffs of the associated decreased vigilance rates. Decreased vigi-lance is an important selective factor that affects time allocation not only forgrooming, but for a range of other time consuming activities and, thus, canpotentially have a more general impact on fitness (Dukas & Clark, 1995).Thus, the trade-off between vigilance and grooming could select not only

20 Behaviour (2013) DOI:10.1163/1568539X-00003096
for reduced grooming rates, but also against very long grooming bouts, thuspositing a higher cost for these longer routines.
4.5. Concluding remarks
The organisation of behavioural units into modules of coordinated patternsfirst emerged around large body parts, such as the head and the trunk, mod-ules that were later fused. The evolution of higher complexity within thissystem is possibly connected to the selective pressures promoting size reduc-tion within rodents, with indirect effects on the increase of selective pressuresrelated to parasite load. The increase in the complexity of grooming seemssubjected to upper limits, which may be due to physiological and/or vigi-lance constraints.
The modular organisation, and the evolution of the connectivity within be-havioural systems, show that we can move from the use of reflex-like unitsof behaviour as taxonomic indicators, a very successful tradition reminiscentfrom classical ethology (McLennan, 1993; Paterson et al., 1995; Alberts,1996; Slikas, 1998; Stuart & Hunter, 1998; Bucheli et al., 2002; Noll, 2002;Quadros, 2002, 2008; Desutter-Grandcolas & Robillard, 2003; Robillard etal., 2006; Cap et al., 2008), to an evolutionary analysis of the complex be-havioural systems, such as foraging or mating systems, a tradition more akinto behavioural ecology studies. This approximation between the researchagendas of classical ethology and behavioural ecology will require an ex-pansion of the focus in this last discipline: from the modelling of alternativeand evolutionarily stable behavioural strategies in a few, to the comparativestudy of large groups of species, within an explicitly phylogenetic frame-work.
Acknowledgements
The authors wish to thank Fernando Noll, César Ades (in memoriam) andtwo anonymous referees for helpful comments on previous versions of themanuscript. We also thank Dr. Paulo Sérgio D’Andrea, Dr. Paulo Manaf,Dr. Nina Furnari, Dr. Patrícia Monticelli, MsC. Evelyn O. Sarmento andMsC. José A. de Raeffray Barbosa for providing access to some of theiroriginal videos, and for providing the fundamental infrastructure to makenew ones. J.M. and H.F.J. received CNPq grants during the elaboration of thepresent research. This project is part of the INOMEP-PRONEX initiative, amultidisciplinary research program with funding from FAPESB and CNPq.

J. Malange et al. / Behaviour (2013) 21
References
Adkins, R.M., Gelke, E.L., Rowe, D. & Honeycutt, R.L. (2001). Molecular phylogeny andtime estimates for major rodent groups: evidence from multiple genes. — Mol. Biol. Evol.18: 777-791.
Adkins, R.M., Walton, A.H. & Honeycutt, R.L. (2003). Higher-level systematics of rodentsand divergence time estimates based on two congruent nuclear genes. — Mol. Phylogen.Evol. 26: 409-420.
Alberts, C.C. (1996). O comportamento de autolimpeza do gato doméstico (Felis catus) e umaproposta para usá-lo como caractere filogenético. — Tese de doutoramento apresentadaao Instituto de Psicologia da Universidade de São Paulo, São Paulo, 205 pp.
Arnason, U., Adegoke, J.A., Bodin, K., Born, E.W., Esa, Y.B., Gullberg, A., Nilsson, M.,Short, R.V., Xu, X. & Janke, A. (2002). Mammalian mitogenomic relationships and theroot of the eutherian tree. — Proc. Natl. Acad. Sci. USA 99: 8151-8156.
Atz, J.W. (1970). The application of the idea of homology to behaviour. — In: Developmentand evolution of behaviour: essays in memory of TC Schneirla (Aronson, L.R., Lehrman,D.S. & Rosenblatt, J.S., eds). W.H. Freeman, San Francisco, CA, p. 53-74.
Baroni Urbani, C. (1989). Phylogeny and behavioural evolution in ants, with a discussion ofbehaviour in evolutionary processes. — Ethol. Ecol. Evol. 1: 137-168.
Basile, P.A. (2003). Taxonomia de Thrichomys Trouessart 1880 (Rodentia, Echimyidae).— Dissertação (Mestrado em Biologia Comparada), Faculdade de Filosofia, Ciências eLetras de Ribeirão Preto, USP, Ribeirão Preto.
Berridge, K.C. (1990). Comparative fine structure of action: rules of form and sequence inthe grooming patterns of six rodent species. — Behaviour 113: 21-56.
Berridge, K.C., Aldridge, J.W., Houchard, K.R. & Zhuang, X. (2005). Sequential super-stereotypy of an instinctive fixed action pattern in hyper-dopaminergic mutant mice:a model of obsessive compulsive disorder and Tourette’s. — BMC Biol. 3: 1-16.
Blanga-Kanfi, S., Miranda, H., Penn, O., Pupko, T., DeBry, R.W. & Huchon, D. (2009).Rodent phylogeny revised: analysis of six nuclear genes from all major rodents clade. —BMC Evol. Biol. 9: 71.
Borchelt, P.L., Griswold, J.G. & Branchek, R.S. (1976). An analysis of sand bathing andgrooming in the kangaroo rat (Dipodomys merriami). — Anim. Behav. 24: 347-353.
Brenowitz, E.A. (2004). Plasticity of the adult avian song control system. — Ann. NY Acad.Sci. 1016: 560-585.
Bucheli, S., Landry, J.F. & Wenzel, J. (2002). Larval case architecture and implications ofhost-plant associations for North American Coleophora (Lepidoptera: Coleophoridae). —Cladistics 18: 71-93.
Bursten, S.N., Owings, D.H. & Berridge, K.C. (2000). Do California ground squirrels (Sper-mophilus beecheyi) use ritualized syntactic cephalocaudal grooming as an agonistic sig-nal? — J. Comp. Psychol. 114: 281-290.
Cap, H., Deleporte, P., Joachim, J. & Reby, D. (2008). Male vocal behaviour and phylogenyin deer. — Cladistics 24: 917-931.

22 Behaviour (2013) DOI:10.1163/1568539X-00003096
Carleton, M.D. & Musser, G.G. (2005). Order Rodentia. — In: Mammal species of the world:a taxonomic and geographic reference (Wilson, D.E. & Reeder, D.M., eds). Johns HopkinsUniversity Press, Baltimore, MD, p. 745-752.
Catchpole, C.K. & Slater, P.J.B. (2008). Bird song: biological themes and variations. —Cambridge University Press. Cambridge.
Chou, L.S., Lin, Y.S. & Mok, H.K. (1985). Study of the maintenance behaviour of the red-bellied tree squirrel, Callosciurus erythraeus. — Bull. Inst. Zool. Acad. Sin. 24: 39-50.
Colonnese, M.T., Stallman, E.L. & Berridge, K.C. (1996). Ontogeny of action syntax in altri-cial and precocial rodents: grooming sequence of rat and guinea pig pups. — Behaviour133: 1165-1195.
Cromwell, H.C. & Berridge, K.C. (1996). Implementation of action sequences by a neostri-atal site: a lesion mapping study of grooming syntax. — J. Neurosci. 16: 3444-3458.
Da Silva Neto, E.J. (2000). Morphology of the regiones ethmoidalis and orbitotemporalis inGalea musteloides Meyen 1832 and Kerodon rupestris (Wied-Neuwied 1820) (Rodentia:Caviidae) with comments on the phylogenetic systematics of the Caviidae. — J. Zool.Syst. Evol. Res. 38: 219-229.
Dawkins, R. & Dawkins, M. (1976). Hierarchical organisation and postural facilitation: rulesfor grooming in flies. — Anim. Behav. 24: 739-755.
DeBry, R.W. & Sagel, R.M. (2001). Phylogeny of Rodentia (Mammalia) inferred from thenuclear-encoded gene IRBP. — Mol. Phylogenet. Evol. 19: 290-301.
D’Erchia, A.M., Gissi, C., Pesole, G., Saccone, C. & Arnason, U. (1996). The guinea pig isnot a rodent. — Nature 381: 597-600.
Desutter-Grandcolas, L. & Robillard, T. (2003). Phylogeny and the evolution of calling songsin Gryllus (Insecta, Orthoptera, Gryllidae). — Zool. Scripta 32: 173-183.
Dukas, R. & Clark, C. (1995). Sustained vigilance and animal performance. — Anim. Behav.49: 1259-1267.
Eisenberg, J.F. (1963). The behaviour of heteromyid rodents. — Univ. Calif. Publ. Zool. 69:1-100.
Fabre, P.-H., Hautier, L., Dimitrov, D. & Douzery, E.J.P. (2012). A glimpse on the pat-tern of rodent diversification: a phylogenetic approach. — BMC Evol. Biol. 12: 88,DOI:10.1186/1471-2148-12-88.
Farish, D.J. (1972). The evolutionary implications of qualitative variation in the groomingbehaviour of the Hymenoptera (Insecta). — Anim. Behav. 20: 662-676.
Farris, J.S., Albert, V.A., Källersjö, M., Lipscomb, D. & Kluge, A.G. (1996). Parsimonyjackknifing outperforms neighbor-joining. — Cladistics 12: 99-124.
Felsenstein, J. (1985). Confidence limits on phylogenies: an approach using the bootstrap. —Evolution 39: 783-791.
Fentress, J.C. (1972). Development and patterning of movement sequences in inbred mice.— In: The biology of behaviour (Kiger, J.A., ed.). Oregon State University Press, Eugene,OR, p. 83-132.
Fentress, J.C. (1973). Development of grooming in mice with amputed forelimbs. — Science179: 704-705.

J. Malange et al. / Behaviour (2013) 23
Fentress, J.C. (1987). Ethological invariants: boxes, rubber bands, and biological processes.— Behav. Brain Sci. 10: 377-378.
Ferkin, M.H., Sorokin, E.S. & Johnston, R.E. (1996). Self-grooming as a sexually dimorphiccommunicative behaviour in meadow voles, Microtus pennsylvanicus. — Anim. Behav.51: 801-810.
Ferkin, M.H., Heath, L.A. & Paz-Y-Miño, G. (2001). Self-grooming as a tactic used by prairievoles Microtus ochrogaster to enhance sexual communication. — Ethology 107: 939-949.
Freeberg, T.M., Dunbar, R.I.M. & Ord, T.J. (2012). Social complexity as a proximate andultimate factor in communicative complexity. — Phil. Trans. Roy. Soc. B 367: 1785-1801.
Freeberg, T.M. & Lucas, J.R. (2012). Information theoretical approaches to chick-a-dee callsof Carolina chickadees (Poecile carolinensis). — J. Comp. Psychol. 126: 68-81.
Geyer, L.A. & Kornet, C.A. (1982). Auto and allo-grooming in pine voles (Microtus pineto-rum) and meadow voles (Microtus pennsylvanicus). — Physiol. Behav. 28: 409-412.
Goloboff, P.A., Farris, J.S. & Nixon, K.C. (2008). TNT, a free program for phylogeneticanalysis. — Cladistics 24: 774-786.
Graur, D., Hide, W.A. & Li, W.H. (1991). Is the guinea pig a rodent? — Nature 351: 649-652.Greer, J.M. & Capecchi, M.R. (2002). Hoxb8 is required for normal grooming behaviour in
mice. — Neuron 33: 23-24.Gustison, M.L., le Roux, A. & Bergman, T.J. (2012). Derived vocalisations of geladas (Thero-
pithecus gelada) and the evolution of vocal complexity in primates. — Phil. Trans. Roy.Soc. B 367: 1847-1859.
Hailman, J.P., Ficken, M.S. & Ficken, R. (1985). The “chick-a-dee” calls of Parus atricapil-lus: a recombinant system of animal communication compared with written English. —Semiotica 56: 191-224.
Hart, B.L., Hart, L.A., Mooring, M.S. & Olubayo, R. (1992). Biological basis of groomingbehaviour in antelope: the body size, vigilance and habitat principles. — Anim. Behav.44: 615-631.
Hawlena, H., Bashary, D., Abramsky, Z. & Krasnov, B.R. (2007). Benefits, costs and con-straints of anti-parasitic grooming in adult and juvenile rodents. — Ethology 113: 394-402.
Huchon, D. & Douzery, E.J.P. (2001). From the Old World to the New World: a molecularchronicle of the phylogeny and biogeography of Hystricognathi rodents. — Mol. Phylo-genet. Evol. 20: 238-251.
Japyassú, H.F. & Caires, R.A. (2008). Hunting tactics in a cobweb spider (Araneae-Theridiidae) and the evolution of behavioural plasticity. — J. Insect Behav. 21: 258-284.
Japyassú, H.F. & Machado, F.A. (2010). Coding behavioural data for cladistic analysis: usingdynamic homology without parsimony. — Cladistics 26: 625-642.
Japyassú, H.F. & Viera, C. (2002). Predatory plasticity in Nephilengys cruentata (Araneae:Tetragnathidae): relevance for phylogeny reconstruction. — Behaviour 139: 529-544.
Japyassú, H.F., Alberts, C.C., Izar, P. & Sato, T. (2006). EthoSeq: a tool for phylogeneticanalysis and data mining on behavioural sequences. — Behav. Res. Methods 38: 549-556.

24 Behaviour (2013) DOI:10.1163/1568539X-00003096
Kalueff, A.V. & Tuohimaa, P. (2004). Grooming analysis algorithm for neurobehaviouralstress research. — Brain Res. Protocols 13: 151-158.
Lacher Jr., T.E. (1981). The comparative social behaviour of Kerodon rupestris and Galeaspixii and the evolution of behaviour in the Caviidae. — Bull. Carnegie Mus. Nat. Hist.17: 5-71.
Legendre, F., Robillard, T., Desutter-Grandcolas, L., Whiting, M.F. & Grandcolas, P. (2008).Phylogenetic analysis of non-stereotyped behavioural sequences with a successive event-pairing method. — Biol. J. Linn. Soc. 94: 853-867.
Lehner, P.N. (1996). Handbook of ethological methods. — Cambridge University Press,Cambridge.
Li, W.H., Hide, W.A., Zharkikh, A., Ma, D.P. & Graur, D. (1992). The molecular taxonomyand evolution of the guinea pig. — J. Hered. 83: 174-181.
Lin, Y., Waddell, P. & Penny, D. (2002). Pika and vole mitochondrial genomes increasesupport for both rodent monophyly and glires. — Gene 294: 119-129.
Lorenz, K. (1941). Vergleichende Bewegungsstudien an Anatinen. — J. Ornithol. 89: 194-294.
Manaf, P. & Spinelli Oliveira, E. (2000). Behavioural repertoire of the spiny rat Proechimys[Trinomys] yonenagae (Rodentia: Echimyidae) in captivity. — Rev. Etol. 2: 3-15.
Martignetti, J.A. & Brosius, J. (1993). Neural BC1 RNA as an evolutionary marker; guineapigs remains a rodent. — Proc. Natl. Acad. Sci. USA 90: 9698-9702.
McComb, K. & Semple, S. (2005). Coevolution of vocal communication and sociality inprimates. — Biol. Lett. 1: 381-385.
McKenna, M.C. & Bell, S.K. (1997). Classification of mammals above the species level. —Columbia University Press, New York, NY.
McLennan, D.A. (1993). Phylogenetic relationships in the Gasterosteidae: an updated treebased on behavioural characters with a discussion of homoplasy. — Copeia: 318-326.
Mendes, L.A.F., Rocha, P.L.B., Ribeiro, M.F.S., Perry, S.F. & Spinelli Oliveira, E. (2004).Differences in ingestive balance of two populations of neotropical Thrichomys apereoides(Rodentia, Echimyidae). — Comp. Biochem. Physiol. 138: 327-332.
Messias, M.R. (1995). Biologia comportamental de Cavia aperea (Mammalia: Caviidae) emárea aberta no Estado de São Paulo. — Dissertação de Mestrado. Instituto de Biociências,Universidade Estadual Paulista, Rio Claro.
Michot, B., Qu, L.H. & Bachellerie, J.P. (1990). Evolution of large-subunit rRNA structure:the diversification of divergent D3 domain among major phylogenetic groups. — Eur. J.Biochem. 188: 219-229.
Mooring, M.S. & Hart, B.L. (1997). Self-grooming in impala mothers and lambs: testing thebody size and tick challenge principles. — Anim. Behav. 53: 925-934.
Mooring, M.S., Benjamin, J.E., Harte, C.R. & Herzog, N.B. (2000). Testing the interspecificbody size principle in ungulates: the smaller they come, the harder they groom. — Anim.Behav. 60: 35-45.
Mooring, M.S., Blumstein, D.T. & Stoner, C.J. (2004). The evolution of parasite-defencegrooming in ungulates. — Biol. J. Linn. Soc. 81: 17-37.

J. Malange et al. / Behaviour (2013) 25
Narita, Y., Oda, S. & Kageyama, T. (2006). Rodent monophyly deduced from the uniquegastric proteinase constitution and molecular phylogenetic analyses using pepsinogen-CcDNA sequences. — Comp. Biochem. Physiol. D 1: 273-282.
Nedbal, M.A., Honeycutt, R.L. & Schlitter, D.A. (1996). Higher-level systematics of rodents(Mammalia, Rodentia): evidence from the mitochondrial 12S rRNA gene. — J. Mammal.Evol. 3: 201-237.
Nee, S., Holmes, E.C., Rambaut, A. & Harvey, P.H. (1998). Inferring population history frommolecular phylogenies. — In: New uses for new phylogenies (Harvey, P.H., Leigh Borwn,J.J., Maynard Smith, J. & Nee, S., eds). Oxford University Press, Oxford, p. 66-80.
Noll, F.B. (2002). Behavioural phylogeny of Corbiculate Apidae (Hymenoptera; Apinae),with special reference to social behaviour. — Cladistics 18: 137-153.
Ord, T.J. & Blumstein, D.T. (2002). Size constraints and the evolution of display complexity:why do large lizards have simple displays? — Biol. J. Linn. Soc. 76: 145-161.
Ord, T.J. & Garcia-Porta, J. (2012). Is sociality required for the evolution of communica-tive complexity? Evidence weighed against alternative hypotheses in diverse taxonomicgroups. — Phil. Trans. Roy. Soc. B 367: 1811-1828.
Ottoni, E.B. (2000). EthoLog 2.2: a tool for the transcription and timing of behaviour obser-vation sessions. — Behav. Res. Methods Instr. Comput. 32: 446-449.
Paterson, A.M., Wallis, G.P. & Gray, R.D. (1995). Penguins, petrels and parsimony: doescladistic analysis of behaviour reflect seabird phylogeny? — Evolution 49: 974-989.
Pollard, K.A. & Blumstein, D.T. (2012). Evolving communicative complexity: insights fromrodents and beyond. — Phil. Trans. Roy. Soc. B 367: 1869-1878.
Poux, C., Chevret, P., Huchon, D., De Jong, W.W. & Douzery, E.J.P. (2006). Arrival anddiversification of caviomorph rodents and platyrrhine primates in South America. — Syst.Biol. 55: 228-244.
Price, J.J., Clapp, M.K. & Omland, K.E. (2011). Where have all trees gone? The declininguse of phylogenies in animal behaviour journals. — Anim. Behav. 81: 667-670.
Quintana, C.C. (1998). Relaciones filogenéticas de roedores Caviinae (Caviomorpha, Cavi-idae), de América del Sur. — Boletín de la Real Sociedad Española de Historia Natural(Sección Biológica) 94: 125-134.
Quadros, A.H. (2002). Filogenia da Família Cathartidae (Aves) baseada em comportamentode autolimpeza. — Dissertação (Mestrado em Psicologia Experimental), Instituto de Psi-cologia, Universidade de São Paulo, São Paulo, 90 pp.
Quadros, A.H. (2008). Filogenia de falconiformes (aves) baseada em comportamento deautolimpeza. — Tese (Doutorado em Psicologia Experimental), Instituto de Psicologia,Universidade de São Paulo, São Paulo, 158 pp.
Randall, J.A. (1981). Comparison of sand bathing and grooming in two species of kangaroorat. — Anim. Behav. 29: 1213-1219.
Reis, S.F., Duarte, L.C., Monteiro, L.R. & Zuben, F.J.V. (2002). Geographic variation incranial morphology in Thrichomys apereoides: I. Geometric descriptors and patterns ofvariation in shape. — J. Mammal. 83: 333-344.

26 Behaviour (2013) DOI:10.1163/1568539X-00003096
Reyes, A., Gissi, C., Pesole, G., Catzeflis, F.M. & Saccone, C. (2000). Where do rodentsfit? Evidence from the complete mitochondrial genome of Sciurus vulgaris. — Mol. Biol.Evol. 17: 979-983.
Reyes, A., Gissi, C., Catzeflis, F.M., Nevo, E., Pesole, G. & Saccone, C. (2004). Congruentmammalian trees from mitochondrial and nuclear genes using Bayesian methods. — Mol.Biol. Evol. 21: 397-403.
Robillard, T., Legendre, F., Desutter-Grandcolas, L. & Grandcolas, P. (2006). Phylogeneticanalysis and alignment of behavioural sequences by direct optimization. — Cladistics 22:602-633.
Rocha, P.L.B. (1995). Proechimys yonenagae, a new species of spiny rat (Rodentia: Echimyi-dae) from fossil sand dunes in the Brazilian Caatinga. — Mammalia 59: 537-549.
Roth, L.L. & Rosenblatt, J.S. (1966). Mammary glands of pregnant rats: development stimu-lated by licking. — Science 151: 1403-1404.
Roth, L.L. & Rosenblatt, J.S. (1967). Changes in self-licking during pregnancy in the rat. —J. Comp. Physiol. Psychol. 63: 397-400.
Rowe, D.L. & Honeycutt, R.L. (2002). Phylogenetic relationships, ecological correlates, andmolecular evolution within the Cavioidea (Mammalia, Rodentia). — Mol. Biol. Evol. 19:263-277.
Sabatini, V. & Costa, M.J.R.P. (2001). Etograma da paca (Agouti paca, Linnaeus 1766) emcativeiro. — Rev. Etol. 3: 3-14.
Sachs, B.D. (1988). The development of grooming and its expression in adult animals. —Ann. NY Acad. Sci. 525: 1-17.
Sakata, J.T. & Vehrencamp, S.L. (2012). Integrating perspectives on vocal performance andconsistency. — J. Exp. Biol. 215: 201-209.
Shanas, U. & Terkel, J. (1996). Grooming secretions and seasonal adaptations in the blindmole rat (Spalax ehrenbergi). — Physiol. Behav. 60: 653-656.
Silverman, P. (1978). Animal behaviour in the laboratory. — Chapman and Hall, London.
Slikas, B. (1998). Recognizing and testing homology of courtship displays in storks (Aves:Ciconiiformes: Ciconiidae). — Evolution 52: 882-893.
Spearman, C. (1904). The proof and measurement of association between two things. — Am.J. Psychol. 15: 72-101.
Steiner, A.L. (1973). Self- and allo-grooming behaviour in some ground squirrels (Sciuridae),a descriptive study. — Can. J. Zool. 51: 151-161.
Stopka, P. & Graciasová, R. (2001). Conditional allogrooming in the herb-field mouse. —Behav. Ecol. 12: 584-589.
Stuart, A.E. & Hunter, F.F. (1998). End-products of behaviour versus behavioural charac-ters: a phylogenetic investigation of pupal cocoon construction and form in some NorthAmerican black flies (Diptera: Simuliidae). — Syst. Entomol. 23: 387-398.
Van der Linden, A., Van Meir, V., Boumans, T., Poirier, C. & Balthazart, J. (2009). MRI insmall brains displaying extensive plasticity. — Trends Neurosci. 32: 257-266.
West-Eberhard, M.J. (2003). Developmental plasticity and evolution. — Oxford UniversityPress, New York, NY.

J. Malange et al. / Behaviour (2013) 27
Wheeler, W.C. (2003). Implied alignment: a synapomorphy-based multiple-sequence align-ment method and its use in cladogram search. — Cladistics 19: 261-268.
Wolff, J.O., Watson, M.H. & Thomas, S.A. (2002). Is self-grooming by prairie voles a pre-dictor of mate choice? — Ethology 108: 169-179.

28 Behaviour (2013) DOI:10.1163/1568539X-00003096
Appendix A.The ethogram with 27 behavioural categories was accomplished through the observation ofgrooming behaviour in the 12 taxa studied, and also from descriptions available in the liter-ature. Silverman (1978) defined the major units of grooming behaviour described for Rattusnorvegicus (Wistar), supported by the descriptions of Grant, Mackintosh and Draper (apudSilverman, 1978). According to them, washing is the alternation among licking forelimbs andrubbing head with them; combing is lick and comb (bite) flank hair, hindfoot, belly and tailwith teeth; scratching is defined as the action of combing flank and back hairs with hindfeet;lick urogenital area is followed by covering the female and, often, by ejaculation.
Lick forelimb(lick_forelimb)
On its hind feet, the animal raises its forelimb(s) to the mouth (togetheror one limb at a time) and lick them, or the animal turns down its headand conduces its mouth and tongue to the forelimb(s). Lick is the sameto “wash” according to Silverman’s (1978) terminology.
Lick hind foot(lick_hindf)
The animal raises its hind foot (feet) to the mouth or it can turn downto the foot (feet) region and lick them, always one foot at a time.
Lick belly(lick_belly)
The animal displays a dorsoventral curvature and lick belly region atthe same time that it moves its head.
Lick back(lick_back)
The animal turns over its body and head in order to reach its body backand lick this region.
Lick flank(lick_flank)
The animal turns over the body to the flank region and licks it.
Lick tail(lick_tail)
The animal holds its tail by forelimbs or it can turn down to tail andlick it. This movement is often performed in a distal-proximaldirection.
Bitingforelimb(bit_forelimb)
On its hind feet, the animal raises its forelimbs to the mouth (togetheror one limb at a time) and bite them, or the animal turns down its headand conduces its mouth and tongue to forelimb(s). This behaviouralcategory is almost interpolated to “lick forelimb” and correspond to the“comb” according to Silverman’s (1978) terminology.
Biting hind foot(bit_hindf)
Raising hind foot (feet) to the mouth or turn down to foot (feet), theanimal bites foot, always one at a time.
Bitting belly(bit_belly)
On its hind feet, the animal displays a dorsoventral curvature in orderto bite its ventral region and comb ventral hair in the direction the hairgrows.
Biting back(bit_back)
On its hind feet, the animal turns over the body and head in order toreach its back. The animal bites its dorsal region and combs the hair inthe direction the hair grows.
Biting flank(bit_flank)
The animal turns over its body and head in order to reach flank regionand bites and combs it in the direction the hair grows.
Biting tail(bit_tail)
The animal holds its tail with the forelimbs or it can turn down to thetail and bite it. This movement is often performed in a distal toproximal direction.
Scratch head(stch_head)
The animal scratches its head, including the ears, the snout and the topof the head, always by one foot at a time. The animal can displays ahead curvature to hind feet.

J. Malange et al. / Behaviour (2013) 29
Appendix A.(Continued.)
Scratch belly(stch_belly)
The animal scratches its belly always by one foot at a time.
Scratch back(stch_back)
The animal scratches its back always by one foot at a time.
Scratch flank(stch_flank)
The animal scratches its flank always by one foot at a time.
Wash anogenitalregion(wash_anoge)
The animal displays a dorsoventral curvature and turns over its neck tothe genital region and licks it, following a down to up direction.Coprophagy (faeces intake) can be observed in this context.
Shake (shake) The animal shakes the head or the wide body one to another side (rightand left) in a rapid way.
Rubbing eye(rub_eye)
On its hind feet, the animal bends the forelimb(s), touches the eye areaand rubs the eyes with ventral side of its limb(s). The movements areperformed in a distal proximal direction by one or both forelimbs at atime.
Rubbing ear(rub_ear)
On its hind feet, the animal bends the forelimb(s), touches the ear areaand rubs the ears with ventral side of its limbs while its head exhibit adorsoventral motion. This behavioural category is performed in a distalproximal direction.
Rubbingvibrissae(rub_vibss)
On its hind feet, the animal bends the forelimb(s), touch face area andrubs vibrissae and nose with ventral side of its limbs in circularmovements.
Drinking(drinking)
The animal puts the tongue out and licks the drinking place.Movements on water surface can help to observe this behaviour.
Eating (eating) The animal reaches the feedbox or takes a food by its forelimbs on thecage and eats them. It is possible to observe chewing performance(chewing behaviour). Coprophagy (faeces intake) wasn’t included inthis behavioural category.
Environmentalexploration(exploration)
The animal can be foraging in the cage or just explore the place whenit is making steady movements with the nose and vibrissae, walking oris stopped on its hind feet in the ground. The animal can displayrearing, and sometimes scratches the ground too.
Walking(walking)
The animal gets around the cage without exploring or foraging ordisplaying rearing.
Stopping andfreezing(stopped)
There’s no observable movement by the animal who can be sleeping ordisplaying freezing behaviour. This behavioural category wasconsidered when the animal stays fixed for more than 30 s (random).Freezing can occur in the middle of a complete grooming sequence,while sleeping can only end such a sequence.
Pause (pause) Pause was considered when the animal stays in inactivity by a periodshorter than 30 s. This behavioural category was included to admit thecounting of the same behavioural categories more than once in asequence.

30 Behaviour (2013) DOI:10.1163/1568539X-00003096
App
endi
xB
.Acc
umul
atio
ncu
rves
indi
catin
gth
atal
lter
min
als
reac
hed
apl
atea
u,in
dica
ting
asu
ffici
enta
ndco
mpa
rabl
esa
mpl
ing
effo
rt.E
ach
grap
hic
belo
wre
pres
ents
the
cum
ulat
ive
num
ber
ofbe
havi
oral
units
type
s(f
ora
desc
ript
ion
ofea
chbe
havi
oral
unit
see
App
endi
xA
),as
afu
nctio
nof
the
tota
lnum
ber
ofbe
havi
oral
units
(eve
nts)
obse
rved
and
regi
ster
ed,f
orea
chte
rmin
al(a
ppro
xim
atel
y20
00ev
ents
in76
h).





























