Embed Size (px)
Citation preview

ZOOTAXA
ISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2015 Magnolia Press
Zootaxa 3931 (1): 001–026
www.mapress.com/zootaxa/Article
http://dx.doi.org/10.11646/zootaxa.3931.1.1
http://zoobank.org/urn:lsid:zoobank.org:pub:6E15EB4A-215E-4F99-B7D2-782BDC981CDE
Systematic revision of the ormiine genera Aulacephala Macquart and
Phasioormia Townsend (Diptera, Tachinidae)
SILVIO S. NIHEI
Department of Zoology, Institute of Biosciences, University of São Paulo, Rua do Matão, Travessa 14, n.101, São Paulo, SP, 05508-
090, Brazil. E-mail: [email protected]
Abstract
The tribe Ormiini comprises 64 species in six genera. In the present paper, the ormiine genera Aulacephala Macquart and
Phasioormia Townsend are revised, with two valid species recognized in the former and three valid species recognized in
the latter. All available nominal species in Aulacephala and Phasioormia were examined and are revised herein, so that
previous synonymies could be confirmed. Furthermore, Phasioormia papuana sp. nov. is described from Papua New
Guinea and Indonesia, and Therobia punctigera (Paramonov, 1955) is proposed as a new synonym of Aulacephala hervei
Bequaert, 1922, syn. nov. Keys to species and illustrations of male and female terminalia are provided for both genera.
Key words: new species, new synonymy, Ormiini, systematics, taxonomy
Introduction
The tribe Ormiini includes 64 species and six genera in the world, namely Aulacephala Macquart, 1851 (2 species;
Afrotropical, Oriental and Palaearctic), Homotrixa Villeneuve, 1914 (3 species; Australasian and Oriental),
Mediosetiger Barraclough, 1983 (1 species; Afrotropical), Ormia Robineau-Desvoidy, 1830 (27 species;
Neotropical and Nearctic), Ormiophasia Townsend, 1919 (9 species; Neotropical), Phasioormia Townsend, 1933
(2 species; Oriental), and Therobia Brauer, 1862 (20 species; Afrotropical, Australasian, Oriental and Palaearctic)
(Guimarães 1971, Crosskey 1977, 1980, Cantrell & Crosskey 1989, Herting & Dely-Draskovits 1993, O’Hara &
Wood 2004).
Aulacephala Macquart, 1851 comprises two species: A. maculithorax Macquart, 1851 (type species), which is
widespread in the Afrotropical Region, and A. hervei Bequaert, 1922, distributed in Japan, China and Indonesia.
Other species were described, but subsequently synonymized with A. maculithorax or A. hervei (Crosskey 1976,
1980). Table 1 summarizes all nominal species available in the genus and their current status (valid or not), as well
as the type and non-type specimens studied herein.
Phasioormia Townsend, 1933 includes two species: P. pallida Townsend, 1933 (type species), distributed in
Singapore, Philippines, Sri Lanka and China, and P. bicornis (Malloch, 1932), occurring in India, Malaysia and
China.
In the present paper, the ormiine genera Aulacephala and Phasioormia are revised. A brief generic description
and diagnosis are provided for Aulacephala and Phasioormia, along with identification keys to species.
Descriptions and illustrations of all species recognized in both genera, including the characterization of the male
and female terminalia, are presented. A new species of Phasioormia from Papua New Guinea and Indonesia is
described. For this systematic revision, all nominal species available in Aulacephala and Phasioormia were
examined (Table 1).
Accepted by J. O'Hara: 16 Feb. 2015; published: 11 Mar. 2015 1

TABLE 1. Nominal species of Aulacephala and Phasioormia treated in this revision, with current status and examined
material. (Legends: valid names in bold; repository acronyms: see Material and methods; abbreviations: HT, holotype;
PT, paratype; M, male; F, female.)
Material and methods
The examined material is deposited at the following institutions: Australian National Insect Collection (ANIC);
Biosystematics Laboratory, Kyushu University, Fukuoka, Japan (BLKU); The Natural History Museum, London,
United Kingdom (BMNH); Ehime University, Matsuyama, Japan (EUMJ); Natal Museum, Pietermaritzburg, South
Africa (NMSA); and National Museum of Natural History, Washington DC, USA (USNM). Other repositories
cited in the text: Muséum National d’Histoire Naturelle, Paris, France (MNHN); and Museum für Naturkunde,
Berlin, Germany (ZMHB).
Males and females of Phasioormia pallida and P. papuana sp. nov. were dissected and illustrated.
Phasioormia bicornis was not dissected, because only the holotype male and one female were available for study.
Males and females of Aulacephala maculithorax were dissected and examined, while only the female of A. hervei
was dissected. To my knowledge, only one male of A. hervei, collected from Yokohama, Japan, and deposited in
the BMNH is represented in collections.
Aulacephala and Phasioormia specimens were photographed using a digital camera Leica DFC240 attached to
a stereomicroscope Leica MZ16. Images were captured with the software Leica LAS v4.1 and then stacked and
scaled using the software Helicon Focus v5.3.14.
Photographs were edited using the software Adobe Photoshop CS5, and line drawings were prepared and
edited in Adobe Illustrator CS5. All plates were prepared in Adobe Illustrator CS5.
For the study of male and female terminalia, the terminalia were detached from the abdomen and put into 10%
solution of KOH at room temperature for about 24 hours, followed by neutralization in acetic acid, and then cleared
and washed with distilled water and a series of ethanol solutions of increasing concentrations. The terminalia were
examined in temporary slides with glycerin, and subsequently stored in plastic microvials with glycerin and
attached to the respective specimen. Measurements from head were taken as follows: the frons width and head
width at the level of lunula taken from frontal view, and at the vertex level from dorsal view; head height is the
vertical axis from the lower head margin (genal level) to the upper head margin (vertical level) in lateral view;
fronto-orbital plate width on upper half is the width taken from frontal view at the level of first ocellus, and its
width at the lowermost level was taken from frontal view at level of lunula; parafacial width taken at its mid height
Genus Nominal species Status Material examined
Type-material examined
Aulacephala maculithorax Macquart, 1851 Available, Valid NMSA BMNH USNM
badia Gerstaecker, 1863 (Aulacocephala)
Available, Invalid (=maculithorax)
USNM HT F (ZMHB) (by photos)
hervei Bequaert, 1922 Available, Valid BMNH EUMJ BLKU USNM
HT F (BMNH)
karnyi Malloch, 1925 (Aulacocephala) Available, Invalid (=hervei)
HT F (USNM)
punctigera Paramonov, 1955 (Ormiominda)
Available, Invalid (=hervei) Syn. nov.
HT F (ANIC)
Phasioormia pallida Townsend, 1933 Available, Valid USNM HT F (BMNH) PTs (BMNH)
bicornis Malloch, 1932 (Ormia) Available, Valid BMNH HT M (BMNH)
papuana sp. nov. Sp. nov. HT M (BMNH) PTs (BMNH)
NIHEI 2 · Zootaxa 3931 (1) © 2015 Magnolia Press

from frontolateral view; facial ridge width was taken from frontal view; antennal measurements (pedicel,
postpedicel, arista) were taken from lateral view; antennal axis is the horizontal axis from the posterior margin of
head to the point of antennal insertion in lateral view; oral (epistomal) axis is the horizontal axis from the posterior
margin of head to the anteriormost point of lower facial margin in lateral view; vibrissal axis is the horizontal axis
from the posterior margin of head to the vibrissal insertion in lateral view; and genal height is the vertical measure
from lower head margin to lower eye margin.
The morphological terminology follows Cumming & Wood (2009) and Wood & Zumbado (2010), but for
antennal morphology follows Stuckenberg (1999).
Label data of type specimens are quoted within double quotation marks, and different lines, if present, are
separated by a slash.
Geographical records were gathered based on label data and then georeferenced by searching in Google Maps.
Distributional maps were prepared using software QGIS 2.4.
Results
TRIBE ORMIINI
Aulacephala Macquart
Aulacephala Macquart, 1851: 138 (165). Type species: Aulacephala maculithorax Macquart, by monotypy.Aulacocephala Gerstaecker, 1863: 1033. Unjustified emendation.Aulacocephalopsis Townsend, 1918: 165. Type species: Aulacocephala badia Gerstaecker [=maculithorax Macquart], by
original designation.
Distribution (2 species) (Fig. 39). Afrotropical, Oriental, Palaearctic.
References. Macquart, 1851: 138 (described Aulacephala and the type species A. maculithorax); Gerstaecker,
1863: 1035 (described Aulacocephala badia); Bequaert, 1922: 303 (synonymized badia with maculithorax,
synonymized Aulacocephalopsis with Aulacephala); Bequaert, 1922: 305 (described A. hervei); Malloch, 1925:
147 (described Aulacocephala karnyi); Bequaert, 1929: 164 (synonymized karnyi with hervei); Townsend, 1936:
115 (key to world Aulacephalini); Mesnil, 1973: 1229 (diagnosis and key to species); Crosskey, 1976: 64 (key to
Oriental genera of Ormiini); Crosskey, 1984: 243 (key to Afrotropical genera of Ormiini); Chao & Xue, 1998:
1954 (cat., key to Chinese genera of Ormiini).
Diagnosis. Aulacephala can be distinguished from other ormiine genera by having cell r4+5
closed far before
the wing margin with a very long petiole (about twice as long as r-m) (Figs 11−12); prealar seta absent; ocelli
present (but reduced) and noticeable infuscation on wing (Figs 11−12). Aulacephala closely resembles
Mediosetiger, with which it shares the above listed characters, but differs from the latter by having abdominal
discal setae absent; face oestriform (i.e., forming a long narrow strip from the oral cavity to the antennal apices and
flanked on each side by extremely broad, flattened and densely setulose facial ridges) (Figs 3−4); and the
postsutural intra-alar seta absent (sometimes weakly developed).
Description. Head: Male holoptic (Figs 1, 3), female dichoptic (Fig. 4). Ocelli present. Female frons
conspicuously broad, while male holoptic with extremely narrow frons. Antenna very short, its length much less
than 1/4 of eye-height. Face oestriform, i.e., forming a long narrow strip from oral cavity to antennal apex and
flanked on each side by broad, flattened and densely short setulose facial ridges (Fig. 3). Vibrissa not differentiated,
as well as subvibrissal and subcranial setae (Fig. 3). Oral cavity very reduced and subcircular; palpus very short,
with short black setulae apically; labella and prementum reduced. Multiple irregular rows of black postocular
setulae from outer vertex ventrally to postgena, and then with pale occipital setulae towards center; with no setulae
(either black or pale) at back of vertex and with only pale setulae behind oral cavity. Occiput concave middorsally.
Thorax: Densely covered with short black setulae on dorsum and laterally, except for paired stripes on scutum that
are non-pruinose, bare, one between acrostichal and dorsocentral rows and another between dorsocentral and intra-
alar rows. Usual thoracic bristling not strongly developed. Acrostichal setae 0+1; dorsocentral setae 2+4 (weakly
developed, only the last postsutural setae conspicuous); postpronotal setae 2 (outer one stronger than inner one,
accompanied by long fine setulae); intra-alar setae absent (sometimes with weak adventitious setae); supra-alars
Zootaxa 3931 (1) © 2015 Magnolia Press · 3SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

1+1 or 1+2 (prealar seta not differentiated). Notopleural setae 2, with dense short black setulae on notopleuron.
Postalar setae 2, subequal in length, with dense short black setulae on postalar callus. Prosternum bare and notably
developed as bilobed (bilaterally symmetrical) inflated apparatus with hemispherical depression on each side.
Proepisternum bare, 2 strong proepisternal setae; 1 strong proepimeral seta; katepisternal setae 2; katepimeron
(barette) bare. Six strong anepisternal setae on posterior margin, and several long upcurved setulae on
upperanterior corner, anepisternum covered with black fine setulae. Anepimeron with several long fine black
setulae on upper and lower portions, without any developed seta. Scutellum with 2–3 lateral and one apical pairs of
setae. Meron with tuft of long fine setae, but no regular row distinguishable.
Wing (Figs 11–12): Tegula (epaulet) densely short setulose; costal spine not differentiated; cell r4+5
closed far
before wing margin with very long petiole (over twice as long as r-m). Rs node setulose dorsally and ventrally;
distance of M bend to wing margin subequal to length of r-m. Noticeable infuscation on some wing veins.
Legs: Fore femur with dorsal, posterodorsal and posteroventral rows of setae, posterodorsal row irregular. Mid
femur without developed setae, except for some basoventral setae and 2 weak preapical posteroventral setae; no
median anterior seta. Mid tibia with one anterodorsal seta at apical third, and one ventral seta at apical third. Hind
tibia with one anteroventral seta at apical third.
Abdomen: Globose in both dorsal and lateral views, conspicuously broader than thorax (Fig. 1), densely
covered with fine black setulae, those setulae longer on tergite 5. Syntergite 1+2 with medial excavation extending
to posterior margin. Abdominal setae not differentiated (median marginals, lateral marginals, and discals all
absent). Sternite 1 widely setulose.
Male terminalia (Figs 6–7) (based on A. maculithorax): Cerci strongly fused (no suture recognizable) with the
posterior apex acuminate in both dorsal and lateral views; surstylus strongly arcuate inwardly (Fig. 6); bacilliform
sclerite (processus longus) broad, articulated (not fused) with surstylus and hypandrium; phallus (aedeagus) (Fig.
7) with basiphallus connected to phallapodeme at 180° angle, then distiphallus ventrally directed; phallic guide
elongate and slightly curved anteriorly, between the pregonites, at 180° angle with phallapodeme (Fig. 7);
postgonite slender and acuminate; pregonites free (not fused with each other and not fused to hypandrium) and
long, reaching lower posterior margin of hypandrium in lateral view; ejaculatory apodeme narrow and curved;
hypandrium deeply concave, half-bowl shaped, and longer than phallapodeme.
Female terminalia (Fig. 8) (based on A. maculithorax): Tergite 6 separated into hemitergites, with row of
spiniform straight setae on inner margin, the right hemitergite without spiracle and the left hemitergite with one
spiracle and a second spiracle segregated in the membrane (probably originated from tergite 7 sec Herting 1957);
tergite 8 separated into narrow hemitergites (not connected to each other, and not connected or fused to sternite 10);
sternite 10 (postgenital plate or hypoproct) covered with fine setulae and without any spiniform or strong setae/
setulae; cerci partially fused to each other basomedially, widely covered with fine long setulae, and basolaterally
with dense rather stronger setulae.
Key to species of Aulacephala
The key below was prepared based upon characters previously reported by Bequaert (1922), Uéda (1960), Mesnil
(1973), as well as my present observations.
1. Crossvein dm-cu strongly bisinuate (Fig. 12); final section of M vein (distal to bend) strongly arcuate inwards (Fig. 12); base
of wing, r-m, dm-cu and final section of M dark brownish coloured and conspicuously infuscated around (Fig. 12) (widespread
in continental Afrotropics) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . A. maculithorax Macquart
- Crossvein dm-cu weakly bisinuate (Fig. 11); final section of M vein (distal to bend) is straight or only slightly arcuate inwards
(Fig. 11); base of wing, r-m, dm-cu and final section of M light brownish to brownish coloured (at most r-m dark brownish),
and r-m with conspicuous infuscation around, while dm-cu, final section of M, and wing base with some weak infuscation
(Fig. 11) (Japan, China, Indonesia) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .A. hervei Bequaert
Aulacephala maculithorax Macquart
(Figs 1–8, 12, 39)
maculithorax Macquart, 1851: 139 (166), pl. XV, fig. 6. Lectotype female (MNHN) desig. by Crosskey, 1971: 264 (not examined). Type locality: Madagascar.
NIHEI 4 · Zootaxa 3931 (1) © 2015 Magnolia Press

Aulacocephala badia Gerstaecker, 1863: 1035. Holotype female (ZMHB) (examined by photos). Type locality: South Africa (as “Caffraria”).
Aulacocephala badia; Brauer, 1863: 170, fig. 9 (female); Malloch, 1925: 147, fig 1 (comments, wing illustration); Bau, 1928: 298 (comments).
Aulacephala badia; Ricardo, 1901: 109 (list).Aulacocephalopsis badia; Townsend, 1918: 165 (comments on male); Townsend, 1936: 115 (list, key to world Aulacephalini).Aulacocephala maculithorax; Brauer, 1863: 191, pl. III, fig. 5 (female); Verbeke, 1963: pl. II, fig. 17 (male phallus).Aulacephala maculithorax; Bigot, 1859: 533 (comments on female, Madagascar); Bequaert, 1922: 303, fig 1a (synonymy of
badia, synonymy of Aulacocephalopsis, wing illustration); Séguy, 1926: 6, figs 8 and 9 (head and wing); Bequaert, 1929: 163 (comments, ?Madagascar); Townsend, 1936: 115 (list, key to world Aulacephalini); Townsend, 1938: 256 (incorrect information about the type being in Lille Museum or lost); Crosskey, 1971: 264 (lectotype designation, type data); Mesnil, 1973: 1229 (key to species); Crosskey, 1980: 836 (catalogue).
Distribution (Fig. 39). Botswana, Cameroon, Democratic Republic of the Congo, Kenya, Liberia, ?Madagascar
(see comments below), Malawi, Mozambique, Nigeria, South Africa, Tanzania, Uganda, and Zambia.
Redescription. Male. Body length: 9.5–11.8 mm (mean=10.9), wing length: 9.0–11.2 mm (mean=10.4) (n=7).
Coloration: Head brown with whitish pruinosity on face, parafacial, fronto-orbital plate, facial ridge and gena;
ocellar triangle brown (Fig. 3). Scape and pedicel yellowish, postpedicel light reddish brown but base and apex
yellowish, arista yellow (Fig. 3). Palpus and proboscis yellowish. Thorax brown with whitish pruinosity, scutum
with non-pruinose, bare paired stripes, one between acrostichal and dorsocentral rows and another between
dorsocentral and intra-alar rows (Fig. 1). Wing hyaline (Figs 1, 12), wing veins yellow but basal parts of veins, r-m,
dm-cu and final section of M (distal to bend) brown to dark brownish; with conspicuous infuscation around basal
part of veins, r-m, dm-cu and final section of M; tegula (epaulet) brown to dark brown; calypters light brownish on
disc and borders; halter yellow, knob brownish. Legs yellowish, with whitish pruinosity on coxae, femora and
tibiae; tarsomeres yellow (Fig. 2). Claws light brown with black apex; pulvilli yellow. Abdomen brown, with
posterior margins of tergites 1+2, 3 and 4 darkened, some specimens also with posterior margin of tergites 1+2, 3
and 4 noticeably marked with narrow yellow transverse line (like A. hervei); tergites 3, 4 and 5 with dense whitish
pruinosity on anterior 2/3 (Fig. 1).
Head: Eye bare, at most with very short and sparse setulae; inner anterior ommatidia enlarged. Frontal vitta
extremely narrow so that fronto-orbital plates touch each other on upper half, although broadening ventrally (Fig.
3). Fronto-orbital plate covered with short and fine setulae from midlevel downwards to level of antennal insertion;
no developed setae on frontal vitta and fronto-orbital plate (Fig. 3). Ratio of frons width/head width at level of
lunula 0.10–0.14; ratio of frons width/head width at vertex level 0.02–0.035 (Fig. 3). Ocellar triangle constricted
and swollen (visible in profile), with few setulae; with three developed ocelli, shining-coloured, and easier to
recognize than in female. Fronto-orbital plate as narrow as arista width on upper half, and about 2x arista width at
lowermost level (Fig. 3). Parafacial bare and as wide as upper portion of fronto-orbital plate (Fig. 3). Antennal axis/
head height ratio 0.49–0.59 (Fig. 2). Facial ridge narrower than in females (compare Figs 3 and 4). Antenna short;
postpedicel about 1.5x pedicel length in lateral view; arista elongate and slender, about 3.5x postpedicel length
(Fig. 3). Genal dilation less developed (more discrete, more flattened) than in female, and densely covered with
short black setulae. Genal height/head height ratio 0.07–0.11 (Fig. 2).
Thorax: Postsutural intra-alar setae not differentiated, although some specimens might have one developed
seta posteriorly. Supra-alar setae 1+1, postsutural one in median position. Mesothoracic spiracle partially exposed
dorsally and ventrally, with light brownish branched hairs on anterior and posterior edges, but with longer hairs on
lower 2/3; metathoracic spiracle not exposed, completely covered by short anterior lappet and opercular posterior
lappet with brown branched hairs. Scutellum with three lateral and one apical pairs of setae, anteriormost lateral
and apical pairs stronger, and with dense fine black setulae covering dorsal, lateral and lateroventral areas (the
latter with long setulae).
Wing (Figs 2, 12): R1 with some tiny setulae on the very apex, and not sinuous on basal third; final section of M
vein (distal to bend) strongly convex inwards; dm-cu strongly bisinuate; anterior end of dm-cu closer to r-m than to
wing margin, but in some specimens about same distance from r-m and wing margin; and r-m placed about halfway
between dm-cu and bm-cu.
Legs: Fore tibia with no anterodorsal seta, and 2 developed posterodorsal setae at middle. All claws slightly
shorter than last tarsomere, and pulvilli slightly shorter than claws. Mid tibia with 1–2 posterodorsal setae on apical
half. Hind femur with dorsal row of setae, with longer setae at base; anteroventral row of setae, with longer setae at
Zootaxa 3931 (1) © 2015 Magnolia Press · 5SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA
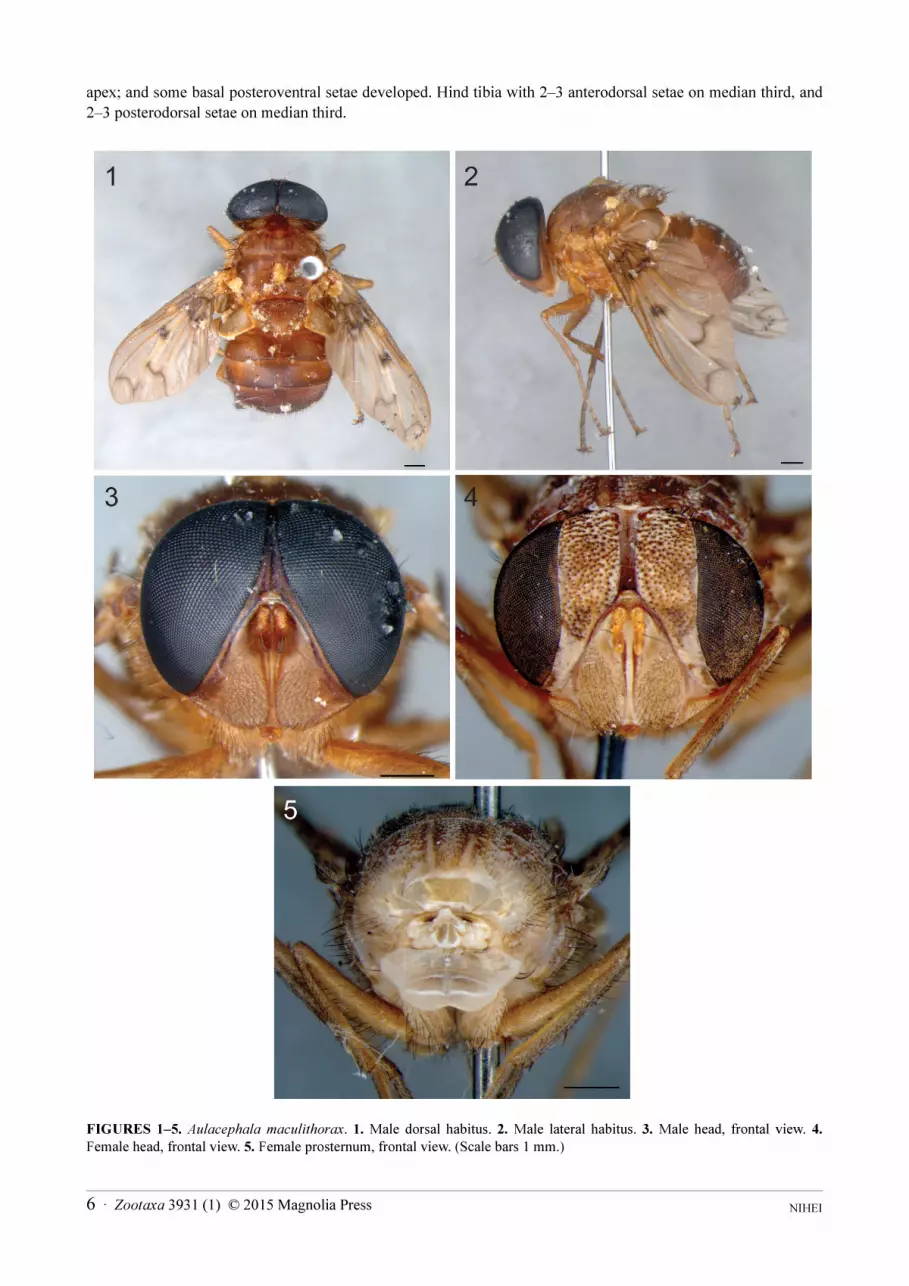
apex; and some basal posteroventral setae developed. Hind tibia with 2–3 anterodorsal setae on median third, and
2–3 posterodorsal setae on median third.
FIGURES 1–5. Aulacephala maculithorax. 1. Male dorsal habitus. 2. Male lateral habitus. 3. Male head, frontal view. 4.
Female head, frontal view. 5. Female prosternum, frontal view. (Scale bars 1 mm.)
NIHEI 6 · Zootaxa 3931 (1) © 2015 Magnolia Press

Terminalia (Figs 6–7): Cerci strongly fused (no suture recognizable) with posterior apexes free and strongly
divergent (Fig. 6); apex acuminate in both dorsal and lateral views (Figs 6–7); cerci slightly longer than wide
(about 1.4x). Surstylus strongly arcuate inwardly (Fig. 6); with broad and rounded apex in both dorsal and lateral
views (Figs 6–7), and with basomedial lobes bearing fine setulae hidden behind the cerci (Fig. 6). Bacilliform
sclerite (processus longus) broad, articulated (not fused) with surstylus and hypandrium. Phallus (aedeagus) (Fig.
7) with basiphallus connected to phallapodeme at 180° angle, then distiphallus ventrally directed. Phallic guide
elongate and slightly curved anteriorly, between the pregonites, at 180° angle with phallapodeme. Postgonite
slender and acuminate. Pregonites free (not fused with each other and not fused to hypandrium) and long, reaching
lower posterior margin of hypandrium in lateral view. Ejaculatory apodeme narrow and curved. Hypandrium
deeply concave, half-bowl shaped, and longer than phallapodeme.
FIGURES 6–8. Aulacephala maculithorax. 6. Male terminalia, dorsal view. 7. Male terminalia, lateral view. 8. Female terminalia, posterior view. (Legends: bac scl, bacilliform sclerite; basph, basiphallus; cer, cerci; distph, distiphallus; ej apod, ejaculatory apodeme; epand, epandrium; hypd, hypandrium; phapod, phallapodeme; ph gd, phallic guide; posgt, postgonite; pregt, pregonite; spr, spiracle; s8, sternite 8; s10, sternite 10; sur, surstylus; t6, tergite 6; t8, tergite 8.)
Female. Body length: 9.8–13.7 mm (mean=11.9), wing length: 9.5–12.1 mm (mean=11.1) (n=7).
Differs from male by the following: Coloration: Ocellar triangle yellow to light brownish; frontal vitta reddish
brown on upper half (Fig. 4); fronto-orbital plate darker on upper portion and some yellowish to reddish brown at
antennal level (Fig. 4); facial ridge with some yellowish areas (Fig. 4); gena yellow to light brownish; tegula
(epaulet) brown to dark brownish. Head. Dichoptic (Fig. 4); inner anterior ommatidia not enlarged. Frons
extremely broad, and narrowing very slightly at vertex (Fig. 4). Ratio of frons width/head width at level of lunula
0.48–0.50 (Fig. 4); ratio of frons width/head width at vertex level 0.45 (Fig. 4). Frontal setae very weakly
developed and medioclinate (Fig. 4). Fronto-orbital plate covered with short and fine setulae from vertex ventrally
and continuing until upper half of parafacials, setulae on inner upper half rather longer (Fig. 4); no developed setae
Zootaxa 3931 (1) © 2015 Magnolia Press · 7SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

on frons. Frontal vitta narrow, about 1.5x pedicel width at midlevel, broadening dorsally when approaching ocellar
triangle, about or slightly over 2.0x pedicel width, and ventrally broadening slightly at level of lunula hardly 2.0x
pedicel width (Fig. 4). Ocellar triangle developed and flattened (not constricted and swollen as in males) (Fig. 4),
with few setulae, bearing three ocelli that are not as developed as in males, ocelli pale in colour and not
conspicuously convex (although not flattened). Fronto-orbital plate notably broad and narrowing inconspicuously
towards vertex, about 3.0–4.0x width of frontal vitta at midlevel (Fig. 4). Parafacial densely setulose on upper half
(continued from fronto-orbital plate), narrowing notably from level of antennal insertion towards gena, upper width
2.0–2.5x lower width (Fig. 4). Facial ridge about 4/5 width of fronto-orbital plate. Antennal axis/head height ratio
0.49–0.56. Genal height/head height ratio 0.19–0.23. Legs: Fore tibia with 1–2 developed anterodorsal setae. Claws
shorter than in males. Terminalia (Fig. 8): Tergite 6 separated into hemitergites, with continuous row of spiniform
straight setae on inner margin from median third until half of apical third and two outer setae at middle, the right
hemitergite without spiracle and the left hemitergite with one spiracle and a second spiracle segregated in the
membrane (probably originated from tergite 7 sec Herting 1957). Tergite 8 separated into narrow hemitergites (not
connected to each other, and not connected or fused to sternite 10). Sternite 10 (postgenital plate or hypoproct)
covered with fine setulae, without any spiniform or strong setae/setulae. Cerci partially fused to each other
basomedially, widely covered with fine long setulae, and basolaterally with dense rather stronger setulae. Three
oblong spermathecae, two equal-sized and one smaller.
Type material examined. Holotype female of Aulacocephala badia Gerstaecker (ZMHB), labeled “Afr. austr.
Drége” (dark-green label); “773”; “Type” (red label); “badia / Gerst.” (dark-green label); “Aulacephala /
maculithorax Mcq. / det. Zumpt 57”. Specimen examined by photos kindly sent by James O’Hara (Canadian
National Collection, Ottawa). Specimen in good condition, only left foreleg missing.
Examined material. BOTSWANA: Xugana Is., B. Lamoral Woodland, 19°04′S: 23°03′E, Malaise trap, 1
male, 18–25.xi.1980, 1 male, 22–26.xi.1979 (NMSA); CAMEROON, Rég. de Dchang, Plateaux Volcaniques,
1400m, 1 male, Gromier leg. (USNM); DEMOCRATIC REPUBLIC OF CONGO: N. Lac Kivu, Rwankwi, 1
female, v.1948, J.V. Leroy leg. (BMNH); Kasai River, 1 female, xii.1927, Lt. J. Ghesquière leg. (BMNH);
KENYA, Nairobi, 1 male, iii.1926, van Someren leg. (BMNH); LIBERIA, 1 female, 16.v.1945, M.S. Briscoe leg.
(USNM); MALAWI (“Nyassaland”): Fort Johnston [Mangochi], 1 female, 1911, P. Rendall leg. (BMNH); near
Kaporo, N. Nyassa, 1 male, 13.viii.1909, J.B. Davey leg. (BMNH); Maiwale, 1 female, 22.x.1928, W.A. Lamborn
leg. (BMNH); “Nyassaland”, 1 female, 1912, E.d. Rhoades leg. (BMNH); Cholo, 2700ft, 1 male, v.1920, R.C.
Wood leg. (BMNH); MOZAMBIQUE (“PORTUG. E. AFR”), east of Mt. Mlanje [Mt. Mulanje], 1 male,
3–7.x.1913, S.A. Neave leg. (BMNH); NIGERIA, Baro, 1 female, x.1910, Dr. Ingram leg. (BMNH); SOUTH
AFRICA: Transvaal, Potchefstroom (“Potchestm”), 1 female, no date, T. Ayres leg. (BMNH); Pretoria, 1 female,
07.iii.1917, G.A.H. Bedford leg. (BMNH); Pretoria, 1 male, no additional data (USNM); Natal, Weenen, 1 male,
x–xi.1927, H.P. Thomasset leg. (BMNH); Natal, Lindeque Spruit, 14km SE Bergville, 1 female, 19.ii.1978, D. &
M. Davis & B. Akerbergs leg. (USNM); ?country/location, “Caught in Harris”, O’Paort (?leg), 1 female,
04.iii.1960 (BMNH); Pietermaritzburg, 1 male, 18.x.1961, B. & P. Stuckenberg leg. (NMSA); Tongaland,
Ingwavuma District, Ndumu Reserve, 2 females, 1–10.xii.1963, B. & P. Stuckenberg leg. (NMSA); Merrivale,
Stuckenberg House, 30°05′E, 29°30′S, 1051m, 1 male, 3.xi.2001 (NMSA); Cape Good Hope Nature Reserve, 1
female, 7–10.iii.1968, P. Spangler leg. (USNM); Cape Province, Wilderness, 1 female, 12–13.iii.1968, P. Spangler
leg. (USNM); Cape Province, Stormsrivier, 2.5km S, 3 females, 11–12.iii.1978, D. & M. Davis & B. Akerbergs
leg. (USNM); Cape Province, Thomas Baines Nature Reserve, 13km SW Grahamstown, Palmiet R., 2 females,
6.iii.1978, D. & M. Davis & B. Akerbergs leg. (USNM); Cape Province, Brackenhill Falls, 9km E Knysna, 5
females, 15–16.iii.1978, D. & M. Davis & B. Akerbergs leg. (USNM); Cape Province, Diepwalle Plantation, 39km
NE Knysna, 1 female, 13.iii.1978, D. & M. Davis & B. Akerbergs leg. (USNM); Mpumalanga Province, Barberton
District, Farm Alfa 448JU, 1 female, 22–26.xi.1972, A. Strydom leg. (NMSA); Algeria Forestry, Clanwilliam
District, 1 female, 4–10.iii.1969, Potgieter & Strydom leg. (NMSA); Harkerville, Forest Reserve, 1 female,
1–5.iii.1972, L. & G. Vari leg. (NMSA); Groenkloof, 1 male, 19.xi.1921, A. Roberts leg. (NMSA); TANZANIA,
Tabora, Igombe river, 1 female, x.1948, van Someren leg. (BMNH); UGANDA: Kampala, 1 female, i.1963,
A.W.R. McRae leg. (at light) (BMNH); Kawanda, 1 male, 16.iv.1965, 4 females, 29.ix.1972, D.J. Greathead leg.
(BMNH); Kalinzu Forest, 1 male, 1935, T.H.E. Jackson leg. (BMNH); and ZAMBIA: Lochinvar Nat. Park, 1 male,
10.v.1979, 1 female, 13.v.1978, G.W. Howard leg. (BMNH); Roma, Lusaka, 1 male, 29.xi.1979, G.W. Howard leg.
(BMNH). [One male from Zambia/Roma-Lukasa and one female from Zambia/Lochinvar were dissected and
examined.]
NIHEI 8 · Zootaxa 3931 (1) © 2015 Magnolia Press

Synonymy of Aulacocephala badia Gerstaecker: During the course of my revision, I had not seen the female
holotype of Aulacocephala badia Gerstaecker, 1863 deposited at Berlin (ZMHB), but only specimens studied and
determined by Bezzi (1911), Townsend (1918) and Sabrosky, which were examined during a visit to the USNM
collection. Recently, James O’Hara (Canadian National Collection, Ottawa) very kindly provided me some
photographs of the holotype taken during his visit to the Berlin museum in summer 2014. With those photographs
at hand, I could then confirm the synonymy of A. badia Gerstaecker with A. maculithorax Macquart.
Madagascar as the type locality. Bequaert (1919: 163) doubted that the type specimens studied by Macquart
to describe A. maculithorax were really collected from Madagascar: “no specimen of Aulacephala has been taken
in that island since”. And he goes further in suggesting that probably Macquart’s specimens were from South
Africa. And then, since Bequart’s comment in 1919, no material from Madagascar has been reported.
Aulacephala hervei Bequaert
(Figs 9–11, 39)
hervei Bequaert, 1922: 305, fig 1b (female description, wing illustration). Holotype female (BMNH). Type locality: Japan, Honshu, Yokohama District.
Aulacocephala karnyi Malloch, 1925: 147, fig 2 (male description, wing illustration). Holotype female (USNM). Type locality: Indonesia, Sumatra, Way Lima.
Aulacephala hervei; Bequaert, 1929: 164 (synonymy of karnyi with hervei); Uéda, 1960: 14, figs 1–6 (redescription, comments, 1st instar larva description and illustrations, Japan); Mesnil, 1973: 1229 (key to species); Crosskey, 1976: 63, 185, 280 (comments, cat.); Herting & Dely-Draskovits, 1993: 354 (cat.); Chao & Xue, 1998: 1954 (cat., China); O’Hara et al. 2009: 160 (cat., China); Shima, 2014: 875 (cat., Japan).
Aulacephala karnyi; Mesnil, 1973: 1229 (synonymy with hervei); Crosskey, 1976: 63, 185, 280 (comments, cat., mentions type as lost – “not located”).
Ormiominda punctigera Paramonov, 1955: 128. Holotype female (ANIC). Type locality: Papua New Guinea, Wewak. Syn.
nov.
Therobia punctigera; Crosskey, 1966: 103 (new combination, synonymy of Ormiominda Paramonov with Therobia); Cantrell & Crosskey, 1989: 749 (cat.).
Distribution (Fig. 39). China, Indonesia (Sumatra), Japan, and Philippines.
Redescription. Female. Body length (holotype): 10.9 mm, wing length (holotype): 10.3 mm. Body length (non-
types): 8.8–11.3 mm (mean=10.2), wing length (non-types): 8.3–10.4 mm (mean=9.3) (n=8).
Coloration: Head brown with whitish pruinosity on face, parafacial and fronto-orbital plate; ocellar triangle
yellowish; parafacial yellowish at level of antennal insertion; facial ridge mostly brown but yellowish dorsally;
gena yellowish. Scape and pedicel yellow. Palpus and proboscis yellowish. Thorax brown dorsally with whitish
pruinosity, but yellowish between dorsocentral and intra-alar rows, and laterally brownish and partially yellowish
with whitish pruinosity. Wing hyaline, wing veins yellow but basal parts of veins, final section of M (distal to
bend), and r-m and dm-cu crossveins light brownish to brownish (at most r-m dark brown); r-m conspicuously
infuscated around and some basal parts of veins, final section of M (distal to bend) and dm-cu only weakly
infuscasted; tegula (epaulet) light brown; calypters light brownish on disc and borders; halter yellow, knob slightly
brownish. Legs yellowish, pruinosity not apparent as they are covered with fungi hyphae (see male description
below); fore tarsomeres yellow (other tarsomeres missing). Fore claws yellow with black apex; pulvilli yellow
(other claws and pulvilli missing). Abdomen brown, with posterior margins of tergites 1+2, 3 and 4 darkened and
noticeably marked with narrow yellow transverse line; all tergites with whitish pruinosity but tergites 3 and 4 with
notable denser pruinosity on anterior 2/3.
Head: Eye bare. Frons extremely broad, narrowing very slightly at vertex. Ratio of frons width/head width at
level of lunula 0.55 (non-types: 0.54–0.58); ratio of frons width/head width at vertex level 0.45 (non-types:
0.44–0.49). Frontal setae very weakly developed and medioclinate. Fronto-orbital plate densely covered with short
setulae from vertex ventrally until middle of parafacial; no developed setae on fronto-orbital plate. Fronto-orbital
plate notably broad and narrowing inconspicuously towards vertex, about 3.5–4.0x frontal vitta width at midlevel.
Frontal vitta notably narrow, subequal in width to pedicel at uppermost level (approaching ocellar triangle) and
broadening quite slightly at level of lunula with hardly 1.5x pedicel width. Three reduced ocelli, hardly
recognizable (reduced in size, pale in colour and flattened); with a few sparse setulae on ocellar triangle. Parafacial
Zootaxa 3931 (1) © 2015 Magnolia Press · 9SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

densely setulose on upper half (continued from fronto-orbital plate), narrowing notably from level of antennal
insertion towards gena, upper width about 3x lower width. Antenna presumably short, with antennal depression on
face very small; postpedicel missing (see male description below). Antennal axis/head height ratio 0.61 (non-types:
0.44–0.58). Genal dilation densely covered with short setulae. Genal height/head height ratio 0.25 (non-types:
0.23–0.26).
Thorax: Dorsal and lateral chaetotaxy very damaged (some missing, others covered with fungi), but the
following features are recognizable (see the complete male description below): thorax with dense coverage of short
black setulae on dorsum and laterally; presutural seta 1, aligned with supra-alar row; postsutural supra-alar setae 2
(some specimens with 1, but always in median position), prealar seta not differentiated (although some specimens
with one rather developed setula found at same position), anterior seta slightly stronger than posterior. Scutellum
with 2 lateral and 1 apical pairs of setae (all recognized by insertions), with dense fine black setulae dorsally,
laterally and lateroventrally (the latter with long setulae).
Wing: R1 not sinuous on basal third; final section of M vein (distal to bend) straight or only slightly convex
inwards; dm-cu weakly bisinuate; anterior end of dm-cu closer to wing margin than to r-m; and r-m placed about
halfway between dm-cu and bm-cu.
Legs: Fore claws short, about 2/3 length of last tarsomere; and pulvilli slightly shorter than claws. All other
legs covered with fungi, please see male description below for other leg features.
Terminalia: female holotype not dissected.
FIGURES 9–12. Aulacephala hervei. 9. Male dorsal habitus. 10. Male lateral habitus. 11. Female wing. Aulacephala
maculithorax. 12. Male wing. (Male specimen from Japan, Yokohama; female from Japan, Matsuyama.) (Scale bars 1 mm.)
Male. Body length: 8.9 mm, wing length: 8.0 mm (n=1).
Coloration: Head brown with whitish pruinosity on face, parafacial, fronto-orbital plate, facial ridge and gena;
ocellar triangle brown. Scape and pedicel yellow, postpedicel reddish yellow, arista yellow. Palpus and proboscis
yellowish. Thorax brown with whitish pruinosity, scutum with non-pruinose, bare paired stripes, one between
acrostichal and dorsocentral rows (restricted to prescutum) and another between dorsocentral and intra-alar rows.
Wing hyaline, wing veins yellow but basal parts of veins, r-m and final section of M (distal to bend) brownish; r-m
conspicuously infuscated around while some basal parts of veins, final section of M (distal to bend) and dm-cu
only weakly infuscasted; tegula (epaulet) yellow; calypters light brownish on disc and borders; halter yellow, knob
brownish. Legs yellowish, with whitish pruinosity on coxae, femora and tibiae; tarsomeres yellow. Claws yellow
NIHEI 10 · Zootaxa 3931 (1) © 2015 Magnolia Press

with black apex; pulvilli yellow. Abdomen brown, with posterior margins of tergites 1+2, 3 and 4 darkened and
noticeably marked with narrow yellow transverse line; tergites with whitish pruinosity that is notably denser on
anterior 2/3 of tergite 4.
Head: Eye bare, at most very short and sparse setulae; inner anterior ommatidia enlarged. Fronto-orbital plate
covered with short and fine setulae from midlevel downwards to level of antennal insertion; no developed setae on
frons. Frontal vitta extremely narrow so that fronto-orbital plates almost touch each other on upper half, although
broadening ventrally. Ratio of frons width/head width at vertex level 0.05; ratio of frons width/head width at level
of lunula 0.20. Ocellar triangle constricted and swollen (visible in profile, although not as swollen as in A.
maculithorax), and with few setulae, with three reduced ocelli, shinning-coloured, not flattened and easier to
recognize than in female. Fronto-orbital plate as narrow as arista width at upper half, and about 2x arista width at
lowermost level. Parafacial bare and as wide as upper level of fronto-orbital plate. Antennal axis/head height ratio
0.86. Facial ridge less broad than in females. Antenna short; postpedicel as long as pedicel; arista elongate and
slender, about 4x postpedicel length. Genal dilation less developed (more flattened) than in female, and densely
covered with short black setulae. Gena heigh/head height ratio 0.08.
Thorax: Supra-alar setae 1+1, postsutural one in median position. Mesothoracic spiracle partially exposed
dorsally and ventrally, with light brownish branched hairs on anterior and posterior edges, but with longer hairs on
lower half; metathoracic spiracle not exposed, completely covered with short anterior lappet and opercular
posterior lappet with brown branched hairs. Scutellum with three lateral and one apical pairs of setae, anteriormost
lateral and apical pairs stronger, and with dense fine black setulae dorsally, laterally and lateroventrally (the latter
with long setulae).
Wing: Similar to female except for apex of R1 with some few setulae dorsally.
Legs: Fore tibia with 1 weak submedian anterodorsal seta, and posterodorsal irregular row with longer setae at
middle. All claws slightly shorter than last tarsomere; and pulvilli slightly shorter than claws. Mid tibia with
posterodorsal irregular row with some longer setae on median third. Hind femur with anteroventral row of setae
with the longest setae at apex; posteroventral row of setae with the longest setae at base. Hind tibia with 2
anterodorsal setae on median third, and 2 posterodorsal setae on median third.
Abdomen: Similar to female, clearly broader than thorax, but not as conspicuously as in females.
Terminalia: Not dissected.
Type material examined. Holotype female of Aulacephala hervei (BMNH), “HOLO- / TYPE” (white round
label with red border), “Yokohama District. / Japan / H. Prior. / 1901-13.”, “Aulacephala / hervei J. Beq. / type / J.
Bequaert. det.”. Specimen in bad condition, the right wing, foreleg and midleg missing, the left wing with the tip
damaged, antennae missing, ventral side completely covered by fungal hyphae.
Holotype female of Aulacocephala karny (USNM) labeled “Wai [Way] Lima, Z Sum. / Lampongs [Lampung]
/ Karny / XI.XII 1921. No.451”, “Type No / USNM” [red label], Aulacocephala / karny / Type / Det. / J.R.
Malloch” [Malloch’s handwriting].
Holotype female of Ormiominda punctigera (ANIC) labeled “WEWAK / N. Guinea / FHTaylor”,
“Ormiominda / punctigera / sp. nov. [female symbol] Typus / Paramonov det.” [Paramonov’s handwriting];
“ANIC” [light green label with black border].
Additional material examined. CHINA: S. [South] of Suifu, 1 female, D.C. Graham leg. (USNM); Suifu, Sz.
[Sichuan], 1 female, 1.vi.30, D.C. Graham leg. (USNM); JAPAN: Yokohama, 1 male, v.1906, Brunetti leg.
(BMNH); Shikoku, Matsuyama: 1 female, 25.ix.1951, T.I, M.M. & T.E. leg.; 1 female, 2.viii.1948, M. Miyatake
leg.; 1 female, 28.vi.1949, Miyatake leg. (EUMJ); Matsuyama, Tarumi: 1 female, 4.x.1953, M. Miyatake leg.; 1
female, 28.ix.1955, light trap; 1 female, 10.ix.1955, light trap (EUMJ); Shinmura, Amami-Ooshima: 1 female,
17.vii.1954, S.H., S.U. & T.E. leg.; 1 female, 21.vii.1954, 2 females, 19.vii.1954, S. Hiramatsu leg. (EUMJ);
Kyushu, Fukuoka, Mt. Tachibana, 1 female, 14.viii.1978, T. Goto leg. (BLKU); Okinawa, Hentona, 1 female,
10.ix.1945, light trap, G.E. Bohart leg. (USNM); PHILIPPINES, Luzon, Mt. Makiling, 1 female, Baker leg.
(USNM).
Remarks. Besides the holotype female (BMNH) from Yokohama described by Bequaert, a reasonably large
and representative series of specimens from Japan were gathered and available for examination: one male from the
type locality, Yokohama, but collected by Brunetti; one female from Fukuoka (kindly sent by Hiroshi Shima); and
10 additional females from Matsuyama and Shinmura studied by Uéda (1960), who had examined 11 specimens,
all them females captured at light. To my knowledge, the only male available in collections is the Yokohama male
Zootaxa 3931 (1) © 2015 Magnolia Press · 11SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

collected by Brunetti and deposited at BMNH. Furthermore, the holotype female of Aulacocephala karnyi Malloch
was examined in the USNM and the synonymy between hervei and karnyi is confirmed. I also examined the
holotype female (ANIC) of Ormiominda punctigera Paramonov, and concluded that it is conspecific with A.
hervei. This new synonymy is proposed herein.
Phasioormia Townsend
Phasioormia Townsend, 1933: 447. Type species: Phasioormia pallida Townsend, 1933, by original designation.Distribution (3 species) (Fig. 39). Oriental and Australasian.References. Townsend, 1936: 101 (key to world Ormiini); Crosskey, 1976: 64 (key to Oriental genera of Ormiini); Dear &
Crosskey, 1982: 119 (key to Philippine genera of Ormiini); Chao & Xue, 1998: 1954 (cat., key to Chinese genera of Ormiini).
Diagnosis. Phasioormia differs from other ormiine genera by having the ocelli absent (present in Aulacephala,
Homotrixa, Mediosetiger, Ormiophasia and most Ormia; absent also in Therobia and some Neotropical and
Nearctic Ormia); facial region not oestriform (not like Aulacephala and Therobia that have the face forming a long
narrow strip from the oral cavity to the antennal apices and flanked on each side by broad, flattened and densely
setulose facial ridges) (Figs 17–18, 23, 29–30); wing with cell r4+5
open, R4+5
ending at wing margin (cell r4+5
closed
in Aulacephala and Mediosetiger) (Figs 14, 25–26); in profile view, head hemispherical (Figs 19–20, 24, 28,
31–32) rather than the trapezoidal head found in Homotrixa (e.g., epistomal axis longer than antennal axis), which
has the lower facial margin conspicuously warped forward (unlike all the other Ormiini).
Description. Head: Male holoptic (Fig. 17), female dichoptic (Fig. 18). Upper ommatidia enlarged in male,
and not in female. Eye bare. Ocelli absent. Female with two pairs of proclinate fronto-orbital setae on upper half,
and one reclinate pair before ocellar triangle. Antenna short. Face broad and normally developed (not oestriform
like in Aulacephala and Therobia); facial ridge rather broadened, but not strongly flattened, about 2x pedicel width
at vibrissal level. Vibrissa developed, inserted well above lower facial margin (Fig. 17, 23); subvibrissal and
subcranial setae differentiated, fine long setulae on facial ridge also continuing and accompanying subvibrissal and
subcranial setae (Figs 23–24, 29, 31). Oral cavity and mouthparts developed, but palpus very short, filiform and
clavate at apex, not spatulate, with short black setulae apically. Postocular row of black setulae from behind outer
vertex ventrally until genal dilation, starting in single row but multiplied and mixed with occipital black setulae on
lower half, and then with black and pale occipital setulae towards the center, with pale setulae behind oral cavity.
No occipital setulae behind vertex.
Thorax: Coloration reddish-brown but slightly lighter on postpronotal lobe, notopleuron and lateral parts, with
whitish pruinosity but scutum with non-pruinose, bare paired stripes, one between acrostichal and dorsocentral
rows (restricted to prescutum) and another between dorsocentral and intra-alar rows. Acrostichal setae 0+2 or 1+2;
dorsocentral setae 2+3 or 3+3; postpronotal setae 2, outer one stronger than inner one, accompanied by long fine
setulae; intra-alar setae 1+1, presutural one lateral to postpronotal lobe; intrapostalar seta not differentiated; supra-
alar setae 1+2, prealar seta strongly developed, about 4/5 length of posterior one, and as long as postsutural intra-
alar seta. Postalar setae 2, posterior one stronger, with dense short black setulae on postalar callus. Nine strong
anepisternal setae on posterior margin, and several long upcurved setulae on upperanterior corner, anepisternum
covered with black fine setulae. Katepisternal setae 2, subequal, with black fine setulae above and below.
Proepisternum bare; one long proepisternal seta; 2 long proepimeral setae; katepisternal setae 2; katepimeron
(barette) with few setulae anteriorly. Anepimeron with several long fine black setulae on upper and lower portions,
upper portion with longer setulae but without any developed setae. Meron with posterior row of long setae
accompanied by several shorter, finer setae anteriorly. Scutellum with one basal, one lateral, one apical and one
discal pairs of setae, discal pair weaker; scutellum with dense fine black setulae dorsally, laterally and
lateroventrally.
Wing (Figs 13, 14, 25, 26): Tegula (epaulet) yellowish-brown, densely short setulose; calypters light brownish
on disc and borders; halter yellow. Wing hyaline, veins yellowish-brown, with no strong infuscation, but membrane
somewhat tinged with light brown. Costa spine not differentiated; R1 not sinuous on basal third. M vein ending at
wing margin, distance between apexes of M and R4+5
shorter than r-m; bend of M curved at right angle, then
continuing very convex inwards until wing margin; distance of M bend to wing margin subequal to length of r-m;
NIHEI 12 · Zootaxa 3931 (1) © 2015 Magnolia Press

dm-cu bisinuate; anterior end of dm-cu closer to wing margin than to r-m; and r-m placed about mid distance
between dm-cu and bm-cu.
Legs: Coloration yellowish-brown, with whitish pruinosity on coxae, femora and tibiae (Fig. 15); tarsomeres
yellowish-brown. Fore femur with dorsal, posterodorsal and posteroventral rows of setae, the posterodorsal row
irregular. Claws about 2/3 length of last tarsomere, pulvilli slightly shorter than claws. Mid femur without
developed setae, except for one preapical posterodorsal seta; no median anterior seta. Mid tibia with 1 ventral seta
at apical third.
Abdomen: Globose in both dorsal and lateral views, broader than thorax. Tergites 1+2, 3 and 4 without
developed setae (median marginal, lateral marginal or discal setae), although their posterior margins with rather
elongate fine setulae. Syntergite 1+2 with medial excavation extending to posterior margin. Tergite 5 entirely
covered with elongate fine setulae, but with no differentiated setae. Sternite 1 widely setulose.
Male terminalia (Figs 33, 34, 36, 37): Cerci strongly fused (no suture recognizable) (Figs 33, 36) with the
posterior apex acuminate and curved in lateral view (Figs 34, 37); surstylus slightly arcuate inwardly (Figs 33, 36);
bacilliform sclerite (processus longus) broad, articulated with surstylus and hypandrium; phallus (aedeagus) (Figs
34, 37) with basiphallus connected with phallapodeme at 180° angle, then distiphallus ventrally directed; phallic
guide elongate and straight, between the pregonites, at 180° angle with phallapodeme (Figs 34, 37); postgonite
slender and acuminate; pregonites free (not fused with each other and not fused to hypandrium) and elongate,
reaching lower posterior margin of hypandrium in lateral view; ejaculatory apodeme narrow and curved;
hypandrium deeply concave, half-bowl shaped, and as long as phallapodeme.
Female terminalia (Figs 35, 38): Tergite 6 separated into hemitergites, with row of spiniform setae on inner
margin, and with single spiracle on each side basally; tergite 8 formed as a single narrow arc (fused
medioventrally) not fused or partially fused with sternite 10; sternite 8 with pair of lateral rounded protuberances
bearing strong setae; sternite 10 (postgenital plate or hypoproct) covered with fine setulae and bearing 2 spiniform
lateral setae and a medial row of 3–4 spiniform setae decreasing inwards; cerci partially fused with each other
basomedially, widely covered with fine long setulae, and laterobasally with dense rather stronger setulae.
Systematic affinities
According to some authors (Mesnil 1973, Crosskey 1976), Phasioormia and Homotrixa would be undoubtedly
similar to each other. In Homotrixa ocelli are present, but those authors questioned whether this character is a valid
generic character or not in Ormiini (ocelli are also absent in Therobia and some Neotropical and Nearctic Ormia).
At that time, Homotrixa was a monotypic genus with only the Taiwanese type species known and its holotype was
lost. Only recently, Barraclough & Allen (1996) described two Southwestern Australian species in Homotrixa.
Despite the long distance in range between the Australian species and the type species, at this moment they are our
only source of morphological knowledge for Homotrixa. Unfortunately, contacts with Chinese curators and
researchers have provided no clues to finding additional material of Homotrixa either from Taiwan or from
continental Chinese localities. David Barraclough (in Barraclough & Allen, 1996: 138) examined Phasioormia
pallida (BMNH) and, based on external morphology, considered Phasioormia distinct from the Australian
Homotrixa. A detailed, illustrated study of the male and female terminalia of Phasioormia and Homotrixa has not
been published before now. Here, the analysis of male and female terminalia supports the validity of Phasioormia
and Homotrixa as two valid genera. Their male terminalia share several similarities, but some differences are
readily observed: in Phasioormia the postgonite has an acuminate apex (round apex in Homotrixa), and the anterior
end of the hypandrium is as long as the phallapodeme (shorter than the phallapodeme in Homotrixa) (Figs 34, 37).
Comparison of the female terminalia shows even more substantial differences: in Phasioormia (Figs 35, 38) there
is one single spiracle on each hemitergite of tergite 6 (while in Homotrixa there are two spiracles on each plate;
Barraclough & Allen, 1996: 142, fig. 11, and present dissections); sternite 8 has two conspicuous lateral
protuberances (sternite 8 flat in Homotrixa); and tergite 6 with a single row of sparse spiniform setae on its inner
margin (with dense spiniform setae in Homotrixa).
Zootaxa 3931 (1) © 2015 Magnolia Press · 13SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

Key to species of Phasioormia
1. One presutural acrostichal seta moderately developed. Female: frontal margins, in anterior view, parallel-sided from midlevel
dorsally to vertex (Figs 18, 30); anteroventral margin of head (bearing vibrissal, subvibrissal and subcranial setae/setulae), in
profile, subparallel to anteroventral margin of eye (apparently diverging only slightly) (Figs 20, 32); gena short (Figs 20, 32);
ocellar triangle hardly recognizable as narrow, slightly protuberant strip, with some weak setulae (Figs 18, 30). Male: ocellar
triangle extremely constricted between eyes, and not visible, bare (Figs 17, 29); ratio of frons width/head width at level of
lunula 0.18 (Figs 17, 29). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
- No presutural acrostichal seta (although developed in the single female available). Female: frontal margins, in anterior view,
converging from the midlevel dorsally to vertex (Fig. 23); anteroventral margin of head, in profile, not subparallel to
anteroventral margin of eye, closer to eye margin at vibrissal level (anteriorly) and then diverging ventrally toward gena (pos-
teriorly), becoming conspicuously distant (Fig. 24); gena high (Fig. 24); ocellar triangle easily recognizable, broad and protu-
berant, with dense weak setulae (Figs 23–24). Male: ocellar triangle extremely constricted between eyes, but still recognizable,
with a few minute setulae; ratio of frons width/head width at level of lunula 0.26 (India, Malaysia, and China) . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .P. bicornis (Malloch)
2. Abdomen reddish-yellow on tergites 1+2, 3 and 4, but with the last two with dark brown posterior margins, concolorous to
tergite 5 that is entirely dark brown (Figs 13–14); notopleuron densely covered with fine black setulae. Female: frontal setae
numerous and irregularly spaced, not forming single linear row (Fig. 18); fronto-orbital plate broader, about 2.5x pedicel width
at lowermost level (Fig. 18); fronto-orbital plate with dense setulae on its entire length (Figs 18, 20) (Singapore, Philippines,
Sri Lanka, and China) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .P. pallida Townsend
- Abdomen entirely yellowish-brown to brown (Figs 25–26); notopleuron bare or with some sparse black setulae. Female: fron-
tal setae more or less forming a linear row and not strongly developed (Fig. 23); fronto-orbital plate narrower, about 2.0x pedi-
cel width at lowermost level (Fig. 23); fronto-orbital plate with sparse setulae on its entire length (Figs 23–24) (Indonesia and
Papua New Guinea). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . P. papuana sp. nov.
Phasioormia pallida Townsend
(Figs 13–20, 33–35, 39)
pallida Townsend, 1933: 448. Holotype female (BMNH). Type locality: Singapore.Phasioormia pallida; Crosskey, 1976: 185 (cat.); Dear & Crosskey, 1982: 119 (key, list, Philippines); Preston, 1993
(misidentification, =Ormia ochracea sec Evenhuis, 2003: 34); Nishida, 1997, 2002 (misidentification, = Ormia ochraceasec Evenhuis, 2003: 34); Chao & Xue, 1998: 1955 (cat., China); O’Hara et al. 2009: 161 (cat., China).
Distribution (Fig. 39). China, Philippines, Singapore, and Sri Lanka.
Redescription. Male. Body length: 9.5–10.2 mm (n=2) (paratype 10.2 mm), wing length: 7.2–10.3 mm (n=3)
(paratype 10.3 mm).
Coloration (Figs 13, 15, 17, 19): Head brown with whitish pruinosity on face, parafacial, fronto-orbital plate,
facial ridge and gena; lower margin of face and facial ridge yellowish-brown (Figs 17, 19). Scape and pedicel light
brownish, postpedicel dark brown, arista brown (Figs 17, 19). Palpus and proboscis light brownish (Figs 17, 19).
Thorax reddish-brown (Figs 13, 15) but tending to reddish-yellow on postpronotal lobe and notopleuron and also
on some lateral parts. Claws light brown with black apex; pulvilli yellow. Abdomen reddish-yellow on tergites 1+2,
3 and 4, with the last two with dark brown posterior margins (concolorous to tergite 5), with whitish pruinosity
apparent on all tergites (Figs 13, 15).
Head: Frons extremely narrow, on upper half as eyes approach each other only fronto-orbital plates visible as
quite narrow strips, but frons broadening ventrally (Figs 13, 17). Setae and/or setulae on frontal vitta and fronto-
orbital plate not apparent due to fungi covering paratype’s head dorsally, but on additional non-type specimens with
about 9 pairs of frontal setae on lower 2/3, shortening dorsally. Ocellar triangle extremely constricted and not
recognizable (Fig. 17). Fronto-orbital plate as narrow as pedicel width at lowermost level, without setulae (Fig. 17).
Ratio of frons width/head width 0.02 at vertex level; ratio of frons width/head width at level of lunula 0.18 (Fig.
17). Parafacial bare and as wide as fronto-orbital plate (Fig 17). Facial ridge rather broadened, about 2x pedicel
width at vibrissal level, and with dense fine long setulae on lower portion (Fig. 17). Antenna short; postpedicel
about 1.3x pedicel length; arista elongate and slender, about 3.5x postpedicel length (Figs 17, 19). Antenna inserted
below mid height of head in frontal view, and also below mid height of eye (Fig. 19). Antennal axis/head height
ratio 0.68 (Fig. 19). Antennal axis/vibrissal axis ratio 1.13; antennal axis/oral (epistomal) axis ratio 1.13; vibrissal
axis/oral axis ratio 1.00 (Fig. 19). Genal dilation not apparent, but gena densely covered with short black setulae.
Genal height/head height ratio 0.12 (Fig. 19). Occiput strongly concave middorsally.
NIHEI 14 · Zootaxa 3931 (1) © 2015 Magnolia Press

Thorax: Acrostichal setae 1+2, presutural pair anteriorly placed and not close to suture (a third anteriormost
postsutural seta weakly developed or undeveloped), prescutellar pair stronger. Dorsocentral setae 2+3. Notopleural
setae 2, subequal and close to each other, with dense short black setulae. Mesothoracic spiracle conspicuously
exposed, with light brownish branched hairs on whole anterior and posterior edges, anterior hairs becoming longer
towards upper second quarter, spiracle is exposed dorsally and ventrally with just the middle somewhat covered;
metathoracic spiracle not exposed, completely covered by anterior lappet of short light brown branched hairs and
opercular posterior lappet of brown branched hairs.
Wing (Fig. 13): Rs node setulose dorsally and ventrally, with setulae extending at most on basal sixth of R4+5
(these
setulae not always recognizable).
Legs: Legs of male paratype covered with fungi, for leg features see below under female description.
Terminalia (Figs 33–34): Cerci with posterior apex subtruncate in dorsal view (Fig. 33), and acuminate and
curved in lateral view (Fig. 34). Surstylus slightly arcuate inwards (Fig. 33), elongate and tapering at apex in dorsal
view (Fig. 33), with subparallel margins in lateral view (Fig. 34), and with basomedial lobes bearing fine setulae
hidden behind the cerci (Fig. 33). Pregonite narrow and long.
Female. Body length: 9.4–9.6 mm (n=4) (holotype 9.6 mm), wing length: 9.6–10.0 mm (n=4) (holotype 9.6
mm).
FIGURES 13–16. Phasioormia pallida. 10. Male dorsal habitus. 11. Female dorsal habitus. 12. Male lateral habitus. 13.
Female lateral habitus. (Male and female from Sri Lanka.) (Scale bars 1 mm.)
Zootaxa 3931 (1) © 2015 Magnolia Press · 15SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA
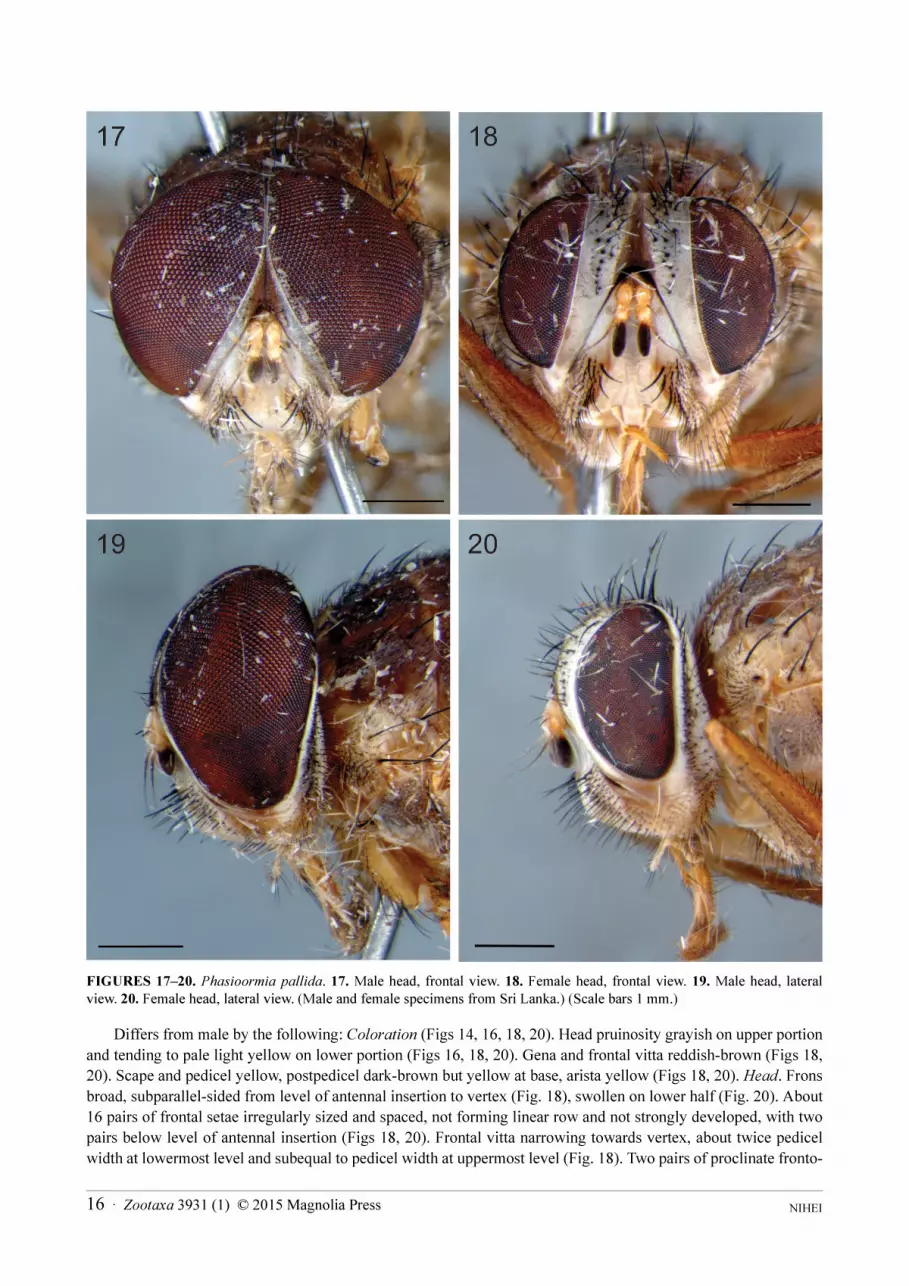
FIGURES 17–20. Phasioormia pallida. 17. Male head, frontal view. 18. Female head, frontal view. 19. Male head, lateral view. 20. Female head, lateral view. (Male and female specimens from Sri Lanka.) (Scale bars 1 mm.)
Differs from male by the following: Coloration (Figs 14, 16, 18, 20). Head pruinosity grayish on upper portion
and tending to pale light yellow on lower portion (Figs 16, 18, 20). Gena and frontal vitta reddish-brown (Figs 18,
20). Scape and pedicel yellow, postpedicel dark-brown but yellow at base, arista yellow (Figs 18, 20). Head. Frons
broad, subparallel-sided from level of antennal insertion to vertex (Fig. 18), swollen on lower half (Fig. 20). About
16 pairs of frontal setae irregularly sized and spaced, not forming linear row and not strongly developed, with two
pairs below level of antennal insertion (Figs 18, 20). Frontal vitta narrowing towards vertex, about twice pedicel
width at lowermost level and subequal to pedicel width at uppermost level (Fig. 18). Two pairs of proclinate fronto-
NIHEI 16 · Zootaxa 3931 (1) © 2015 Magnolia Press

orbital setae on upper half, and one reclinate pair before ocellar triangle (Figs 18, 20). Ocellar triangle hardly
recognizable as narrow slightly protuberant strip, with some weak setulae, and bearing no ocelli (Fig. 18). Fronto-
orbital plate broad, about 2.5x pedicel width at lowermost level (Fig. 18). Fronto-orbital plate with weak setulae on
its entire length (Figs 18, 20). Ratio of frons width/head width at vertex level 0.31; ratio of frons width/head width
at level of lunula 0.37 (Fig. 18). Parafacial bare and narrower than fronto-orbital plate, about 2.0x pedicel width
above and narrowing towards gena (Fig. 18). Facial ridge rather broadened, over 2x pedicel width at vibrissal level,
and with dense fine long setulae on lower portion (Figs 18, 20). Antennal axis/head height ratio 0.61 (Fig. 20).
Antennal axis/vibrissal axis ratio 1.08; antennal axis/oral (epistomal) axis ratio 1.13; vibrissal axis/oral axis ratio
1.05 (Fig. 20). In profile, anteroventral margin of head (bearing vibrissal, subvibrissal and subcranial setae/setulae)
subparallel to anteroventral margin of eye (Fig. 20). Antenna inserted clearly above mid height of head in frontal
view, and about mid height of eye (Fig. 20). Genal height/head height ratio 0.28. Genal dilation not apparent; gena
covered with short black setulae. Legs: Fore tibia with 2–4 differentiated anterodorsal setae; 2–4 differentiated
posterodorsal setae; one submedian posteroventral seta. Mid tibia with 2 anterodorsal setae on apical half, and 2
posterodorsal setae on apical half. Hind femur with dorsal row of setae, with longer setae at base; and anteroventral
and posteroventral rows of weak setae. Hind tibia with 2 anterodorsal setae on median third, 3 posterodorsal setae
on median third, and 1 anteroventral seta at apical third.
Terminalia (Fig. 35): Tergite 6 separated into hemitergites, with row of spiniform straight setae on median third
of inner margin and two isolated spiniform setae on apical third, and with single spiracle on each hemitergite.
Tergite 8 formed as a single narrow arc (fused medioventrally), and basally connected and partially fused with
sternite 10. Sternite 10 (postgenital plate or hypoproct) covered with fine setulae, bearing two spiniform lateral
setae and medial row of 4 spiniform setae decreasing inwards.
Type material examined. Holotype female (BMNH), “Holo- / type” (white round label with red border),
“Syngapore. / H. N. Ridley. / 1904-160”, “Phasioormia / pallida TT ♀ / Holotype”; specimen in good condition.
Paratype female (BMNH), “Para- / type” (white round label with yellow border), “Syngapore. / H. N. Ridley. /
1904-160”, “Phasioormia / pallida / Det. DA Barraclough, 1995”, “Phasioormia pallida 26.n”; specimen in
reasonably good condition, but the head detached from body and glued on a pinned paper triangle, and the right
foreleg and all left legs missing. Paratype male (BMNH), “Para- / type” (white round label with yellow border),
“Syngapore. / H. N. Ridley. / 98-114”; specimen in good condition, but left postpedicel and forelegs missing and
the ventral side with some fungi. Paratype female (BMNH), “Para- / type” (white round label with yellow border),
“Syngapore. / H. N. Ridley. / 1903-231”; specimen in good condition.
Additional material examined. SRI LANKA, Kan. Dist., Kandy, Udawattakele, 1 male, 1–3.x.1973, black
light, K.V. Krombein, P.B. Karunaratne & P. Fernando leg. (USNM); same locality, 2 females, 6–8.vi.1978, black
light, K.V. Krombein, P.B. Karunaratne, T. Wijesinhe, V. Kulasekare & L. Jayawickrema leg. (USNM); same
locality, 2 females, 26–30.vii.1978, black light, K.V. Krombein, P.B., T. Wijesinhe, V. Kulasekare & L.
Jayawickrema leg. (USNM); Kan. Dist., Peradeniya, upper Hantane Hill, 2300ft, 3 females, 12–16.i.1970, Davis &
Rowe leg. (USNM); Mon. Dist., Angunakolapelessa, 1 male, 17–19.vi.1978, Malaise trap, K.V. Krombein, P.B., T.
Wijesinhe, V. Kulasekare & L. Jayawickrema leg. (USNM). [One male from Udawattakele and one female from
Peradeniya were dissected and examined.]
Remarks. With previous records from the Philippines and Singapore, this species was newly recorded in the
Chinese provinces of Hainan and Jiangxi (O’Hara et al. 2009). I have not seen Chinese material to confirm its
identity, but so far those are the northernmost records for P. pallida. Here the species is also recorded in Sri Lanka.
Phasioormia bicornis (Malloch)
(Figs 21–24, 39)
bicornis Malloch, 1932: 313 (Ormia). Holotype male (BMNH). Type locality: Malaysia, Selangor, Bukit Kitu [Bukit Kutu].Phasioormia bicornis; Townsend, 1938: 237 (states that it should be assigned to a new genus); Crosskey, 1976: 64, 185
(combination with Phasioormia, cat.); O’Hara et al. 2009: 161 (cat., China).
Distribution (Fig. 39). China, India (Assam), and Malaysia.
Redescription. Male. Body length: 8.0 mm (n=1, holotype), wing length: not measured (both wings damaged
at tip).
Zootaxa 3931 (1) © 2015 Magnolia Press · 17SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

Coloration: Head dark-brown with silver pruinosity on face, parafacial, fronto-orbital plate, facial ridge, and
gena; frontal vitta black. Scape and pedicel yellow, postpedicel blackish, arista brown but light-brown on basal
fourth. Palpus yellowish, proboscis light brownish. Thorax reddish-brown but tending to yellowish-brown on
postpronotal lobe, notopleuron and lateral parts. Claws light brown with black apex; pulvilli yellow. Abdomen with
tergites 1+2 and basal half of 3 light-brown, apical half of tergite 3 and tergite 4 brown, and with whitish pruinosity
on tergites 4 and 5.
Head: Frons extremely narrow, on upper 1/3 eyes close to each other and only fronto-orbital plates visible as
narrow strips, but frons broadening ventrally. About 12 pairs of frontal setae on lower 2/3, shortening dorsally.
Ocellar triangle extremely constricted, but still recognizable, with a few minute setulae, no ocelli. Fronto-orbital
plate subequal to pedicel width at lowermost level in frontal view, without setulae. Ratio of frons width/head width
at vertex level 0.04; ratio of frons width/head width at level of lunula 0.26. Parafacial bare and as wide as fronto-
orbital plate. Antenna short; postpedicel about 1.3x pedicel length; arista elongate and slender, about 3.5x length of
postpedicel. Antenna inserted at mid height of head in frontal view, and slightly below mid height of eye. Antennal
axis/head height ratio 0.51. Antennal axis/vibrissal axis ratio 1.17; antennal axis/oral (epistomal) axis ratio 1.12;
vibrissal axis/oral axis ratio 0.96. Genal dilation developed and densely covered with short black setulae. Genal
height/head height ratio 0.15. Occiput strongly concave middorsally.
Thorax: Acrostichal setae 0+2, prescutellar pair stronger. Dorsocentral setae 3+3 (anteriormost presutural seta
weak). Notopleural setae 2, subequal and close to each other, with few short black setulae. Mesothoracic spiracle
conspicuously exposed, with light brownish branched hairs on whole anterior and posterior edges, anterior hairs
becoming longer towards upper second quarter, spiracle exposed dorsally and ventrally with just the middle
somewhat covered; metathoracic spiracle completely exposed, with very short light brown branched hairs on
anterior lappet, and without any hairs or lappet on posterior edge (this was observed on the male holotype and in
the female available, and until more material is examined, I am not certain whether this is common to the species or
is an artifact of these two specimens).
Wing: Rs node setulose dorsally and ventrally.
Legs: Fore tibia with one differentiated anterodorsal setae at apical third; 2 erect differentiated posterodorsal
setae on apical half. Mid femur with 1 basoventral seta. Mid tibia with 1 anterodorsal seta at apical third, and 2
posterodorsal setae on apical half. Hind femur with dorsal and anteroventral rows of setae, and posteroventral row
with weak setae. Hind tibia with 2–3 anterodorsal setae on apical half, 2–3 posterodorsal setae on apical half, and 2
anteroventral seta on apical half.
Terminalia: Not dissected, only the male holotype available.
Female. Body length: 8.6 mm (n=1), wing length: 8.0 mm (n=1).
Differs from male by the following: Coloration (Figs 21–24). Head pruinosity grayish on upper portion and
tending to pale light-yellow on lower portion (Figs 23–24). Gena and frontal vitta reddish-brown (Figs 23–24).
Scape and pedicel yellow, postpedicel entirely orangish, arista yellow (Figs 23–24). Head. Upper ommatidia not
enlarged. Frons broad, margins converging from level of antennal insertion towards vertex (Fig. 23), slightly
swollen on lower half (Fig. 24). About 16 pairs of frontal setae irregularly sized (Fig. 23), but forming linear row
and not strongly developed (the present and observable ones, and based on the insertions of broken setae), with two
pairs below level of antennal insertion (Fig. 24). Frontal vitta narrowing towards vertex, about twice pedicel width
at lowermost level and subequal to pedicel width at uppermost level (Fig 23). Two pairs of proclinate fronto-orbital
setae on upper half, and one reclinate pair before ocellar triangle. Ocellar triangle recognizable, broad and notably
protuberant, with dense weak setulae, and bearing no ocelli (Figs 23–24). Fronto-orbital plate broad, about 2.5x
pedicel width at lowermost level (Fig. 23). Fronto-orbital plate with weak setulae on its entire length. Ratio of frons
width/head width at vertex level 0.30; ratio of frons width/head width at level of lunula 0.37 (Fig. 23). Parafacial
bare and narrower than fronto-orbital plate, about 2.0x pedicel width above and narrowing towards gena (Fig. 23).
Facial ridge over 2x pedicel width at vibrissal level, with dense fine long setulae on lower portion (Fig. 23).
Antennal axis/head height ratio 0.53. Antennal axis/vibrissal axis ratio 1.12; antennal axis/oral (epistomal) axis
ratio 1.12; vibrissal axis/oral axis ratio 1.05 (Fig. 24). In profile, anteroventral margin of head (bearing vibrissal,
subvibrissal and subcranial setae/setulae) not subparallel to anteroventral margin of eye, closer to eye margin at
vibrissal level (anteriorly) and diverging ventrally toward gena (posteriorly), becoming conspicuously distant (Fig.
24). Antenna inserted clearly above mid height of head in frontal view, and slightly above mid height of eye (Fig.
24). Genal height/head height ratio 0.26 (Fig. 24). Genal dilation not apparent; gena covered with short black
setulae.
NIHEI 18 · Zootaxa 3931 (1) © 2015 Magnolia Press

Terminalia: Not dissected, only one female available.
Type material examined. Holotype male (BMNH), “Holo- / type” (white round label with red border),
“MALAY PENIN; / Selangor. / Bukit Kitu / at height 3500 ft. / 12-3-1931. / H. M. Pendlebury.”, on the underside
of latter label “EX:COLL: / F. M. S. / MUSEUMS.”, white label with underside “Pres. by / Fed. Malay States Mus.
/ Brit. Mus. 1934-363.”, “Ormia / bicornis / type / det. JRMALLOCH”. Specimen in good condition, except the
wing tips are damaged.
Additional material examined. INDIA, Assam, Gauhati [Guwahati], 1 female, 21.x.1920, B.M. 1924–100,
R. Senior-White leg. (BMNH).
Remarks. This species is extremely poorly represented in collections. If P. pallida is ‘well known’ from a few
dozen specimens, P. bicornis was previously known only from the male holotype. I have associated the female
from India (Assam) with the type from Malaysia, despite the geographical distance between them, and despite
some slight differences noticed between them (see above). I did not find any additional specimens in collections
that were visited or contacted, therefore, this is the only known female at present. Nevertheless, Chao & Zhou
(2003) extended the distribution of P. bicornis northwards to the Chinese coastal province of Fujian (see also
O’Hara et al. 2009). I have not seen this material.
FIGURES 21–24. Phasioormia bicornis, female. 21. Dorsal habitus. 22. Lateral habitus. 23. Head, frontal view. 24. Head, lateral view. (Female specimen from India.) (Scale bars 1 mm.)
Zootaxa 3931 (1) © 2015 Magnolia Press · 19SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

Phasioormia papuana sp. nov.
(Figs 25–32, 36–38, 39)
Type locality. Indonesia, Papua, Humboldt Bay District, Bewani Mts.
Distribution (Fig. 39). Indonesia and Papua New Guinea.
Diagnosis. P. papuana sp. nov. can be distinguished from P. bicornis by having the female ocellar triangle
hardly recognizable as a narrow strip and with weak, sparse setulae (ocellar triangle broad and protuberant, with
weak, dense setulae in P. bicornis), male ocellar triangle extremely constricted between eyes, not visible and bare
(extremely constricted between eyes, but still visible, with a few minute setulae in P. bicornis). The
abovementioned characters are shared with P. pallida, from which P. papuana sp. nov. can be differentiated by the
abdomen entirely yellowish-brown to brown (while P. pallida has abdomen reddish-yellow on tergites 1+2, 3 and
4, and tergite 5 dark brown), notopleuron bare or at most with sparse setulae (densely setulose in P. pallida), female
fronto-orbital plate narrower (2x the pedicel width at the lowermost level) than in P. pallida (2.5x the pedicel width
at the lowermost level), and female fronto-orbital plate with sparse setulae (dense setulose in P. pallida). There are
also distinguishing features in the male and female terminalia (Figs 33–38).
Description. Male. Body length: 8.1–9.2 mm (n=2) (holotype 9.2 mm), wing length: 8.4–9.1mm (n=2)
(holotype 9.1 mm).
FIGURES 25–28. Phasioormia papuana sp. nov. 25. Male dorsal habitus. 26. Female dorsal habitus. 27. Male lateral habitus. 28. Female lateral habitus. (Scale bars 1 mm.)
NIHEI 20 · Zootaxa 3931 (1) © 2015 Magnolia Press
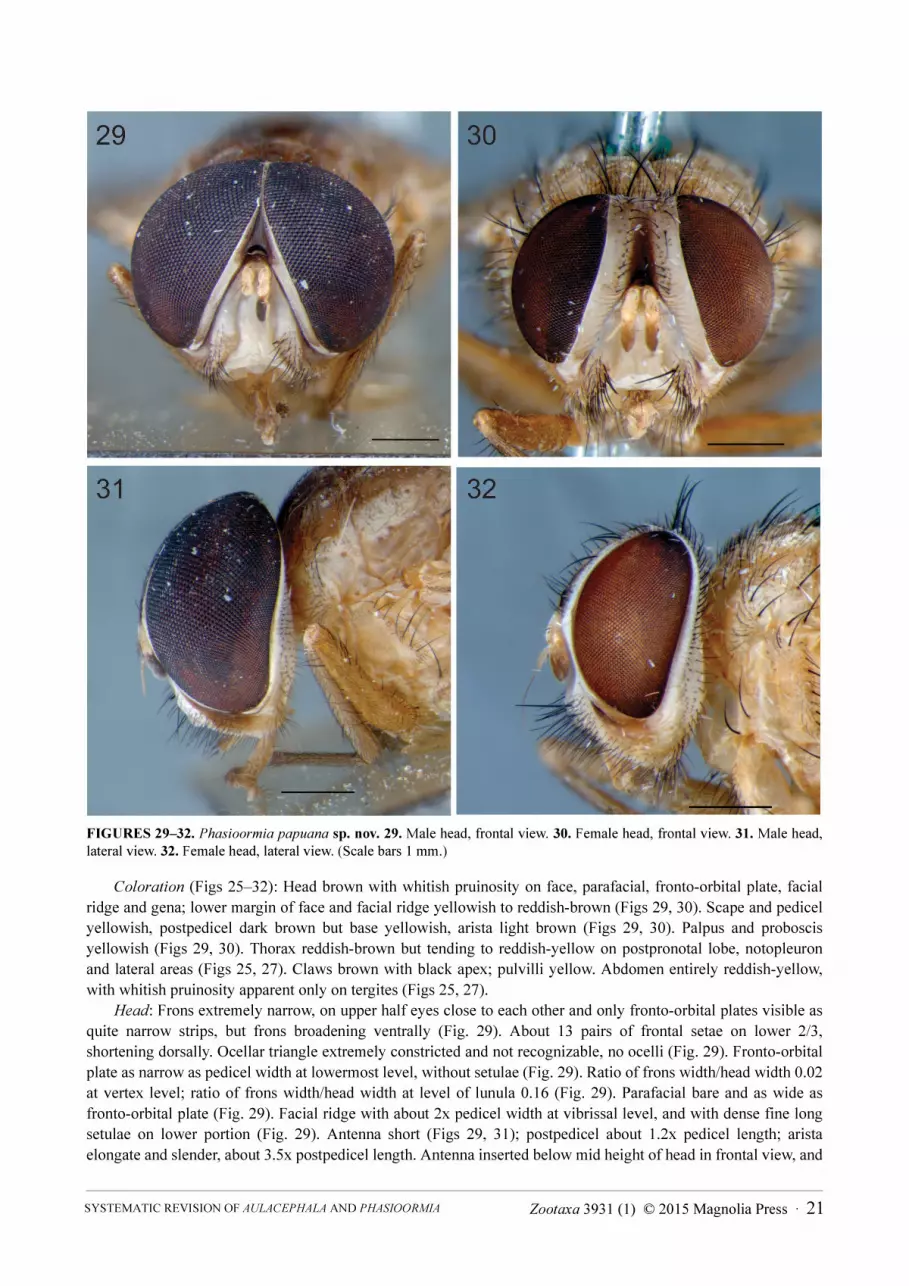
FIGURES 29–32. Phasioormia papuana sp. nov. 29. Male head, frontal view. 30. Female head, frontal view. 31. Male head, lateral view. 32. Female head, lateral view. (Scale bars 1 mm.)
Coloration (Figs 25–32): Head brown with whitish pruinosity on face, parafacial, fronto-orbital plate, facial
ridge and gena; lower margin of face and facial ridge yellowish to reddish-brown (Figs 29, 30). Scape and pedicel
yellowish, postpedicel dark brown but base yellowish, arista light brown (Figs 29, 30). Palpus and proboscis
yellowish (Figs 29, 30). Thorax reddish-brown but tending to reddish-yellow on postpronotal lobe, notopleuron
and lateral areas (Figs 25, 27). Claws brown with black apex; pulvilli yellow. Abdomen entirely reddish-yellow,
with whitish pruinosity apparent only on tergites (Figs 25, 27).
Head: Frons extremely narrow, on upper half eyes close to each other and only fronto-orbital plates visible as
quite narrow strips, but frons broadening ventrally (Fig. 29). About 13 pairs of frontal setae on lower 2/3,
shortening dorsally. Ocellar triangle extremely constricted and not recognizable, no ocelli (Fig. 29). Fronto-orbital
plate as narrow as pedicel width at lowermost level, without setulae (Fig. 29). Ratio of frons width/head width 0.02
at vertex level; ratio of frons width/head width at level of lunula 0.16 (Fig. 29). Parafacial bare and as wide as
fronto-orbital plate (Fig. 29). Facial ridge with about 2x pedicel width at vibrissal level, and with dense fine long
setulae on lower portion (Fig. 29). Antenna short (Figs 29, 31); postpedicel about 1.2x pedicel length; arista
elongate and slender, about 3.5x postpedicel length. Antenna inserted below mid height of head in frontal view, and
Zootaxa 3931 (1) © 2015 Magnolia Press · 21SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

below mid height of eye (Fig. 31). Antennal axis/head height ratio 0.63 (Fig. 31). Antennal axis/vibrissal axis ratio
1.28; antennal axis/oral (epistomal) axis ratio 1.36; vibrissal axis/oral axis ratio 1.14 (Fig. 31). Genal dilation not
apparent, but gena densely covered with short black setulae. Genal height/head height ratio 0.14. Occiput strongly
concave middorsally.
Thorax: Acrostichal setae 1+2, presutural pair anteriorly placed and not close to suture, prescutellar pair
stronger. Dorsocentral setae 2+3. Notopleural setae 2, subequal and close to each other, notopleuron bare or with
some sparse black setulae. Mesothoracic spiracle conspicuously exposed, with light brownish branched hairs on
whole anterior and posterior edges, anterior hairs becoming longer towards upper second quarter, spiracle exposed
dorsally and ventrally with just the middle somewhat covered; metathoracic spiracle not exposed, completely
covered by anterior lappet of short light brown branched hairs and opercular posterior lappet of brown branched
hairs.
Wing (Fig. 25): Rs node setulose dorsally and ventrally, with setulae extending at most on basal sixth of R4+5
.
Legs: One male paratype with the forelegs only, and a second male paratype lacking all legs; based on the
remnant legs, the male can be characterized as: fore tibia with 1 anterodorsal seta at apical third, 2 erect
posterodorsal setae on apical half; claws about 2/3 length of last tarsomere; and pulvilli slightly shorter than claws.
For other details, refer to the female, as other species of Phasioormia have not been observed to have sexually
dimorphic features in the legs.
Terminalia (Figs 36–37): Cerci with posterior apexes conspicuously separated and divergent (Fig. 36); apex
thumb-shaped in dorsal view (Fig. 36), and acuminate and curved in lateral view (Fig. 37). Cerci conspicuously
longer than wide (about twice). Surstylus slightly arcuate inwardly (Fig. 36), elongate, with broad and rounded
apex in both dorsal and lateral views (Figs 36 and 37), and with basomedial lobes bearing a few fine setulae hidden
behind the cerci (Fig. 36). Pregonite broad and long.
Female. Body length: 9.4–9.6 mm (n=4), wing length: 9.6–10.0 mm (n=4).
Differs from male by the following: Head (Figs 30, 32). Dichoptic. Upper ommatidia not enlarged. Frons
broad, subparallel sided from level of antennal insertion to vertex (Fig. 30), swollen on lower half (Fig. 32). About
16 pairs of frontal setae forming a linear row and not strongly developed (Fig. 30), with two pairs below level of
antennal insertion. Frontal vitta narrowing towards vertex, about twice pedicel width at lowermost level and
subequal to pedicel width at uppermost level (Fig. 30). Two pairs of proclinate fronto-orbital setae on upper half,
and one reclinate pair before ocellar triangle (Figs 30, 32). Ocellar triangle hardly recognizable as narrow slightly
protuberant strip, with some weak setulae, and bearing no ocelli (Fig. 30). Fronto-orbital plate broad, about 2.0x
pedicel width at lowermost level (Fig. 30). Fronto-orbital plate with weak setulae on its entire length (but not dense
as in P. pallida and P. bicornis). Ratio of frons width/head width at vertex level 0.25; ratio of frons width/head
width at level of lunula 0.34 (Fig. 30). Parafacial bare and narrower than fronto-orbital plate, about 2.0x pedicel
width above and narrowing towards gena (Fig. 30). Face broad and developed (not oestriform like in Aulacephala
and Therobia) (Fig. 30). Facial ridge rather broadened although not flattened (not oestriform like in Aulacephala
and Therobia), more than 2x pedicel width at vibrissal level, and with dense fine long setulae on lower portion (Fig.
30). Antennal axis/head height ratio 0.57 (Fig. 32). Antennal axis/vibrissal axis ratio 1.16; antennal axis/oral
(epistomal) axis ratio 1.28; vibrissal axis/oral axis ratio 1.10 (Fig. 32). In profile, anteroventral margin of head
(bearing vibrissal, subvibrissal and subcranial setae/setulae) subparallel to anteroventral margin of eye (Fig. 32).
Antenna inserted clearly above mid height of head in frontal view, and about mid height of eye (Fig. 32). Genal
height/head height ratio 0.25. Genal dilation not apparent; gena covered with short black setulae. Legs: Fore tibia
with 2–4 differentiated anterodorsal setae; 2 differentiated posterodorsal setae; one submedian posteroventral seta.
Mid tibia with 1–2 anterodorsal setae on apical half, and 1–2 posterodorsal setae on apical half. Hind femur with
dorsal row of setae, with longer setae at base, and anteroventral and posteroventral rows of weak setae. Hind tibia
with 2 anterodorsal setae on median third, 2–3 posterodorsal setae on median third, and 1–2 anteroventral seta on
apical third.
Terminalia (Fig. 38): Tergite 6 separated into hemitergites, with continuous row of spiniform straight setae on
inner margin from median third until half of apical third, and with single spiracle on each side. Tergite 8 formed as
a single narrow arc (fused medioventrally), but not fused with sternite 10. Sternite 10 (postgenital plate or
hypoproct) covered with fine setulae, bearing 2 spiniform lateral setae and medial row of 3 spiniform setae
decreasing inwards.
NIHEI 22 · Zootaxa 3931 (1) © 2015 Magnolia Press
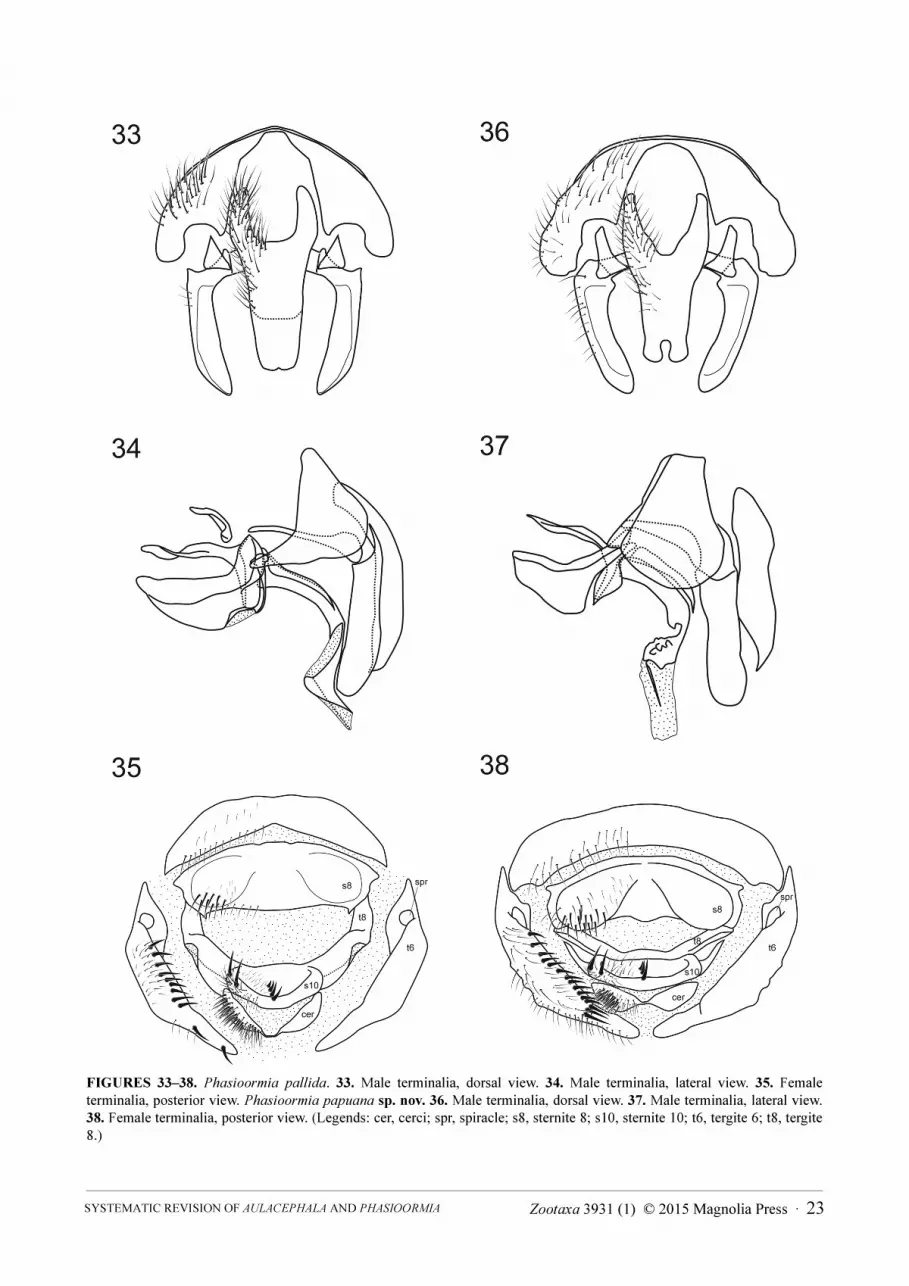
FIGURES 33–38. Phasioormia pallida. 33. Male terminalia, dorsal view. 34. Male terminalia, lateral view. 35. Female terminalia, posterior view. Phasioormia papuana sp. nov. 36. Male terminalia, dorsal view. 37. Male terminalia, lateral view. 38. Female terminalia, posterior view. (Legends: cer, cerci; spr, spiracle; s8, sternite 8; s10, sternite 10; t6, tergite 6; t8, tergite 8.)
Zootaxa 3931 (1) © 2015 Magnolia Press · 23SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

FIGURE 39. Geographical distribution of the genera Aulacephala (black circles: A. maculithorax; black squares: A. hervei) and Phasioormia (blue circles: P. bicornis; blue squares: P. pallida; blue stars: P. papuana sp. nov.). [Empty symbols denote inexact locality records, where the country or province capitals were arbitrarily used to plot the occurrence.]
Type material examined. Holotype male, DUTCH NEW GUINEA [INDONESIAN PAPUA], Humboldt Bay
Dist. [Yos Sudarso Bay], Bewani Mts., ix.1937, W. Stüber leg. (BMNH). Paratypes: same data as holotype, 5
females (BMNH); DUTCH NEW GUINEA [INDONESIAN PAPUA]: Cyclops Mts., Sabron, Camp 2, 1 female,
vii.1936 (2000ft), L.E. Cheesman leg. (BMNH); Fak-Fak, 1 female, A.E. Pratt leg. (C.J. Wainwright coll.), B.M.
1948-488 (BMNH); PAPUA [INDONESIAN PAPUA], Kokoda, 1200ft, 1 female, viii.1933, L.E. Cheesman leg.
(BMNH); [PAPUA] NEW GUINEA: Madang Dist., Damanti, Pinisterre Mts. (3550ft), 1 female, 2–11.x.1964,
M.E. Bacchus leg. (BMNH); NE Wau, 1200m, 1 male, 9.iii.1962 (light trap), J. & M. Sedlacek leg. (BMNH). [The
male from NE Wau and the female from Fak-Fak were dissected and examined.]
Acknowledgements
Thanks to Yoshitomi Hiroyuki (EUMJ), Hiroshi Shima (BLKU), Nigel Wyatt (BMNH), Mikhail Mostovski
(NMSA), Norman Woodley (USNM) and David Yeates (ANIC), for the loan of material. I am indebted to Erica
McAllister and Nigel Wyatt (BMNH) and to Norman Woodley and Torsten Dikow (USNM) for the facilities and
kind assistance during my visits. Thanks to Chun-tian Zhang (Shenyang Normal University) for kindly providing
useful information about Chinese Ormiini. Thanks to Rodrigo Dios for the digital drawings of male and female
terminalia of Aulacephala and Phasioormia; to Rodrigo Dios, Priscylla Moll, Filipe Gudin and Pedro Dias for
suggestions on an earlier version of this manuscript; and to Norman Woodley (USNM) and an anonymous reviewer
for valuable criticisms and suggestions that improved the submitted manuscript. Thanks to James O’Hara
(Canadian National Collection, Ottawa) for kindly providing photographs of the holotype of Aulacephala badia.
Financial support from FAPESP (proc. n. 2007/50836–7, and 2013/05131–6) and CNPq (proc. n. 562199/2010–1).
References
Barraclough, D.A. & Allen, G.R. (1996) Two new species of Homotrixa Villeneuve (Diptera: Tachinidae: Ormiini) from Southwestern Australia, with data on biology and ecology. Australian Journal of Entomology, 35, 135–145.
NIHEI 24 · Zootaxa 3931 (1) © 2015 Magnolia Press

http://dx.doi.org/10.1111/j.1440-6055.1996.tb01377.xBau, A. (1928) Ueber Aulacocephala und Rogenhofera (Diptera, olim Oestridae). Konowia, 7, 298–302.Bequaert, J. (1922) Sur le genre Aulacephala Macquart avec la description d’une espèce nouvelle de l’Extrème-Orient. Revue
Zoologique Africaine, 10, 301–308.Bequaert, J. (1929) A note on the genus Aulacephala Macquart (Diptera). Konowia, 8, 163–164.Bezzi, M. (1911) Miodarii superiori raccolti dal signor C. W. Howard nell’Africa austral orientale. Bollettino del Laboratorio di
Zoologia Generale e Agraria della Regia Scuola Superiore d'Agricoltura in Portici, 6, 45–104.Bigot, J.M.F. (1859) Diptères de Madagascar. 3e partie. Annales de la Société Entomologique de France, Série 3, 7, 533–558.Brauer, F. (1863) Monographie der Oestriden. C. Ueberreuter, Wien, 292 pp.Cantrell, B.K. & Crosskey, R.W. (1989) Family Tachinidae. In: Evenhuis, N. (Ed.), Catalog of the Diptera of the Australasian
and Oceanian Regions. Bishop Museum Press, Honolulu, pp. 733–784.Chao, C.-m. & Xue, W.-Q. (1998) Tachinidae. In: Xue, W.-Q. & Chao, C.-M. (Eds.), Flies of China. Vol. 2. Liaoning Science
and Technology Press, Shenyang, pp. 1661–2206 + pls. 1–30. [in Chinese]Chao, C.-M. & Zhou, S.-X. (2003) Tachinidae. In: Huang, B.-K. (Ed.), Fauna of insects in Fujian Province of China. Vol. 8.
Fujian Science and Technology Press, Fuzhou, pp. 443–498. [in Chinese]Crosskey, R.W. (1966) New generic and specific synonymy in Australian Tachinidae (Diptera). Proceedings of the Royal
Entomological Society of London (B), 35, 101–110.Crosskey, R.W. (1971) The type-material of Australasian, Oriental and Ethiopian Tachinidae (Diptera) described by Macquart
and Bigot. Bulletin of the British Museum (Natural History), Entomology, 25, 251–305.Crosskey, R.W. (1976) A taxonomic conspectus of the Tachinidae (Diptera) of the Oriental Region. Bulletin of the British
Museum (Natural History), Entomology Supplement, 26, 1–357.Crosskey, R.W. (1977) Family Tachinidae. In: Delfinado, M.D. & Hardy, D.E. (Eds.), A catalog of the Diptera of the Oriental
Region. Vol. III. Suborder Cyclorrhapha (excluding Aschiza). University Press of Hawaii, Honolulu, pp. 586–697.Crosskey, R.W. (1980) Family Tachinidae. In: Crosskey, R.W. (Ed.), Catalogue of the Diptera of the Afrotropical Region.
British Museum (Natural History), London, pp. 822–888.Crosskey, R.W. (1984) Annotated keys to the genera of Tachinidae (Diptera) found in tropical and southern Africa. Annals of
Natal Museum, 26, 189–337.Cumming, J.M. & Wood, D.M. (2009) Adult morphology and terminology. In: Brown, B.V., Borkent, A., Cumming, J.M.,
Wood, D.M., Woodley, N.E. & Zumbado, M.A. (Eds.), Manual of Central American Diptera. Vol. 1. NRC Research Press, Ottawa, pp. 9–50.
Dear, J.P. & Crosskey, R.W. (1982) A taxonomic review of the Tachinidae (Insecta, Diptera) of the Philippines. Steenstrupia, 8, 105–155.
Evenhuis, N.L. (2003) The status of the cricket parasites Ormia ochracea and Phasioormia pallida in the Hawaiian Islands (Diptera: Tachinidae). Bishop Museum Occasional Papers, 74, 34–35.
Gerstaecker, A. (1863) Eine neue Oestriden-Art, Aulacocephala badia. Verhandlungen der Kaiserlich-Königlichen Zoologisch-Botanischen Gesellschaft in Wien, 13, 1033–1035.
Guimarães, J.H. (1971) 104. Family Tachinidae. In: A catalogue of the Diptera of the Americas South of the United States. Museu de Zoologia, Universidade de São Paulo, São Paulo, pp. 1–333.
Herting, B. (1957) Das weibliche Postabdomen der calyptraten Fliegen (Diptera) und sein Merkmalswert für die Systematik der gruppe. Zeitschrift für Morphologie und Ökologie der Tiere, 45, 429–461. http://dx.doi.org/10.1007/BF00389872
Herting, B. & Dely-Draskovits, Á. (1993) Family Tachinidae. In: Soós, Á. & Papp, L. (Eds.), Catalogue of Palaearctic Diptera. Vol. 13. Anthomyiidae-Tachinidae. Hungarian Natural History Museum, Budapest, pp. 118–458.
Macquart, J. (1851) Diptères exotiques nouveaux ou peu connus. Suite du 4e supplement publié dans les memoires de 1849. Mémoires de la Société Royale des Sciences, de l’Agriculture et des Arts de Lille, 1850, 134–294. [Also published as “Diptères exotiques nouveaux ou peu connus”, Supplément IV [part], pp. 161–336. Paris, 1851.]
Malloch, J.R. (1925) A new species of the genus Aulacocephala (Dipt.) from Sumatra. Treubia, 6, 146–147.Malloch, J.R. (1932) Exotic Muscaridae (Diptera). – XXXVII. Annals and Magazine of Natural History, Series 10, 10,
297–330.Mesnil, L.P. (1973) 64g. Larvaevorinae (Tachininae). Die Fliegen der Palaearktischen Region, 10 (Lieferung 299), 1169–1232.Nishida, G.M. (1997) Hawaiian terrestrial arthropod checklist. 3rd edition. Bishop Museum Technical Reports, 12, 1–263.Nishida, G.M. (2002) Hawaiian terrestrial arthropod checklist. 4th edition. Bishop Museum Technical Reports, 22, 1–313.O’Hara, J.E. & Wood, D.M. (2004) Catalogue of the Tachinidae (Diptera) of America north of Mexico. Memoirs on
Entomology, International, 18, 1–410.O’Hara, J.E., Shima, H. & Zhang, C.-t. (2009) Annotated catalogue of the Tachinidae (Insecta: Diptera) of China. Zootaxa,
2190, 1–236.Paramonov, S.J. (1955) Notes on Australian Diptera. XVI. Ormiominda, gen. nov. – First representative of the tribe Ormiini
(Tachinidae) in Australia. Annals and Magazine of Natural History, Series 12, 8, 125–130.Preston, D.J. (1993) Notes and exhibitions: Phasioormia pallida Townsend. Proceedings of the Hawaiian Entomological
Society, 31 [1992], 16.Ricardo, G. (1901) Notes on Diptera from South Africa. Annals and Magazine of Natural History, Series 7, 8, 89–110.
Zootaxa 3931 (1) © 2015 Magnolia Press · 25SYSTEMATIC REVISION OF AULACEPHALA AND PHASIOORMIA

Séguy, M.E. (1926) Sur une forme nouvelle se rapportant aux “Oestridae dubiosae”. Diptera, Tome III (Fasc. 1), pp. 1–10. [Paris]
Shima, H. (2014) Family Tachinidae. In: Editorial Committee of the Insects of Japan (Eds.), Catalogue of the Insects of Japan. Vol. II. Part 2. Entomological Society of Japan, Tokyo, pp. 832–882.
Stuckenberg, B.R. (1999) Antennal evolution in the Brachycera (Diptera), with a reassessment of terminology relating to the flagellum. Studia dipterologica, 6, 33–48.
Townsend, C.H.T. (1918) New muscoid genera, species and synonymy (Diptera). Insecutor Inscitiae Menstruus, 6, 157–182.Townsend, C.H.T. (1933) New genera and species of Old World oestromuscoid flies. Journal of the New York Entomological
Society, 40 [1932], 439–479.Townsend, C.H.T. (1936) Manual of myiology, in twelve parts. Part III. Townsend & Filhos, Itaquaquecetuba, 250 pp.Townsend, C.H.T. (1938) Manual of myiology, in twelve parts. Part VII. Townsend & Filhos, Itaquaquecetuba, 246 pp.Uéda, S. (1960) Description of a new species of the genus Plesiooestrus Villeneuve, with notes on Aulacephala hervei
Bequaert (Diptera: Larvaevoridae). Insecta Matsumurana, 23, 14–20.Verbeke, J. (1963) The structure of the male genitalia in Tachinidae (Diptera) and their taxonomic value. Stuttgarter Beiträge
zur Naturkunde, 114, 1–8.Wood, D.M. & Zumbado, M.A. (2010) Tachinidae. In: Brown, B.V., Borkent, A., Cumming, J.M., Wood, D.M., Woodley, N.E.
& Zumbado, M.A. (Eds.), Manual of Central American Diptera. Vol. 2. NRC Research Press, Ottawa, pp. 1343–1417.
NIHEI 26 · Zootaxa 3931 (1) © 2015 Magnolia Press





























