Embed Size (px)
Citation preview

A
Ri
TNM
a
ARRA
KDMCI
C
fTf
c
0d
Molecular and Cellular Endocrinology 331 (2011) 1–10
Contents lists available at ScienceDirect
Molecular and Cellular Endocrinology
journa l homepage: www.e lsev ier .com/ locate /mce
t the Cutting Edge
ole of curcumin in the prevention of cholinergic mediated cortical dysfunctionsn streptozotocin-induced diabetic rats
. Peeyush Kumar, Sherin Antony, Smijin Soman, Korah P. Kuruvilla,aijil George, C.S. Paulose ∗
olecular Neurobiology and Cell Biology Unit, Centre for Neuroscience, Cochin University of Science and Technology, Cochin 682022, Kerala, India
r t i c l e i n f o
rticle history:eceived 23 January 2010eceived in revised form 3 June 2010ccepted 8 July 2010
a b s t r a c t
Diabetes exacerbates neuronal injury mediated through neurotransmitters deregulation in cerebral cor-tex. Our study analyzed the neuroprotective effect of curcumin to prevent cortical dysfunction associatedwith diabetes. Our study revealed decreased gene expression of muscarinic M1, insulin receptor, SOD,choline acetyl transferase and increased gene expression of muscarinic M3, �7-nicotinic acetylcholine
eywords:iabetesuscarinic
urcuminnsulin and cerebral cortex
receptor, acetylcholine esterase and GLUT3 in cerebral cortex of diabetic rats. Curcumin and insulin treat-ment reversed this altered parameters to near control. Immunohistochemistry studies of muscarinic M1and M3 confirmed the gene expression at protein level. Decreased novel arm entry of diabetic rats inY-maze test, improved in treatment group. These results suggest that cholinergic dysfunction, impairedglucose transport and oxidative stress contributes to learning and memory deficits in diabetes and fur-ther suggest that antioxidant curcumin has potential therapeutic role in preventing and/or delaying the
diabetic complications associated with brain.© 2010 Elsevier Ireland Ltd. All rights reserved.
ontents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22. Materials and methods. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
2.1. Estimation of blood glucose . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22.2. Y-maze test . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22.3. Total muscarinic, muscarinic M1 and M3 receptor binding studies in the cerebral cortex . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.4. Protein determination . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.5. Receptor data analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.6. Analysis of gene expression by real-time PCR . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32.7. Muscarinic M1 and M3 receptor expression studies in the cerebral cortex of control and experimental rats using confocal microscope 32.8. Statistics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
3. Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33.1. Y-maze performance of control and experimental groups of rats . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 33.2. Total muscarinic receptor analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
3.2.1. Scatchard analysis of [3H] QNB binding against atropine in the cerebral cortex of control and experimental rats . . . . . . . . . . . . . . 33.3. Muscarinic M1 receptor analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
3.3.1. Scatchard analysis of [3H] QNB binding against pirenzep3.4. Muscarinic M3 receptor analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
3.4.1. Scatchard analysis of [3H] DAMP binding against 4-DAM3.5. Real-time PCR analysis of muscarinic M1 receptor . . . . . . . . . . . . . . . .
∗ Corresponding author at: Molecular Neurobiology and Cell Biology Unit, Centreor Neuroscience, Department of Biotechnology, Cochin University of Science andechnology, Cochin 682022, Kerala, India. Tel.: +91 484 2576267/257588;ax: +91 484 2575588/2576699.
E-mail addresses: [email protected], [email protected],[email protected] (C.S. Paulose).
303-7207/$ – see front matter © 2010 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.mce.2010.07.004
ine in the cerebral cortex of control and experimental rats . . . . . . . . . . 4. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4P mustard in the cerebral cortex of control and experimental rats . . . 4. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4

2 T. Peeyush Kumar et al. / Molecular and Cellular Endocrinology 331 (2011) 1–10
3.6. Real-time PCR analysis of muscarinic M3 receptor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43.7. Real-time PCR analysis of �7-nicotinic acetylcholine receptor. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43.8. Real-time PCR analysis of acetylcholine esterase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 43.9. Real-time PCR analysis of choline acetyl transferase . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53.10. Real-time PCR analysis of insulin receptor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53.11. Real-time PCR analysis of GLUT3 receptor . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53.12. Real-time PCR analysis of super oxide dismutase (SOD) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53.13. Muscarinic M1 and M3 receptor antibody staining in the cerebral cortex of control and experimental groups of rats . . . . . . . . . . . . . . . . . . 6
. . . . .. . . . . .
1
tlitiwatim(rnism
id2ma(ac2gitfs
slemVaw2c(
cetddfoi
4. Discussion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Introduction
Prolonged exposure to chronic hyperglycaemia in diabetes leadso various complications, affecting the neurological, cardiovascu-ar, renal and visual systems (Brownlee, 2001). Previous studiesnvolving behavioural and electrophysiological analysis indicatehat diabetes mellitus induces cognitive impairment and defectsn long-term potentiation (Hasanein and Shahidi, 2010). People
ith diabetes apparently face a greater risk of Alzheimer’s diseasend depression (Biessels et al., 2006). Gold et al. (2007) reportedhat type 2 diabetes mellitus causes neuronal damage and memorympairments. Also, abnormalities in executive functions which are
ediated by the frontal lobe are noted in patients with diabetesQiu et al., 2006). Oxidative stress is believed to play an importantole in the development of complications in diabetes associatedeuronal disorders (Osawa and Kato, 2005). Greater understand-
ng of central nervous system (CNS) involvement could lead to newtrategies to prevent or reverse the damage caused by diabetesellitus.Antioxidant agents from diet have a significant therapeutic
nfluence on various neurodegenerative disorders associated withiabetes and oxidative stress (Ahmad et al., 2005; Ishrat et al.,006). Curcumin, a yellow pigment from Curcuma longa, is aajor component of turmeric and exhibits a powerful antioxidant,
n anti-diabetic, anti-inflammatory and anti-cancer propertiesCommandeur and Vermeulen, 1996; Surh et al., 2001; Miller etl., 2001). A number of experimental studies have demonstratedurcumin’s antioxidant and neuroprotective potential (Bala et al.,006; Kuhad and Chopra, 2007). Curcumin antagonise the deficit oflucose energy metabolism or oxidative stress related to cognitivempairment associated with diabetes. Also, curcumin modulateshe expression of various molecular targets such as transcriptionactors, enzymes, cytokines, cell cycle proteins, receptors and adhe-ion molecules (Shishodia et al., 2005).
Diabetes is also found to be associated with changes in somaticensations which involve the cerebral cortex, cerebellum and tha-amus. The cholinergic innervation of the cerebral cortex has beenxtensively investigated because of its role in arousal, learning andemory (Bartus et al., 1985; McCormick, 1989; Olton et al., 1991;oytko et al., 1994). Alterations in glucose transport and utilizationre known to occur in the important regions of brain connectedith learning and memory (Krebs and Parent, 2005; McNay et al.,
000). The brain glucose uptake is dependent on facilitative glu-ose transporters. GLUT3 is the main neuronal glucose transporterMaher et al., 1993) abundant in the brain.
Cognitive deficits and neurophysiological and structuralhanges are reported in the brain during diabetes mellitus (Brandst al., 2003; Mijnhout et al., 2006). However, the mechanisms ofhese changes remain obscure. Factors that contribute to cognitive
eficits as well as the protective factors that reduce the impact ofiabetes on brain functions are not completely elucidated. There-ore, this study was designed to investigate the beneficial effectf curcumin, a neuroprotective agent, in modulating cholinergic,nsulin receptors and GLUT3 in the cerebral cortex of streptozotocin. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 6. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 9
(STZ)-induced diabetic rats. Present study will certainly enlightennovel therapeutic possibilities for diabetes associated neurologicaldysfunctions.
2. Materials and methods
Bio chemicals used in the present study were purchased from Sigma ChemicalCo., St. Louis, USA. All other reagents of analytical grade were purchased locally.Quinuclidinyl benzilate, L-[benzilic-4,4′-3H], ([3H] QNB)(sp. activity 42 Ci/mmol)and 4-DAMP, [N-methyl-3H] (sp. activity 83 Ci/mmol) were purchased from NENLife Sciences Products Inc., Boston, USA. Streptozotocin, pirenzepine, 4-DAMP mus-tard, and curcumin from Sigma Chemical Co., USA. Tri-reagent kit was purchasedfrom MRC, USA. Real-time PCR Taqman probe assays on demand were from AppliedBiosystems, Foster City, CA, USA.
Male adult Wistar rats of 180–240 g body weight were used for all experiments.They were housed in separate cages under 12 h light and 12 h dark periods. Ratshad free access to standard food and water ad libitum. All animal care and pro-cedures were done in accordance with the Institutional and National Institute ofHealth guidelines. Diabetes was induced in rats by single intrafemoral vein injec-tion of STZ freshly dissolved in 0.1 M citrate buffer, pH 4.5, under anaesthesia(Junod et al., 1969). STZ was given at a dose of 55 mg/kg body weight (Hoheneggerand Rudas, 1971; Arison et al., 1967). Animals were divided into the followinggroups: (I) control, (ii) diabetic, (iii) insulin-treated diabetic and (iv) curcumin-treated diabetic rats. Each group consisted of 6–8 animals. The insulin-treateddiabetic group received subcutaneous injections (1 U/kg body weight) of Lenteand Plain insulin (Boots India) daily during the entire period of the experiment.The last injection was given 24 h before sacrificing the rats. Curcumin-treatedgroups received 60 mg/kg suspension of curcumin orally for the entire period ofthe experiment. Curcumin was suspended in 0.5% (w/v) sodium carboxymethyl-cellulose immediately before administration in constant volume of 5 ml/kg bodyweight (Sharma et al., 2006). Rats were sacrificed on 15th day by decapitation. Thecerebral cortex was dissected out quickly over ice according to the procedure ofGlowinski and Iversen (1966) and the tissues collected were stored at −80 ◦C untilassayed.
2.1. Estimation of blood glucose
Glucose was measured by GOD-POD glucose estimation kit (Biolab DiagnosticsPvt Ltd). Blood samples were collected from the tail vein at 0 h (before the start ofthe experiment), 3rd, 6th, 10th and 14th day and the glucose levels were estimatedsubsequently. Along with this blood samples were collected 3 h after the admin-istration of morning dose of insulin and curcumin. The results were expressed interms of milligram per decilitre of blood.
2.2. Y-maze test
For Y-maze test, the experimental rats were retained for five weeks and the ratswere subjected to training and test on day time to assess short-term and spatialmemory performance in Y-maze as per Nitta et al. (2002). The Y-maze was made ofgrey wood, covered with black paper, and consisted of three arms with an angle of120◦ between each of the arms. Each arm was 8 cm width × 30 cm length × 15 cmheight. The three identical arms were randomly designated: start arm, in which therat started to explore (always open); novel arm, which was blocked at the 1st trial,but open at the 2nd trial; and the other arm (always open). The maze was placed ina separate room with enough light. The floor of the maze was covered with sawdust,which was mixed after each individual trial in order to eliminate olfactory stimuli.Visual cues were placed on the walls of the maze.
The Y-maze test consisted of two trials separated by an inter-trial interval (ITI).The first trial (training) was of 10 min duration and allowed the rat to explore only
two arms (start arm and the other arm) of the maze, with the third arm (novelarm) blocked. After a 1 h ITI (Ma et al., 2007), the second trial (retention) wasconducted, during which all three arms were accessible and novelty vs. familiaritywas analyzed through comparing behaviour in all three arms. For the second trial,the rat was placed back in the maze in the same starting arm, with free access to allthree arms for 5 min. The time spent in each arm was analyzed. Data was expressed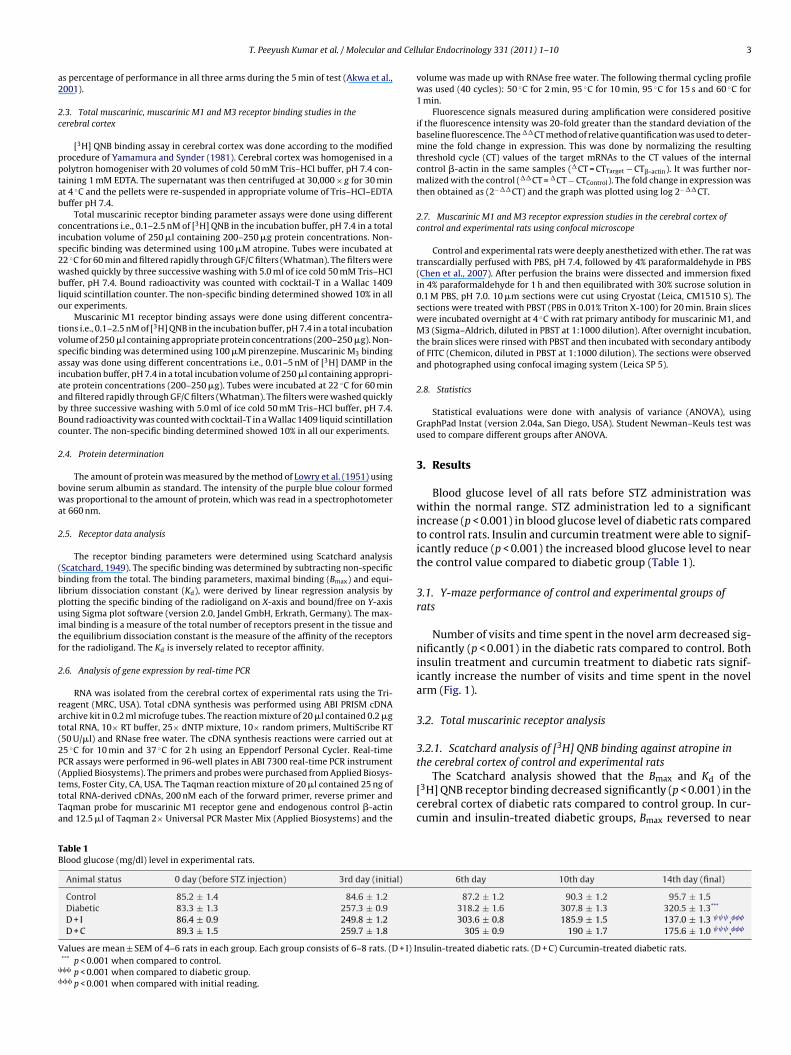
d Cell
a2
2c
pptab
cis2wblo
tvsaiaabBc
2
bwa
2
(blpuitf
2
rat(2P(ttTa
TB
V
�
�
T. Peeyush Kumar et al. / Molecular an
s percentage of performance in all three arms during the 5 min of test (Akwa et al.,001).
.3. Total muscarinic, muscarinic M1 and M3 receptor binding studies in theerebral cortex
[3H] QNB binding assay in cerebral cortex was done according to the modifiedrocedure of Yamamura and Synder (1981). Cerebral cortex was homogenised in aolytron homogeniser with 20 volumes of cold 50 mM Tris–HCl buffer, pH 7.4 con-aining 1 mM EDTA. The supernatant was then centrifuged at 30,000 × g for 30 mint 4 ◦C and the pellets were re-suspended in appropriate volume of Tris–HCl–EDTAuffer pH 7.4.
Total muscarinic receptor binding parameter assays were done using differentoncentrations i.e., 0.1–2.5 nM of [3H] QNB in the incubation buffer, pH 7.4 in a totalncubation volume of 250 �l containing 200–250 �g protein concentrations. Non-pecific binding was determined using 100 �M atropine. Tubes were incubated at2 ◦C for 60 min and filtered rapidly through GF/C filters (Whatman). The filters wereashed quickly by three successive washing with 5.0 ml of ice cold 50 mM Tris–HCl
uffer, pH 7.4. Bound radioactivity was counted with cocktail-T in a Wallac 1409iquid scintillation counter. The non-specific binding determined showed 10% in allur experiments.
Muscarinic M1 receptor binding assays were done using different concentra-ions i.e., 0.1–2.5 nM of [3H] QNB in the incubation buffer, pH 7.4 in a total incubationolume of 250 �l containing appropriate protein concentrations (200–250 �g). Non-pecific binding was determined using 100 �M pirenzepine. Muscarinic M3 bindingssay was done using different concentrations i.e., 0.01–5 nM of [3H] DAMP in thencubation buffer, pH 7.4 in a total incubation volume of 250 �l containing appropri-te protein concentrations (200–250 �g). Tubes were incubated at 22 ◦C for 60 minnd filtered rapidly through GF/C filters (Whatman). The filters were washed quicklyy three successive washing with 5.0 ml of ice cold 50 mM Tris–HCl buffer, pH 7.4.ound radioactivity was counted with cocktail-T in a Wallac 1409 liquid scintillationounter. The non-specific binding determined showed 10% in all our experiments.
.4. Protein determination
The amount of protein was measured by the method of Lowry et al. (1951) usingovine serum albumin as standard. The intensity of the purple blue colour formedas proportional to the amount of protein, which was read in a spectrophotometer
t 660 nm.
.5. Receptor data analysis
The receptor binding parameters were determined using Scatchard analysisScatchard, 1949). The specific binding was determined by subtracting non-specificinding from the total. The binding parameters, maximal binding (Bmax) and equi-
ibrium dissociation constant (Kd), were derived by linear regression analysis bylotting the specific binding of the radioligand on X-axis and bound/free on Y-axissing Sigma plot software (version 2.0, Jandel GmbH, Erkrath, Germany). The max-
mal binding is a measure of the total number of receptors present in the tissue andhe equilibrium dissociation constant is the measure of the affinity of the receptorsor the radioligand. The Kd is inversely related to receptor affinity.
.6. Analysis of gene expression by real-time PCR
RNA was isolated from the cerebral cortex of experimental rats using the Tri-eagent (MRC, USA). Total cDNA synthesis was performed using ABI PRISM cDNArchive kit in 0.2 ml microfuge tubes. The reaction mixture of 20 �l contained 0.2 �gotal RNA, 10× RT buffer, 25× dNTP mixture, 10× random primers, MultiScribe RT50 U/�l) and RNase free water. The cDNA synthesis reactions were carried out at5 ◦C for 10 min and 37 ◦C for 2 h using an Eppendorf Personal Cycler. Real-time
CR assays were performed in 96-well plates in ABI 7300 real-time PCR instrumentApplied Biosystems). The primers and probes were purchased from Applied Biosys-ems, Foster City, CA, USA. The Taqman reaction mixture of 20 �l contained 25 ng ofotal RNA-derived cDNAs, 200 nM each of the forward primer, reverse primer andaqman probe for muscarinic M1 receptor gene and endogenous control �-actinnd 12.5 �l of Taqman 2× Universal PCR Master Mix (Applied Biosystems) and theable 1lood glucose (mg/dl) level in experimental rats.
Animal status 0 day (before STZ injection) 3rd day (initial)
Control 85.2 ± 1.4 84.6 ± 1.2Diabetic 83.3 ± 1.3 257.3 ± 0.9D + I 86.4 ± 0.9 249.8 ± 1.2D + C 89.3 ± 1.5 259.7 ± 1.8
alues are mean ± SEM of 4–6 rats in each group. Each group consists of 6–8 rats. (D + I) I*** p < 0.001 when compared to control.�� p < 0.001 when compared to diabetic group.�� p < 0.001 when compared with initial reading.
ular Endocrinology 331 (2011) 1–10 3
volume was made up with RNAse free water. The following thermal cycling profilewas used (40 cycles): 50 ◦C for 2 min, 95 ◦C for 10 min, 95 ◦C for 15 s and 60 ◦C for1 min.
Fluorescence signals measured during amplification were considered positiveif the fluorescence intensity was 20-fold greater than the standard deviation of thebaseline fluorescence. The��CT method of relative quantification was used to deter-mine the fold change in expression. This was done by normalizing the resultingthreshold cycle (CT) values of the target mRNAs to the CT values of the internalcontrol �-actin in the same samples (�CT = CTTarget − CT�-actin). It was further nor-malized with the control (��CT =�CT − CTControl). The fold change in expression wasthen obtained as (2−��CT) and the graph was plotted using log 2−��CT.
2.7. Muscarinic M1 and M3 receptor expression studies in the cerebral cortex ofcontrol and experimental rats using confocal microscope
Control and experimental rats were deeply anesthetized with ether. The rat wastranscardially perfused with PBS, pH 7.4, followed by 4% paraformaldehyde in PBS(Chen et al., 2007). After perfusion the brains were dissected and immersion fixedin 4% paraformaldehyde for 1 h and then equilibrated with 30% sucrose solution in0.1 M PBS, pH 7.0. 10 �m sections were cut using Cryostat (Leica, CM1510 S). Thesections were treated with PBST (PBS in 0.01% Triton X-100) for 20 min. Brain sliceswere incubated overnight at 4 ◦C with rat primary antibody for muscarinic M1, andM3 (Sigma–Aldrich, diluted in PBST at 1:1000 dilution). After overnight incubation,the brain slices were rinsed with PBST and then incubated with secondary antibodyof FITC (Chemicon, diluted in PBST at 1:1000 dilution). The sections were observedand photographed using confocal imaging system (Leica SP 5).
2.8. Statistics
Statistical evaluations were done with analysis of variance (ANOVA), usingGraphPad Instat (version 2.04a, San Diego, USA). Student Newman–Keuls test wasused to compare different groups after ANOVA.
3. Results
Blood glucose level of all rats before STZ administration waswithin the normal range. STZ administration led to a significantincrease (p < 0.001) in blood glucose level of diabetic rats comparedto control rats. Insulin and curcumin treatment were able to signif-icantly reduce (p < 0.001) the increased blood glucose level to nearthe control value compared to diabetic group (Table 1).
3.1. Y-maze performance of control and experimental groups ofrats
Number of visits and time spent in the novel arm decreased sig-nificantly (p < 0.001) in the diabetic rats compared to control. Bothinsulin treatment and curcumin treatment to diabetic rats signif-icantly increase the number of visits and time spent in the novelarm (Fig. 1).
3.2. Total muscarinic receptor analysis
3.2.1. Scatchard analysis of [3H] QNB binding against atropine in
the cerebral cortex of control and experimental ratsThe Scatchard analysis showed that the Bmax and Kd of the[3H] QNB receptor binding decreased significantly (p < 0.001) in thecerebral cortex of diabetic rats compared to control group. In cur-cumin and insulin-treated diabetic groups, Bmax reversed to near
6th day 10th day 14th day (final)
87.2 ± 1.2 90.3 ± 1.2 95.7 ± 1.5318.2 ± 1.6 307.8 ± 1.3 320.5 ± 1.3***
303.6 ± 0.8 185.9 ± 1.5 137.0 ± 1.3 ,���
305 ± 0.9 190 ± 1.7 175.6 ± 1.0 ,���
nsulin-treated diabetic rats. (D + C) Curcumin-treated diabetic rats.

4 T. Peeyush Kumar et al. / Molecular and Cellular Endocrinology 331 (2011) 1–10
Table 2Scatchard analysis of [3H] QNB binding against atropine in the cerebral cortex ofcontrol, and experimental rats.
Animal status Bmax (fmoles/mg protein) Kd (nM)
Control 316 ± 8.5 0.21 ± 0.02Diabetic 131 ± 6.2*** 0.17 ± 0.02*
D + I 300 ± 8.2 ��� 0.3 ± 0.01 �
D + C 308.6 ± 8.6 ��� 0.32 ± 0.03�
Values are mean ± SEM of 4–6 separate experiments. Each group consists of 6–8rats. (D + I) Insulin-treated diabetic rats. (D + C) Curcumin-treated diabetic rats.
* p < 0.05 when compared to control group.*** p < 0.001 when compared to control.
� p < 0.05 when compared to diabetic group.��� p < 0.001 when compared to diabetic group.
Table 3Scatchard analysis of [3H] QNB binding against pirenzepine in the cerebral cortex ofcontrol, and experimental rats.
Animal status Bmax (fmoles/mg protein) Kd (nM)
Control 180 ± 12.4 1.6 ± 0.2Diabetic 65 ± 13.2*** 0.58 ± 0.02***
D + I 225 ± 8.6��� 1.8 ± 0.01���
D + C 205 ± 9.2��� 1.78 ± 0.03���
Vr
�
ct
3
3i
c(an
3
3m
c(ct
Fcm(r
Table 4Scatchard analysis of [3H] DAMP binding against 4-DAMP mustard in the cerebralcortex of control, and experimental rats.
Animal status Bmax (fmoles/mg protein) Kd (nM)
Control 56 ± 1.4 0.20 ± 0.02Diabetic 202 ± 2.2*** 0.49 ± 0.02***
D + I 52 ± 0.5��� 0.25 ± 0.01@@
D + C 75 ± 0.4��� 0.20 ± 0.03���
Values are mean ± SEM of 4–6 separate experiments. Each group consist of 6–8 rats.(D + I) Insulin-treated diabetic rats. (D + C) Curcumin-treated diabetic rats.
Real-time PCR analysis showed that the acetylcholine esterasegene expression in the cerebral cortex was increased significantly(p < 0.001) in diabetic condition and it reversed to near control valuein insulin and curcumin-treated diabetic rats (Fig. 5).
alues are mean ± SEM of 4–6 separate experiments. Each group consists of 6–8ats. (D + I) Insulin-treated diabetic rats. (D + C) Curcumin-treated diabetic rats.*** p < 0.001 when compared to control.�� p < 0.001 when compared to diabetic group.
ontrol value. Kd of insulin-treated and curcumin group reversedo near control (Table 2).
.3. Muscarinic M1 receptor analysis
.3.1. Scatchard analysis of [3H] QNB binding against pirenzepinen the cerebral cortex of control and experimental rats
The Scatchard analysis showed that the Bmax and Kd of mus-arinic M1 receptors of cerebral cortex were decreased significantlyp < 0.001) in diabetic condition compared to control group. Insulinnd curcumin-treated diabetic rats Bmax and Kd were reversed toear control value compared to diabetic group (Table 3).
.4. Muscarinic M3 receptor analysis
.4.1. Scatchard analysis of [3H] DAMP binding against 4-DAMPustard in the cerebral cortex of control and experimental rats
The Scatchard analysis showed that the B and K of mus-
max darinic M3 receptors of cerebral cortex were increased significantlyp < 0.001) in diabetic rats compared to control group. Insulin andurcumin-treated diabetic rats showed Bmax and Kd were reversedo near control value compared to diabetic group (Table 4).ig. 1. Y-maze performance of control, diabetic, insulin-treated diabetic andurcumin-treated diabetic rats. Values are mean ± SEM of 4–6 separate experi-ents (n = 5–6 rats per group) ANOVA followed by Students–Newman–Keuls’ test.
a) p < 0.01 when compared to control and (b) p < 0.01 when compared to diabeticats.
*** p < 0.001 when compared to control.��� p < 0.001 when compared to diabetic group.@@ p < 0.01 when compared to diabetic group.
3.5. Real-time PCR analysis of muscarinic M1 receptor
Real-time PCR analysis showed that the muscarinic M1 receptorgene expression in the cerebral cortex was decreased significantly(p < 0.001) in diabetic condition and it reversed to near control valuein insulin and curcumin-treated diabetic rats (Fig. 2).
3.6. Real-time PCR analysis of muscarinic M3 receptor
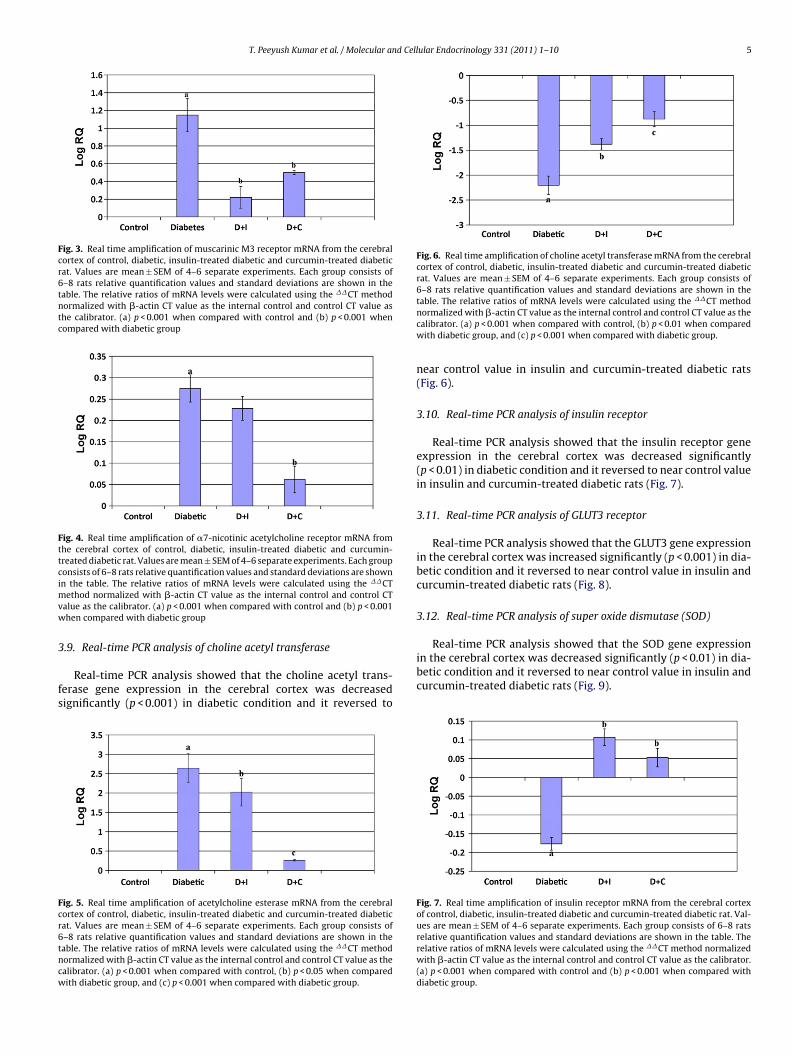
Real-time PCR analysis showed that the muscarinic M3 receptorgene expression in the cerebral cortex was increased significantly(p < 0.001) in diabetic condition and it reversed to near control valuein insulin and curcumin-treated diabetic rats (Fig. 3).
3.7. Real-time PCR analysis of ˛7-nicotinic acetylcholine receptor
Real-time PCR analysis showed that �7-nicotinic acetylcholinereceptor gene expression in the cerebral cortex was increased sig-nificantly (p < 0.001) in diabetic condition and it reversed to nearcontrol value in curcumin-treated diabetic rats (Fig. 6). But insulintreatment did not show any significant change in �7-nicotinicacetylcholine receptor gene expression in the cerebral cortex whencompared to diabetes (Fig. 4).
3.8. Real-time PCR analysis of acetylcholine esterase
Fig. 2. Real time amplification of muscarinic M1 receptor mRNA from the cerebralcortex of control, diabetic, insulin-treated diabetic and curcumin-treated diabeticrat. Values are mean ± SEM of 4–6 separate experiments. Each group consists of6–8 rats relative quantification values and standard deviations are shown in thetable. The relative ratios of mRNA levels were calculated using the ��CT methodnormalized with �-actin CT value as the internal control and control CT value asthe calibrator. (a) p < 0.001 when compared with control and (b) p < 0.001 whencompared with diabetic group
T. Peeyush Kumar et al. / Molecular and Cellular Endocrinology 331 (2011) 1–10 5
Fig. 3. Real time amplification of muscarinic M3 receptor mRNA from the cerebralcortex of control, diabetic, insulin-treated diabetic and curcumin-treated diabeticrat. Values are mean ± SEM of 4–6 separate experiments. Each group consists of6–8 rats relative quantification values and standard deviations are shown in thetable. The relative ratios of mRNA levels were calculated using the ��CT methodnormalized with �-actin CT value as the internal control and control CT value asthe calibrator. (a) p < 0.001 when compared with control and (b) p < 0.001 whencompared with diabetic group
Fig. 4. Real time amplification of �7-nicotinic acetylcholine receptor mRNA fromthe cerebral cortex of control, diabetic, insulin-treated diabetic and curcumin-treated diabetic rat. Values are mean ± SEM of 4–6 separate experiments. Each groupconsists of 6–8 rats relative quantification values and standard deviations are shownimvw
3
fs
Fcr6tncw
Fig. 6. Real time amplification of choline acetyl transferase mRNA from the cerebralcortex of control, diabetic, insulin-treated diabetic and curcumin-treated diabeticrat. Values are mean ± SEM of 4–6 separate experiments. Each group consists of6–8 rats relative quantification values and standard deviations are shown in the
��
Real-time PCR analysis showed that the SOD gene expression
n the table. The relative ratios of mRNA levels were calculated using the ��CTethod normalized with �-actin CT value as the internal control and control CT
alue as the calibrator. (a) p < 0.001 when compared with control and (b) p < 0.001hen compared with diabetic group
.9. Real-time PCR analysis of choline acetyl transferase
Real-time PCR analysis showed that the choline acetyl trans-erase gene expression in the cerebral cortex was decreasedignificantly (p < 0.001) in diabetic condition and it reversed to
ig. 5. Real time amplification of acetylcholine esterase mRNA from the cerebralortex of control, diabetic, insulin-treated diabetic and curcumin-treated diabeticat. Values are mean ± SEM of 4–6 separate experiments. Each group consists of–8 rats relative quantification values and standard deviations are shown in theable. The relative ratios of mRNA levels were calculated using the ��CT methodormalized with �-actin CT value as the internal control and control CT value as thealibrator. (a) p < 0.001 when compared with control, (b) p < 0.05 when comparedith diabetic group, and (c) p < 0.001 when compared with diabetic group.
table. The relative ratios of mRNA levels were calculated using the CT methodnormalized with �-actin CT value as the internal control and control CT value as thecalibrator. (a) p < 0.001 when compared with control, (b) p < 0.01 when comparedwith diabetic group, and (c) p < 0.001 when compared with diabetic group.
near control value in insulin and curcumin-treated diabetic rats(Fig. 6).
3.10. Real-time PCR analysis of insulin receptor
Real-time PCR analysis showed that the insulin receptor geneexpression in the cerebral cortex was decreased significantly(p < 0.01) in diabetic condition and it reversed to near control valuein insulin and curcumin-treated diabetic rats (Fig. 7).
3.11. Real-time PCR analysis of GLUT3 receptor
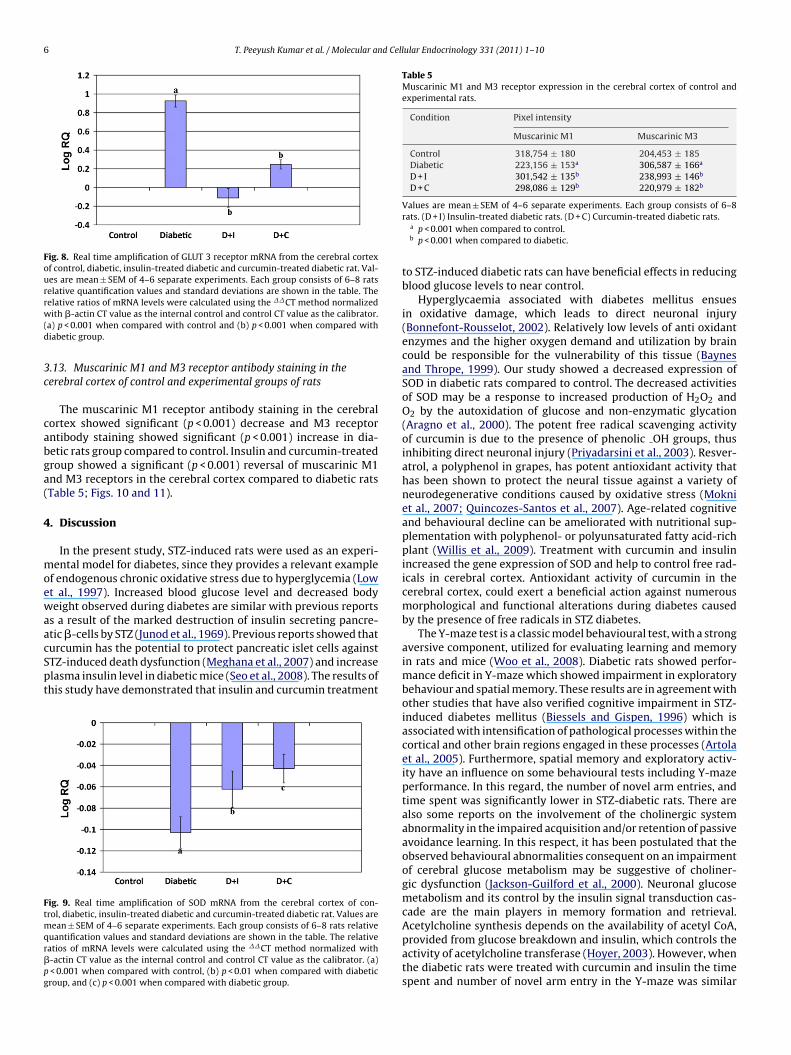
Real-time PCR analysis showed that the GLUT3 gene expressionin the cerebral cortex was increased significantly (p < 0.001) in dia-betic condition and it reversed to near control value in insulin andcurcumin-treated diabetic rats (Fig. 8).
3.12. Real-time PCR analysis of super oxide dismutase (SOD)
in the cerebral cortex was decreased significantly (p < 0.01) in dia-betic condition and it reversed to near control value in insulin andcurcumin-treated diabetic rats (Fig. 9).
Fig. 7. Real time amplification of insulin receptor mRNA from the cerebral cortexof control, diabetic, insulin-treated diabetic and curcumin-treated diabetic rat. Val-ues are mean ± SEM of 4–6 separate experiments. Each group consists of 6–8 ratsrelative quantification values and standard deviations are shown in the table. Therelative ratios of mRNA levels were calculated using the ��CT method normalizedwith �-actin CT value as the internal control and control CT value as the calibrator.(a) p < 0.001 when compared with control and (b) p < 0.001 when compared withdiabetic group.
6 T. Peeyush Kumar et al. / Molecular and Cellular Endocrinology 331 (2011) 1–10
Fig. 8. Real time amplification of GLUT 3 receptor mRNA from the cerebral cortexof control, diabetic, insulin-treated diabetic and curcumin-treated diabetic rat. Val-ues are mean ± SEM of 4–6 separate experiments. Each group consists of 6–8 ratsrelative quantification values and standard deviations are shown in the table. Therw(d
3c
cabga(
4
moewaacSpt
Ftmqr�pg
Table 5Muscarinic M1 and M3 receptor expression in the cerebral cortex of control andexperimental rats.
Condition Pixel intensity
Muscarinic M1 Muscarinic M3
Control 318,754 ± 180 204,453 ± 185Diabetic 223,156 ± 153a 306,587 ± 166a
D + I 301,542 ± 135b 238,993 ± 146b
D + C 298,086 ± 129b 220,979 ± 182b
elative ratios of mRNA levels were calculated using the ��CT method normalizedith �-actin CT value as the internal control and control CT value as the calibrator.
a) p < 0.001 when compared with control and (b) p < 0.001 when compared withiabetic group.
.13. Muscarinic M1 and M3 receptor antibody staining in theerebral cortex of control and experimental groups of rats
The muscarinic M1 receptor antibody staining in the cerebralortex showed significant (p < 0.001) decrease and M3 receptorntibody staining showed significant (p < 0.001) increase in dia-etic rats group compared to control. Insulin and curcumin-treatedroup showed a significant (p < 0.001) reversal of muscarinic M1nd M3 receptors in the cerebral cortex compared to diabetic ratsTable 5; Figs. 10 and 11).
. Discussion
In the present study, STZ-induced rats were used as an experi-ental model for diabetes, since they provides a relevant example
f endogenous chronic oxidative stress due to hyperglycemia (Lowt al., 1997). Increased blood glucose level and decreased bodyeight observed during diabetes are similar with previous reports
s a result of the marked destruction of insulin secreting pancre-
tic �-cells by STZ (Junod et al., 1969). Previous reports showed thaturcumin has the potential to protect pancreatic islet cells againstTZ-induced death dysfunction (Meghana et al., 2007) and increaselasma insulin level in diabetic mice (Seo et al., 2008). The results ofhis study have demonstrated that insulin and curcumin treatmentig. 9. Real time amplification of SOD mRNA from the cerebral cortex of con-rol, diabetic, insulin-treated diabetic and curcumin-treated diabetic rat. Values are
ean ± SEM of 4–6 separate experiments. Each group consists of 6–8 rats relativeuantification values and standard deviations are shown in the table. The relativeatios of mRNA levels were calculated using the ��CT method normalized with-actin CT value as the internal control and control CT value as the calibrator. (a)< 0.001 when compared with control, (b) p < 0.01 when compared with diabeticroup, and (c) p < 0.001 when compared with diabetic group.
Values are mean ± SEM of 4–6 separate experiments. Each group consists of 6–8rats. (D + I) Insulin-treated diabetic rats. (D + C) Curcumin-treated diabetic rats.
a p < 0.001 when compared to control.b p < 0.001 when compared to diabetic.
to STZ-induced diabetic rats can have beneficial effects in reducingblood glucose levels to near control.
Hyperglycaemia associated with diabetes mellitus ensuesin oxidative damage, which leads to direct neuronal injury(Bonnefont-Rousselot, 2002). Relatively low levels of anti oxidantenzymes and the higher oxygen demand and utilization by braincould be responsible for the vulnerability of this tissue (Baynesand Thrope, 1999). Our study showed a decreased expression ofSOD in diabetic rats compared to control. The decreased activitiesof SOD may be a response to increased production of H2O2 andO2 by the autoxidation of glucose and non-enzymatic glycation(Aragno et al., 2000). The potent free radical scavenging activityof curcumin is due to the presence of phenolic OH groups, thusinhibiting direct neuronal injury (Priyadarsini et al., 2003). Resver-atrol, a polyphenol in grapes, has potent antioxidant activity thathas been shown to protect the neural tissue against a variety ofneurodegenerative conditions caused by oxidative stress (Mokniet al., 2007; Quincozes-Santos et al., 2007). Age-related cognitiveand behavioural decline can be ameliorated with nutritional sup-plementation with polyphenol- or polyunsaturated fatty acid-richplant (Willis et al., 2009). Treatment with curcumin and insulinincreased the gene expression of SOD and help to control free rad-icals in cerebral cortex. Antioxidant activity of curcumin in thecerebral cortex, could exert a beneficial action against numerousmorphological and functional alterations during diabetes causedby the presence of free radicals in STZ diabetes.
The Y-maze test is a classic model behavioural test, with a strongaversive component, utilized for evaluating learning and memoryin rats and mice (Woo et al., 2008). Diabetic rats showed perfor-mance deficit in Y-maze which showed impairment in exploratorybehaviour and spatial memory. These results are in agreement withother studies that have also verified cognitive impairment in STZ-induced diabetes mellitus (Biessels and Gispen, 1996) which isassociated with intensification of pathological processes within thecortical and other brain regions engaged in these processes (Artolaet al., 2005). Furthermore, spatial memory and exploratory activ-ity have an influence on some behavioural tests including Y-mazeperformance. In this regard, the number of novel arm entries, andtime spent was significantly lower in STZ-diabetic rats. There arealso some reports on the involvement of the cholinergic systemabnormality in the impaired acquisition and/or retention of passiveavoidance learning. In this respect, it has been postulated that theobserved behavioural abnormalities consequent on an impairmentof cerebral glucose metabolism may be suggestive of choliner-gic dysfunction (Jackson-Guilford et al., 2000). Neuronal glucosemetabolism and its control by the insulin signal transduction cas-cade are the main players in memory formation and retrieval.
Acetylcholine synthesis depends on the availability of acetyl CoA,provided from glucose breakdown and insulin, which controls theactivity of acetylcholine transferase (Hoyer, 2003). However, whenthe diabetic rats were treated with curcumin and insulin the timespent and number of novel arm entry in the Y-maze was similar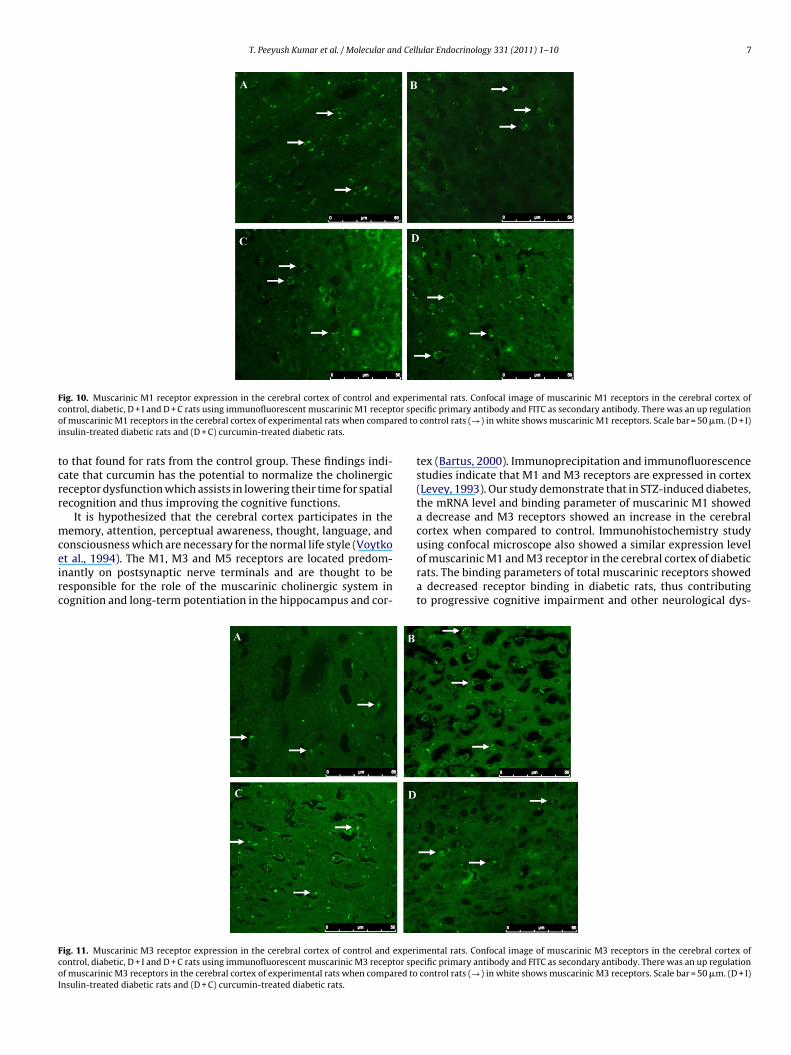
T. Peeyush Kumar et al. / Molecular and Cellular Endocrinology 331 (2011) 1–10 7
F experic or speo red toi
tcrr
mceirc
FcoI
ig. 10. Muscarinic M1 receptor expression in the cerebral cortex of control andontrol, diabetic, D + I and D + C rats using immunofluorescent muscarinic M1 receptf muscarinic M1 receptors in the cerebral cortex of experimental rats when compansulin-treated diabetic rats and (D + C) curcumin-treated diabetic rats.
o that found for rats from the control group. These findings indi-ate that curcumin has the potential to normalize the cholinergiceceptor dysfunction which assists in lowering their time for spatialecognition and thus improving the cognitive functions.
It is hypothesized that the cerebral cortex participates in theemory, attention, perceptual awareness, thought, language, and
onsciousness which are necessary for the normal life style (Voytkot al., 1994). The M1, M3 and M5 receptors are located predom-nantly on postsynaptic nerve terminals and are thought to beesponsible for the role of the muscarinic cholinergic system inognition and long-term potentiation in the hippocampus and cor-
ig. 11. Muscarinic M3 receptor expression in the cerebral cortex of control and experiontrol, diabetic, D + I and D + C rats using immunofluorescent muscarinic M3 receptor spef muscarinic M3 receptors in the cerebral cortex of experimental rats when compared tonsulin-treated diabetic rats and (D + C) curcumin-treated diabetic rats.
mental rats. Confocal image of muscarinic M1 receptors in the cerebral cortex ofcific primary antibody and FITC as secondary antibody. There was an up regulationcontrol rats (→) in white shows muscarinic M1 receptors. Scale bar = 50 �m. (D + I)
tex (Bartus, 2000). Immunoprecipitation and immunofluorescencestudies indicate that M1 and M3 receptors are expressed in cortex(Levey, 1993). Our study demonstrate that in STZ-induced diabetes,the mRNA level and binding parameter of muscarinic M1 showeda decrease and M3 receptors showed an increase in the cerebralcortex when compared to control. Immunohistochemistry study
using confocal microscope also showed a similar expression levelof muscarinic M1 and M3 receptor in the cerebral cortex of diabeticrats. The binding parameters of total muscarinic receptors showeda decreased receptor binding in diabetic rats, thus contributingto progressive cognitive impairment and other neurological dys-mental rats. Confocal image of muscarinic M3 receptors in the cerebral cortex ofcific primary antibody and FITC as secondary antibody. There was an up regulationcontrol rats (→) in white shows muscarinic M3 receptors. Scale bar = 50 �m. (D + I)

8 d Cell
fsg(mtmchrd
oapco(1mohiStcibraa1rnSlrri
echSaamHfsu1tmglndemYweii
T. Peeyush Kumar et al. / Molecular an
unctions associated with cortex seen in diabetes. Earlier reportshowed significant alterations in neurotransmitters during hyper-lycaemia and causes degenerative changes in neurons of the CNSGarris, 1990; Bhardwaj et al., 1999). Curcumin and insulin treat-
ent was able to significantly reverse these altered parameterso near the control value. Previous reports show that activation of
uscarinic M1 cholinergic receptors produced an increase in glu-ose utilization (Hosey, 1992). Thus, we speculated that curcuminas an ability to modulate muscarinic receptors thereby amelio-ating the impaired cognitive performance shown in STZ-inducediabetes.
�7-Nicotinic receptor is highly expressed during brain devel-pment (Court et al., 1995, 1997; Hellstrom-Lindahl et al., 1999)nd has been implicated in memory, attention and informationrocessing (Picciotto et al., 2000). It is well established that mus-arinic receptors as well as nicotinic receptors are present mainlyn pyramidal neurons in the human and rodent cerebral cortexSchroder et al., 1989; Van der Zee et al., 1992; Nakayama et al.,995; Wevers et al., 1999). However, the potential of curcumin inodulating the mRNA of �7-nicotinic receptor and involvement
f the �7-nicotinic receptor in the prevention of cognitive deficitsave not been addressed. The current data reveal a significant
ncrease in the expression of �7-nicotinic receptor in the cortex ofTZ-induced diabetic rats. These receptors significantly influencehe activity within the cortex circuitry and diabetes mellitus asso-iated deregulation of this activity could contribute to disordersnvolving the cerebral cortex. �7-nicotinic receptor agonists haveeen shown to improve performance on a variety of cognitive testselated to working, short-term, and long-term memory function innimal models (Bitner et al., 2007; Boess et al., 2006; Hashimoto etl., 2008). As nicotine increases cerebral blood flow (Uchida et al.,997), glucose utilization (McNamara et al., 1990), acetylcholineelease (Armitage and Hall, 1969) in the brain; alterations in �7-icotinic receptor expression may be related to cognitive deficits inTZ-Induced diabetes. Curcumin treatment reverted this up regu-ation of �7-nicotinic receptor in the cortex of STZ-induced diabeticats. In line with this, we suggest that regulation of cholinergiceceptors function by curcumin can contribute consequently tomprove the cognitive functions, such as learning and memory.
In the present work, we observed an increased acetylcholinesterase and decreased choline acetyl transferase mRNA level inerebral cortex of diabetic rats. It has been demonstrated that AChEas a fundamental role in learning and memory (Das et al., 2002;ato et al., 2004) and alterations in its activity as well as in thecetylcholine neurotransmitter level are neurochemically associ-ted with cognitive deficits observed in patients and in animalodels of diabetes mellitus (Kuhad and Chopra, 2007; Ghareeb andussen, 2008). Increased AChE and decreased choline acetyl trans-
erase mRNA level indicates fast acetylcholine degradation and aubsequent down stimulation of acetylcholine receptors causingndesirable effects on cognitive functions (Tõugu and Kesvatera,996; Soreq and Seidman, 2001). Based on our results we suggesthat the increased AChE and decreased choline acetyl transferase
RNA level caused by diabetes leads to a reduction of choliner-ic neurotransmission efficiency due to a decrease in acetylcholineevels in the synaptic cleft, thus contributing to progressive cog-itive impairment and other neurological dysfunctions seen iniabetic patients. Furthermore, we may infer that the activatorffect elicited by diabetic state on AChE level could be one of theechanisms involved on the memory impairment observed in the
-maze performance in this study. In the present study, treatment
ith curcumin and insulin substantially regularize the increasedxpression of acetylcholine esterase and choline acetyl transferasen diabetic to a control level and consequently in cognitive deficitsnduced by the diabetic state.
ular Endocrinology 331 (2011) 1–10
Insulin receptor in peripheral tissues participates mainly inglucose metabolism; however its role in the CNS appears not tobe related to glucose metabolism but to other neuronal activi-ties such as memory (Zhao et al., 1999). Recently, much evidencehas been presented regarding the role of brain insulin or insulinreceptors in memory formation (Frolich et al., 1998). Experimentshave shown the ability of small doses of insulin (0.4–0.8 U/kg) toreverse the amnesia produced by a 2 mg/kg scopolamine injec-tion (Blanchard and Duncan, 1997; Messier and Destrade, 1994).In the higher limbic system including the hippocampus, piryformcortex and amygdala, the insulin receptor has been demonstratedto play an important role in spatial and emotional memory viaeffects on a variety of signaling pathways (Zhao and Alkon, 2001).Our study reports that STZ-induced diabetes exhibits a significantdown regulation of insulin receptor mRNA in the cerebral cortex.Cognitive impairments associated with diabetes mellitus causedby inadequate insulin/insulin receptor functions have also beendocumented. Treatment with curcumin and insulin normalize theimpaired expression of insulin receptors in diabetic state to nearcontrol. Uptake of insulin into the CNS appears to occur via anactive transendothelial transport across the blood–brain barrier(Schwartz et al., 1990; Banks et al., 1997). This transport processappears to be specific for insulin (Banks and Kastin, 1998) and issaturable at blood insulin levels that are found during euglycemia(Banks et al., 1997). In the present study, we present evidence thatcurcumin have stimulatory actions on memory formation with theregulatory role in insulin receptors of cerebral cortex.
Glucose transport into the brain is critical for the maintenanceof brain metabolism. Although under basal conditions the rateof glucose transport is not the rate-limiting step for glycolysis inthe CNS (Gruefter et al., 1992), hypoglycaemia or hyperglycaemiais known to change the glucose transport system in the brain(Pramming et al., 1986; Devivo et al., 1991), suggesting that thereshould be glucose-regulatable mechanisms associated with thetransport of glucose. Our study investigated the effect of diabeteson cortical glucose transport mediated through GLUT3 and the pro-tective effect of curcumin in modulating GLUT3. Result showed theincreased mRNA content of GLUT3 in the cerebral cortex of STZ-induced diabetes, which showed impaired glucose transport. Also,curcumin treatment improved the glucose transport in cerebralcortex of diabetes rats by controlling the increased GLUT3 expres-sion. Alterations in glucose utilization are known to occur in theimportant regions of brain connected with learning and memory(Krebs and Parent, 2005; McNay et al., 2000). Learning and mem-ory processing is found to produce increases of glucose metabolismin the cortical brain regions that is functionally related to mem-ory processing as well as to the sensorimotor task requirements(Friedman and Goldman-Rakic, 1994; Balabanova et al., 1984).
Diabetes associated complications result in morbidity andmortality (Daneman, 2001). Elucidation of the risk factors andpathophysiological mechanisms underlying diabetic complicationsis important. Deterioration in glucose homeostasis that resultsfrom diabetes induced hyperglycaemia trigger neuronal injuries(Bonnefont-Rousselot, 2002); the molecular basis of this neuronalvulnerability is not completely elucidated. In addition, curcuminhas recently received considerable attention since it has beenshown to protect neurons against a variety of experimental neu-rodegenerative conditions (Cole et al., 2007). The results of thisstudy have demonstrated that the supplementation of curcuminto STZ-induced diabetic rats has beneficial effects in reducing thealterations in cholinergic, insulin receptors, oxidative stress and
imbalanced glucose transport through GLUT3 in the cerebral cortex.Our findings provide confirmatory evidence for the neuroprotec-tive role of curcumin and pave way for the better management ofdiabetes mediated neurological complications
d Cell
R
A
A
A
A
A
A
B
B
B
B
B
B
B
B
B
B
B
B
B
B
B
B
B
C
C
C
C
T. Peeyush Kumar et al. / Molecular an
eferences
hmad, M., Saleem, S., Ahmad, A.S., Yousuf, S., Ansari, M.A., Khan, M.B., Ishrat,T., Chaturvedi, R.K., Agrawal, A.K., Islam, F., 2005. Ginkgo biloba affords dose-dependent protection against 6-hydroxydopamine-induced parkinsonism inrats: neurobehavioural, neurochemical and immunohistochemical evidences.J. Neurochem. 93, 94–104.
kwa, Y., Ladurelle, N., Covey, D.F., Baulieu, E.E., 2001. The synthetic enantiomerof pregnenolone sulfate is very active on memory in rats and mice, even moreso than its physiological neurosteroid counterpart: distinct mechanisms. Proc.Natl. Acad. Sci. U.S.A. 98, 14033–14037.
ragno, M., Parola, S., Tamagno, E., Brignardello, E., Manti, R., Danni, O., Boccuzzi, G.,2000. Oxidative derangement in rat synaptosomes induced by hyperglycemia:restorative effect of dehydroepiandrosterone treatment. Biochem. Pharmacol.60, 389–395.
rison, R.N., Ciaccio, E.I., Glitzer, M.S., Cassaro, A.V., Pruss, M., 1967. Light andelectron microscopy of lesions in rats rendered diabetic with streptozotocin.Diabetes 16, 51–56.
rmitage, A.K., Hall, G.H., 1969. Sellers CM effects of nicotine on electrocortical activ-ity and acetylcholine release from the cat cerebral cortex. Br. J. Pharmacol. 35,152–160.
rtola, A., Kamal, A., Ramakers, G.M., Biessels, G.J., Gispen, W.H., 2005. Diabetesmellitus concomitantly facilitates the induction of long-term depression andinhibits that of long-term potentiation in hippocampus. Eur. J. Neurosci. 22,169–178.
ala, K., Tripathy, B.C., Sharma, D., 2006. Neuroprotective and antiageing effects ofcurcumin in aged rat brain regions. Biogerontology 7, 81–89.
alabanova, S., Richter, H.P., Antoniadis, G., Homoki, J., Kremmer, N., Hanle,J., Teller, W.M., 1984. 25-Hydroxyvitamin D, 24,25-dihydroxyvitamin D and1,25-dihydroxyvitamin D in human cerebrospinal fluid. Klin. Wochenschr. 62,1086–1090.
anks, W.A., Jaspan, J.B., Huang, W., Kastin, A.J., 1997a. Transport of insulin acrossthe blood–brain barrier: saturability at euglycemic doses of insulin. Peptides 18,1423–1429.
anks, W.A., Jaspan, J.B., Kastin, A.J., 1997b. Selective, physiological transport ofinsulin across the blood–brain barrier: novel demonstration by species-specificradioimmunoassay. Peptides 28, 1257–1262.
anks, W.A., Kastin, A.J., 1998. Differential permeability of the blood–brain barrierto two pancreatic peptides: insulin and amylin. Peptides 19, 883–889.
artus, R.T., 2000. On neurodegenerative diseases, models, and treatment strategies:lessons learned and lessons forgotten a generation following the cholinergichypothesis. Exp. Neurol. 163, 495–529.
artus, R.T., Dean, R.L., Pontecorvo, M.J., Flicker, C., 1985. The cholinergic hypothesis:a historical overview, current perspective, and future directions. Ann. N.Y. Acad.Sci. 444, 332–358.
aynes, J.W., Thrope, S.R., 1999. Role of oxidative stress in diabetic complications.Diabetes 48, 1–9.
hardwaj, S.K., Sandhu, S.K., Sharma, P., Kaur, G., 1999. Impact of diabetes on CNS.Role of signal transduction cascade. Brain Res. Bull. 49, 155–162.
iessels, G., Gispen, W.H., 1996. The calcium hypothesis of brain aging and neu-rodegenerative disorders: significance in diabetic neuropathy. Life. Sci. 59 (5–6),379–387.
iessels, G.J., Staekenborg, S., Brunner, E., Brayne, C., Scheltens, P., 2006. Risk ofdementia in diabetes mellitus: a systematic review. Lancet Neurol. 5, 64–74.
itner, R.S., Bunnelle, W.H., Anderson, D.J., Briggs, C.A., Buccafusco, J., Curzon, P.,Decker, M.W., Frost, J.M., Gronlien, J.H., Gubbins, E., Li, J., Malysz, J., Markosyan,S., Marsh, K., Meyer, M.D., Nikkel, A.L., Radek, R.J., Robb, H.M., Timmermann,D., Sullivan, J.P., Gopalakrishnan, M., 2007. Broad-spectrum efficacy across cog-nitive domains by alpha7 nicotinic acetylcholine receptor agonism correlateswith activation of ERK1/2 and CREB phosphorylation pathways. J. Neurosci. 27,10578–10587.
lanchard, J.G., Duncan, P.M., 1997. Effect of combinations of insulin, glucose andscopolamine on radial arm maze performance. Pharmacol. Biochem. Behav. 58,209–214.
oess, F.G., De Vry, J., Erb, C., Flessner, T., Hendrix, M., Luithle, J., Methfessel, C., 2006.Brain insulin receptors and spatial memory. J. Biol. Chem. 274, 34893–34902.
onnefont-Rousselot, D., 2002. Glucose and reactive oxygen species. Curr. Opin. Clin.Nutr. Metab. Care 5, 561–568.
rands, A.M., Henselmans, J.M., de Haan, E.H., Biessels, G.J., 2003. Diabeticencephalopathy: an underexposed complication of diabetes mellitus. Ned. Tijd-schr. Geneeskd. 147, 11–14.
rownlee, M., 2001. Biochemistry and molecular cell biology of diabetic complica-tions. Nature 414, 813–820.
hen, S., Kobayashi, M., Honda, Y., Kakuta, S., Sato, F., Kishi, K., 2007. Preferen-tial neuron loss in the rat piriform cortex following pilocarpine-induced statusepilepticus. Epilepsy Res. 74, 1–18.
ole, G.M., Bruce, T., Sally, A.F., 2007. Neuroprotective effects of curcumin. Adv. Exp.Med. Biol. 595, 197–212.
ommandeur, J.N.M., Vermeulen, N.P.E., 1996. Cytotoxicity and cytoprotective activ-ities of natural compounds the case of curcumin. Xenobiotica 26 (7), 667–
680.ourt, J.A., Lloyd, S., Johnson, M., Griffiths, M., Birdsall, N.J., Piggott, M.A., Oakley,A.E., Ince, P.G., Perry, E.K., Perry, R.H., 1997. Nicotinic and muscarinic cholinergicreceptor binding in the human hippocampal formation during development andaging. Brain. Res. Dev. Brain. Res. 101 (1–2), 93–105.
ular Endocrinology 331 (2011) 1–10 9
Court, J.A., Perry, E.K., Spurden, D., Griffiths, M., Kerwin, J.M., Morris, C.M., et al., 1995.The role of the cholinergic system in the development of the human cerebellum.Brain Res. Dev. Brain Res. 90, 159–167.
Daneman, Denis, 2001. Diabetes-related mortality. Diabetes Care 24, 5.Das, A., Shanker, G., Nath, C., Pal, R., Singh, S., Singh, H.K., 2002. A comparative study
rodents of standardized extracts of Bacopa monniera and ginkgo biloba anti-cholinesterase and cognitive enhancing activities. Pharmacol. Biochem. Behav.73, 893–900.
Devivo, D.C., Trifiletti, R.R., Jacobson, R.I., Rosen, G.M., Behmand, R.A., Harik, S.I.,1991. Defective glucose transport across the blood-brain barrier as a cause ofpersistent hypoglycorrhachia, seizures, and developmental delay. N. Engl. J. Med.325, 703–709.
Friedman, H.R., Goldman-Rakic, P.S., 1994. Coactivation of prefrontal cortex andinferior parietal cortex in working memory tasks revealed by 2DG functionalmapping in the rhesus monkey. J. Neurosci. 14, 2775–2788.
Frolich, L., Blum-degen, D., Bernstein, H.G., Engelsberger, S., Humrich, J., Laufer,S., Muschner, D., Thalheimer, A., Turk, A., Hoyer, S., Zochling, R., Boissl, K.W.,Jellinger, K., Piederer, P., 1998. Brain insulin and insulin receptors in aging andsporadic Alzheimer’s disease. J. Neural Transm. 105, 423–438.
Garris, 1990. Age diabetes associated alterations in regional brain norepinephrineconcentrations and adrenergic populations in C57BL/KsL mice. Dev. Brain Res.51, 161–166.
Ghareeb, D.A., Hussen, H.M., 2008. Vanadium improves brain acetylcholinesteraseactivity on early stage alloxan-diabetic rats. Neurosci. Lett. 436, 44–47.
Glowinski, J., Iversen, L.L., 1966. Regional studies of catecholamines in the rat brain,the disposition of [3H] norepinephrine, [3H]dopa in various regions of brain. J.Neurochem. 13, 655–669.
Gold, S.M., Dziobek, I., Sweat, V., Tirsi, A., Rogers, K., Bruehl, H., Tsui, W., Richardson,S., Javier, E., Convit, A., 2007. Hippocampal damage and memory impairmentsas possible early brain complications of type 2 diabetes. Diabetologia 50 (4),711–719.
Gruefter, R., Novotny, E.J., Boulware, S.D., Rothman, D.L., Mason, G.G., Shulman,G.I., Shulman, R.G., Tamborlane, W.V., 1992. Proc. Natl. Acad. Sci. U.S.A. 89,1109–1112.
Hasanein, P., Shahidi, S., 2010. Effects of combined treatment with vitamins C andE on passive avoidance learning and memory in diabetic rats. Neurobiol. LearnMem. 93, 472–478.
Hashimoto, K., Ishima, T., Fujita, Y., Matsuo, M., Kobashi, T., Takahagi, M., Tsukada,H., Iyo, M., 2008. Phencyclidine-induced cognitive deficits in mice are improvedby subsequent subchronic administration of the novel selective alpha7 nicotinicreceptor agonist SSR180711. Biol. Psychiatry 63, 92–97.
Hellstrom-Lindahl, E., Mousavi, M., Zhang, X., Ravid, R., Nordberg, A., 1999. Regionaldistribution of nicotinic receptor subunit mRNAs in human brain: comparisonbetween Alzheimer and normal brain. Brain Res. Mol. Brain Res. 66, 94–103.
Hohenegger, M., Rudas, B., 1971. Kidney functions in experimental diabetic ketosis.Diabetologia 17, 334–338.
Hosey, M.M., 1992. Diversity of structure, signaling and regulation within the familyof muscarinic cholinergic receptors. FASEB J. 6, 845–852.
Hoyer, S., 2003. Memory function and brain glucose. Metab. Pharmacopsychiat. 36(Suppl. 1), S62–S67.
Ishrat, T., Khan, M.B., Hoda, M.N., Yousuf, S., Ahmad, M., Ansari, M.A., Ahmad, A.S.,Islam, F., 2006. Coenzyme Q10 modulates cognitive impairment against intrac-erebroventricular injection of streptozotocin in rats. Behav. Brain Res. 171, 9–16.
Jackson-Guilford, J., Leander, J.D., Nisenbaum, L.K., 2000. The effect of streptozotocin-induced diabetes on cell proliferation in the rat dentate gyrus. Neurosci. Lett.293, 91–94.
Junod, A., Lambert, A.E., Staufferacher, W., Renold, A.E., 1969. Diabetogenic actionof streptozotocin relationship of dose to metabolic response. J. Clin. Invest. 48,2129–2139.
Krebs, D.L., Parent, M.B., 2005. The enhancing effects of hippocampal infusions ofglucose are not restricted to spatial working memory. Neurobiol. Learn Mem.83, 168–172.
Kuhad, A., Chopra, K., 2007. Curcumin attenuates diabetic encephalopathy in rats:behavioral and biochemical evidences. Eur. J. Pharmacol. 576, 34–42.
Levey, A.I., 1993. Immunological localization of M1-M5 muscarinic acetylcholinereceptors in peripheral tissues and brain. J. Life Sci. 52, 441–448.
Low, P.A., Nickander, K.K., Tritschler, H.J., 1997. The role of oxidative stress andantioxidant treatment in experimental diabetic neuropathy. Diabetes 46, 38.
Lowry, O.H., Roserbrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith Folin Phenol reagent. J. Biol. Chem. 193, 265–275.
Ma, J., Yu, L.J., Ma, R.D., Zhang, Y.P., Fang, J.Z., Zhang, X.Y., Yu, T.X., 2007. Repair ofglutamate-induced excitotoxic neuronal damage mediated by intracerebroven-tricular transplantation of neural stem cells in adult mice. Neurosci. Bull. 23 (4),209–214.
Maher, F., Vannucci, S.J., Simpson, I.A., 1993. Glucose transporter isoforms in brain:absence of GLUT3 from the blood–brain barrier. J. Cereb. Blood Flow Metab. 13,342–345.
McCormick, D.A., 1989. Cholinergic and noradrenergic modulation of thalamocorti-cal processing. Trends Neurosci. 12, 215–221.
McNamara, D., Larson, D.M., Rapoport, S.I., Soncrant, T.T., 1990. Preferential
metabolic activation of subcortical brain areas by acute administration of nico-tine to rats. J. Cereb. Blood Flow Metab. 10, 48–56.McNay, E.C., Fries, T.M., Gold, P.E., 2000. Decreases in rat extracellular hippocampalglucose concentration associated with cognitive demand during a spatial task.Proc. Natl. Acad. Sci. 97, 2881–2885.

1 d Cell
M
M
M
M
M
N
N
O
O
P
P
P
Q
Q
S
S
S
0 T. Peeyush Kumar et al. / Molecular an
eghana, K., Sanjeev, G., Ramesh, B., 2007. Curcumin prevents streptozotocin-induced islet damage by scavenging free radicals: a prophylactic and protectiverole. Eur. J. Pharmacol. 577 (1–3), 183–191.
essier, C., Destrade, C., 1994. Insulin attenuates scopolamine-induced memorydeficits. Psychobiology 22, 16–21.
ijnhout, G.S., Scheltens, P., Diamant, M., Biessels, G.J., Wessels, A.M., Simsek, S.,Snoek, F.J., Heine, R.J., 2006. Diabetic encephalopathy: a concept in need of adefinition. Diabetologia 49, 1447–1448.
iller, J.S.M., Angeles, F.M., Reuter, B.K., Bobrowski, P., Sandoval, M., 2001. Dietaryantioxidants protect gut epithelial cells from oxidant-induced apoptosis. BMCComplement. Altern. Med. 1, 11.
okni, M., Elkahoui, S., Limam, F., Amri, M., Aouani, E., 2007. Effect of resveratrol onantioxidant enzyme activities in the brain of healthy rat. Neurochem. Res. 32,981–987.
akayama, H., Shioda, S., Okuda, H., Nakashima, T., Nakai, Y., 1995. Immunocyto-chemical localization of nicotinic acetylcholine receptor in rat cerebral cortex.Mol. Brain. Res. 32, 321–328.
itta, A., Murai, R., Suzuki, N., Ito, H., Nomoto, H., Katoh, G., Furukawa, Y., Furukawa,S., 2002. Diabetic neuropathies in brain are induced by deficiency of BDNF.Neurotoxicol. Teratol. 24, 695–701.
lton, D., Markowska, A., Voytko, M.L., Givens, B., Gorman, L., Wenk, G., 1991. Basalforebrain cholinergic system: a functional analysis. Adv. Exp. Med. Biol. 295,353–372.
sawa, T., Kato, Y., 2005. Protective role of antioxidative food factors in oxidativestress caused by hyperglycemia. Ann. N.Y. Acad. Sci. 1043, 440–451.
icciotto, M.R., Caldarone, B.J., King, S.L., Zachariou, V., 2000. Nicotinic receptors inthe brain. Links between molecular biology and behavior. Neuropsychopharma-cology 22, 451–465.
ramming, S., Thorsteinsson, B., Theilgaard, A., Pinner, B.M., Binder, C., 1986. Cogni-tive function during hypoglycaemia in type I diabetes mellitus. Br. Med. J. 292,647–650.
riyadarsini, K., Maity, D., Naik, G., Kumar, M., Unnikrishnan, M., Satav, J., Mohan,H., 2003. Role of phenolic O H and methylene hydrogen on the free radicalreactions and antioxidant activity of curcumin. Free Radic. Biol. Med. 35, 475–484.
iu, W.Q., Price, L.L., Hibberd, P., Buell, J., Collins, L., Leins, D., Mwamburi, D.M.,Rosenberg, I., Smaldone, L., Scott, T.M., Siegel, R.D., Summergrad, P., Sun, X., Wag-ner, C., Wang, L., Yee, J., Tucker, K.L., Folstein, M., 2006. Executive dysfunction inhomebound older people with diabetes mellitus. J. Am. Geriatr. Soc. 54, 496–501.
uincozes-Santos, Andreazzaa, A.C., Nardin, A.P., Funchala, C., Goncalves, C.A., Got-tfried, C., 2007. Resveratrol attenuates oxidative-induced DNA damage in C6Glioma cells A. Neurotoxicology 28, 886–891.
ato, A., Sato, Y., Uchida, S., 2004. Activation of the intracerebral cholinergic nerve
fibers originating in the basal forebrain. Neurosci. Lett. 361, 90–93.catchard, G., 1949. The attraction of proteins for small molecules and ions. Ann.N.Y. Acad. Sci. 51, 660–672.
chroder, H., Zilles, K., Maelicke, A., Hajo, S.F., 1989. Immunohisto and cytochemicallocalization of cortical nicotinic cholinoceptors in rat and man. Brain. Res. 502,287–295.
ular Endocrinology 331 (2011) 1–10
Schwartz, M.W., Sipols, A.J., Kahn, S.E., Lattemann, D.P., Taborsky Jr., G.J., Bergman,R.N., Woods, S.C., Porte Jr., D., 1990. Kinetics and specificity of insulin uptakefrom plasma into cerebrospinal fluid. Am. J. Physiol. 259, E378–E383.
Seo, K.-II, Choi, M.-S., Jung Un, J., Kim, H-J., Yeo, J., Jeon, Seon-Min., Lee, M.-K., 2008.Effect of curcumin supplementation on blood glucose, plasma insulin, and glu-cose homeostasis related enzyme activities in diabetic db/db mice. Mol. Nutr.Food Res. 52 (9), 995–1004.
Sharma, S., Kulkarni, S.K., Chopra, K., 2006. Curcumin, the active principle of turmeric(Curcuma longa), ameliorates diabetic nephropathy in rats. Clin. Exp. Pharmacol.Physiol. 33, 940–945.
Shishodia, S., Sethi, G., Aggarwal, B.B., 2005. Curcumin: getting back to the roots.Ann. N.Y. Acad. Sci. Nov. 1056, 206–217.
Soreq, H., Seidman, S., 2001. Acetylcholinesterase—new roles for and old actor. Nat.Rev. Neurosci. 2, 294–302.
Surh, Y.J., Chun, K.S., Cha, H.H., Han, S.S., Keum, Y.S., Park, K.K., Lee, S.S., 2001. Molec-ular mechanisms underlying chemopreventive activities of anti-inflammatoryphytochemicals, down-regulation of COX-2 and iNOS through suppression ofNF kappa B activation. Mutat. Res. 480, 243–268.
Tõugu, V., Kesvatera, T., 1996. Role of ionic interactions in cholinesterase catalysis.Biochim. Biophys. Acta 1298, 12–30.
Uchida, S., Kagitani, F., Nakayama, H., Sato, A., 1997. Effect of stimulation of nicotiniccholinergic receptors on cortical cerebral blood flow and changes in the effectduring aging in anesthetized rats. Neurosci. Lett. 228, 203–206.
Van der Zee, E.A., Streefland, C., Strosberg, A.D., Schro der, H., Luiten, P.G.M., 1992.Visualization of cholinoceptive neurons in the rat neocortex: colocalization ofmuscarinic and nicotinic acetylcholine receptors. Mol. Brain Res. 14, 326–336.
Voytko, M.L., Olton, D.S., Richardson, R.T., Gorman, L.K., Tobin, J.R., Price, D.L., 1994.Basal forebrain lesions in monkeys disrupt attention but not learning and mem-ory. J. Neurosci. 14, 167–186.
Wevers, A., Monteggia, L., Nowacki, S., Bloch, W., Schu, U., Lindstrom, J., Pereira, E.F.R.,Eisenberg, H., Giacobini, E., de Vos, R.A.I., Jansen Steur, E.N.H., Maelicke, A., Albu-querque, E.X., Schro der, H., 1999. Expression of nicotinic acetylcholine receptorsubunits in the cerebral cortex in Alzheimer’s disease—histotopographical cor-relation with amyloid plaques and hyperphosphorylated tau-protein. Eur. J.Neurosci. 11, 2551–2565.
Willis, L.M., Shukitt-Hale, B., Joseph, J.A., 2009. Modulation of cognition and behaviorin aged animals: role for antioxidant- and essential fatty acid-rich plant foods.Am. J. Clin. Nutr. 89, 1602S–1606S.
Woo, E., Schmitter-Edgecombe, M., Fancher, J.B., 2008. Memory prediction accu-racy in younger and older adults: a cross-sectional and longitudinal analysis.Neuropsychol. Dev. Cogn. B. Aging. Neuropsychol. Cogn. 15 (1), 68–94.
Yamamura, H.I., Synder, G., 1981. Binding of [3H] QNB in rat brain. Proc. Natl. Acad.Sci. U.S.A. 71, 1725–1729.
Zhao, W.-Q., Alkon, D.L., 2001. Role of insulin and insulin receptor in learning andmemory. Mol. Cell. Endocrinol. 177, 125–134.
Zhao, W., Chen, H., Xu, H., Moore, E., Meiri, N., Quon, M.J., Alkon, D.L., 1999. Braininsulin receptors and spatial memory. Correlated changes in gene expression,tyrosine phosphorylation, and signaling molecules in the hippocampus of watermaze trained rats. J. Biol. Chem. 274 (49), 34893–34902.





























