Embed Size (px)
Citation preview

Research ArticleKrill Oil Ameliorates Mitochondrial Dysfunctions in RatsTreated with High-Fat Diet
Alessandra Ferramosca Annalea Conte and Vincenzo Zara
Dipartimento di Scienze e Tecnologie Biologiche ed Ambientali Universita del SalentoVia Provinciale Lecce-Monteroni 73100 Lecce Italy
Correspondence should be addressed to Alessandra Ferramosca alessandraferramoscaunisalentoit
Received 15 October 2014 Revised 9 December 2014 Accepted 11 January 2015
Academic Editor Karsten Weylandt
Copyright copy Alessandra Ferramosca et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
In recent years several studies focused their attention on the role of dietary fats in the pathogenesis of hepatic steatosis It has beendemonstrated that a high-fat diet is able to induce hyperglycemia hyperinsulinemia obesity and nonalcoholic fatty liver diseaseOn the other hand krill oil a novel dietary supplement of n-3 PUFAs has the ability to improve lipid and glucose metabolismexerting possible protective effects against hepatic steatosis In this studywe have investigated the effects of krill oil onmitochondrialenergetic metabolism in animals fed a high-fat diet To this end male Sprague-Dawley rats were divided into three groups and fedfor 4 weeks with a standard diet (control group) a diet with 35 fat (HF group) or a high-fat diet supplemented with 25 krilloil (HF+KO group) The obtained results suggest that krill oil promotes the burning of fat excess introduced by the high-fat dietThis effect is obtained by stimulating mitochondrial metabolic pathways such as fatty acid oxidation Krebs cycle and respiratorychain complexes activity Modulation of the expression of carrier proteins involved inmitochondrial uncoupling was also observedOverall krill oil counteracts the negative effects of a high-fat diet on mitochondrial energetic metabolism
1 Introduction
Nonalcoholic fatty liver disease (NAFLD) is a condition inwhich fat accumulates in the cells of liver It occurs when therate of hepatic fatty acid uptake from plasma and de novolipogenesis is greater than the rate of fatty acid oxidationand export Traditionally NAFLD is a histological diseasewhich progresses from pure fatty liver (simple steatosis)throughnonalcoholic steatohepatitis (NASH) to liver fibrosiscirrhosis and eventually hepatocellular carcinoma
Several evidences indicate that hepatic mitochondrialdysfunction is crucial to the pathogenesis of NAFLD [1ndash3]However the whole spectrum of mitochondrial adaptationsduring hepatic steatosis remains to be characterized A recentstudy [4] proposed a model describing the development ofNAFLD in which overflow of free fatty acids to hepatocytesleads to an increase in mitochondrial energetic metabo-lism This seems necessary to supply the energy needed fortriglycerides storage Later mitochondrial oxidative stress
morphological abnormalities and finally hepatocytes apop-tosis lead to the development of the second hit of steatosis
As liver fat accumulation can originate from dietaryintake it is of critical importance to understand how differentdiets and their macronutrient composition can influencethe development of NAFLD Despite contradictory resultsregarding the role of different diets on NAFLD it is rea-sonable to propose that overconsumption of either fat orcarbohydrates may promote the development of NAFLD [5]It seems that specific fatty acids or carbohydrates are moreprone to induce or improveNAFLD Indeed diets rich in fattyacids particularly saturated or in refined carbohydrates canactually exacerbate NAFLD [5]
Interestingly it has been demonstrated that dietary fatsnot only influence the pathogenesis of NAFLD or NASH butalso may prevent or reverse its expression [6] Therefore acondition of fatty liver was induced in rodent animal modelsby the use of high-fat (HF) diets [2 7] whereas supplemen-tation of the diet with n-3 PUFAs helped in the prevention
Hindawi Publishing CorporationBioMed Research InternationalArticle ID 645984
2 BioMed Research International
andor in the treatment of steatotic liver [2] Moreoverthere is much stronger evidence that polyunsaturated oilsare responsible for the progression from simple steatosis toNASH and these effects may result from a high intake of totalPUFAs or may result from a high ratio of n-6 to n-3 fattyPUFAs [8]
In this context it has been reported that a dietary krilloil (KO) supplementation has the capacity to reduce fattyliver in mice and rats [2 9 10] Recently it has been alsodemonstrated that this oil stimulated the catabolization of fatexcess introduced by hypercaloric diets while inhibiting denovo fatty acid synthesis and therefore preventing the onsetof fatty liver [2 10]
KO is a novel dietary supplement extracted from Antarc-tic krill (Euphausia superba) a shrimp-like zooplanktonwhich has become increasingly popular during the lastdecade This oil contains two n-3 PUFAs eicosapentaenoicacid (EPA 20 5) and docosahexanoic acid (DHA 22 6) inamounts similar to those present in fish oil However in KOthese long-chain n-3 fatty acids are present in the form ofphospholipids rather than triglycerides as in the case of fishoil [11] Furthermore the ratio of EPA to DHA present inKO is higher than that in fish oil It has been proposed thatthese peculiar characteristics of KO composition render itmore efficient than fish oil in the modulation of activity andexpression of many enzymes involved in lipid metabolism inanimal models [10 12]
In this study we have investigated the molecular mech-anism by which KO exerts a possible protective effect onmitochondrial energetic metabolism in animals fed a dietenriched in fat To this end we have thoroughly analyzedmitochondrial respiration by monitoring oxygen consump-tion and the activity of the single respiratory chain complexesinmitochondriaThe expression of proteins related to the res-piratory chain function was also investigated The obtainedresults led us to depict a possible framework for the protectiveeffects of KO against hepatic steatosis
2 Materials and Methods
21 Materials Bio-Rad protein assay kit was purchased fromBio-Rad sodium pyruvate malic acid succinic acid ascorbicacid NNN1015840N1015840-tetramethyl-p-phenylenediamine (TMPD)palmitoyl-L-carnitine rotenone antimycin A oligomy-cin coenzyme A trilithium salt (CoA-SH) acetyl-CoA oxa-loacetic acid thiamine pyrophosphate (TPP) 55-dithio-bis-(2-nitrobenzoic acid) (DTNB) 26-dichlorophenolin-dophenol (DCPIP) carbonyl cyanide-4-(trifluorome-thoxy)phenylhydrazone (CCCP) phosphoenolpyruvate(PEP) ADP ATP NAD+ NADH NADPH KCNpyruvate kinase lactate dehydrogenase cytochrome 119888and decylubiquinone were from Sigma KO was a generousgift of Aker BioMarine ASA (Oslo Norway) Antibodiesagainst AAC andUCP2 were from Santa Cruz Biotechnology(sc-11433 and sc-6526) antibodies against OXPHOS proteinswere from Mitosciences (ab110413) Kits for the assay oftriglycerides and total cholesterol were purchased fromFutura System Plasma insulin concentration was analyzedwith a Mercodia Ultrasensitive Mouse Insulin kit Luciferase
ATP assay kit was from Sigma and Lipid Hydroperoxide(LPO) assay kit was from Merck All other reagents were ofanalytical grade
22 Ethics Statement This study was carried out in strictaccordance with the European Committee Council 106Directive (86609EEC) and with the Italian animal welfarelegislation (art 4 and 5 of DL 11692)The Italian Ministry ofHealth specifically approved this study
23 Animals andDiets Male Sprague-Dawley rats (70ndash100 g)were obtained from Harlan (Carezzana Italy) and housedindividually in animal cages at a temperature of 22 plusmn 1∘Cwith a 12 12 hour light-dark cycle and 30ndash40 humidityAfter 1 week of acclimatization 15 rats were divided into threegroups of 5 animals each The first group (control group)received a standard diet also known as standard rodent chowcontaining 62 fat and 442 carbohydrates (Global Diet2018S from Harlan Teklad) The second group received a dietwith a higher content (35) of fat (and therefore referred to asHF group) and 361 of carbohydrates (Diet TD03584 fromHarlan Teklad) The third group of animals (HF+KO group)was fed with the above reported HF diet supplemented with25 (ww)KODiets composition is described inTable 1Theanimals were treated for 4 weeks with ad libitum access todiets and water
At the end of dietary treatment (after 4 weeks) bodyand liver weight were determined when rats were killed bydecapitation according to the guidelines for the care and useof laboratory animals For the determination of plasma lipidsglucose and insulin control and treated rats were starvedovernight before killing Homeostasis model assessment(HOMA) index was calculated according to the followingformula HOMA = (fasting glucose (mM) times fasting insulin(120583UmL))225 [13]
Liver lipids were extracted and quantified as described in[14]
24 Assay of Enzymes Involved in Glucose and Fatty Acid Oxi-dation Rat liver mitochondria were prepared by standardprocedures and protein concentration was determined by theBradford method [15]
Mitochondria (025mg of mitochondrial proteinmL)were incubated in a buffer containing 25mM MOPS and005 Triton X-100 at pH 74 Solubilization of mitochondriawith 005 Triton X-100 inhibited complex I of the respi-ratory chain preventing consumption of NADH Pyruvatedehydrogenase (PDH) activity was measured spectrophoto-metrically as the rate of NAD+ reduction to NADH uponaddition of 25mMpyruvate 01mMCoA-SH 02mMTPP1mMNAD+ and 5mMMgCl
2(pH 74) [16]
Citrate synthase (CS) activity was determinedin a medium containing 100mMTris-HCl (pH 81)01mMDTNB and 025mMacetyl-CoA The reaction wasstarted by adding 025mMoxaloacetate followed by moni-toring changes at 412 nm for 3min [17]
Total carnitine palmitoyl-CoA transferase (CPT) andCPT I activity was determined spectrophotometrically at
BioMed Research International 3
Table 1 Composition of diets ()
Control HF HF+KOProteins 186 204 199Lipids 62 352 368
Fatty acids140 mdash 05 06160 07 87 90180 02 43 43161 mdash mdash 01181 (n9-7-5) 12 158 160182 (n6) 31 35 35183 (n3) 03 mdash mdash205 (n3) EPA mdash mdash 03226 (n3) DHA mdash mdash 02sum SFA 09 135 139sumMUFA 13 158 161sumPUFA 34 35 41sumPUFA n-3 03 mdash 06
Carbohydrates 442 361 352kcal100 g 310 540 549Calories from protein () 240 150 140Calories from fat () 180 580 600Calories from carbohydrates () 580 270 260The control group of animals received a standard diet (Global Diet 2018Sfrom Harlan Teklad) The HF group received a diet with 35 fat (DietTD03584 from Harlan Teklad) and the KO group was fed with the abovereported HF diet supplemented with 25 (ww) KO Fatty acids wereextracted from the three diets and analyzed by gas-liquid chromatographyEnergy content was calculated using 4 kcalg for protein and carbohydrateand 9 kcalg for lipid
412 nm in freshly isolated rat liver mitochondria essentiallyas described previously [18]
25 Mitochondrial Respiration Efficiency Mitochondrial res-piration (03mg ofmitochondrial proteinmL) wasmeasuredin a medium consisting of 220mM sucrose 20mMKCl25mMKH
2PO4 1 mMEDTA 20mMHepes 5mMMgCl
2
and 01 BSA pH 74 by a Clark oxygen electrode at 25∘C[2] The addition of different substrates permitted to evalu-ate mitochondrial respiration when respiratory complexes I(5mM pyruvate 25mM malate) II (5mM K-succinate5 120583M rotenone) and IV (10mM ascorbate 02mM TMPD5 120583M rotenone 5 120583M antimycin A) were stimulated [19 20]Instead the addition of 40 120583M palmitoyl-L-carnitine and25mM malate permitted revealing oxygen consumptionfrom 120573-oxidation pathway
For each substrate after 2min state 3 respiration wasinduced by the addition of 03mM ADP Respiratory controlratio (RCR) was calculated as the ratio of the rate of oxygenuptake in the presence of added ADP (state 3) to the rateobserved when added ADP had been completely phospho-rylated to ATP (state 4)
26 Assay of Mitochondrial Respiratory Complex ActivityComplex I activity was determined using decylubiquinone
as electron acceptor and NADH as donor Rat liver mito-chondria (40 120583g of protein) were pretreated first by freeze-thawing the samples two or three times in hypotonicmedium consisting of 25mMKH
2PO4(pH 72) 5mMMgCl
2
followed by a hypotonic shock in H2O to disrupt mito-
chondrial membranes The reaction started after the addi-tion of 50mMTris (pH 80) medium supplemented with5mgmLBSA 08mMNADH as donor 240120583MKCN 4 120583MantimycinA and 50120583Mof the acceptor decylubiquinoneTheoxidation of NADH was measured following the decrease inabsorbance at 340 nm for 3min [20]
Complex II activity assay was performed following thedecrease in absorbance at 600 nm resulting from the reduc-tion of DCPIP as electron acceptor and using succinateas donor Rat liver mitochondria (40 120583g of protein) werepretreated first by freeze-thawing following by an incubationwith succinate 10min at 30∘C in a medium containing10mMKH
2PO4(pH 78) 80 120583M DCPIP as acceptor 4 120583M
rotenone and 02mM ATP The reaction started by additionof 80 120583M decylubiquinone and the activity was measured for5min [20]
Complex III activity was determined by measuring thereduction of oxidized cytochrome 119888 at 550 nm Rat livermitochondria (40 120583g protein) were incubated for 1min at30∘C in a reaction medium containing 50mMKH
2PO4
(pH 72) 001 Tween-20 50mM EDTA 4mMKCN and40mM oxidized cytochrome 119888 The reaction was initiatedby adding the ubiquinol analog decylubiquinol (DBH
2) to
a final concentration of 50 120583M and the rate of cytochrome119888 reduction was calculated from the absorbance increase at550 nm [2]
Complex IV activity assay was performed following thedecrease in absorbance at 550 nm resulting from the oxida-tion of reduced cytochrome 119888 Rat liver mitochondria (20120583gof protein) were incubated in 1mL of isosmotic mediumcontaining 250mM sucrose 10mM KH
2PO4(pH 65) and
10 120583M reduced cytochrome 119888 Cytochrome 119888 solution wasfreshly reduced by adding some crystals of sodiumdithioniteThe addition of 25mM laurylmaltoside permitted permeabi-lization of the outer mitochondrial membrane to cytochrome119888 and the decrease in absorbance at 550 nm was followed for3min [20]
ATP-synthase activity (ATP hydrolysis) was measuredspectrophotometrically by a coupled assay using lactatedehydrogenase and pyruvate kinase as the coupling enzyme[20] For each ATP molecule that is hydrolyzed a moleculeof NADH is oxidized Rat liver mitochondria were incubatedfirst in distilled water for 30 s at 37∘C and then in a mediumcontaining 200 120583L of 50mMTris (pH 80) 5mgmLBSA20mMMgCl
2 50mMKCl 15120583MCCCP 5 120583Mantimycin A
10mMPEP 25mMATP 4 units of lactate dehydrogenaseand pyruvate kinase and 1mMNADH previously incubatedfor 5min at 37∘C Reaction was started by the addition of themedium to mitochondria and monitored for 3min Finallythe addition of 3 120583Moligomycin permitted distinguishing theATPase activity coupled to the respiratory chain [20]
27 Western Blot Analysis The expression of oxidative phos-phorylation system (OXPHOS) proteins and mitochondrial
4 BioMed Research International
Table 2 Body and liver weight serum parameters and liver fat content
Control HF HF+KOBody weight (g) 2458 plusmn 78 2772 plusmn 158lowast 2352 plusmn 252
Liver weight (g100 g body weight) 39 plusmn 06 35 plusmn 05 41 plusmn 03Glycemia (mM) 29 plusmn 01 49 plusmn 07lowast 41 plusmn 04lowast
Insulin (120583UmL) 501 plusmn 75 1428 plusmn 100lowast 777 plusmn 50lowast
HOMA 64 plusmn 01 312 plusmn 03lowast 142 plusmn 01lowast
Serum triglycerides (mgdL) 1373 plusmn 129 1471 plusmn 88 1386 plusmn 92Serum cholesterol (mgdL) 926 plusmn 83 956 plusmn 42 941 plusmn 62Liver triglycerides (mgg liver) 86 plusmn 04 136 plusmn 10lowast 83 plusmn 05
Liver cholesterol (mgg liver) 27 plusmn 04 66 plusmn 04lowast 35 plusmn 05
Each point represents the mean plusmn SD for 5 animals lowast119875 lt 005 versus rats fed a control diet 119875 lt 005 versus rats fed HF diet
carrier proteins AAC (ADPATP carrier) and UCP2 (uncou-pling protein 2) was determined by western blotting analysisPolyacrylamide gel electrophoresis was performed in thepresence of 01 SDS (SDS-PAGE) according to standardprocedures The mitochondrial proteins that have been sepa-rated by SDS-PAGEwere transferred to a nitrocellulosemem-brane For the OXPHOS proteins a cocktail directed againstNDUFB8 (Complex I) SDHB (Complex II) COR2 (ComplexIII) COX1 (Complex IV) and ATP5A (ATP-synthase) wasused at a dilution 1 1times101 and the immunoreaction was car-ried out with monoclonal antibody by chemiluminescenceFor AAC UCP2 and porin (a protein used as a control)detection antisera were used at a dilution of 1 1times103 and theimmunoreaction was carried out with polyclonal antibody bychemiluminescence
28 ATP and Lipid Hydroperoxide (LPO) Levels ATP levelsin liver homogenates were determined by using the luciferaseATP assay kit The concentration of ATP in the samples wascalculated from interpolation of a standard curve preparedwith known amounts of ATP Results were expressed asnmoles of ATP per mg of protein
Lipid peroxidation levels in liver samples were deter-mined using a lipid hydroperoxide assay kit which measuresthe redox reactions with ferrous ions LPO content wasexpressed as LPO per mg of protein
29 Statistical Analysis Experimental data represent themeans plusmn SD The data were analyzed by one-way ANOVAand a Tukey-Kramer post hoc analysis was used to detectsignificant differences between the means at a level of 119875 gt005
3 Results
31 Body and Liver Weight Serum Parameters and Liver FatContent As shown in Table 2 a significant increase in bodyweight was recorded in HF animals after 4 weeks of dietarytreatment in agreement with the high caloric content ofthe diet Interestingly the supplementation of the HF dietwith 25 KO significantly prevented this effect Insteadno statistically significant variations were found in the liverweight of animals fed with the different experimental diets
The blood glucose concentration increased in both HFand HF+KO animals after 4 weeks of dietary administrationin comparison to control rats Such an increase was lessevident in the case of the HF+KO fed animals A massiveincrease in the levels of insulin was revealed in the plasmaof HF animals in comparison to control rats Interestinglythe KO supplementation significantly mitigated this increaseHF rats showed a marked increase in HOMA index demon-strating typical insulin resistance induced by the high-fat dietIn the HF+KO animals HOMA decreased showing reducedinsulin resistance induced by the addition of KO
No significant difference was detected in the serumlipid (triglyceride and cholesterol) levels between the threegroups of animals On the contrary liver lipid content wasdramatically enhanced in HF animals thus reflecting thepresence of hepatic steatosis (Figure 1) The supplementationwith KO of the HF diet reversed this effect thereby keepingthe lipid concentrations similar to those found in controlrats Accordingly the liver histologic examination revealed abeginning of microvesicular fat depositions only in HF rats
32 Pyruvate Dehydrogenase (PDH) Complex CarnitinePalmitoyltransferase (CPT) and Citrate Synthase (CS) Activ-ity Mitochondria play a central role in energy productionSugars and fatty acids undergo glycolysis and mitochon-drial 120573-oxidation respectively to produce acetyl-CoA (Fig-ure 2(a)) PDH complex activity is the major determinantof glucose oxidation in animal cells We found that PDHactivity remained unchanged in the three groups of animalssuggesting that glucose catabolism was not affected by a HFdiet or by KO supplementation (Figure 2(b))
The limiting step for catabolic pathway of fatty acidoxidation is represented by the activity of CPT I which isresponsible for the entry of fatty acids into the mitochondrialmatrix where 120573-oxidation of fatty acids occurs As shown inFigure 2(c) in the HF animals a clear decrease (about 40)in the CPT I activity was detected On the contrary a strongincrease in CPT I activity was found in theHF+KO ratsThusKO supplemented to theHF diet strongly stimulates fatty acidoxidation
Acetyl-CoA generated essentially from glucose or fromfatty acids is channeled into the Krebs cycle where CS(the rate-limiting enzyme of the Krebs cycle) catalyzes its
BioMed Research International 5
Control HF HF+KO
Figure 1 Liver samples were stored in neutral formaldehyde and embedded in paraffin wax Sections (15120583m) were stained with hematoxylinand eosin
Glucose
Fatty acids
CPT I
Pyruvate
NAD+
120573-oxidationAcetyl-CoA Acyl-CoA
CSCO2
PDH
Oxaloacetate
Krebscycle
FADH2
Mitochondrion
Cytosol
NADH + H+
NADH + H+ NADH + H+
NADH + H+
(a)
0
5
10
15
20
Control HF
PDH
activ
ity (n
molmiddotm
inminus1middotm
g pr
otei
nminus1)
HF+KO
(b)
Control HF0
25
50
75
100
125
150
CPT
I act
ivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(c)
Control HF0
50
100
150
200
250
300
CS ac
tivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(d)
Figure 2Mitochondrialmetabolic pathways fromglucose and fatty acid oxidation Schematic picture of themetabolic pathways fromglucoseand fatty acid oxidation (a) Effect of KO on the activity of mitochondrial enzymes PDH (b) CPT I (c) and CS (d) in control HF or HF+KO-fed rats liver The values reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed a control diet 119875 lt 005 versusrats fed a HF diet
6 BioMed Research International
Table 3 Mitochondrial respiratory efficiency
Control HF HF+KO
Palmitoyl-L-carnitine + malate (120573 oxidation)1198813(nmol O2sdotmLminus1sdotminminus1) 858 plusmn 78 597 plusmn 85lowast 1140 plusmn 112lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 143 plusmn 24 229 plusmn 15lowast 185 plusmn 52lowast
RCR 60 plusmn 04 26 plusmn 03lowast 61 plusmn 08
Pyruvate + malate (Complex I)1198813(nmol O2sdotmLminus1sdotminminus1) 170 plusmn 03 159 plusmn 04lowast 199 plusmn 09lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 54 plusmn 01 76 plusmn 02lowast 66 plusmn 06lowast
RCR 32 plusmn 03 21 plusmn 03lowast 30 plusmn 04
Succinate + rotenone (Complex II)1198813(nmol O2sdotmLminus1sdotminminus1) 615 plusmn 33 531 plusmn 37lowast 783 plusmn 40lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 101 plusmn 03 250 plusmn 12lowast 150 plusmn 19lowast
RCR 61 plusmn 09 21 plusmn 08lowast 52 plusmn 10
Ascorbate + TMPD + rotenone + antimycin A (Complex IV)1198813(nmol O2sdotmLminus1sdotminminus1) 551 plusmn 53 475 plusmn 65lowast 686 plusmn 78lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 93 plusmn 07 172 plusmn 09lowast 128 plusmn 08lowast
RCR 59 plusmn 06 28 plusmn 03lowast 54 plusmn 09
Respiratory control ratio (RCR) was calculated as the ratio of the rate of oxygen uptake in the presence of added ADP (1198813) to the rate observed when added
ADP had been completely phosphorylated to ATP (1198814)
lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
condensation with oxaloacetate to synthesize citrate (Fig-ure 2(a)) A significant decrease (about 55) in the activityof the CS was found in HF animals (Figure 2(d)) whileonly a moderate decrease (about 35) was observed inHF+KO animals in comparison to the control groups Thepartial reversal of CS inhibition observed in HF animals afterKO supplementation could be explained by a compensatoryresponse of this enzyme to sustain oxidation of acetyl-CoAgenerated from fatty acid oxidation
33 Mitochondrial Oxygen Consumption Respiratory ChainComplex Activities and ATP Synthesis The increased fattyacid oxidation observed in HF+KO animals produces higherlevels of reducing equivalents which are normally addressedtowards the mitochondrial oxidative phosphorylation forATP production We therefore analyzed the respiratoryefficiency of freshly isolated mitochondria by oxygraphicmethods Mitochondrial oxygen consumption was measuredwith various substrates in the presence (state 3) or the absence(state 4) of ADP (Table 3) According to the results shownin Figure 2(c) when we used palmitoyl-L-carnitine andmalate as substrates 119881
3(mitochondrial respiration state 3)
decreased inHF animals thus indicating lowermitochondrial120573-oxidation On the other hand the increase in the119881
4(mito-
chondrial respiration state 4) values suggests that the excess ofdietary fat most probably is the cause of a partial uncouplingbetween respiration and phosphorylation in mitochondriaInterestingly the dietary KO supplementation is able toefficiently abolish this effect keeping the mitochondrialrespiratory efficiency unaltered
When we added respiratory substrates for mitochondrialcomplexes I II and IV (Table 3) a little yet significantdecrease in 119881
3was observed in the HF groups in comparison
to control animals In the HF+KO animals a significantincrease in the 119881
3values was found in comparison to HF
and control animals 1198814 also known as the resting state of
respiration showed an increase in theHF andHF+KO groupeven if the increase observed after KO supplementation was
less evident than that found inHF animals As a consequencethe RCRvalueswere profoundly and significantly lower inHFanimals with respect to those calculated for control animalsThe KO supplementation of the HF diet reversed this effectthereby keeping RCR values similar to those calculated forcontrol group
In a more selective approach for investigating the func-tionality of the mitochondrial oxidative phosphorylation weassayed the activity of single components of the respiratorychain complexes IndashIV andATP-synthase (Figure 3(a)) A sig-nificant decrease (about 30) in the activity of all respiratorychain complexes was found in HF animals On the contrarythe values of complexes activity of the HF+KO rats werepractically identical to those measured in control animals
Surprisingly western blotting analysis of the OXPHOSproteins showed significantly higher protein levels in bothHFand HF+KO animals compared to the control group (Figures3(b) and 3(c)) Such an increase was more evident for com-plexes I III and IV which are associated as supercomplexesin the mitochondrial inner membrane [21] This effect couldbe determined by a compensatory mechanism related to thereduction of the activities of the respiratory complexes
In order to prove biological relevance of alteration inmitochondrial respiratory chain activity we measured ATPconcentration and oxidative damage in liver samples fromthe three groups of animals (Figure 3(d)) ATP quantificationdemonstrated that aHFdiet caused a slight but not significantdecrease in ATP content an effect that was rescued by KOInterestingly in parallel experiments we found a significantincrease (about 30) in the LPO levels inHF animals and thiseffect was efficiently reversed by KO supplementation of theHF diet
34 Expression of Proteins Related to the Respiratory ChainFunction Uncoupling protein 2 (UCP2) uncouples respira-tion from oxidative phosphorylation We therefore analyzedthe expression profile of this carrier protein in the threegroups of animals (Figure 4(a)) After 4 weeks of HF dietary
BioMed Research International 7
0
30
60
90
120
Con
trol
HF
Resp
irato
ry co
mpl
exes
activ
ity (
)
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
lowast lowast lowastlowast
lowast
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(a)
CV-ATP5A
CIII-COR2CIV-COX1
CII-SDHB
CI-NDUFB8
Porin
(b)
0
50
100
150
250
OXP
HO
S pr
otei
ns (
) 200lowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
Con
trol
HF
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(c)
30
90
0
60
150
( o
f con
trol)
120
ATP LPO
lowast
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
(d)
Figure 3 Effect of KO on respiratory complexes activity expression and function Effect of KO on respiratory complexes activity (a)Enzymatic activities measured in the control group were set to 100 Effect of KO on OXPHOS proteins levels (b) Liver mitochondrialproteins from control HF or HF+KO-fed rats were separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with acocktail against NDUFB8 SDHB COR2 COX1 ATP5A or the mammalian porin as a controlThe amount of OXPHOS proteins revealed byimmunodecoration in the control group was set to 100 (c) ATP and LPO quantification of liver samples isolated from animals fed a controldiet a HF diet or a HF+KO (d) The amount of ATP or LPO (expressed as nmoles per mg protein) in the control group was set to 100Thevalues reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
treatment the amount of this protein increased about 5-foldcompared to the control group Interestingly a significantsmaller increase in the expression of UCP2 was found afterKO supplementation In fact in the presence of KO in theHF diet the level of this carrier protein was about 50 of thatfound in HF rats
The AAC (ADPATP carrier) is another mitochondrialcarrier which is able to regulate the basal proton conductanceThis protein exchanges ADP for ATP across the mitochon-drial inner membrane and may also play an important rolein the mitochondrial permeability transition pore When weanalyzed the expression of this carrier protein (Figure 4(b))we found an increase of about 22-fold in HF animalscompared to the control group AAC levels in the HF+KOanimals even if higher than those found in the controlrats were significantly lower than those found in HF group
The amount of porin an outer membrane protein tested as acontrol did not change in any group of animals (Figure 4(c))
These results suggest that the KO supplementation is ableto modulate the expression of mitochondrial carrier proteininvolved in the regulation of proton conductance reducingthe uncoupling effect caused by a HF diet
4 Discussions
In animal experiments a diet rich in fat or carbohydrates hasbeen shown to induce hyperglycemia and hyperinsulinemiaand to contribute to the development of obesity and NAFLD[2 6 7 22] This last condition is recognized as the mostcommon chronic liver disease in the Western world NASHis a more severe form of NAFLD that is broadly defined bythe presence of steatosis with inflammation and progressive
8 BioMed Research International
0
100
200
300
500
Control HF
UCP
2 (
)
UCP2
400
lowast
lowast
HF+KO
(a)
0
60
120
180
300
AAC
()
AAC
240
Control HF
lowast
lowast
HF+KO
(b)
0
25
50
75
125
Porin
()
Porin
100
Control HF HF+KO
(c)
Figure 4 Effect of KO on proteins related to the respiratory chain function Effect of KO on proteins levels of UCP2 (a) and AAC (b) Theamount of porin an outer membrane protein was tested as a control (c) Liver mitochondrial proteins from control HF or HF+KO-fed ratswere separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with antisera against UCP2 AAC or the mammalianporinThe amount of proteins revealed by immunodecoration in the control groupwas set to 100The values reported in the figure representthe means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
fibrosis leading then to cirrhosis and hepatocellular carci-noma Some patients with NAFLD develop NASH throughpoorly understood mechanisms Since NAFLD and NASHare considered lifestyle-associated diseases diet interventionis an important approach to their treatment
In this context it is important to underline that theldquoobesogenicrdquoWestern diet has a ratio of fatty acids and carbo-hydrates of approximately 5050Moreover it is characterizedby an insufficient n-3 PUFAs consumption and a high n-6n-3 ratio which are associated with the progression fromsimple steatosis to NASH [8] along with modifications of gutmicrobiota [23]
Although the mechanisms responsible for increasedlipid accumulation in the liver are not yet fully elucidateddecreased capacity to oxidize fatty acids increased transportof fatty acids to liver and increased hepatic fatty acidsynthesis seem to be important factors involved in thepathogenesis of NAFLD [2 6 7 24]Moreover abnormalitiesin mitochondrial morphology and functions seem to beassociated with fatty liver [1ndash3 25ndash27] This finding suggeststhat regulation of mitochondria function and of molecularmechanisms involved in lipid accumulation in the liver couldprovide useful targets to prevent the development of NAFLD
In recent years the use of dietary supplements hasrapidly increased [2 6 10 24 28] with the aim to preventNAFLD KO a dietary supplement of n-3 PUFAs has becomeincreasingly popular because of its capability of reducing
hepatic lipogenesis and of stimulating catabolization ofexcess fat introduced by a HF diet [2 10] The analysisof hepatic transcriptome in mice demonstrated that KOsupplementation was able to regulate genes involved in hep-atic energy metabolism including glucose metabolism lipidbiosynthesis fatty acid metabolism cholesterol biosynthesisand mitochondrial electron transport more efficiently thanfish oil [10 12] These differential results were tentativelyascribed to the influence of the structural form of n-3PUFAs esterified to either phospholipids or triglycerideson a biological response Moreover PUFAs as phospholipids(KO) or triglycerides (fish oil)may differentially influence gutmicrobiota or synthesis of bacterial metabolites
A previous study by our group [2] suggested that KOsupplementation to a HF diet resulted in a retention of nor-malmitochondrial respiration efficiency whichwas impairedin HF animals The aim of the present study was thereforeto analyze the molecular mechanisms by which dietary fatsinfluence mitochondrial energetic metabolismThis new andintriguing aspect was evaluated in hepatocytes from rats feda HF diet or a HF diet supplemented with 25 KO with theaim to investigate the role of mitochondria in themodulationof nutrient metabolism exerted by KO
No change in PDH activity which essentially producesacetyl-CoA from carbohydrate was seen after administrationof HF or HF+KO diets Nevertheless fasting serum glucoseand insulin were higher after feeding a HF diet suggesting
BioMed Research International 9
an altered insulin sensitivity as indicated by HOMA indexchanges In fact an excess of energy supplied by fat corre-sponds to a scarce tissutal utilization of glucose thereby caus-ing hyperglycemia and hyperinsulinemia Insulin resistancewas reduced after KO supplementation
CPT I activity which is involved in fatty acid oxidationwas inhibited by a HF diet and stimulated after KO supple-mentationThe increase inCPT I activity observed inHF+KOgroup could be explained by the lower levels ofmalonyl-CoAametabolic intermediate of hepatic lipogenesis which in thiscase is instead strongly inhibited [2 10] Interestingly thecapability of KO to stimulate fatty acid oxidation seems tobe associated to the presence of a high lipid content (35)in the experimental diet used in this study In fact whenKO was added to a standard diet the expression of enzymesinvolved inmitochondrial fatty acid 120573-oxidation significantlydecreased [12]
The modulation of the CPT I activity observed in HFand HF+KO animals seems to be accompanied by a parallelmodulation of the activity of CS a Krebs cycle enzyme thatcatalyzes the synthesis of citrate fromoxaloacetate and acetyl-CoA Recently a strong and significant correlation of CSwith obesity was noted with a reduced enzyme activity inmitochondria of human omental adipose tissue in the stateof obesity [28] According to this observation the increasein CS activity revealed after KO supplementation could beexplained as a compensatory response to maintain the entryof acetyl-CoA produced by fatty acid oxidation into the Krebscycle It is interesting to underline that citrate generated byCS continues in the Krebs cycle In fact in animals fed aHF diet supplemented with KO a reduction in the activityand in the expression of citrate carrier protein (CIC) whichtransports citrate from mitochondria to the cytosol for fattyacid synthesis was observed [2 10 24]
NADH and FADH2generated by 120573-oxidation and Krebs
cycle are used in the final common oxidative phosphorylationsystem (OXPHOS) to generate ATP This process is coupledwith the transfer of electrons along the respiratory chainOur data indicate that a HF diet caused a strong decrease inthe mitochondrial respiratory efficiency and this effect couldbe due to a possible uncoupling effect exerted by the excessof fatty acids present in this diet In fact we found a littledecrease in 119881
3and a strong increase in 119881
4values in liver
mitochondria isolated fromHF animals1198814is the resting state
of mitochondrial respiration and an increase in this valuesuggests a stimulus ofmitochondrial respiration independentof ADP phosphorylation In other terms during HF feedinga lower coupling betweenmitochondrial respiration andATPsynthesis occurs Interestingly the addition of KO to the HFdiet almost completely reversed this effect thereby leadingmitochondria respiration efficiency to be comparable to thatof control rats
When we assayed in a more selective approach theactivity of single components of the respiratory chain weobserved a significant decrease in the activity of all respira-tory chain complexes in HF animals according to a recentstudy [29] inwhich an impairment ofmitochondrial functionand in particular of complex I activity was found AfterKO supplementation the values of complex activities were
identical to those measured in control animals Expressionof OXPHOS proteins significantly increased in HF animalscompared to the control group and this evidence was accom-panied by the presence of ATP levels in HF animals thatwere only slightly decreased in comparison to those foundin control animals According to this result previous studies[30 31] demonstrated that a high fat content in the dietinduced higher level of these proteins as a compensatoryresponse to the reduced activity of the respiratory chaincomplexes When we performed western blotting analysis inmitochondria isolated from HF+KO animals liver we founda higher level of the OXPHOS proteins compared to thecontrol and theHF groups probably caused by the synergisticeffect of the HF diet and the KO supplementation Indeedit is already known that genes involved in mitochondrialOXPHOS proteins expression were upregulated in miceconsuming KO as revealed from the transcriptomic analysis[12] Overall thismeans that KO is able to induce the burningof excess of fat introduced by diet stimulating fatty acidoxidation Krebs cycle and respiratory chain complex activityand preventing a condition of oxidative damage
Uncoupling between mitochondrial respiration and ATPsynthesis observed in HF animals could be caused bya different expression of mitochondrial carrier proteinsinvolved in the regulation of proton conductance It is knownthat uncoupling proteins (UCPs) homologues uncouplemitochondrial respiration from oxidative phosphorylationincreasing thermogenesis while reducing the efficiency ofATP synthesis In a previous study [32] an increase in theexpression of the UCP2 isoform was found in rats withNAFLD Interestingly in this study we found a massiveincrease of this carrier protein inHF animals but the additionof KO to the diet was able to strongly reduce the expressionof hepatic UCP2 Therefore it is reasonable to assume thatthe addition of KO to the HF diet can increase the couplingbetween mitochondrial respiration and ATP synthesis by adownregulation of UCP2 expression
Not only are UCPs involved in mitochondrial uncou-pling butAAC is another carrier protein of particular interestin the understanding of the mitochondrial energy couplingprocess [33]This carrier catalyzes the exchange of ADP3minus forATP4minus and therefore utilizes as its driving force the electro-chemical potential generated across the inner membrane bythe respiratory chain The exchange reaction is electrogenicand driven by the electrical component of the proton gradi-ent so that entry of ADP into mitochondria and exit of ATPare favored We can therefore speculate that if mitochondriaare uncoupled and proton gradient was dissipated by theentry of H+ into the matrix by UCP2 an upregulation ofthe expression of AAC could compensate a reduced fluxof ATP from mitochondria when membrane potential wasreduced This hypothesized compensatory mechanism wasobserved in animals fed a HF diet The increase in theexpression of AAC was less evident in HF+KO animals inwhich livermitochondria respiratory efficiencywas unalteredin comparison to the control animals
In conclusion the present study indicates that KO coun-teracts the negative effects of a HF diet In particular our
10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
andor in the treatment of steatotic liver [2] Moreoverthere is much stronger evidence that polyunsaturated oilsare responsible for the progression from simple steatosis toNASH and these effects may result from a high intake of totalPUFAs or may result from a high ratio of n-6 to n-3 fattyPUFAs [8]
In this context it has been reported that a dietary krilloil (KO) supplementation has the capacity to reduce fattyliver in mice and rats [2 9 10] Recently it has been alsodemonstrated that this oil stimulated the catabolization of fatexcess introduced by hypercaloric diets while inhibiting denovo fatty acid synthesis and therefore preventing the onsetof fatty liver [2 10]
KO is a novel dietary supplement extracted from Antarc-tic krill (Euphausia superba) a shrimp-like zooplanktonwhich has become increasingly popular during the lastdecade This oil contains two n-3 PUFAs eicosapentaenoicacid (EPA 20 5) and docosahexanoic acid (DHA 22 6) inamounts similar to those present in fish oil However in KOthese long-chain n-3 fatty acids are present in the form ofphospholipids rather than triglycerides as in the case of fishoil [11] Furthermore the ratio of EPA to DHA present inKO is higher than that in fish oil It has been proposed thatthese peculiar characteristics of KO composition render itmore efficient than fish oil in the modulation of activity andexpression of many enzymes involved in lipid metabolism inanimal models [10 12]
In this study we have investigated the molecular mech-anism by which KO exerts a possible protective effect onmitochondrial energetic metabolism in animals fed a dietenriched in fat To this end we have thoroughly analyzedmitochondrial respiration by monitoring oxygen consump-tion and the activity of the single respiratory chain complexesinmitochondriaThe expression of proteins related to the res-piratory chain function was also investigated The obtainedresults led us to depict a possible framework for the protectiveeffects of KO against hepatic steatosis
2 Materials and Methods
21 Materials Bio-Rad protein assay kit was purchased fromBio-Rad sodium pyruvate malic acid succinic acid ascorbicacid NNN1015840N1015840-tetramethyl-p-phenylenediamine (TMPD)palmitoyl-L-carnitine rotenone antimycin A oligomy-cin coenzyme A trilithium salt (CoA-SH) acetyl-CoA oxa-loacetic acid thiamine pyrophosphate (TPP) 55-dithio-bis-(2-nitrobenzoic acid) (DTNB) 26-dichlorophenolin-dophenol (DCPIP) carbonyl cyanide-4-(trifluorome-thoxy)phenylhydrazone (CCCP) phosphoenolpyruvate(PEP) ADP ATP NAD+ NADH NADPH KCNpyruvate kinase lactate dehydrogenase cytochrome 119888and decylubiquinone were from Sigma KO was a generousgift of Aker BioMarine ASA (Oslo Norway) Antibodiesagainst AAC andUCP2 were from Santa Cruz Biotechnology(sc-11433 and sc-6526) antibodies against OXPHOS proteinswere from Mitosciences (ab110413) Kits for the assay oftriglycerides and total cholesterol were purchased fromFutura System Plasma insulin concentration was analyzedwith a Mercodia Ultrasensitive Mouse Insulin kit Luciferase
ATP assay kit was from Sigma and Lipid Hydroperoxide(LPO) assay kit was from Merck All other reagents were ofanalytical grade
22 Ethics Statement This study was carried out in strictaccordance with the European Committee Council 106Directive (86609EEC) and with the Italian animal welfarelegislation (art 4 and 5 of DL 11692)The Italian Ministry ofHealth specifically approved this study
23 Animals andDiets Male Sprague-Dawley rats (70ndash100 g)were obtained from Harlan (Carezzana Italy) and housedindividually in animal cages at a temperature of 22 plusmn 1∘Cwith a 12 12 hour light-dark cycle and 30ndash40 humidityAfter 1 week of acclimatization 15 rats were divided into threegroups of 5 animals each The first group (control group)received a standard diet also known as standard rodent chowcontaining 62 fat and 442 carbohydrates (Global Diet2018S from Harlan Teklad) The second group received a dietwith a higher content (35) of fat (and therefore referred to asHF group) and 361 of carbohydrates (Diet TD03584 fromHarlan Teklad) The third group of animals (HF+KO group)was fed with the above reported HF diet supplemented with25 (ww)KODiets composition is described inTable 1Theanimals were treated for 4 weeks with ad libitum access todiets and water
At the end of dietary treatment (after 4 weeks) bodyand liver weight were determined when rats were killed bydecapitation according to the guidelines for the care and useof laboratory animals For the determination of plasma lipidsglucose and insulin control and treated rats were starvedovernight before killing Homeostasis model assessment(HOMA) index was calculated according to the followingformula HOMA = (fasting glucose (mM) times fasting insulin(120583UmL))225 [13]
Liver lipids were extracted and quantified as described in[14]
24 Assay of Enzymes Involved in Glucose and Fatty Acid Oxi-dation Rat liver mitochondria were prepared by standardprocedures and protein concentration was determined by theBradford method [15]
Mitochondria (025mg of mitochondrial proteinmL)were incubated in a buffer containing 25mM MOPS and005 Triton X-100 at pH 74 Solubilization of mitochondriawith 005 Triton X-100 inhibited complex I of the respi-ratory chain preventing consumption of NADH Pyruvatedehydrogenase (PDH) activity was measured spectrophoto-metrically as the rate of NAD+ reduction to NADH uponaddition of 25mMpyruvate 01mMCoA-SH 02mMTPP1mMNAD+ and 5mMMgCl
2(pH 74) [16]
Citrate synthase (CS) activity was determinedin a medium containing 100mMTris-HCl (pH 81)01mMDTNB and 025mMacetyl-CoA The reaction wasstarted by adding 025mMoxaloacetate followed by moni-toring changes at 412 nm for 3min [17]
Total carnitine palmitoyl-CoA transferase (CPT) andCPT I activity was determined spectrophotometrically at
BioMed Research International 3
Table 1 Composition of diets ()
Control HF HF+KOProteins 186 204 199Lipids 62 352 368
Fatty acids140 mdash 05 06160 07 87 90180 02 43 43161 mdash mdash 01181 (n9-7-5) 12 158 160182 (n6) 31 35 35183 (n3) 03 mdash mdash205 (n3) EPA mdash mdash 03226 (n3) DHA mdash mdash 02sum SFA 09 135 139sumMUFA 13 158 161sumPUFA 34 35 41sumPUFA n-3 03 mdash 06
Carbohydrates 442 361 352kcal100 g 310 540 549Calories from protein () 240 150 140Calories from fat () 180 580 600Calories from carbohydrates () 580 270 260The control group of animals received a standard diet (Global Diet 2018Sfrom Harlan Teklad) The HF group received a diet with 35 fat (DietTD03584 from Harlan Teklad) and the KO group was fed with the abovereported HF diet supplemented with 25 (ww) KO Fatty acids wereextracted from the three diets and analyzed by gas-liquid chromatographyEnergy content was calculated using 4 kcalg for protein and carbohydrateand 9 kcalg for lipid
412 nm in freshly isolated rat liver mitochondria essentiallyas described previously [18]
25 Mitochondrial Respiration Efficiency Mitochondrial res-piration (03mg ofmitochondrial proteinmL) wasmeasuredin a medium consisting of 220mM sucrose 20mMKCl25mMKH
2PO4 1 mMEDTA 20mMHepes 5mMMgCl
2
and 01 BSA pH 74 by a Clark oxygen electrode at 25∘C[2] The addition of different substrates permitted to evalu-ate mitochondrial respiration when respiratory complexes I(5mM pyruvate 25mM malate) II (5mM K-succinate5 120583M rotenone) and IV (10mM ascorbate 02mM TMPD5 120583M rotenone 5 120583M antimycin A) were stimulated [19 20]Instead the addition of 40 120583M palmitoyl-L-carnitine and25mM malate permitted revealing oxygen consumptionfrom 120573-oxidation pathway
For each substrate after 2min state 3 respiration wasinduced by the addition of 03mM ADP Respiratory controlratio (RCR) was calculated as the ratio of the rate of oxygenuptake in the presence of added ADP (state 3) to the rateobserved when added ADP had been completely phospho-rylated to ATP (state 4)
26 Assay of Mitochondrial Respiratory Complex ActivityComplex I activity was determined using decylubiquinone
as electron acceptor and NADH as donor Rat liver mito-chondria (40 120583g of protein) were pretreated first by freeze-thawing the samples two or three times in hypotonicmedium consisting of 25mMKH
2PO4(pH 72) 5mMMgCl
2
followed by a hypotonic shock in H2O to disrupt mito-
chondrial membranes The reaction started after the addi-tion of 50mMTris (pH 80) medium supplemented with5mgmLBSA 08mMNADH as donor 240120583MKCN 4 120583MantimycinA and 50120583Mof the acceptor decylubiquinoneTheoxidation of NADH was measured following the decrease inabsorbance at 340 nm for 3min [20]
Complex II activity assay was performed following thedecrease in absorbance at 600 nm resulting from the reduc-tion of DCPIP as electron acceptor and using succinateas donor Rat liver mitochondria (40 120583g of protein) werepretreated first by freeze-thawing following by an incubationwith succinate 10min at 30∘C in a medium containing10mMKH
2PO4(pH 78) 80 120583M DCPIP as acceptor 4 120583M
rotenone and 02mM ATP The reaction started by additionof 80 120583M decylubiquinone and the activity was measured for5min [20]
Complex III activity was determined by measuring thereduction of oxidized cytochrome 119888 at 550 nm Rat livermitochondria (40 120583g protein) were incubated for 1min at30∘C in a reaction medium containing 50mMKH
2PO4
(pH 72) 001 Tween-20 50mM EDTA 4mMKCN and40mM oxidized cytochrome 119888 The reaction was initiatedby adding the ubiquinol analog decylubiquinol (DBH
2) to
a final concentration of 50 120583M and the rate of cytochrome119888 reduction was calculated from the absorbance increase at550 nm [2]
Complex IV activity assay was performed following thedecrease in absorbance at 550 nm resulting from the oxida-tion of reduced cytochrome 119888 Rat liver mitochondria (20120583gof protein) were incubated in 1mL of isosmotic mediumcontaining 250mM sucrose 10mM KH
2PO4(pH 65) and
10 120583M reduced cytochrome 119888 Cytochrome 119888 solution wasfreshly reduced by adding some crystals of sodiumdithioniteThe addition of 25mM laurylmaltoside permitted permeabi-lization of the outer mitochondrial membrane to cytochrome119888 and the decrease in absorbance at 550 nm was followed for3min [20]
ATP-synthase activity (ATP hydrolysis) was measuredspectrophotometrically by a coupled assay using lactatedehydrogenase and pyruvate kinase as the coupling enzyme[20] For each ATP molecule that is hydrolyzed a moleculeof NADH is oxidized Rat liver mitochondria were incubatedfirst in distilled water for 30 s at 37∘C and then in a mediumcontaining 200 120583L of 50mMTris (pH 80) 5mgmLBSA20mMMgCl
2 50mMKCl 15120583MCCCP 5 120583Mantimycin A
10mMPEP 25mMATP 4 units of lactate dehydrogenaseand pyruvate kinase and 1mMNADH previously incubatedfor 5min at 37∘C Reaction was started by the addition of themedium to mitochondria and monitored for 3min Finallythe addition of 3 120583Moligomycin permitted distinguishing theATPase activity coupled to the respiratory chain [20]
27 Western Blot Analysis The expression of oxidative phos-phorylation system (OXPHOS) proteins and mitochondrial
4 BioMed Research International
Table 2 Body and liver weight serum parameters and liver fat content
Control HF HF+KOBody weight (g) 2458 plusmn 78 2772 plusmn 158lowast 2352 plusmn 252
Liver weight (g100 g body weight) 39 plusmn 06 35 plusmn 05 41 plusmn 03Glycemia (mM) 29 plusmn 01 49 plusmn 07lowast 41 plusmn 04lowast
Insulin (120583UmL) 501 plusmn 75 1428 plusmn 100lowast 777 plusmn 50lowast
HOMA 64 plusmn 01 312 plusmn 03lowast 142 plusmn 01lowast
Serum triglycerides (mgdL) 1373 plusmn 129 1471 plusmn 88 1386 plusmn 92Serum cholesterol (mgdL) 926 plusmn 83 956 plusmn 42 941 plusmn 62Liver triglycerides (mgg liver) 86 plusmn 04 136 plusmn 10lowast 83 plusmn 05
Liver cholesterol (mgg liver) 27 plusmn 04 66 plusmn 04lowast 35 plusmn 05
Each point represents the mean plusmn SD for 5 animals lowast119875 lt 005 versus rats fed a control diet 119875 lt 005 versus rats fed HF diet
carrier proteins AAC (ADPATP carrier) and UCP2 (uncou-pling protein 2) was determined by western blotting analysisPolyacrylamide gel electrophoresis was performed in thepresence of 01 SDS (SDS-PAGE) according to standardprocedures The mitochondrial proteins that have been sepa-rated by SDS-PAGEwere transferred to a nitrocellulosemem-brane For the OXPHOS proteins a cocktail directed againstNDUFB8 (Complex I) SDHB (Complex II) COR2 (ComplexIII) COX1 (Complex IV) and ATP5A (ATP-synthase) wasused at a dilution 1 1times101 and the immunoreaction was car-ried out with monoclonal antibody by chemiluminescenceFor AAC UCP2 and porin (a protein used as a control)detection antisera were used at a dilution of 1 1times103 and theimmunoreaction was carried out with polyclonal antibody bychemiluminescence
28 ATP and Lipid Hydroperoxide (LPO) Levels ATP levelsin liver homogenates were determined by using the luciferaseATP assay kit The concentration of ATP in the samples wascalculated from interpolation of a standard curve preparedwith known amounts of ATP Results were expressed asnmoles of ATP per mg of protein
Lipid peroxidation levels in liver samples were deter-mined using a lipid hydroperoxide assay kit which measuresthe redox reactions with ferrous ions LPO content wasexpressed as LPO per mg of protein
29 Statistical Analysis Experimental data represent themeans plusmn SD The data were analyzed by one-way ANOVAand a Tukey-Kramer post hoc analysis was used to detectsignificant differences between the means at a level of 119875 gt005
3 Results
31 Body and Liver Weight Serum Parameters and Liver FatContent As shown in Table 2 a significant increase in bodyweight was recorded in HF animals after 4 weeks of dietarytreatment in agreement with the high caloric content ofthe diet Interestingly the supplementation of the HF dietwith 25 KO significantly prevented this effect Insteadno statistically significant variations were found in the liverweight of animals fed with the different experimental diets
The blood glucose concentration increased in both HFand HF+KO animals after 4 weeks of dietary administrationin comparison to control rats Such an increase was lessevident in the case of the HF+KO fed animals A massiveincrease in the levels of insulin was revealed in the plasmaof HF animals in comparison to control rats Interestinglythe KO supplementation significantly mitigated this increaseHF rats showed a marked increase in HOMA index demon-strating typical insulin resistance induced by the high-fat dietIn the HF+KO animals HOMA decreased showing reducedinsulin resistance induced by the addition of KO
No significant difference was detected in the serumlipid (triglyceride and cholesterol) levels between the threegroups of animals On the contrary liver lipid content wasdramatically enhanced in HF animals thus reflecting thepresence of hepatic steatosis (Figure 1) The supplementationwith KO of the HF diet reversed this effect thereby keepingthe lipid concentrations similar to those found in controlrats Accordingly the liver histologic examination revealed abeginning of microvesicular fat depositions only in HF rats
32 Pyruvate Dehydrogenase (PDH) Complex CarnitinePalmitoyltransferase (CPT) and Citrate Synthase (CS) Activ-ity Mitochondria play a central role in energy productionSugars and fatty acids undergo glycolysis and mitochon-drial 120573-oxidation respectively to produce acetyl-CoA (Fig-ure 2(a)) PDH complex activity is the major determinantof glucose oxidation in animal cells We found that PDHactivity remained unchanged in the three groups of animalssuggesting that glucose catabolism was not affected by a HFdiet or by KO supplementation (Figure 2(b))
The limiting step for catabolic pathway of fatty acidoxidation is represented by the activity of CPT I which isresponsible for the entry of fatty acids into the mitochondrialmatrix where 120573-oxidation of fatty acids occurs As shown inFigure 2(c) in the HF animals a clear decrease (about 40)in the CPT I activity was detected On the contrary a strongincrease in CPT I activity was found in theHF+KO ratsThusKO supplemented to theHF diet strongly stimulates fatty acidoxidation
Acetyl-CoA generated essentially from glucose or fromfatty acids is channeled into the Krebs cycle where CS(the rate-limiting enzyme of the Krebs cycle) catalyzes its
BioMed Research International 5
Control HF HF+KO
Figure 1 Liver samples were stored in neutral formaldehyde and embedded in paraffin wax Sections (15120583m) were stained with hematoxylinand eosin
Glucose
Fatty acids
CPT I
Pyruvate
NAD+
120573-oxidationAcetyl-CoA Acyl-CoA
CSCO2
PDH
Oxaloacetate
Krebscycle
FADH2
Mitochondrion
Cytosol
NADH + H+
NADH + H+ NADH + H+
NADH + H+
(a)
0
5
10
15
20
Control HF
PDH
activ
ity (n
molmiddotm
inminus1middotm
g pr
otei
nminus1)
HF+KO
(b)
Control HF0
25
50
75
100
125
150
CPT
I act
ivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(c)
Control HF0
50
100
150
200
250
300
CS ac
tivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(d)
Figure 2Mitochondrialmetabolic pathways fromglucose and fatty acid oxidation Schematic picture of themetabolic pathways fromglucoseand fatty acid oxidation (a) Effect of KO on the activity of mitochondrial enzymes PDH (b) CPT I (c) and CS (d) in control HF or HF+KO-fed rats liver The values reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed a control diet 119875 lt 005 versusrats fed a HF diet
6 BioMed Research International
Table 3 Mitochondrial respiratory efficiency
Control HF HF+KO
Palmitoyl-L-carnitine + malate (120573 oxidation)1198813(nmol O2sdotmLminus1sdotminminus1) 858 plusmn 78 597 plusmn 85lowast 1140 plusmn 112lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 143 plusmn 24 229 plusmn 15lowast 185 plusmn 52lowast
RCR 60 plusmn 04 26 plusmn 03lowast 61 plusmn 08
Pyruvate + malate (Complex I)1198813(nmol O2sdotmLminus1sdotminminus1) 170 plusmn 03 159 plusmn 04lowast 199 plusmn 09lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 54 plusmn 01 76 plusmn 02lowast 66 plusmn 06lowast
RCR 32 plusmn 03 21 plusmn 03lowast 30 plusmn 04
Succinate + rotenone (Complex II)1198813(nmol O2sdotmLminus1sdotminminus1) 615 plusmn 33 531 plusmn 37lowast 783 plusmn 40lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 101 plusmn 03 250 plusmn 12lowast 150 plusmn 19lowast
RCR 61 plusmn 09 21 plusmn 08lowast 52 plusmn 10
Ascorbate + TMPD + rotenone + antimycin A (Complex IV)1198813(nmol O2sdotmLminus1sdotminminus1) 551 plusmn 53 475 plusmn 65lowast 686 plusmn 78lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 93 plusmn 07 172 plusmn 09lowast 128 plusmn 08lowast
RCR 59 plusmn 06 28 plusmn 03lowast 54 plusmn 09
Respiratory control ratio (RCR) was calculated as the ratio of the rate of oxygen uptake in the presence of added ADP (1198813) to the rate observed when added
ADP had been completely phosphorylated to ATP (1198814)
lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
condensation with oxaloacetate to synthesize citrate (Fig-ure 2(a)) A significant decrease (about 55) in the activityof the CS was found in HF animals (Figure 2(d)) whileonly a moderate decrease (about 35) was observed inHF+KO animals in comparison to the control groups Thepartial reversal of CS inhibition observed in HF animals afterKO supplementation could be explained by a compensatoryresponse of this enzyme to sustain oxidation of acetyl-CoAgenerated from fatty acid oxidation
33 Mitochondrial Oxygen Consumption Respiratory ChainComplex Activities and ATP Synthesis The increased fattyacid oxidation observed in HF+KO animals produces higherlevels of reducing equivalents which are normally addressedtowards the mitochondrial oxidative phosphorylation forATP production We therefore analyzed the respiratoryefficiency of freshly isolated mitochondria by oxygraphicmethods Mitochondrial oxygen consumption was measuredwith various substrates in the presence (state 3) or the absence(state 4) of ADP (Table 3) According to the results shownin Figure 2(c) when we used palmitoyl-L-carnitine andmalate as substrates 119881
3(mitochondrial respiration state 3)
decreased inHF animals thus indicating lowermitochondrial120573-oxidation On the other hand the increase in the119881
4(mito-
chondrial respiration state 4) values suggests that the excess ofdietary fat most probably is the cause of a partial uncouplingbetween respiration and phosphorylation in mitochondriaInterestingly the dietary KO supplementation is able toefficiently abolish this effect keeping the mitochondrialrespiratory efficiency unaltered
When we added respiratory substrates for mitochondrialcomplexes I II and IV (Table 3) a little yet significantdecrease in 119881
3was observed in the HF groups in comparison
to control animals In the HF+KO animals a significantincrease in the 119881
3values was found in comparison to HF
and control animals 1198814 also known as the resting state of
respiration showed an increase in theHF andHF+KO groupeven if the increase observed after KO supplementation was
less evident than that found inHF animals As a consequencethe RCRvalueswere profoundly and significantly lower inHFanimals with respect to those calculated for control animalsThe KO supplementation of the HF diet reversed this effectthereby keeping RCR values similar to those calculated forcontrol group
In a more selective approach for investigating the func-tionality of the mitochondrial oxidative phosphorylation weassayed the activity of single components of the respiratorychain complexes IndashIV andATP-synthase (Figure 3(a)) A sig-nificant decrease (about 30) in the activity of all respiratorychain complexes was found in HF animals On the contrarythe values of complexes activity of the HF+KO rats werepractically identical to those measured in control animals
Surprisingly western blotting analysis of the OXPHOSproteins showed significantly higher protein levels in bothHFand HF+KO animals compared to the control group (Figures3(b) and 3(c)) Such an increase was more evident for com-plexes I III and IV which are associated as supercomplexesin the mitochondrial inner membrane [21] This effect couldbe determined by a compensatory mechanism related to thereduction of the activities of the respiratory complexes
In order to prove biological relevance of alteration inmitochondrial respiratory chain activity we measured ATPconcentration and oxidative damage in liver samples fromthe three groups of animals (Figure 3(d)) ATP quantificationdemonstrated that aHFdiet caused a slight but not significantdecrease in ATP content an effect that was rescued by KOInterestingly in parallel experiments we found a significantincrease (about 30) in the LPO levels inHF animals and thiseffect was efficiently reversed by KO supplementation of theHF diet
34 Expression of Proteins Related to the Respiratory ChainFunction Uncoupling protein 2 (UCP2) uncouples respira-tion from oxidative phosphorylation We therefore analyzedthe expression profile of this carrier protein in the threegroups of animals (Figure 4(a)) After 4 weeks of HF dietary
BioMed Research International 7
0
30
60
90
120
Con
trol
HF
Resp
irato
ry co
mpl
exes
activ
ity (
)
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
lowast lowast lowastlowast
lowast
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(a)
CV-ATP5A
CIII-COR2CIV-COX1
CII-SDHB
CI-NDUFB8
Porin
(b)
0
50
100
150
250
OXP
HO
S pr
otei
ns (
) 200lowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
Con
trol
HF
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(c)
30
90
0
60
150
( o
f con
trol)
120
ATP LPO
lowast
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
(d)
Figure 3 Effect of KO on respiratory complexes activity expression and function Effect of KO on respiratory complexes activity (a)Enzymatic activities measured in the control group were set to 100 Effect of KO on OXPHOS proteins levels (b) Liver mitochondrialproteins from control HF or HF+KO-fed rats were separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with acocktail against NDUFB8 SDHB COR2 COX1 ATP5A or the mammalian porin as a controlThe amount of OXPHOS proteins revealed byimmunodecoration in the control group was set to 100 (c) ATP and LPO quantification of liver samples isolated from animals fed a controldiet a HF diet or a HF+KO (d) The amount of ATP or LPO (expressed as nmoles per mg protein) in the control group was set to 100Thevalues reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
treatment the amount of this protein increased about 5-foldcompared to the control group Interestingly a significantsmaller increase in the expression of UCP2 was found afterKO supplementation In fact in the presence of KO in theHF diet the level of this carrier protein was about 50 of thatfound in HF rats
The AAC (ADPATP carrier) is another mitochondrialcarrier which is able to regulate the basal proton conductanceThis protein exchanges ADP for ATP across the mitochon-drial inner membrane and may also play an important rolein the mitochondrial permeability transition pore When weanalyzed the expression of this carrier protein (Figure 4(b))we found an increase of about 22-fold in HF animalscompared to the control group AAC levels in the HF+KOanimals even if higher than those found in the controlrats were significantly lower than those found in HF group
The amount of porin an outer membrane protein tested as acontrol did not change in any group of animals (Figure 4(c))
These results suggest that the KO supplementation is ableto modulate the expression of mitochondrial carrier proteininvolved in the regulation of proton conductance reducingthe uncoupling effect caused by a HF diet
4 Discussions
In animal experiments a diet rich in fat or carbohydrates hasbeen shown to induce hyperglycemia and hyperinsulinemiaand to contribute to the development of obesity and NAFLD[2 6 7 22] This last condition is recognized as the mostcommon chronic liver disease in the Western world NASHis a more severe form of NAFLD that is broadly defined bythe presence of steatosis with inflammation and progressive
8 BioMed Research International
0
100
200
300
500
Control HF
UCP
2 (
)
UCP2
400
lowast
lowast
HF+KO
(a)
0
60
120
180
300
AAC
()
AAC
240
Control HF
lowast
lowast
HF+KO
(b)
0
25
50
75
125
Porin
()
Porin
100
Control HF HF+KO
(c)
Figure 4 Effect of KO on proteins related to the respiratory chain function Effect of KO on proteins levels of UCP2 (a) and AAC (b) Theamount of porin an outer membrane protein was tested as a control (c) Liver mitochondrial proteins from control HF or HF+KO-fed ratswere separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with antisera against UCP2 AAC or the mammalianporinThe amount of proteins revealed by immunodecoration in the control groupwas set to 100The values reported in the figure representthe means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
fibrosis leading then to cirrhosis and hepatocellular carci-noma Some patients with NAFLD develop NASH throughpoorly understood mechanisms Since NAFLD and NASHare considered lifestyle-associated diseases diet interventionis an important approach to their treatment
In this context it is important to underline that theldquoobesogenicrdquoWestern diet has a ratio of fatty acids and carbo-hydrates of approximately 5050Moreover it is characterizedby an insufficient n-3 PUFAs consumption and a high n-6n-3 ratio which are associated with the progression fromsimple steatosis to NASH [8] along with modifications of gutmicrobiota [23]
Although the mechanisms responsible for increasedlipid accumulation in the liver are not yet fully elucidateddecreased capacity to oxidize fatty acids increased transportof fatty acids to liver and increased hepatic fatty acidsynthesis seem to be important factors involved in thepathogenesis of NAFLD [2 6 7 24]Moreover abnormalitiesin mitochondrial morphology and functions seem to beassociated with fatty liver [1ndash3 25ndash27] This finding suggeststhat regulation of mitochondria function and of molecularmechanisms involved in lipid accumulation in the liver couldprovide useful targets to prevent the development of NAFLD
In recent years the use of dietary supplements hasrapidly increased [2 6 10 24 28] with the aim to preventNAFLD KO a dietary supplement of n-3 PUFAs has becomeincreasingly popular because of its capability of reducing
hepatic lipogenesis and of stimulating catabolization ofexcess fat introduced by a HF diet [2 10] The analysisof hepatic transcriptome in mice demonstrated that KOsupplementation was able to regulate genes involved in hep-atic energy metabolism including glucose metabolism lipidbiosynthesis fatty acid metabolism cholesterol biosynthesisand mitochondrial electron transport more efficiently thanfish oil [10 12] These differential results were tentativelyascribed to the influence of the structural form of n-3PUFAs esterified to either phospholipids or triglycerideson a biological response Moreover PUFAs as phospholipids(KO) or triglycerides (fish oil)may differentially influence gutmicrobiota or synthesis of bacterial metabolites
A previous study by our group [2] suggested that KOsupplementation to a HF diet resulted in a retention of nor-malmitochondrial respiration efficiency whichwas impairedin HF animals The aim of the present study was thereforeto analyze the molecular mechanisms by which dietary fatsinfluence mitochondrial energetic metabolismThis new andintriguing aspect was evaluated in hepatocytes from rats feda HF diet or a HF diet supplemented with 25 KO with theaim to investigate the role of mitochondria in themodulationof nutrient metabolism exerted by KO
No change in PDH activity which essentially producesacetyl-CoA from carbohydrate was seen after administrationof HF or HF+KO diets Nevertheless fasting serum glucoseand insulin were higher after feeding a HF diet suggesting
BioMed Research International 9
an altered insulin sensitivity as indicated by HOMA indexchanges In fact an excess of energy supplied by fat corre-sponds to a scarce tissutal utilization of glucose thereby caus-ing hyperglycemia and hyperinsulinemia Insulin resistancewas reduced after KO supplementation
CPT I activity which is involved in fatty acid oxidationwas inhibited by a HF diet and stimulated after KO supple-mentationThe increase inCPT I activity observed inHF+KOgroup could be explained by the lower levels ofmalonyl-CoAametabolic intermediate of hepatic lipogenesis which in thiscase is instead strongly inhibited [2 10] Interestingly thecapability of KO to stimulate fatty acid oxidation seems tobe associated to the presence of a high lipid content (35)in the experimental diet used in this study In fact whenKO was added to a standard diet the expression of enzymesinvolved inmitochondrial fatty acid 120573-oxidation significantlydecreased [12]
The modulation of the CPT I activity observed in HFand HF+KO animals seems to be accompanied by a parallelmodulation of the activity of CS a Krebs cycle enzyme thatcatalyzes the synthesis of citrate fromoxaloacetate and acetyl-CoA Recently a strong and significant correlation of CSwith obesity was noted with a reduced enzyme activity inmitochondria of human omental adipose tissue in the stateof obesity [28] According to this observation the increasein CS activity revealed after KO supplementation could beexplained as a compensatory response to maintain the entryof acetyl-CoA produced by fatty acid oxidation into the Krebscycle It is interesting to underline that citrate generated byCS continues in the Krebs cycle In fact in animals fed aHF diet supplemented with KO a reduction in the activityand in the expression of citrate carrier protein (CIC) whichtransports citrate from mitochondria to the cytosol for fattyacid synthesis was observed [2 10 24]
NADH and FADH2generated by 120573-oxidation and Krebs
cycle are used in the final common oxidative phosphorylationsystem (OXPHOS) to generate ATP This process is coupledwith the transfer of electrons along the respiratory chainOur data indicate that a HF diet caused a strong decrease inthe mitochondrial respiratory efficiency and this effect couldbe due to a possible uncoupling effect exerted by the excessof fatty acids present in this diet In fact we found a littledecrease in 119881
3and a strong increase in 119881
4values in liver
mitochondria isolated fromHF animals1198814is the resting state
of mitochondrial respiration and an increase in this valuesuggests a stimulus ofmitochondrial respiration independentof ADP phosphorylation In other terms during HF feedinga lower coupling betweenmitochondrial respiration andATPsynthesis occurs Interestingly the addition of KO to the HFdiet almost completely reversed this effect thereby leadingmitochondria respiration efficiency to be comparable to thatof control rats
When we assayed in a more selective approach theactivity of single components of the respiratory chain weobserved a significant decrease in the activity of all respira-tory chain complexes in HF animals according to a recentstudy [29] inwhich an impairment ofmitochondrial functionand in particular of complex I activity was found AfterKO supplementation the values of complex activities were
identical to those measured in control animals Expressionof OXPHOS proteins significantly increased in HF animalscompared to the control group and this evidence was accom-panied by the presence of ATP levels in HF animals thatwere only slightly decreased in comparison to those foundin control animals According to this result previous studies[30 31] demonstrated that a high fat content in the dietinduced higher level of these proteins as a compensatoryresponse to the reduced activity of the respiratory chaincomplexes When we performed western blotting analysis inmitochondria isolated from HF+KO animals liver we founda higher level of the OXPHOS proteins compared to thecontrol and theHF groups probably caused by the synergisticeffect of the HF diet and the KO supplementation Indeedit is already known that genes involved in mitochondrialOXPHOS proteins expression were upregulated in miceconsuming KO as revealed from the transcriptomic analysis[12] Overall thismeans that KO is able to induce the burningof excess of fat introduced by diet stimulating fatty acidoxidation Krebs cycle and respiratory chain complex activityand preventing a condition of oxidative damage
Uncoupling between mitochondrial respiration and ATPsynthesis observed in HF animals could be caused bya different expression of mitochondrial carrier proteinsinvolved in the regulation of proton conductance It is knownthat uncoupling proteins (UCPs) homologues uncouplemitochondrial respiration from oxidative phosphorylationincreasing thermogenesis while reducing the efficiency ofATP synthesis In a previous study [32] an increase in theexpression of the UCP2 isoform was found in rats withNAFLD Interestingly in this study we found a massiveincrease of this carrier protein inHF animals but the additionof KO to the diet was able to strongly reduce the expressionof hepatic UCP2 Therefore it is reasonable to assume thatthe addition of KO to the HF diet can increase the couplingbetween mitochondrial respiration and ATP synthesis by adownregulation of UCP2 expression
Not only are UCPs involved in mitochondrial uncou-pling butAAC is another carrier protein of particular interestin the understanding of the mitochondrial energy couplingprocess [33]This carrier catalyzes the exchange of ADP3minus forATP4minus and therefore utilizes as its driving force the electro-chemical potential generated across the inner membrane bythe respiratory chain The exchange reaction is electrogenicand driven by the electrical component of the proton gradi-ent so that entry of ADP into mitochondria and exit of ATPare favored We can therefore speculate that if mitochondriaare uncoupled and proton gradient was dissipated by theentry of H+ into the matrix by UCP2 an upregulation ofthe expression of AAC could compensate a reduced fluxof ATP from mitochondria when membrane potential wasreduced This hypothesized compensatory mechanism wasobserved in animals fed a HF diet The increase in theexpression of AAC was less evident in HF+KO animals inwhich livermitochondria respiratory efficiencywas unalteredin comparison to the control animals
In conclusion the present study indicates that KO coun-teracts the negative effects of a HF diet In particular our
10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 3
Table 1 Composition of diets ()
Control HF HF+KOProteins 186 204 199Lipids 62 352 368
Fatty acids140 mdash 05 06160 07 87 90180 02 43 43161 mdash mdash 01181 (n9-7-5) 12 158 160182 (n6) 31 35 35183 (n3) 03 mdash mdash205 (n3) EPA mdash mdash 03226 (n3) DHA mdash mdash 02sum SFA 09 135 139sumMUFA 13 158 161sumPUFA 34 35 41sumPUFA n-3 03 mdash 06
Carbohydrates 442 361 352kcal100 g 310 540 549Calories from protein () 240 150 140Calories from fat () 180 580 600Calories from carbohydrates () 580 270 260The control group of animals received a standard diet (Global Diet 2018Sfrom Harlan Teklad) The HF group received a diet with 35 fat (DietTD03584 from Harlan Teklad) and the KO group was fed with the abovereported HF diet supplemented with 25 (ww) KO Fatty acids wereextracted from the three diets and analyzed by gas-liquid chromatographyEnergy content was calculated using 4 kcalg for protein and carbohydrateand 9 kcalg for lipid
412 nm in freshly isolated rat liver mitochondria essentiallyas described previously [18]
25 Mitochondrial Respiration Efficiency Mitochondrial res-piration (03mg ofmitochondrial proteinmL) wasmeasuredin a medium consisting of 220mM sucrose 20mMKCl25mMKH
2PO4 1 mMEDTA 20mMHepes 5mMMgCl
2
and 01 BSA pH 74 by a Clark oxygen electrode at 25∘C[2] The addition of different substrates permitted to evalu-ate mitochondrial respiration when respiratory complexes I(5mM pyruvate 25mM malate) II (5mM K-succinate5 120583M rotenone) and IV (10mM ascorbate 02mM TMPD5 120583M rotenone 5 120583M antimycin A) were stimulated [19 20]Instead the addition of 40 120583M palmitoyl-L-carnitine and25mM malate permitted revealing oxygen consumptionfrom 120573-oxidation pathway
For each substrate after 2min state 3 respiration wasinduced by the addition of 03mM ADP Respiratory controlratio (RCR) was calculated as the ratio of the rate of oxygenuptake in the presence of added ADP (state 3) to the rateobserved when added ADP had been completely phospho-rylated to ATP (state 4)
26 Assay of Mitochondrial Respiratory Complex ActivityComplex I activity was determined using decylubiquinone
as electron acceptor and NADH as donor Rat liver mito-chondria (40 120583g of protein) were pretreated first by freeze-thawing the samples two or three times in hypotonicmedium consisting of 25mMKH
2PO4(pH 72) 5mMMgCl
2
followed by a hypotonic shock in H2O to disrupt mito-
chondrial membranes The reaction started after the addi-tion of 50mMTris (pH 80) medium supplemented with5mgmLBSA 08mMNADH as donor 240120583MKCN 4 120583MantimycinA and 50120583Mof the acceptor decylubiquinoneTheoxidation of NADH was measured following the decrease inabsorbance at 340 nm for 3min [20]
Complex II activity assay was performed following thedecrease in absorbance at 600 nm resulting from the reduc-tion of DCPIP as electron acceptor and using succinateas donor Rat liver mitochondria (40 120583g of protein) werepretreated first by freeze-thawing following by an incubationwith succinate 10min at 30∘C in a medium containing10mMKH
2PO4(pH 78) 80 120583M DCPIP as acceptor 4 120583M
rotenone and 02mM ATP The reaction started by additionof 80 120583M decylubiquinone and the activity was measured for5min [20]
Complex III activity was determined by measuring thereduction of oxidized cytochrome 119888 at 550 nm Rat livermitochondria (40 120583g protein) were incubated for 1min at30∘C in a reaction medium containing 50mMKH
2PO4
(pH 72) 001 Tween-20 50mM EDTA 4mMKCN and40mM oxidized cytochrome 119888 The reaction was initiatedby adding the ubiquinol analog decylubiquinol (DBH
2) to
a final concentration of 50 120583M and the rate of cytochrome119888 reduction was calculated from the absorbance increase at550 nm [2]
Complex IV activity assay was performed following thedecrease in absorbance at 550 nm resulting from the oxida-tion of reduced cytochrome 119888 Rat liver mitochondria (20120583gof protein) were incubated in 1mL of isosmotic mediumcontaining 250mM sucrose 10mM KH
2PO4(pH 65) and
10 120583M reduced cytochrome 119888 Cytochrome 119888 solution wasfreshly reduced by adding some crystals of sodiumdithioniteThe addition of 25mM laurylmaltoside permitted permeabi-lization of the outer mitochondrial membrane to cytochrome119888 and the decrease in absorbance at 550 nm was followed for3min [20]
ATP-synthase activity (ATP hydrolysis) was measuredspectrophotometrically by a coupled assay using lactatedehydrogenase and pyruvate kinase as the coupling enzyme[20] For each ATP molecule that is hydrolyzed a moleculeof NADH is oxidized Rat liver mitochondria were incubatedfirst in distilled water for 30 s at 37∘C and then in a mediumcontaining 200 120583L of 50mMTris (pH 80) 5mgmLBSA20mMMgCl
2 50mMKCl 15120583MCCCP 5 120583Mantimycin A
10mMPEP 25mMATP 4 units of lactate dehydrogenaseand pyruvate kinase and 1mMNADH previously incubatedfor 5min at 37∘C Reaction was started by the addition of themedium to mitochondria and monitored for 3min Finallythe addition of 3 120583Moligomycin permitted distinguishing theATPase activity coupled to the respiratory chain [20]
27 Western Blot Analysis The expression of oxidative phos-phorylation system (OXPHOS) proteins and mitochondrial
4 BioMed Research International
Table 2 Body and liver weight serum parameters and liver fat content
Control HF HF+KOBody weight (g) 2458 plusmn 78 2772 plusmn 158lowast 2352 plusmn 252
Liver weight (g100 g body weight) 39 plusmn 06 35 plusmn 05 41 plusmn 03Glycemia (mM) 29 plusmn 01 49 plusmn 07lowast 41 plusmn 04lowast
Insulin (120583UmL) 501 plusmn 75 1428 plusmn 100lowast 777 plusmn 50lowast
HOMA 64 plusmn 01 312 plusmn 03lowast 142 plusmn 01lowast
Serum triglycerides (mgdL) 1373 plusmn 129 1471 plusmn 88 1386 plusmn 92Serum cholesterol (mgdL) 926 plusmn 83 956 plusmn 42 941 plusmn 62Liver triglycerides (mgg liver) 86 plusmn 04 136 plusmn 10lowast 83 plusmn 05
Liver cholesterol (mgg liver) 27 plusmn 04 66 plusmn 04lowast 35 plusmn 05
Each point represents the mean plusmn SD for 5 animals lowast119875 lt 005 versus rats fed a control diet 119875 lt 005 versus rats fed HF diet
carrier proteins AAC (ADPATP carrier) and UCP2 (uncou-pling protein 2) was determined by western blotting analysisPolyacrylamide gel electrophoresis was performed in thepresence of 01 SDS (SDS-PAGE) according to standardprocedures The mitochondrial proteins that have been sepa-rated by SDS-PAGEwere transferred to a nitrocellulosemem-brane For the OXPHOS proteins a cocktail directed againstNDUFB8 (Complex I) SDHB (Complex II) COR2 (ComplexIII) COX1 (Complex IV) and ATP5A (ATP-synthase) wasused at a dilution 1 1times101 and the immunoreaction was car-ried out with monoclonal antibody by chemiluminescenceFor AAC UCP2 and porin (a protein used as a control)detection antisera were used at a dilution of 1 1times103 and theimmunoreaction was carried out with polyclonal antibody bychemiluminescence
28 ATP and Lipid Hydroperoxide (LPO) Levels ATP levelsin liver homogenates were determined by using the luciferaseATP assay kit The concentration of ATP in the samples wascalculated from interpolation of a standard curve preparedwith known amounts of ATP Results were expressed asnmoles of ATP per mg of protein
Lipid peroxidation levels in liver samples were deter-mined using a lipid hydroperoxide assay kit which measuresthe redox reactions with ferrous ions LPO content wasexpressed as LPO per mg of protein
29 Statistical Analysis Experimental data represent themeans plusmn SD The data were analyzed by one-way ANOVAand a Tukey-Kramer post hoc analysis was used to detectsignificant differences between the means at a level of 119875 gt005
3 Results
31 Body and Liver Weight Serum Parameters and Liver FatContent As shown in Table 2 a significant increase in bodyweight was recorded in HF animals after 4 weeks of dietarytreatment in agreement with the high caloric content ofthe diet Interestingly the supplementation of the HF dietwith 25 KO significantly prevented this effect Insteadno statistically significant variations were found in the liverweight of animals fed with the different experimental diets
The blood glucose concentration increased in both HFand HF+KO animals after 4 weeks of dietary administrationin comparison to control rats Such an increase was lessevident in the case of the HF+KO fed animals A massiveincrease in the levels of insulin was revealed in the plasmaof HF animals in comparison to control rats Interestinglythe KO supplementation significantly mitigated this increaseHF rats showed a marked increase in HOMA index demon-strating typical insulin resistance induced by the high-fat dietIn the HF+KO animals HOMA decreased showing reducedinsulin resistance induced by the addition of KO
No significant difference was detected in the serumlipid (triglyceride and cholesterol) levels between the threegroups of animals On the contrary liver lipid content wasdramatically enhanced in HF animals thus reflecting thepresence of hepatic steatosis (Figure 1) The supplementationwith KO of the HF diet reversed this effect thereby keepingthe lipid concentrations similar to those found in controlrats Accordingly the liver histologic examination revealed abeginning of microvesicular fat depositions only in HF rats
32 Pyruvate Dehydrogenase (PDH) Complex CarnitinePalmitoyltransferase (CPT) and Citrate Synthase (CS) Activ-ity Mitochondria play a central role in energy productionSugars and fatty acids undergo glycolysis and mitochon-drial 120573-oxidation respectively to produce acetyl-CoA (Fig-ure 2(a)) PDH complex activity is the major determinantof glucose oxidation in animal cells We found that PDHactivity remained unchanged in the three groups of animalssuggesting that glucose catabolism was not affected by a HFdiet or by KO supplementation (Figure 2(b))
The limiting step for catabolic pathway of fatty acidoxidation is represented by the activity of CPT I which isresponsible for the entry of fatty acids into the mitochondrialmatrix where 120573-oxidation of fatty acids occurs As shown inFigure 2(c) in the HF animals a clear decrease (about 40)in the CPT I activity was detected On the contrary a strongincrease in CPT I activity was found in theHF+KO ratsThusKO supplemented to theHF diet strongly stimulates fatty acidoxidation
Acetyl-CoA generated essentially from glucose or fromfatty acids is channeled into the Krebs cycle where CS(the rate-limiting enzyme of the Krebs cycle) catalyzes its
BioMed Research International 5
Control HF HF+KO
Figure 1 Liver samples were stored in neutral formaldehyde and embedded in paraffin wax Sections (15120583m) were stained with hematoxylinand eosin
Glucose
Fatty acids
CPT I
Pyruvate
NAD+
120573-oxidationAcetyl-CoA Acyl-CoA
CSCO2
PDH
Oxaloacetate
Krebscycle
FADH2
Mitochondrion
Cytosol
NADH + H+
NADH + H+ NADH + H+
NADH + H+
(a)
0
5
10
15
20
Control HF
PDH
activ
ity (n
molmiddotm
inminus1middotm
g pr
otei
nminus1)
HF+KO
(b)
Control HF0
25
50
75
100
125
150
CPT
I act
ivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(c)
Control HF0
50
100
150
200
250
300
CS ac
tivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(d)
Figure 2Mitochondrialmetabolic pathways fromglucose and fatty acid oxidation Schematic picture of themetabolic pathways fromglucoseand fatty acid oxidation (a) Effect of KO on the activity of mitochondrial enzymes PDH (b) CPT I (c) and CS (d) in control HF or HF+KO-fed rats liver The values reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed a control diet 119875 lt 005 versusrats fed a HF diet
6 BioMed Research International
Table 3 Mitochondrial respiratory efficiency
Control HF HF+KO
Palmitoyl-L-carnitine + malate (120573 oxidation)1198813(nmol O2sdotmLminus1sdotminminus1) 858 plusmn 78 597 plusmn 85lowast 1140 plusmn 112lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 143 plusmn 24 229 plusmn 15lowast 185 plusmn 52lowast
RCR 60 plusmn 04 26 plusmn 03lowast 61 plusmn 08
Pyruvate + malate (Complex I)1198813(nmol O2sdotmLminus1sdotminminus1) 170 plusmn 03 159 plusmn 04lowast 199 plusmn 09lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 54 plusmn 01 76 plusmn 02lowast 66 plusmn 06lowast
RCR 32 plusmn 03 21 plusmn 03lowast 30 plusmn 04
Succinate + rotenone (Complex II)1198813(nmol O2sdotmLminus1sdotminminus1) 615 plusmn 33 531 plusmn 37lowast 783 plusmn 40lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 101 plusmn 03 250 plusmn 12lowast 150 plusmn 19lowast
RCR 61 plusmn 09 21 plusmn 08lowast 52 plusmn 10
Ascorbate + TMPD + rotenone + antimycin A (Complex IV)1198813(nmol O2sdotmLminus1sdotminminus1) 551 plusmn 53 475 plusmn 65lowast 686 plusmn 78lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 93 plusmn 07 172 plusmn 09lowast 128 plusmn 08lowast
RCR 59 plusmn 06 28 plusmn 03lowast 54 plusmn 09
Respiratory control ratio (RCR) was calculated as the ratio of the rate of oxygen uptake in the presence of added ADP (1198813) to the rate observed when added
ADP had been completely phosphorylated to ATP (1198814)
lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
condensation with oxaloacetate to synthesize citrate (Fig-ure 2(a)) A significant decrease (about 55) in the activityof the CS was found in HF animals (Figure 2(d)) whileonly a moderate decrease (about 35) was observed inHF+KO animals in comparison to the control groups Thepartial reversal of CS inhibition observed in HF animals afterKO supplementation could be explained by a compensatoryresponse of this enzyme to sustain oxidation of acetyl-CoAgenerated from fatty acid oxidation
33 Mitochondrial Oxygen Consumption Respiratory ChainComplex Activities and ATP Synthesis The increased fattyacid oxidation observed in HF+KO animals produces higherlevels of reducing equivalents which are normally addressedtowards the mitochondrial oxidative phosphorylation forATP production We therefore analyzed the respiratoryefficiency of freshly isolated mitochondria by oxygraphicmethods Mitochondrial oxygen consumption was measuredwith various substrates in the presence (state 3) or the absence(state 4) of ADP (Table 3) According to the results shownin Figure 2(c) when we used palmitoyl-L-carnitine andmalate as substrates 119881
3(mitochondrial respiration state 3)
decreased inHF animals thus indicating lowermitochondrial120573-oxidation On the other hand the increase in the119881
4(mito-
chondrial respiration state 4) values suggests that the excess ofdietary fat most probably is the cause of a partial uncouplingbetween respiration and phosphorylation in mitochondriaInterestingly the dietary KO supplementation is able toefficiently abolish this effect keeping the mitochondrialrespiratory efficiency unaltered
When we added respiratory substrates for mitochondrialcomplexes I II and IV (Table 3) a little yet significantdecrease in 119881
3was observed in the HF groups in comparison
to control animals In the HF+KO animals a significantincrease in the 119881
3values was found in comparison to HF
and control animals 1198814 also known as the resting state of
respiration showed an increase in theHF andHF+KO groupeven if the increase observed after KO supplementation was
less evident than that found inHF animals As a consequencethe RCRvalueswere profoundly and significantly lower inHFanimals with respect to those calculated for control animalsThe KO supplementation of the HF diet reversed this effectthereby keeping RCR values similar to those calculated forcontrol group
In a more selective approach for investigating the func-tionality of the mitochondrial oxidative phosphorylation weassayed the activity of single components of the respiratorychain complexes IndashIV andATP-synthase (Figure 3(a)) A sig-nificant decrease (about 30) in the activity of all respiratorychain complexes was found in HF animals On the contrarythe values of complexes activity of the HF+KO rats werepractically identical to those measured in control animals
Surprisingly western blotting analysis of the OXPHOSproteins showed significantly higher protein levels in bothHFand HF+KO animals compared to the control group (Figures3(b) and 3(c)) Such an increase was more evident for com-plexes I III and IV which are associated as supercomplexesin the mitochondrial inner membrane [21] This effect couldbe determined by a compensatory mechanism related to thereduction of the activities of the respiratory complexes
In order to prove biological relevance of alteration inmitochondrial respiratory chain activity we measured ATPconcentration and oxidative damage in liver samples fromthe three groups of animals (Figure 3(d)) ATP quantificationdemonstrated that aHFdiet caused a slight but not significantdecrease in ATP content an effect that was rescued by KOInterestingly in parallel experiments we found a significantincrease (about 30) in the LPO levels inHF animals and thiseffect was efficiently reversed by KO supplementation of theHF diet
34 Expression of Proteins Related to the Respiratory ChainFunction Uncoupling protein 2 (UCP2) uncouples respira-tion from oxidative phosphorylation We therefore analyzedthe expression profile of this carrier protein in the threegroups of animals (Figure 4(a)) After 4 weeks of HF dietary
BioMed Research International 7
0
30
60
90
120
Con
trol
HF
Resp
irato
ry co
mpl
exes
activ
ity (
)
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
lowast lowast lowastlowast
lowast
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(a)
CV-ATP5A
CIII-COR2CIV-COX1
CII-SDHB
CI-NDUFB8
Porin
(b)
0
50
100
150
250
OXP
HO
S pr
otei
ns (
) 200lowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
Con
trol
HF
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(c)
30
90
0
60
150
( o
f con
trol)
120
ATP LPO
lowast
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
(d)
Figure 3 Effect of KO on respiratory complexes activity expression and function Effect of KO on respiratory complexes activity (a)Enzymatic activities measured in the control group were set to 100 Effect of KO on OXPHOS proteins levels (b) Liver mitochondrialproteins from control HF or HF+KO-fed rats were separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with acocktail against NDUFB8 SDHB COR2 COX1 ATP5A or the mammalian porin as a controlThe amount of OXPHOS proteins revealed byimmunodecoration in the control group was set to 100 (c) ATP and LPO quantification of liver samples isolated from animals fed a controldiet a HF diet or a HF+KO (d) The amount of ATP or LPO (expressed as nmoles per mg protein) in the control group was set to 100Thevalues reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
treatment the amount of this protein increased about 5-foldcompared to the control group Interestingly a significantsmaller increase in the expression of UCP2 was found afterKO supplementation In fact in the presence of KO in theHF diet the level of this carrier protein was about 50 of thatfound in HF rats
The AAC (ADPATP carrier) is another mitochondrialcarrier which is able to regulate the basal proton conductanceThis protein exchanges ADP for ATP across the mitochon-drial inner membrane and may also play an important rolein the mitochondrial permeability transition pore When weanalyzed the expression of this carrier protein (Figure 4(b))we found an increase of about 22-fold in HF animalscompared to the control group AAC levels in the HF+KOanimals even if higher than those found in the controlrats were significantly lower than those found in HF group
The amount of porin an outer membrane protein tested as acontrol did not change in any group of animals (Figure 4(c))
These results suggest that the KO supplementation is ableto modulate the expression of mitochondrial carrier proteininvolved in the regulation of proton conductance reducingthe uncoupling effect caused by a HF diet
4 Discussions
In animal experiments a diet rich in fat or carbohydrates hasbeen shown to induce hyperglycemia and hyperinsulinemiaand to contribute to the development of obesity and NAFLD[2 6 7 22] This last condition is recognized as the mostcommon chronic liver disease in the Western world NASHis a more severe form of NAFLD that is broadly defined bythe presence of steatosis with inflammation and progressive
8 BioMed Research International
0
100
200
300
500
Control HF
UCP
2 (
)
UCP2
400
lowast
lowast
HF+KO
(a)
0
60
120
180
300
AAC
()
AAC
240
Control HF
lowast
lowast
HF+KO
(b)
0
25
50
75
125
Porin
()
Porin
100
Control HF HF+KO
(c)
Figure 4 Effect of KO on proteins related to the respiratory chain function Effect of KO on proteins levels of UCP2 (a) and AAC (b) Theamount of porin an outer membrane protein was tested as a control (c) Liver mitochondrial proteins from control HF or HF+KO-fed ratswere separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with antisera against UCP2 AAC or the mammalianporinThe amount of proteins revealed by immunodecoration in the control groupwas set to 100The values reported in the figure representthe means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
fibrosis leading then to cirrhosis and hepatocellular carci-noma Some patients with NAFLD develop NASH throughpoorly understood mechanisms Since NAFLD and NASHare considered lifestyle-associated diseases diet interventionis an important approach to their treatment
In this context it is important to underline that theldquoobesogenicrdquoWestern diet has a ratio of fatty acids and carbo-hydrates of approximately 5050Moreover it is characterizedby an insufficient n-3 PUFAs consumption and a high n-6n-3 ratio which are associated with the progression fromsimple steatosis to NASH [8] along with modifications of gutmicrobiota [23]
Although the mechanisms responsible for increasedlipid accumulation in the liver are not yet fully elucidateddecreased capacity to oxidize fatty acids increased transportof fatty acids to liver and increased hepatic fatty acidsynthesis seem to be important factors involved in thepathogenesis of NAFLD [2 6 7 24]Moreover abnormalitiesin mitochondrial morphology and functions seem to beassociated with fatty liver [1ndash3 25ndash27] This finding suggeststhat regulation of mitochondria function and of molecularmechanisms involved in lipid accumulation in the liver couldprovide useful targets to prevent the development of NAFLD
In recent years the use of dietary supplements hasrapidly increased [2 6 10 24 28] with the aim to preventNAFLD KO a dietary supplement of n-3 PUFAs has becomeincreasingly popular because of its capability of reducing
hepatic lipogenesis and of stimulating catabolization ofexcess fat introduced by a HF diet [2 10] The analysisof hepatic transcriptome in mice demonstrated that KOsupplementation was able to regulate genes involved in hep-atic energy metabolism including glucose metabolism lipidbiosynthesis fatty acid metabolism cholesterol biosynthesisand mitochondrial electron transport more efficiently thanfish oil [10 12] These differential results were tentativelyascribed to the influence of the structural form of n-3PUFAs esterified to either phospholipids or triglycerideson a biological response Moreover PUFAs as phospholipids(KO) or triglycerides (fish oil)may differentially influence gutmicrobiota or synthesis of bacterial metabolites
A previous study by our group [2] suggested that KOsupplementation to a HF diet resulted in a retention of nor-malmitochondrial respiration efficiency whichwas impairedin HF animals The aim of the present study was thereforeto analyze the molecular mechanisms by which dietary fatsinfluence mitochondrial energetic metabolismThis new andintriguing aspect was evaluated in hepatocytes from rats feda HF diet or a HF diet supplemented with 25 KO with theaim to investigate the role of mitochondria in themodulationof nutrient metabolism exerted by KO
No change in PDH activity which essentially producesacetyl-CoA from carbohydrate was seen after administrationof HF or HF+KO diets Nevertheless fasting serum glucoseand insulin were higher after feeding a HF diet suggesting
BioMed Research International 9
an altered insulin sensitivity as indicated by HOMA indexchanges In fact an excess of energy supplied by fat corre-sponds to a scarce tissutal utilization of glucose thereby caus-ing hyperglycemia and hyperinsulinemia Insulin resistancewas reduced after KO supplementation
CPT I activity which is involved in fatty acid oxidationwas inhibited by a HF diet and stimulated after KO supple-mentationThe increase inCPT I activity observed inHF+KOgroup could be explained by the lower levels ofmalonyl-CoAametabolic intermediate of hepatic lipogenesis which in thiscase is instead strongly inhibited [2 10] Interestingly thecapability of KO to stimulate fatty acid oxidation seems tobe associated to the presence of a high lipid content (35)in the experimental diet used in this study In fact whenKO was added to a standard diet the expression of enzymesinvolved inmitochondrial fatty acid 120573-oxidation significantlydecreased [12]
The modulation of the CPT I activity observed in HFand HF+KO animals seems to be accompanied by a parallelmodulation of the activity of CS a Krebs cycle enzyme thatcatalyzes the synthesis of citrate fromoxaloacetate and acetyl-CoA Recently a strong and significant correlation of CSwith obesity was noted with a reduced enzyme activity inmitochondria of human omental adipose tissue in the stateof obesity [28] According to this observation the increasein CS activity revealed after KO supplementation could beexplained as a compensatory response to maintain the entryof acetyl-CoA produced by fatty acid oxidation into the Krebscycle It is interesting to underline that citrate generated byCS continues in the Krebs cycle In fact in animals fed aHF diet supplemented with KO a reduction in the activityand in the expression of citrate carrier protein (CIC) whichtransports citrate from mitochondria to the cytosol for fattyacid synthesis was observed [2 10 24]
NADH and FADH2generated by 120573-oxidation and Krebs
cycle are used in the final common oxidative phosphorylationsystem (OXPHOS) to generate ATP This process is coupledwith the transfer of electrons along the respiratory chainOur data indicate that a HF diet caused a strong decrease inthe mitochondrial respiratory efficiency and this effect couldbe due to a possible uncoupling effect exerted by the excessof fatty acids present in this diet In fact we found a littledecrease in 119881
3and a strong increase in 119881
4values in liver
mitochondria isolated fromHF animals1198814is the resting state
of mitochondrial respiration and an increase in this valuesuggests a stimulus ofmitochondrial respiration independentof ADP phosphorylation In other terms during HF feedinga lower coupling betweenmitochondrial respiration andATPsynthesis occurs Interestingly the addition of KO to the HFdiet almost completely reversed this effect thereby leadingmitochondria respiration efficiency to be comparable to thatof control rats
When we assayed in a more selective approach theactivity of single components of the respiratory chain weobserved a significant decrease in the activity of all respira-tory chain complexes in HF animals according to a recentstudy [29] inwhich an impairment ofmitochondrial functionand in particular of complex I activity was found AfterKO supplementation the values of complex activities were
identical to those measured in control animals Expressionof OXPHOS proteins significantly increased in HF animalscompared to the control group and this evidence was accom-panied by the presence of ATP levels in HF animals thatwere only slightly decreased in comparison to those foundin control animals According to this result previous studies[30 31] demonstrated that a high fat content in the dietinduced higher level of these proteins as a compensatoryresponse to the reduced activity of the respiratory chaincomplexes When we performed western blotting analysis inmitochondria isolated from HF+KO animals liver we founda higher level of the OXPHOS proteins compared to thecontrol and theHF groups probably caused by the synergisticeffect of the HF diet and the KO supplementation Indeedit is already known that genes involved in mitochondrialOXPHOS proteins expression were upregulated in miceconsuming KO as revealed from the transcriptomic analysis[12] Overall thismeans that KO is able to induce the burningof excess of fat introduced by diet stimulating fatty acidoxidation Krebs cycle and respiratory chain complex activityand preventing a condition of oxidative damage
Uncoupling between mitochondrial respiration and ATPsynthesis observed in HF animals could be caused bya different expression of mitochondrial carrier proteinsinvolved in the regulation of proton conductance It is knownthat uncoupling proteins (UCPs) homologues uncouplemitochondrial respiration from oxidative phosphorylationincreasing thermogenesis while reducing the efficiency ofATP synthesis In a previous study [32] an increase in theexpression of the UCP2 isoform was found in rats withNAFLD Interestingly in this study we found a massiveincrease of this carrier protein inHF animals but the additionof KO to the diet was able to strongly reduce the expressionof hepatic UCP2 Therefore it is reasonable to assume thatthe addition of KO to the HF diet can increase the couplingbetween mitochondrial respiration and ATP synthesis by adownregulation of UCP2 expression
Not only are UCPs involved in mitochondrial uncou-pling butAAC is another carrier protein of particular interestin the understanding of the mitochondrial energy couplingprocess [33]This carrier catalyzes the exchange of ADP3minus forATP4minus and therefore utilizes as its driving force the electro-chemical potential generated across the inner membrane bythe respiratory chain The exchange reaction is electrogenicand driven by the electrical component of the proton gradi-ent so that entry of ADP into mitochondria and exit of ATPare favored We can therefore speculate that if mitochondriaare uncoupled and proton gradient was dissipated by theentry of H+ into the matrix by UCP2 an upregulation ofthe expression of AAC could compensate a reduced fluxof ATP from mitochondria when membrane potential wasreduced This hypothesized compensatory mechanism wasobserved in animals fed a HF diet The increase in theexpression of AAC was less evident in HF+KO animals inwhich livermitochondria respiratory efficiencywas unalteredin comparison to the control animals
In conclusion the present study indicates that KO coun-teracts the negative effects of a HF diet In particular our
10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 BioMed Research International
Table 2 Body and liver weight serum parameters and liver fat content
Control HF HF+KOBody weight (g) 2458 plusmn 78 2772 plusmn 158lowast 2352 plusmn 252
Liver weight (g100 g body weight) 39 plusmn 06 35 plusmn 05 41 plusmn 03Glycemia (mM) 29 plusmn 01 49 plusmn 07lowast 41 plusmn 04lowast
Insulin (120583UmL) 501 plusmn 75 1428 plusmn 100lowast 777 plusmn 50lowast
HOMA 64 plusmn 01 312 plusmn 03lowast 142 plusmn 01lowast
Serum triglycerides (mgdL) 1373 plusmn 129 1471 plusmn 88 1386 plusmn 92Serum cholesterol (mgdL) 926 plusmn 83 956 plusmn 42 941 plusmn 62Liver triglycerides (mgg liver) 86 plusmn 04 136 plusmn 10lowast 83 plusmn 05
Liver cholesterol (mgg liver) 27 plusmn 04 66 plusmn 04lowast 35 plusmn 05
Each point represents the mean plusmn SD for 5 animals lowast119875 lt 005 versus rats fed a control diet 119875 lt 005 versus rats fed HF diet
carrier proteins AAC (ADPATP carrier) and UCP2 (uncou-pling protein 2) was determined by western blotting analysisPolyacrylamide gel electrophoresis was performed in thepresence of 01 SDS (SDS-PAGE) according to standardprocedures The mitochondrial proteins that have been sepa-rated by SDS-PAGEwere transferred to a nitrocellulosemem-brane For the OXPHOS proteins a cocktail directed againstNDUFB8 (Complex I) SDHB (Complex II) COR2 (ComplexIII) COX1 (Complex IV) and ATP5A (ATP-synthase) wasused at a dilution 1 1times101 and the immunoreaction was car-ried out with monoclonal antibody by chemiluminescenceFor AAC UCP2 and porin (a protein used as a control)detection antisera were used at a dilution of 1 1times103 and theimmunoreaction was carried out with polyclonal antibody bychemiluminescence
28 ATP and Lipid Hydroperoxide (LPO) Levels ATP levelsin liver homogenates were determined by using the luciferaseATP assay kit The concentration of ATP in the samples wascalculated from interpolation of a standard curve preparedwith known amounts of ATP Results were expressed asnmoles of ATP per mg of protein
Lipid peroxidation levels in liver samples were deter-mined using a lipid hydroperoxide assay kit which measuresthe redox reactions with ferrous ions LPO content wasexpressed as LPO per mg of protein
29 Statistical Analysis Experimental data represent themeans plusmn SD The data were analyzed by one-way ANOVAand a Tukey-Kramer post hoc analysis was used to detectsignificant differences between the means at a level of 119875 gt005
3 Results
31 Body and Liver Weight Serum Parameters and Liver FatContent As shown in Table 2 a significant increase in bodyweight was recorded in HF animals after 4 weeks of dietarytreatment in agreement with the high caloric content ofthe diet Interestingly the supplementation of the HF dietwith 25 KO significantly prevented this effect Insteadno statistically significant variations were found in the liverweight of animals fed with the different experimental diets
The blood glucose concentration increased in both HFand HF+KO animals after 4 weeks of dietary administrationin comparison to control rats Such an increase was lessevident in the case of the HF+KO fed animals A massiveincrease in the levels of insulin was revealed in the plasmaof HF animals in comparison to control rats Interestinglythe KO supplementation significantly mitigated this increaseHF rats showed a marked increase in HOMA index demon-strating typical insulin resistance induced by the high-fat dietIn the HF+KO animals HOMA decreased showing reducedinsulin resistance induced by the addition of KO
No significant difference was detected in the serumlipid (triglyceride and cholesterol) levels between the threegroups of animals On the contrary liver lipid content wasdramatically enhanced in HF animals thus reflecting thepresence of hepatic steatosis (Figure 1) The supplementationwith KO of the HF diet reversed this effect thereby keepingthe lipid concentrations similar to those found in controlrats Accordingly the liver histologic examination revealed abeginning of microvesicular fat depositions only in HF rats
32 Pyruvate Dehydrogenase (PDH) Complex CarnitinePalmitoyltransferase (CPT) and Citrate Synthase (CS) Activ-ity Mitochondria play a central role in energy productionSugars and fatty acids undergo glycolysis and mitochon-drial 120573-oxidation respectively to produce acetyl-CoA (Fig-ure 2(a)) PDH complex activity is the major determinantof glucose oxidation in animal cells We found that PDHactivity remained unchanged in the three groups of animalssuggesting that glucose catabolism was not affected by a HFdiet or by KO supplementation (Figure 2(b))
The limiting step for catabolic pathway of fatty acidoxidation is represented by the activity of CPT I which isresponsible for the entry of fatty acids into the mitochondrialmatrix where 120573-oxidation of fatty acids occurs As shown inFigure 2(c) in the HF animals a clear decrease (about 40)in the CPT I activity was detected On the contrary a strongincrease in CPT I activity was found in theHF+KO ratsThusKO supplemented to theHF diet strongly stimulates fatty acidoxidation
Acetyl-CoA generated essentially from glucose or fromfatty acids is channeled into the Krebs cycle where CS(the rate-limiting enzyme of the Krebs cycle) catalyzes its
BioMed Research International 5
Control HF HF+KO
Figure 1 Liver samples were stored in neutral formaldehyde and embedded in paraffin wax Sections (15120583m) were stained with hematoxylinand eosin
Glucose
Fatty acids
CPT I
Pyruvate
NAD+
120573-oxidationAcetyl-CoA Acyl-CoA
CSCO2
PDH
Oxaloacetate
Krebscycle
FADH2
Mitochondrion
Cytosol
NADH + H+
NADH + H+ NADH + H+
NADH + H+
(a)
0
5
10
15
20
Control HF
PDH
activ
ity (n
molmiddotm
inminus1middotm
g pr
otei
nminus1)
HF+KO
(b)
Control HF0
25
50
75
100
125
150
CPT
I act
ivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(c)
Control HF0
50
100
150
200
250
300
CS ac
tivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(d)
Figure 2Mitochondrialmetabolic pathways fromglucose and fatty acid oxidation Schematic picture of themetabolic pathways fromglucoseand fatty acid oxidation (a) Effect of KO on the activity of mitochondrial enzymes PDH (b) CPT I (c) and CS (d) in control HF or HF+KO-fed rats liver The values reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed a control diet 119875 lt 005 versusrats fed a HF diet
6 BioMed Research International
Table 3 Mitochondrial respiratory efficiency
Control HF HF+KO
Palmitoyl-L-carnitine + malate (120573 oxidation)1198813(nmol O2sdotmLminus1sdotminminus1) 858 plusmn 78 597 plusmn 85lowast 1140 plusmn 112lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 143 plusmn 24 229 plusmn 15lowast 185 plusmn 52lowast
RCR 60 plusmn 04 26 plusmn 03lowast 61 plusmn 08
Pyruvate + malate (Complex I)1198813(nmol O2sdotmLminus1sdotminminus1) 170 plusmn 03 159 plusmn 04lowast 199 plusmn 09lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 54 plusmn 01 76 plusmn 02lowast 66 plusmn 06lowast
RCR 32 plusmn 03 21 plusmn 03lowast 30 plusmn 04
Succinate + rotenone (Complex II)1198813(nmol O2sdotmLminus1sdotminminus1) 615 plusmn 33 531 plusmn 37lowast 783 plusmn 40lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 101 plusmn 03 250 plusmn 12lowast 150 plusmn 19lowast
RCR 61 plusmn 09 21 plusmn 08lowast 52 plusmn 10
Ascorbate + TMPD + rotenone + antimycin A (Complex IV)1198813(nmol O2sdotmLminus1sdotminminus1) 551 plusmn 53 475 plusmn 65lowast 686 plusmn 78lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 93 plusmn 07 172 plusmn 09lowast 128 plusmn 08lowast
RCR 59 plusmn 06 28 plusmn 03lowast 54 plusmn 09
Respiratory control ratio (RCR) was calculated as the ratio of the rate of oxygen uptake in the presence of added ADP (1198813) to the rate observed when added
ADP had been completely phosphorylated to ATP (1198814)
lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
condensation with oxaloacetate to synthesize citrate (Fig-ure 2(a)) A significant decrease (about 55) in the activityof the CS was found in HF animals (Figure 2(d)) whileonly a moderate decrease (about 35) was observed inHF+KO animals in comparison to the control groups Thepartial reversal of CS inhibition observed in HF animals afterKO supplementation could be explained by a compensatoryresponse of this enzyme to sustain oxidation of acetyl-CoAgenerated from fatty acid oxidation
33 Mitochondrial Oxygen Consumption Respiratory ChainComplex Activities and ATP Synthesis The increased fattyacid oxidation observed in HF+KO animals produces higherlevels of reducing equivalents which are normally addressedtowards the mitochondrial oxidative phosphorylation forATP production We therefore analyzed the respiratoryefficiency of freshly isolated mitochondria by oxygraphicmethods Mitochondrial oxygen consumption was measuredwith various substrates in the presence (state 3) or the absence(state 4) of ADP (Table 3) According to the results shownin Figure 2(c) when we used palmitoyl-L-carnitine andmalate as substrates 119881
3(mitochondrial respiration state 3)
decreased inHF animals thus indicating lowermitochondrial120573-oxidation On the other hand the increase in the119881
4(mito-
chondrial respiration state 4) values suggests that the excess ofdietary fat most probably is the cause of a partial uncouplingbetween respiration and phosphorylation in mitochondriaInterestingly the dietary KO supplementation is able toefficiently abolish this effect keeping the mitochondrialrespiratory efficiency unaltered
When we added respiratory substrates for mitochondrialcomplexes I II and IV (Table 3) a little yet significantdecrease in 119881
3was observed in the HF groups in comparison
to control animals In the HF+KO animals a significantincrease in the 119881
3values was found in comparison to HF
and control animals 1198814 also known as the resting state of
respiration showed an increase in theHF andHF+KO groupeven if the increase observed after KO supplementation was
less evident than that found inHF animals As a consequencethe RCRvalueswere profoundly and significantly lower inHFanimals with respect to those calculated for control animalsThe KO supplementation of the HF diet reversed this effectthereby keeping RCR values similar to those calculated forcontrol group
In a more selective approach for investigating the func-tionality of the mitochondrial oxidative phosphorylation weassayed the activity of single components of the respiratorychain complexes IndashIV andATP-synthase (Figure 3(a)) A sig-nificant decrease (about 30) in the activity of all respiratorychain complexes was found in HF animals On the contrarythe values of complexes activity of the HF+KO rats werepractically identical to those measured in control animals
Surprisingly western blotting analysis of the OXPHOSproteins showed significantly higher protein levels in bothHFand HF+KO animals compared to the control group (Figures3(b) and 3(c)) Such an increase was more evident for com-plexes I III and IV which are associated as supercomplexesin the mitochondrial inner membrane [21] This effect couldbe determined by a compensatory mechanism related to thereduction of the activities of the respiratory complexes
In order to prove biological relevance of alteration inmitochondrial respiratory chain activity we measured ATPconcentration and oxidative damage in liver samples fromthe three groups of animals (Figure 3(d)) ATP quantificationdemonstrated that aHFdiet caused a slight but not significantdecrease in ATP content an effect that was rescued by KOInterestingly in parallel experiments we found a significantincrease (about 30) in the LPO levels inHF animals and thiseffect was efficiently reversed by KO supplementation of theHF diet
34 Expression of Proteins Related to the Respiratory ChainFunction Uncoupling protein 2 (UCP2) uncouples respira-tion from oxidative phosphorylation We therefore analyzedthe expression profile of this carrier protein in the threegroups of animals (Figure 4(a)) After 4 weeks of HF dietary
BioMed Research International 7
0
30
60
90
120
Con
trol
HF
Resp
irato
ry co
mpl
exes
activ
ity (
)
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
lowast lowast lowastlowast
lowast
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(a)
CV-ATP5A
CIII-COR2CIV-COX1
CII-SDHB
CI-NDUFB8
Porin
(b)
0
50
100
150
250
OXP
HO
S pr
otei
ns (
) 200lowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
Con
trol
HF
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(c)
30
90
0
60
150
( o
f con
trol)
120
ATP LPO
lowast
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
(d)
Figure 3 Effect of KO on respiratory complexes activity expression and function Effect of KO on respiratory complexes activity (a)Enzymatic activities measured in the control group were set to 100 Effect of KO on OXPHOS proteins levels (b) Liver mitochondrialproteins from control HF or HF+KO-fed rats were separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with acocktail against NDUFB8 SDHB COR2 COX1 ATP5A or the mammalian porin as a controlThe amount of OXPHOS proteins revealed byimmunodecoration in the control group was set to 100 (c) ATP and LPO quantification of liver samples isolated from animals fed a controldiet a HF diet or a HF+KO (d) The amount of ATP or LPO (expressed as nmoles per mg protein) in the control group was set to 100Thevalues reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
treatment the amount of this protein increased about 5-foldcompared to the control group Interestingly a significantsmaller increase in the expression of UCP2 was found afterKO supplementation In fact in the presence of KO in theHF diet the level of this carrier protein was about 50 of thatfound in HF rats
The AAC (ADPATP carrier) is another mitochondrialcarrier which is able to regulate the basal proton conductanceThis protein exchanges ADP for ATP across the mitochon-drial inner membrane and may also play an important rolein the mitochondrial permeability transition pore When weanalyzed the expression of this carrier protein (Figure 4(b))we found an increase of about 22-fold in HF animalscompared to the control group AAC levels in the HF+KOanimals even if higher than those found in the controlrats were significantly lower than those found in HF group
The amount of porin an outer membrane protein tested as acontrol did not change in any group of animals (Figure 4(c))
These results suggest that the KO supplementation is ableto modulate the expression of mitochondrial carrier proteininvolved in the regulation of proton conductance reducingthe uncoupling effect caused by a HF diet
4 Discussions
In animal experiments a diet rich in fat or carbohydrates hasbeen shown to induce hyperglycemia and hyperinsulinemiaand to contribute to the development of obesity and NAFLD[2 6 7 22] This last condition is recognized as the mostcommon chronic liver disease in the Western world NASHis a more severe form of NAFLD that is broadly defined bythe presence of steatosis with inflammation and progressive
8 BioMed Research International
0
100
200
300
500
Control HF
UCP
2 (
)
UCP2
400
lowast
lowast
HF+KO
(a)
0
60
120
180
300
AAC
()
AAC
240
Control HF
lowast
lowast
HF+KO
(b)
0
25
50
75
125
Porin
()
Porin
100
Control HF HF+KO
(c)
Figure 4 Effect of KO on proteins related to the respiratory chain function Effect of KO on proteins levels of UCP2 (a) and AAC (b) Theamount of porin an outer membrane protein was tested as a control (c) Liver mitochondrial proteins from control HF or HF+KO-fed ratswere separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with antisera against UCP2 AAC or the mammalianporinThe amount of proteins revealed by immunodecoration in the control groupwas set to 100The values reported in the figure representthe means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
fibrosis leading then to cirrhosis and hepatocellular carci-noma Some patients with NAFLD develop NASH throughpoorly understood mechanisms Since NAFLD and NASHare considered lifestyle-associated diseases diet interventionis an important approach to their treatment
In this context it is important to underline that theldquoobesogenicrdquoWestern diet has a ratio of fatty acids and carbo-hydrates of approximately 5050Moreover it is characterizedby an insufficient n-3 PUFAs consumption and a high n-6n-3 ratio which are associated with the progression fromsimple steatosis to NASH [8] along with modifications of gutmicrobiota [23]
Although the mechanisms responsible for increasedlipid accumulation in the liver are not yet fully elucidateddecreased capacity to oxidize fatty acids increased transportof fatty acids to liver and increased hepatic fatty acidsynthesis seem to be important factors involved in thepathogenesis of NAFLD [2 6 7 24]Moreover abnormalitiesin mitochondrial morphology and functions seem to beassociated with fatty liver [1ndash3 25ndash27] This finding suggeststhat regulation of mitochondria function and of molecularmechanisms involved in lipid accumulation in the liver couldprovide useful targets to prevent the development of NAFLD
In recent years the use of dietary supplements hasrapidly increased [2 6 10 24 28] with the aim to preventNAFLD KO a dietary supplement of n-3 PUFAs has becomeincreasingly popular because of its capability of reducing
hepatic lipogenesis and of stimulating catabolization ofexcess fat introduced by a HF diet [2 10] The analysisof hepatic transcriptome in mice demonstrated that KOsupplementation was able to regulate genes involved in hep-atic energy metabolism including glucose metabolism lipidbiosynthesis fatty acid metabolism cholesterol biosynthesisand mitochondrial electron transport more efficiently thanfish oil [10 12] These differential results were tentativelyascribed to the influence of the structural form of n-3PUFAs esterified to either phospholipids or triglycerideson a biological response Moreover PUFAs as phospholipids(KO) or triglycerides (fish oil)may differentially influence gutmicrobiota or synthesis of bacterial metabolites
A previous study by our group [2] suggested that KOsupplementation to a HF diet resulted in a retention of nor-malmitochondrial respiration efficiency whichwas impairedin HF animals The aim of the present study was thereforeto analyze the molecular mechanisms by which dietary fatsinfluence mitochondrial energetic metabolismThis new andintriguing aspect was evaluated in hepatocytes from rats feda HF diet or a HF diet supplemented with 25 KO with theaim to investigate the role of mitochondria in themodulationof nutrient metabolism exerted by KO
No change in PDH activity which essentially producesacetyl-CoA from carbohydrate was seen after administrationof HF or HF+KO diets Nevertheless fasting serum glucoseand insulin were higher after feeding a HF diet suggesting
BioMed Research International 9
an altered insulin sensitivity as indicated by HOMA indexchanges In fact an excess of energy supplied by fat corre-sponds to a scarce tissutal utilization of glucose thereby caus-ing hyperglycemia and hyperinsulinemia Insulin resistancewas reduced after KO supplementation
CPT I activity which is involved in fatty acid oxidationwas inhibited by a HF diet and stimulated after KO supple-mentationThe increase inCPT I activity observed inHF+KOgroup could be explained by the lower levels ofmalonyl-CoAametabolic intermediate of hepatic lipogenesis which in thiscase is instead strongly inhibited [2 10] Interestingly thecapability of KO to stimulate fatty acid oxidation seems tobe associated to the presence of a high lipid content (35)in the experimental diet used in this study In fact whenKO was added to a standard diet the expression of enzymesinvolved inmitochondrial fatty acid 120573-oxidation significantlydecreased [12]
The modulation of the CPT I activity observed in HFand HF+KO animals seems to be accompanied by a parallelmodulation of the activity of CS a Krebs cycle enzyme thatcatalyzes the synthesis of citrate fromoxaloacetate and acetyl-CoA Recently a strong and significant correlation of CSwith obesity was noted with a reduced enzyme activity inmitochondria of human omental adipose tissue in the stateof obesity [28] According to this observation the increasein CS activity revealed after KO supplementation could beexplained as a compensatory response to maintain the entryof acetyl-CoA produced by fatty acid oxidation into the Krebscycle It is interesting to underline that citrate generated byCS continues in the Krebs cycle In fact in animals fed aHF diet supplemented with KO a reduction in the activityand in the expression of citrate carrier protein (CIC) whichtransports citrate from mitochondria to the cytosol for fattyacid synthesis was observed [2 10 24]
NADH and FADH2generated by 120573-oxidation and Krebs
cycle are used in the final common oxidative phosphorylationsystem (OXPHOS) to generate ATP This process is coupledwith the transfer of electrons along the respiratory chainOur data indicate that a HF diet caused a strong decrease inthe mitochondrial respiratory efficiency and this effect couldbe due to a possible uncoupling effect exerted by the excessof fatty acids present in this diet In fact we found a littledecrease in 119881
3and a strong increase in 119881
4values in liver
mitochondria isolated fromHF animals1198814is the resting state
of mitochondrial respiration and an increase in this valuesuggests a stimulus ofmitochondrial respiration independentof ADP phosphorylation In other terms during HF feedinga lower coupling betweenmitochondrial respiration andATPsynthesis occurs Interestingly the addition of KO to the HFdiet almost completely reversed this effect thereby leadingmitochondria respiration efficiency to be comparable to thatof control rats
When we assayed in a more selective approach theactivity of single components of the respiratory chain weobserved a significant decrease in the activity of all respira-tory chain complexes in HF animals according to a recentstudy [29] inwhich an impairment ofmitochondrial functionand in particular of complex I activity was found AfterKO supplementation the values of complex activities were
identical to those measured in control animals Expressionof OXPHOS proteins significantly increased in HF animalscompared to the control group and this evidence was accom-panied by the presence of ATP levels in HF animals thatwere only slightly decreased in comparison to those foundin control animals According to this result previous studies[30 31] demonstrated that a high fat content in the dietinduced higher level of these proteins as a compensatoryresponse to the reduced activity of the respiratory chaincomplexes When we performed western blotting analysis inmitochondria isolated from HF+KO animals liver we founda higher level of the OXPHOS proteins compared to thecontrol and theHF groups probably caused by the synergisticeffect of the HF diet and the KO supplementation Indeedit is already known that genes involved in mitochondrialOXPHOS proteins expression were upregulated in miceconsuming KO as revealed from the transcriptomic analysis[12] Overall thismeans that KO is able to induce the burningof excess of fat introduced by diet stimulating fatty acidoxidation Krebs cycle and respiratory chain complex activityand preventing a condition of oxidative damage
Uncoupling between mitochondrial respiration and ATPsynthesis observed in HF animals could be caused bya different expression of mitochondrial carrier proteinsinvolved in the regulation of proton conductance It is knownthat uncoupling proteins (UCPs) homologues uncouplemitochondrial respiration from oxidative phosphorylationincreasing thermogenesis while reducing the efficiency ofATP synthesis In a previous study [32] an increase in theexpression of the UCP2 isoform was found in rats withNAFLD Interestingly in this study we found a massiveincrease of this carrier protein inHF animals but the additionof KO to the diet was able to strongly reduce the expressionof hepatic UCP2 Therefore it is reasonable to assume thatthe addition of KO to the HF diet can increase the couplingbetween mitochondrial respiration and ATP synthesis by adownregulation of UCP2 expression
Not only are UCPs involved in mitochondrial uncou-pling butAAC is another carrier protein of particular interestin the understanding of the mitochondrial energy couplingprocess [33]This carrier catalyzes the exchange of ADP3minus forATP4minus and therefore utilizes as its driving force the electro-chemical potential generated across the inner membrane bythe respiratory chain The exchange reaction is electrogenicand driven by the electrical component of the proton gradi-ent so that entry of ADP into mitochondria and exit of ATPare favored We can therefore speculate that if mitochondriaare uncoupled and proton gradient was dissipated by theentry of H+ into the matrix by UCP2 an upregulation ofthe expression of AAC could compensate a reduced fluxof ATP from mitochondria when membrane potential wasreduced This hypothesized compensatory mechanism wasobserved in animals fed a HF diet The increase in theexpression of AAC was less evident in HF+KO animals inwhich livermitochondria respiratory efficiencywas unalteredin comparison to the control animals
In conclusion the present study indicates that KO coun-teracts the negative effects of a HF diet In particular our
10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 5
Control HF HF+KO
Figure 1 Liver samples were stored in neutral formaldehyde and embedded in paraffin wax Sections (15120583m) were stained with hematoxylinand eosin
Glucose
Fatty acids
CPT I
Pyruvate
NAD+
120573-oxidationAcetyl-CoA Acyl-CoA
CSCO2
PDH
Oxaloacetate
Krebscycle
FADH2
Mitochondrion
Cytosol
NADH + H+
NADH + H+ NADH + H+
NADH + H+
(a)
0
5
10
15
20
Control HF
PDH
activ
ity (n
molmiddotm
inminus1middotm
g pr
otei
nminus1)
HF+KO
(b)
Control HF0
25
50
75
100
125
150
CPT
I act
ivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(c)
Control HF0
50
100
150
200
250
300
CS ac
tivity
(nm
olmiddotm
inminus1middotm
g pr
otei
nminus1)
lowast
lowast
HF+KO
(d)
Figure 2Mitochondrialmetabolic pathways fromglucose and fatty acid oxidation Schematic picture of themetabolic pathways fromglucoseand fatty acid oxidation (a) Effect of KO on the activity of mitochondrial enzymes PDH (b) CPT I (c) and CS (d) in control HF or HF+KO-fed rats liver The values reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed a control diet 119875 lt 005 versusrats fed a HF diet
6 BioMed Research International
Table 3 Mitochondrial respiratory efficiency
Control HF HF+KO
Palmitoyl-L-carnitine + malate (120573 oxidation)1198813(nmol O2sdotmLminus1sdotminminus1) 858 plusmn 78 597 plusmn 85lowast 1140 plusmn 112lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 143 plusmn 24 229 plusmn 15lowast 185 plusmn 52lowast
RCR 60 plusmn 04 26 plusmn 03lowast 61 plusmn 08
Pyruvate + malate (Complex I)1198813(nmol O2sdotmLminus1sdotminminus1) 170 plusmn 03 159 plusmn 04lowast 199 plusmn 09lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 54 plusmn 01 76 plusmn 02lowast 66 plusmn 06lowast
RCR 32 plusmn 03 21 plusmn 03lowast 30 plusmn 04
Succinate + rotenone (Complex II)1198813(nmol O2sdotmLminus1sdotminminus1) 615 plusmn 33 531 plusmn 37lowast 783 plusmn 40lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 101 plusmn 03 250 plusmn 12lowast 150 plusmn 19lowast
RCR 61 plusmn 09 21 plusmn 08lowast 52 plusmn 10
Ascorbate + TMPD + rotenone + antimycin A (Complex IV)1198813(nmol O2sdotmLminus1sdotminminus1) 551 plusmn 53 475 plusmn 65lowast 686 plusmn 78lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 93 plusmn 07 172 plusmn 09lowast 128 plusmn 08lowast
RCR 59 plusmn 06 28 plusmn 03lowast 54 plusmn 09
Respiratory control ratio (RCR) was calculated as the ratio of the rate of oxygen uptake in the presence of added ADP (1198813) to the rate observed when added
ADP had been completely phosphorylated to ATP (1198814)
lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
condensation with oxaloacetate to synthesize citrate (Fig-ure 2(a)) A significant decrease (about 55) in the activityof the CS was found in HF animals (Figure 2(d)) whileonly a moderate decrease (about 35) was observed inHF+KO animals in comparison to the control groups Thepartial reversal of CS inhibition observed in HF animals afterKO supplementation could be explained by a compensatoryresponse of this enzyme to sustain oxidation of acetyl-CoAgenerated from fatty acid oxidation
33 Mitochondrial Oxygen Consumption Respiratory ChainComplex Activities and ATP Synthesis The increased fattyacid oxidation observed in HF+KO animals produces higherlevels of reducing equivalents which are normally addressedtowards the mitochondrial oxidative phosphorylation forATP production We therefore analyzed the respiratoryefficiency of freshly isolated mitochondria by oxygraphicmethods Mitochondrial oxygen consumption was measuredwith various substrates in the presence (state 3) or the absence(state 4) of ADP (Table 3) According to the results shownin Figure 2(c) when we used palmitoyl-L-carnitine andmalate as substrates 119881
3(mitochondrial respiration state 3)
decreased inHF animals thus indicating lowermitochondrial120573-oxidation On the other hand the increase in the119881
4(mito-
chondrial respiration state 4) values suggests that the excess ofdietary fat most probably is the cause of a partial uncouplingbetween respiration and phosphorylation in mitochondriaInterestingly the dietary KO supplementation is able toefficiently abolish this effect keeping the mitochondrialrespiratory efficiency unaltered
When we added respiratory substrates for mitochondrialcomplexes I II and IV (Table 3) a little yet significantdecrease in 119881
3was observed in the HF groups in comparison
to control animals In the HF+KO animals a significantincrease in the 119881
3values was found in comparison to HF
and control animals 1198814 also known as the resting state of
respiration showed an increase in theHF andHF+KO groupeven if the increase observed after KO supplementation was
less evident than that found inHF animals As a consequencethe RCRvalueswere profoundly and significantly lower inHFanimals with respect to those calculated for control animalsThe KO supplementation of the HF diet reversed this effectthereby keeping RCR values similar to those calculated forcontrol group
In a more selective approach for investigating the func-tionality of the mitochondrial oxidative phosphorylation weassayed the activity of single components of the respiratorychain complexes IndashIV andATP-synthase (Figure 3(a)) A sig-nificant decrease (about 30) in the activity of all respiratorychain complexes was found in HF animals On the contrarythe values of complexes activity of the HF+KO rats werepractically identical to those measured in control animals
Surprisingly western blotting analysis of the OXPHOSproteins showed significantly higher protein levels in bothHFand HF+KO animals compared to the control group (Figures3(b) and 3(c)) Such an increase was more evident for com-plexes I III and IV which are associated as supercomplexesin the mitochondrial inner membrane [21] This effect couldbe determined by a compensatory mechanism related to thereduction of the activities of the respiratory complexes
In order to prove biological relevance of alteration inmitochondrial respiratory chain activity we measured ATPconcentration and oxidative damage in liver samples fromthe three groups of animals (Figure 3(d)) ATP quantificationdemonstrated that aHFdiet caused a slight but not significantdecrease in ATP content an effect that was rescued by KOInterestingly in parallel experiments we found a significantincrease (about 30) in the LPO levels inHF animals and thiseffect was efficiently reversed by KO supplementation of theHF diet
34 Expression of Proteins Related to the Respiratory ChainFunction Uncoupling protein 2 (UCP2) uncouples respira-tion from oxidative phosphorylation We therefore analyzedthe expression profile of this carrier protein in the threegroups of animals (Figure 4(a)) After 4 weeks of HF dietary
BioMed Research International 7
0
30
60
90
120
Con
trol
HF
Resp
irato
ry co
mpl
exes
activ
ity (
)
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
lowast lowast lowastlowast
lowast
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(a)
CV-ATP5A
CIII-COR2CIV-COX1
CII-SDHB
CI-NDUFB8
Porin
(b)
0
50
100
150
250
OXP
HO
S pr
otei
ns (
) 200lowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
Con
trol
HF
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(c)
30
90
0
60
150
( o
f con
trol)
120
ATP LPO
lowast
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
(d)
Figure 3 Effect of KO on respiratory complexes activity expression and function Effect of KO on respiratory complexes activity (a)Enzymatic activities measured in the control group were set to 100 Effect of KO on OXPHOS proteins levels (b) Liver mitochondrialproteins from control HF or HF+KO-fed rats were separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with acocktail against NDUFB8 SDHB COR2 COX1 ATP5A or the mammalian porin as a controlThe amount of OXPHOS proteins revealed byimmunodecoration in the control group was set to 100 (c) ATP and LPO quantification of liver samples isolated from animals fed a controldiet a HF diet or a HF+KO (d) The amount of ATP or LPO (expressed as nmoles per mg protein) in the control group was set to 100Thevalues reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
treatment the amount of this protein increased about 5-foldcompared to the control group Interestingly a significantsmaller increase in the expression of UCP2 was found afterKO supplementation In fact in the presence of KO in theHF diet the level of this carrier protein was about 50 of thatfound in HF rats
The AAC (ADPATP carrier) is another mitochondrialcarrier which is able to regulate the basal proton conductanceThis protein exchanges ADP for ATP across the mitochon-drial inner membrane and may also play an important rolein the mitochondrial permeability transition pore When weanalyzed the expression of this carrier protein (Figure 4(b))we found an increase of about 22-fold in HF animalscompared to the control group AAC levels in the HF+KOanimals even if higher than those found in the controlrats were significantly lower than those found in HF group
The amount of porin an outer membrane protein tested as acontrol did not change in any group of animals (Figure 4(c))
These results suggest that the KO supplementation is ableto modulate the expression of mitochondrial carrier proteininvolved in the regulation of proton conductance reducingthe uncoupling effect caused by a HF diet
4 Discussions
In animal experiments a diet rich in fat or carbohydrates hasbeen shown to induce hyperglycemia and hyperinsulinemiaand to contribute to the development of obesity and NAFLD[2 6 7 22] This last condition is recognized as the mostcommon chronic liver disease in the Western world NASHis a more severe form of NAFLD that is broadly defined bythe presence of steatosis with inflammation and progressive
8 BioMed Research International
0
100
200
300
500
Control HF
UCP
2 (
)
UCP2
400
lowast
lowast
HF+KO
(a)
0
60
120
180
300
AAC
()
AAC
240
Control HF
lowast
lowast
HF+KO
(b)
0
25
50
75
125
Porin
()
Porin
100
Control HF HF+KO
(c)
Figure 4 Effect of KO on proteins related to the respiratory chain function Effect of KO on proteins levels of UCP2 (a) and AAC (b) Theamount of porin an outer membrane protein was tested as a control (c) Liver mitochondrial proteins from control HF or HF+KO-fed ratswere separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with antisera against UCP2 AAC or the mammalianporinThe amount of proteins revealed by immunodecoration in the control groupwas set to 100The values reported in the figure representthe means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
fibrosis leading then to cirrhosis and hepatocellular carci-noma Some patients with NAFLD develop NASH throughpoorly understood mechanisms Since NAFLD and NASHare considered lifestyle-associated diseases diet interventionis an important approach to their treatment
In this context it is important to underline that theldquoobesogenicrdquoWestern diet has a ratio of fatty acids and carbo-hydrates of approximately 5050Moreover it is characterizedby an insufficient n-3 PUFAs consumption and a high n-6n-3 ratio which are associated with the progression fromsimple steatosis to NASH [8] along with modifications of gutmicrobiota [23]
Although the mechanisms responsible for increasedlipid accumulation in the liver are not yet fully elucidateddecreased capacity to oxidize fatty acids increased transportof fatty acids to liver and increased hepatic fatty acidsynthesis seem to be important factors involved in thepathogenesis of NAFLD [2 6 7 24]Moreover abnormalitiesin mitochondrial morphology and functions seem to beassociated with fatty liver [1ndash3 25ndash27] This finding suggeststhat regulation of mitochondria function and of molecularmechanisms involved in lipid accumulation in the liver couldprovide useful targets to prevent the development of NAFLD
In recent years the use of dietary supplements hasrapidly increased [2 6 10 24 28] with the aim to preventNAFLD KO a dietary supplement of n-3 PUFAs has becomeincreasingly popular because of its capability of reducing
hepatic lipogenesis and of stimulating catabolization ofexcess fat introduced by a HF diet [2 10] The analysisof hepatic transcriptome in mice demonstrated that KOsupplementation was able to regulate genes involved in hep-atic energy metabolism including glucose metabolism lipidbiosynthesis fatty acid metabolism cholesterol biosynthesisand mitochondrial electron transport more efficiently thanfish oil [10 12] These differential results were tentativelyascribed to the influence of the structural form of n-3PUFAs esterified to either phospholipids or triglycerideson a biological response Moreover PUFAs as phospholipids(KO) or triglycerides (fish oil)may differentially influence gutmicrobiota or synthesis of bacterial metabolites
A previous study by our group [2] suggested that KOsupplementation to a HF diet resulted in a retention of nor-malmitochondrial respiration efficiency whichwas impairedin HF animals The aim of the present study was thereforeto analyze the molecular mechanisms by which dietary fatsinfluence mitochondrial energetic metabolismThis new andintriguing aspect was evaluated in hepatocytes from rats feda HF diet or a HF diet supplemented with 25 KO with theaim to investigate the role of mitochondria in themodulationof nutrient metabolism exerted by KO
No change in PDH activity which essentially producesacetyl-CoA from carbohydrate was seen after administrationof HF or HF+KO diets Nevertheless fasting serum glucoseand insulin were higher after feeding a HF diet suggesting
BioMed Research International 9
an altered insulin sensitivity as indicated by HOMA indexchanges In fact an excess of energy supplied by fat corre-sponds to a scarce tissutal utilization of glucose thereby caus-ing hyperglycemia and hyperinsulinemia Insulin resistancewas reduced after KO supplementation
CPT I activity which is involved in fatty acid oxidationwas inhibited by a HF diet and stimulated after KO supple-mentationThe increase inCPT I activity observed inHF+KOgroup could be explained by the lower levels ofmalonyl-CoAametabolic intermediate of hepatic lipogenesis which in thiscase is instead strongly inhibited [2 10] Interestingly thecapability of KO to stimulate fatty acid oxidation seems tobe associated to the presence of a high lipid content (35)in the experimental diet used in this study In fact whenKO was added to a standard diet the expression of enzymesinvolved inmitochondrial fatty acid 120573-oxidation significantlydecreased [12]
The modulation of the CPT I activity observed in HFand HF+KO animals seems to be accompanied by a parallelmodulation of the activity of CS a Krebs cycle enzyme thatcatalyzes the synthesis of citrate fromoxaloacetate and acetyl-CoA Recently a strong and significant correlation of CSwith obesity was noted with a reduced enzyme activity inmitochondria of human omental adipose tissue in the stateof obesity [28] According to this observation the increasein CS activity revealed after KO supplementation could beexplained as a compensatory response to maintain the entryof acetyl-CoA produced by fatty acid oxidation into the Krebscycle It is interesting to underline that citrate generated byCS continues in the Krebs cycle In fact in animals fed aHF diet supplemented with KO a reduction in the activityand in the expression of citrate carrier protein (CIC) whichtransports citrate from mitochondria to the cytosol for fattyacid synthesis was observed [2 10 24]
NADH and FADH2generated by 120573-oxidation and Krebs
cycle are used in the final common oxidative phosphorylationsystem (OXPHOS) to generate ATP This process is coupledwith the transfer of electrons along the respiratory chainOur data indicate that a HF diet caused a strong decrease inthe mitochondrial respiratory efficiency and this effect couldbe due to a possible uncoupling effect exerted by the excessof fatty acids present in this diet In fact we found a littledecrease in 119881
3and a strong increase in 119881
4values in liver
mitochondria isolated fromHF animals1198814is the resting state
of mitochondrial respiration and an increase in this valuesuggests a stimulus ofmitochondrial respiration independentof ADP phosphorylation In other terms during HF feedinga lower coupling betweenmitochondrial respiration andATPsynthesis occurs Interestingly the addition of KO to the HFdiet almost completely reversed this effect thereby leadingmitochondria respiration efficiency to be comparable to thatof control rats
When we assayed in a more selective approach theactivity of single components of the respiratory chain weobserved a significant decrease in the activity of all respira-tory chain complexes in HF animals according to a recentstudy [29] inwhich an impairment ofmitochondrial functionand in particular of complex I activity was found AfterKO supplementation the values of complex activities were
identical to those measured in control animals Expressionof OXPHOS proteins significantly increased in HF animalscompared to the control group and this evidence was accom-panied by the presence of ATP levels in HF animals thatwere only slightly decreased in comparison to those foundin control animals According to this result previous studies[30 31] demonstrated that a high fat content in the dietinduced higher level of these proteins as a compensatoryresponse to the reduced activity of the respiratory chaincomplexes When we performed western blotting analysis inmitochondria isolated from HF+KO animals liver we founda higher level of the OXPHOS proteins compared to thecontrol and theHF groups probably caused by the synergisticeffect of the HF diet and the KO supplementation Indeedit is already known that genes involved in mitochondrialOXPHOS proteins expression were upregulated in miceconsuming KO as revealed from the transcriptomic analysis[12] Overall thismeans that KO is able to induce the burningof excess of fat introduced by diet stimulating fatty acidoxidation Krebs cycle and respiratory chain complex activityand preventing a condition of oxidative damage
Uncoupling between mitochondrial respiration and ATPsynthesis observed in HF animals could be caused bya different expression of mitochondrial carrier proteinsinvolved in the regulation of proton conductance It is knownthat uncoupling proteins (UCPs) homologues uncouplemitochondrial respiration from oxidative phosphorylationincreasing thermogenesis while reducing the efficiency ofATP synthesis In a previous study [32] an increase in theexpression of the UCP2 isoform was found in rats withNAFLD Interestingly in this study we found a massiveincrease of this carrier protein inHF animals but the additionof KO to the diet was able to strongly reduce the expressionof hepatic UCP2 Therefore it is reasonable to assume thatthe addition of KO to the HF diet can increase the couplingbetween mitochondrial respiration and ATP synthesis by adownregulation of UCP2 expression
Not only are UCPs involved in mitochondrial uncou-pling butAAC is another carrier protein of particular interestin the understanding of the mitochondrial energy couplingprocess [33]This carrier catalyzes the exchange of ADP3minus forATP4minus and therefore utilizes as its driving force the electro-chemical potential generated across the inner membrane bythe respiratory chain The exchange reaction is electrogenicand driven by the electrical component of the proton gradi-ent so that entry of ADP into mitochondria and exit of ATPare favored We can therefore speculate that if mitochondriaare uncoupled and proton gradient was dissipated by theentry of H+ into the matrix by UCP2 an upregulation ofthe expression of AAC could compensate a reduced fluxof ATP from mitochondria when membrane potential wasreduced This hypothesized compensatory mechanism wasobserved in animals fed a HF diet The increase in theexpression of AAC was less evident in HF+KO animals inwhich livermitochondria respiratory efficiencywas unalteredin comparison to the control animals
In conclusion the present study indicates that KO coun-teracts the negative effects of a HF diet In particular our
10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
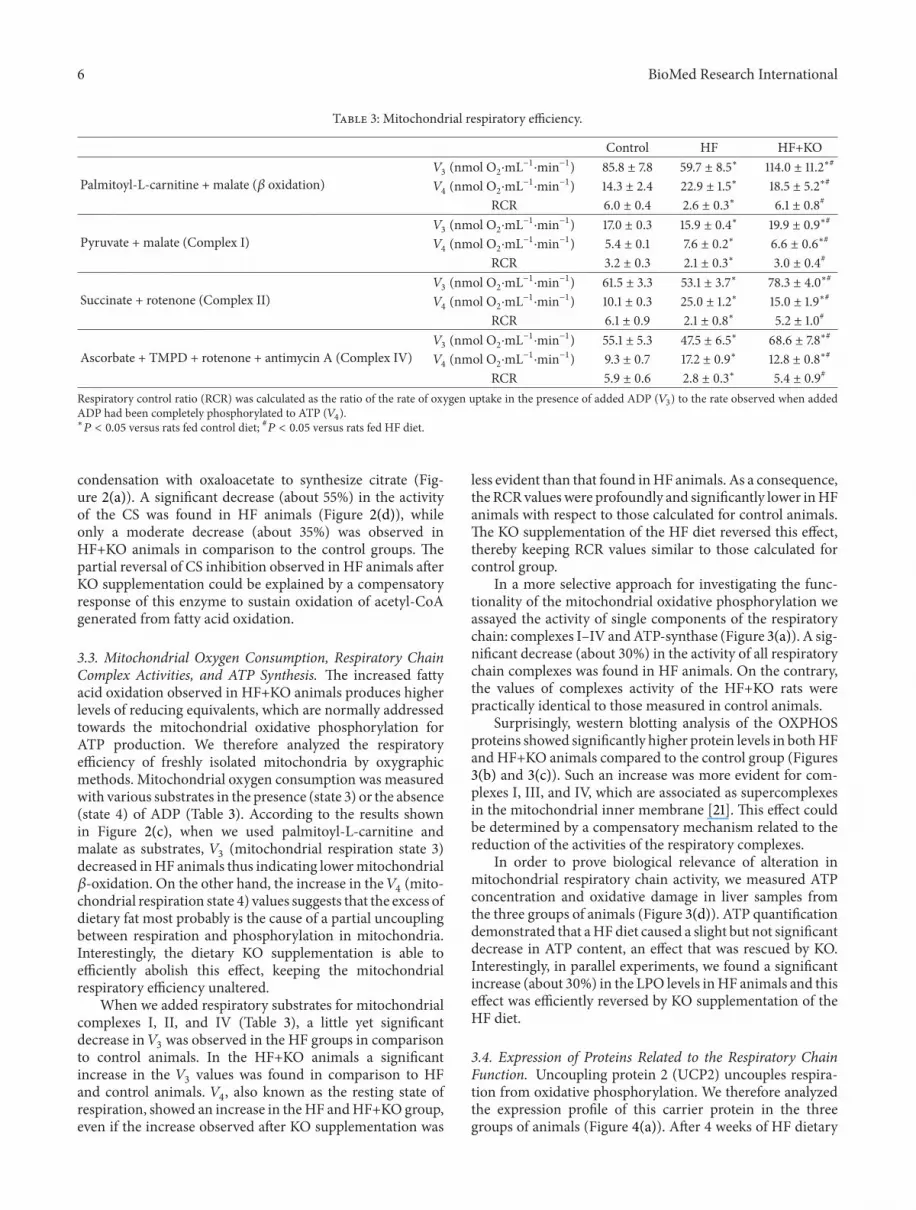
6 BioMed Research International
Table 3 Mitochondrial respiratory efficiency
Control HF HF+KO
Palmitoyl-L-carnitine + malate (120573 oxidation)1198813(nmol O2sdotmLminus1sdotminminus1) 858 plusmn 78 597 plusmn 85lowast 1140 plusmn 112lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 143 plusmn 24 229 plusmn 15lowast 185 plusmn 52lowast
RCR 60 plusmn 04 26 plusmn 03lowast 61 plusmn 08
Pyruvate + malate (Complex I)1198813(nmol O2sdotmLminus1sdotminminus1) 170 plusmn 03 159 plusmn 04lowast 199 plusmn 09lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 54 plusmn 01 76 plusmn 02lowast 66 plusmn 06lowast
RCR 32 plusmn 03 21 plusmn 03lowast 30 plusmn 04
Succinate + rotenone (Complex II)1198813(nmol O2sdotmLminus1sdotminminus1) 615 plusmn 33 531 plusmn 37lowast 783 plusmn 40lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 101 plusmn 03 250 plusmn 12lowast 150 plusmn 19lowast
RCR 61 plusmn 09 21 plusmn 08lowast 52 plusmn 10
Ascorbate + TMPD + rotenone + antimycin A (Complex IV)1198813(nmol O2sdotmLminus1sdotminminus1) 551 plusmn 53 475 plusmn 65lowast 686 plusmn 78lowast
1198814(nmol O2sdotmLminus1sdotminminus1) 93 plusmn 07 172 plusmn 09lowast 128 plusmn 08lowast
RCR 59 plusmn 06 28 plusmn 03lowast 54 plusmn 09
Respiratory control ratio (RCR) was calculated as the ratio of the rate of oxygen uptake in the presence of added ADP (1198813) to the rate observed when added
ADP had been completely phosphorylated to ATP (1198814)
lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
condensation with oxaloacetate to synthesize citrate (Fig-ure 2(a)) A significant decrease (about 55) in the activityof the CS was found in HF animals (Figure 2(d)) whileonly a moderate decrease (about 35) was observed inHF+KO animals in comparison to the control groups Thepartial reversal of CS inhibition observed in HF animals afterKO supplementation could be explained by a compensatoryresponse of this enzyme to sustain oxidation of acetyl-CoAgenerated from fatty acid oxidation
33 Mitochondrial Oxygen Consumption Respiratory ChainComplex Activities and ATP Synthesis The increased fattyacid oxidation observed in HF+KO animals produces higherlevels of reducing equivalents which are normally addressedtowards the mitochondrial oxidative phosphorylation forATP production We therefore analyzed the respiratoryefficiency of freshly isolated mitochondria by oxygraphicmethods Mitochondrial oxygen consumption was measuredwith various substrates in the presence (state 3) or the absence(state 4) of ADP (Table 3) According to the results shownin Figure 2(c) when we used palmitoyl-L-carnitine andmalate as substrates 119881
3(mitochondrial respiration state 3)
decreased inHF animals thus indicating lowermitochondrial120573-oxidation On the other hand the increase in the119881
4(mito-
chondrial respiration state 4) values suggests that the excess ofdietary fat most probably is the cause of a partial uncouplingbetween respiration and phosphorylation in mitochondriaInterestingly the dietary KO supplementation is able toefficiently abolish this effect keeping the mitochondrialrespiratory efficiency unaltered
When we added respiratory substrates for mitochondrialcomplexes I II and IV (Table 3) a little yet significantdecrease in 119881
3was observed in the HF groups in comparison
to control animals In the HF+KO animals a significantincrease in the 119881
3values was found in comparison to HF
and control animals 1198814 also known as the resting state of
respiration showed an increase in theHF andHF+KO groupeven if the increase observed after KO supplementation was
less evident than that found inHF animals As a consequencethe RCRvalueswere profoundly and significantly lower inHFanimals with respect to those calculated for control animalsThe KO supplementation of the HF diet reversed this effectthereby keeping RCR values similar to those calculated forcontrol group
In a more selective approach for investigating the func-tionality of the mitochondrial oxidative phosphorylation weassayed the activity of single components of the respiratorychain complexes IndashIV andATP-synthase (Figure 3(a)) A sig-nificant decrease (about 30) in the activity of all respiratorychain complexes was found in HF animals On the contrarythe values of complexes activity of the HF+KO rats werepractically identical to those measured in control animals
Surprisingly western blotting analysis of the OXPHOSproteins showed significantly higher protein levels in bothHFand HF+KO animals compared to the control group (Figures3(b) and 3(c)) Such an increase was more evident for com-plexes I III and IV which are associated as supercomplexesin the mitochondrial inner membrane [21] This effect couldbe determined by a compensatory mechanism related to thereduction of the activities of the respiratory complexes
In order to prove biological relevance of alteration inmitochondrial respiratory chain activity we measured ATPconcentration and oxidative damage in liver samples fromthe three groups of animals (Figure 3(d)) ATP quantificationdemonstrated that aHFdiet caused a slight but not significantdecrease in ATP content an effect that was rescued by KOInterestingly in parallel experiments we found a significantincrease (about 30) in the LPO levels inHF animals and thiseffect was efficiently reversed by KO supplementation of theHF diet
34 Expression of Proteins Related to the Respiratory ChainFunction Uncoupling protein 2 (UCP2) uncouples respira-tion from oxidative phosphorylation We therefore analyzedthe expression profile of this carrier protein in the threegroups of animals (Figure 4(a)) After 4 weeks of HF dietary
BioMed Research International 7
0
30
60
90
120
Con
trol
HF
Resp
irato
ry co
mpl
exes
activ
ity (
)
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
lowast lowast lowastlowast
lowast
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(a)
CV-ATP5A
CIII-COR2CIV-COX1
CII-SDHB
CI-NDUFB8
Porin
(b)
0
50
100
150
250
OXP
HO
S pr
otei
ns (
) 200lowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
Con
trol
HF
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(c)
30
90
0
60
150
( o
f con
trol)
120
ATP LPO
lowast
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
(d)
Figure 3 Effect of KO on respiratory complexes activity expression and function Effect of KO on respiratory complexes activity (a)Enzymatic activities measured in the control group were set to 100 Effect of KO on OXPHOS proteins levels (b) Liver mitochondrialproteins from control HF or HF+KO-fed rats were separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with acocktail against NDUFB8 SDHB COR2 COX1 ATP5A or the mammalian porin as a controlThe amount of OXPHOS proteins revealed byimmunodecoration in the control group was set to 100 (c) ATP and LPO quantification of liver samples isolated from animals fed a controldiet a HF diet or a HF+KO (d) The amount of ATP or LPO (expressed as nmoles per mg protein) in the control group was set to 100Thevalues reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
treatment the amount of this protein increased about 5-foldcompared to the control group Interestingly a significantsmaller increase in the expression of UCP2 was found afterKO supplementation In fact in the presence of KO in theHF diet the level of this carrier protein was about 50 of thatfound in HF rats
The AAC (ADPATP carrier) is another mitochondrialcarrier which is able to regulate the basal proton conductanceThis protein exchanges ADP for ATP across the mitochon-drial inner membrane and may also play an important rolein the mitochondrial permeability transition pore When weanalyzed the expression of this carrier protein (Figure 4(b))we found an increase of about 22-fold in HF animalscompared to the control group AAC levels in the HF+KOanimals even if higher than those found in the controlrats were significantly lower than those found in HF group
The amount of porin an outer membrane protein tested as acontrol did not change in any group of animals (Figure 4(c))
These results suggest that the KO supplementation is ableto modulate the expression of mitochondrial carrier proteininvolved in the regulation of proton conductance reducingthe uncoupling effect caused by a HF diet
4 Discussions
In animal experiments a diet rich in fat or carbohydrates hasbeen shown to induce hyperglycemia and hyperinsulinemiaand to contribute to the development of obesity and NAFLD[2 6 7 22] This last condition is recognized as the mostcommon chronic liver disease in the Western world NASHis a more severe form of NAFLD that is broadly defined bythe presence of steatosis with inflammation and progressive
8 BioMed Research International
0
100
200
300
500
Control HF
UCP
2 (
)
UCP2
400
lowast
lowast
HF+KO
(a)
0
60
120
180
300
AAC
()
AAC
240
Control HF
lowast
lowast
HF+KO
(b)
0
25
50
75
125
Porin
()
Porin
100
Control HF HF+KO
(c)
Figure 4 Effect of KO on proteins related to the respiratory chain function Effect of KO on proteins levels of UCP2 (a) and AAC (b) Theamount of porin an outer membrane protein was tested as a control (c) Liver mitochondrial proteins from control HF or HF+KO-fed ratswere separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with antisera against UCP2 AAC or the mammalianporinThe amount of proteins revealed by immunodecoration in the control groupwas set to 100The values reported in the figure representthe means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
fibrosis leading then to cirrhosis and hepatocellular carci-noma Some patients with NAFLD develop NASH throughpoorly understood mechanisms Since NAFLD and NASHare considered lifestyle-associated diseases diet interventionis an important approach to their treatment
In this context it is important to underline that theldquoobesogenicrdquoWestern diet has a ratio of fatty acids and carbo-hydrates of approximately 5050Moreover it is characterizedby an insufficient n-3 PUFAs consumption and a high n-6n-3 ratio which are associated with the progression fromsimple steatosis to NASH [8] along with modifications of gutmicrobiota [23]
Although the mechanisms responsible for increasedlipid accumulation in the liver are not yet fully elucidateddecreased capacity to oxidize fatty acids increased transportof fatty acids to liver and increased hepatic fatty acidsynthesis seem to be important factors involved in thepathogenesis of NAFLD [2 6 7 24]Moreover abnormalitiesin mitochondrial morphology and functions seem to beassociated with fatty liver [1ndash3 25ndash27] This finding suggeststhat regulation of mitochondria function and of molecularmechanisms involved in lipid accumulation in the liver couldprovide useful targets to prevent the development of NAFLD
In recent years the use of dietary supplements hasrapidly increased [2 6 10 24 28] with the aim to preventNAFLD KO a dietary supplement of n-3 PUFAs has becomeincreasingly popular because of its capability of reducing
hepatic lipogenesis and of stimulating catabolization ofexcess fat introduced by a HF diet [2 10] The analysisof hepatic transcriptome in mice demonstrated that KOsupplementation was able to regulate genes involved in hep-atic energy metabolism including glucose metabolism lipidbiosynthesis fatty acid metabolism cholesterol biosynthesisand mitochondrial electron transport more efficiently thanfish oil [10 12] These differential results were tentativelyascribed to the influence of the structural form of n-3PUFAs esterified to either phospholipids or triglycerideson a biological response Moreover PUFAs as phospholipids(KO) or triglycerides (fish oil)may differentially influence gutmicrobiota or synthesis of bacterial metabolites
A previous study by our group [2] suggested that KOsupplementation to a HF diet resulted in a retention of nor-malmitochondrial respiration efficiency whichwas impairedin HF animals The aim of the present study was thereforeto analyze the molecular mechanisms by which dietary fatsinfluence mitochondrial energetic metabolismThis new andintriguing aspect was evaluated in hepatocytes from rats feda HF diet or a HF diet supplemented with 25 KO with theaim to investigate the role of mitochondria in themodulationof nutrient metabolism exerted by KO
No change in PDH activity which essentially producesacetyl-CoA from carbohydrate was seen after administrationof HF or HF+KO diets Nevertheless fasting serum glucoseand insulin were higher after feeding a HF diet suggesting
BioMed Research International 9
an altered insulin sensitivity as indicated by HOMA indexchanges In fact an excess of energy supplied by fat corre-sponds to a scarce tissutal utilization of glucose thereby caus-ing hyperglycemia and hyperinsulinemia Insulin resistancewas reduced after KO supplementation
CPT I activity which is involved in fatty acid oxidationwas inhibited by a HF diet and stimulated after KO supple-mentationThe increase inCPT I activity observed inHF+KOgroup could be explained by the lower levels ofmalonyl-CoAametabolic intermediate of hepatic lipogenesis which in thiscase is instead strongly inhibited [2 10] Interestingly thecapability of KO to stimulate fatty acid oxidation seems tobe associated to the presence of a high lipid content (35)in the experimental diet used in this study In fact whenKO was added to a standard diet the expression of enzymesinvolved inmitochondrial fatty acid 120573-oxidation significantlydecreased [12]
The modulation of the CPT I activity observed in HFand HF+KO animals seems to be accompanied by a parallelmodulation of the activity of CS a Krebs cycle enzyme thatcatalyzes the synthesis of citrate fromoxaloacetate and acetyl-CoA Recently a strong and significant correlation of CSwith obesity was noted with a reduced enzyme activity inmitochondria of human omental adipose tissue in the stateof obesity [28] According to this observation the increasein CS activity revealed after KO supplementation could beexplained as a compensatory response to maintain the entryof acetyl-CoA produced by fatty acid oxidation into the Krebscycle It is interesting to underline that citrate generated byCS continues in the Krebs cycle In fact in animals fed aHF diet supplemented with KO a reduction in the activityand in the expression of citrate carrier protein (CIC) whichtransports citrate from mitochondria to the cytosol for fattyacid synthesis was observed [2 10 24]
NADH and FADH2generated by 120573-oxidation and Krebs
cycle are used in the final common oxidative phosphorylationsystem (OXPHOS) to generate ATP This process is coupledwith the transfer of electrons along the respiratory chainOur data indicate that a HF diet caused a strong decrease inthe mitochondrial respiratory efficiency and this effect couldbe due to a possible uncoupling effect exerted by the excessof fatty acids present in this diet In fact we found a littledecrease in 119881
3and a strong increase in 119881
4values in liver
mitochondria isolated fromHF animals1198814is the resting state
of mitochondrial respiration and an increase in this valuesuggests a stimulus ofmitochondrial respiration independentof ADP phosphorylation In other terms during HF feedinga lower coupling betweenmitochondrial respiration andATPsynthesis occurs Interestingly the addition of KO to the HFdiet almost completely reversed this effect thereby leadingmitochondria respiration efficiency to be comparable to thatof control rats
When we assayed in a more selective approach theactivity of single components of the respiratory chain weobserved a significant decrease in the activity of all respira-tory chain complexes in HF animals according to a recentstudy [29] inwhich an impairment ofmitochondrial functionand in particular of complex I activity was found AfterKO supplementation the values of complex activities were
identical to those measured in control animals Expressionof OXPHOS proteins significantly increased in HF animalscompared to the control group and this evidence was accom-panied by the presence of ATP levels in HF animals thatwere only slightly decreased in comparison to those foundin control animals According to this result previous studies[30 31] demonstrated that a high fat content in the dietinduced higher level of these proteins as a compensatoryresponse to the reduced activity of the respiratory chaincomplexes When we performed western blotting analysis inmitochondria isolated from HF+KO animals liver we founda higher level of the OXPHOS proteins compared to thecontrol and theHF groups probably caused by the synergisticeffect of the HF diet and the KO supplementation Indeedit is already known that genes involved in mitochondrialOXPHOS proteins expression were upregulated in miceconsuming KO as revealed from the transcriptomic analysis[12] Overall thismeans that KO is able to induce the burningof excess of fat introduced by diet stimulating fatty acidoxidation Krebs cycle and respiratory chain complex activityand preventing a condition of oxidative damage
Uncoupling between mitochondrial respiration and ATPsynthesis observed in HF animals could be caused bya different expression of mitochondrial carrier proteinsinvolved in the regulation of proton conductance It is knownthat uncoupling proteins (UCPs) homologues uncouplemitochondrial respiration from oxidative phosphorylationincreasing thermogenesis while reducing the efficiency ofATP synthesis In a previous study [32] an increase in theexpression of the UCP2 isoform was found in rats withNAFLD Interestingly in this study we found a massiveincrease of this carrier protein inHF animals but the additionof KO to the diet was able to strongly reduce the expressionof hepatic UCP2 Therefore it is reasonable to assume thatthe addition of KO to the HF diet can increase the couplingbetween mitochondrial respiration and ATP synthesis by adownregulation of UCP2 expression
Not only are UCPs involved in mitochondrial uncou-pling butAAC is another carrier protein of particular interestin the understanding of the mitochondrial energy couplingprocess [33]This carrier catalyzes the exchange of ADP3minus forATP4minus and therefore utilizes as its driving force the electro-chemical potential generated across the inner membrane bythe respiratory chain The exchange reaction is electrogenicand driven by the electrical component of the proton gradi-ent so that entry of ADP into mitochondria and exit of ATPare favored We can therefore speculate that if mitochondriaare uncoupled and proton gradient was dissipated by theentry of H+ into the matrix by UCP2 an upregulation ofthe expression of AAC could compensate a reduced fluxof ATP from mitochondria when membrane potential wasreduced This hypothesized compensatory mechanism wasobserved in animals fed a HF diet The increase in theexpression of AAC was less evident in HF+KO animals inwhich livermitochondria respiratory efficiencywas unalteredin comparison to the control animals
In conclusion the present study indicates that KO coun-teracts the negative effects of a HF diet In particular our
10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
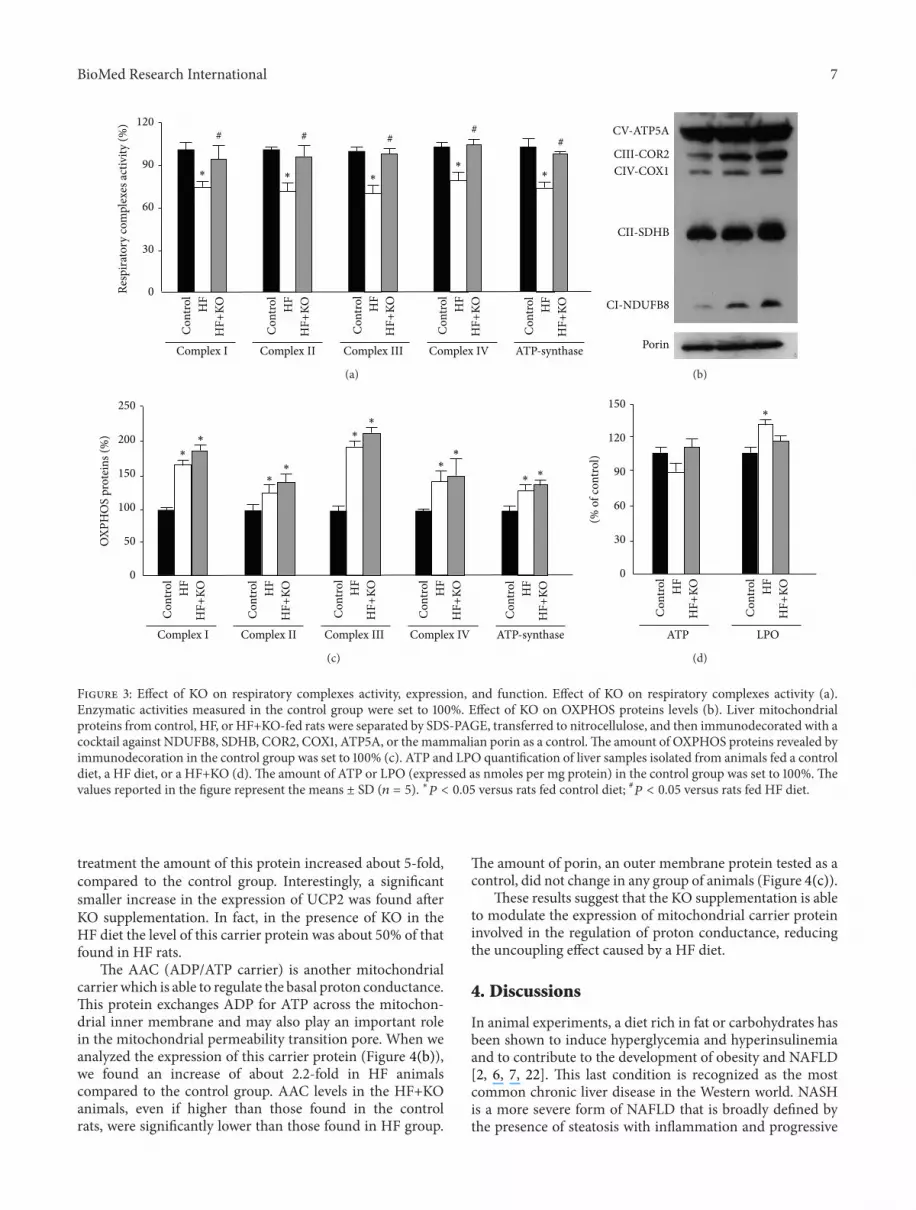
BioMed Research International 7
0
30
60
90
120
Con
trol
HF
Resp
irato
ry co
mpl
exes
activ
ity (
)
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
lowast lowast lowastlowast
lowast
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(a)
CV-ATP5A
CIII-COR2CIV-COX1
CII-SDHB
CI-NDUFB8
Porin
(b)
0
50
100
150
250
OXP
HO
S pr
otei
ns (
) 200lowast
lowast
lowastlowast
lowastlowast
lowastlowast
lowast lowast
Con
trol
HF
Complex I Complex II Complex III Complex IV ATP-synthase
Con
trol
HF
Con
trol
HF
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
HF+
KO
HF+
KO
HF+
KO
(c)
30
90
0
60
150
( o
f con
trol)
120
ATP LPO
lowast
Con
trol
HF
Con
trol
HF
HF+
KO
HF+
KO
(d)
Figure 3 Effect of KO on respiratory complexes activity expression and function Effect of KO on respiratory complexes activity (a)Enzymatic activities measured in the control group were set to 100 Effect of KO on OXPHOS proteins levels (b) Liver mitochondrialproteins from control HF or HF+KO-fed rats were separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with acocktail against NDUFB8 SDHB COR2 COX1 ATP5A or the mammalian porin as a controlThe amount of OXPHOS proteins revealed byimmunodecoration in the control group was set to 100 (c) ATP and LPO quantification of liver samples isolated from animals fed a controldiet a HF diet or a HF+KO (d) The amount of ATP or LPO (expressed as nmoles per mg protein) in the control group was set to 100Thevalues reported in the figure represent the means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
treatment the amount of this protein increased about 5-foldcompared to the control group Interestingly a significantsmaller increase in the expression of UCP2 was found afterKO supplementation In fact in the presence of KO in theHF diet the level of this carrier protein was about 50 of thatfound in HF rats
The AAC (ADPATP carrier) is another mitochondrialcarrier which is able to regulate the basal proton conductanceThis protein exchanges ADP for ATP across the mitochon-drial inner membrane and may also play an important rolein the mitochondrial permeability transition pore When weanalyzed the expression of this carrier protein (Figure 4(b))we found an increase of about 22-fold in HF animalscompared to the control group AAC levels in the HF+KOanimals even if higher than those found in the controlrats were significantly lower than those found in HF group
The amount of porin an outer membrane protein tested as acontrol did not change in any group of animals (Figure 4(c))
These results suggest that the KO supplementation is ableto modulate the expression of mitochondrial carrier proteininvolved in the regulation of proton conductance reducingthe uncoupling effect caused by a HF diet
4 Discussions
In animal experiments a diet rich in fat or carbohydrates hasbeen shown to induce hyperglycemia and hyperinsulinemiaand to contribute to the development of obesity and NAFLD[2 6 7 22] This last condition is recognized as the mostcommon chronic liver disease in the Western world NASHis a more severe form of NAFLD that is broadly defined bythe presence of steatosis with inflammation and progressive
8 BioMed Research International
0
100
200
300
500
Control HF
UCP
2 (
)
UCP2
400
lowast
lowast
HF+KO
(a)
0
60
120
180
300
AAC
()
AAC
240
Control HF
lowast
lowast
HF+KO
(b)
0
25
50
75
125
Porin
()
Porin
100
Control HF HF+KO
(c)
Figure 4 Effect of KO on proteins related to the respiratory chain function Effect of KO on proteins levels of UCP2 (a) and AAC (b) Theamount of porin an outer membrane protein was tested as a control (c) Liver mitochondrial proteins from control HF or HF+KO-fed ratswere separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with antisera against UCP2 AAC or the mammalianporinThe amount of proteins revealed by immunodecoration in the control groupwas set to 100The values reported in the figure representthe means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
fibrosis leading then to cirrhosis and hepatocellular carci-noma Some patients with NAFLD develop NASH throughpoorly understood mechanisms Since NAFLD and NASHare considered lifestyle-associated diseases diet interventionis an important approach to their treatment
In this context it is important to underline that theldquoobesogenicrdquoWestern diet has a ratio of fatty acids and carbo-hydrates of approximately 5050Moreover it is characterizedby an insufficient n-3 PUFAs consumption and a high n-6n-3 ratio which are associated with the progression fromsimple steatosis to NASH [8] along with modifications of gutmicrobiota [23]
Although the mechanisms responsible for increasedlipid accumulation in the liver are not yet fully elucidateddecreased capacity to oxidize fatty acids increased transportof fatty acids to liver and increased hepatic fatty acidsynthesis seem to be important factors involved in thepathogenesis of NAFLD [2 6 7 24]Moreover abnormalitiesin mitochondrial morphology and functions seem to beassociated with fatty liver [1ndash3 25ndash27] This finding suggeststhat regulation of mitochondria function and of molecularmechanisms involved in lipid accumulation in the liver couldprovide useful targets to prevent the development of NAFLD
In recent years the use of dietary supplements hasrapidly increased [2 6 10 24 28] with the aim to preventNAFLD KO a dietary supplement of n-3 PUFAs has becomeincreasingly popular because of its capability of reducing
hepatic lipogenesis and of stimulating catabolization ofexcess fat introduced by a HF diet [2 10] The analysisof hepatic transcriptome in mice demonstrated that KOsupplementation was able to regulate genes involved in hep-atic energy metabolism including glucose metabolism lipidbiosynthesis fatty acid metabolism cholesterol biosynthesisand mitochondrial electron transport more efficiently thanfish oil [10 12] These differential results were tentativelyascribed to the influence of the structural form of n-3PUFAs esterified to either phospholipids or triglycerideson a biological response Moreover PUFAs as phospholipids(KO) or triglycerides (fish oil)may differentially influence gutmicrobiota or synthesis of bacterial metabolites
A previous study by our group [2] suggested that KOsupplementation to a HF diet resulted in a retention of nor-malmitochondrial respiration efficiency whichwas impairedin HF animals The aim of the present study was thereforeto analyze the molecular mechanisms by which dietary fatsinfluence mitochondrial energetic metabolismThis new andintriguing aspect was evaluated in hepatocytes from rats feda HF diet or a HF diet supplemented with 25 KO with theaim to investigate the role of mitochondria in themodulationof nutrient metabolism exerted by KO
No change in PDH activity which essentially producesacetyl-CoA from carbohydrate was seen after administrationof HF or HF+KO diets Nevertheless fasting serum glucoseand insulin were higher after feeding a HF diet suggesting
BioMed Research International 9
an altered insulin sensitivity as indicated by HOMA indexchanges In fact an excess of energy supplied by fat corre-sponds to a scarce tissutal utilization of glucose thereby caus-ing hyperglycemia and hyperinsulinemia Insulin resistancewas reduced after KO supplementation
CPT I activity which is involved in fatty acid oxidationwas inhibited by a HF diet and stimulated after KO supple-mentationThe increase inCPT I activity observed inHF+KOgroup could be explained by the lower levels ofmalonyl-CoAametabolic intermediate of hepatic lipogenesis which in thiscase is instead strongly inhibited [2 10] Interestingly thecapability of KO to stimulate fatty acid oxidation seems tobe associated to the presence of a high lipid content (35)in the experimental diet used in this study In fact whenKO was added to a standard diet the expression of enzymesinvolved inmitochondrial fatty acid 120573-oxidation significantlydecreased [12]
The modulation of the CPT I activity observed in HFand HF+KO animals seems to be accompanied by a parallelmodulation of the activity of CS a Krebs cycle enzyme thatcatalyzes the synthesis of citrate fromoxaloacetate and acetyl-CoA Recently a strong and significant correlation of CSwith obesity was noted with a reduced enzyme activity inmitochondria of human omental adipose tissue in the stateof obesity [28] According to this observation the increasein CS activity revealed after KO supplementation could beexplained as a compensatory response to maintain the entryof acetyl-CoA produced by fatty acid oxidation into the Krebscycle It is interesting to underline that citrate generated byCS continues in the Krebs cycle In fact in animals fed aHF diet supplemented with KO a reduction in the activityand in the expression of citrate carrier protein (CIC) whichtransports citrate from mitochondria to the cytosol for fattyacid synthesis was observed [2 10 24]
NADH and FADH2generated by 120573-oxidation and Krebs
cycle are used in the final common oxidative phosphorylationsystem (OXPHOS) to generate ATP This process is coupledwith the transfer of electrons along the respiratory chainOur data indicate that a HF diet caused a strong decrease inthe mitochondrial respiratory efficiency and this effect couldbe due to a possible uncoupling effect exerted by the excessof fatty acids present in this diet In fact we found a littledecrease in 119881
3and a strong increase in 119881
4values in liver
mitochondria isolated fromHF animals1198814is the resting state
of mitochondrial respiration and an increase in this valuesuggests a stimulus ofmitochondrial respiration independentof ADP phosphorylation In other terms during HF feedinga lower coupling betweenmitochondrial respiration andATPsynthesis occurs Interestingly the addition of KO to the HFdiet almost completely reversed this effect thereby leadingmitochondria respiration efficiency to be comparable to thatof control rats
When we assayed in a more selective approach theactivity of single components of the respiratory chain weobserved a significant decrease in the activity of all respira-tory chain complexes in HF animals according to a recentstudy [29] inwhich an impairment ofmitochondrial functionand in particular of complex I activity was found AfterKO supplementation the values of complex activities were
identical to those measured in control animals Expressionof OXPHOS proteins significantly increased in HF animalscompared to the control group and this evidence was accom-panied by the presence of ATP levels in HF animals thatwere only slightly decreased in comparison to those foundin control animals According to this result previous studies[30 31] demonstrated that a high fat content in the dietinduced higher level of these proteins as a compensatoryresponse to the reduced activity of the respiratory chaincomplexes When we performed western blotting analysis inmitochondria isolated from HF+KO animals liver we founda higher level of the OXPHOS proteins compared to thecontrol and theHF groups probably caused by the synergisticeffect of the HF diet and the KO supplementation Indeedit is already known that genes involved in mitochondrialOXPHOS proteins expression were upregulated in miceconsuming KO as revealed from the transcriptomic analysis[12] Overall thismeans that KO is able to induce the burningof excess of fat introduced by diet stimulating fatty acidoxidation Krebs cycle and respiratory chain complex activityand preventing a condition of oxidative damage
Uncoupling between mitochondrial respiration and ATPsynthesis observed in HF animals could be caused bya different expression of mitochondrial carrier proteinsinvolved in the regulation of proton conductance It is knownthat uncoupling proteins (UCPs) homologues uncouplemitochondrial respiration from oxidative phosphorylationincreasing thermogenesis while reducing the efficiency ofATP synthesis In a previous study [32] an increase in theexpression of the UCP2 isoform was found in rats withNAFLD Interestingly in this study we found a massiveincrease of this carrier protein inHF animals but the additionof KO to the diet was able to strongly reduce the expressionof hepatic UCP2 Therefore it is reasonable to assume thatthe addition of KO to the HF diet can increase the couplingbetween mitochondrial respiration and ATP synthesis by adownregulation of UCP2 expression
Not only are UCPs involved in mitochondrial uncou-pling butAAC is another carrier protein of particular interestin the understanding of the mitochondrial energy couplingprocess [33]This carrier catalyzes the exchange of ADP3minus forATP4minus and therefore utilizes as its driving force the electro-chemical potential generated across the inner membrane bythe respiratory chain The exchange reaction is electrogenicand driven by the electrical component of the proton gradi-ent so that entry of ADP into mitochondria and exit of ATPare favored We can therefore speculate that if mitochondriaare uncoupled and proton gradient was dissipated by theentry of H+ into the matrix by UCP2 an upregulation ofthe expression of AAC could compensate a reduced fluxof ATP from mitochondria when membrane potential wasreduced This hypothesized compensatory mechanism wasobserved in animals fed a HF diet The increase in theexpression of AAC was less evident in HF+KO animals inwhich livermitochondria respiratory efficiencywas unalteredin comparison to the control animals
In conclusion the present study indicates that KO coun-teracts the negative effects of a HF diet In particular our
10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology
8 BioMed Research International
0
100
200
300
500
Control HF
UCP
2 (
)
UCP2
400
lowast
lowast
HF+KO
(a)
0
60
120
180
300
AAC
()
AAC
240
Control HF
lowast
lowast
HF+KO
(b)
0
25
50
75
125
Porin
()
Porin
100
Control HF HF+KO
(c)
Figure 4 Effect of KO on proteins related to the respiratory chain function Effect of KO on proteins levels of UCP2 (a) and AAC (b) Theamount of porin an outer membrane protein was tested as a control (c) Liver mitochondrial proteins from control HF or HF+KO-fed ratswere separated by SDS-PAGE transferred to nitrocellulose and then immunodecorated with antisera against UCP2 AAC or the mammalianporinThe amount of proteins revealed by immunodecoration in the control groupwas set to 100The values reported in the figure representthe means plusmn SD (119899 = 5) lowast119875 lt 005 versus rats fed control diet 119875 lt 005 versus rats fed HF diet
fibrosis leading then to cirrhosis and hepatocellular carci-noma Some patients with NAFLD develop NASH throughpoorly understood mechanisms Since NAFLD and NASHare considered lifestyle-associated diseases diet interventionis an important approach to their treatment
In this context it is important to underline that theldquoobesogenicrdquoWestern diet has a ratio of fatty acids and carbo-hydrates of approximately 5050Moreover it is characterizedby an insufficient n-3 PUFAs consumption and a high n-6n-3 ratio which are associated with the progression fromsimple steatosis to NASH [8] along with modifications of gutmicrobiota [23]
Although the mechanisms responsible for increasedlipid accumulation in the liver are not yet fully elucidateddecreased capacity to oxidize fatty acids increased transportof fatty acids to liver and increased hepatic fatty acidsynthesis seem to be important factors involved in thepathogenesis of NAFLD [2 6 7 24]Moreover abnormalitiesin mitochondrial morphology and functions seem to beassociated with fatty liver [1ndash3 25ndash27] This finding suggeststhat regulation of mitochondria function and of molecularmechanisms involved in lipid accumulation in the liver couldprovide useful targets to prevent the development of NAFLD
In recent years the use of dietary supplements hasrapidly increased [2 6 10 24 28] with the aim to preventNAFLD KO a dietary supplement of n-3 PUFAs has becomeincreasingly popular because of its capability of reducing
hepatic lipogenesis and of stimulating catabolization ofexcess fat introduced by a HF diet [2 10] The analysisof hepatic transcriptome in mice demonstrated that KOsupplementation was able to regulate genes involved in hep-atic energy metabolism including glucose metabolism lipidbiosynthesis fatty acid metabolism cholesterol biosynthesisand mitochondrial electron transport more efficiently thanfish oil [10 12] These differential results were tentativelyascribed to the influence of the structural form of n-3PUFAs esterified to either phospholipids or triglycerideson a biological response Moreover PUFAs as phospholipids(KO) or triglycerides (fish oil)may differentially influence gutmicrobiota or synthesis of bacterial metabolites
A previous study by our group [2] suggested that KOsupplementation to a HF diet resulted in a retention of nor-malmitochondrial respiration efficiency whichwas impairedin HF animals The aim of the present study was thereforeto analyze the molecular mechanisms by which dietary fatsinfluence mitochondrial energetic metabolismThis new andintriguing aspect was evaluated in hepatocytes from rats feda HF diet or a HF diet supplemented with 25 KO with theaim to investigate the role of mitochondria in themodulationof nutrient metabolism exerted by KO
No change in PDH activity which essentially producesacetyl-CoA from carbohydrate was seen after administrationof HF or HF+KO diets Nevertheless fasting serum glucoseand insulin were higher after feeding a HF diet suggesting
BioMed Research International 9
an altered insulin sensitivity as indicated by HOMA indexchanges In fact an excess of energy supplied by fat corre-sponds to a scarce tissutal utilization of glucose thereby caus-ing hyperglycemia and hyperinsulinemia Insulin resistancewas reduced after KO supplementation
CPT I activity which is involved in fatty acid oxidationwas inhibited by a HF diet and stimulated after KO supple-mentationThe increase inCPT I activity observed inHF+KOgroup could be explained by the lower levels ofmalonyl-CoAametabolic intermediate of hepatic lipogenesis which in thiscase is instead strongly inhibited [2 10] Interestingly thecapability of KO to stimulate fatty acid oxidation seems tobe associated to the presence of a high lipid content (35)in the experimental diet used in this study In fact whenKO was added to a standard diet the expression of enzymesinvolved inmitochondrial fatty acid 120573-oxidation significantlydecreased [12]
The modulation of the CPT I activity observed in HFand HF+KO animals seems to be accompanied by a parallelmodulation of the activity of CS a Krebs cycle enzyme thatcatalyzes the synthesis of citrate fromoxaloacetate and acetyl-CoA Recently a strong and significant correlation of CSwith obesity was noted with a reduced enzyme activity inmitochondria of human omental adipose tissue in the stateof obesity [28] According to this observation the increasein CS activity revealed after KO supplementation could beexplained as a compensatory response to maintain the entryof acetyl-CoA produced by fatty acid oxidation into the Krebscycle It is interesting to underline that citrate generated byCS continues in the Krebs cycle In fact in animals fed aHF diet supplemented with KO a reduction in the activityand in the expression of citrate carrier protein (CIC) whichtransports citrate from mitochondria to the cytosol for fattyacid synthesis was observed [2 10 24]
NADH and FADH2generated by 120573-oxidation and Krebs
cycle are used in the final common oxidative phosphorylationsystem (OXPHOS) to generate ATP This process is coupledwith the transfer of electrons along the respiratory chainOur data indicate that a HF diet caused a strong decrease inthe mitochondrial respiratory efficiency and this effect couldbe due to a possible uncoupling effect exerted by the excessof fatty acids present in this diet In fact we found a littledecrease in 119881
3and a strong increase in 119881
4values in liver
mitochondria isolated fromHF animals1198814is the resting state
of mitochondrial respiration and an increase in this valuesuggests a stimulus ofmitochondrial respiration independentof ADP phosphorylation In other terms during HF feedinga lower coupling betweenmitochondrial respiration andATPsynthesis occurs Interestingly the addition of KO to the HFdiet almost completely reversed this effect thereby leadingmitochondria respiration efficiency to be comparable to thatof control rats
When we assayed in a more selective approach theactivity of single components of the respiratory chain weobserved a significant decrease in the activity of all respira-tory chain complexes in HF animals according to a recentstudy [29] inwhich an impairment ofmitochondrial functionand in particular of complex I activity was found AfterKO supplementation the values of complex activities were
identical to those measured in control animals Expressionof OXPHOS proteins significantly increased in HF animalscompared to the control group and this evidence was accom-panied by the presence of ATP levels in HF animals thatwere only slightly decreased in comparison to those foundin control animals According to this result previous studies[30 31] demonstrated that a high fat content in the dietinduced higher level of these proteins as a compensatoryresponse to the reduced activity of the respiratory chaincomplexes When we performed western blotting analysis inmitochondria isolated from HF+KO animals liver we founda higher level of the OXPHOS proteins compared to thecontrol and theHF groups probably caused by the synergisticeffect of the HF diet and the KO supplementation Indeedit is already known that genes involved in mitochondrialOXPHOS proteins expression were upregulated in miceconsuming KO as revealed from the transcriptomic analysis[12] Overall thismeans that KO is able to induce the burningof excess of fat introduced by diet stimulating fatty acidoxidation Krebs cycle and respiratory chain complex activityand preventing a condition of oxidative damage
Uncoupling between mitochondrial respiration and ATPsynthesis observed in HF animals could be caused bya different expression of mitochondrial carrier proteinsinvolved in the regulation of proton conductance It is knownthat uncoupling proteins (UCPs) homologues uncouplemitochondrial respiration from oxidative phosphorylationincreasing thermogenesis while reducing the efficiency ofATP synthesis In a previous study [32] an increase in theexpression of the UCP2 isoform was found in rats withNAFLD Interestingly in this study we found a massiveincrease of this carrier protein inHF animals but the additionof KO to the diet was able to strongly reduce the expressionof hepatic UCP2 Therefore it is reasonable to assume thatthe addition of KO to the HF diet can increase the couplingbetween mitochondrial respiration and ATP synthesis by adownregulation of UCP2 expression
Not only are UCPs involved in mitochondrial uncou-pling butAAC is another carrier protein of particular interestin the understanding of the mitochondrial energy couplingprocess [33]This carrier catalyzes the exchange of ADP3minus forATP4minus and therefore utilizes as its driving force the electro-chemical potential generated across the inner membrane bythe respiratory chain The exchange reaction is electrogenicand driven by the electrical component of the proton gradi-ent so that entry of ADP into mitochondria and exit of ATPare favored We can therefore speculate that if mitochondriaare uncoupled and proton gradient was dissipated by theentry of H+ into the matrix by UCP2 an upregulation ofthe expression of AAC could compensate a reduced fluxof ATP from mitochondria when membrane potential wasreduced This hypothesized compensatory mechanism wasobserved in animals fed a HF diet The increase in theexpression of AAC was less evident in HF+KO animals inwhich livermitochondria respiratory efficiencywas unalteredin comparison to the control animals
In conclusion the present study indicates that KO coun-teracts the negative effects of a HF diet In particular our
10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 9
an altered insulin sensitivity as indicated by HOMA indexchanges In fact an excess of energy supplied by fat corre-sponds to a scarce tissutal utilization of glucose thereby caus-ing hyperglycemia and hyperinsulinemia Insulin resistancewas reduced after KO supplementation
CPT I activity which is involved in fatty acid oxidationwas inhibited by a HF diet and stimulated after KO supple-mentationThe increase inCPT I activity observed inHF+KOgroup could be explained by the lower levels ofmalonyl-CoAametabolic intermediate of hepatic lipogenesis which in thiscase is instead strongly inhibited [2 10] Interestingly thecapability of KO to stimulate fatty acid oxidation seems tobe associated to the presence of a high lipid content (35)in the experimental diet used in this study In fact whenKO was added to a standard diet the expression of enzymesinvolved inmitochondrial fatty acid 120573-oxidation significantlydecreased [12]
The modulation of the CPT I activity observed in HFand HF+KO animals seems to be accompanied by a parallelmodulation of the activity of CS a Krebs cycle enzyme thatcatalyzes the synthesis of citrate fromoxaloacetate and acetyl-CoA Recently a strong and significant correlation of CSwith obesity was noted with a reduced enzyme activity inmitochondria of human omental adipose tissue in the stateof obesity [28] According to this observation the increasein CS activity revealed after KO supplementation could beexplained as a compensatory response to maintain the entryof acetyl-CoA produced by fatty acid oxidation into the Krebscycle It is interesting to underline that citrate generated byCS continues in the Krebs cycle In fact in animals fed aHF diet supplemented with KO a reduction in the activityand in the expression of citrate carrier protein (CIC) whichtransports citrate from mitochondria to the cytosol for fattyacid synthesis was observed [2 10 24]
NADH and FADH2generated by 120573-oxidation and Krebs
cycle are used in the final common oxidative phosphorylationsystem (OXPHOS) to generate ATP This process is coupledwith the transfer of electrons along the respiratory chainOur data indicate that a HF diet caused a strong decrease inthe mitochondrial respiratory efficiency and this effect couldbe due to a possible uncoupling effect exerted by the excessof fatty acids present in this diet In fact we found a littledecrease in 119881
3and a strong increase in 119881
4values in liver
mitochondria isolated fromHF animals1198814is the resting state
of mitochondrial respiration and an increase in this valuesuggests a stimulus ofmitochondrial respiration independentof ADP phosphorylation In other terms during HF feedinga lower coupling betweenmitochondrial respiration andATPsynthesis occurs Interestingly the addition of KO to the HFdiet almost completely reversed this effect thereby leadingmitochondria respiration efficiency to be comparable to thatof control rats
When we assayed in a more selective approach theactivity of single components of the respiratory chain weobserved a significant decrease in the activity of all respira-tory chain complexes in HF animals according to a recentstudy [29] inwhich an impairment ofmitochondrial functionand in particular of complex I activity was found AfterKO supplementation the values of complex activities were
identical to those measured in control animals Expressionof OXPHOS proteins significantly increased in HF animalscompared to the control group and this evidence was accom-panied by the presence of ATP levels in HF animals thatwere only slightly decreased in comparison to those foundin control animals According to this result previous studies[30 31] demonstrated that a high fat content in the dietinduced higher level of these proteins as a compensatoryresponse to the reduced activity of the respiratory chaincomplexes When we performed western blotting analysis inmitochondria isolated from HF+KO animals liver we founda higher level of the OXPHOS proteins compared to thecontrol and theHF groups probably caused by the synergisticeffect of the HF diet and the KO supplementation Indeedit is already known that genes involved in mitochondrialOXPHOS proteins expression were upregulated in miceconsuming KO as revealed from the transcriptomic analysis[12] Overall thismeans that KO is able to induce the burningof excess of fat introduced by diet stimulating fatty acidoxidation Krebs cycle and respiratory chain complex activityand preventing a condition of oxidative damage
Uncoupling between mitochondrial respiration and ATPsynthesis observed in HF animals could be caused bya different expression of mitochondrial carrier proteinsinvolved in the regulation of proton conductance It is knownthat uncoupling proteins (UCPs) homologues uncouplemitochondrial respiration from oxidative phosphorylationincreasing thermogenesis while reducing the efficiency ofATP synthesis In a previous study [32] an increase in theexpression of the UCP2 isoform was found in rats withNAFLD Interestingly in this study we found a massiveincrease of this carrier protein inHF animals but the additionof KO to the diet was able to strongly reduce the expressionof hepatic UCP2 Therefore it is reasonable to assume thatthe addition of KO to the HF diet can increase the couplingbetween mitochondrial respiration and ATP synthesis by adownregulation of UCP2 expression
Not only are UCPs involved in mitochondrial uncou-pling butAAC is another carrier protein of particular interestin the understanding of the mitochondrial energy couplingprocess [33]This carrier catalyzes the exchange of ADP3minus forATP4minus and therefore utilizes as its driving force the electro-chemical potential generated across the inner membrane bythe respiratory chain The exchange reaction is electrogenicand driven by the electrical component of the proton gradi-ent so that entry of ADP into mitochondria and exit of ATPare favored We can therefore speculate that if mitochondriaare uncoupled and proton gradient was dissipated by theentry of H+ into the matrix by UCP2 an upregulation ofthe expression of AAC could compensate a reduced fluxof ATP from mitochondria when membrane potential wasreduced This hypothesized compensatory mechanism wasobserved in animals fed a HF diet The increase in theexpression of AAC was less evident in HF+KO animals inwhich livermitochondria respiratory efficiencywas unalteredin comparison to the control animals
In conclusion the present study indicates that KO coun-teracts the negative effects of a HF diet In particular our
10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 BioMed Research International
Acetyl-CoA
Krebscycle
Pyruvate
Glucose Fatty acids
Acyl-CoA
Respiratory chaincomplexes
Q
ATP
ADP
AACIII IVI
ATP
UCP2
ADP ATP
ATP-syn
H+
H+
120573-oxidation
FADH2 Pi + ADP
4H+ 4H+ 2H+
NADH + H+
c
Figure 5 Mitochondrial pathways influenced by the addition of KO to a HF diet 119888 cytochrome 119888 Q ubiquinone
results contribute to elucidate the molecular mechanism(Figure 5) by which KO positively influences manymetabolicsteps and ameliorates mitochondrial dysfunctions causedby the administration of a hyperlipidic diet which oftencharacterizes the nutritional habits of western populationsNevertheless additional studies in humans are required toconfirm this view since rodents may respond to the dietarymacronutrients composition in a different manner thanhuman subjects
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
References
[1] Y Wei R S Rector J P Thyfault and J A Ibdah ldquoNonalco-holic fatty liver disease and mitochondrial dysfunctionrdquoWorldJournal of Gastroenterology vol 14 no 2 pp 193ndash199 2008
[2] A Ferramosca A Conte L Burri et al ldquoA krill oil supple-mented diet suppresses hepatic steatosis in high-fat fed ratsrdquoPLoS ONE vol 7 no 6 Article ID e38797 2012
[3] C Garcıa-Ruiz A Baulies M Mari P M Garcıa-Roves andJ C Fernandez-Checa ldquoMitochondrial dysfunction in non-alcoholic fatty liver disease and insulin resistance cause orconsequencerdquo Free Radical Research vol 47 no 11 pp 854ndash868 2013
[4] M Flamment J Rieusset H Vidal et al ldquoRegulation ofhepatic mitochondrial metabolism in response to a high fatdiet a longitudinal study in ratsrdquo Journal of Physiology andBiochemistry vol 68 no 3 pp 335ndash344 2012
[5] M Asrih and F R Jornayvaz ldquoDiets and nonalcoholic fatty liverdisease the good and the badrdquo Clinical Nutrition vol 33 no 2pp 186ndash190 2014
[6] A Ferramosca and V Zara ldquoModulation of hepatic steatosis bydietary fatty acidsrdquo World Journal of Gastroenterology vol 20no 7 pp 1746ndash1755 2014
[7] A Ferramosca A Conte F Damiano L Siculella and V ZaraldquoDifferential effects of high-carbohydrate and high-fat diets onhepatic lipogenesis in ratsrdquo European Journal of Nutrition vol53 no 4 pp 1103ndash1114 2014
[8] G S Lee J S Yan R K Ng S Kakar and J J MaherldquoPolyunsaturated fat in the methionine-choline-deficient dietinfluences hepatic inflammation but not hepatocellular injuryrdquoJournal of Lipid Research vol 48 no 8 pp 1885ndash1896 2007
BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 11
[9] S Tandy R W S Chung E Wat et al ldquoDietary krill oil supple-mentation reduces hepatic steatosis glycemia and hypercholes-terolemia in high-fat-fedmicerdquo Journal of Agricultural and FoodChemistry vol 57 no 19 pp 9339ndash9345 2009
[10] A Ferramosca L Conte and V Zara ldquoA krill oil supplementeddiet reduces the activities of the mitochondrial tricarboxylatecarrier and of the cytosolic lipogenic enzymes in ratsrdquo Journalof Animal Physiology and Animal Nutrition vol 96 no 2 pp295ndash306 2012
[11] L Burri N Hoem S Banni and K Berge ldquoMarine omega-3phospholipids metabolism and biological activitiesrdquo Interna-tional Journal of Molecular Sciences vol 13 no 11 pp 15401ndash15419 2012
[12] L Burri K Berge K Wibrand R K Berge and J L BargerldquoDifferential effects of krill oil and fish oil on the hepatictranscriptome in micerdquo Frontiers in Genetics vol 2 article 452011
[13] E Bonora G Targher M Alberiche et al ldquoHomeostasis modelassessment closely mirrors the glucose clamp technique in theassessment of insulin sensitivity studies in subjects with variousdegrees of glucose tolerance and insulin sensitivityrdquo DiabetesCare vol 23 no 1 pp 57ndash63 2000
[14] A Ferramosca V Savy L Conte S Colombo A W CEinerhand and V Zara ldquoConjugated linoleic acid and hepaticlipogenesis in mouse role of the mitochondrial citrate carrierrdquoJournal of Lipid Research vol 47 no 9 pp 1994ndash2003 2006
[15] M M Bradford ldquoA rapid and sensitive method for the quanti-tation of microgram quantities of protein utilizing the principleof protein dye bindingrdquoAnalytical Biochemistry vol 72 no 1-2pp 248ndash254 1976
[16] C Crewe M Kinter and L I Szweda ldquoRapid inhibition ofpyruvate dehydrogenase an initiating event in high dietary fat-induced loss of metabolic flexibility in the heartrdquo PLoS ONEvol 8 no 10 Article ID e77280 2013
[17] MV ZakharchenkoAV ZakharchenkoNVKhunderyakovaet al ldquoBurst of succinate dehydrogenase and 120572-ketoglutaratedehydrogenase activity in concert with the expression of genescoding for respiratory chain proteins underlies short-term ben-eficial physiological stress in mitochondriardquo The InternationalJournal of Biochemistry amp Cell Biology vol 45 no 1 pp 190ndash200 2013
[18] A Ferramosca V Savy and V Zara ldquoOlive oil increases thehepatic triacylglycerol content in mice by a distinct influenceon the synthesis and oxidation of fatty acidsrdquo BioscienceBiotechnology and Biochemistry vol 72 no 1 pp 62ndash69 2008
[19] A Ferramosca R Focarelli P Piomboni L Coppola andV Zara ldquoOxygen uptake by mitochondria in demembranatedhuman spermatozoa a reliable tool for the evaluation of spermrespiratory efficiencyrdquo International Journal of Andrology vol31 no 3 pp 337ndash345 2008
[20] A Barrientos F Fontanesi and F Dıaz ldquoEvaluation of themitochondrial respiratory chain and oxidative phosphorylationsystem using polarography and spectrophotometric enzymeassaysrdquo inCurrent Protocols in Human Genetics chapter 19 unit193 2009
[21] Y Chaban E J Boekema and N V Dudkina ldquoStructures ofmitochondrial oxidative phosphorylation supercomplexes andmechanisms for their stabilisationrdquo Biochimica et BiophysicaActamdashBioenergetics vol 1837 no 4 pp 418ndash426 2014
[22] B A Neuschwander-Tetri ldquoCarbohydrate intake and nonalco-holic fatty liver diseaserdquo Current Opinion in Clinical Nutritionand Metabolic Care vol 16 no 4 pp 446ndash452 2013
[23] B D Pachikian A M Neyrinck L Portois et al ldquoInvolvementof gut microbial fermentation in the metabolic alterationsoccurring in n-3 polyunsaturated fatty acids-depleted micerdquoNutrition amp Metabolism vol 8 no 1 article 44 2011
[24] A Ferramosca andV Zara ldquoDietary fat and hepatic lipogenesismitochondrial citrate carrier as a sensor of metabolic changesrdquoAdvances in Nutrition vol 5 no 3 pp 217ndash225 2014
[25] G Vendemiale I Grattagliano P Caraceni et al ldquoMitochon-drial oxidative injury energy metabolism alteration in rat fattyliver effect of the nutritional statusrdquo Hepatology vol 33 no 4pp 808ndash815 2001
[26] S K Mantena D P Vaughn Jr K K Andringa et al ldquoHighfat diet induces dysregulation of hepatic oxygen gradients andmitochondrial function in vivordquo Biochemical Journal vol 417no 1 pp 183ndash193 2009
[27] K Begriche J Massart M A Robin F Bonnet and BFromenty ldquoMitochondrial adaptations and dysfunctions innonalcoholic fatty liver diseaserdquo Hepatology vol 58 no 4 pp1497ndash1507 2013
[28] A Ferramosca V Savy L Conte and V Zara ldquoDietarycombination of conjugated linoleic acid (CLA) and pine nut oilprevents CLA-induced fatty liver in micerdquo Journal of Agricul-tural and Food Chemistry vol 56 no 17 pp 8148ndash8158 2008
[29] T Garnol R Endlicher O Kucera Z Drahota and ZCervinkova ldquoImpairment of mitochondrial function of rathepatocytes by high fat diet and oxidative stressrdquo PhysiologicalResearch vol 63 no 2 pp 271ndash274 2014
[30] J Hoeks J de Wilde M F M Hulshof et al ldquoHigh fat diet-induced changes inmousemusclemitochondrial phospholipidsdo not impair mitochondrial respiration despite insulin resis-tancerdquo PLoS ONE vol 6 no 11 Article ID e27274 2011
[31] C Ruggiero M Ehrenshaft E Cleland and K Stadler ldquoHigh-fat diet induces an initial adaptation of mitochondrial bioen-ergetics in the kidney despite evident oxidative stress andmitochondrial ROS productionrdquo The American Journal ofPhysiologymdashEndocrinology and Metabolism vol 300 no 6 ppE1047ndashE1058 2011
[32] Y Jiang H Zhang L-Y Dong DWang andW An ldquoIncreasedhepatic UCP2 expression in rats with nonalcoholic steato-hepatitis is associated with upregulation of Sp1 binding to itsmotif within the proximal promoter regionrdquo Journal of CellularBiochemistry vol 105 no 1 pp 277ndash289 2008
[33] P Schonfeld M R Wieckowski and L Wojtczak ldquoThyroidhormone-induced expression of the ADPATP carrier and itseffect on fatty acid-induced uncoupling of oxidative phospho-rylationrdquo FEBS Letters vol 416 no 1 pp 19ndash22 1997
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology





























