Embed Size (px)
Citation preview

R
La
b
a
ARRA
KPPPREDP
I
e(to2(gPAoBtaWbPm(t
ET
0d
European Journal of Cell Biology 90 (2011) 844– 853
Contents lists available at ScienceDirect
European Journal of Cell Biology
jou rn al homepage: www.elsev ier .de /e jcb
NAi knockdown of parafusin inhibits the secretory pathway
i Liua, Elzbieta Wyrobab, Birgit H. Satira,∗
Department of Anatomy and Structural Biology, Albert Einstein College of Medicine, New York, USADepartment of Cell Biology, Nencki Institute of Experimental Biology, Warsaw, Poland
r t i c l e i n f o
rticle history:eceived 20 July 2009eceived in revised form 27 May 2011ccepted 11 June 2011
eywords:arafusin
a b s t r a c t
Several glycolytic enzymes and their isoforms have been found to be important in cell signaling unre-lated to glycolysis. The involvement of parafusin (PFUS), a member of the phosphoglucomutase (PGM)superfamily with no phosphoglucomutase activity, in Ca2+-dependent exocytosis has been controversial.This protein was first described in Paramecium tetraurelia, but is widely found. Earlier work showed thatparafusin is a secretory vesicle scaffold component with unusual post-translational modifications (cyclicphosphorylation and phosphoglucosylation) coupled to stages in the exocytic process. Using RNAi, we
FUSarameciumNAixocytosisCSVhosphoglumutase
demonstrate that parafusin synthesis can be reversibly blocked, with minor or no effect on other PGMisoforms. PFUS knockdown produces an inhibition of dense core secretory vesicle (DCSV) synthesis lead-ing to an exo− phenotype. Although cell growth is unaffected, vesicle content is not packaged properlyand no new DCSVs are formed. We conclude that PFUS and its orthologs are necessary for proper scaffoldmaturation. Because of this association, parafusin is an important signaling component for regulatorycontrol of the secretory pathway.
ntroduction
Parafusin (PFUS), first described in Paramecium tetraurelia (Satirt al., 1989) has a 51% sequence identity to phosphoglucomutasePGM), with 4 insertions and a longer N-terminal as comparedo rabbit muscle PGM (Subramanian et al., 1994). PFUS or itsrthologs are present in most eukaryotic cells (Wyroba and Satir,000; Wyroba et al., 1995). PFUS and PRP1, its Toxoplasma orthologMatthiesen et al., 2001, 2003) have been shown to be phospho-lycoproteins and members of PGM superfamily with little or noGM enzymatic activity (Satir et al., 1989; Subramanian et al., 1994;ndersen et al., 1994; Levin et al., 1999). Similarly other isoformsf PGM in various organisms have little PGM activity (Belkin andurridge, 1995; Fu et al., 2000; Narayanan and Xu, 1997). Like cer-ain isoforms of other glycolytic enzymes including GAPDH andldolase (Daubenberger et al., 2003; Hessler et al., 1998; Rogalski-ilk and Cohen, 1997), these non-enzymatic forms of PGM may
e involved in cell signaling processes, rather than in glycolysis.FUS and PRP1 function via post-translational modifications in
embrane scaffolds of respectively, dense core secretory vesiclesDCSVs) in Paramecium (Zhao and Satir, 1998) and micronemes,he Toxoplasma secretory vesicles to facilitate Ca2+-dependent pro-
∗ Corresponding author at: Department of Anatomy & Structural Biology, Albertinstein College of Medicine, 1300 Morris Park Ave., Bronx, NY 10461, USA.el.: +1 718 430 4063; fax: +1 718 430 8577.
E-mail address: [email protected] (B.H. Satir).
171-9335/$ – see front matter © 2011 Elsevier GmbH. All rights reserved.oi:10.1016/j.ejcb.2011.06.002
© 2011 Elsevier GmbH. All rights reserved.
cesses related to regulated exocytosis (Matthiesen et al., 2001,2003; Liu et al., 2009). When cells are stimulated to exocytose,in conjunction with a Ca2+-dependent dephosphoglucosylation,PFUS/PRP1 dissociates from the vesicle scaffold and can no longerbe specifically localized. PFUS/PRP1 re-associates with newly form-ing vesicles in the cytosol. Isolated DCSVs contain glycosylated PFUSin their scaffold (Liu et al., 2009).
Neither exocytosis, dephosphoglucosylation (Subramanian andSatir, 1992) or dissociation of the PFUS of the DCSV scaffold (Satirand Zhao, 1999) is seen in the temperature sensitive exo− mutantcells (nd9 – 27 ◦C) of Paramecium after stimulation at the non-permissive temperature. The nd9 gene product was found to bea novel protein with armadillo-like repeats that may play a rolein possible protein–protein interactions, like the SNARE proteins(Froissard et al., 2001).
In a recent study using a heterologous system, Alexa-labeledHis-PRP1 electroporated into living wild type Paramecium associ-ated with DCSVs and co-localized with PFUS upon fixation. Uponexocytosis PRP1 dissociated from the DCSV only to re-associate asnew organelles formed. Association of PRP1 with the vesicle scaf-fold could be blocked by site-specific mutagenesis of PRP1 (Liu et al.,2009). Taken together, these results suggest that PFUS is a signifi-cant component for proper scaffold formation around ParameciumDCSVs, and that after scaffold formation and organelle docking, the
correct covalent modification of PFUS is essential for membranefusion and exocytosis.The importance of PFUS as both a component of the DCSV scaf-fold and a crucial component involved in exocytosis, but not with

L. Liu et al. / European Journal of Cell Biology 90 (2011) 844– 853 845
Fig. 1. 2D gel analysis of Paramecium PGM superfamily members. (A) Coomassie blue stained 2-D gel of immunoprecipitated PGM superfamily members with a pan-PGMa 63 kDb US ants e note
gmaistfactmg
M
C
(a1wK
A
twsIDH
ntibody from wt exo+ cells (left panel) in comparison to nd9 cells (right panel). Six ∼lot: Western blot analysis of the above 2D gels for wt and nd9 cells probed with PFpots at pI 6.7 and 6.5. (C) Intensity ratio of the spots at pI 6.5/6.7 shown in B. Pleas
lycolysis, has been a somewhat controversial idea. Now, RNAiethods (Bastin et al., 2001; Galvani and Sperling, 2002) have
llowed us to establish that independent of other PGM superfam-ly members, PFUS knockdown produces an inhibition of DCSVynthesis leading to an exo− phenotype. These results supporthe conclusion that proper incorporation of PFUS into the scaf-old is necessary for maintenance and maturation of the DCSVsnd subsequently for Ca2+ regulated exocytosis. Orthologs of PFUSould serve similar roles in other cells. The results also suggesthat, via even small changes or a post-translational modification,
embers of the PGM superfamily, can assume roles unrelated tolycolysis.
aterials and methods
ells, media, and culture conditions
Axenic cultures of wt Paramecium tetraurelia or mutant nd9Sonneborn, 1974) and nd6 (Lefort-Tran et al., 1981) were grownt 27 ◦C in Cephalin-Fatty Acid (CFA) media (Satir and Bleyman,993) to early stationary phase, harvested (400 g, 30 s) andashed twice in Mg2+-buffer (10 mM MgCl2, 5 mM Tris–HCl, 1 mMCl, pH 7.0).
ntibodies
Pan-PGM antibody is a rabbit polyclonal antibody raised against
he isolated parafusin (Murtaugh et al., 1987). PFUS antibodyas raised against the synthetic peptide (CDYEFEKHINLDQ) corre-ponding to the insertion-2 region of PFUS by Immuno-dynamics,nc. T8 antibody was a monoclonal antibody raised againstCSV content T8, courtesy from Dr. Richard Allen, University ofawaii.
a spots at pI 6.9–6.4 are present. Arrows indicate spots at pI 6.7 and 6.5. (B) Westernibody reveals that only two of the six spots are recognized, arrows indicate the two
that the intensity ratio (n = 3) of these spots is 0.75 for wt and 2.1 for nd9-27 cells.
Preparation of cell homogenate and immunoprecipitation
Collected and washed Paramecium cells were mechanicallylysed on ice in glass homogenizer in lysis buffer (50 mM Tris–HClpH 7.5, 150 mM NaCl, 1% Triton X-100, 5 mM EDTA, 2 �M leu-peptin, 1.5 �M antipain, 0.5 mM PMSF, 0.012 TIU/ml aprotinin,5 mM Na3VO4) Homogenate was centrifuged (12,000 × g, 5 min,4 ◦C) twice and the supernatant (1 ml) was collected. Pan-PGM anti-body (1:125) was added into the supernatant and the mixture wasincubated at room temperature for 1 h. 50 �l of Protein G beads(Sigma, St. Louis, MO) were washed with lysis buffer and trans-ferred into the homogenate–antibody mixture. After incubation(30 min, 4 ◦C on shaker), the beads were washed (with lysis buffer,1500 g, 2 min, 4 ◦C) three times and collected.
Two-dimensional electrophoresis and Western blot
The proteins were resolved according to the manufacturer’smanual for the Multiphor II horizontal two-dimensional gel elec-trophoresis system (Amersham Pharmacia Biotech, Piscataway, NJ,USA). The first dimension (isoelectric focusing) was performed withImmobiline Dry Strip, pH 3–10 linear pH gradient and the sec-ond dimension (SDS-PAGE) was performed with ExcelGel SDS XL12–14% Tris–glycine. The gel was either stained with Coomassieblue, or for western blot, transferred to nitrocellulose membrane(0.8 mA/cm2 membrane, 1 h) by Semi-dry Blotting System (IMM-1,W.E.P. Co. Seattle, WA, USA). The membrane was blocked with 5%milk in washing buffer (10 mM Tris–HCl, 0.9% NaCl, pH 7.3) for 1 h,rinsed with 0.5% milk three times, and then incubated with 1:1000PFUS antibody for 2 h. The membrane was washed three times with
washing buffer and incubated for 1.5 h in 1:3000 HRP-conjugatedgoat anti-rabbit secondary antibody (Chemicon, Billerica, MA, USA).Membrane was then washed three times and developed with ECL(GE Healthcare, Little Chalfont, UK).
846 L. Liu et al. / European Journal of Cell Biology 90 (2011) 844– 853
F m of
e rt (DaP show
fI1
G
OabrIwdoa2iasgaPrDfath
ig. 2. Southern blot analysis of parafusin (PGM/PFUS superfamily genes. (A) Diagranzymes: Bgl II, Eco RI, and Hind III are shown. Note that the Eco RI site is in the insearamecium DNA cut with Bgl II, Eco RI or Hind III and probed with the 1.6 kb probe
Quantification on scanned-in Western blot images was per-ormed using ImageJ (Rasband, W.S., ImageJ, U.S. Nationalnstitutes of Health, Bethesda, MD, USA, http://rsb.info.nih.gov/ij/,997–2009).
ene silencing by feeding
N-PFUS (PstI and BglII), C-PFUS (NcoI and BglII) and the wholeRF of PFUS (W-PFUS, GenBank Accession #AY970820.1) (PstInd NcoI) were subcloned into the LITMUS 28i feeding vectoretween two T7 promoters by the restriction sites indicated. Theesulting plasmids were transformed into HT115 (DE3), an RNaseII-deficient strain of Escherichia coli (Timmons et al., 2001). Bacteria
ere fed to wt Paramecium using the following feeding proce-ure. HT115(DE3) bacteria containing vector with RNAi constructsr control vector were grown in LBAT (LB plate with 50 �g/mlmpicillin and 10 �g/ml Tetracyclin) plate before inoculated into5 LBA (LB liquid media with 50 �g/ml ampicillin). Bacteria were
nduced with IPTG (125 ng/ml) to reach the density at OD600 = 0.4nd induced for next 2–4 h, centrifuged (10 min, 3000 rpm) and re-uspended in the same volume of N medium (filtered 1 g/l wheatrass powder after boiling; 5 mM Tris–HCl, 4 mM Na2HPO4·12H2O,nd 1.5 mM NaH2PO4·H2O). This is 10× feeding bacteria. 3-Dayaramecium cells were washed in Dryl’s Buffer (0.1 M sodium cit-ate, 0.1 M NaH2PO4, 0.1 M Na2HPO4, 0.1 M CaCl2), and purged 2 h inryl’s Buffer. After next wash with Dryl’s Buffer, cells were trans-
erred to 20 ml sterile N plus medium (N medium with 50 �g/mlmpicillin, 2.5 mM IPTG, 8 �g/ml stigmasterol and 1× feeding bac-eria) to final concentration of 500–1000 cells/ml and grown inumid chamber in dark at 27 ◦C. Phenotypes were observed every
PFUS genomic DNA and the probe used. The positions of the cut sites for restrictionubenberger et al., 2003) that is not found in PGM. (B) Southern blot analysis of totaln above.
24 h. Feed Paramecium cells were fed with freshly plated andinduced bacteria every 24 h so long as the phenotype was observed.
Negative controls used HT115(DE3) carrying the LITMUS 28iplasmid without insertion. For recovery, cells at 72 h in the RNAiwere collected and fed with bacteria containing no RNAi vector.
Indirect immunofluorescence staining
Paramecium cells were collected, washed twice (Mg2+-buffer),fixed (3% paraformaldehyde in PBS, 30 min), and permeabilized(1% Triton X-100 in PBS, 15 min). Cells were blocked in PBST (5%BSA and 0.3% Tween-20 in PBS) for 30 min, and then incubatedwith PFUS antibody (1:300) in PBST for 2 h. After washed twicewith PBST cells were incubated (1 h) with Alexa488 labeled sec-ondary antibody, goat anti-rabbit IgG (Invitrogen, Carlsbad, CA) inPBST 1:400. Cells were then washed three times, mounted in 15 �l50% glycerol in PBS onto glass slides and observed under BioRadRadiance 2000 Laser Scanning Confocal Microscope. For double-labeling, mixed primary antibodies (mouse and rabbit) were usedfor primary antibody incubation step and mixed secondary anti-bodies (goat anti-mouse IgG, goat anti-rabbit IgG) were used forsecondary antibody incubation step.
Induction of exocytosis by secretagogue
Exocytosis was induced with 100 �M lysozyme as modified afterHarumoto (Harumoto, 1994; Hennessey et al., 1995).
In order to demonstrate molecularly that PFUS is important forsecretion and exocytosis, we have performed a series of RNAi exper-

l of Ce
i–
iP(uPla
S
tawtMoanel
R
sPo(ibtiS
piwtsgottNtsim
L. Liu et al. / European Journa
ments to demonstrate that a phenotype – inhibition of exocytosis is associated with loss of PFUS.
Three different regions of the PFUS sequence were used for RNAnterference: the C-terminal (C-PFUS, 0.8 kb), the N-terminal (N-FUS, 1.0 kb) and the entire open reading frame (W-PFUS, 1.8 kb)Fig. 2A). As a control the empty RNAi vector (LITMUS 28i) wassed. PFUS in pGEM vector was digested by PstI and NcoI (W-PFUS),stI and BglII (N-PFUS) or BglII and NcoI (C-PFUS). They were eachigated into LITMUS 28i respectively between the two T7 promotorss shown on the scheme below:
outhern hybridization analysis
Paramecium total DNA was isolated and digested with restric-ion enzymes: EcoRI, HindIII and BglII, electrophoresed and blotteds described previously (Subramanian et al., 1994). Hybridizationas performed at 55 ◦C with PCR amplified parafusin cDNA as
he probe that was labeled with digoxigenin-dUTP (Boehringer-anheim) by nick translation method followed by purification
n Bio-Spin chromatography columns (Bio-Rad) as in Wyroband Satir (2000). Southern blot analysis was performed using theon-radioactive detection system as described previously (Wyrobat al., 1995) with the CDP-Star (Boehringer-Mannheim) as a chemi-uminescent substrate.
esults
Paramecium tetraurelia contains several members of the PGMuperfamily in addition to PFUS (Hauser et al., 1997). A pan-GM antibody (made against isolated PFUS) recognizes PFUS andther PGM superfamily members while a specific PFUS antibodysynthetic peptide antibody to insertion 2 in PFUS, not presentn mammalian PGM), recognizes only PFUS. Unlike PGM, PFUSecomes post-translationally modified by two covalent modifica-ions: phosphorylation or phosphoglucosylation, via pathways thatncorporate 32Pi into the molecule in living cells (Subramanian andatir, 1992; Gilligan and Satir, 1982).
To establish how many isoforms of the PGM superfamily wereresent in this cell and which of these were PFUS, immunoprecip-
tation experiments were performed using the pan-PGM antibodyith wt (exo+). The immunoprecipitates were subjected to 2D elec-
rophoresis. Six distinct spots, ranging from pI ∼ 6.2 to 6.8 at veryimilar molecular weights ∼63 kDa, are seen in Coomassie stainedels (Fig. 1A). However, in all gels the PFUS antibody recognizednly two spots, pIs ∼ 6.7 and 6.5 (Fig. 1B). These results suggest thathese two isoforms represent PFUS, probably with different post-ranslational modifications related to the phosphorylation cycle.ote that the spot patterns are similar in wt and nd9, however,
he relative intensity of the two spots on immunoblots at pI 6.5/6.7hown in Fig. 1B is different (Fig. 1C). Their ratio is over 2-fold highern the nd9/27 mutant than in wt suggesting that these species
ay represent the phosphoglycosylated form of parafusin since
ll Biology 90 (2011) 844– 853 847
mutant blocked both in exocytosis and dephosphoglycosylation(Subramanian and Satir, 1992).
Southern hybridization analysis was used to analyze PGM/PFUS-related genes in Paramecium. This was performed on the total DNAdigested with the Eco RI, Bgl II or Hind III, respectively and analyzedwith the 1.6 kb cloned PFUS cDNA as the probe (Subramanian et al.,1994) as shown in Fig. 2A and B. There is only one restriction site forall these enzymes in the open reading frame (ORF) of the parafusingene at position 553, 1081 and 1687,
respectively (Fig. 2A). There are four hybridizing DNA fragmentsin EcoRI DNA digest and three in Hind III – cut DNA. Bgl II siteis located almost in the middle of ORF, so if one gene exists,this should yield two DNA fragments in Southern hybridization.Analysis revealed eight distinct DNA fragments ranging from12.1 kb to 1 kb in size hybridizing to the probe that suggests thepresence of four copies of related PFUS/PGM genes in Paramecium.These genes in Paramecium characterized by the presence ofone restriction site for Bgl II, besides parafusin are: P. tetraureliaphosphoglucomutase 1 (Accession #Y09969) identified by Hauseret al. (1997) and two other genes related to phosphoglucomutaseand described as coding for hypothetical proteins from the Geno-scope database (www.genoscope.cns.fr) GSPATT00032405001(Accession #XM 001429534) and GSPATT00023467001(Accession #XM 001457652) The protein product (Accession#XP 001429571.1) of GSPATT00032405001 shows 99% identityto parafusin. The presence of these multiple PGM/PFUS-relatedgenes was further confirmed by comparison of their introns (datanot shown) and may explain the presence of four hybridizing DNAfragments (Fig. 2B) after Eco RI digestion since they possess singlerestriction site for this enzyme.
In order to demonstrate that PFUS is necessary for exocy-tosis, we have used RNAi. In Paramecium, RNAi silencing usingthe entire open reading frame of various proteins, as originallydescribed in plants and fungi, has been applied successfully tostudy gene knockdowns (Bastin et al., 2001; Galvani and Sperling,2002; Elbashir et al., 2001; Froissard et al., 2002; Meyer andCohen, 1999; Wassmer et al., 2005). RNA interference is gener-ated by feeding. Briefly, as outlined in the Paramecium database(http://Paramecium.cgm.cnrs-gif.fr/RNAi), cDNA constructs weredesigned based on three different regions of the PFUS sequence:the C-terminal (0.8 kb), the N-terminal (1.0 kb) and the entire openreading frame (1.8 kb) for insertion into the LITMUS 28i vector andsubsequent transfection in E. coli strain HT115 (DE3) as describedin “Materials and methods” section. The bacteria were grown inselective media and vector incorporation was tested by PCR. In
the bacteria, the DNAs generate double stranded RNAs which sur-vive because the RNase is inactivated by a deletion. Then the wtParamecium culture, washed in inorganic medium, was inoculatedinto the vector-containing bacteria and the ciliate was allowed to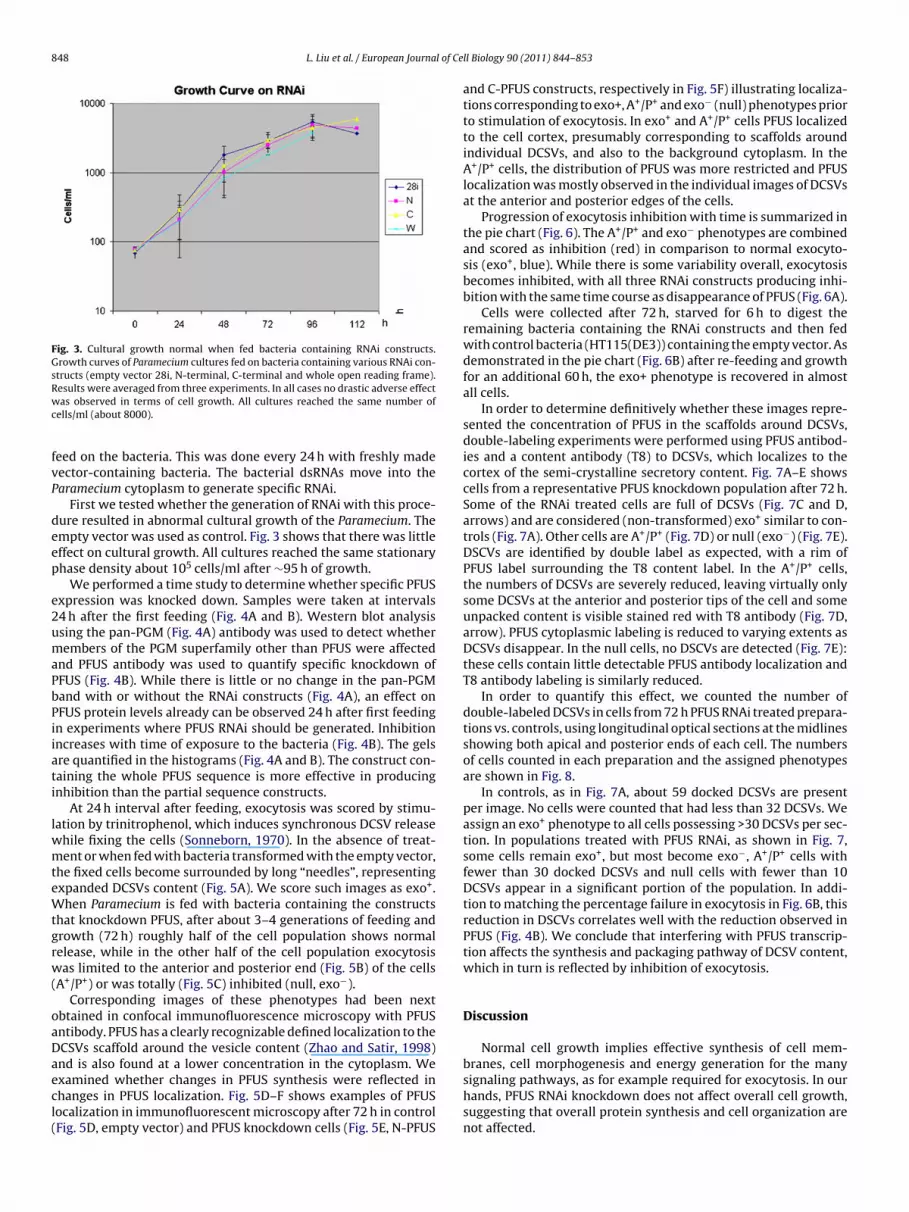
848 L. Liu et al. / European Journal of Ce
Fig. 3. Cultural growth normal when fed bacteria containing RNAi constructs.Growth curves of Paramecium cultures fed on bacteria containing various RNAi con-structs (empty vector 28i, N-terminal, C-terminal and whole open reading frame).Results were averaged from three experiments. In all cases no drastic adverse effectwc
fvP
deep
e2umaPbPiiati
lwmteWtgrw(
oaDaecl(
as observed in terms of cell growth. All cultures reached the same number ofells/ml (about 8000).
eed on the bacteria. This was done every 24 h with freshly madeector-containing bacteria. The bacterial dsRNAs move into thearamecium cytoplasm to generate specific RNAi.
First we tested whether the generation of RNAi with this proce-ure resulted in abnormal cultural growth of the Paramecium. Thempty vector was used as control. Fig. 3 shows that there was littleffect on cultural growth. All cultures reached the same stationaryhase density about 105 cells/ml after ∼95 h of growth.
We performed a time study to determine whether specific PFUSxpression was knocked down. Samples were taken at intervals4 h after the first feeding (Fig. 4A and B). Western blot analysissing the pan-PGM (Fig. 4A) antibody was used to detect whetherembers of the PGM superfamily other than PFUS were affected
nd PFUS antibody was used to quantify specific knockdown ofFUS (Fig. 4B). While there is little or no change in the pan-PGMand with or without the RNAi constructs (Fig. 4A), an effect onFUS protein levels already can be observed 24 h after first feedingn experiments where PFUS RNAi should be generated. Inhibitionncreases with time of exposure to the bacteria (Fig. 4B). The gelsre quantified in the histograms (Fig. 4A and B). The construct con-aining the whole PFUS sequence is more effective in producingnhibition than the partial sequence constructs.
At 24 h interval after feeding, exocytosis was scored by stimu-ation by trinitrophenol, which induces synchronous DCSV release
hile fixing the cells (Sonneborn, 1970). In the absence of treat-ent or when fed with bacteria transformed with the empty vector,
he fixed cells become surrounded by long “needles”, representingxpanded DCSVs content (Fig. 5A). We score such images as exo+.hen Paramecium is fed with bacteria containing the constructs
hat knockdown PFUS, after about 3–4 generations of feeding androwth (72 h) roughly half of the cell population shows normalelease, while in the other half of the cell population exocytosisas limited to the anterior and posterior end (Fig. 5B) of the cells
A+/P+) or was totally (Fig. 5C) inhibited (null, exo−).Corresponding images of these phenotypes had been next
btained in confocal immunofluorescence microscopy with PFUSntibody. PFUS has a clearly recognizable defined localization to theCSVs scaffold around the vesicle content (Zhao and Satir, 1998)nd is also found at a lower concentration in the cytoplasm. We
xamined whether changes in PFUS synthesis were reflected inhanges in PFUS localization. Fig. 5D–F shows examples of PFUSocalization in immunofluorescent microscopy after 72 h in controlFig. 5D, empty vector) and PFUS knockdown cells (Fig. 5E, N-PFUSll Biology 90 (2011) 844– 853
and C-PFUS constructs, respectively in Fig. 5F) illustrating localiza-tions corresponding to exo+, A+/P+ and exo− (null) phenotypes priorto stimulation of exocytosis. In exo+ and A+/P+ cells PFUS localizedto the cell cortex, presumably corresponding to scaffolds aroundindividual DCSVs, and also to the background cytoplasm. In theA+/P+ cells, the distribution of PFUS was more restricted and PFUSlocalization was mostly observed in the individual images of DCSVsat the anterior and posterior edges of the cells.
Progression of exocytosis inhibition with time is summarized inthe pie chart (Fig. 6). The A+/P+ and exo− phenotypes are combinedand scored as inhibition (red) in comparison to normal exocyto-sis (exo+, blue). While there is some variability overall, exocytosisbecomes inhibited, with all three RNAi constructs producing inhi-bition with the same time course as disappearance of PFUS (Fig. 6A).
Cells were collected after 72 h, starved for 6 h to digest theremaining bacteria containing the RNAi constructs and then fedwith control bacteria (HT115(DE3)) containing the empty vector. Asdemonstrated in the pie chart (Fig. 6B) after re-feeding and growthfor an additional 60 h, the exo+ phenotype is recovered in almostall cells.
In order to determine definitively whether these images repre-sented the concentration of PFUS in the scaffolds around DCSVs,double-labeling experiments were performed using PFUS antibod-ies and a content antibody (T8) to DCSVs, which localizes to thecortex of the semi-crystalline secretory content. Fig. 7A–E showscells from a representative PFUS knockdown population after 72 h.Some of the RNAi treated cells are full of DCSVs (Fig. 7C and D,arrows) and are considered (non-transformed) exo+ similar to con-trols (Fig. 7A). Other cells are A+/P+ (Fig. 7D) or null (exo−) (Fig. 7E).DSCVs are identified by double label as expected, with a rim ofPFUS label surrounding the T8 content label. In the A+/P+ cells,the numbers of DCSVs are severely reduced, leaving virtually onlysome DCSVs at the anterior and posterior tips of the cell and someunpacked content is visible stained red with T8 antibody (Fig. 7D,arrow). PFUS cytoplasmic labeling is reduced to varying extents asDCSVs disappear. In the null cells, no DSCVs are detected (Fig. 7E):these cells contain little detectable PFUS antibody localization andT8 antibody labeling is similarly reduced.
In order to quantify this effect, we counted the number ofdouble-labeled DCSVs in cells from 72 h PFUS RNAi treated prepara-tions vs. controls, using longitudinal optical sections at the midlinesshowing both apical and posterior ends of each cell. The numbersof cells counted in each preparation and the assigned phenotypesare shown in Fig. 8.
In controls, as in Fig. 7A, about 59 docked DCSVs are presentper image. No cells were counted that had less than 32 DCSVs. Weassign an exo+ phenotype to all cells possessing >30 DCSVs per sec-tion. In populations treated with PFUS RNAi, as shown in Fig. 7,some cells remain exo+, but most become exo−, A+/P+ cells withfewer than 30 docked DCSVs and null cells with fewer than 10DCSVs appear in a significant portion of the population. In addi-tion to matching the percentage failure in exocytosis in Fig. 6B, thisreduction in DSCVs correlates well with the reduction observed inPFUS (Fig. 4B). We conclude that interfering with PFUS transcrip-tion affects the synthesis and packaging pathway of DCSV content,which in turn is reflected by inhibition of exocytosis.
Discussion
Normal cell growth implies effective synthesis of cell mem-branes, cell morphogenesis and energy generation for the many
signaling pathways, as for example required for exocytosis. In ourhands, PFUS RNAi knockdown does not affect overall cell growth,suggesting that overall protein synthesis and cell organization arenot affected.
L. Liu et al. / European Journal of Cell Biology 90 (2011) 844– 853 849
Fig. 4. Western blot analysis and quantification of PFUS during RNAi treatment demonstrating knockdown of only PFUS. (A) Western blot of cultures exposed to differentPFUS constructs (H: feeding bacteria without RNAi vector added in, 28i, N, C and W as in (B) for 24, 48 and 72 h and probed with the pan-PGM antibody, n = 3 except for 72 hdata that were averaged from two experiments. Quantification (done as described in “Materials and methods” section) was standardized to amount of protein after HT115feeding (no vector incorporated). There appears to be no major changes in amount of protein detected with the pan-PGM antibody irrespective of the construct and time ofexposure to the individual constructs. (B) A different result is obtained when the samples are probed with the anti-PFUS antibody. In this case all three constructs produceknockdown of protein as also seen in the quantification. PFUS knockdown is already observed after 24 h exposure with all RNAi constructs.
Fig. 5. Exocytosis phenotypes after PFUS knockdown observed after 72 h of RNAi treatment. (A–C) Exocytosis phenotypes in the light microscopy. (A) Exo+ phenotype: cellsurrounded by secreted content (needle-like structures, arrows). (B) A+/P+: an intermediate exo− phenotype. Cell secretion only takes place from anterior and posterior polesof the cell (arrow). (C) Null: phenotype corresponds to no obvious secretion upon stimulation of exocytosis (no trichocyst content visible, only cilia). (D–F) Correspondingimages of phenotypes in confocal immunofluorescence microscopy showing PFUS localization after 72 h of RNAi treatment. (D) Control cells exo+ empty vector. (E) Exampleof N-terminal construct showing A+/P+ PFUS localization and C-terminal null (F). Note that content of DSCVs is unstained while a PFUS rim is present in the DSCV scaffold.All cells show some cytoplasmic label, but this is diminished in exo− cells.

850 L. Liu et al. / European Journal of Cell Biology 90 (2011) 844– 853
Fig. 6. Pie chart showing progression of inhibition of exocytosis during RNAi treatment. (A) Cells were stimulated and scored for secretory response at different time pointsduring RNAi treatment. Exo+ (full release) is shown in blue and exo− (null plus A+/P+) shown in red. Percentage of inhibition is shown next to corresponding areas. Theseresults show that on average after 72 h of RNAi treatment with any of the constructs about 60% of the cells are inhibited, n = 3. (B) Recovery after RNAi treatment. After 72 hR HT11e his fig
sctp(pstteSipP(otccP
NAi, cells were collected, washed and transferred to fresh medium supplied withxo+ phenotype recovery was 90%. (For interpretation of the references to color in t
Earlier experiments demonstrated that the biosynthesis of theecretory content (trichocyst matrix proteins, TMP) in Parameciumonsists of a series of many polypeptides synthesized and packagedogether in large immature vesicles that sequentially assemble viaroteolytic processing and scaffold formation into mature DCSVsVayssie et al., 2000). The general secretory pathway for TMProteins such as T8, follows the well established steps: RNA tran-cription – synthesis on bound ribosomes of the ER – translocationo the ER lumen and subsequently to the Golgi – vesicle forma-ion – content packaging and – transport – docking – and finallyxocytosis (membrane fusion and release of content) (Gerber andudhof, 2002; Sudhof and Rothman, 2009). Unlike T8, PFUS itselfs not a secretory protein and it is synthesized in the cytoplasm byolyribosomes and released there. Earlier data demonstrated thatFUS was found associated with fully formed cytoplasmic DCSVs15) and recent information has shown that PFUS is a componentf the scaffold of isolated DCSVs (Liu et al., 2009). This suggests
hat PFUS is added to the maturing secretory vesicle as a specificomponent of the membrane scaffold during the packaging pro-ess before transport and docking. Consistent with this explanation,FUS is found on DSCVs of tam8 mutant Paramecium, which cannot5(DE3) bacteria (without RNAi vector). After 72 h further growth in this medium,ure legend, the reader is referred to the web version of the article.)
transport the DCSVs to their cortical docking sites (Satir and Zhao,1999).
Earlier studies showed that PFUS in isolated DSCVs is glycosy-lated (Liu et al., 2009). More precisely, they suggest that PFUS whenproperly covalently modified by phosphoglucosylation is crucialfor scaffold integrity, and that dephosphoglucosylation accompa-nied by liberation of PFUS from the scaffold back to the cytoplasmis necessary for successful exocytosis to occur (Subramanian andSatir, 1992). This is demonstrated by the results from the exo−
temperature sensitive mutant, nd9. In these cells at the restrictivetemperature, DCSVs with PFUS containing scaffolds reach their cor-rect docking sites at the cell membrane (fusion rosettes) but uponstimulation, Ca2+-dependent dephosphoglucosylation of PFUS anddissociation of the vesicle scaffold does not take place and exocy-tosis cannot be induced (Subramanian and Satir, 1992; Satir andZhao, 1999).
During PFUS RNAi knockdown, with each generation the num-
ber of DCSVs diminishes. The feeding protocol produces many cellswhere knockdown is successful as measured by diminished DSCVsand a corresponding exo− phenotype after stimulation of exocy-tosis. However, there are always some cells in the population that
L. Liu et al. / European Journal of Cell Biology 90 (2011) 844– 853 851
Fig. 7. Localization of PFUS and vesicle content antibodies and decrease in numbers of DCSVs with RNAi treatment (72 h). After double labeling, localization of PFUS (green)is seen in scaffold around T8 content (red). (A) Illustrates control cells (empty vector 28i). Note the presence of numerous docked DCSVs aligned below the cell membrane,all showing PFUS surrounding each vesicle containing T8. Images illustrating results of RNAi treated cells: (B) N-terminal – note absence of unpackaged content areas (red)in the cytoplasm and C-terminal (C and D), respectively. There are two cells in C illustrating how some ciliates are not affected (arrow) whereas others are exhibiting a nullp conteo ls. (Fot
lcptTtI
henotype (C) or are partially affected as in D: note the presence of un-packaged T8pen reading frame. Overall RNAi treatment reduces the number of DCSVs in the celo the web version of the article.)
ook and respond as controls, possibly because they do not pro-ess the bacterial dsRNA properly. One explanation of the A+/P+
henotype is that RNAi knockdown of PFUS does not affect DCSVs
hat already are docked and have PFUS in their membrane scaffold.here are roughly 2000 DCSVs per cell of which our images of con-rols showed about 60 per optical section through the cell midline.f no new mature DCSVs are being formed in the knockdown cells,nt (arrow). (E) A null phenotype from treatment with the construct from the wholer interpretation of the references to color in this figure legend, the reader is referred
then in 4 generations, with average doubling time of 12 h, the num-ber of DCSVs will drop to 125 per cell or about 7 per cell in an opticalsection, evidently mainly in clusters at the cell poles.
When PFUS transcription is knocked down via RNAi, the concen-tration of this protein decreases, probably so that it can no longerassociate with the maturing DCSVs in sufficient concentration tostabilize the scaffold. Also when PFUS mRNA or protein concen-

852 L. Liu et al. / European Journal of Ce
Fig. 8. Number of docked DCSVs per medial longitudinal confocal sections in con-trol (28i) and PFUS RNAi 72 h treated cells. This histogram characterizes the numberof DCSVs associated with each assigned phenotype in various populations. A cellis defined as exo+ with >30 docked DCSVs/image, A+/P+ 10–30/image and null<10/image. For each phenotype the average number of DCSVs per cell for the pop-ulation counted is given. In control wt cell populations (empty vector) there are noce
tsttaiTiTaiitu
ctnrwChtcTecoe2
A
iFboaosU
ells with A+/P+ or null phenotypes. RNAi treated cell populations have cells withxo+, A+/P+ and null phenotypes.
ration decreases, this may signal feedback to the nucleus to stopynthesis of secretory contents and vesicle formation which even-ually leads to a shut down of the secretory pathway of DCSVs inhis cell. Alternatively, TMPs that are synthesized and not pack-ged properly are rapidly degraded. In any event, as shown bymmunofluorescent labeling of T8 in exo− cells, although someMPs are synthesized, new mature DCSVs are not produced, result-ng in an exo− phenotype after several days of PFUS knockdown.his dramatic change is not reflected in the total PGM, since only
small decrease in pan-PGM (≈12% average) is observed, whichs completely accounted for by the knockdown of PFUS, suggest-ng that other PGM superfamily members, some almost identicalo PFUS in amino acid composition and base composition (31) arenaffected by the PFUS RNAi knockdown procedure.
We cannot formally exclude a direct effect of the PFUS RNAionstructs on some residual DCSV component, although we thinkhis highly unlikely. More probably, as suggested above, PFUS isecessary at some point in DCSV scaffold assembly on which matu-ation of the DCSV depends. This interpretation would be consistentith the release of PFUS from the scaffold as a critical step in thea2+-dependent exocytosis of the DCSVs. In addition, PFUS clearlyas a different functional role from the original PGM, indicatinghat minor nucleotide changes and/or post-translational modifi-ations can evolutionarily add new dimensions to an old enzyme.his finding is probably not unprecedented, since isoforms of glyc-raldehyde phosphate dehydrogenase (GAPDH) and aldolase alsoan assume roles in signal transduction within the cell, in additionr in place of the function in glycolysis (Bosch et al., 2007; Csutorat al., 2005; Glaser et al., 2002; Kim et al., 2002; Takahashi et al.,004).
cknowledgements
We thank Drs. P. Satir and M. Rapport for suggestions and crit-cal reading of the manuscript. We thank Drs. R.D. Allen and A.K.ok for generously providing us with the vesicle content (T8) anti-ody. Microscopy was performed in the Analytical Imaging Facilityf the Albert Einstein College of Medicine. The data in this paperre in part from a thesis submitted by Li Liu in partial fulfillment
f the requirements for the Ph.D. in the Sue Golding Graduate Divi-ion of Medical Science, Albert Einstein College of Medicine, Yeshivaniversity, Bronx, NY, USA.ll Biology 90 (2011) 844– 853
References
Andersen, A.P., Wyroba, E., Reichman, M., Zhao, H., Satir, B.H., 1994. The activity ofparafusin is distinct from that of phosphoglucomutase in the unicellular eukary-ote Paramecium. Biochem. Biophys. Res. Commun. 200, 1353–1358.
Bastin, P., Galvani, A., Sperling, L., 2001. Genetic interference in protozoa. Res. Micro-biol. 152, 123–129.
Belkin, A.M., Burridge, K., 1995. Association of aciculin with dystrophin and utrophin.J. Biol. Chem. 270, 6328–6337.
Bosch, J., Buscaglia, C.A., Krumm, B., Ingason, B.P., Lucas, R., Roach, C., Cardozo, T.,Nussenzweig, V., Hol, W.G., 2007. Aldolase provides an unusual binding site forthrombospondin-related anonymous protein in the invasion machinery of themalaria parasite. Proc. Natl. Acad. Sci. U. S. A. 104, 7015–7020.
Csutora, P., Strassz, A., Boldizsar, F., Nemeth, P., Sipos, K., Aiello, D.P., Bedwell, D.M.,Miseta, A., 2005. Inhibition of phosphoglucomutase activity by lithium alterscellular calcium homeostasis and signaling in Saccharomyces cerevisiae. Am. J.Physiol. Cell Physiol. 289, C58–C67.
Daubenberger, C.A., Tisdale, E.J., Curcic, M., Diaz, D., Silvie, O., Mazier, D., Eling,W., Bohrmann, B., Matile, H., Pluschke, G., 2003. The N′-terminal domain ofglyceraldehyde-3-phosphate dehydrogenase of the apicomplexan Plasmodiumfalciparum mediates GTPase Rab2-dependent recruitment to membranes. Biol.Chem. 384, 1227–1237.
Elbashir, S.M., Lendeckel, W., Tuschl, T., 2001. RNA interference is mediated by 21-and 22-nucleotide RNAs. Genes Dev. 15, 188–200.
Froissard, M., Keller, A.M., Cohen, J., 2001. ND9P, a novel protein with armadillo-like repeats involved in exocytosis: physiological studies using allelic mutantsin Paramecium. Genetics 157, 611–620.
Froissard, M., Kissmehl, R., Dedieu, J.C., Gulik-Krzywicki, T., Plattner, H., Cohen, J.,2002. N-ethylmaleimide-sensitive factor is required to organize functional exo-cytotic microdomains in Paramecium. Genetics 161, 643–650.
Fu, L., Miseta, A., Hunton, D., Marchase, R.B., Bedwell, D.M., 2000. Loss of the majorisoform of phosphoglucomutase results in altered calcium homeostasis in Sac-charomyces cerevisiae. J. Biol. Chem. 275, 5431–5440.
Galvani, A., Sperling, L., 2002. RNA interference by feeding in Paramecium. TrendsGenet. 18, 11–12.
Gerber, S.H., Sudhof, T.C., 2002. Molecular determinants of regulated exocytosis.Diabetes 51 (Suppl. 1), 3–11.
Gilligan, D.M., Satir, B.H., 1982. Protein phosphorylation/dephosphorylation andstimulus-secretion coupling in wild type and mutant Paramecium. J. Biol. Chem.257, 13903–13906.
Glaser, P.E., Han, X., Gross, R.W., 2002. Tubulin is the endogenous inhibitor of theglyceraldehyde 3-phosphate dehydrogenase isoform that catalyzes membranefusion: implications for the coordinated regulation of glycolysis and membranefusion. Proc. Natl. Acad. Sci. U. S. A. 99, 14104–14109.
Harumoto, T., 1994. The role of trichocyst discharge and backward swimming inescaping behavior of Paramecium from Dileptus margaritifer. J. Eukaryotic Micro-biol. 41, 560–564.
Hauser, K., Kissmehl, R., Linder, J., Schultz, J.E., Lottspeich, F., Plattner, H., 1997. Iden-tification of isoforms of the exocytosis-sensitive phosphoprotein PP63/parafusinin Paramecium tetraurelia and demonstration of phosphoglucomutase activity.Biochem. J. 323, 289–296.
Hennessey, T.M., Kim, M.Y., Satir, B.H., 1995. Lysozyme acts as a chemorepellentand secretagogue in Paramecium by activating a novel receptor-operated Ca++
conductance. J. Membr. Biol. 148, 13–25.Hessler, R.J., Blackwood, R.A., Brock, T.G., Francis, J.W., Harsh, D.M., Smolen, J.E.,
1998. Identification of glyceraldehyde-3-phosphate dehydrogenase as a Ca2+-dependent fusogen in human neutrophil cytosol. J. Leukoc. Biol. 63, 331–336.
Kim, J.H., Lee, S., Lee, T.G., Hirata, M., Suh, P.G., Ryu, S.H., 2002. Phospholipase D2directly interacts with aldolase via its PH domain. Biochemistry 41, 3414–3421.
Lefort-Tran, M., Aufderheide, K., Pouphile, M., Rossignol, M., Beisson, J., 1981. Con-trol of exocytotic processes: cytological and physiological studies of trichocystmutants in Paramecium tetraurelia. J. Cell Biol. 88, 301–311.
Levin, S., Almo, S.C., Satir, B.H., 1999. Functional diversity of the phosphoglucomu-tase superfamily: structural implications. Protein Eng. 12, 737–746.
Liu, L., Tucker, S.C., Satir, B.H., 2009. Toxoplasma PRP1 is an ortholog of parafusin(PFUS) in vesicle scaffold assembly in Ca(2+)-regulated exocytosis. Eur. J. CellBiol. 88, 301–313.
Matthiesen, S.H., Shenoy, S.M., Kim, K., Singer, R.H., Satir, B.H., 2001. A parafusin-related Toxoplasma protein in Ca2+-regulated secretory organelles. Eur. J. CellBiol. 80, 775–783.
Matthiesen, S.H., Shenoy, S.M., Kim, K., Singer, R.H., Satir, B.H., 2003. Role of the para-fusin orthologue PRP1, in microneme exocytosis and cell invasion in Toxoplasmagondii. Cell Microbiol. 5, 613–624.
Meyer, E., Cohen, J., 1999. Paramecium molecular genetics: functional complemen-tation and homology-dependent gene inactivation. Protist 150, 11–16.
Murtaugh, T.J., Gilligan, D.M., Satir, B.H., 1987. Purification of and production of anantibody against a 63,000 Mr stimulus-sensitive phosphoprotein in Paramecium.J. Biol. Chem. 262, 15734–15739.
Narayanan, N., Xu, A., 1997. Phosphorylation and regulation of the Ca(2+)-pumpingATPase in cardiac sarcoplasmic reticulum by calcium/calmodulin-dependentprotein kinase. Basic Res. Cardiol. 92 (Suppl. 1), 25–35.
Rogalski-Wilk, A.A., Cohen, R.S., 1997. Glyceraldehyde-3-phosphate dehydrogenaseactivity and F-actin associations in synaptosomes and postsynaptic densities ofporcine cerebral cortex. Cell. Mol. Neurobiol. 17, 51–70.
Satir, B.H., Bleyman, L.K., 1993. Use of Tetrahymena and Paramecium in studies ofexocytosis. Methods Enzymol. 221, 174–190.

l of Ce
S
S
S
S
S
S
S
L. Liu et al. / European Journa
atir, B.H., Zhao, H., 1999. Parafusin and calcium-induced signal transduction inexocytosis. Protoplasma 206, 228–233.
atir, B.H., Hamasaki, T., Reichman, M., Murtaugh, T.J., 1989. Species distribution ofa phosphoprotein (parafusin) involved in exocytosis. Proc. Natl. Acad. Sci. U. S.A. 86, 930–932.
onneborn, T.M., 1970. Methods in Paramecium research. Methods Cell Physiol. 4,242–339.
onneborn, T.M., 1974. Paramecium aurelia. In: King, R.C. (Ed.), Handbook of Genetics.Plenum Press, New York, pp. 496–594.
ubramanian, S.V., Satir, B.H., 1992. Carbohydrate cycling in signal transduction:parafusin, a phosphoglycoprotein and possible Ca(2+)-dependent transducermolecule in exocytosis in Paramecium. Proc. Natl. Acad. Sci. U. S. A. 89,11297–11301.
ubramanian, S.V., Wyroba, E., Andersen, A.P., Satir, B.H., 1994. Cloning
and sequencing of parafusin, a calcium-dependent exocytosis-related phosphoglycoprotein. Proc. Natl. Acad. Sci. U. S. A. 91,9832–9836.udhof, T.C., Rothman, J.E., 2009. Membrane fusion: grappling with SNARE and SMproteins. Science 323, 474–477.
ll Biology 90 (2011) 844– 853 853
Takahashi, K., Inuzuka, M., Ingi, T., 2004. Cellular signaling mediated by calphoglin-induced activation of IPP and PGM. Biochem. Biophys. Res. Commun. 325,203–214.
Timmons, L., Court, D.L., Fire, A., 2001. Ingestion of bacterially expressed dsRNAs canproduce specific and potent genetic interference in Caenorhabditis elegans. Gene263, 103–112.
Vayssie, L., Skouri, F., Sperling, L., Cohen, J., 2000. Molecular genetics of regulatedsecretion in Paramecium. Biochimie 82, 269–288.
Wassmer, T., Froissard, M., Plattner, H., Kissmehl, R., Cohen, J., 2005. The vacuolarproton-ATPase plays a major role in several membrane-bounded organelles inParamecium. J. Cell Sci. 118, 2813–2825.
Wyroba, E., Satir, B.H., 2000. A comparative hybridization analysis of yeast DNAwith Paramecium parafusin- and different phosphoglucomutase-specific probes.Biochem. Cell Biol. 78, 683–690.
Wyroba, E., Widding Hoyer, A., Storgaard, P., Satir, B.H., 1995. Mammalian homo-logue of the calcium-sensitive phosphoglycoprotein, parafusin. Eur. J. Cell Biol.68, 419–426.
Zhao, H., Satir, B.H., 1998. Parafusin is a membrane and vesicle associated proteinthat cycles at exocytosis. Eur. J. Cell Biol. 75, 46–53.





























