Embed Size (px)
Citation preview

FISHERIES MANAGEMENT
Managing Macrophytes to Improve FishGrowth: A Multi-lake ExperimentBy Mark H. Olson, Stephen R. Carpenter, Paul Cunningham, Sarig Gafny, Brian R. Herwig,Nathan P. Nibbelink, Tom Pellett, Christine Storlie, Anett S. Trebitz, and Karen A. Wilson
ABSTRACTMacrophyte harvesting often has been suggested as a way to improve fish growth and size struc-ture in lakes with high densities of submergent macrophytes and stunted fish populations. How-ever, previous experimental tests have provided no clear consensus on whether the techniqueworks for management. We conducted a series of whole-lake manipulations to test the effects ofmacrophyte removal on growth of bluegill and largemouth bass. We selected four lakes in south-ern and central Wisconsin for experimental manipulation and nine others for controls. In August1994, we removed macrophytes from approximately 20% of the littoral zone by cutting a series ofevenly spaced, deep channels throughout each treatment lake. In the first year after manipulation,we observed substantially increased growth rates of some age classes of both bluegill and large-mouth bass in treatment lakes relative to controls. Growth rates of other age classes were lessresponsive to manipulation. We observed increased bluegill and largemouth bass growth despiterapid regrowth of macrophytes in our treatment lakes. By May 1996, fewer than 25% of the chan-nels remained. Our results suggest that harvesting macrophytes in a series of deep channels maybe a valuable tool for integrated management of fish and macrophytes.
any lakes and reservoirsacross North America suf-fer from high densities ofsubmergent macrophytes.
The thick beds of vegetation thatcover the littoral zone are perennialproblems in some lakes and recentdevelopments in others (Colle andShireman 1980; Cooke et al. 1993).Excessive macrophyte growth oftencan be traced to the invasion of exoticmacrophytes such as Eurasian milfoil(Myriophyllum spicatum) (Aiken et al.1979) and hydrilla (Hydrilla verticil-lata) (Haller 1979). As these invadersspread, they displace the diversecommunity of native species andcreate a near-monoculture of densemacrovhvtes (Haller and Sutton
* 1975; Nichols 1994). Consequently,
the aesthetic quality and recreationalvalue of a lake can decline severely.
High densities of macrophytesalso can harm the quality of a fish-ery (Wiley et al. 1984; Bettoli et al.1992). Dense macrophytes cancause panfish and game fish to be-come stunted via two pathways.First, feeding rates generally arereduced in lakes with dense macro-phytes (Crowder and Cooper 1982).Although prey abundance (eitherinsect or fish) may increase asmacrophyte density increases,macrophytes also reduce predatorforaging efficiency by providing arefuge for prey. Therefore, feedingrate (determined by the combina-tion of prey abundance and forag-ing efficiency) is maximized at
intermediate macrophyte densitiesand reduced as densities increasebeyond that point (Heck and Crow-der 1991; Savino et al. 1992). Sec-ond, dense vegetation generallyreduces foraging efficiency of pisci-vores and lowers predator-inducedmortality rates of small fishes(Savino and Stein 1982; Gotceitasand Colgan 1989). This reduction inmortality leads to a greater popula-tion density and stronger competi-tive interactions among small fishes(Mittelbach 1988).
Macrophyte Harvesting asa Management Strategy
The potential for improvinggrowth and size-structure of fishesby reducing macrophyte densities
Mark H. Olson is a senior research associate at the Cornell Biological Field Station, 900 Shackelton Point Road, Bridge-port, NY 13030; [email protected]; FAX 315/633-2358. Stephen R. Carpenter is a Halverson professor of limnology at theCenter for Limnology, University of Wisconsin. Paul Cunningham is a fisheries ecologist at the Bureau of Fish Manage-ment, Wisconsin Department of Natural Resources. Sarig Gafny is a research scientist at the Institute for Nature Conserva-tion Research, Tel Aviv University, Israel. Brian R. Herwig is a research technician at the Center for Limnology, Universityof Wisconsin. Nathan P. Nibbelink is a research assistant at the Department of Zoology and Physiology, University ofWyoming. Tom Pellett is an aquatic ecosystems ecologist for the Bureau of Integrated Science Services, Wisconsin Depart-ment of Natural Resources. Christine Storlie is a fisheries ecologist at the Bureau of Integrated Science Services, WisconsinDepartment of Natural Resources. Anett S. Trebitz is a research ecologist at the Environmental Protection Agency in Min-nesota. Karen A. Wilson is a research assistant at the Center for Limnology, University of Wisconsin.
Vol. 23, No. 26 Fisheries

FISHERIES MANAGEMENT
has long been recognized. As earlyas 1941, Swingle and Smith recom-mended macrophyte control as astrategy "for correcting conditionsthat produce stunted populations inimpounded waters" (Swingle andSmith 1941:104). Since that time, theidea has been proposed and testedmany times (reviewed in Engel1995). However, in spite of this re-search the current status of macro-phyte removal as a fishery manage-ment tool is unclear. One reason forthe uncertainty is the lack of con-sensus from previous experiments.Even studies conducted with thesame fish and macrophyte specieshave yielded variable results (e.g.,Shireman et al. 1985; Cross et al.1992; Bettoli et al. 1993), in part be-cause experiments were conductedat different scales (e.g., aquaria, cages,experimental ponds) to examinedifferent mechanisms of fish-macro-phyte interactions. Furthermore,few studies have provided any prac-tical advice on macrophyte removal.Thus, the logistic challenges oflarge-scale macrophyte manipula-tions have not been assessed.
Resolving the status of macro-phyte removal as a managementtool requires explicit considerationof the scale of experimentation(Levin 1992). To test whether macro-phytes can be managed to improvefish growth, the experiment mustbe conducted on whole lakes. Atthis scale, all direct and indirectpathways between fish and macro-phytes will be affected, and theoverall effect of a manipulation canbe investigated (Frost et al. 1988).Whole lakes also are the typicalunit of management (Kitchell 1992).Therefore, whole-lake experimentscan be used as field tests of poten-tial management techniques.
Another important considerationin experimental design is the levelof replication (McAllister and Peter-man 1992). Replication provides thestatistical power necessary to detectan effect, if the effect exists (Cohen1988). Because the interaction be-tween fish and macrophytes in-volves many potentially conflictingmechanisms, the possibility of a
February 1998
negative result (i.e., no effect ofmacrophyte manipulations) isstrong. Replication is necessary toallow a conclusion of no effect to beinterpreted as a lack of biologicalchange and not a lack of statisticalpower (Peterman 1990).
A Multi-lake ExperimentIn 1994, we initiated a replicated
whole-lake experiment to test thehypothesis that submergent macro-phytes can be managed to improvefish growth. Our experiment wasconducted in southern and centralWisconsin, where Eurasian milfoilhas become a nuisance and reducedthe recreational value of many lakes(Nichols 1994). High macrophytedensities also have harmed growthrates of important fishes for therecreational fishery such as bluegill(Lepomis macrochirus) and large-mouth bass (Micropterus salmoides),causing concern among anglers(Engel 1987).
The early stages of our experi-ment involved several planningexercises (Carpenter et al., in press).First, we chose a design that assignedexperimental lakes into two groups.We designated one group of lakesto receive a uniform manipulationof littoral zone macrophytes and asecond group of lakes to serve as
unmanipulated controls. In subse-quent analyses, we compared dif-ferences between pre- and postma-nipulation growth rates of bluegilland largemouth bass in manipulatedlakes to control lakes. We focusedour analyses on growth responsesbecause changes in fish density canbe difficult to detect (Carpenter etal. 1995). Next, we developed aseries of simulation models to pre-dict fish growth responses to arange of manipulation intensities(Trebitz and Nibbelink 1996; Trebitzet al. 1997). These models indicatedthat clearing approximately 20% ofthe littoral zone of macrophyteswould provide meaningful im-provements in fish growth and sizestructure. Finally, we conducted apower analysis to determine thenumber of experimental lakes need-ed to detect these predictedresponses (Carpenter et al. 1995). Todetect changes in growth, the poweranalysis suggested that we wouldrequire at least three lakes per treat-ment (manipulated or control).
Lakes were selected for theexperiment based on several criteria(Carpenter et al., in press). Usinginsights of state biologists and man-agers, we chose a set of 13 lakes(Table 1) with similar size structuresof bluegill and largemouth bass
Table 1 describes our study lakes in southern and central Wisconsin. Littoral zone areawas calculated as the area between shore and the lower depth limit of macrophytegrowth. The value for White Mound Lake was not available (NA). Sample sizes reportedfor bluegill (BG) and largemouth bass (LMB) are for all age classes combined.
% ofareain Sample Size
Treat- Area littoral Max. BG LMBLake County ment (ha) zone depth 1993 1995 1993 1995
FishHeidmannSilverTumaEast AlaskaHarptHorseshoeKuselMautheNapowanSheaWingraWhite Mound
DaneKewauneeWatipacaManitowocKewauneeManitowocManitowocWausharaFond du LacWausharaKewauneeDaneSauk
CutCutCutCutControlControlControlControlControlControlControlControlControl
102.010.528.37.7
21.512.68.9
32.031.620.713.0
140.042.1
54.567.277.776.949.040.014.076.042.087.072.089.0
NA
19.09.25.29.2
15.37.3
16.58.97.15.57.34.38.5
112 103 17269 112 6039 118 955 125 7458 87 2256 115 6643 75 1138 106 9437 78 542 89 3673 110 2534 124 4768 114 109
Fisheries * 7
241316182825213625546
4550
FISHERIES MANAGEMENT
(Olson 1996; Nibbelink and Carpenter,in press), macrophyte-dominated lit-toral zones (i.e., >90% coverage) withhigh densities of Eurasian milfoil, nohistory of winterkill (to avoid con-founding effects; Tonn and Paszkowski1986), and no other significant man-agement actions (e.g., chemical reha-bilitation, predator management, etc.).
We manipulated macrophytes infour study lakes (cut lakes) inAugust-September 1994 (Table 1). Theother nine lakes served as unmanipu-lated controls (Table 1; control lakesoutnumbered cut lakes because theycould be monitored with little effort).In each cut lake, we removed macro-phytes in a series of evenly spacedchannels that extended from shore tothe edge of the littoral zone. In addi-tion to being the most practicalmethod for macrophyte removal(Engel 1995), channels also increasethe amount of vegetation-open wateredge, which has been hypothesized tobe an important zone of interaction inaquatic systems (Smith 1993; Jeppesenet al., in press). Channels were createdwith a mechanical harvester equippedwith a 3-m-wide conventional cuttingbar and a second custom-designedcutting bar that could cut a 2-m swathto a depth of 5 m. Macrophytes werecut just above the root crown (to slowregrowth), collected, and transportedaway from the lake to nearby agricul-tural fields. Due to variation in lakesize and morphometry, the length,number, and width of channels, andthe amount of effort varied amonglakes (Table 2). However, the percent-age of littoral zone cleared of macro-phytes from each lake was close toour goal of 20%.
To investigate the effect of macro-phyte harvesting on bluegill and
1
EE
o0o
-C
0a
0()
-5
a) Blueill40
EE- 30o
0
, 20O
a, 10
o0
C
(.9
-10
b) Largemouth bass
F TTZ0 2 3 4 5 6 2 3 4 5 6
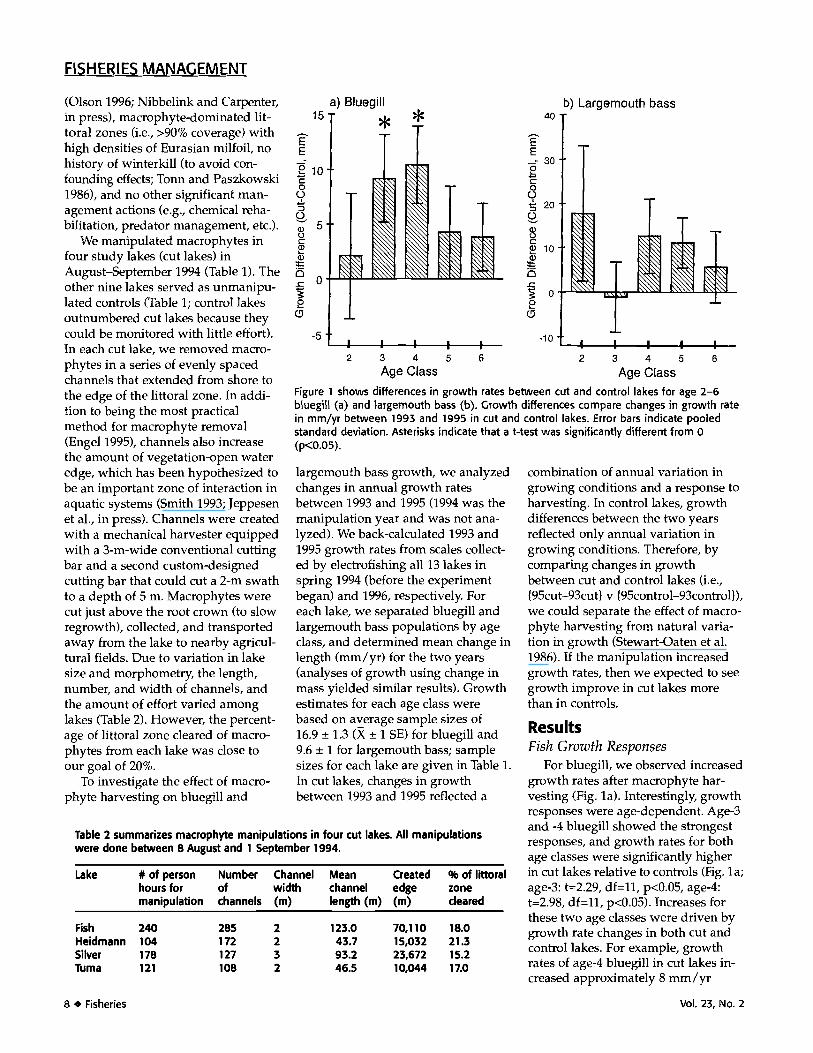
Age Class Age ClassFigure 1 shows differences in growth rates between cut and control lakes for age 2-6bluegill (a) and largemouth bass (b). Growth differences compare changes in growth ratein mm/yr between 1993 and 1995 in cut and control lakes. Error bars indicate pooledstandard deviation. Asterisks indicate that a t-test was significantly different from 0(p<0.05).
largemouth bass growth, we analyzedchanges in annual growth ratesbetween 1993 and 1995 (1994 was themanipulation year and was not ana-lyzed). We back-calculated 1993 and1995 growth rates from scales collect-ed by electrofishing all 13 lakes inspring 1994 (before the experimentbegan) and 1996, respectively. Foreach lake, we separated bluegill andlargemouth bass populations by ageclass, and determined mean change inlength (mm/yr) for the two years(analyses of growth using change inmass yielded similar results). Growthestimates for each age class werebased on average sample sizes of16.9 + 1.3 (X 1 SE) for bluegill and9.6 1 for largemouth bass; samplesizes for each lake are given in Table 1.In cut lakes, changes in growthbetween 1993 and 1995 reflected a
Table 2 summarizes macrophyte manipulations in four cut lakes. All manipulationswere done between 8 August and 1 September 1994.
Lake # of person Number Channel Mean Created 9b of littoralhours for of width channel edge zonemanipulation channels (m) length (m) (m) cleared
FishHeidmannSilverTuma
240104178121
285172127108
2232
123.043.793.246.5
70,11015,03223,67210,044
18.021.315.217.0
8 * Fisheries
combination of annual variation ingrowing conditions and a response toharvesting. In control lakes, growthdifferences between the two yearsreflected only annual variation ingrowing conditions. Therefore, bycomparing changes in growthbetween cut and control lakes (i.e.,(95cut-93cut) v 95control-93control}),we could separate the effect of macro-phyte harvesting from natural varia-tion in growth (Stewart-Oaten et al.1986). If the manipulation increasedgrowth rates, then we expected to seegrowth improve in cut lakes morethan in controls.
ResultsFish Growth Responses
For bluegill, we observed increasedgrowth rates after macrophyte har-vesting (Fig. la). Interestingly, growthresponses were age-dependent. Age-3and -4 bluegill showed the strongestresponses, and growth rates for bothage classes were significantly higherin cut lakes relative to controls (Fig. la;age-3: t=2.29, df=ll, p<0.05, age-4:t=2.98, df=11, p<0.05). Increases forthese two age classes were driven bygrowth rate changes in both cut andcontrol lakes. For example, growthrates of age-4 bluegill in cut lakes in-creased approximately 8 mm/yr
Vol. 23, No. 2
c
F 11,
FISHERIES MANAGEMENT
(from 22.4 mm/yr to 30.9 mm/yr),whereas growth in control lakes de-creased 2 mm/yr (from 28.6 mm/yrto 26.8 mm/yr). Consequently, therewas a 10 mm/yr increase in growthof age-4 bluegill in cut lakes relativeto controls. Other age classes also hadimproved growth rates in cut lakes,but these increases were not statisti-cally significant.
Unlike bluegill, growth rates oflargemouth bass did not respond con-sistently to macrophyte harvesting(Fig. lb). Although growth rates ofmost age classes tended to increase incut lakes (age-3 was the lone excep-tion), these changes were not statisti-cally significant (t-tests; p>0.10 for alllargemouth bass age classes).
To better understand and interpretthe effects of macrophyte harvesting,we also analyzed changes in bluegilland largemouth bass growth from aBayesian perspective (Ellison 1996).Conventional statistical analyses (e.g.,t-tests) give the probability of reject-ing the null hypothesis that themanipulation has no effect when thenull is true (i.e., Type I error), provid-ing a simple yes-no answer (Bergerand Berry 1988). In contrast, a Bayesian
a) Bluegill0.140.120.10
i.co
_d
0.080.060.040.020.00 F.
-30 -20Growth
5-
2 /,- ,,
6II
.''.
o3'n
0F,
a
M
zC,C,
0
C,
Dense macrophyte beds in the littoral zone of White Mound Lake, Wisconsin, harm thefishery and reduce the lake's recreational value.
analysis estimates probabilities for allpossible outcomes (Gelman et al.1995). Using a noninformative priordistribution (i.e., all outcomes haveequal probability in the prior proba-bility distribution), this analysis calcu-lates posterior probability distribu-tions from the difference in mean
growthresponsesamong cutand controllakes calculat-
r4 ed between
3I-'
-10 0 10 20Difference (Cut - Control,
30mm)
b) Largemouth bass
n0C
on
5
1993 and 1995and thepooled stan-dard devia-tion. Thesedistributionscould then beinterpreteddirectly as theprobability ofa specifiedeffect (Gel-man et al.1995). Because
the distribution describes all out-comes, the area under the curve sumsto one. Therefore, combined probabil-ities for a range of outcomes could bedetermined. For example, the proba-bility of growth increasing is equal tothe area under the curve to the rightof zero. Similarly, the probability of adesired (e.g., a 10-mm increase ingrowth) or undesired result (e.g., adecrease in growth) may be calculat-ed directly.
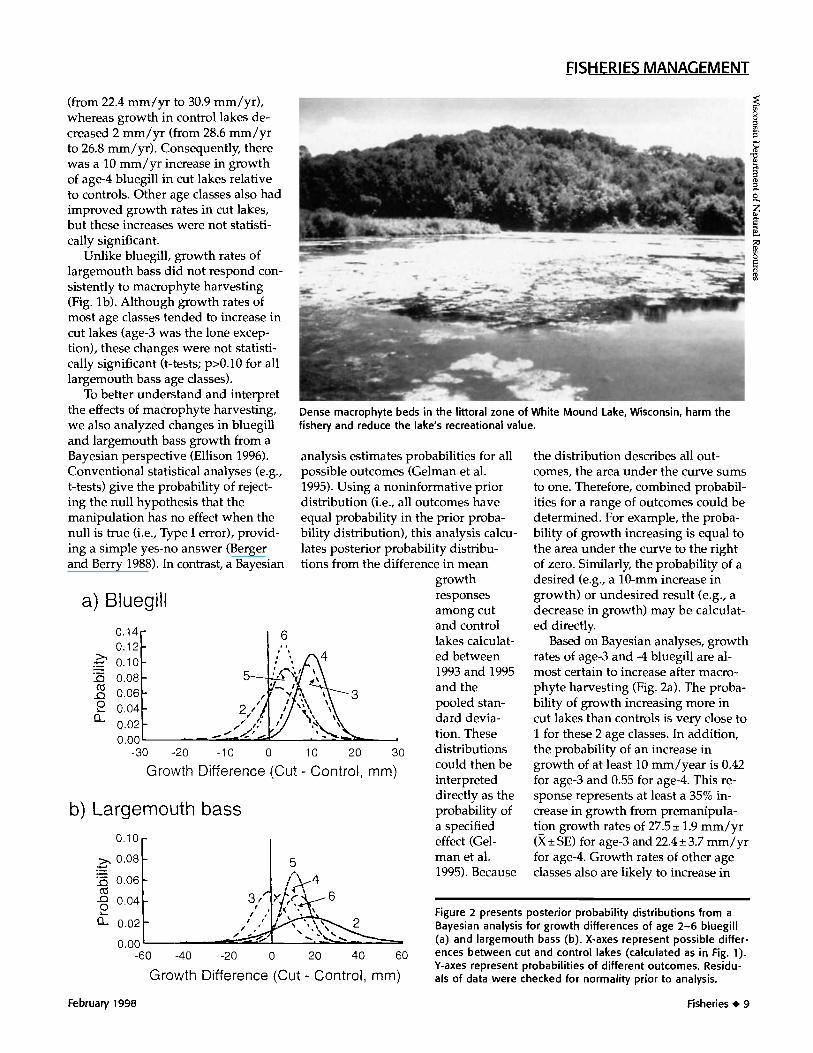
Based on Bayesian analyses, growthrates of age-3 and -4 bluegill are al-most certain to increase after macro-phyte harvesting (Fig. 2a). The proba-bility of growth increasing more incut lakes than controls is very close to1 for these 2 age classes. In addition,the probability of an increase ingrowth of at least 10 mm/year is 0.42for age-3 and 0.55 for age-4. This re-sponse represents at least a 35% in-crease in growth from premanipula-tion growth rates of 27.5 + 1.9 mm/yr(X+ SE) for age-3 and 22.4 + 3.7 mm/yrfor age-4. Growth rates of other ageclasses also are likely to increase in
-6Figure 2 presents posterior probability distributions from aBayesian analysis for growth differences of age 2-6 bluegill(a) and largemouth bass (b). X-axes represent possible differ-
0 ences between cut and control lakes (calculated as in Fig. 1).Y-axes represent probabilities of different outcomes. Residu-als of data were checked for normality prior to analysis.
February 1998
-60 -40 -20 0 20 40 61Growth Difference (Cut - Control, mm)
I .. _ i
/ 11k, 11 1
Fisheries * 9

FISHERIES MANAGEMENT
response to macrophyte harvesting(probability that growth increasesmore in cut lakes than controls is >0.6for ages 2, 5, and 6). However, increasesfor these age classes are likely to bebetween 0 and 10 mm/yr (Fig. 2a).
For largemouth bass, Bayesiananalyses indicate a high probability ofgrowth rates increasing for age-4 andage-5 (Fig. 2b; probability of growthincreasing more in cut lakes than con-trols is >0.9 for both age classes). In-creases of 10 mm/yr from premanipu-lation growth rates of 41.7 + 8.3 mm/yrfor age-4 and 38.4 + 3.0 mm/yr forage-5 are likely for both age classes(probability is >0.5 for both age class-es). Ages 2 and 6 also are likely togrow better after harvesting, andincreases of more than 20 mm/yr arepossible for age-2 (probability =0.44).In contrast, growth rates of age-3largemouth bass could increase ordecrease with equal probability afterharvesting. This result is expectedwhen the manipulation has no effect,and growth rates vary among years.
The variation in growth responsesamong age classes of both bluegilland largemouth bass is not surprisingconsidering diets of both species arestrongly size-dependent. Bluegill typi-cally undergo a diet shift as a result ofchanges in predator-mediated habitatuse (Mittelbach 1981). Up to 80 mmlong, bluegill feed primarily on vege-tation-dwelling invertebrates, where-as bluegill above 125 mm feed on
zooplankton (Mittelbach 1981; Wernerand Hall 1988). Between these twosizes, bluegill consume a combinationof both prey types (Werner and Hall1988) and, therefore, would be expect-ed to benefit from an increase in theamount of edge between vegetationand open water. This size range corre-sponds closely to the two age classesthat responded most strongly to ourmanipulation (age-3: X=92.8 ± 5.6 mmat the start of 1995; age-4: X=111.7+6.7 mm). Largemouth bass diets alsochange with size. In our study lakes,largemouth bass switch from a diet ofinvertebrates to primarily bluegill at alength of 100 mm (Olson 1996). Be-cause largemouth bass are gape-limit-ed predators, they can consume onlybluegill that are <40% their ownlength (Lawrence 1958; Timmons etal. 1980). The two age classes of large-mouth bass that responded moststrongly to macrophyte harvesting,age-4 and age-5, are capable of con-suming bluegill up to 106 mm and17 mm, respectively. Growth respons-es of bluegill suggest that fish of thesesizes use channels. Therefore, ourexperiment suggests that harvestingmacrophytes may lead to increasedprey availability and growth of age-4and -5 largemouth bass.
Macrophyte RegrowthMacrophytes recovered quickly
from harvesting. From aerial countsof channels, we estimated that an
u0
oz
This aerial photo shows the spatial arrangement of channels after macrophyte harvesting.
10 * Fisheries
400 1994
= 300 1996
O-Z 200
E= 100
0
·o` ·"" , 0 k e
Figure 3 shows the number of channels incut lakes. Channels were counted from aerialphotos of each lake in 1994 (after harvest-ing) and 1996. No channels were visible inHeidmanns Lake in 1996.
average of 23% of the original chan-nels remained in May 1996, 21 monthsafter our manipulation occurred (Fig. 3).Remaining channels also were muchshorter in 1996, with the most signifi-cant regrowth occurring in shallowwater (0-2 m). Channels persisted moststrongly in Fish Lake, where Eurasianmilfoil grows to depths of 2 m-5 m.
ConclusionsBecause macrophytes quickly
returned to premanipulation densities,our experiment was a short-term per-turbation. Our August 1994 manipu-lation of macrophytes lasted onlythrough the 1995 growing season, andby summer 1996 the manipulationwas essentially over in all but onelake. Nevertheless, we observedstrong positive responses in growthrates of some age classes of both blue-gill and largemouth bass. Althoughthese increases were temporary, theireffects will persist as an increase inbluegill and largemouth bass sizestructure for the lifetime of the affect-ed age classes. We did not measurethe longer-term effects of repeatedmacrophyte harvesting.
Results of this study suggest thatmacrophyte harvesting can be a use-ful technique for improving growthrates and size structures of panfishand game fish. Substantial increasesin fish growth are possible with a
Vol. 23, No. 2

FISHERIES MANAGEMENT
manageable level of effort. In ourexperiment, harvesting macrophytesfrom 20% of the littoral zone was suf-ficient to temporarily increase growthrates of bluegill and largemouth bass.Models suggest that similar resultsshould occur with macrophyte har-vests of up to 40% of the littoral zone(Trebitz and Nibbelink 1996; Trebitzet al. 1997). In many lakes, mechani-cal harvesters are already used tocontrol nuisance macrophytes. Ourstudy suggests that changing thestrategy of harvesting from clear-cut-ting the top meter of vegetation toselectively cutting deep channelsthroughout the lake may simultane-ously improve the fishery and recre-ational value of a lake. )_
AcknowledgmentsThis project was a collaborative
effort between the Wisconsin Depart-ment of Natural Resources and the Cen-ter for Limnology, University of Wis-consin. It was funded by the FederalAid to Sport Fish Restoration Actthrough the Wisconsin Department ofNatural Resources, Project F-95-P. Wethank the Dane County Departmentof Public Works for building the macro-phyte harvester and conducting theharvests. Dave Dreikosen, JonathonHricik, Sarah Huhta, CatherineMcClellan, Steve Mrazik, Terry Schenk,Dave Sloey, Kyle Tanger, Jean Unmuth,Marty Wessels, and Beth Zaniewskiprovided valuable assistance in thefield and/or lab. Kathy Cottingham,Janet Fischer, Lars Rudstam, DanielSchindler, and Craig Stow providedhelpful reviews of the manuscript.
ReferencesAiken, S. G., P. R. Newroth, and 1. Wile.
1979. The biology of Canadian weeds.Myriophyllum spicatum. L. Can. J.Plant Sci. 59:201-215.
Berger, J. O., and D. A. Berry. 1988. Statis-tical analysis and the illusion of objec-tivity. Am. Sci. 76:159-165.
Bettoli, P. W., M. J. Maceina, R. L. Noble,and R. K. Betsill. 1992. Piscivory inlargemouth bass as a function of aquat-ic vegetation abundance. N. Am. J.Fish. Manage. 12:509-516.
Bettoli, P. W., M. J. Maceina, R. L. Noble,and R. K. Betsill. 1993. Response of areservoir fish community to aquatic
February 1998
vegetation removal. N. Am. J. Fish.Manage. 13:110-124.
Carpenter, S. R., and eight coauthors.1995. Responses of bluegill to habitatmanipulations: power to detect effects.N. Am. J. Fish. Manage. 15:519-527.
. In press. Macrophyte structureand growth of bluegill (Lepomis macro-chirus): design of a multi-lake experi-ment. In E. Jeppesen, K. Christoffersen,and M. Sondergaard, eds. The role ofsubmerged macrophytes in structuringlakes. Springer-Verlag, Berlin.
Cohen, J. 1988. Statistical power analysisfor the behavioral sciences, 2nd ed.L. Erlbaum Associates, Hillsdale, NJ.
Colle, D. E., and I. V. Shireman. 1980.Coefficients of condition for large-mouth bass, bluegill, and redear sun-fish in Hydrilla-infested lakes. Trans.Am. Fish. Soc. 109:521-531.
Cooke, G. D, E. B. Welch, S. A. Peterson,and P. R. Newroth. 1993. Lake andreservoir restoration. Lewis Publishers,Boca Raton, FL.
Cross, T. K., M. C. Mclnerny, and R. A.Davis. 1992. Macrophyte removal toenhance bluegill, largemouth bass, andnorthern pike populations. Section ofTechnical Services InvestigationalReport 415. Minnesota Department ofNatural Resources, Division of Gameand Fish, St. Paul.
Crowder, L. B., and W. E. Cooper. 1982.Habitat structural complexity and theinteraction between bluegills and theirprey. Ecology 63:1,802-1,813.
Ellison, A. M. 1996. An introduction toBayesian inference for ecological re-search and environmental decision-making. Ecol. Appl. 6:1,036-1,046.
Engel, S. 1987. The impact of submergedmacrophytes on largemouth bass andbluegills. Lake Reserv. Manage.3:227-234.
. 1995. Eurasian watermilfoil as afishery management tool. Fisheries20(3):20-27.
Frost, T. M., D. L. DeAngelis, S. M. Bar-tell, D. J. Hall, and S. H. Hurlbert.1988. Scale in the design and interpre-tation of aquatic community research.Pages 229-258 in S. R. Carpenter, ed.Complex interactions in lake commu-nities. Springer-Verlag, New York.
Gelman, A., J. B. Carlin, H. S. Stern, andD. B. Rubin. 1995. Bayesian data an-alysis. Chapman and Hall, New York.
Gotceitas, V., and P. Colgan. 1989. Preda-tor foraging success and habitat com-plexity: quantitative test of thethreshold hypothesis. Oecologia80:158-166.
Haller, W. T. 1979. Aquatic weeds in Florida.Pages 1-14 in J. V. Shireman, ed. Pro-ceedings of the Grass Carp Conference.University of Florida, Gainesville.
Haller, W. T., and D. L. Sutton. 1975.Community structure and competitionbetween hydrilla and vallisneria. Hya-cinth Control Journal 13:48-50.
Heck, K. L., and L. B. Crowder. 1991.Habitat structure and predator-preyinteractions in vegetated aquatic sys-tems. Pages 281-299 in S. S. Ball, E. D.McCoy, and H. R. Mushinsky, eds.Habitat structure: the physical arrange-ment of objects in space. Chapman andHall, London.
Jeppesen, E., K. Christoffersen, and M.Sondergaard, eds. In press. The role ofsubmerged macrophytes in structuringlakes. Springer-Verlag, Berlin.
Kitchell, J. E, ed. 1992. Food web manage-ment: a case study of Lake Mendota.Springer-Verlag, New York.
Lawrence, J. M. 1958. Estimated sizes ofvarious forage fishes largemouth basscan swallow. Proc. Ann. Conf. SEAssoc. Game Fish Comm. 11:220-225.
Levin, S. A. 1992. The problem of patternand scale in ecology. Ecology73:1,943-1,967.
Fisheries * 11

FISHERIES MANAGEMENTMittelbach, G. G. 1981. Foraging efficien-
cy and body size: a study of optimaldiet and habitat use by bluegills.Ecology 62:1,370-1,386.
·. 1988. Competition among refugingsunfishes and effects of fish density onlittoral zone invertebrates. Ecology69:614-623.
McAllister, M. K., and R. M. Peterman.1992. Experimental design in the man-agement of fisheries: a review. N. Am.J. Fish. Manage. 12:1-18.
Nibbelink, N. P., and S. R. Carpenter. Inpress. Interlake variation in growthand size structure of bluegill: inverseanalysis of an individual-based model.Can. J. Fish. Aquat. Sci.
Nichols, S. A. 1994. Evaluation of inva-sions and declines of submersedmacrophytes for the Upper Great Lakesregion. Lake Reserv. Manage. 10:29-33.
Olson, M. H. 1996. Predator-prey interac-tions in size-structured fish communi-ties: implications of prey growth.Oecologia 108:757-763.
Peterman, R. M. 1990. Statistical poweranalysis can improve fisheries researchand management. Can. J. Fish. Aquat.Sci. 47:2-15.
Savino, J. E, E. A. Marschall, and R. A.Stein. 1992. Bluegill growth as modi-fied by plant density: an exploration ofunderlying mechanisms. Oecologia89:153-160.
Savino, J. F., and R. A. Stein. 1982. Preda-tor-prey interaction between large-mouth bass and bluegills as influencedby simulated, submersed vegetation.Trans. Am. Fish. Soc. 111:255-266.
Shireman, J. V., M. V. Hoyer, M. J.Maceina, and D. E. Canfield, Jr. 1985.The water quality and fishery of LakeBaldwin, Florida, four years aftermacrophyte removal by grass carp.Lake Reserv. Manage. 1:201-206.
Smith, K. D. 1993. Vegetation-open waterinterface and the predator-prey interac-tion between largemouth bass andbluegills. Doctoral dissertation. Univer-sity of Michigan, Ann Arbor.
Stewart-Oaten, A., W. W. Murdoch, andK. R. Parker. 1986. Environmental im-pact assessment: "pseudoreplication"in time? Ecology 67:929-940.
Swingle, H. S., and E. V. Smith. 1941. Themanagement of ponds with stuntedfish populations. Trans. Am. Fish. Soc.71:102-105.
Timmons, T. J., W. J. Shelton, and W. D.Davies. 1980. Differential growth oflargemouth bass in West Point Reser-voir, Alabama-Georgia. Trans. Am.Fish. Soc. 1098:176-186.
Tonn, W. M., and C. A. Paszkowski. 1986.Size-limited predation, winter-kill, andthe organization of Umbra-Percaassemblages. Can. J. Fish. Aquat. Sci.43:194-202.
Trebitz, A. S., and N. P. Nibbelink. 1996.Effect of pattern of vegetation removalon growth of bluegill: a simple model.Can. J. Fish. Aquat. Sci. 53:1,844-1,851.
Trebitz, A. S., and 11 coauthors. 1997. Amodel of bluegill-largemouth bassinteractions in relation to aquatic vege-tation and its management. Ecol.Model. 94:139-156.
Werner, E. E., and D. J. Hall. 1988. Ontoge-netic habitat shifts in bluegill: the forag-ing rate-predation risk trade-off.Ecology 69:1,352-1,366.
Wiley, M. J., R. W. Gorden, S. W. Waite,and T. Powless. 1984. The relationshipbetween aquatic macrophytes andsport-fish production in Illinois ponds:a simple model. N. Am. J. Fish.Manage. 4:111-119.
12 * Fisheries Vol. 23, No. 2





























