Embed Size (px)
Citation preview

Carctnogenesls vol.17 no.5 pp.1101-1106, 1996
Rat liver epithelial cells express functional cytochrome P450 2E1
Carole Lerche, Catherine Le Jossic, Alain Fautrel,Isabelle de Waziers1, Francois Ballet2, Andre Guillouzoand Laurent Corcos3
INSERM U49, CHRU Pontchaillou, 35033 Renncs Cedex, 'INSERM U75,CHU Necker, 75730 Paris Cedcx 15 and 2Rh6ne-Poulenc Rorer SA.94403 Vitry sur Seine, France
^To whom correspondence should be addressed
Rat liver epithelial cells (RLECs) isolated by trypsinizationof the h'vers of normal 10 day old rats are largely used inco-culture with primary hepatocytes. The aim of the presentstudy was to investigate the expression of biotransformationenzyme-encoding genes in three preparations of RLEClines. Although no expression of cytochrome P4S0 1A1/2(CYP1A1/2), CYP2B1/2, CYP2C6 or CYP3A mRNAs couldbe detected, we found that all of the different preparationsof RLECs expressed a high level of CYP2E1 mRNA. Wedemonstrated the presence of the CYP2E1 apoprotein inmicrosomes of RLECs by immunoblot analyses, togetherwith chlorzoxazone 6-hydroxylation, an activity known tobe mainly catalyzed by CYP2E1. In addition, acetonetreatment of these cells resulted in an increase in bothCYP2E1 apoprotein and chlorzoxazone 6-hydroxylationactivity levels. Finally, we showed the susceptibility ofRLECs to JV-methyl formamide- and diethylnitrosamine-induced toxicity, suggesting metabolic activation byCYP2E1. Thus, RLECs may cooperate with hepatocytesto CYP2E1-mediated metabolism in the co-culture model.In addition, transfection experiments with a CYP2E1 pro-moter construct, in which the proximal 539 bp containingthe binding site for HNFla were inserted upstream of thechloramphenicol acetyl transferase gene, demonstrated astrong induction upon co-transfection with an HNFlaexpression plasmid. Thus, RLECs provide a useful tool forstudying metabolism and cytotoxicity of CYP2E1 substratesin the absence of other expressed CYPs, and for analyzingCYP2E1 promoter function.
Introduction
Hepatocytes express an array of essential functions such asproduction of plasma proteins and metabolism of endogenousand exogenous compounds. Disturbances of their pericellularenvironment, occurring during isolation and plating in vitro,result in early decrease or loss of most of their liver-specificfunctions. Such phenotypic changes can be delayed and/orreduced by culturing hepatocytes either on matrigel, a mixtureof basement membrane proteins (1,2) or in association withundifferentiated rat liver epithelial cells (RLECs*) (3). RLEC
•Abbreviations: RLEC, rat liver epithelial cell; CYP, cytochrome P45O,NMF, yV-methylformamide; DEN, diethylnitrosamine; SD, Sprague-Dawley;F, Fisher; WEM, Williams' E medium; FCS, fetal calf serum; RPE, relativeplating efficiency; TBS, Tris-buffered saline; EROD, ethoxyresorufin O-deethylase; CAT, chloramphenicol acetyl transferase; (Jgal, fj-galactosidase.
lines have been isolated by many investigators (for review,see ref. 4). They bear no morphological resemblance toauthentic hepatocytes or bile duct cells (5,6). In addition, thesecells express low amounts of y-glutamyltranspeptidase, andmay contain low amounts of glucocorticoid-inducible tyrosineaminotransferase (5). These RLEC lines, which can be propa-gated in long-term culture, have been the object of manystudies with regard to their origin in vivo and their involvementin chemical-induced hepatocarcinogenesis (4,7,8). However,the diversity of RLEC lines, which may result from differencesin the differentiation status of the cell of origin in vivo or incell purity following isolation, makes it difficult to define aunique phenotype and requires extensive analysis of individu-ally cloned populations.
The participation of RLECs in liver-specific functions in vitrohas usually been considered negligible. However, xenobioticbiotransformation activities, such as sulfotransferase (9) andglutathione S-transferase (10), have been reported in RLEClines. In addition, benzo[a]pyrene metabolism has beendetected in an epithelial cell line isolated from the liver of anadult female Wistar rat (12). These observations led us toreconsider the potential participation of RLECs in the metabol-ism of xenobiotics in vitro. Our results demonstrate that,among the cytochromes P450 (CYP) surveyed, only CYP2E1mRNA, protein and enzyme activity were expressed in thesecells. Moreover, in vitro cytotoxicity was induced followingN-methylformamide (NMF) and diethylnitrosamine (DEN)activation by CYP2E1. Consequently, RLECs are likely toparticipate, in concert with hepatocytes, in CYP2E1-mediatedxenobiotic metabolism in vitro.
Materials and methodsCell isolation and cultureRLECs have been obtained by trypsinization of the livers of normal 10 dayold Sprague-Dawley (SD) or Fisher (F) rats, as described elsewhere (5). TheRLECs (three preparations named SDII, SDVI and F388) and H5 rat hepatomacells (13) were cultured in Williams' E medium (WEM) supplemented with10% fetal calf serum (FCS). Hepatocytes were isolated from SD male ratsusing the two-step collagenase perfusion method and cultured in a mediumconsisting of 75% minimum essential medium and 25% 199 medium, aspreviously described (11), except that the cells were seeded at a density of18X106 cells/175 cm2 flasks. FAO cells were cultured in a mixture of Ham'sF-12 and NCTC 135 (50%/50%, v/v) with 10% FCS as reported previously(13). Acetone was added for 72 h at a final concentration of 30 mM, 4 h aftercell seeding for hepatocyte cultures and 2 days before confluency for RLECcultures. Acetone treatment had no apparent effect on cell growth. Cytotoxicityof both NMF and DEN was initially assayed by determination of lactatedehydrogenase release, followed by determination of the relative platingefficiency (RPE) (see legend to Table I).Isolation of total RNA and blot analysesCell monolayers were scraped in 0.1 M PBS. Total RNA was prepared asdescribed previously (14). RNA blot analyses and hybridization reactionswere performed as described by Abdel-Razzak et al. (15), except that Hybond-N nylon filters were used for transfer. cDNA probes were 32P-labeled byrandom priming using cDNAs to rat hepatic mRNAs and included: CYP 2E1(full-length rat and human cDNA probes) (16), CYP 2C6 (pTF2, partialcDNA) (17), tyrosine aminotransferase (pcTAT3, partial cDNA) (18), epoxidehydrolase (full-length cDNA) (19). An 18S genomic DNA probe was used asRNA loading control. Filters were autoradiographed at -80°C.
© Oxford University Press 1101
at Beijing U
niversity of Technology on July 15, 2011
carcin.oxfordjournals.orgD
ownloaded from
CLerche et al
Tbble I. Comparison of CYP2E1 -mediated cytotoxicity in H5 and SDVI cells, respectively negative and positive for CYP2E1 expression
Cells
H5
SDVI
Exp. 1Exp. 2'Exp. 1Exp. 21
-NMF
1010b
2210114495
+NMF
8501855
48169
RPE
0.840.840.420.34
-DEN
ndnd209141
+ DEN
ndnd8677
RPE
_
_0410.55
•Represents the average of duplicate plates.bNumber of colonies.nd, Not determined.Cells were treated with 250 mM NMF or 40 mM DEN for 24 h. After treatment, cultures were rinsed, cells detached, counted and inoculated fordetermination of the relative plating efficiency (RPE, number of colonies from treated/untreated cultures). One to two weeks later, colonies were fixed, stainedand counted.
Immunoblot detection of CYP2E1
Microsomal fractions were obtained as described elsewhere (20), except thatcell pellets were sonicated for 10 s and homogenized for 45 s with an Ultra-turrax (Janke and Kunkel, IKA-labortechnik).
Microsomal proteins were separated by SDS-PAGE (9% acrylamide)according to Laemmli (21). Proteins were transferred onto nitrocellulose filtersas described (22). Subsequently, the filters were blocked with Tris-bufferedsaline (TBS) containing 3% creamed milk for 2 h at 20°C. The filters werethen incubated overnight, at 4°C, with an antiserum containing anti-peptideantibodies directed against a 35 kDa peptide derived from the human CYP2E1protein (amino acids 145—463) (23), and known to recognize the rat protein(unpublished observations). After five washes at 20°C with TBS/0.3% Tween20, the filters were incubated with alkaline phosphatase-conjugated anti-antibody for 2 h. The blots were then washed and alkaline phosphataseconjugates were detected as described by Blake et al. (24).
Monooxygenase activities
Ethoxyresorufin O-deethylase (EROD) activity was assayed on microsomalpreparations from both cultured cells and liver tissue according to the methodof Burke et al. (25). Chlorzoxazone 6-hydroxylation activity assays wereperformed on microsoma] preparations as described previously (26), exceptthat 1 mM NADPH was used instead of an NADPH-generating system andthat incubation time was 25 min.
Mutagenesis experiments
Cells in growth phase at a density ranging between 0.4X105 and O.8XIO5/cm2 were exposed to DEN for 24 or 72 h at a final concentration of 40 mM.Cells were then trypsinized and reseeded for determination of the RPE or formutation expression time. Finally, they were used for a new determination ofthe RPE and submitted to ouabain selection by plating 0.02X105 cells/cm2.Ouabain was added 4 days later at the final concentration of 2, 3 or 5 mMand the medium was changed every 2 days. After 10 days, colonies werefixed, stained and counted.
Transfection experiments
pRSV-HNFloc (HNFloc cDNA transcribed from the Rous sarcoma viruspromoter), pSBI-CAT (promoterless plasmid), pSVE-CAT [SV40 early pro-moter driving chloramphenicol acetyl transferase (CAT) expression] and thepRSV-P-galactosidase (Pgal) plasmid were generous gifts from M.Yaniv. TheCYP2E1 promoter vector contained a 539 bp fragment from the rat genepromoter driving CAT (27).
SDVI cells were transfected with plasmid DNA by electroporation using aGene Pulser apparatus (Eurogentec, Seraing, Belgium). Four hundred microlit-ersofcell suspension containing 25X106 cells, in WEM with 10% FCS, weremixed with 30 jig plasmid DNA. The mixture was subjected to a single pulseof 1800 (iF at 230 V. After electroporation, the cells were transferred to25 cm2 flasks containing WEM supplemented with 10% FCS, and thenincubated for 48 h at 37*C. Cells were harvested and extracts prepared forCAT and Pgal activity assays (28).
Results
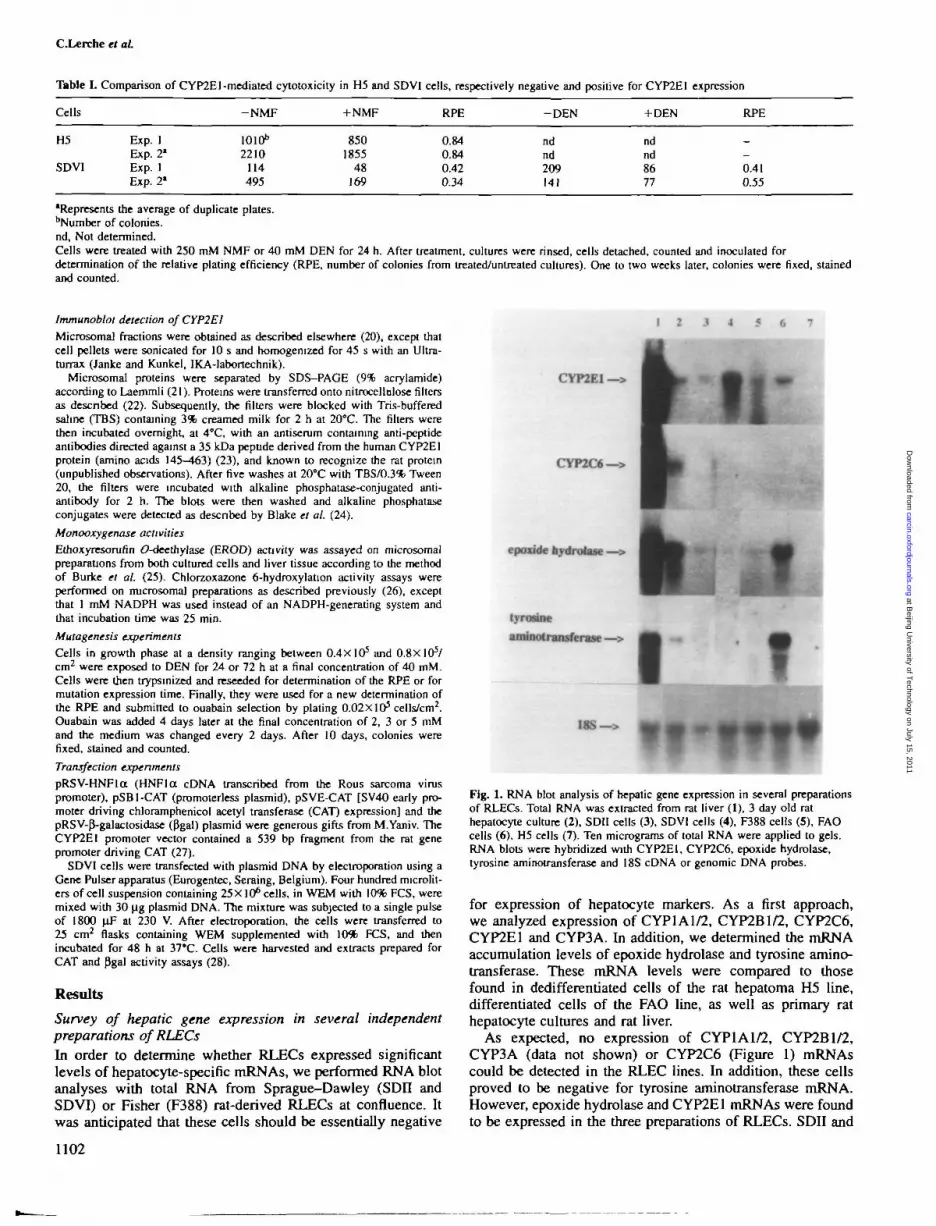
Survey of hepatic gene expression in several independentpreparations of RLECsIn order to determine whether RLECs expressed significantlevels of hepatocyte-specific mRNAs, we performed RNA blotanalyses with total RNA from Sprague-Dawley (SDH andSDVI) or Fisher (F388) rat-derived RLECs at confluence. Itwas anticipated that these cells should be essentially negative
1 2 3 4 5 6
CYP2E1—>
CYP2C6—>
epoxide hydrolase —>
It
tyrortneaminotransferasc —>
J f18S—>
Fig. 1. RNA blot analysis of hepatic gene expression in several preparationsof RLECs. Total RNA was extracted from rat liver (1), 3 day old rathepatocyte culture (2), SDII cells (3), SDVI cells (4), F388 cells (5), FAOcells (6), H5 cells (7). Ten micrograms of total RNA were applied to gels.RNA blots were hybridized with CYP2E1, CYP2C6, epoxide hydrolase,tyrosine aminotransferase and 18S cDNA or genomic DNA probes.
for expression of hepatocyte markers. As a first approach,we analyzed expression of CYP1A1/2, CYP2B1/2, CYP2C6,CYP2E1 and CYP3A. In addition, we determined the mRNAaccumulation levels of epoxide hydrolase and tyrosine amino-transferase. These mRNA levels were compared to thosefound in dedifferentiated cells of the rat hepatoma H5 line,differentiated cells of the FAO line, as well as primary rathepatocyte cultures and rat liver.
As expected, no expression of CYP1A1/2, CYP2B1/2,CYP3A (data not shown) or CYP2C6 (Figure 1) mRNAscould be detected in the RLEC lines. In addition, these cellsproved to be negative for tyrosine aminotransferase mRNA.However, epoxide hydrolase and CYP2E1 mRNAs were foundto be expressed in the three preparations of RLECs. SDII and
1102
at Beijing U
niversity of Technology on July 15, 2011
carcin.oxfordjournals.orgD
ownloaded from

Rat liver epithelial cells express functional CYP2E1
I 2 3 4 5 6
fffttfttfftffFig. 2. Stability of expression of CYP2E1 mRNA in SDVT cells. Total RNAwas extracted from SDV1 cells studied between passages 11 (3) and 22(14). Ten micrograms of RNA were applied to gels. 1 and 2 refer to ratliver and 3 day old rat hepatocyte cultures, respectively.
F388 cells expressed the same level of CYP2E1 mRNA ashepatocytes cultured for 3 days, whereas SDVI cells expresseda 7-fold higher level of this mRNA. In addition, specificamplification of exonic sequences followed by HaeUldiagnostic restriction enzyme digestion demonstrated that thesignal observed on RNA filters was indeed that expected fromthe known structural features of the rat CYP2E1 mRNA(data not shown). SDVI cells were then used for subsequentexperiments.
The observation of a strong expression level of CYP2E1mRNA species prompted us to ask whether long-term stabilitycould be demonstrated. To this end, we determined, by RNAblot analysis, CYP2E1 expression over 12 passages of SDVIcells analyzed between passages 11 and 22 (Figure 2). Overall,no differences in CYP2E1 mRNA expression levels werefound, supporting the idea of a strong phenotypic stabilitywith respect to expression of this CYP gene.
Detection of CYP2E1 apoprotein by Western blot analysesWe then investigated presence of the CYP2E1 protein inmicrosomes from SDVI cells, in comparison with rat hepato-cytes from 3 day old cultures, as well as rat liver. To this end,a specific anti-peptide antibody directed against a peptidesequence derived from human CYP2E1, and known to recog-nize the rat protein very efficiently, was used.
As shown in Figure 3(A), a strong signal was observed inrat liver microsomes. After 3 days of culture, the level ofCYP2E1 protein was markedly reduced. Acetone, a knowninducer of the CYP2E1 protein, induced an increase, after72 h of treatment with 30 mM, of the apoprotein level in rathepatocytes (~ 10-fold). Similarly, a signal was observed inSDVI cell microsomes, which was increased by acetone (2-fold). By contrast with RNA blot analyses, the CYP2E1apoprotein signal was slightly higher in hepatocytes than inSDVI cells.Functional activity of CYP2E1 in SDVI cellsFinally, we looked to see if CYP2E1-associated enzymeactivity could be evidenced in SDVI cells. Chlorzoxazone 6-hydroxylation activity has been shown to be supported mainlyby CYP2E1, although high levels of CYP1A1 could result insignificant activity (26). However, since no CYP1A1 mRNAor EROD activity could ever be detected in both controland acetone-treated SDVI cells, it was expected that anychlorzoxazone 6-hydroxylation activity should only be due topresence of the CYP2E1 enzyme.
As seen in Figure 3(B), rat liver microsomes showed astrong activity toward chlorzoxazone. This activity was mark-
KIM
CYP2E1-50-35-
B
Rat liver
Hepatocytes
SDVI cells
Basal activity
Basal activity
Induction level
Basal activity
Induction level
Chlorzoxazone 6-hydroxylation(pmoles/min/mgofmicrosomalprotein)
1214.43 ±40.63'
25.89 ±3.25*
1.49
17.85 ±7.42b
1.40
EROD(pmolesAnin/mgof microsonialprotein)
75J5±9JO»
10.9211.92"
1.14
Fig. 3. Expression of CYP2E1 apoprotein in SDVI cells. Microsomalproteins were extracted from rat liver, primary rat hepatocyte cultures andSDVI cells. Forty micrograms of proteins were analyzed byimmunoblotting. (A) The following samples were analyzed: lane 2, ratliver, lanes 3 and 4, untreated and acetone-treated 3 day old hepatocytecultures; lanes 5 and 6, untreated and acetone-treated SDVI cells. Theposition of ovalbumin (mol. wt 50 000) and CYP2E1 peptide (lane 1, mol.wt 35 000) are indicated. (B) Chlorzoxazone 6-hydroxylation and ERODactivities were measured as described in Materials and methods. 'Activityrepresents the average of two separate experiments ± SD. bActivityrepresents the average of three separate experiments ± SD.
edly reduced in 3 day old hepatocyte cultures. Acetonetreatment resulted in both hepatocyte and SDVI cell culturesin a slight increase of chlorzoxazone 6-hydroxylation (1.5-and 1.4-fold respectively). In addition, since each determinationof this activity required at least 800 (ig of microsomal proteins,we decided to perform three independent experiments forSDVI cells, rather than multiplicate determinations from asingle experiment
As an ultimate proof of the functional activity of CYP2E1in SDVI cells, we analyzed their susceptibility to NMF- andDEN-induced toxicity. Indeed, NMF and DEN have beenshown to exhibit hepatotoxic activity following metabolicactivation into cytotoxic derivatives by CYP2E1 (29,30).Moreover, numerous studies have reported the genotoxicity ofDEN (30,31). Increased lactate dehydrogenase release wasobserved in SDVI cell culture medium following exposure to500 mM NMF or 40 mM DEN for 24 h. In addition, no effectwas observed in NMF-treated H5 cells lacking CYP2E1expression (data not shown). Plating efficiency experimentsshowed a significant decrease in cloning ability of NMF(250 mM)-treated SDVI cells, as compared to the resistant H5cells (Table I). Similar results were obtained for DEN (40 mM>treated SDVI cells. These results strongly suggest that SDVIcells express a functional CYP2E1 enzyme, and are able toactivate hepatotoxic compounds into cytotoxic products.Finally, to test if SDVI cells could be used for the detectionof potential genotoxic compounds, involving CYP2E1 activa-tion, we performed a 3 day DEN treatment of SDVI cellsfollowed by the selection of mutant colonies in the presence
1103
at Beijing U
niversity of Technology on July 15, 2011
carcin.oxfordjournals.orgD
ownloaded from

C.Lerche el at
Plaonids
CAT activity/p-gal activity(irbitniy units)
%ofSVE-CAT
activity
Exptl
Expt2
Expt3
Exptl
Expt2
Expt3
SB 1 CAT
3.06
3.54
10.5
1.96
2.82
2.40
SVE-CAT
155.65
125.50
437.50
100
100
100
CYP2E1-CAT
4.13
1.36
5.43
2.65
1.08
1.24
CYP2E1-CAT+ HNFla
9.38
15.82
16.00
6.03
12.60
3.65
Fig. 4. Transfection experiments in SDVI cells. Expression plasmids(30 |ig) together with 10 ng of pRSVpgal were introduced into SDVI cellsby electroporation. After 48 h of incubation, cells were scraped forchloramphenicol acetyltransferase (CAT) and P-galactosidase ((3gal) assays.
of 3 mM ouabain. This selection of putative ouabain resistantcolonies was made after 2, 3 or 5 days of mutation expressiontime. Although DEN-induced cytotoxicity was observed aftertreatment (and during the first three days of expression time),very few resistant colonies were observed after selection inouabain-containing medium, suggesting only a small mutageniceffect of DEN in SDVI cells (data not shown).
Regulation of CYP2E1 promoter activity in SDVI cellsPrevious studies have shown that the CYP2E1 gene wascontrolled by a protein displaying DNA binding propertiessimilar to those of HNFla (32). We wondered whether HNFlacould take part in the regulation of the CYP2E1 gene in SDVIcells. We performed transfection experiments with a CYP2E1promoter construct in which the proximal 539 bp, containingthe binding site for HNFla, were inserted upstream of thebacterial CAT gene (27). Figure 4 shows that basal CYP2E1promoter activity was similar or lower than that of promoterlessplasmid activity in SDVI cells, but co-transfection of theC YP2E1 promoter construct and an HNF 1 a expression plasmidled to an increase of CAT activity, reaching up to 11.6-fold.These results suggest that overexpression of HNFla cantransactivate the CYP2E1 promoter in SDVI cells.
Discussion
The RLEC lines characterized in this study have been com-monly used in co-culture with primary hepatocytes (3). Insuch a system, expression of a large set of hepatic functionshas been shown to be stably maintained over several weekswithin hepatocytes. Recent data have suggested that the ter-minal ductule cells connecting the canals of Hering with bile
1104
canaliculi and/or a distinct population of periductal cellsconstitute a hepatic stem cell compartment (7,8,33,34). Thepossibility that RLECs are derived from this compartment hasbeen suggested. Moreover, Marceau (35) has studied the celltyping of liver ECs isolated by various tissue dispersionprocedures and concluded that some liver ECs represent anearly sublineage of progenitors committed along theintrahepatic biliary EC lineage. However, it should be noticedthat some EC lines have been shown to express a few ofthe hepatocytic metabolic activities and to differentiate intohepatocytes after intrahepatic transplantation (36,37).
RLECs can be propagated in long-term culture and theirphenotypic stability and homogeneity have permitted to studyseveral of their characteristics, such as morphology and liver-specific function expression. Similarly to our observations,CYP-dependent monooxygenase activities (EROD and 7-ethoxycoumarin (9-deethylase) were undetectable in RLECsand were not inducible by benz[a]anthracene (9), whereascytochrome b5 and NADPH cytochrome P450 reductase weredetected (38). Furthermore, epoxide hydrolase (this work),glucuronosyltransferase, sulfotransferase (9) and glutathioneS-transferase activities (10) were found in these cells. Inaddition, our results showed that RLECs stably expressCYP2E1 mRNA, protein and enzyme activity. Of the threeRLEC preparations, only SDVI cells express a high levelof CYP2E1 mRNA. Differences in carcinogen susceptibilitybetween RLEC lines have already been reported by Tong et al.(39). Therefore, our results could conceivably be explained byintrinsic differences between lines in the biotransformationcapacities of the various RLEC preparations. In addition,strain-specific differences in CYP2E1 expression could explainthe low CYP2E1 mRNA level observed in F388 cells. To ourknowledge, this paper is the first reporting the induction byacetone of CYP2E1 in vitro in rat hepatocytes and liverepithelial cells. By contrast with immunoblot analyses, acetonetreatment of hepatocytes cultures led to only a slight increasein chlorzoxazone 6-hydroxylation activity. In line with theseresults, it has been shown that treatment of human hepatocytecultures with ethanol resulted in a 7.7-fold increase in CYP2E1apoprotein but only in a 2.7-fold increase in chlorzoxazone 6-hydroxylation activity (26).
The fact that RLECs stably express basal and acetone-inducible CYP2E1 make these cells likely to represent agood model system for CYP2E1-related metabolism studies.Moreover, we have shown that RLECs were able to activateNMF and DEN, some known CYP2E1 substrates, into cyto-toxic metabolites. Furthermore, we tried to look for possiblemutagenic effects of CYP2E1-activated DEN. However,although appearance of micronuclei in RLECs has beenreported (12), we failed to demonstrate a significant genotoxicactivity of DEN in SDVI cells, although we were able to scorea few ouabain-resistant colonies. This observation could bedue to the particular type of pro-mutagen used or to lowstability within the RLECs of the mutagenic metabolites ofDEN. Alternatively, this result may suggest a marked capacityfor the repair of DEN-induced lesions in RLECs. Othermutagen compounds could be tested in RLECs. Nevertheless,although our results suggest that RLECs could be used inCYP2E1-mediated cytotoxicity studies, this assay system can-not be readily applied to routine screening of suspectedenvironmental mutagens.
Obviously, several other in vitro systems have beendeveloped that would also fulfil some of the proposed tasks
at Beijing U
niversity of Technology on July 15, 2011
carcin.oxfordjournals.orgD
ownloaded from

Rat liver epithelial cells express functional CYP2E1
using RLECs. Our results suggest that co-cultures betweenRLECs and primary hepatocytes may be a useful model forCYP2E1-related pharmaco-toxicological studies, in view ofthe high CYP2E1 expression in both cell types in co-cultures(data not illustrated). Furthermore, the expression of clonedcDNAs coding for each CYP isoform in heterologous systems(for review, see ref. 40) permits high production of the enzymesas well as metabolism studies. However, several of thesesystems have the disadvantages of being transient, havingonly relatively low activities of NADPH cytochrome P450reductase. Moreover, some of them require modifications ofthe amino-terminal region of the enzyme or need a long timefor isolating an expressing clone (40).
RLECs may be a useful model for CYP2E1 promoteranalyses since co-introduction of an HNFla expressionplasmid and a CYP2E1 promoter in SDVI cells leads to astrong increase in reporter gene activity. It remains to bedetermined whether endogenous HNFla is responsible fordirecting expression of the CYP2E1 gene in these cells, sinceour preliminary experiments failed to show substantial HNFlaDNA binding activity in gel-retardation assays (data notshown). Recently, Liu et al. (41) have shown that HNFlaplays a key role in the activation of the CYP2E1 gene inhepatoma cells. The possible influence of xenobiotics on thevarious allelic forms of the CYP2E1 promoter, one of whichappears to alter HNFla binding, could be investigated follow-ing stable integration in RLECs. In addition, the members ofsuch transcriptional regulators are known to be implicated inthe control of cell differentiation and proliferation and in theliver-specific expression of many genes. The studies, in RLECs,of expression of the fra/ts-acting factors could explain theabsence of the other detoxication functions surveyed, particu-larly the other CYPs like CYP2C, which is known to requireHNF3 and HNF4 for liver-specific expression (42,43). Inaddition, the fact that less CYP2E1 apoprotein and enzymeactivity could be detected in RLECs as compared to hepatocytecultures, despite the higher mRNA levels, might suggestdifferences in regulation pathways controlling both transcrip-tional and post-transcriptional steps in the two cell types.
Finally, a promising area of investigation is the in vivoanalysis of CYP2E1 expression and activity in RLECs. In thatrespect, it should be first determined whether the enzyme canindeed activate pro-carcinogens into their ultimate harmfulmetabolites. In this case, one could expect the appearance ofliver tumors to originate in this subcellular population followingCYP2E1-mediated carcinogen activation.
Acknowledgements
We thank B.Clement, A.Corlu, N.Ferry, D.Glaise, G.IIyin, O.Loreal andC.Guillouzo for helpful discussions, and M.Yaniv and F.J.Gonzalez for theirgifts of cDNA and promoter expression plasmids. This work was supportedby the INSERM and the Association pour la Recherche Contre le Cancer.C.L. is the recipient of a doctoral fellowship from Rhone-Poulenc Rorer.
References
1. Schuetz,E.G., Li.D., Omiecinski,CJ., Muller-Eberhard,U., Kleinman.H.K.,Elswick.B. and Guzelian.P.S. (1988) Regulation of gene expression inadult rat hepatocytes cultured on a basement membrane matrix. J. CellPhysiol., 134, 309-323.
2.Hunt,C.M., Guzelian.P.S., Molowa,D.T. and Wright,S.A. (1991) Regulationof rat hepatic cytochrome P450 2EI in primary monolayer hepatocyteculture. Xenobiotica, 21, 1621-1631.
3.Guguen-Guillouzo,C, Clement,B., Baffet,G., Beaumont,C, Morel-Chany.E., Glaise.D. and Guillouzo.A. (1983) Maintenance and reversibility
of active albumin secretion by adult rat hepatocytes co-cultured withanother liver epithelial cell type. Exp. Cell Res., 143, 47-54.
4.MarceauJM., Blouin,M-J, Noel.M., T6r0k,N. and Loranger.A. (1992) Therole of bipotential progenitor cells in liver ontogenesis and neoplasia . InSirica^A.E. (ed.), The Role of Cell Types in Hepatocarcinogenesis. CRCPress, Boca Raton, FL, pp. 121-150.
5.Morel-Chany,E., Guillouzo.C, Trincal.G. and Szajnert,MF. (1978)'Spontaneous' neoplastic transformation in vitro of epithelial cell strainsof rat liver cytology, growth and enzymatic activities. Eur. J. Cancer, 14,1341-1352.
6.Tsao,M.-S., SmithJ.D., Nelson.K.G. and GrishamJ.W. (1984) A diploidepithelial cell line from normal adult rat liver with phenotypic propertiesof 'oval' cells. Exp. Cell Res., 154, 38-52.
7.Sell,S. (1994) Liver stem cells. Mod. Pathol., 7, 105-112.8.Thorgeirsson,S.S. (1993) Hepatic stem cells. Am. J. Pathol, 142, 1331-
1333.9. Schrenk.D., Eisenmann-Tappe,I., Gebhardt,R., MayerJD., El Mouelhijvi.,
Rohrdanz.E., MunzelJ". and Bock.ICW. (1991) Drug metabolism enzymeactivities in rat liver epithelial cell lines, hepatocytes and bile duct cells.Biochem. Pharmacol, 41, 1751-1757.
lO.Vandenberghe.Y., Morel.F., Pemble.S., TaylorJ.B., Rogiers.V,Ratanasavanh.D., Vercruysse,A., Ketterer.B. and Guillouzo.A. (1990)Changes in expression of mRNA coding for glutathione S-transferasesubunits 1-2 and 7 in cultured rat hepatocytes. Mol Pharmacol, 37,372-376.
ll.Gugucn-Guillouzo.C. (1992) Isolation and culture of animal and humanhepatocytes. In Freshney.R.I. (ed.). Culture of Epithelial Cells. AlanR.Liss, Glasgow, pp. 197-223.
12.Glatt,H., Gemperlein.I., Setiabudi,R, Platt.K.L. and Oesch.F. (1990)Expression of xenobiotic-metabolizing enzymes in propagatable cellcultures and induction of micronuclei by 13 compounds. Mutagenesis, 5,241-249.
13.Deschartrctte,J. and Weiss,M.C. (1974) Characterization of differentiatedand dedifferentiated clones from a rat hepatoma. Biochimie, 56,1603-1611.
14.Chomczynski,P. and Sacchi,N. (1987) Single-step method of RNA isolationby acid guanidinium thiocyanate-phenol-chloroform extraction. Anal.Biochem., 162, 156-159.
15.Abdel-Razzak,Z., Corcos.L., Fautrel.A., CampionJ.P. and Guillouzo^A.(1994) Transforming growth factor-fil down-regulates basal and polycyclicaromatic hydrocarbon-induced cytochromes P-450 1A1 and 1A2 in adulthuman hepatocytes in primary culture. Mol. Pharmacol, 46, 1100-1110.
16.Song,B.-J., Gelboin.H.V., Park,S.S, Yang.C.S. and Gonzalez,F.J. (1986)Complementary DNA and protein sequences of ethanol-inducible rat andhuman cytochrome P^50s. J. Biol. Chem., 261, 16689-16697.
17.Friedberg,T., Waxman.DJ., Atchinson.M., Kumar^A., Haaparanta,T.,Raphael.C. and Adesnikjvl. (1986) Isolation and characterization of cDNAclones for cytochromes P-450 immunochemically related to rat hepatic P-450 form PB-1. Biochemistry, 25, 7975-7983.
18.Scherer,G., Schmid.W., Strange.C, Rowckamp.W. and SchUtz.G. (1982)Isolation of cDNA clones coding for rat tyrosine aminotransferase. Proc.Natl Acad. Sci. USA, 79, 7205-7208.
19.Porter,T.D., Beck,T.W. and Kasper.C.B. (1986) Complementary DNA andamino acid sequence of rat liver microsomal, xenobiotic epoxide hydrolase.Arch. Biochem. Biophys., 248, 121-129.
20.Morel,F, Beaune.P.H., Ratanasavanh.D., FlinoisJ.-P., Yang.C.S.,Guengerich,F.P. and Guillouzo,A. (1990) Expression of cytochrome P^»50enzymes in cultured human hepatocytes. Eur. J. Biochem., 191, 437-444.
21.Laemmli,U.K. (1970) Cleavage of structural proteins during the assemblyof the head of the bacteriophage T4. Nature, 227, 680-685.
22.LoyerJ"., Glaise.D., Cariou.S., Baffet,G., Meijer.L. and Guguen-Guillouzo.C. (1994) Expression and activation of cdks (1 and 2) andcyclins in the cell cycle progression during liver regeneration. J. BiolChem., 269, 1-10.
23.Belloc,C, Baird.S., Lecoeur.S., CosmeJ., GautierJ.-C., ChallineJD., deWaziersX, FlinoisJ.-P. and BeauneJ».H. (1996) Human cytochromes P-450 expressed in Eschenchia coli: production of specific antibodies.Toxicology, 106, 207-219.
24.Blake,M.S., Johnston.K.H., Russel-Jones.GJ. and Gotschlich.E.C. (1984)A rapid, sensitive method for detection of alkaline phosphatase conjugatedanti-antibody on western blots. Anal. Biochem., 136, 175-179.
25.Burke,M.D. and Mayer.R.T. (1983) Differential effects of phenobarbitoneand 3-methylcholanthrene induction on the hepatic microsomal metabolismand cytochrome P-450-binding of phenoxazone and a homologous seriesof its Malkyl ethers (allcoxyresorufin). Chem.-Biol. Interactions, 45,243-258.
26. Carriere.V., Goasduff.T, Ratanasavanh.D., Morel.F., GautierJ.-C.,Guillouzo,A., Beaune,P.F. and Berthou.F. (1993) Both cytochromes P450
1105
at Beijing U
niversity of Technology on July 15, 2011
carcin.oxfordjournals.orgD
ownloaded from

CLerehe et at
2E1 and 1A1 are involved in the metabolism of chlorzoxazone. Chem,Res. Toxicol., 6, 852-857.
27.Umeno,M., McBride.O.W., Yang.C.S., Gelboin.H.V. and Gonzalez,FJ.(1988) Human ethanol-inducible P450HE1: complete gene sequence,promoter characterization, chromosome mapping, and cDNA-directedexpression. Biochemistry, 27, 9006-9013.
28.Maniatis,T., Fritsch.E.F. and SambrookJ. (1982) Molecular Cloning: ALaboratory Manual. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
29.Hyland,R., Gescher,A., Thummel.K., Schiller.C, Jheeta,P., Mynett,K.,Smith,A.W. and MrazJ. (1992) Metabolic oxidation and toxification ofAf-methylformamide catalysed by the cytochrome P450 isoenzymeCYP2E1. Mol. Pharmacol., 41, 259-266.
30.Loquet,C. and Wiebel.FJ. (1982) Geno- and cytotoxicity of nitrosamines,aflatoxin Bl, and benzo[a]pyrene in continuous cultures of rat hepatomacells. Carcinogenesis, 3, 1213-1218.
31.Yamazaki,H., Oda,Y., Funae.Y, Imaoka,S., Inui.Y, Guengerich.F.P. andShimadaJ". (1992) Participation of rat liver cytochrome P450 2E1 inthe activation of A'-nitrosodimethylamine and W-nitrosodiethylamine toproducts genotoxic in an acetyltransferase-overexpressing Salmonellatyphimurium strain (NM2009). Carcinogenesis, 13, 979-985.
32.Ueno,T. and GonzalezJU. (1990) Transcriptional control of the rat hepaticCYP2E1 gene. Mol. Cell. Biol, 10, 4495-4505.
33.Hixson,D.C, Faris.R.A., Yang.L. and Novikoff.P. (1992) Antigenic cluesto liver development, renewal, and carcinogenesis: an integrated model.In Sirica^.E. (ed.), The Role of Cell Types in Hepatocarcinogenesis. CRCPress, Boca Raton, FL, pp. 152-181.
34.YavorkovskyX-, Lai,E-, Ilic,Z. and Sell.S. (1995) Participation of smallintraportal stem cells in the restitutive response of the liver to penportalnecrosis induced by ally! alcohol. Hepatology, 21, 1702-1712.
35.Marceau,N. (1994) Epithelial cell lineages in developing, restoring, andtransforming liver evidence for the existence of a "differentiation window'.Gut, 35, 294-296.
36.Coleman,W.B., Wennerberg.A.E., Smith.CJ. and GrishamJ.W. (1993)Regulation of the differentiation of diploid and some aneuploid rat liverepithelial (stemlike) cells by the hepatic microenvironmenL Am. J. Pathoi,142, 1373-1382.
37. Shiojiri,N. and Mizuno.T. (1993) Differentiation of functional hepatocytesand biliary epithelial cells from immature hepatocytes of the fetal mousein vitro. Anal. EmbryoL, 187, 221-229.
38.Akrawi,M., Rogiers.V., Vandenberghe,Y., Palmer.C.N.A., Vercruysse^A.,Shephard^.A. and Phillips,I.R. (1993) Maintenance and induction in co-cultured rat hepatocytes of components of the cytochrome P45O-mediatedmono-oxygenase. Biochem. Pharmacol, 45, 1583-1591.
39.Tong,C, Telang.S. and Williams.G.M. (1984) Differences in responses of4 adult rat-liver epithelial cell lines to a spectrum of chemical mutagens.Mutal. Res., 130,53-61.
40. Gonzalez,FJ. and Korzekwa,K.R. (1995) Cytochromes P450 expressionsystems. Annu. Rev. Pharmacol Toxicol, 35, 369-390.
41.Liu,S.Y. and Gonzalez,FJ. (1995) Role of the liver-enriched transcriptionfactor HNF-la in expression of the CYP2E1 gene. DNA Cell Biol, 14,285-293.
42.ChenJ)., Lepar.G. and Kemper3. (1994) Transcriptional regulatoryelement common to a large family of hepatic cytochrome P450 genes isa functional binding site of the orphan receptor HNF-4. J. Biol Chem.,269, 5420-5427.
43. ShawJ'.M., Weiss,M.C. and Adesnik,M. (1994) Hepatocyte nuclear factor3 is a major determinant of CYP2C6 promoter activity in hepatoma cells.Mol Pharmacol, 46, 79-87.
Received on July 25, 1995; revised on January 19, 1996; accepted on January23, 1996
1106
at Beijing U
niversity of Technology on July 15, 2011
carcin.oxfordjournals.orgD
ownloaded from





























