Embed Size (px)
Citation preview

d n a r e p a i r 7 ( 2 0 0 8 ) 1250–1261
avai lab le at www.sc iencedi rec t .com
journa l homepage: www.e lsev ier .com/ locate /dnarepai r
Radiation induction of delayed recombination inSchizosaccharomyces pombe
Jun Takedaa,1, Norio Uematsub, Satomi Shiraishi c, Megumi Toyoshimad,Tomohiro Matsumotoa,2, Ohtsura Niwae,∗
a Department of Radiation System Biology, Radiation Biology Center, Kyoto University, Yoshida Konoe,Sakyo-ku, Kyoto 606-8315, Japanb Department of Genome Dynamics, Radiation Biology Center, Kyoto University, Yoshida Konoe, Sakyo-ku, Kyoto 606-8315, Japanc Department of Physics, University of Chicago, 5640 S. Ellis Avenue, Chicago, IL 60637, USAd Department of Experimental Oncology, Research Institute for Radiation Biology and Medicine, Hiroshima University, 1-2-3 Kasumi,Minami-ku, Hiroshima 734-0037, Japane Research Center for Charged Particle Therapy, National Institute of Radiological Sciences,4-9-1 Anagawa, Inage-ku, Chiba 263-8555, Japan
a r t i c l e i n f o
Article history:
Received 30 September 2007
Received in revised form
15 February 2008
Accepted 5 April 2008
Published on line 10 June 2008
Keywords:
Ionizing radiation
Delayed recombination
Untargeted recombination
DNA damage memory
Rad22
Schizosaccharomyces pombe
a b s t r a c t
Ionizing radiation is known to induce delayed chromosome and gene mutations in the
descendants of the irradiated tissue culture cells. Molecular mechanisms of such delayed
mutations are yet to be elucidated, since high genomic complexity of mammalian cells
makes it difficult to analyze. We now tested radiation induction of delayed recombination
in the fission yeast Schizosaccharomyces pombe by monitoring the frequency of homologous
recombination after X-irradiation. A reporter with 200 bp tandem repeats went through
spontaneous recombination at a frequency of 1.0 × 10−4, and the frequency increased dose-
dependently to around 10 × 10−4 at 500 Gy of X-irradiation. Although the repair of initial
DNA damage was thought to be completed before the restart of cell division cycle, the ele-
vation of the recombination frequency persisted for 8–10 cell generations after irradiation
(delayed recombination). The delayed recombination suggests that descendants of the irra-
diated cells keep a memory of the initial DNA damage which upregulates recombination
machinery for 8–10 generations even in the absence of DNA double-strand breaks (DSBs).
Since radical scavengers were ineffective in inhibiting the delayed recombination, a memory
by continuous production of DNA damaging agents such as reactive oxygen species (ROS)
was excluded. Recombination was induced in trans in a reporter on chromosome III by a
DNA DSB at a site on chromosome I, suggesting the untargeted nature of delayed recombi-
nation. Interestingly, Rad22 foci persisted in the X-irradiated population in parallel with the
elevation of the recombination frequency. These results suggest that the epigenetic dam-
age memory induced by D
pombe.
∗ Corresponding author. Tel.: +81 43 206 3482; fax: +81 43 206 4531.E-mail addresses: [email protected] (T. Matsumot
1 Present address: Laboratory of Biochemistry, Department of Chemistan University, 1-1 Minami-Osawa, Hachioji-shi, Tokyo 192-0397, Japan
2 Contact for the experimental materials in this article. Tel.: +81 75 71568-7864/$ – see front matter © 2008 Elsevier B.V. All rights reserved.doi:10.1016/j.dnarep.2008.04.006
NA DSB upregulates untargeted and delayed recombination in S.
© 2008 Elsevier B.V. All rights reserved.
o), [email protected] (O. Niwa).try, Graduate School of Science and Engineering, Tokyo Metropoli-.53 7553; fax: +81 75 753 7564.

0 0 8
1
Iwtocyot
hnietgr
itswiRwtlsmaecupidrfsfc
tsecd
etnrndodsr
d n a r e p a i r 7 ( 2
. Introduction
onizing radiations induce DNA double-strand breaks (DSBs),hich are potentially lethal to cells if not repaired [1]. In order
o avoid such a detriment, cells are endowed with a varietyf DNA damage responses such as DNA repair and cell cycleheckpoints. Even a single DSB can activate these responses ineasts and in mammalian cells [2–4]. Consequently, activationf damage responses takes place immediately after introduc-ion of DSB [5,6].
DSBs are repaired mainly by two mechanisms: non-omologous endjoining (NHEJ) and homologous recombi-ation (HR) [7]. NHEJ is error prone in theory because of
ncapability of recovering correct information at some DSBnds. In contrast, HR is supposed to be error free, becausehe sequence information can be retrieved from a homolo-ous strand, although HR at tandem repeats may frequentlyesult in gain or loss of repeat units [8,9].
HR is initiated by DSB; hence the DSB is a cis-actingnducer of HR. In addition, DSB is known to induce HR inrans [10–12]. Fabre and Roman were the first to demon-trate the trans effect of DSB on HR in Saccharomyces cerevisiaehere the introduction of radiation-damaged chromosomes
nduced HR in undamaged chromosomes of the host cells [10].ecently, the trans-activation of HR was also found in murinehole body systems. Length change mutations were reported
o occur at the maternally derived allele of a minisatelliteocus in F1 mice born to X-irradiated spermatozoa [11]. Theame group reported another study in which the rate of HR-ediated reversion was upregulated at the maternally derived
llele of the pink-eyed unstable (pun) locus in retinal pigmentpithelium of sperm-irradiated F1 mice [12]. These reportslearly demonstrate the trans effect in which intact allelendergoes HR when DNA damage is present in a same cyto-lasmic and/or nucleoplasmic environment. The trans effect
n mice is likely to be mediated by p53, because the newlyiscovered p53-dependent S-phase checkpoint decreased theate of replication fork progression in trans in undamagedemale pronuclei when zygotes were fertilized by irradiatedperms [13,14]. This p53-dependent slowdown of replicationork progression was proposed to facilitate HR between sisterhromatids, or sister chromatid exchange [15].
The pun experiment [12] is of particular interest, becausehe introduction of DNA damage into zygotes by X-irradiatedpermatozoa and the elevation of HR in the retinal pigmentpithelium on days 11–12 are separated by many cell divisionycles. This time gap indicates a long-lasting memory of DNAamage in addition to the trans-acting HR induction.
For its complexity, the murine whole body system, how-ver, is not ideal in analyzing molecular mechanisms of therans effect and the memory of DNA damage. Therefore, weow used the fission yeast Schizosaccharomyces pombe to studyadiation induction of trans-acting HR and delayed recombi-ation. We report here that HR was upregulated for 8–10 cellivision cycles (generations) after X-irradiation in the absence
f remaining and/or newly arising DSBs. This duration, 8–10ivisions, was independent of X-ray dose. We also demon-trate that a single DSB on a chromosome induced HR at aeporter on another chromosome in trans. Focus formation of) 1250–1261 1251
Rad22, a homologue of S. cerevisiae and vertebrate Rad52 [16],also perpetuated for about 10 generations after X-irradiation.Altogether, this is the first report to demonstrate the delayedand trans-acting HR operating downstream of the DNA damagememory in S. pombe.
2. Materials and methods
2.1. Yeast strains and media
Yeast strains used in this study were 972 (h−), IR101 (h−, leu1-32ura4::RDUX100(+)), IR208 (h−, leu1-32 ura4::RDUX200(+)), IR302(h−, leu1-32 ura4::RDUX300(+)), RDK21 (h+, ura4::RDUX200(+)),RDK22 (h+, ura4::RDUX200(−)), RDK76 (h−, ura4::Pnmt81-HOleu1-32 his3::RDUX200(+) ade7ins::pLEU2-ade7TR), RDK78 (h−,ura4::Pnmt81-HO leu1-32 his3::RDUX200(+) ade7ins::pLEU2-ade7TR-HOcs), RDK88 (h−, smt-0 ura4::RDUX200(+)lys3::hph leu1-32 his3-D1 pREP81X-HO), RDK89 (h−, smt-0ura4::RDUX200(+) lys3::HOcs-hph leu1-32 his3-D1 pREP81X-HO)and RDK90 (h−, leu1-32 ura4::RDUX200(+) rad22-Venus). Allstrains except for 972 were constructed in this study by thestandard procedures [17] with several modifications in theprotocols of PCR and transformation of yeast cells [18,19].Sequences of DNA primers used for strain construction werelisted in Supplementary Table S1. Edinburgh minimal mediawith supplements (EMMS) were used for liquid culturesand tester plates. EMMS with 500 mg/l of G-418 was usuallyused for liquid culture of G-418 resistant strains. In theexperiments to introduce a DSB by HO endonuclease (HO),EMM conditioned by culture of the wild-type (WT) strain 972was added at a final concentration of 25% to fresh EMMSfor the preparation of tester plates. Plates with yeast extract(YE) media were used as rich media for routine passage ofcells and also used as low adenine media to distinguish ade7mutants from ade7+ revertants. Liquid culture was performedat 30 ◦C, plates were incubated at 32 ◦C, and experiments inthe present study were repeated at least three times unlessotherwise stated.
2.2. X-irradiation
S. pombe cells at a concentration of 107 to 109 cells/ml weretransferred in 0.6 ml microtubes and X-irradiated at a dose rateof 32.6 Gy/min (250 kV, 15 mA, without filter; Rigaku RadioflexX-ray Generator, Rigaku-denki, Japan). This condition permit-ted low energy components of X-rays to reach the samples.
2.3. Analysis of colony survival and frequency of HR
Three reporter strains each of which carried a recombinationreporter cassette ura4::kanMX6 (RDUX) were constructed. Thecassette had a duplication of a central portion of the ura4+
gene with the length of either 100, 200 or 300 bp bracketingthe neomycin-resistance module kanMX6 [20]. These cassettesintegrated into the endogenous ura4+ locus were referred to as
RDUX100(+), RDUX200(+) and RDUX300(+), named after theirrepeat length and orientation on the genome, and the corre-sponding reporter strains were designated as IR101, IR208 andIR302, respectively. Upon HR of the duplication, the RDUX con-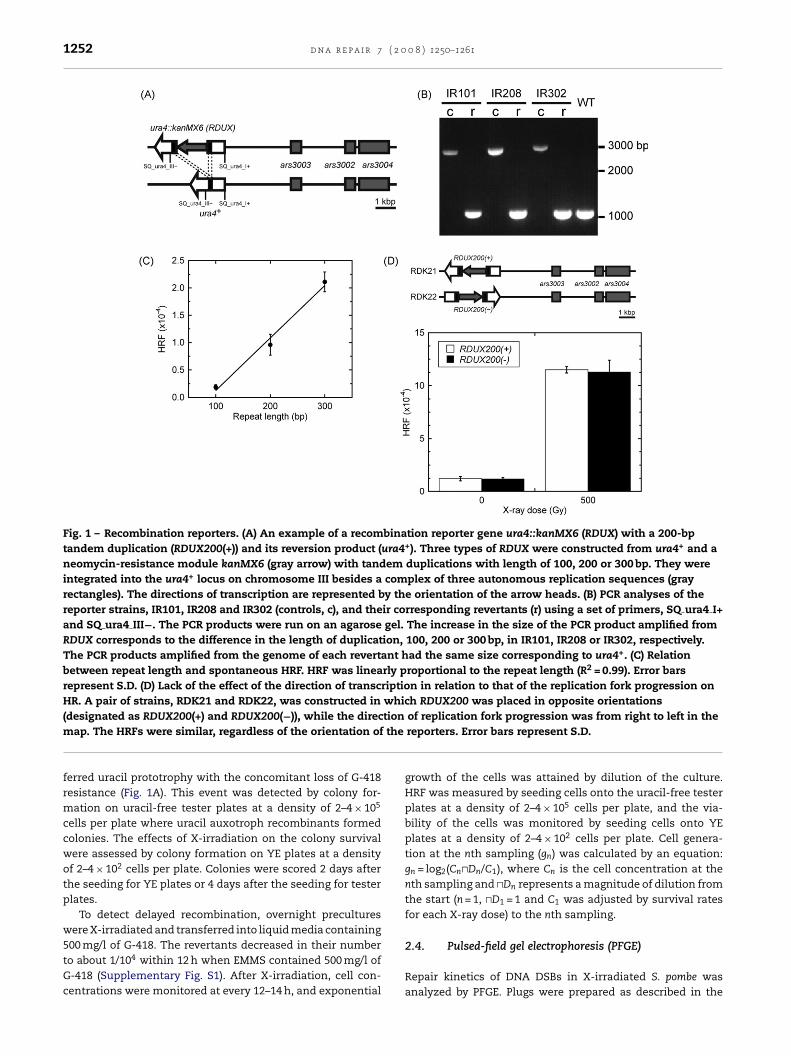
1252 d n a r e p a i r 7 ( 2 0 0 8 ) 1250–1261
Fig. 1 – Recombination reporters. (A) An example of a recombination reporter gene ura4::kanMX6 (RDUX) with a 200-bptandem duplication (RDUX200(+)) and its reversion product (ura4+). Three types of RDUX were constructed from ura4+ and aneomycin-resistance module kanMX6 (gray arrow) with tandem duplications with length of 100, 200 or 300 bp. They wereintegrated into the ura4+ locus on chromosome III besides a complex of three autonomous replication sequences (grayrectangles). The directions of transcription are represented by the orientation of the arrow heads. (B) PCR analyses of thereporter strains, IR101, IR208 and IR302 (controls, c), and their corresponding revertants (r) using a set of primers, SQ ura4 I+and SQ ura4 III−. The PCR products were run on an agarose gel. The increase in the size of the PCR product amplified fromRDUX corresponds to the difference in the length of duplication, 100, 200 or 300 bp, in IR101, IR208 or IR302, respectively.The PCR products amplified from the genome of each revertant had the same size corresponding to ura4+. (C) Relationbetween repeat length and spontaneous HRF. HRF was linearly proportional to the repeat length (R2 = 0.99). Error barsrepresent S.D. (D) Lack of the effect of the direction of transcription in relation to that of the replication fork progression onHR. A pair of strains, RDK21 and RDK22, was constructed in which RDUX200 was placed in opposite orientations
tionthe
(designated as RDUX200(+) and RDUX200(−)), while the direcmap. The HRFs were similar, regardless of the orientation of
ferred uracil prototrophy with the concomitant loss of G-418resistance (Fig. 1A). This event was detected by colony for-mation on uracil-free tester plates at a density of 2–4 × 105
cells per plate where uracil auxotroph recombinants formedcolonies. The effects of X-irradiation on the colony survivalwere assessed by colony formation on YE plates at a densityof 2–4 × 102 cells per plate. Colonies were scored 2 days afterthe seeding for YE plates or 4 days after the seeding for testerplates.
To detect delayed recombination, overnight precultureswere X-irradiated and transferred into liquid media containing
500 mg/l of G-418. The revertants decreased in their numberto about 1/104 within 12 h when EMMS contained 500 mg/l ofG-418 (Supplementary Fig. S1). After X-irradiation, cell con-centrations were monitored at every 12–14 h, and exponentialof replication fork progression was from right to left in thereporters. Error bars represent S.D.
growth of the cells was attained by dilution of the culture.HRF was measured by seeding cells onto the uracil-free testerplates at a density of 2–4 × 105 cells per plate, and the via-bility of the cells was monitored by seeding cells onto YEplates at a density of 2–4 × 102 cells per plate. Cell genera-tion at the nth sampling (gn) was calculated by an equation:gn = log2(Cn�Dn/C1), where Cn is the cell concentration at thenth sampling and �Dn represents a magnitude of dilution fromthe start (n = 1, �D1 = 1 and C1 was adjusted by survival ratesfor each X-ray dose) to the nth sampling.
2.4. Pulsed-field gel electrophoresis (PFGE)
Repair kinetics of DNA DSBs in X-irradiated S. pombe wasanalyzed by PFGE. Plugs were prepared as described in the

0 0 8
mRw(d1iapw0wbass
2(
D[ctaaeclw1
2
AwTrsuwgpwac1(
ntwafvs(apw
in two opposite orientations (Fig. 1D, upper diagram). Both
d n a r e p a i r 7 ( 2
anufacture’s instruction (CHEF Genomic DNA Plug Kits, Bio-ad Laboratories, Inc., USA) with the modifications below. Cellall digestion was performed by 1 mg/ml zymolyase 100T
Seikagaku Corporation, Japan) in spheroplast buffer (1.0 M-sorbitol, 100 mM EDTA, pH 7.5, 30 mM 2-mercaptoethanol,0 mM sodium azide) at 37 ◦C for 90 min before suspend-ng cells in Cell Suspension Buffer. After solidification, 0.5%garose plugs were incubated with SDS buffer (0.25 M EDTA,H 8.0, 50 mM Tris–HCl, pH 7.6, 1% SDS) at 55 ◦C for 90 min,ashed twice with TE buffer warmed at 37 ◦C, and treated with
.5 mg/ml proteinase K at 50 ◦C for 24 h. PFGE was carried outith 0.8% chromosomal grade agarose (Bio-Rad) and 1× TAEuffer for 48 h at 14 ◦C using CHEF-DR III System (Bio-Rad) atn angle of 106◦ with the voltage of 2 V/cm and 30 min of awitching time. Images were obtained after ethidium bromidetaining of the gels.
.5. Detection of intracellular reactive oxygen speciesROS) and the effect of ROS scavengers
ihydrorhodamin 123 (DHR) was used to detect cellular ROS21]. DHR at a final concentration of 2.5 �M was added to yeastulture in 1.5–2 h before analysis. The fluorescence was quan-ified by a flowcytometer (BD FACSCalibur, Becton, Dickinsonnd Company, USA) using FL-1 filter. Ascorbate (vitamin C)nd Trolox (a derivative of vitamin E) have been used to scav-nge ROS in yeast cells [21,22]. Ascorbate scavenges ROS in theytosol, while Trolox inhibits membrane peroxidation. Sodium-ascorbate was used at a final concentration of 10 mM. Troloxas dissolved in ethanol and used at a final concentration ofmM.
.6. Induction of DSB by HO
part of the Y˛-Z region in the MAT locus of S. cerevisiaeas used as the target sequence of HO (5′-CACGGAATA-GGGACTACTTCG-CGCAACAGTATAATTTTATAAA-3′) andeferred to as HOcs (HO cutting site) [23]. The HO gene wasupplied by the plasmid pREP81X-HO [8] or by a targeted allelera4::Pnmt81-HO-Tnmt1 (ura4::Pnmt81-HO) in which the ura4+ alleleas completely replaced with the nmt81 promoter-driven HO
ene. Depletion of thiamine from the culture derepresses theromoter. For HO induction, overnight precultures in EMMSith 150 �M of thiamine were washed three times with water
nd then cultured in EMMS devoid of thiamine with mediumhanges at every 8–12 h. Production of HO peaked at around5 h after induction and HRF increased linearly thereafterSupplementary Fig. S3).
For the analysis of the cis effect of DSB, a pair of recombi-ation reporters was constructed. Plasmid pLEU2-ade7TR andhe same construct except with HOcs, pLEU2-ade7TR-HOcs,ere introduced into ade7+ locus [24] to disrupt the gene by500-bp tandem duplication derived from the open reading
rame (ORF) of the ade7+ gene using the LEU2 gene of S. cere-isiae as a selectable marker (Fig. 4A, upper diagram). Thetrains with the duplication, RDK76 (without HOcs) and RDK78
with HOcs), were unable to grow on adenine deficient platesnd formed red colonies when they grew on low adeninelates, while recombinants without the duplication formedhite colonies [25].) 1250–1261 1253
For the analysis of the trans effect of a DSB, a pair of strains,RDK88 (without HOcs) and RDK89 (with HOcs), were con-structed. They carried RDUX200(+) on chromosome III togetherwith or without HOcs at the lys3 locus on chromosome I(Fig. 4B, upper diagram). To measure HRF, RDK88 and RDK89cells were plated on uracil-free tester plates and on YE platesas described above at 20 h after HO induction. YE plates wereincubated for 2 days, and the tester plates were incubated for5–6 days.
2.7. Analysis of Rad22 focus formation
First, a plasmid pVUv-C was constructed, which carries ura4+
as a removable selectable marker together with a tandemduplication of an EYFP variant Venus (RIKEN, Japan) [26]. Then,a C-terminus fragment of the rad22+ ORF and a fragment ofrad22 terminator were cloned into pVUv-C to generate pKI-rad22-VUv. Finally, the NcoI-digested pKI-rad22-VUv was usedfor the targeted integration to the rad22+ locus (SupplementaryFig. S4A). Transformants were screened on uracil-depletedEMMS plates followed by secondary screening of rad22+-Venuson a YE plate containing 150 mg/l uracil and 0.1% 5-fluorooroticacid (Supplementary Fig. S4B and C). Correct integration wasconfirmed by PCR.
Before observation of Rad22-Venus, cells were cooled onice. Photos of the cells were taken under a fluorescent micro-scope (Leica DM5500B, Leica Microsystems GmbH, Germany)to score the frequency of cells with focus. Cell length wasmeasured from the photos computationally with image-processing software (IPLab for Windows 4.04, BD Biosciences,USA).
3. Results
3.1. Recombination in reporter strains
PCR analysis was conducted on the three reporter strains(IR101, IR208 and IR302) and on their corresponding uracil-prototrophic revertants (Fig. 1B), using a primer set, SQ ura4 I+and SQ ura4 III− (Fig. 1A and Supplementary Table S1). Thelength of each pair of PCR products matched with the recombi-nation reporters with disrupted ura4 (RDUX100(+), RDUX200(+)and RDUX300(+)) and with the reverted WT ura4+. The spon-taneous HRF was linearly proportional to the length of eachrepeat (Fig. 1C). Subsequent experiments were done withthe reporter RDUX200 unless otherwise indicated, since thisreporter enabled us to detect a wider range of increase in HRFby X-irradiation than the other two.
A complex of three strong replication origins, ars3002,ars3003 and ars3004, is located near the ura4+ locus (Fig. 1Aand D) [27]. The direction of replication fork progression inrelation to that of transcription of a gene may affect HRF.Therefore, we constructed a pair of strains, RDK21 and RDK22,in which the recombination reporter RDUX200 were placed
strains were found to exhibit similar frequencies of sponta-neous and X-ray induced HR (Fig. 1D, lower diagram). Thisresult indicated that the orientation of transcription did notaffect HR.
( 2 0
1254 d n a r e p a i r 73.2. Induction of delayed recombination by X-rays
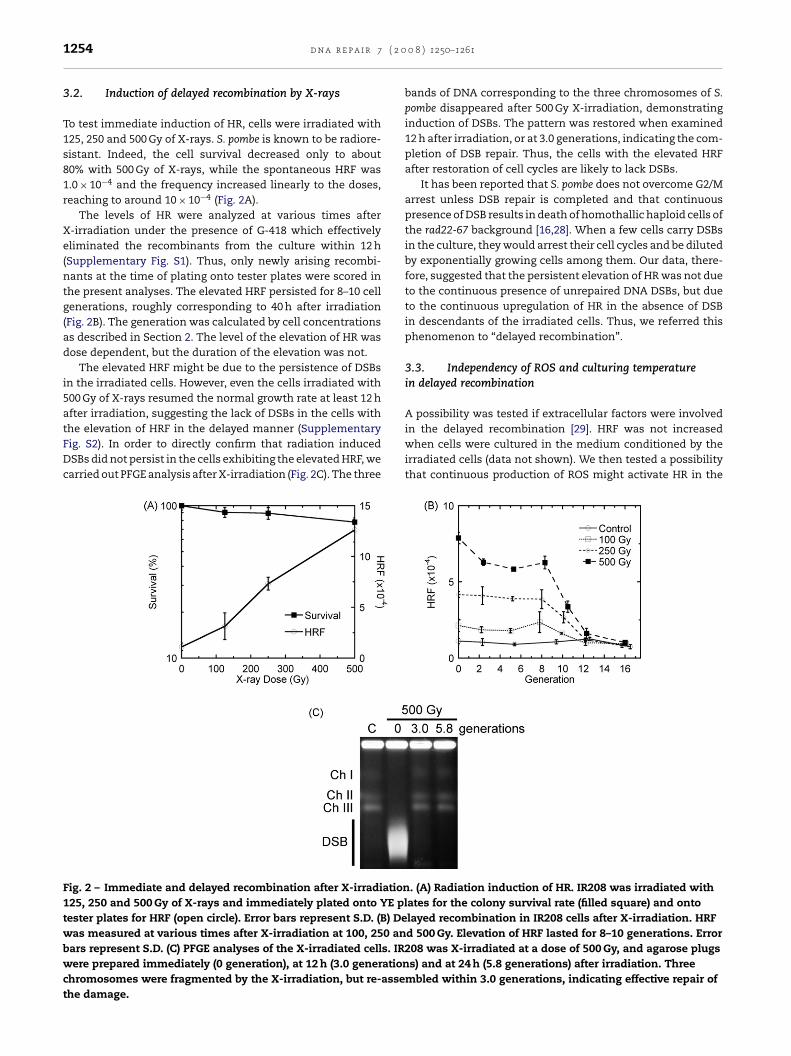
To test immediate induction of HR, cells were irradiated with125, 250 and 500 Gy of X-rays. S. pombe is known to be radiore-sistant. Indeed, the cell survival decreased only to about80% with 500 Gy of X-rays, while the spontaneous HRF was1.0 × 10−4 and the frequency increased linearly to the doses,reaching to around 10 × 10−4 (Fig. 2A).
The levels of HR were analyzed at various times afterX-irradiation under the presence of G-418 which effectivelyeliminated the recombinants from the culture within 12 h(Supplementary Fig. S1). Thus, only newly arising recombi-nants at the time of plating onto tester plates were scored inthe present analyses. The elevated HRF persisted for 8–10 cellgenerations, roughly corresponding to 40 h after irradiation(Fig. 2B). The generation was calculated by cell concentrationsas described in Section 2. The level of the elevation of HR wasdose dependent, but the duration of the elevation was not.
The elevated HRF might be due to the persistence of DSBsin the irradiated cells. However, even the cells irradiated with500 Gy of X-rays resumed the normal growth rate at least 12 hafter irradiation, suggesting the lack of DSBs in the cells with
the elevation of HRF in the delayed manner (SupplementaryFig. S2). In order to directly confirm that radiation inducedDSBs did not persist in the cells exhibiting the elevated HRF, wecarried out PFGE analysis after X-irradiation (Fig. 2C). The threeFig. 2 – Immediate and delayed recombination after X-irradiation125, 250 and 500 Gy of X-rays and immediately plated onto YE pltester plates for HRF (open circle). Error bars represent S.D. (B) Dewas measured at various times after X-irradiation at 100, 250 anbars represent S.D. (C) PFGE analyses of the X-irradiated cells. IRwere prepared immediately (0 generation), at 12 h (3.0 generationchromosomes were fragmented by the X-irradiation, but re-assethe damage.
0 8 ) 1250–1261
bands of DNA corresponding to the three chromosomes of S.pombe disappeared after 500 Gy X-irradiation, demonstratinginduction of DSBs. The pattern was restored when examined12 h after irradiation, or at 3.0 generations, indicating the com-pletion of DSB repair. Thus, the cells with the elevated HRFafter restoration of cell cycles are likely to lack DSBs.
It has been reported that S. pombe does not overcome G2/Marrest unless DSB repair is completed and that continuouspresence of DSB results in death of homothallic haploid cells ofthe rad22-67 background [16,28]. When a few cells carry DSBsin the culture, they would arrest their cell cycles and be dilutedby exponentially growing cells among them. Our data, there-fore, suggested that the persistent elevation of HR was not dueto the continuous presence of unrepaired DNA DSBs, but dueto the continuous upregulation of HR in the absence of DSBin descendants of the irradiated cells. Thus, we referred thisphenomenon to “delayed recombination”.
3.3. Independency of ROS and culturing temperaturein delayed recombination
A possibility was tested if extracellular factors were involved
in the delayed recombination [29]. HRF was not increasedwhen cells were cultured in the medium conditioned by theirradiated cells (data not shown). We then tested a possibilitythat continuous production of ROS might activate HR in the. (A) Radiation induction of HR. IR208 was irradiated withates for the colony survival rate (filled square) and ontolayed recombination in IR208 cells after X-irradiation. HRFd 500 Gy. Elevation of HRF lasted for 8–10 generations. Error208 was X-irradiated at a dose of 500 Gy, and agarose plugss) and at 24 h (5.8 generations) after irradiation. Three
mbled within 3.0 generations, indicating effective repair of
0 0 8 ) 1250–1261 1255
irXwtbcnt
tt3r(saa
3
TDroc
RgooHiwR1Retdcei
RomobT
3X
Ricm[s
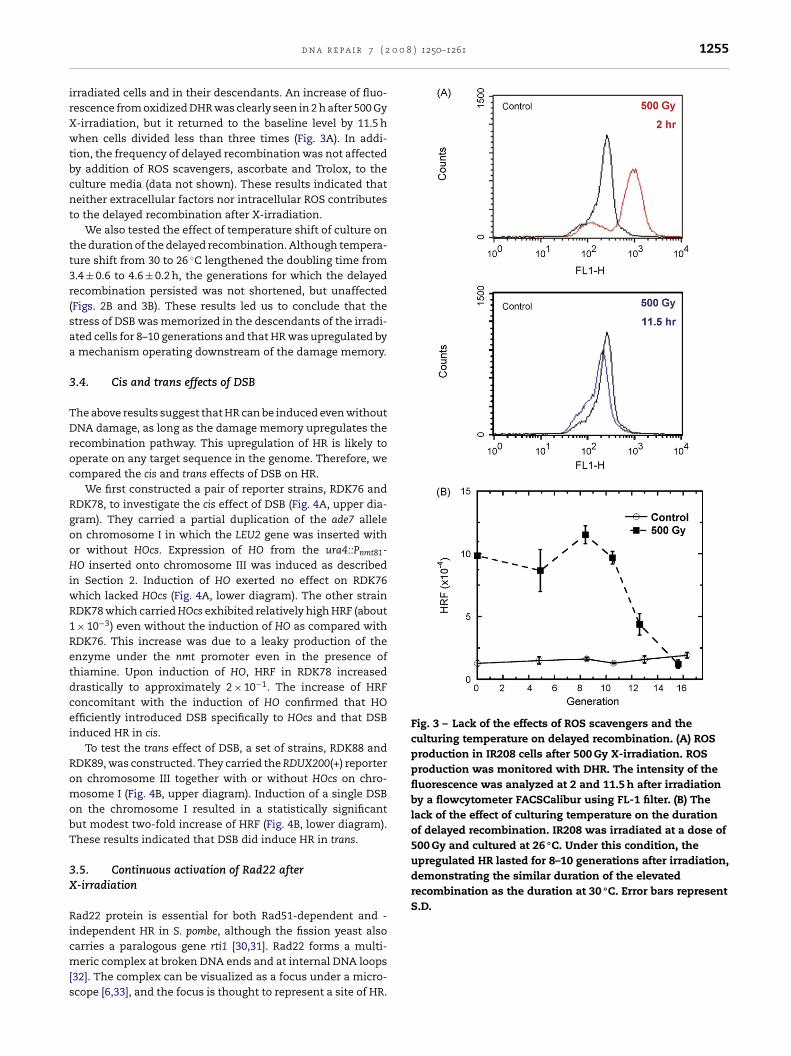
Fig. 3 – Lack of the effects of ROS scavengers and theculturing temperature on delayed recombination. (A) ROSproduction in IR208 cells after 500 Gy X-irradiation. ROSproduction was monitored with DHR. The intensity of thefluorescence was analyzed at 2 and 11.5 h after irradiationby a flowcytometer FACSCalibur using FL-1 filter. (B) Thelack of the effect of culturing temperature on the durationof delayed recombination. IR208 was irradiated at a dose of500 Gy and cultured at 26 ◦C. Under this condition, theupregulated HR lasted for 8–10 generations after irradiation,demonstrating the similar duration of the elevatedrecombination as the duration at 30 ◦C. Error bars representS.D.
d n a r e p a i r 7 ( 2
rradiated cells and in their descendants. An increase of fluo-escence from oxidized DHR was clearly seen in 2 h after 500 Gy-irradiation, but it returned to the baseline level by 11.5 hhen cells divided less than three times (Fig. 3A). In addi-
ion, the frequency of delayed recombination was not affectedy addition of ROS scavengers, ascorbate and Trolox, to theulture media (data not shown). These results indicated thateither extracellular factors nor intracellular ROS contributeso the delayed recombination after X-irradiation.
We also tested the effect of temperature shift of culture onhe duration of the delayed recombination. Although tempera-ure shift from 30 to 26 ◦C lengthened the doubling time from.4 ± 0.6 to 4.6 ± 0.2 h, the generations for which the delayedecombination persisted was not shortened, but unaffectedFigs. 2B and 3B). These results led us to conclude that thetress of DSB was memorized in the descendants of the irradi-ted cells for 8–10 generations and that HR was upregulated bymechanism operating downstream of the damage memory.
.4. Cis and trans effects of DSB
he above results suggest that HR can be induced even withoutNA damage, as long as the damage memory upregulates the
ecombination pathway. This upregulation of HR is likely toperate on any target sequence in the genome. Therefore, weompared the cis and trans effects of DSB on HR.
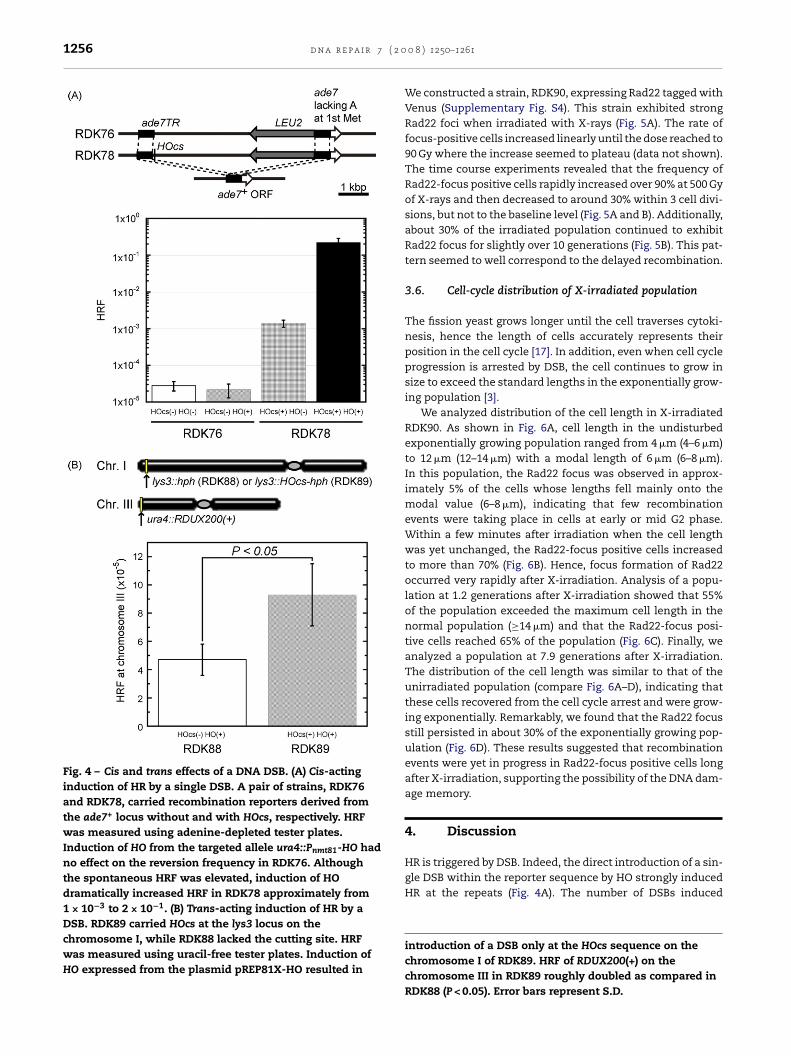
We first constructed a pair of reporter strains, RDK76 andDK78, to investigate the cis effect of DSB (Fig. 4A, upper dia-ram). They carried a partial duplication of the ade7 allelen chromosome I in which the LEU2 gene was inserted withr without HOcs. Expression of HO from the ura4::Pnmt81-O inserted onto chromosome III was induced as described
n Section 2. Induction of HO exerted no effect on RDK76hich lacked HOcs (Fig. 4A, lower diagram). The other strainDK78 which carried HOcs exhibited relatively high HRF (about× 10−3) even without the induction of HO as compared withDK76. This increase was due to a leaky production of thenzyme under the nmt promoter even in the presence ofhiamine. Upon induction of HO, HRF in RDK78 increasedrastically to approximately 2 × 10−1. The increase of HRFoncomitant with the induction of HO confirmed that HOfficiently introduced DSB specifically to HOcs and that DSBnduced HR in cis.
To test the trans effect of DSB, a set of strains, RDK88 andDK89, was constructed. They carried the RDUX200(+) reportern chromosome III together with or without HOcs on chro-osome I (Fig. 4B, upper diagram). Induction of a single DSB
n the chromosome I resulted in a statistically significantut modest two-fold increase of HRF (Fig. 4B, lower diagram).hese results indicated that DSB did induce HR in trans.
.5. Continuous activation of Rad22 after-irradiation
ad22 protein is essential for both Rad51-dependent and -ndependent HR in S. pombe, although the fission yeast also
arries a paralogous gene rti1 [30,31]. Rad22 forms a multi-eric complex at broken DNA ends and at internal DNA loops32]. The complex can be visualized as a focus under a micro-cope [6,33], and the focus is thought to represent a site of HR.
1256 d n a r e p a i r 7 ( 2 0
Fig. 4 – Cis and trans effects of a DNA DSB. (A) Cis-actinginduction of HR by a single DSB. A pair of strains, RDK76and RDK78, carried recombination reporters derived fromthe ade7+ locus without and with HOcs, respectively. HRFwas measured using adenine-depleted tester plates.Induction of HO from the targeted allele ura4::Pnmt81-HO hadno effect on the reversion frequency in RDK76. Althoughthe spontaneous HRF was elevated, induction of HOdramatically increased HRF in RDK78 approximately from1 × 10−3 to 2 × 10−1. (B) Trans-acting induction of HR by aDSB. RDK89 carried HOcs at the lys3 locus on thechromosome I, while RDK88 lacked the cutting site. HRFwas measured using uracil-free tester plates. Induction ofHO expressed from the plasmid pREP81X-HO resulted in
0 8 ) 1250–1261
We constructed a strain, RDK90, expressing Rad22 tagged withVenus (Supplementary Fig. S4). This strain exhibited strongRad22 foci when irradiated with X-rays (Fig. 5A). The rate offocus-positive cells increased linearly until the dose reached to90 Gy where the increase seemed to plateau (data not shown).The time course experiments revealed that the frequency ofRad22-focus positive cells rapidly increased over 90% at 500 Gyof X-rays and then decreased to around 30% within 3 cell divi-sions, but not to the baseline level (Fig. 5A and B). Additionally,about 30% of the irradiated population continued to exhibitRad22 focus for slightly over 10 generations (Fig. 5B). This pat-tern seemed to well correspond to the delayed recombination.
3.6. Cell-cycle distribution of X-irradiated population
The fission yeast grows longer until the cell traverses cytoki-nesis, hence the length of cells accurately represents theirposition in the cell cycle [17]. In addition, even when cell cycleprogression is arrested by DSB, the cell continues to grow insize to exceed the standard lengths in the exponentially grow-ing population [3].
We analyzed distribution of the cell length in X-irradiatedRDK90. As shown in Fig. 6A, cell length in the undisturbedexponentially growing population ranged from 4 �m (4–6 �m)to 12 �m (12–14 �m) with a modal length of 6 �m (6–8 �m).In this population, the Rad22 focus was observed in approx-imately 5% of the cells whose lengths fell mainly onto themodal value (6–8 �m), indicating that few recombinationevents were taking place in cells at early or mid G2 phase.Within a few minutes after irradiation when the cell lengthwas yet unchanged, the Rad22-focus positive cells increasedto more than 70% (Fig. 6B). Hence, focus formation of Rad22occurred very rapidly after X-irradiation. Analysis of a popu-lation at 1.2 generations after X-irradiation showed that 55%of the population exceeded the maximum cell length in thenormal population (≥14 �m) and that the Rad22-focus posi-tive cells reached 65% of the population (Fig. 6C). Finally, weanalyzed a population at 7.9 generations after X-irradiation.The distribution of the cell length was similar to that of theunirradiated population (compare Fig. 6A–D), indicating thatthese cells recovered from the cell cycle arrest and were grow-ing exponentially. Remarkably, we found that the Rad22 focusstill persisted in about 30% of the exponentially growing pop-ulation (Fig. 6D). These results suggested that recombinationevents were yet in progress in Rad22-focus positive cells longafter X-irradiation, supporting the possibility of the DNA dam-age memory.
4. Discussion
HR is triggered by DSB. Indeed, the direct introduction of a sin-gle DSB within the reporter sequence by HO strongly inducedHR at the repeats (Fig. 4A). The number of DSBs induced
introduction of a DSB only at the HOcs sequence on thechromosome I of RDK89. HRF of RDUX200(+) on thechromosome III in RDK89 roughly doubled as compared inRDK88 (P < 0.05). Error bars represent S.D.
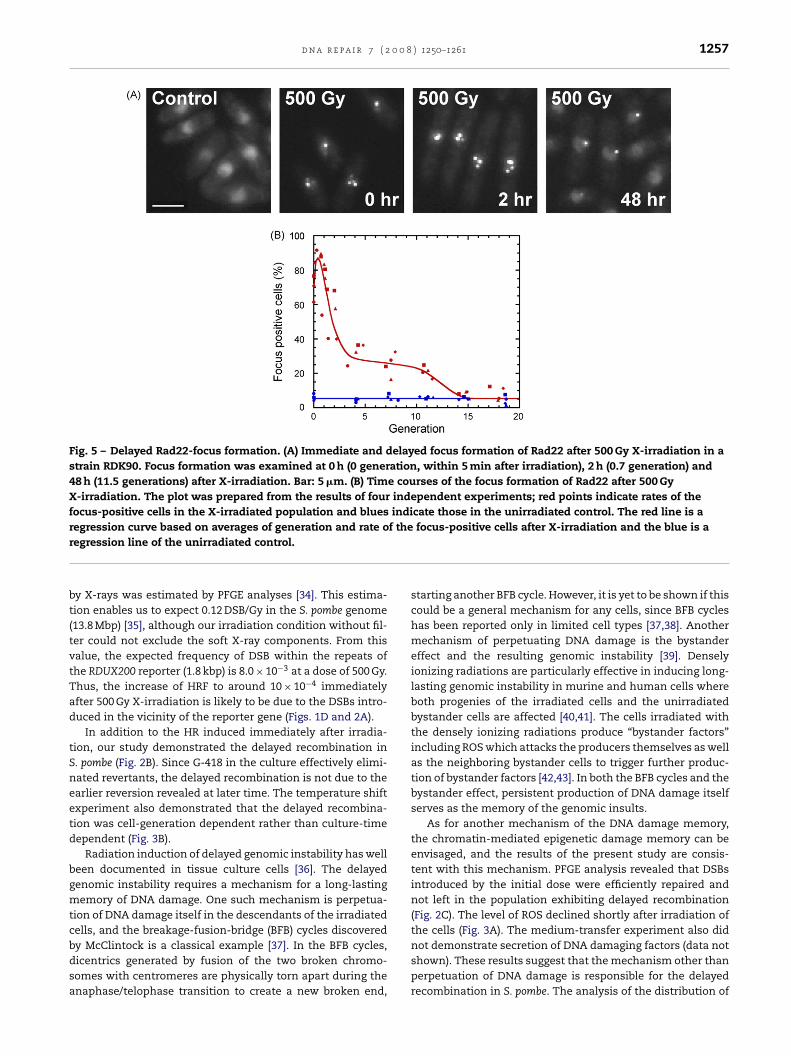
d n a r e p a i r 7 ( 2 0 0 8 ) 1250–1261 1257
Fig. 5 – Delayed Rad22-focus formation. (A) Immediate and delayed focus formation of Rad22 after 500 Gy X-irradiation in astrain RDK90. Focus formation was examined at 0 h (0 generation, within 5 min after irradiation), 2 h (0.7 generation) and48 h (11.5 generations) after X-irradiation. Bar: 5 �m. (B) Time courses of the focus formation of Rad22 after 500 GyX-irradiation. The plot was prepared from the results of four independent experiments; red points indicate rates of thefocus-positive cells in the X-irradiated population and blues indicate those in the unirradiated control. The red line is aregression curve based on averages of generation and rate of the focus-positive cells after X-irradiation and the blue is ar
bt(tvtTad
tSneetd
bgmtcbdsa
egression line of the unirradiated control.
y X-rays was estimated by PFGE analyses [34]. This estima-ion enables us to expect 0.12 DSB/Gy in the S. pombe genome13.8 Mbp) [35], although our irradiation condition without fil-er could not exclude the soft X-ray components. From thisalue, the expected frequency of DSB within the repeats ofhe RDUX200 reporter (1.8 kbp) is 8.0 × 10−3 at a dose of 500 Gy.hus, the increase of HRF to around 10 × 10−4 immediatelyfter 500 Gy X-irradiation is likely to be due to the DSBs intro-uced in the vicinity of the reporter gene (Figs. 1D and 2A).
In addition to the HR induced immediately after irradia-ion, our study demonstrated the delayed recombination in. pombe (Fig. 2B). Since G-418 in the culture effectively elimi-ated revertants, the delayed recombination is not due to thearlier reversion revealed at later time. The temperature shiftxperiment also demonstrated that the delayed recombina-ion was cell-generation dependent rather than culture-timeependent (Fig. 3B).
Radiation induction of delayed genomic instability has welleen documented in tissue culture cells [36]. The delayedenomic instability requires a mechanism for a long-lastingemory of DNA damage. One such mechanism is perpetua-
ion of DNA damage itself in the descendants of the irradiatedells, and the breakage-fusion-bridge (BFB) cycles discovered
y McClintock is a classical example [37]. In the BFB cycles,icentrics generated by fusion of the two broken chromo-omes with centromeres are physically torn apart during thenaphase/telophase transition to create a new broken end,starting another BFB cycle. However, it is yet to be shown if thiscould be a general mechanism for any cells, since BFB cycleshas been reported only in limited cell types [37,38]. Anothermechanism of perpetuating DNA damage is the bystandereffect and the resulting genomic instability [39]. Denselyionizing radiations are particularly effective in inducing long-lasting genomic instability in murine and human cells whereboth progenies of the irradiated cells and the unirradiatedbystander cells are affected [40,41]. The cells irradiated withthe densely ionizing radiations produce “bystander factors”including ROS which attacks the producers themselves as wellas the neighboring bystander cells to trigger further produc-tion of bystander factors [42,43]. In both the BFB cycles and thebystander effect, persistent production of DNA damage itselfserves as the memory of the genomic insults.
As for another mechanism of the DNA damage memory,the chromatin-mediated epigenetic damage memory can beenvisaged, and the results of the present study are consis-tent with this mechanism. PFGE analysis revealed that DSBsintroduced by the initial dose were efficiently repaired andnot left in the population exhibiting delayed recombination(Fig. 2C). The level of ROS declined shortly after irradiation ofthe cells (Fig. 3A). The medium-transfer experiment also did
not demonstrate secretion of DNA damaging factors (data notshown). These results suggest that the mechanism other thanperpetuation of DNA damage is responsible for the delayedrecombination in S. pombe. The analysis of the distribution of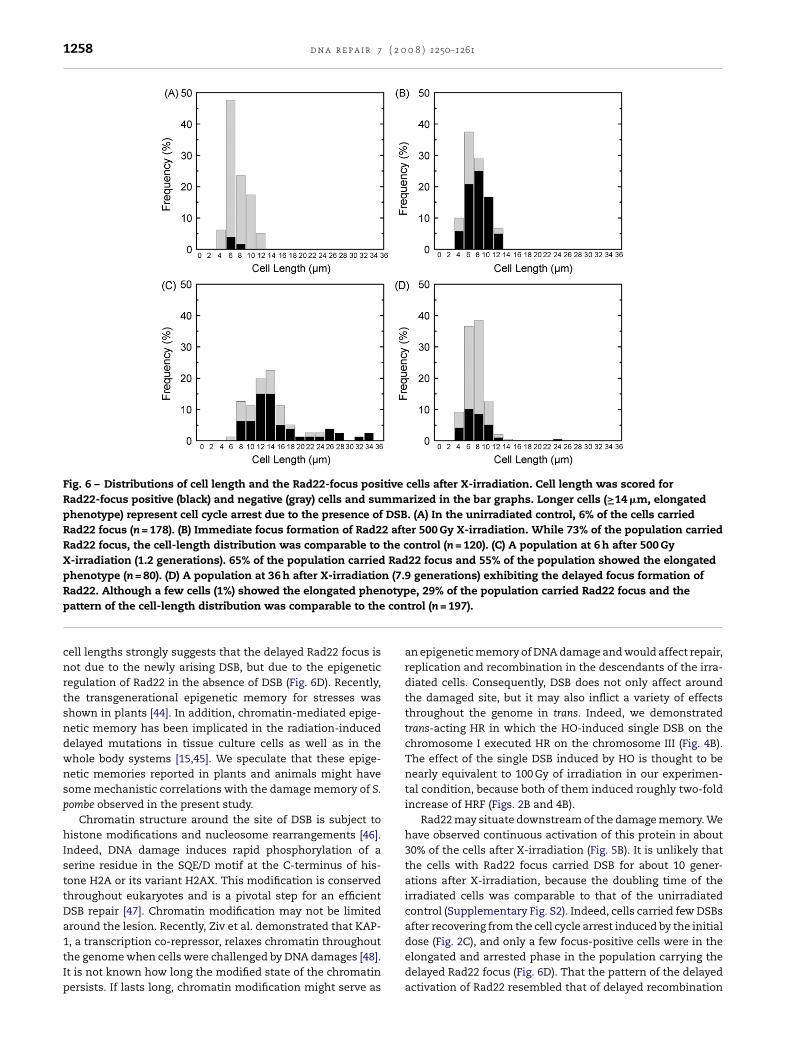
1258 d n a r e p a i r 7 ( 2 0 0 8 ) 1250–1261
Fig. 6 – Distributions of cell length and the Rad22-focus positive cells after X-irradiation. Cell length was scored forRad22-focus positive (black) and negative (gray) cells and summarized in the bar graphs. Longer cells (≥14 �m, elongatedphenotype) represent cell cycle arrest due to the presence of DSB. (A) In the unirradiated control, 6% of the cells carriedRad22 focus (n = 178). (B) Immediate focus formation of Rad22 after 500 Gy X-irradiation. While 73% of the population carriedRad22 focus, the cell-length distribution was comparable to the control (n = 120). (C) A population at 6 h after 500 GyX-irradiation (1.2 generations). 65% of the population carried Rad22 focus and 55% of the population showed the elongatedphenotype (n = 80). (D) A population at 36 h after X-irradiation (7.9 generations) exhibiting the delayed focus formation of
otypcon
Rad22. Although a few cells (1%) showed the elongated phenpattern of the cell-length distribution was comparable to the
cell lengths strongly suggests that the delayed Rad22 focus isnot due to the newly arising DSB, but due to the epigeneticregulation of Rad22 in the absence of DSB (Fig. 6D). Recently,the transgenerational epigenetic memory for stresses wasshown in plants [44]. In addition, chromatin-mediated epige-netic memory has been implicated in the radiation-induceddelayed mutations in tissue culture cells as well as in thewhole body systems [15,45]. We speculate that these epige-netic memories reported in plants and animals might havesome mechanistic correlations with the damage memory of S.pombe observed in the present study.
Chromatin structure around the site of DSB is subject tohistone modifications and nucleosome rearrangements [46].Indeed, DNA damage induces rapid phosphorylation of aserine residue in the SQE/D motif at the C-terminus of his-tone H2A or its variant H2AX. This modification is conservedthroughout eukaryotes and is a pivotal step for an efficientDSB repair [47]. Chromatin modification may not be limitedaround the lesion. Recently, Ziv et al. demonstrated that KAP-
1, a transcription co-repressor, relaxes chromatin throughoutthe genome when cells were challenged by DNA damages [48].It is not known how long the modified state of the chromatinpersists. If lasts long, chromatin modification might serve ase, 29% of the population carried Rad22 focus and thetrol (n = 197).
an epigenetic memory of DNA damage and would affect repair,replication and recombination in the descendants of the irra-diated cells. Consequently, DSB does not only affect aroundthe damaged site, but it may also inflict a variety of effectsthroughout the genome in trans. Indeed, we demonstratedtrans-acting HR in which the HO-induced single DSB on thechromosome I executed HR on the chromosome III (Fig. 4B).The effect of the single DSB induced by HO is thought to benearly equivalent to 100 Gy of irradiation in our experimen-tal condition, because both of them induced roughly two-foldincrease of HRF (Figs. 2B and 4B).
Rad22 may situate downstream of the damage memory. Wehave observed continuous activation of this protein in about30% of the cells after X-irradiation (Fig. 5B). It is unlikely thatthe cells with Rad22 focus carried DSB for about 10 gener-ations after X-irradiation, because the doubling time of theirradiated cells was comparable to that of the unirradiatedcontrol (Supplementary Fig. S2). Indeed, cells carried few DSBsafter recovering from the cell cycle arrest induced by the initial
dose (Fig. 2C), and only a few focus-positive cells were in theelongated and arrested phase in the population carrying thedelayed Rad22 focus (Fig. 6D). That the pattern of the delayedactivation of Rad22 resembled that of delayed recombination
0 0 8
sF
oHtplgu(otaanD[tad
mdeFmm
C
N
A
WPTTtMwtbfSD
A
Si
r
d n a r e p a i r 7 ( 2
uggests a possible mechanistic correlation of them (compareigs. 2B, 3B and 5B).
If there operates the chromatin-mediated damage mem-ry, it may well affect a mode of the global transcription.iCEP (High Coverage Expression Profiling) analysis is prac-
ically suited for studying subtle changes in the expressionrofiles of the irradiated population [49]. We identified that at
east 43 transcripts upregulated their expression levels for 5–6enerations in addition to over 100 transcripts which upreg-lated the expressions for 13 generations after X-irradiation
Takeda et al., in preparation). These long-lasting changesf the expression profiles strongly support the existence ofhe damage memory which is possibly chromatin-mediatednd may affect transcription and recombination in almostll the descendants of the X-irradiated cells. Whereas Bren-an and Schiestl demonstrated that ionizing radiation andNA damaging agents induced unstable clones in S. cerevisiae
50], we postulate from our results that delayed recombina-ion in S. pombe is a population-wise phenomenon occurrings genome-wide epigenetic changes brought about by DNAamage.
In summary, our data demonstrate that S. pombe has aechanism to memorize genomic insults. The memory is evi-
enced by the delayed recombination, prolonged changes ofxpression profiles and the delayed focus formation of Rad22.urther analyses are underway to elucidate the molecularechanisms of the delayed recombination and the damageemory.
onflict of interest
one.
cknowledgements
e thank Dr. Fikret Osman for the plasmid REP81X-HO androf. Antony M. Carr for providing important information.hanks are also to Mr. and Drs. Kohei Dohke, Tatsuki Kunoh,oshiyuki Habu, Ryoichi Hirayama and Yota Murakami forheir helps in the analysis of repair kinetics. We also thankr. Hideki Nagai for his excellent technical help. This workas supported by a grant-in-aid from the Ministry of Educa-
ion, Culture, Sports, Science and Technology of Japan, andy a grant from Nuclear Safety Research Association (NSRA)or ON. JT was supported by a research fellowship of the Japanociety for the Promotion of Science for Young Scientists (JSPS,C1).
ppendix A. Supplementary data
upplementary data associated with this article can be found,n the online version, at doi:10.1016/j.dnarep.2008.04.006.
e f e r e n c e s
[1] M. Frankenberg-Schwager, D. Frankenberg, DNAdouble-strand breaks: their repair and relationship to cell
) 1250–1261 1259
killing in yeast, Int. J. Radiat. Biol. 58 (1990)569–575.
[2] M. Lisby, U.H. Mortensen, R. Rothstein, Colocalization ofmultiple DNA double-strand breaks at a single Rad52 repaircentre, Nat. Cell Biol. 5 (2003) 572–577.
[3] L.-L. Du, T.M. Nakamura, B.A. Moser, P. Russell, Retention butnot recruitment of Crb2 at double-strand breaks requiresRad1 and Rad3 complexes, Mol. Cell. Biol. 23 (2003)6150–6158.
[4] L.-C. Huang, K.C. Clarkin, G.M. Wahl, Sensitivity andselectivity of the DNA damage sensor responsible foractivating p53-dependent G1 arrest, Proc. Natl. Acad. Sci.U.S.A. 93 (1996) 4827–4832.
[5] T.A. Kato, H. Nagasawa, M.M. Weil, J.B. Little, J.S. Bedford,Levels of �-H2AX foci after low-dose-rate irradiation reveal aDNA DSB rejoining defect in cells from human ATMheterozygotes in two at families and in another apparentlynormal individual, Radiat. Res. 166 (2006)443–453.
[6] T.M. Nakamura, L.-L. Du, C. Redon, P. Russell, Histone H2Aphosphorylation controls Crb2 recruitment at DNA breaks,maintains checkpoint arrest, and influences DNA repair infission yeast, Mol. Cell. Biol. 24 (2004) 6215–6230.
[7] T. Helleday, J. Lo, D.C. van Gent, B.P. Engelward, DNAdouble-strand break repair: from mechanistic understandingto cancer treatment, DNA Repair 6 (2007) 923–935.
[8] F. Osman, E.A. Fortunato, S. Subramani, Double-strandbreak-induced mitotic intrachromosomal recombination inthe fission yeast Schizosaccharomyces pombe, Genetics 142(1996) 341–357.
[9] F. Paques, J.E. Haber, Multiple pathways of recombinationinduced by double-strand breaks in Saccharomyces cerevisiae,Microbiol. Mol. Biol. Rev. 63 (1999) 349–404.
[10] F. Fabre, H. Roman, Genetic evidence for inducibility ofrecombination competence in yeast, Proc. Natl. Acad. Sci.U.S.A. 74 (1977) 1667–1671.
[11] O. Niwa, R. Kominami, Untargeted mutation of thematernally derived mouse hypervariable minisatellite allelein F1 mice born to irradiated spermatozoa, Proc. Natl. Acad.Sci. U.S.A. 98 (2001) 1705–1710.
[12] K. Shiraishi, T. Shimura, M. Taga, N. Uematsu, Y. Gondo, M.Ohtaki, R. Kominami, O. Niwa, Persistent induction ofsomatic reversions of the pink-eyed unstable mutation in F1
mice born to fathers irradiated at the spermatozoa stage,Radiat. Res. 157 (2002) 661–667.
[13] T. Shimura, M. Inoue, M. Taga, K. Shiraishi, N. Uematsu, N.Takei, Z.-M. Yuan, T. Shinohara, O. Niwa, p53-dependentS-phase damage checkpoint and pronuclear cross talk inmouse zygotes with X-irradiated sperm, Mol. Cell. Biol. 22(2002) 2220–2228.
[14] T. Shimura, M. Toyoshima, S.K. Adiga, T. Kunoh, H. Nagai, N.Shimizu, M. Inoue, O. Niwa, Suppression of replication forkprogression in low-dose-specific p53-dependent S-phaseDNA damage checkpoint, Oncogene 25 (2006)5921–5932.
[15] O. Niwa, Indirect mechanisms of genomic instability and thebiological significance of mutations at tandem repeat loci,Mutat. Res. 598 (2006) 61–72.
[16] K. Ostermann, A. Lorentz, H. Schmidt, The fission yeastrad22 gene, having a function in mating-type switching andrepair of DNA damages, encodes a protein homolog to Rad52of Saccharomyces cerevisiae, Nucleic Acids Res. 21 (1993)5940–5944.
[17] C. Alfa, P. Fantes, J. Hyams, M. McLeod, E. Warbrick,Experiments with Fission Yeast, Cold Spring Harbor
Laboratory Press, New York, 1993.[18] A. Wach, PCR-synthesis of marker cassettes with longflanking homology regions for gene disruptions in S.cerevisiae, Yeast 12 (1996) 259–265.

( 2 0
1260 d n a r e p a i r 7[19] M. Suga, T. Hatakeyama, A rapid and simple procedure forhigh-efficiency lithium acetate transformation ofcryopreserved Schizosaccharomyces pombe cells, Yeast 22(2005) 799–804.
[20] J. Bahler, J.-Q. Wu, M.S. Longtine, N.G. Shah, A. McKenzie III,A.B. Steever, A. Wach, P. Philippsen, J.R. Pringle,Heterologous modules for efficient and versatile PCR-basedgene targeting in Schizosaccharomyces pombe, Yeast 14 (1998)943–951.
[21] P. Raspor, S. Plesnicar, Z. Gazdag, M. Pesti, M. Miklavcic, B.Lah, R. Logar-Marinsek, B. Poljsak, Prevention of intracellularoxidation in yeast: the role of vitamin E analogue, Trolox(6-hydroxy-2,5,7,8-tetramethylkroman-2-carboxyl acid), CellBiol. Int. 29 (2005) 57–63.
[22] E. Zyracka, R. Zadrag, S. Koziol, A. Krzepilko, G. Bartosz, T.Bilinski, Ascorbate abolishes auxotrophy caused by the lackof superoxide dismutase in Saccharomyces cerevisiae. Yeastcan be a biosensor for antioxidants, J. Biotechnol. 115 (2005)271–278.
[23] R. Wang, Y. Jin, D. Norris, Identification of a protein thatbinds to the HO endonuclease recognition sequence at theyeast mating type locus, Mol. Cell. Biol. 17 (1997) 770–777.
[24] Y. Fujita, Y. Giga-Hama, K. Takegawa, Development of agenetic transformation system using new selectablemarkers for fission yeast Schizosaccharomyces pombe, Yeast 22(2005) 193–202.
[25] B. Chaudhuri, S. Ingavale, A.K. Bachhawat, apd1+, a generequired for red pigment formation in ade6 mutants ofSchizosaccharomyces pombe, encodes an enzyme required forglutathione biosynthesis: a role for glutathione and aglutathione-conjugate pump, Genetics 145 (1997)75–83.
[26] T. Nagai, K. Ibata, E.S. Park, M. Kubota, K. Mikoshiba, A.Miyawaki, A variant of yellow fluorescent protein with fastand efficient maturation for cell-biological applications, Nat.Biotechnol. 20 (2002) 87–90.
[27] D.D. Dubey, J. Zhu, D.L. Carlson, K. Sharma, J.A. Huberman,Three ARS elements contribute to the ura4 replication originregion in the fission yeast, Schizosaccharomyces pombe, EMBOJ. 13 (1994) 3638–3647.
[28] R. Rowley, Radiation-induced mitotic delay: a geneticcharacterization in the fission yeast, Radiat. Res. 132 (1992)144–152.
[29] C. Shao, F.M. Lyng, M. Folkard, K.M. Prise, Calcium fluxesmodulate the radiation-induced bystander responses intargeted glioma and fibroblast cells, Radiat. Res. 166 (2006)479–487.
[30] M. van den Bosch, K. Vreeken, J.B. Zonneveld, J.A. Brandsma,M. Lombaerts, J.M. Murray, P.H. Lohman, A. Pastink,Characterization of RAD52 homologs in the fission yeastSchizosaccharomyces pombe, Mutat. Res. 461 (2001)311–323.
[31] C.L. Doe, F. Osman, J. Dixon, M.C. Whitby, DNA repair by aRad22-Mus81-dependent pathway that is independent ofRhp51, Nucleic Acids Res. 32 (2004) 5570–5581.
[32] W.J. Kim, S. Lee, M.S. Park, Y.K. Jang, J.B. Kim, S.D. Park,Rad22 protein, a Rad52 homologue in Schizosaccharomycespombe, binds to DNA double-strand breaks, J. Biol. Chem. 275(2000) 35607–35611.
[33] P. Meister, M. Poidevin, S. Francesconi, I. Tratner, P. Zarzov, G.Baldacci, Nuclear factories for signalling and repairing DNAdouble strand breaks in living fission yeast, Nucleic AcidsRes. 31 (2003) 5064–5073.
[34] K.M. Prise, G. Ahnstrom, M. Belli, J. Carlsson, D. Frankenberg,
J. Kiefer, M. Lobrich, B.D. Michael, J. Nygren, G. Simone, B.Stenerlow, A review of dsb induction data for varying qualityradiations, Int. J. Radiat. Biol. 74 (1998)173–184.0 8 ) 1250–1261
[35] V. Wood, R. Gwilliam, M.-A. Rajandream, M. Lyne, R. Lyne, A.Stewart, J. Sgouros, N. Peat, J. Hayles, S. Baker, D. Basham, S.Bowman, K. Brooks, D. Brown, S. Brown, T. Chillingworth, C.Churcher, M. Collins, R. Connor, A. Cronin, P. Davis, T.Feltwell, A. Fraser, S. Gentles, A. Goble, N. Hamlin, D. Harris,J. Hidalgo, G. Hodgson, S. Holroyd, T. Hornsby, S. Howarth,E.J. Huckle, S. Hunt, K. Jagels, K. James, L. Jones, M. Jones, S.Leather, S. McDonald, J. McLean, P. Mooney, S. Moule, K.Mungall, L. Murphy, D. Niblett, C. Odell, K. Oliver, S. O’Neil,D. Pearson, M.A. Quail, E. Rabbinowitsch, K. Rutherford, S.Rutter, D. Saunders, K. Seeger, S. Sharp, J. Skelton, M.Simmonds, R. Squares, S. Squares, K. Stevens, K. Taylor, R.G.Taylor, A. Tivey, S. Walsh, T. Warren, S. Whitehead, J.Woodward, G. Volckaert, R. Aert, J. Robben, B. Grymonprez, I.Weltjens, E. Vanstreels, M. Rieger, M. Schafer, S. Muller-Auer,C. Gabel, M. Fuchs, A. Dusterhoft, C. Fritzc, E. Holzer, D.Moestl, H. Hilbert, K. Borzym, I. Langer, A. Beck, H. Lehrach,R. Reinhardt, T.M. Pohl, P. Eger, W. Zimmermann, H. Wedler,R. Wambutt, B. Purnelle, A. Goffeau, E. Cadieu, S. Dreano, S.Gloux, V. Lelaure, S. Mottier, F. Galibert, S.J. Aves, Z. Xiang, C.Hunt, K. Moore, S.M. Hurst, M. Lucas, M. Rochet, C.Gaillardin, V.A. Tallada, A. Garzon, G. Thode, R.R. Daga, L.Cruzado, J. Jimenez, M. Sanchez, F. del Rey, J. Benito, A.Domınguez, J.L. Revuelta, S. Moreno, J. Armstrong, S.L.Forsburg, L. Cerutti, T. Lowe, W.R. McCombie, I. Paulsen, J.Potashkin, G.V. Shpakovski, D. Ussery, B.G. Barrell, P. Nurse,The genome sequence of Schizosaccharomyces pombe, Nature415 (2002) 871–880.
[36] J.B. Little, Genomic instability and bystander effects: ahistorical perspective, Oncogene 22 (2003) 6978–6987.
[37] B. McClintock, The stability of broken ends of chromosomesin Zea mays, Genetics 26 (1941) 234–282.
[38] S.M. Bailey, J.P. Murnane, Telomeres, chromosome instabilityand cancer, Nucleic Acids Res. 34 (2006) 2408–2417.
[39] W.F. Morgan, Is there a common mechanism underlyinggenomic instability, bystander effects and other nontargetedeffects of exposure to ionizing radiation? Oncogene 22 (2003)7094–7099.
[40] M.A. Kadhim, D.A. Macdonald, D.T. Goodhead, S.A. Lorimore,S.J. Marsden, E.G. Wright, Transmission of chromosomalinstability after plutonium �-particle irradiation, Nature 355(1992) 738–740.
[41] L. Sabatier, B. Dutrillaux, M.B. Martin, Chromosomalinstability, Nature 357 (1992) 548.
[42] W.F. Morgan, Non-targeted and delayed effects of exposureto ionizing radiation: I. Radiation-induced genomicinstability and bystander effects in vitro, Radiat. Res. 159(2003) 567–580.
[43] M.A. Kadhim, S.R. Moore, E.H. Goodwin, Interrelationshipsamongst radiation-induced genomic instability, bystandereffects, and the adaptive response, Mutat. Res. 568 (2004)21–32.
[44] J. Molinier, G. Ries, C. Zipfel, B. Hohn, Transgenerationmemory of stress in plants, Nature 442 (2006) 1046–1049.
[45] K. Suzuki, M. Ojima, S. Kodama, M. Watanabe, Delayedactivation of DNA damage checkpoint and radiation-inducedgenomic instability, Mutat. Res. 597 (2006) 73–77.
[46] H. van Attikum, S.M. Gasser, The histone code at DNAbreaks: a guide to repair? Nat. Rev. Mol. Cell Biol. 6 (2005)757–765.
[47] C. Redon, D. Pilch, E. Rogakou, O. Sedelnikova, K. Newrock,W. Bonner, Histone H2A variants H2AX and H2AZ, Curr.Opin. Genet. Dev. 12 (2002) 162–169.
[48] Y. Ziv, D. Bielopolski, Y. Galanty, C. Lukas, Y. Taya, D.C.
Schultz, J. Lukas, S. Bekker-Jensen, J. Bartek, Y. Shiloh,Chromatin relaxation in response to DNA double-strandbreaks is modulated by a novel ATM- and KAP-1 dependentpathway, Nat. Cell Biol. 8 (2006) 870–876.
0 0 8
d n a r e p a i r 7 ( 2[49] R. Fukumura, H. Takahashi, T. Saito, Y. Tsutsumi, A.Fujimori, S. Sato, K. Tatsumi, R. Araki, M. Abe, A sensitivetranscriptome analysis method that can detect unknowntranscripts, Nucleic Acids Res. 31 (2003) e94.
) 1250–1261 1261
[50] R.J. Brennan, R.H. Schiestl, Persistent genomic instability inthe yeast Saccharomyces cerevisiae induced by ionizingradiation and DNA-damaging agents, Radiat. Res. 155 (2001)768–777.





























