Embed Size (px)
Citation preview

PDFlib PLOP: PDF Linearization, Optimization, Protection
Page inserted by evaluation versionwww.pdflib.com – [email protected]

Protein Tyrosine Phosphorylation and Estradiol Action a
F. AURICCHIO, A. MIGLIACCIO, G. CASTORIA, M. DI DOMENICO, A. BILANCIO, AND A. ROTONDI
II Cattedra di Patologia Generale Facolta di Medicina e Chirirrgia
II Universita di Napoli 80138 Napoli, Italy
INTRODUCTION
Steriod hormones regulate differentiation and proliferation of several cell types. They interact with intracellular receptors that belong to the family of ligand- inducible transcriptional enhancer factors, In contrast to steroids, peptide growth factors interact with membrane receptors, producing intracellular signals that are responsible for short-term responses. Steroids also produce extragenomic, rapid responses similar to those evoked by the peptide growth factors. Within a few seconds, vitamin D3 treatment activates phospholipase C from intact en terocyte~ .~ Within seconds, progesterone stimulates calcium influx in sperm.5 Within minutes, progesterone directly causes an increase of oxytocin receptor binding in the poste- nor ventromedial hypothalamus.6 TABLE 1 presents some of the nongenomic effects of steroid hormones.
We observed that estradiol complexed with its receptor stimulates in uitro a 67-kDa tyrosine k i n a ~ e . ~ The kinase has been isolated and some of the in uitro substrates have been identified. The enzyme phosphorylates synthesized human wild-type (HEGO) and a point-mutated (HEO) estradiol receptor (ER).'.' Phosphorylation of the tyrosyl residue at position 537 of the hormone binding domain of the synthetic ER confers hormone binding with high affinity to the receptor."' The physiological relevance of this observation has been proved recently by finding the same tyrosyl residue phosphorylation in whole MCF-7 cell ER."
FIGURE 1 shows a model of the 67-kDa tyrosine kinase activation by the receptor occupied by the hormone. This model, extensively verified in uitro in our laboratory, immediately suggests that estradiol directly stimulates tyrosine kinase(s) and protein tyrosine phosphorylation in uiuo. Stimulation of protein phosphorylation on tyrosine is a well-known early effect of peptide growth fac- t o r ~ ~ ~ - " and is also involved in cell transformation induced by retroviral onco- gene~. '~ , ' ' Like these growth factors, estradiol may stimulate tyrosine phosphoryla- tion of intact cells. This possibility has been investigated using human breast cancer MCF-7 cells under conditions of estradiol-stimulated growth.
This work was supported by grants from the Associazione Italiana per la Ricerca sul Cancro (AIRC), from the Consiglio Nazionale delle Ricerche (CNR; Progetti Speciali: Appli- cazioni Cliniche della Ricerca Oncologica ed Ingegneria Genetica), and from the Minister0 dell'Universitl e della Ricerca Scientifica.
149

150 ANNALS NEW YORK ACADEMY OF SCIENCES
TABLE 1. Examples of Nontranscriptional Effects of Steroid Hormones Steroid Effect System
vitamin D, activation of phospholipase C intact enterocytes4 progesterone increase of oxytocin receptor posterior ventromedial hypo-
estradiol stimulation of c-src MCF-7 cells4 binding thalamus6
ESTRADIOL STIMULATES PROTEIN TYROSINE PHOSPHORYLATION AND c-src ACTIVITY OF WHOLE MCF-7 CELLS
Metabolic Labeling of MCF-7 Cells with [3zP]-Orthophosphate
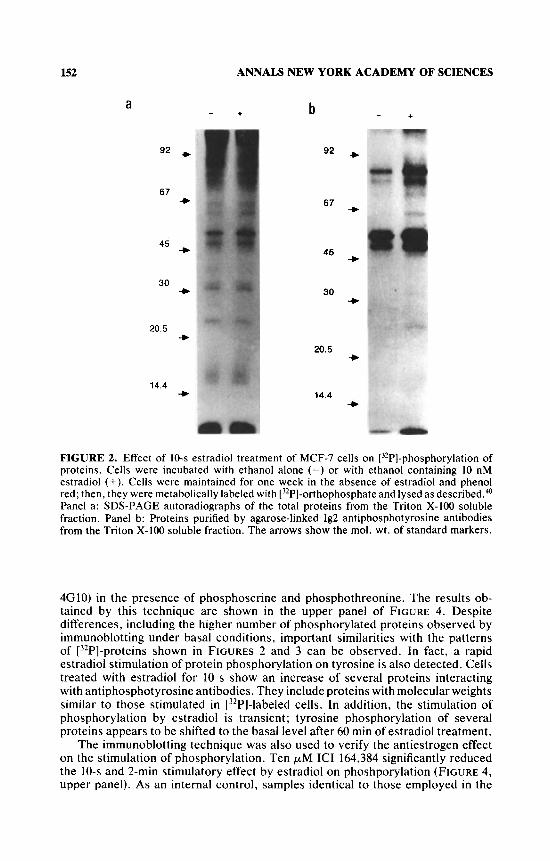
MCF-7 cells were grown for 2-3 weeks in a phenol red-free medium supple- mented with fetal calf serum treated with charcoal. Under these conditions, cell growth is very sensitive to the presence of estradiol.ls Cells were labeled with [32Pl-orthophosphate, and the detergent-soluble fraction of the cell lysate was used as a source to purify phosphotyrosine-containing proteins by interaction with 1G2 antiphosphotyrosine antibodies coupled to agarose and by specific elution with phenyl-phosphate. l9 Purified proteins were analyzed by SDS-PAGE. FIGURE 2b shows that some proteins under basal conditions were phosphorylated. Ten sec- onds (s) of 10 nM estradiol treatment stimulated tyrosine phosphorylation of these proteins (FIGURE 2b). Estradiol treatment only slightly affected the total protein phosphorylation of the Triton X-100 soluble fraction (FIGURE 2a). indicating that after 10 s this steroid mostly stimulates phosphorylation of a specific family of proteins, interacting with the antiphosphotyrosine antibody.
To answer the question of whether estradiol stimulates protein phosphorylation via its own receptor, MCF-7 cells were incubated for 10 s with the vehicle alone or with estradiol in the absence and in the presence of two nonsteroidal antiestro- gens acting through their binding to the ER. We employed 1 pM of OH-tamoxifen or the pure antiestrogen ICI 164,384*O at two different concentrations (1 or 10 pM). Both antiestrogens efficiently antagonized the effect of estradiol (FIGURE 3). Fifty nM of progesterone of 10 nM of OH-testosterone, added to the MC'F cells for 10 s, did not stimulate phosphorylation of proteins interacting with the antiphosphotyrosine antibody (not shown). Therefore, stimulation of tyrosine phosphorylation under these experimental conditions was restricted to estradiol.
Antiphosphotyrosine Immunoblotting Experiments
Stimulation of phosphorylation on tyrosine by estradiol was also investigated using an antiphosphotyrosine antibody immunoblot. This method can be more sensitive than immunoprecipitation with the antiphosphotyrosine antibody.2' MCF-7 cells grown for 1 week in a phenol red-free medium were exposed to 10 nM estradiol at 37°C for 0 time, 10 s, and 2, 15, and 60 min. Cells were quickly cooled, homogenized, and centrifuged at 13,000 rpm. Finally, supernatant aliquots containing equal amounts of proteins were run on SDS-PAGE and were submitted to immunoblotting with Ig2bk antiphosphotyrosine monoclonal antibodies (clone

PTK
ac
tivat
ed P
TK
tYr
subs
trat
e E
stra
diol
-,
P-t
yr
FIGURE 1.
A m
odel
of
the
67-k
Da
tyro
sine
kin
ase
activ
atio
n by
the
rece
ptor
occ
upie
d by
the
hor
mon
e. T
erm
s-R
: re
cept
or;
ER:
estra
diol
- re
cept
or c
ompl
ex; F
TK: p
rote
in t
yros
ine
kina
se;
tyr:
tyr
osin
e; P
-tyr:
phos
phot
yros
ine.
I-
01
I-
152
a
67 +
45 +
30 +
20.5 +
14.4 -+
ANNALS NEW YORK ACADEMY OF SCIENCES
- + b +
92 +
67 -+
45 -w
30 +
20.5 +
14.4 -w
FIGURE 2. Effect of 10-s estradiol treatment of MCF-7 cells on [32P]-phosphorylation of proteins. Cells were incubated with ethanol alone (-) or with ethanol containing 10 nM estradiol (+). Cells were maintained for one week in the absence of estradiol and phenol red; then, they were metabolically labeled with [3'P]-orthophosphate and lysed as described.40 Panel a: SDS-PAGE autoradiographs of the total proteins from the Triton X-100 soluble fraction. Panel b: Proteins purified by agarose-linked Ig2 antiphosphotyrosine antibodies from the Triton X-100 soluble fraction. The arrows show the mol. wt. of standard markers.
4G 10) in the presence of phosphoserine and phosphothreonine. The results ob- tained by this technique are shown in the upper panel of FIGURE 4. Despite differences, including the higher number of phosphorylated proteins observed by immunoblotting under basal conditions, important similarities with the patterns of ["PI-proteins shown in FIGURES 2 and 3 can be observed. In fact, a rapid estradiol stimulation of protein phosphorylation on tyrosine is also detected. Cells treated with estradiol for 10 s show an increase of several proteins interacting with antiphosphotyrosine antibodies. They include proteins with molecular weights similar to those stimulated in [32P]-labeled cells. In addition, the stimulation of phosphorylation by estradiol is transient; tyrosine phosphorylation of several proteins appears to be shifted to the basal level after 60 min of estradiol treatment.
The immunoblotting technique was also used to verify the antiestrogen effect on the stimulation of phosphorylation. Ten pM ICI 164,384 significantly reduced the 10-s and 2-min stimulatory effect by estradiol on phoshporylation (FIGURE 4, upper panel). As an internal control, samples identical to those employed in the

AURICCHIO el al.: PHOSPHORYLATION
1 2 3 4
153
5
45 -It
30 -+
2 0 5 +
14.4 +
FIGURE 3. Stimulation of [3'P]-phosphorylation of proteins interacting with antiphospho- tyrosine antibodies after 10-s estradiol treatment i s inhibited by antiestrogens in MCF-7 cells. MCF-7 cells were maintained for one week in the absence of estradiol and phenol red and proteins were [3ZP]-phosphorylated as de~cribed.~" Proteins from the Triton X- 100 soluble fraction of cell lysates were purified by agarose-linked lg2 antiphosphotyrosine antibodies and then submitted to SDS-PAGE. The gel was fixed and exposed for autoradiography. Lane 1: MCb-7 cells incubated for 10 s with vehicle alone; lane 2: MCF-7 cells incubated with 10 nM estradiol; lanes 3 and 4: as in lane 2 plus either I or 10 p M ICI 164,384, respectively; lane 5: as in lane 2 plus 1 pM OH-tamoxifen. The arrows indicate the mol. wt. of standard markers.

154 ANNALS NEW YORK ACADEMY OF SCIENCES
- - - - + + ICt - E 2 + + + + + + +
time: 0 10s 2m 15m 60m 10s 2m
200 -.
31
actin +
FIGURE 4. Time course of stimulation by estradiol of protein tyrosine phosphorylation followed by a Western blot with antiphosphotyrosine antibodies. Upper panel: MCF-7 cells maintained for one week in the absence of estradiol add phenol red were treated with 10 nM estradiol for 0 time, 10 s, and 2, 15, and 60 min or with estradiol (10 nM) plus 10 pM ICI 164,384 for 10 s and 2 min. Cell lysates were prepared and proteins (80 pg) were resolved by SDS-PAGE, transferred to nitrocellulose, and hybridized with Ig2bk antiphosphotyrosine monoclonal antibodies as described.@ Lower panel: samples identical to those shown in the upper panel were blotted with monoclonal antiactin antibodies.

AURICCHIO er al.: PHOSPHORYLATION 155
experiment shown in the upper panel of FIGURE 4 were submitted in parallel to Western blotting with monoclonal antiactin antihodies. The same amount of actin was detected in each lane (FIGURE 4, lower panel), indicating that the samples of the experiment presented in the upper panel contained the same amount of proteins.
In conclusion, both labeling and immunoblotting studies show that estradiol stimulates protein tyrosine phosphorylation of MCF-7 cells in 10 s. Phosphoryla- tion declines toward the basal level after 60 min of estradiol treatment. Estradiol stimulation requires its occupancy of the receptor. Estradiol does not affect tyro- sine phosphorylation of nontarget cells and its effect on tyrosine phosphorylation is not shared by other steroids.
c-src Activiy
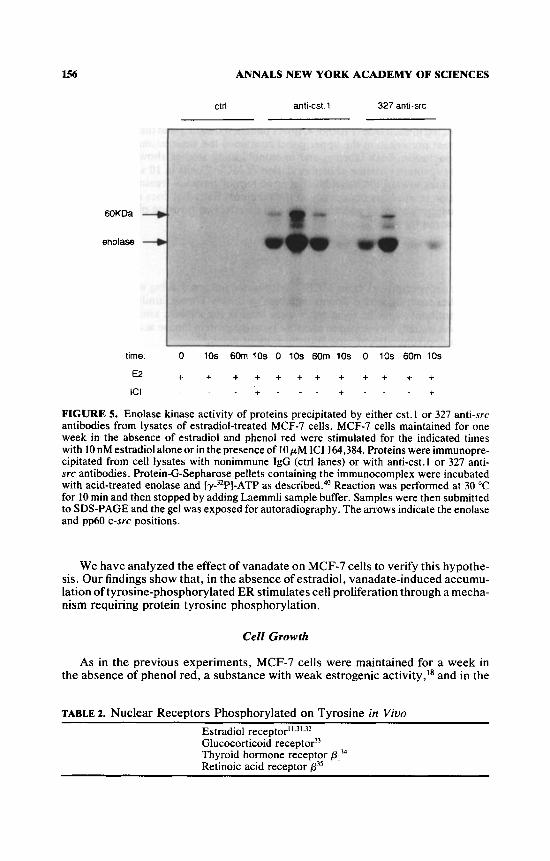
Immunoprecipitates from MCF-7 cell lysates prepared using either anti-cst , 1 antibodies reacting with different members of the src kinase family or 327 mono- clonal antibody, specific for C-STC,~* were used in the enolase kinase assay. This assay was initially developed to measure tyrosine protein kinase activity contained in immunoprecipitates of viral transforming proteins.23
The results observed with this assay are presented in FIGURE 5. The kinase activity immunoprecipitated by the anti-cst. 1 antibody behaves similar to the one immunoprecipitated by the 327 antibody (FIGURE 5). The 327 antibody has been raised against an epitope localized in an src homology-3 domain (SH3) of pp60c-s"'. The enolase kinase activity of both immunoprecipitates was increased after 10 s of estradiol treatment of MCF-7 cells and is greatly reduced after 60 min. Ten pM ICI 164,384 abolishes the 10-s estradiol-treatment stimulatory effect. In addition to the [32P]-protein corresponding to the enolase, a [32P]-protein with a molecular weight of approximately 60 kDa was observed in FIGURE 5 whose phosphorylation pattern mirrors that of enolase. This protein represents the immu- noprecipitated kinase. The association of the enolase kinase activity with the immunoprecipitates is specific. No enzyme activity was observed when a control (ctrl) antibody was used for the immunoprecipitation (FIGURE 5 ) .
VANADATE STIMULATION OF MCF-7 CELL GROWTH IN THE ABSENCE OF ESTRADIOL REQUIRES ER ACTIVITY
The turnover of phosphate in phosphotyrosine is Therefore, the balance between phosphorylation and dephosphorylation can be strongly affected by PTP-ase inhibitors. One of the compounds able to increase the phosphotyrosine content of proteins is vanadate, which inhibits PTP-ases in vitro as well as in uivo.26-Z8 Vanadate is widely distributed in normal biological systems at concentra- tions typically less than 1 pMZ9 and has different properties, including induction of cell transformation.30
Four nuclear receptors, namely, I7P-ER (ER),".31.32 glucocorticoid receptor,33 thyroid hormone receptor PI ,34 and retinoic acid receptor p,35 have been found to be phosphorylated on tyrosine in uiuo (see TABLE 2).
As reported above, estradiol treatment of estradiol-responsive MCF-7 cells immediately induces the reversible activation of c-SYC kinase and the phosphoryla- tion of several proteins. Taken together, these data show that different nuclear receptors including ER are physiological substrates of PTKs and suggest that ER may act on proliferation by stimulating tyrosine phosphorylation pathways.
156 ANNALS NEW YORK ACADEMY OF SCIENCES
ctrl anti-cst.1 327 anti-src
6OKDa +
enolase +
time. 0 10s 6Om 10s 0 10s 60m 10s 0 10s 60m 10s
€2 + + + + + + + + + + + + + - . - + ICI . . + . - .
FIGURE 5. Enolase kinase activity of proteins precipitated by either cst.1 or 327 anti-src antibodies from lysates of estradiol-treated MCF-7 cells. MCF-7 cells maintained for one week in the absence of estradiol and phenol red were stimulated for the indicated times with 10 nM estradiol alone or in the presence of 10 p M ICI 164,384. Proteins were immunopre- cipitated from cell lysates with nonimmune IgG (ctrl lanes) or with anti-cst.1 or 327 anti- src antibodies. Protein-G-Sepharose pellets containing the immunocomplex were incubated with acid-treated enolase and [y-3ZP]-ATP as de~cribed.~' Reaction was performed at 30 "C for 10 min and then stopped by adding Laemmli sample buffer. Samples were then submitted to SDS-PAGE and the gel was exposed for autoradiography. The arrows indicate the enolase and pp60 c-src positions.
We have analyzed the effect of vanadate on MCF-7 cells to verify this hypothe- sis. Our findings show that, in the absence of estradiol, vanadate-induced accumu- lation of tyrosine-phosphorylated ER stimulates cell proliferation through a mecha- nism requiring protein tyrosine phosphorylation.
Cell Growth
As in the previous experiments, MCF-7 cells were maintained for a week in the absence of phenol red, a substance with weak estrogenic activity,I8 and in the
TABLE 2. Nuclear Receptors Phosphorylated on Tyrosine in Viuo Estradiol receptoril,3'.32 Glucocorticoid receptor33 Thyroid hormone receptor p,34 Retinoic acid receptor p''

AURICCHIO ef al.: PHOSPHORYLATION 157
0 2 4 6 8 days
t 1 0
0 1 2 3 4 5 vanadate In the medlurn (pM)
FIGURE 6. Vanadate dose-dependent effect on MCF-7 cell growth. Subconfluent MCF-7 cells were maintained for one week in the absence of estradiol and phenol red.32 Upper panel: MCF-7 cells untreated (0) or treated (0) with 2 p M vanadate were counted at the indicated times. The values of each point are averaged from seven different experiments. The statistical significance of these results was evaluated by a paired t test. P values were <0.001 at the third and fourth days and <0.005 at day eight. Points: mean; bars: SD. Lower panel: MCF-7 cells were treated with increasing amounts of vanadate and were counted after four days.

158 ANNALS NEW YORK ACADEMY OF SCIENCES
presence of charcoal-treated serum. The cell growth under these conditions was slow (FIGURE 6, upper panel). Two pM of vanadate significantly stimulated it (FIGURE 6, upper panel). The study of dose-dependence showed that the prolifera- tive effect of vanadate reached a plateau at 1 pM, declined at 3 pM, and disappeared at 5 p M (FIGURE 6 , lower panel). Interestingly, whcn vanadate was simultaneously added to either 1 pM of OH-tamoxifen (FIGURE 7A) or 10 pM of ICI 164,384 (FIGURE 7B), its effect was abolished. Addition of ICI 164,384 to the medium in the absence of vanadate did not reduce the slow basal growth rate of MCF-7 cells (not shown). The inhibitory effect of 3-day ICI 164,384 treatment on vanadate- induced cell growth was removed by estradiol (FIGURE 7B), indicating that the antiestrogen was acting through its receptor. Ten pM of ICI 182,780 also inhibited vanadate’s proliferative effect. Twenty pM of estradiol in the cell medium removed this inhibition (not shown). It is worthwhile to stress that the proliferative effect of vanadate was abolished by 10 pM of genistein, a well known tyrosine kinase inhibitor (not shown). Therefore, antiestrogens and genistein both prevent cell growth, inhibiting the accumulation of tyrosine-phosphorylated proteins.
Total Protein Tyrosine Phosphorylation
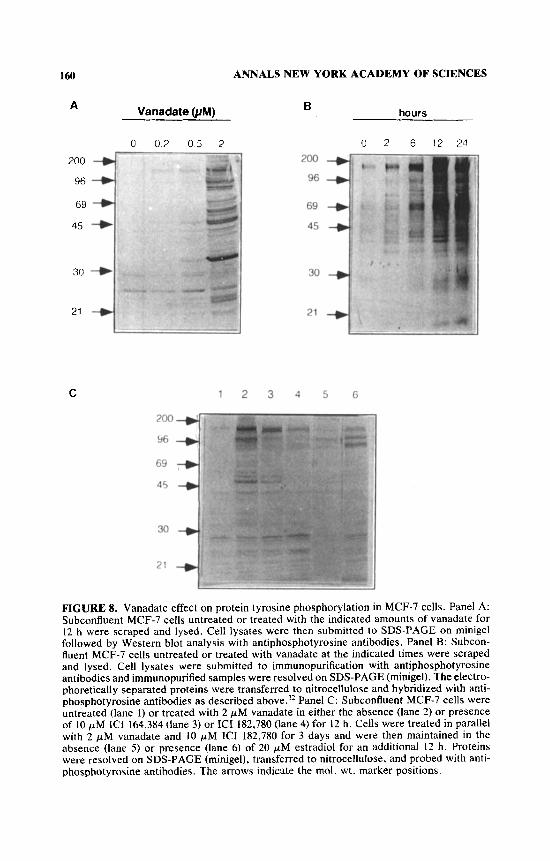
Vanadate was added to the medium in the absence or presence of either ICI 182,780 or ICI 164,384 and, after 12 h, cells were lysed, submitted to SDS-PAGE, and blotted with monoclonal antiphosphotyrosine. Results are shown in FIGURE 8. Both antiestrogens reduced the level of protein phosphorylation on tyrosine in vanadate-treated cells. After 3 days, the inhibitory effect of ICI 182,780 on vana- date stimulation of tyrosine phosphorylation was even more pronounced than after 12 h. By adding an excess of estradiol. the inhibitory effect of the 3-day treatment with antiestrogen was partially removed. This shows that the antiestro- gen was acting through occupancy of its receptor. In a parallel experiment, estra- diol also abolished the inhibitory effect of ICI 182,780 on vanadate-induced cell growth (not shown).
In conclusion, vanadate induces tyrosine phosphorylation and cell growth of MCF-7 cells. Both are inhibited by antiestrogen treatment and estradiol removes this inhibition. This indicates that ER plays a key role in stimulationg tyrosine phosphorylation and proliferation induced by the PTP-ase inhibitor.
ER Tyrosine Phosphorylation
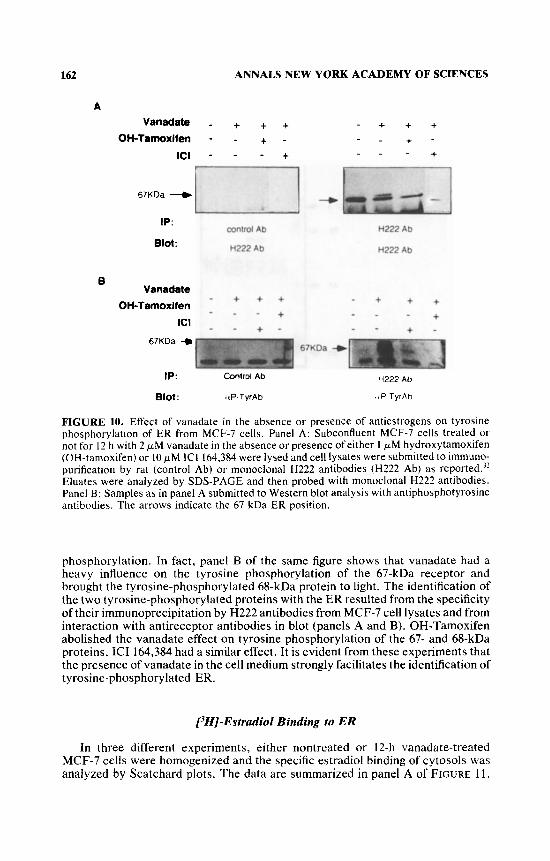
The receptor was purified by either antireceptor H222 antibodies or control antibodies from cell lysate and blotted with either H222 antibodies or antiphospho- tyrosine antibodies. FIGURE 9A shows that the 67-kDa receptor, specifically precip- itated by H222 antibodies, is phosphorylated on tyrosine.
We then studied the effect of 12-h vanadate treatment on the tyrosine phosphor- ylation of ER (FIGURE 9B). This was significantly increased. We also verified the effect of OH-tamoxifen and ICI 164,384 on the receptor phosphorylation induced by vanadate. Purification of the receptor from lysate by either antibodies directed against ER or control antibodies followed by blotting with the antireceptor antibod- ies showed that a minor protein band interacting with antireceptor antibody and migrating at about 68 kDa appeared after vanadate treatment (FIGURE IOA). OH- Tamoxifen and ICI 164,384 (FIGURE 10A) as well as ICI 182,780 (not shown) abolished the 68-kDa protein. The doublet is probably the consequence of receptor

AURICCHIO ef a/.: PHOSPHORYLATION 159
A
l o 5 1
I
4 0 2 6 6 10 O J
/ 3
I
2 4 6 8 10 d.W
0 VanCldauI
+ICI vanadate . vanadate +ICI+E2
0 wntrol
FIGURE 7. Stimulation by vanadate of MCF-7 cell growth. Subconfluent MCF-7 cells were grown as described.32 Panel A: MCF-7 cells untreated (0) or treated with 2 p M vanadate either alone (0) or with 1 pM hydroxytamoxifen (m) were counted at the indicated times. Panel B: MCF-7 cells were grown in the absence (0) and presence of 2 pM vanadate either without (0) or with (a) 10 pM ICI 164,384. Cells were also grown in the presence of vanadate, ICI 164,384, and subsequently (see arrow) 1.25 pM estradiol (+).
ANNALS NEW YORK ACADEMY OF SCIENCES 160
A Vanadate UM)
0 0 2 0 5 2
30 +
C
hours B
0 2 6 12 21
200 + 96 +
1 2 3 4 5 G
FIGURE 8. Vanadate effect on protein tyrosine phosphorylation in MCF-7 cells. Panel A: Subconfluent MCF-7 cells untreated or treated with the indicated amounts of vanadate for 12 h were scraped and lysed. Cell lysates were then submitted to SDS-PAGE on minigel followed by Western blot analysis with antiphosphotyrosine antibodies. Panel B: Subcon- fluent MCF-7 cells untreated or treated with vanadate at the indicated times were scraped and lysed. Cell lysates were submitted to immunopurification with antiphosphotyrosine antibodies and irnrnunopurified samples were resolved on SDS-PAGE (minigel). The electro- phoretically separated proteins were transferred to nitrocellulose and hybridized with anti- phosphotyrosine antibodies as described above.’* Panel C: Subconfluent MCF-7 cells were untreated (lane 1) or treated with 2 pM vanadate in either the absence (lane 2) or presence of 10 pM ICI 164,384 (lane 3) or ICI 182,780 (lane 4) for 12 h. Cells were treated in parallel with 2 p M vanadate and 10 pM ICI 182,780 for 3 days and were then maintained in the absence (lane 5) or presence (lane 6) of 20 pM estradiol for an additional 12 h. Proteins were resolved on SDS-PAGE (minigel), transferred to nitrocellulose, and probed with anti- phosphotyrosine antibodies. The arrows indicate the mol. wt. marker positions.

AURICCHIO el af.: PHOSPHORYLATION 161
A
II
47 -0
97 - R PP 97- R %p 47 - 4? - 30-
1 P H222Ab
Blot: H222Ab
a- m Ab
a P-Qr Ab
FIGURE 9. Tyrosine phosphorylation of ER in untreated or vanadate-treated MCF-7 cells. Panel A: Subconfluent MCF-7 cells were lysed and cell lysates were submitted to ER immunopurification by anti-ER monoclonal H222 (H222 lanes) or rat antibodies (control lanes) as described.” Eluates were resolved on SDS-PAGE and then blotted with either anti-ER monoclonal H222 or antiphosphotyrosine antibodies. Panel B: Subconfluent MCF-7 cells either treated or not for 12 h with 2 p M vanadate were lysed and proteins were submitted to immunopurification with monoclonal H222 antibodies as described.)* Eluates were re- solved on SDS-PAGE and then blotted with monoclonal H222 or antiphosphotyrosine anti- bodies. The arrows indicate the 67-kDa ER position. Standard markers were coelectro- phoresed.
162 ANNALS NEW YORK ACADEMY OF SCIENCES
A Vanadate
OH-Tamoxifen
ICI
67KDa +
IP:
Blot:
B Vanadate
OH-lamoxifen ICI
67KDa +
IP:
Blot:
- + + - - + - - -
control Ab
H222 Ab
- + + - _ -
+ - -
Control Ab
ttP-TyrAb
+
+
+ i
- + + + - - + - - - - +
H222 Ab
H222 Ab
- + + + - + + -
- - - -
67KDa +
H222 Ab
,IP TyrAb
FIGURE 10. Effect of vanadate in the absence or presence of antiestrogens on tyrosine phosphorylation of ER from MCF-7 cells. Panel A: Subconfluent MCF-7 cells treated or not for 12 h with 2 p M vanadate in the absence or presence of either I p M hydroxytamoxifen (OH-tamoxifen) or 10 pM ICI 164,384 were lysed and cell lysates were submitted to immuno- purification by rat (control Ab) or monoclonal H222 antibodies (H222 Ab) as rep~rted.~’ Eluates were analyLed by SDS-PAGE and then probed with monoclonal H222 antibodies. Panel B: Samples as in panel A submitted to Western blot analysis with antiphosphotyrosine antibodies. The arrows indicate the 67-kDa ER position,
phosphorylation. In fact, panel B of the same figure shows that vanadate had a heavy influence on the tyrosine phosphorylation of the 67-kDa receptor and brought the tyrosine-phosphorylated 68-kDa protein to light. The identification of the two tyrosine-phosphorylated proteins with the ER resulted from the specificity of their imrnunoprecipitation by H222 antibodies from MCF-7 cell lysates and from interaction with antireceptor antibodies in blot (panels A and B). OH-Tamoxifen abolished the vanadate effect on tyrosine phosphorylation of the 67- and 68-kDa proteins. ICI 164,384 had a similar effect. It is evident from these experiments that the presence of vanadate in the cell medium strongly facilitates the identification of tyrosine-phosphorylated ER.
[3H]-Estradiol Binding to ER
In three different experiments, either nontreated or 12-h vanadate-treated MCF-7 cells were homogenized and the specific estradiol binding of cytosols was analyzed by Scatchard plots. The data are summarized in panel A of FIGURE 11.
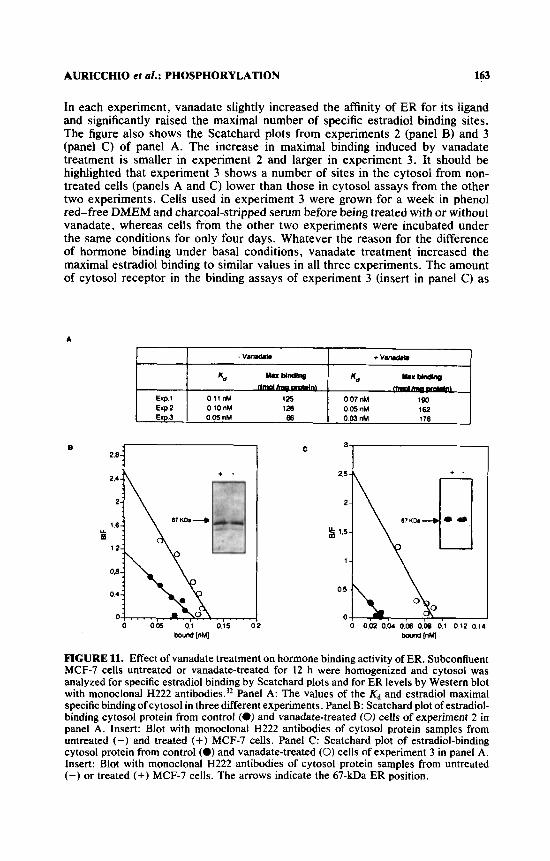
AURlCCHlO el 01.: PHOSPHORYLATION 163
In each experiment, vanadate slightly increased the affinity of ER for its ligand and significantly raised the maximal number of specific estradiol binding sites. The figure also shows the Scatchard plots from experiments 2 (panel B) and 3 (panel C) of panel A. The increase in maximal binding induced by vanadate treatment is smaller in experiment 2 and larger in experiment 3. It should be highlighted that experiment 3 shows a number of sites in the cytosol from non- treated cells (panels A and C) lower than those in cytosol assays from the other two experiments. Cells used in experiment 3 were grown for a week in phenol red-free DMEM and charcoal-stripped serum before being treated with or without vanadate, whereas cells from the other two experiments were incubated under the same conditions for only four days. Whatever the reason for the difference of hormone binding under basal conditions, vanadate treatment increased the maximal estradiol binding to similar values in all three experiments. The amount of cytosol receptor in the binding assays of experiment 3 (insert in panel C) as
B
0 0 . k 0,'l 0.i5 02 lmund [nMl
C 3
2 5 -
2-
1.5-
1 -
0.5 -
0 - h - , , . , 0 002 0.01 0.06 0.08 0.1 012 014 - IW
FIGURE 11. Effect of vanadate treatment on hormone binding activity of ER. Subconfluent MCF-7 cells untreated or vanadate-treated for 12 h were homogenized and cytosol was analyzed for specific estradiol binding by Scatchard plots and for ER levels by Western blot with monoclonal H222 antib~dies.)~ Panel A: The values of the Kd and estradiol maximal specific binding of cytosol in three different experiments. Panel B: Scatchard plot of estradiol- binding cytosol protein from control (8) and vanadate-treated (0) cells of experiment 2 in panel A. Insert: Blot with monoclonal H222 antibodies of cytosol protein samples from untreated (-) and treated (+) MCF-7 cells. Panel C: Scatchard plot of estradiol-binding cytosol protein from control (0) and vanadate-treated (0) cells of experiment 3 in panel A. Insert: Blot with monoclonal H222 antibodies of cytosol protein samples from untreated (-) or treated (+) MCF-7 cells. The arrows indicate the 67-kDa ER position.

164 ANNALS NEW YORK ACADEMY OF SCIENCES
well as in experiment 2 (insert in panel B) and experiment 1 (not shown) was similar in vanadate-treated and untreated cells. This was verified by blotting with anti-ER antibodies. Thus, while the level of hormone-binding ER can fluctuate under different conditions, the total amount of ER is constant and vanadate treat- ment converts most of it to hormone-binding receptor.
NONHORMONE-BINDING ER IN GR MOUSE MAMMARY TUMORS
The presence of nonhormone-binding ER (NHBER) in MCF-7 cells converted by vanadate treatment to hormone-binding receptor through tyrosine phosphoryla- tion in uiuo led us to wonder whether hormone-dependent (HD) and hormone- independent (HI) mammary tumors differ in their phosphorylation of ER. Mam- mary carcinomas induced in mice from the inbred GR strain are a useful model to investigate the changes in ER function in relation to the loss of hormone dependence. Mammary tumors induced in ovariectomized GR mice by hormonal treatment are usually HD, but first become hormone-responsive (HR) and finally HI during serial transplantations in synergic m i ~ e . ~ ~ . ~ ’ We have investigated whether the ERs from HD and HI mammary tumors of GR mice also differ in regard to phosphorylation and whether the in uitro phosphorylation affects the hormone binding to ER in these tumors.
Effect of ATP on [3H]-Estradiol Binding to ER
TABLES 3 and 4 list the results of the experiments investigating the effect of ATP on the hormone-binding ability of the estrogen receptor in tumor cytosol. We used different transplant passages from 13 separate GR mouse mammary tumor lines for this study. In most cases, different tumors from the same passage (transplant generation) were used. In some cases, when the tumor samples were small, they wcre pooled.
In the absence of ATP, HD and HR mammary tumor cytosol, on average, showed a higher specific [3H]-estradiol binding than HI tumors. Three out of 13 HDiHR tumors showed an ATP-induced increase in specific [3H]-estradiol binding of more than 10 fmol/mg protein. The highest increase observed in one of these tumors was 20 fmol of specific bindinglmg cytosol protein (TARLE 3).
The number of ATP-activated estradiol binding sites in 8 out of 20 HI tumors was significantly higher than in HD/HR tumors (values of 84, 50, 24, 53, 124, 79, 32, and 23 fmol of specific bindingimg cytosol protein; TABLE 4).
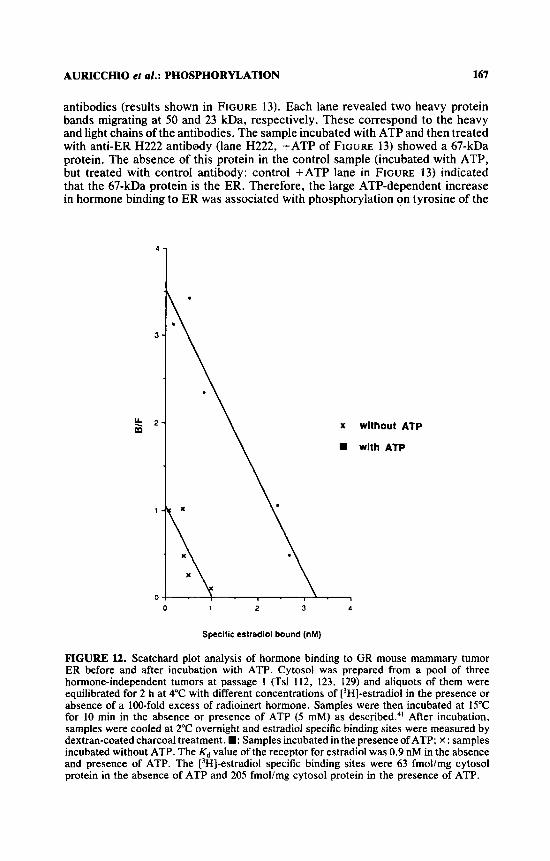
The effect of ATP on estradiol binding to ER was studied by Scatchard plot analysis. A pool of three HI tumors at passage 1, obtained from passages 1 and 2, was used. Cytosol was first incubated with different concentrations of [‘HI- estradiol (from 0.08 to 8 nM) in the absence and presence of an excess of cold hormone, then incubated with or without 5 mM ATP at 15°C for 10 min, and finally assayed for the [3H]-estradiol specific binding activity. Data are presented in FIGURE 12. Incubation with ATP induced a large increase in hormone binding to ER (63 fmol/mg cytosol protein without ATP and 205 fmol/mg cytosol protein with ATP) with no change of affinity (Kd about 0.9 nM in both the absence and presence of ATP).

AURICCHIO et ul.: PHOSPHORYLATLON 165
TABLE 3. Effect of ATP on Specific [3H]-Estradiol Binding in Cytosol of Hormone-dependent and Hormone-responsive GR Mouse Mammary Tumor Transplants
Specific r3H 1-Estradiol Binding Sites' ATP-activated Tumor Hormone Transplant
Line" Responseb Generation -ATP +ATP Binding Sites'
I10 HD 3 58 64 6 88 103 15 73 93 20 59 58 - 32 36 4 61 59 -
29 45 16 52 59 7
I20 120 - 125 119 -
1 1 1 HD 1 130 1 92 96 4
3 I07 HR 5 ad 68 -
" Tsl number is given.
' Expressed as fmolhg cytosol protein. Bold type: tumors with more than 10 fmol of
- I17 HD 2 9 7
Terms: HD, hormone-dependent passage; HR, hormone-responsive passage.
ATP-activated binding sites. Pool of three tumors.
Estradiol Stimulates the ATP-dependent Increase in Hormone Binding
The uterus tyrosine kinase confers the hormone binding capacity to uterine cytosol ER preinactivated by nuclear p h o s p h a t a ~ e , ~ ~ as well as to in uitro synthesized human ER, phosphorylating tyrosine 537.8-'a A very interesting property of the uterus kinase is the strong stimulation or dependence on estradiol.' Therefore, it was important to verify that the ATP-induced activation of ER binding sites is also stimulated by estradiol. In fact, this would support the belief that the enzyme present in the GR mammary tumor cytosol and involved in activation of ER is the same estradiol-stimulated kinase as previously detected in the
Five HI tumors (three belonging to passage 14 of line Tsl 107) and a pool of four HI tumors from lines Tsl 11 1 and 121 were selected to study this problem (TABLE 5). In the previously described experiments, an increase in hormone binding in the presence of ATP was detected when cytosol was preincubated with a physiological concentration of estradiol. Parallel incubations were performed omitting the preincubation with estradiol; this resulted in the disappearance of ATP-activated sites in five out of six tumors and in a lowered increase (from 50 to 22 fmol ATP-activated binding sites/mg cytosol protein) in the Tsl108 HI tumor (TABLE 5).

166 ANNALS NEW YORK ACADEMY OF SCIENCES
TABLE 4. Effect of ATP on Specific [3H]-Estradiol Binding in Cytosol of Hormone-independent GR Mammary Tumor Transplants
~~ ~~~ ~~~~ ~~~~~
Specific [3H]-Estrad~ol Binding Sites' ATP-activated Tumor Hormone Transplant
Line ' Responseb Generation - ATP +ATP Binding Sites' 108 HI 8 37d 121d 844 108 HI 14 24 74 50
32 56 24 13 66 53
112 HI 2' I25 HI 3' 42e 166 ' 124' 123 HI 1 I29 HI I 54d 77d 23d 132 HI 1 111 HI 13
HI IS 121 HI 19 66, 8Of 14f
HI 21 I07 HI 10 29 108 79
36 68 32 37 39 2 34 32 27 20
107 HI 18 21 19 18 19 1 14 14 I 1 16 S 17 16 17 17
37 28
- - - -
- -
106 HI 35 6 21 15 -
a Tsl number is given. HI: hormone-independent. Expressed as fmolimg cytosol protein. Bold type: tumors with more than 10 fmol of
ATP-activated binding sites. Pool of three tumors. ' Pool of two tumors. f pool of four tumors.
Western Blot of ER with Antiphosphotyrosine Antibody
A pool of two tumors, HI at passage 1 , was employed: tumor line Tsl 112 and 125. Their cytosol was incubated with or without ATP under the conditions used to assess ATP-dependent binding activation of ER. ATP induced a shift in hormone- binding receptor from 42 fmol/mg cytosol protein to 166 fmol/mg cytosol protein. The remainder of the same cytosol, previously incubated either with or without ATP, was divided into two aliquots, of which one was added to the anti-ER H222 antibody and the other was added to the control antibody. Both aliquots were subsequently added to anti-rat IgG, incubated in the presence of protein-A-Sepha- rose, and then centrifuged. The immunoprecipitated proteins were eluted from the pellets, submitted to SDS-PAGE, and finally blotted with antiphosphotyrosine
AURICCHIO er al.: PHOSPHORYLATION 167
antibodies (results shown in FIGURE 13). Each lane revealed two heavy protein bands migrating at 50 and 23 kDa, respectively. These correspond to the heavy and light chains of the antibodies. The sample incubated with ATP and then treated with anti-ER H222 antibody (lane H222, +ATP of FIGURE 13) showed a 67-kDa protein. The absence of this protein in the control sample (incubated with ATP, but treated with control antibody: control +ATP lane in FIGURE 13) indicated that the 67-kDa protein is the ER. Therefore, the large ATP-dependent increase in hormone binding to ER was associated with phosphorylation on tyrosine of the
4
3
k 2 m
1
0
x without ATP
with ATP
0 1 2 3 4
Specific eslradiol bound (nM)
FIGURE l2. Scatchard plot analysis of hormone binding to GR mouse mammary tumor ER before and after incubation with ATP. Cytosol was prepared from a pool of three hormone-independent tumors at passage 1 (Tsl 112, 123, 129) and aliquots of them were equilibrated for 2 h at 4°C with different concentrations of [3H]-estradiol in the presence or absence of a 100-fold excess of radioinert hormone. Samples were then incubated at 15°C for 10 min in the absence or presence of ATP (5 mM) as de~cribed.~ ' After incubation, samples were cooled at 2°C overnight and estradiol specific binding sites were measured by dextran-coated charcoal treatment. W: Samples incubated in the presence ofATP; x: samples incubated without ATP. The Kd value of the receptor for estradiol was 0.9 nM in the absence and presence of ATP. The [3H]-estradiol specific binding sites were 63 fmol/mg cytosol protein in the absence of ATP and 205 fmollmg cytosol protein in the presence of ATP.

a I- ?
cent ro I
a I- 7
H222
a a I- F ? 7
200 4
97 -I 69 -b
kER
47 -b
21 +
14.4 -+
FIGURE 13. Western blot with antiphosphotyrosine antibodies of GR mouse mamma tumor estradiol receptor. Cytosol prepared from GR mouse mammary tumor was incubati in the absence or presence of 5 mM ATP and then immunoprecipitated with anti-ER mon clonal H222 antibodies (H222 lanes) or nonimmune antibodies (control lanes) as reported The immunoprecipitated proteins were resolved on SDS-PAGE and then probed with an phosphotyrosine antibodies. In this experiment, a pool of two tumors, HI at passage (tumor lines Tsl 112 and Tsl 125). was used. The hormone binding activity was 42 fmol/n cytosol protein in the absence of ATP and 166 fmollmg cytosol protein in the presence ATP (see TABLE 4). The asterisk indicates the position of the 67-kDa ER in the H2 +ATP lane.

AURICCHIO ef al.: PHOSPHORYLATION 169
TABLE 5. Effect of Estradiol (E2) on the ATP-dependent Activation of Specific [3H]-Estradiol Binding Sites in GR Mouse Mammary Tumor Cytosol
Specific Estradiol ATP-activated Binding SitesC
-ATP +ATP Binding Sites' Tumor Hormone Transplant Line" Responseb Passage -E2 +E, -E2 +E, -E2 +E2 [(+EJ - (-Ell1
108 HI 14 24 24 46 14 22 50 28 35 32 26 56 - 24 24 10 13 11 66 1 53 52
107 HI 10 36 29 29 108 - 19 19 30 36 26 68 - 32 32
111 HI 13
121 HI 19 21
HI I5 ad 66d U d SOd -d 14d 14d
Tsl number is given.
Expressed as fmollmg cytosol protein. POOI of four tumors.
* HI: hormone-independent.
receptor itself. In the sample incubated with ATP, an unidentified phosphorylated protein migrating at 90-100 kDa was specifically immunoprecipitated, in associa- tion with 67-kDa ER, by H222 antibody.
CONCLUSIONS
ER is the first member of the nuclear receptor family whose tyrosine phosphory- lation has been observed in u ~ u o . ~ ' This phosphorylation occurs on tyrosine 537 of the ER1O.tt and confers hormone binding to the receptor.lO It is possible that other functions of the receptor are also associated with phosphorylation of this amino acid, which appears to confer an active conformation to ER.
Vanadate, a largely used phosphotyrosine-phosphatase inhibitor, induces the accumulation of tyrosine-phosphorylated ER and other proteins as well as the proliferation of MCF-7 cells in the absence of estradiol. Different antiestrogens prevent the accumulation of tyrosine phosphorylation of ER and other proteins, as well as the cell proliferation. These findings indicate that ER can be phosphory- lated and activated in the absence of estradiol and can stimulate proliferative tyrosine phosphorylation pathways. Tyrosine phosphorylation of ER in the ab- sence of hormone suggests a cross talk between (receptor-associated) tyrosine kinase(s) activated by vanadate and ER. It is also of interest that antiestrogens inhibit ER proliferative activity in the absence of hormone. NHBER can be con- verted to hormone-binding receptor through tyrosine phosphorylation by vanadate in MCF-7 cells and in mammary tumor cytosol by ATP in uitro.
Some hormone-independent mouse mammary tumors as well as human mam- mary tumors, both with low levels of hormone-binding ER, also contain large amounts of NHBER. This form of receptor probably results from a phosphoryla-

170 ANNALS NEW YORK ACADEMY OF SCIENCES
tion-dephosphorylation imbalance and could be responsible for the loss of hormone dependence as well as malignancy in some mammary tumors.
ACKNOWLEDGMENTS
We are indebted to D. Piccolo for technical assistance and to G. M. La Placa for editorial work.
1 .
2.
3. 4 .
5.
6 .
7.
8.
9.
10.
' I .
2.
13.
14.
15.
16. 17.
in .
REFERENCES
EVANS, R. M. 1988. The steroid and thyroid hormone receptor superfamily. Science
GREEN, S. & P. CHAMBON. 1988. Nuclear receptors enhance our understanding of
BEATO, M. 1989. Gene regulation by steroid hormones. Cell 56: 335-344. LIEBERHERR, M., B. GROSSE, P. DUCHAMBON & T. DRUEKE. 1989. A functional cell
surface type receptor is required for early action of I ,25-dihydroxyvitamin D3 on phosphoinositide metabolism in rat. J . Biol. Chem. 264: 20403-20406.
BLACKMORE. P. F., S. J . BEEBE, D. R. DANFORTH & N. ALEXANDER. 1990. Progester- one and 17-alpha-hydroxyprogesterone: novel stimulators of calcium influx in human sperm. J. Biol. Chem. 265: 1376-1380.
S C H U M A C H t R , M.. H. COIRINI, W. PFAFF & B. S. MCEWAN. 1990. Behavioral effects of progesterone associated with rapid modulation of oxytocin receptors. Science
AURICCHIO, F., A. MIGLIACCIO, M. DI DOMENICO & E. NOLA. 1987. Oestradiol stimulates tyrosine phosphorylation and hormone binding activity of its own receptor in a cell-free system. EMBO J. 6: 2923-2929.
MIGLIACCIO, A.. M. DI DOMENICO, S. GREEN, A. DE FALCO, E. L. KAJTANIAK, F. BLASI, P. CHAMBON & F. AURICCHIO. 1989. Phosphorylation on tyrosine of in uifro synthesized human estrogen receptor activates its hormone binding. Mol. Endocri- nol. 3: 1061-1069.
MIGLIACCIO, A., G. CASTORIA, A. DE FALCO, M. Dr DOMENICO, M. GALDIERO, E. NOLA, P. CHAMEON & F. AURICCHIO. 1990. In uirro phosphorylation and hormone binding activation of the synthetic wild-type human estradiol receptor. J. Steroid Biochem. 38: 407-413.
CASTORIA, G . , A. MIGLIACCIO, S. GREEN, M. DI DOMENICO, P. CHAMBON & F. AURICCHIO. 1993. Properties of a purified estradiol-dependent calf uterus tyrosine kinase. Biochemistry 32 1740-1750.
ARNOLD, S. F., J. D. OBOURN, H. JAFFE & A. C. NOTIDES. 1995. Phosphorylation of the human estrogen receptor on tyrosine 537 in uivo and by src family tyrosine kinases in v i m . Mol. Endocrinol. 9: 24-33.
EK, B. & C. H. ELDIN. 1982. Characterization of a tyrosine specific kinase activity in human fibroblast membranes stimulated by platelet derived growth factor. J. Biol. Chem. 257: 10486-10492.
KASUGA, M., F. A. KARLSSON & C . R. KAHN. 1Y82. Insulin stimulates thephosphoryla- tion of the 95,000 dalton subunit of its own receptor. Science 215: 185-187.
HUNTER, T. & J. A. COOPER. 1985. Protein tyrosine kinases. Annu. Rev. Biochem.
CARPENTER, G. & S. COHEN. 1990. Epidermal growth factor. J. Biol. Chem. 265:
BISHOP, J. M. 1985. Viral oncogenes. Cell 42: 23-38. YARDEN, Y. & A. ULLRICH. 1988. Growth factor receptor tyrosine kinases. Annu.
BERTHOIS, Y., J . A. KATZENELLENBOGEN & B. S. KATZENELLENBOCEN. 1986. Phenol
240 889-895.
transcription. Trends Genet. 4: 309-314.
2 5 0 69 1-693.
5 4 897-930.
7709-7712.
Rev. Biochem. 57: 443-478.

AURICCHIO e l ol.: PHOSPHORYLATION 171
19.
20 *
21.
22.
23.
24,
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
red in tissue culture media is a weak estrogen: implications concerning the study of estrogen-responsive cells in culture. Proc. Natl. Acad. Sci. U.S.A. 83: 2496- 2500.
FRACKELTON, A. R., M. POSNER, B. KANNAN & F. MERMELSTEIN. 1991. Gener- ation of monoclonal antibodies against phosphotyrosine and their use for affinity purification of phosphotyrosine-containing proteins. Methods Enzymol. 201:
WAKELING, A. E. & J. BOWLER. 1988. Biology and model of action of pure antioestro- gens. J. Steroid Biochem. 30: 141-147.
MAYER, B. J. & H. HANAFUSA. 1990. Association of v-crk oncogene product with phosphotyrosine-containing proteins and protein kinase activity. Proc. Natl. Acad. Sci. U.S.A. 87: 2638-2642.
LIPSICH, L. A., A. J. LEWIS & J. S. BRUGGE. 1983. Isolation of monoclonal antibodies that recognize the transforming protein of avian sarcoma virus. J. Virol. 48: 352- 360.
COOPER, J. A., F. S. ESCH, S. S. TAYLOR & T. HUNTER. 1984. Phosphorylation sites in enolase and lactate dehydrogenase utilized by tyrosine protein kinases in uiuo and in uitro. J. Biol. Chem. 259: 7835-7841.
SEFTON, B. M., T. HUNTER, K. BEEMON & W. ECKHART. 1980. Evidence that phos- phorylation of tyrosine is essential for cellular transformation by Rous sarcoma virus. Cell 2 0 807-816.
BEEMON, K., T. RYDEN & E. A. MCNELLY. 1982. Transformation by avian sarcoma viruses leads to phosphorylation of multiple cellular proteins on tyrosine residues. J. Virol. 4 2 742-747.
LEIS, J. F. & N. 0. KAPLAN. 1982. An acid phosphatase in the plasma membranes of human astrocytoma showing marked specificity toward phosphotyrosine protein. Proc. Natl. Acad. Sci. U.S.A. 7 9 6507-651 1.
SWARUP, G., S. COHEN & D. L. GARBERS. 1982. Inhibition of membrane phospho- tyrosyl-phosphatase by vanadate. Biochem. Biophys. Res. Commun. 1W: 1104- 1109.
NELSON, R. L. & P. E. BRANTON. 1984. Identification, purification, and characteriza- tion of phosphotyrosine-specific protein phosphatases from cultured chicken embryo fibroblast. Mol. Cell. Biol. 4: 1003-1012.
RAMASARMA, T. & F. L. CRANE. 1981. Does vanadium play a role in cellular regulation? Curr. Top. Cell. Regul.. 2 0 247-301.
KLARLUND, J. K. 1985. Transformation of cells by an inhibitor of phosphatases acting on phosphotyrosine in proteins. Cell 41: 707-717.
MIGLIACCIO, A., A. ROTONDI & F. AURICCHIO. 1986. Estradiol receptor: phosphoryla- tion on tyrosine in uterus and interaction with antiphosphotyrosine antibodies.
AURICCHIO, F., M. DI DOMENICO, A. MIGLIACCIO, G . CASTORIA & A. BILANCIO. 1995. The role of estradiol receptor in the proliferative activity of vanadate on MCF-7 cells. Cell Growth Differ. 6 105-113.
RAO, K. V. & C. F. Fox. 1987. Epidermal growth factor stimulates tyrosine phosphory- lation of human glucocorticoid receptor in cultured cells. Biochem. Biophys. Res. Commun. 144: 512-519.
L I N , K. H., K. ASHIZAWA & S. Y. CHENG. 1992. Phosphorylation stimulates the transcriptional activity of the human p-1 thyroid nuclear receptor. Proc. Natl. Acad. Sci. U.S.A. 89: 7737-7741.
ROCHETTE-EGLY, C., M. P. GAUB, Y. LUTZ, S. ALI, I. SHEUER & P. CHAMBON. 1992. Retinoic acid receptor-p: immunodetection and phosphorylation on tyrosine residues. Mol. Endocrinol. 6: 2197-2209.
SLUYSER, M. & R. VAN NIL 1974. Estrogen receptor content and hormone-responsive growth of mouse mammary tumors. Cancer Res. 34: 3253-3257.
SLUYSER, M., S. G. EVERS & R. VAN NIE. 1976. Sex hormone receptors in mammary tumors of GR mice. Nature 263: 386-389.
MIGLIACCIO, A., A. ROTONDI & F. AURICCHIO. 1984. Calmodulin-stimulated phosphor-
79-92.
EMBO J. 5: 2867-2872.

172 ANNALS NEW YORK ACADEMY OF SCIENCES
ylation of 17p-estradiol receptor on tyrosine. Proc. Natl. Acad. Sci. U.S.A. 81:
39. AURICCHIO, F., A. MIGLIACCIO, G. CASTORIA, A. ROTONDI & M. DI DOMENICO. 1987. Calmodulin-stimulated estradiol receptor-tyrosine kinase. Methods Enzymol.
40. MIGLIACCIO, A., M. PAGANO & F. AURICCHIO. 1993. Immediate and transient stimula- tion of protein tyrosine phosphorylation by estradiol in MCF-7 cells. Oncogene
41. MIGLIACCIO, A., M. PAGANO, C. C. J. DE Gowi, M. DI DOMENICO, G. CASTORIA, M. SLUYSER & F. AURICCHIO. 1992. Phosphorylation and estradiol binding of estrogen receptor in hormone-dependent and hormone-independent GR mouse mammary tu- mors. Int. J . Cancer 51: 733-739.
5921-5925.
139 731-744.
8: 2183-2191.





























