Embed Size (px)
Citation preview

f1
a
A
RU
Dbd
wtDebaspsop
EAt(nctSs
Neuroscience 195 (2011) 112–127
FORM OF DUAL-SPECIFICITY TYROSINE-(Y)-PHOSPHORYLATION-REGULATED KINASE 1A NONPHOSPHORYLATED AT TYROSINE 145AND 147 IS ENRICHED IN THE NUCLEI OF ASTROGLIAL CELLS,ADULT HIPPOCAMPAL PROGENITORS, AND SOME CHOLINERGIC
AXON TERMINALSir
Kp
E. KIDA,a M. WALUS,a K. JARZABEK,b,c
S. PALMINIELLO,b G. ALBERTINI,b A. RABE,a
Y. W. HWANGd AND A. A. GOLABEKa*aDepartment of Developmental Neurobiology, New York State Instituteor Basic Research in Developmental Disabilities, Staten Island, NY0314, USA
bChild Development Department, IRCCS San Raffaele Pisana, Romend San Raffaele Cassino, Italy
cDepartment of Biology and Pathology of Human Reproduction, Polishcademy of Sciences, 15-276 Bialystok, Poland
dDepartment of Molecular Biology, New York State Institute for Basicesearch in Developmental Disabilities, Staten Island, NY 10314,SA
Abstract—Compelling lines of evidence indicate that overex-pression of dual-specificity tyrosine-(Y)-phosphorylation reg-ulated kinase 1A (DYRK1A) in subjects with trisomy 21 (Downsyndrome[DS]) contributes to the abnormal structure andfunction of the DS brain. In the present study, we used anovel, phospho-dependent antibody recognizing DYRK1Aonly with nonphosphorylated tyrosine 145 and 147 (DYRK1ATyr-145/147P�), to investigate the expression pattern of this
YRK1A species in trisomic and disomic human and mouserains. Immunoblotting and dephosphorylation experimentsemonstrated higher levels of DYRK1A Tyr-145/147P� in
postnatal trisomic brains in comparison with controls (by�40%) than those of the DYRK1A visualized by three other N-and C-terminally directed antibodies to DYRK1A. By immu-nofluorescence, the immunoreactivity to DYRK1A Tyr-145/147P� was the strongest in the nuclei of astroglial cells,
hich contrasted with the predominantly neuronal localiza-ion of DYRK1A visualized by the three other antibodies toYRK1A we used. In addition, DYRK1A Tyr-145/147P� wasnriched in the nuclei of neuronal progenitors and newlyorn neurons in the adult hippocampal proliferative zone andlso occurred in some cholinergic axonal terminals. Our datahow a distinctive expression pattern of DYRK1A forms non-hosphorylated at Tyr-145 and Tyr-147 in the brain tissue anduggest that DS subjects may exhibit not only upregulationf total DYRK1A, but also more subtle differences in phos-horylation levels of this kinase in comparison with control
*Corresponding author. Tel: �1-718-494-5208; fax: �1-718-982-6346.-mail address: [email protected] (A. A. Golabek).bbreviations: BrdU, 5-bromo-2-deoxyuridine; ChAT, choline acetyl-
ransferase; DS, Down syndrome; DYRK1A, dual-specificity tyrosine-Y)-phosphorylation-regulated kinase 1A; ECL, enhanced chemilumi-escence; ELISA, enzyme-linked immunosorbent assay; FCS, fetalalf serum; GFAP, glial fibrillary acidic protein; mAb, monoclonal an-ibody; pAbs, polyclonal antibodies; PBS, phosphate buffered saline;
DS-PAGE, sodium dodecylsulfate polyacrylamide gel electrophore-is; wt, wild type.0306-4522/11 $ - see front matter © 2011 IBRO. Published by Elsevier Ltd. All righdoi:10.1016/j.neuroscience.2011.08.028
112
ndividuals. © 2011 IBRO. Published by Elsevier Ltd. Allights reserved.
ey words: Down syndrome, Ts65Dn mice, DYRK1A, phos-horylation, brain tissue.
Down syndrome (DS), a chromosome 21 (HSA21) trisomy,is the most common, noninheritable genetic cause of men-tal retardation (Epstein, 1986). DS is a complex disorderwith highly variable phenotypes. However, mental retarda-tion with subnormal intellectual functioning is its most con-sistent feature (Pennington et al., 2003). DS brain demon-strates a number of morphologic changes such as hypo-cellularity, delayed myelination, dendritic and synapticalterations, and abnormal neurogenesis (Wisniewski et al.,2006). The cause(s) for the abnormal structure and func-tion of the DS brain on the molecular level is still unclear.
Earlier studies proposed that a small region of HSA21qspanning 3.8–6.5 Mb, the so-called DS critical region, mayplay a major role in DS phenotypes (Korenberg et al.,1990; Delabar et al., 1993). Recent high-resolution map-ping of rare, partial trisomies of HSA21 has provided evi-dence that a single DS critical region being responsible forall or most DS features does not exist (Lyle et al., 2009;Korbel et al., 2009). The presence of a phenotype suscep-tibility region for a given DS feature modified by other locion HSA21 and elsewhere in the genome has been pro-posed instead (Lyle et al., 2009). Accordingly, the mentalretardation of individuals with DS was ascribed to twodifferent genomic regions of HSA21, one of them located inthe telomeric, and the other in the centromeric region (Lyleet al., 2009; Korbel et al., 2009).
One candidate protein that may contribute to the ab-normal structure and function of DS brain is DYRK1A (fordual-specificity tyrosine-(Y)-phosphorylation-regulated ki-nase 1A [EC 2.7.12.1]). DYRK1A is encoded in humans bya gene mapped to the 21q22.13 locus of HSA21 (Guimeráet al., 1996), and to the triplicated segment of Mmu16 inTs65Dn mice (Song et al., 1996), a widely used mousemodel for DS (Davisson et al., 1990), which replicatessome behavioral, cellular, and molecular abnormalities ofthe human disease (Holtzman et al., 1996; Rueda et al.,2005).
DYRK1A is a proline-directed serine/threonine and ty-rosine kinase (Kentrup et al., 1996; Becker et al., 1998;
Himpel et al., 2001). In mammals, DYRK1A is widely ex-ts reserved.
bbintopeom2ocDt
etfiia1ss1eitlmc6(
anWrfgtijDle
fur1
m
ta
mtv
E. Kida et al. / Neuroscience 195 (2011) 112–127 113
pressed in internal organs and the brain with the highestlevels in the prenatal periods (Guimera et al., 1999; Okui etal., 1999). The potential involvement of DYRK1A in braindevelopment suggested observations that mutations inDYRK1A orthologue, the Drosophila melanogaster mini-rain gene (mnb), cause an abnormal spacing of neuro-lasts in the outer proliferation center of larval brain, lead-
ng to the production of less neuronal progeny by theeuroblasts than do wild-type (wt) flies and marked reduc-ion in the size of the optic lobes and central hemispheresf the adult brain (Tejedor et al., 1995). However, it ap-ears that both an increase and decrease in DYRK1Axpression level are detrimental given that overexpressionf DYRK1A in mice causes neurodevelopmental delay withotor abnormalities and cognitive deficits (Altafaj et al.,001), and its haploinsufficiency is associated with devel-pmental delay and abnormal brain morphology with mi-rocephaly (Fotaki et al., 2002). In humans, truncation ofYRK1A gene leads to microcephaly and mental retarda-
ion (Møller et al., 2008).Alternative splicing of DYRK1A gene generates sev-
ral transcript variants differing from each other either inhe 5= UTR or in the 3= coding region and encoding at leastve different isoforms including the most prevalent, longsoform, with a predicted length of 763 amino acids (aa)nd a molecular weigh (Mw) of 85.6 kDa (Kentrup et al.,996; Guimera et al., 1999). The DYRK1A molecule haseveral functional domains. Bipartite nuclear translocationignals in the N-terminus and the linker region (aa 117–34) govern the nuclear localization of the protein (Beckert al., 1998), whereas a C-terminally located motif that
ncludes a histidine stretch determines its splicing specklesargeting (Álvarez et al., 2003). In addition, DYRK1A has aong protein kinase domain (aa 159–479), a leucine zipper
otif (aa 274–295), PEST domain (aa 501–520), a highlyonservative 13-consecutive-histidine repeat (aa 607–19), and a serine/threonine-rich domain (aa 656–672)Kentrup et al., 1996; Guimera et al., 1999).
In brain tissue, DYRK1A was localized to both cell nucleind extra-nuclear cell compartments of neurons predomi-antly (Hämmerle et al., 2003, 2008; Martí et al., 2003;egiel et al., 2004). DYRK1A phosphorylates in vitro a wide
ange of proteins, for example, protein-synthesis initiationactor eIF2B�, nuclear factor of activated T cells (NFAT),lycogen synthase, the microtubule-associated protein
au, the cAMP-response element-binding protein, the splic-ng factor SF3b1, caspase–9, Notch, dynamin 1, synapto-anin I, and amphiphysin 1, pointing to a pleiotropic role forYRK1A, including signaling pathways regulating cell pro-
iferation and brain development (for reviews see Wisemant al., 2009; Park et al., 2009).
Increases in DYRK1A levels by �1.5-fold have beenound previously in DS brain (Dowjat et al., 2007). Here, bysing a phospho-dependent monoclonal antibody (mAb)ecognizing DYRK1A only with nonphosphorylated Tyr-45 and Tyr-147 (DYRK1A Tyr-145/147P�), we report an
even higher increase in the levels of DYRK1A Tyr-145/147P� in the postnatal brain of Ts65Dn mice, a mouse
odel for DS, and DS subjects in comparison with con-
rols, and we present data showing a distinctive cellularnd subcellular distribution of this DYRK1A species.
EXPERIMENTAL PROCEDURES
Human tissues
For immunoblotting analyses, postmortem, frozen brain tissuesections from the frontal lobes from five control (at 19–28 wgs)and three DS fetuses (at 18–22 weeks of gestation [wgs]) andfrom four DS and four control subjects from 8 to 23 years of agewere provided by the Brain and Tissue Bank for DevelopmentalDisorders at the University of Maryland, Baltimore, MD. The post-mortem interval was from 1 to 4 h for fetuses and from 5 to 28 hfor older subjects. The diagnosis of DS was based on karyotypingand discovery of an extra chromosome 21. Genetic tests were notperformed in control cases except for the fetuses, which showednormal chromosome complement. The most common cause ofdeath was cardiorespiratory insufficiency, cardiac arrhythmia, oracute respiratory arrest. The brains included in the study were freeof focal changes and inflammatory processes. The study wasapproved by the IBR Institutional Review Board.
Mouse tissues
Female B6EiC3 Ts65Dn and male C57BL/6JEi x C3SnHeSnJ(B6EiC3) mice obtained from the Jackson Laboratory (Bar Harbor,ME, USA) were used to establish the Ts65Dn mouse colony byone of us (A.R.). Animal breeding and all the following experi-ments were conducted in accordance with the protocol approvedby the Institutional Animal Welfare Committee. All animal experi-ments were carried out in accordance with the National Institute ofHealth Guide for the Care and Use of Laboratory Animals. Allefforts were made to minimize the number of animals used andtheir sufferings.
Females at ages 4–8 months were used for the presentstudy. Animals were weighed and deeply anesthetized with Nem-butal (75 mg/kg i.p.) (Abbott Laboratories, Abbott Park, IL, USA)and transcardially perfused for 10 min with 0.1 M phosphate-buffered saline (PBS), pH 7.4 (50–100 ml) (for immunoblottingexperiments), or 4% paraformaldehyde (for immunofluorescence).After perfusion, the brain was removed and one hemisphere wasimmediately snap-frozen and kept at �80 °C until future analyses.To eliminate the inter-individual differences associated with dis-section of small and thus potentially not always identical frag-ments from brain areas, homogenates were prepared from thewhole cerebral hemispheres. The other hemisphere was fixed in4% paraformaldehyde solution, pH 7.4 for 72 h, and thereafterwas processed for immunofluorescence study. Altogether, braintissue from 42 mice was used for the experiments describedbelow.
All mice were genotyped toward trisomy 16 and retinopathyby using PCR-based amplification of DNA, exactly as we de-scribed previously (Palminiello et al., 2008). Although we alsogenotyped these animals for the presence of rd1 mutation in a rodphotoreceptor cyclic GMP phosphodiesterase �-subunit gene(Pde6brd1), producing retinopathy and blindness in affected ho-
ozygotes, both wt, heterozygotes and homozygotes for rd1 mu-ation were included, because we had no reason to suspect thatision loss might affect the levels of DYRK1A.
Production and characterization of mAb 8D9
mAb 8D9 anti-DYRK1A was produced in the IBR MonoclonalAntibody Facility by one of us (Y. W. H). Mice were immunizedusing the 6�histidine-tagged polypeptide containing the first 160residues of rat DYRK1A that was purified from expressing E. colistrain BL21, as described for another mAb anti-DYRK1A (Wegiel
et al., 2004). mAb 8D9 was either batch-purified from ascites fluid
Elltd
lbAsotala
pACCasf8
p
E. Kida et al. / Neuroscience 195 (2011) 112–127114
using Bakerbond Abx (J.T. Baker, Phillipsburg, NJ, USA) or wasemployed as hybridoma supernatant. The epitope of mAb 8D9was mapped by an M13 phage display system (Ph.D.-7 PhageDisplay Peptide Library Kit, New England Biolabs, Ipswich, MA,USA), as described (Miller et al., 2003).
Titration of mAb 8D9 against the synthetic peptides corre-sponding to amino acid residues 141–150 of human DYRK1Asequence Ac–DDDNpYDYIVKC (pepTyr-145P�), Ac-DDDNYD-pYIVKC (pepTyr-147P�), or Ac–DDDNYDYIVKC (pep-P�) (New
ngland Peptide, Gardner, MA, USA) was done by using enzyme-inked immunosorbent assay (ELISA) (cysteine was added at theast position to facilitate conjugation). Wells of polystyrene microti-ration plates (Immulon 2; Dynatech Laboratories, Inc., Alexan-ria, VA, USA) were coated with 0.1 ml of peptide solution (2 �g
of protein per well in carbonate/bicarbonate buffer [pH 9.6]) bypassive adsorption for 2 h at room temperature. Coated plateswere washed three times with PBS, blocked in 1% nonfat drymilk/PBS solution, pH 7.4, for 1 h at room temperature andwashed three times in 0.05% Tween 20 in PBS (PBST). Serialtwofold dilutions of the antibody were then made in the wells.Plates were incubated overnight at 4 °C and washed with PBST.Alkaline-phosphatase conjugated anti-mouse IgG (Thermo FisherScientific, Fremont, CA, USA) diluted 1:2500 in PBST was addedto each well, and the plates were incubated for 1 h at roomtemperature. After washing, p-nitrophenyl phosphate (substrate)solution in diethanolamine buffer was added at 0.1 ml per well.The enzyme–substrate reaction was allowed to proceed at roomtemperature for 20 min, and afterward it was quenched with 0.1 mlof 0.2 M NaOH, and the optical density at 410 nm was measuredon a plate reader (GENios Tecan, USA, Research Triangle Park,NC, USA).
Injections of 5-bromo-2-deoxyuridine
The mice received two intraperitoneal injections of 5-bromo-2-deoxyuridine (BrdU) (Sigma-Aldrich, St. Louis, MO, USA) at 100mg/kg of weight diluted in 0.9% NaCl/0.007 M NaOH (in a 200-�lvolume) within 4-h intervals and were killed 6 h after the firstinjection. Animals were perfused with 4% paraformaldehyde, andthe brains were postfixed for 24–48 h and cut on vibratome(Bannockburn, IL, USA) at 40-�m sections. The sections werekept until further use in PBS with 0.05% sodium azide at 4 °C.
SDS-PAGE and Western blotting
Brain tissues were homogenized in a buffer containing 50 mMTris, pH 7.4; 1% SDS; and protease inhibitor cocktail (Complete,Roche, Indianapolis, IN, USA) supplemented with pepstatin A andphosphatase inhibitor cocktail (PhosSTOP, Roche); and then son-icated and centrifuged at 15,000�g for 5 min. The protein contentwas measured by using a Coomassie Protein Assay and bovineserum albumin (BSA) as a standard (Thermo Fisher Scientific).
Twenty micrograms of protein per lane was loaded onto 10%Tris/Tricine gels, electrophoretically separated, electrotransferredonto nitrocellulose membranes and developed by using the en-hanced chemiluminescence method (ECL), as described (Gola-bek et al., 2003). The primary antibodies were mAb 8D9 (IBR),diluted 1:2000 (purified ascites fluid) or 1:400 (hybridoma super-natant); mAb 7D10, raised in mouse to 674–763 aa of DYRK1Apolypeptide fused to glutathione S-transferase tag (Novus Biolog-icals, Littleton, CO, USA), diluted 1:10,000; polyclonal antibodies(pAbs) anti-N-terminal DYRK1A raised in rabbit to 32–51 aa ofhuman DYRK1A conjugated to keyhole limpet hemocyanin (Sig-ma-Aldrich), diluted 1:10,000 and pAbs H-143 raised against theC-terminal fragment of DYRK1A, including aa residues 621–763(Santa Cruz Biotechnology, Santa Cruz, CA, USA), diluted 1:200.The secondary, species-specific, horseradish peroxidase (HRP)-linked antibodies (GE Healthcare, Piscataway, NJ, USA) were
diluted 1:5000. To adjust for equal protein load, immunoblots werestripped and reprobed with mAbs to �-actin (Sigma-Aldrich), di-uted 1:100,000. Densitometry analysis of immunoblots was doney 1DScan EX software (Scanalytics Inc., Fairfax, VA, USA).ppropriate exposures that fell within the linear range of antibodytaining were selected for scanning. Bands were scanned, and theptical density of the areas under the bands was determined. Thewo-tailed Student’s t-test for independent samples was used tossess the statistical significance of data. P�0.05 represented the
evel of significance. Preincubation of mAb 8D9 with an excess ofdequate synthetic peptides (20 �g/ml), either pepTyr-145YP� or
pep-P�, served as a control of mAb 8D9 specificity.
Slot blots
Synthetic DYRK1A peptides were diluted in PBS, pH 7.4, andapplied in an amount ranging from 0.003 to 10 �g to nitrocellulosemembrane in slot blot apparatus (Bio-Rad Laboratories, Hercules,CA, USA). The membrane was air-dried, blocked in 5% BSA inPBS, incubated with mAb 8D9 at 1:2000 overnight at 4 °C,washed five times with PBS/0.05% Tween20, incubated with sec-ondary antibody conjugated with HRP (GE Healthcare) diluted at1:5000, washed and developed by using ECL reagents, as de-scribed previously.
Dephosphorylation studies
Dephosphorylation of mouse brain tissues. Brain tissuesfrom three Ts65Dn and three wt mice were homogenized in abuffer containing 50 mM Tris, pH 7.4, and protease inhibitorcocktail EDTA-free (Complete, Roche), and then sonicated andcentrifuged at 15,000�g for 5 min. Samples containing 30 �g ofrotein were incubated either in the presence of 2 U of Shrimplkaline Phosphatase (SAP) in SAP Reaction buffer (USB Corp.,leveland, OH, USA) supplemented with 1 mM Mn2� or 15 U ofalf Intestine Phosphatase (CIP) (New England Biolabs), both in15 �l volume for 30 min at 37 °C, then boiled for 5 min in Tricine
ample buffer, resolved on 10% Tris/Tricine gels, electrotrans-erred onto nitrocellulose membranes and developed with mAbD9 by using ECL method, as described previously.
Dephosphorylation of synthetic peptides. One microgram ofepTyr-145P� and pep-P� was incubated for 30 min at 37 °C
either in the presence of CIP (New England Biolabs), SAP (USBCorp.), or Antarctic Phosphatase (New England Biolabs) in Reac-tion Buffer provided by the manufacturers or in Reaction Bufferwithout the addition of phosphatases (mock-treated). In a sepa-rate set of experiments, 200 ng of pepTyr-145P�, pepTyr-147P�
and pep-P� was incubated with SAP, as described previously.Then, the samples were loaded onto nitrocellulose membrane inthe slot blot apparatus (Bio-Rad Laboratories), air-dried and de-veloped with mAb 8D9, as described previously for slot blots.
Immunofluorescence
Immunofluorescence was done on 40-�m free-floating Vibratomesections from mouse brains. Two different detergents—0.1%–0.3% Triton X-100 and 0.1%–0.3% saponin—were used for tissuepermeabilization, incubation with antibodies and washes. Formost double-labeling experiments, sections were heated at 80 °Cfor 30 min in 10 mM citrate buffer, pH 8.5. For BrdU detection,sections were first incubated in 50% formamide in saline-sodiumcitrate (SSC) for 30 min at 65 °C, washed twice in SSC, thenincubated in 2N HCl for 30 min at 37 °C, neutralized in 0.1 M boricacid, pH 8.5, and washed in PBS.
After antigen retrieval, as described previously, sections wereblocked for 4 h at room temperature in 10% fetal calf serum(FCS)/PBS with a detergent, as described previously, then incu-bated for 24–48 h at 4 °C with primary antibodies. The followingprimary antibodies were used: pAbs to glial fibrillary acidic protein
(GFAP) (from rabbit, DAKO, Carpinteria, CA, USA; 1:400), pAbs
AB(b1Nrsc(dw(mUtaq
a
sbp
tneop8th
lpT
ilDrbtiotwd
aatwb1pm3tsd1fpd
Tni
ApsTspatoDpe(r
mpt8pw
E. Kida et al. / Neuroscience 195 (2011) 112–127 115
to doublecortin (from goat, Santa Cruz Biotechnology, at 1:1000),mAb to BrdU (from rat, Accurate Chemical & Scientific Corp,Westbury, NY, USA, at 1:30), pAbs to choline acetyltransferase(ChAT) (from goat, Affinity BioReagents, Golden, CO, USA, di-luted 1:150), pAbs to synaptophysin (from rabbit, DAKO, at1:200), pAbs to GAD65/67 (from rabbit, Sigma-Aldrich, diluted1:1000), pAbs to �-aminobutyric acid (GABA) (from rabbit, Sigma-
ldrich, 1:5000), pAbs to parvalbumin (from rabbit, from AffinityioReagents, diluted 1:1000), and pAbs to dopamine transporter
from rat, Chemicon, Temecula, CA, USA, diluted 1:5000). Anti-odies to DYRK1A were used at the following dilutions: mAb 8D9:50–1:200 (hybridoma supernatant), mAb7D10 1:200, pAbs anti--terminal DYRK1A 1:500 and pAbs H-143 1:200. After several
inses, sections were incubated for 4 h at room temperature withpecies-specific secondary antibodies conjugated with fluores-ent dyes (Invitrogen, Carlsbad, CA, USA): Alexa Fluor 488green color), diluted at 1:1000 and/or Alexa Fluor 555 (red color),iluted at 1:2000, either raised in goat or rabbit. Sections wereashed 4�10 min, mounted on Superfrost/Plus microscope slides
Thermo Fisher Scientific, Fremont, CA, USA), coverslipped withounting medium (Vector Laboratories Inc., Burlingame, CA,SA), and viewed with a Nikon C1 laser-scanning confocal sys-
em mounted on Nikon 90i microscope. Omission of the primaryntibodies and preincubation of mAb 8D9 with an excess of ade-uate synthetic peptides (20 �g/ml), either pepTyr-145P� or pep-
P�, served as a control of mAb 8D9 specificity.Z-stack major projections were generated by collecting im-
ges at 0.4-�m steps along the Z-axis. Optical images wereprocessed by using Adobe Photoshop 6.0. The number of mAb8D9-positive astroglial nuclei in the neocortex and the stratumradiatum and lacunosum/moleculare of the CA1 hippocampal sec-tor and the stratum moleculare of the hippocampal dentate gyruscovering a tissue area of approximately 1.56 mm2 were countedfrom four sections per region in each of the three trisomic andthree wt mice on optical images from the Z-stack major projectionsby using ImageJ software (National Institutes of Health, Bethesda,MD, USA).
RESULTS
Characterization of mAb 8D9
By using an M13 phage display system, we localized theepitope of mAb 8D9 to the aa residues 142–147 (DDNYDY)of human DYRK1A sequence. This aa sequence is highlyconserved in vertebrates from Zebra fish to humans. Inminibrain kinase—DYRK1A orthologue in Drosophilamelanogaster—the first tyrosine residue in the sequenceDDNYDY is replaced by the histidine (NCBI databases).Because there are two potential phosphorylation siteswithin the epitope (Tyr-145 and Tyr-147), next we usedsynthetic decapeptides with eitherTyr-145 or Tyr-147phosphorylated (pepTyr-145P� and pepTyr-147P�, re-pectively) or nonphosphorylated (pep-P�) to test by slotlots and ELISA the possibility that mAb 8D9 could behospho-dependent.
Slot blot analysis (Fig. 1a) and ELISA (Fig. 1b) showedhat mAb 8D9 recognized DYRK1A decapeptide only inonphosphorylated form. Decapeptides phosphorylatedither at tyrosine 145 or 147 were detectable by mAb 8D9nly after their prior dephosphorylation with the use ofhosphatases (Fig. 1a). Also, only preincubation of mAbD9 with nonphosphorylated decapeptide abolished de-ection of DYRK1A on immunoblots of the brain tissue
omogenates by mAb 8D9 (Fig. 1c, compare lane 2 and tane 3). All these findings indicated that mAb 8D9 is phos-ho-dependent, as it recognizes DYRK1A only when itsyr-145 and Tyr-147 are not phosphorylated.
In this respect, detection of DYRK1A by mAb 8D9 onmmunoblots of mouse brain tissue homogenates (Fig. 1c,ane 1) suggested that either a fraction or the whole pool ofYRK1A in the brain might be not phosphorylated at ty-
osine 145 and 147. To investigate this issue, we incu-ated mouse brain tissue samples with alkaline phospha-ases (CIP and SAP) and analyzed by densitometry thentensity of DYRK1A-specific bands detected by mAb 8D9n immunoblots in samples dephosphorylated and mock-reated. Dephosphorylation of brain tissue homogenatesith CIP (Fig. 1d) and SAP (not shown) enhanced theetection of DYRK1A by mAb 8D9 from �50% to �80% on
immunoblots of brain homogenates from disomic mice andup to �25% from trisomic animals (Fig. 1d, upper panelnd diagram on the right). These calculated values arepproximate only because full dephosphorylation of theyrosine residues in the brain samples was not possible,hich was evidenced after immunostaining of the mem-ranes with antibodies specific to phosphotyrosine (Fig.d, the lowest panel). Exposure of the tissue to alkalinehosphatases for longer than 30 min under the recom-ended experimental conditions (incubation temperature7 °C in the absence of EDTA and inhibitors of phospha-ases) led to significant degradation of DYRK1A in theamples (not shown). Nevertheless, these experimentsemonstrated the presence of DYRK1A species with Tyr-45/147 in both phosphorylated and nonphosphorylatedorm in the brain tissue and suggested that the level ofhosphorylation of Tyr-145/147 is more pronounced inisomic than trisomic brain.
he levels of DYRK1A with Tyr-145/147onphosphorylated are higher in Ts65Dn mice than
n disomic animals
ntibodies specific to DYRK1A with Tyr-145/147 phos-horylated are unavailable at present. Therefore, to furtherupport our data suggesting that DYRK1A species withyr-145/147 nonphosphorylated are more abundant in tri-omic than disomic brain, we compared the levels and theattern of DYRK1A visualized on immunoblots of disomicnd trisomic mouse brain homogenates by mAb 8D9 andhree commercially available antibodies with epitopes non-verlapping with that of mAb 8D9, thereby detectingYRK1A species in both phosphorylated and nonphos-horylated form at Tyr-145/147. Two of the antibodies wemployed were raised to the C-terminal region of DYRK1AmAb 7D10 and pAbs H-143), and one to the N-terminalegion of DYRK1A (Fig. 2a).
As shown in Fig. 2b (upper panel), on immunoblots ofouse brain homogenates, mAb 8D9, mAb 7D10, andAbs H-143 visualize a similar pattern of DYRK1A migra-ion with a strong doublet with an apparent Mw �94- and7-kDa and a weaker band at the level of �80 kDa. Thisattern of DYRK1A migration on SDS-PAGE is in agreementith earlier published studies employing antibodies to the C-
erminal region of DYRK1A (Okui et al., 1999). Interestingly,

d(iDt3i
E. Kida et al. / Neuroscience 195 (2011) 112–127116
Fig. 1. mAb 8D9 recognizes DYRK1A only when Tyr-145 and Tyr-147 are not phosphorylated. (a) On slot blot, mAb 8D9 detects DYRK1Aecapeptide with Tyr-145 and Tyr-147 nonphosphorylated (pep-P�), but it does not recognize DYRK1A decapeptide with Tyr-145 phosphorylatedpepTyr-145P�) (left and right panel) and Tyr-147 phosphorylated (pepTyr-147P�) (right panel) unless it is dephosphorylated after incubation with calfntestine alkaline phosphatase (CIP), shrimp alkaline phosphatase (SAP) or Antarctic Phosphatase (ant). (b) Titering of mAb 8D9 by ELISA. (c)YRK1A decapeptide nonphosphorylated (lane 2) abolishes detection of DYRK1A by mAb 8D9 in the mouse brain tissue homogenates. Please note
hat detection of DYRK1A in the mouse brain tissue by mAb 8D9 (lane 1) is not hampered by DYRK1A decapeptide with Tyr-145 phosphorylated (lane). (d) The amounts of DYRK1A detected by mAb 8D9 are higher on immunoblots of the trisomic and disomic mouse brain homogenates after
ncubation with calf intestine alkaline pbosphatase (CIP) (upper left panel and diagram). The middle panel demonstrates the blot reprobed with �-actinantibody to show the protein load. The immunoblot presented in the lower left panel was probed with antibody specific to phosphotyrosine to assessthe level of tissue dephosphorylation by CIP. Please note that after dephosphorylation, the levels of DYRK1A detected by mAb 8D9 show a higherincrease in disomic than trisomic brain, indicating that Tyr-145/147 phosphorylation is more prominent in disomic than trisomic brain (right diagram).
Wt, wild-type; Ts, trisomic.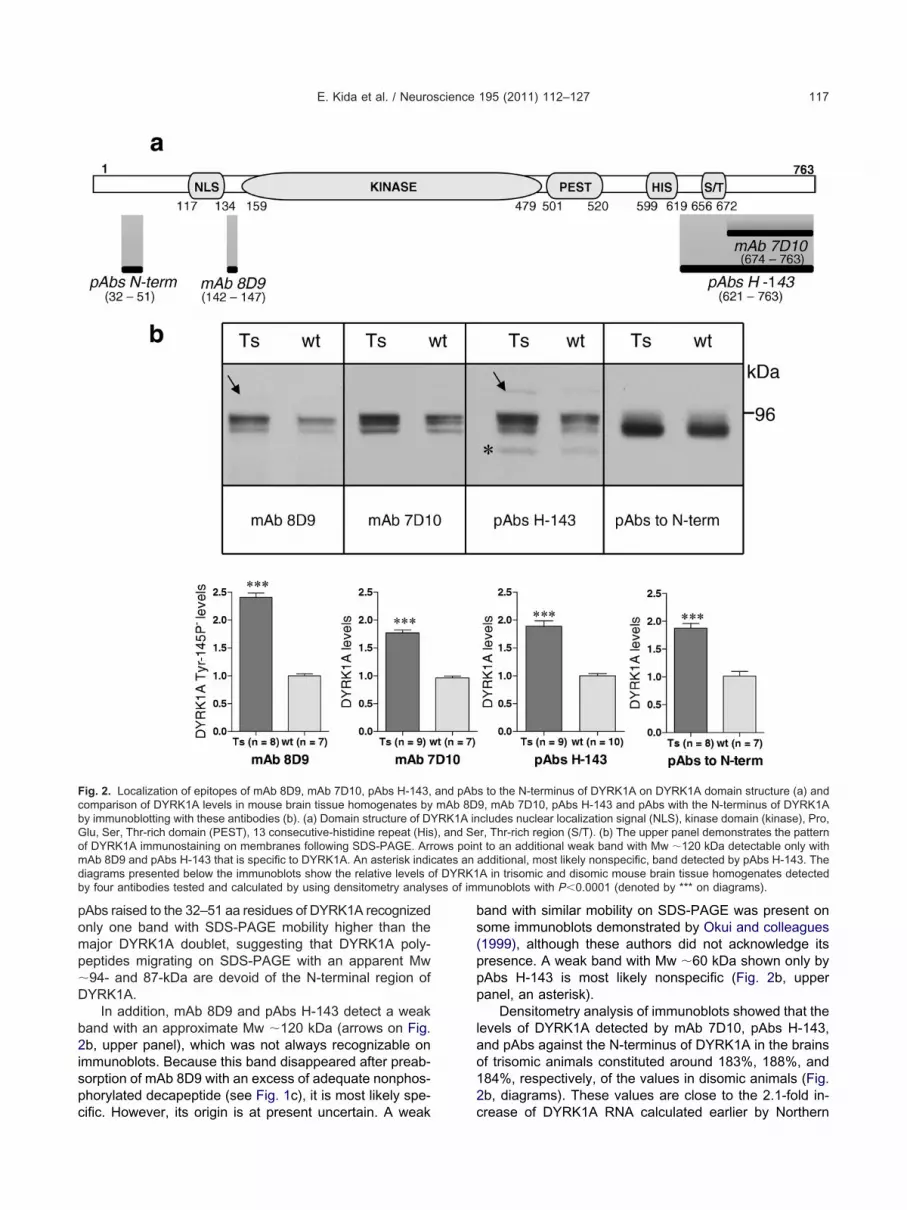
f DYRK1es of im
E. Kida et al. / Neuroscience 195 (2011) 112–127 117
pAbs raised to the 32–51 aa residues of DYRK1A recognizedonly one band with SDS-PAGE mobility higher than themajor DYRK1A doublet, suggesting that DYRK1A poly-peptides migrating on SDS-PAGE with an apparent Mw�94- and 87-kDa are devoid of the N-terminal region ofDYRK1A.
In addition, mAb 8D9 and pAbs H-143 detect a weakband with an approximate Mw �120 kDa (arrows on Fig.2b, upper panel), which was not always recognizable onimmunoblots. Because this band disappeared after preab-sorption of mAb 8D9 with an excess of adequate nonphos-phorylated decapeptide (see Fig. 1c), it is most likely spe-
Fig. 2. Localization of epitopes of mAb 8D9, mAb 7D10, pAbs H-143,comparison of DYRK1A levels in mouse brain tissue homogenates byby immunoblotting with these antibodies (b). (a) Domain structure of DGlu, Ser, Thr-rich domain (PEST), 13 consecutive-histidine repeat (Hisof DYRK1A immunostaining on membranes following SDS-PAGE. ArrmAb 8D9 and pAbs H-143 that is specific to DYRK1A. An asterisk indidiagrams presented below the immunoblots show the relative levels oby four antibodies tested and calculated by using densitometry analys
cific. However, its origin is at present uncertain. A weak
band with similar mobility on SDS-PAGE was present onsome immunoblots demonstrated by Okui and colleagues(1999), although these authors did not acknowledge itspresence. A weak band with Mw �60 kDa shown only bypAbs H-143 is most likely nonspecific (Fig. 2b, upperpanel, an asterisk).
Densitometry analysis of immunoblots showed that thelevels of DYRK1A detected by mAb 7D10, pAbs H-143,and pAbs against the N-terminus of DYRK1A in the brainsof trisomic animals constituted around 183%, 188%, and184%, respectively, of the values in disomic animals (Fig.2b, diagrams). These values are close to the 2.1-fold in-
s to the N-terminus of DYRK1A on DYRK1A domain structure (a) and9, mAb 7D10, pAbs H-143 and pAbs with the N-terminus of DYRK1Acludes nuclear localization signal (NLS), kinase domain (kinase), Pro,r, Thr-rich region (S/T). (b) The upper panel demonstrates the patternt to an additional weak band with Mw �120 kDa detectable only withadditional, most likely nonspecific, band detected by pAbs H-143. TheA in trisomic and disomic mouse brain tissue homogenates detectedmunoblots with P�0.0001 (denoted by *** on diagrams).
and pAbmAb 8D
YRK1A in), and Seows poincates an
crease of DYRK1A RNA calculated earlier by Northern

brhio1d
Do
Tylp
mtsF
(CcprsinTnDAtnbw
1
TnawleDs
mbsD
Dns
IbpDthdMaprt1dtarigrwa4e
E. Kida et al. / Neuroscience 195 (2011) 112–127118
blotting in adult Ts65Dn mice brain in comparison withdisomic brain (Guimera et al., 1999). Interestingly, thelevels of DYRK1A Tyr-145/147P� detected on immuno-lots of trisomic mouse brain homogenates by mAb 8D9eached �240% of the control values; thus they wereigher than the values demonstrated by the other antibod-
es we employed. This observation supports the results ofur dephosphorylation studies suggesting that Tyr-145/47 is less abundantly phosphorylated in trisomic thanisomic mouse brain.
YRK1A Tyr-145/147P� predominates in the nucleif astrocytes
o determine whether the status of Tyr-145/147 phosphor-lation of DYRK1A may affect the distribution and subcel-ular localization of DYRK1A in brain tissue, we next com-ared the immunolocalization of DYRK1A Tyr-145/147P�
visualized by mAb 8D9 with distribution of DYRK1A shownby the three other antibodies directed to DYRK1A used forimmunoblotting experiments.
Laser scanning confocal microscopy analyses of vi-bratome sections of mouse telencephalon showed thatmAb 8D9 labeled both neurons and astroglial cells. Therewere three major differences between the pattern ofDYRK1A Tyr-145/147P� immunoreactivity detected by
Ab 8D9 and DYRK1A immunostaining demonstrated byhe three other antibodies to DYRK1A we employed whenaponin was used for tissue permeabilization (Fig. 3a–f).irst, immunoreactivity to DYRK1A Tyr-145/147P� was
the strongest in the nuclei of astroglial cells (Fig. 3e, f,small arrows), while the immunoreactivity to DYRK1A vi-sualized in astroglial nuclei by the three other antibodieswe used was either markedly weaker than in neuronalnuclei (Fig. 3c, small arrow) or undetectable. Second, inmost neurons, immunoreactivity to DYRK1A Tyr-145/147P� was stronger in the nucleus than in the cytoplasmFig. 3e, f and inset in f) with pyramidal neurons of theA2/CA3 hippocampal sector showing the strongest nu-lear signal. In contrast, mAb 7D10 (Fig. 3a–c) and twoAbs to DYRK1A we used (not shown) showed immuno-eactivity to DYRK1A in the neuronal cytoplasm that wastronger than immunoreactivity in the nucleus (Fig. 3b, c,nset in c), except for individual pyramidal neurons in theeocortex (Fig. 3c, arrow). Third, the signal to DYRK1Ayr-145/147P� was weaker in the dendritic processes ofeurons than that with the three other antibodies toYRK1A we employed (compare Fig. 3b, c with Fig. 3e, f).ll antibodies to DYRK1A we used also immunostained
he initial segments of axons of neocortical large pyramidaleurons (arrowheads in insets in Fig. 3c, f). Heating therain sections before immunostaining increased the signalith mAb 8D9 (not shown).
The pattern of immunoreactivity to DYRK1A Tyr-145/47P� with Triton X-100 (Fig. 3g–i) differed to some extent
from that with saponin, especially when brain sectionswere heated to 80 °C before immunostaining (Fig. 3g–i).These experimental conditions increased the immunore-activity to DYRK1A Tyr-145/147P� in the nuclei of astro-
glial cells (Fig. 3h, i, thin arrows), but reduced the signal in dthe neuronal cells. Furthermore, under these experimentalconditions, neuronal cells showed a distinct, brain region-dependent gradient of immunoreactivity with the strongestsignal in the nuclei of pyramidal neurons of the CA2/CA3hippocampal sectors (Fig. 3h), slightly weaker in the nucleiof the granule cells of the hippocampal dentate gyrus (Fig.3g), markedly weaker in the CA1 hippocampal sector (Fig.3g, h), and only marginal in other brain areas including theneocortex (Fig. 3i). The cytoplasm of neurons was eitherweakly immunoreactive or was unstained. The use of Tri-ton X-100 hampered visualization of DYRK1A by mAb7D10, not affecting the signal obtained with both pAbs toDYRK1A we employed.
Because our immunofluorescence analyses demon-strated the strongest immunoreactivity to DYRK1A Tyr-145/147P� in astroglial nuclei, we also counted DYRK1A
yr-145/147P�-immunoreactive astroglial nuclei in theeocortex, hippocampal stratum lacunosum/molecularend radiatum and dentate stratum moleculare to assesshether astrocytosis could be responsible for the preva-
ence of DYRK1A Tyr-145/147P� in trisomic brain. How-ver, the number of astroglial nuclei immunopositive toYRK1A Tyr-145/147P� was similar in disomic and tri-omic brain in both areas tested (not shown).
We did not find any significant differences in the im-unostaining pattern between trisomic and disomic mouserain, though immunoreactivity to DYRK1A was invariablytronger in trisomic than disomic brain with all of the anti-YRK1A antibodies we employed (not shown).
YRK1A Tyr-145/147P� is strongly expressed in theuclei of the adult neuronal progenitors in theubgranular zone of the hippocampal dentate gyrus
nterestingly, DYRK1A Tyr-145/147P� (Fig. 4a, arrows)ut no other DYRK1A polypeptide(s) visualized either byAbs to the N-terminus (not shown) or other antibodies toYRK1A we employed (not shown) also was enriched in
he nuclei of cells located in the subgranular zone of theippocampal dentate gyrus; thus the brain region thatemonstrates active adult neurogenesis (for review seea et al., 2009). Newly born neurons are generated in thisrea in the adult brain from the radial glial cells (RGs) thatersist here in small amounts from the prenatal period (foreview see Pinto and Götz, 2007; Ma et al., 2009). Hence,o assess whether nuclei immunopositive to DYRK1A Tyr-45/147P� in the subgranular zone of the hippocampalentate gyrus may correspond to RGs, we double-labeledhe sections with mAb 8D9 and GFAP, a marker of bothdult astrocytes and RGs in mouse postnatal brain (foreview see Pinto and Götz, 2007). As shown by co-local-zation studies, GFAP-positive cells located within the sub-ranular zone of the hippocampal dentate gyrus with longadial processes extending through the granular zone, thusith RG morphology, indeed displayed strong immunore-ctivity to DYRK1A Tyr-145/147P� in the nuclei (Fig.a–c, arrows), confirming that DYRK1A Tyr-145/147P� isxpressed at high levels in progenitor cells of the adult
entate gyrus.
ele
ddd
of the Cll other�
E. Kida et al. / Neuroscience 195 (2011) 112–127 119
To determine whether DYRK1A Tyr-145/147P� is alsoxpressed by newly born neurons in this area, we double-
abeled brain sections with mAb 8D9 and doublecortin. The
Trito
S
mAb 7D10
g
d
a
mAb 8D9
mAb 8D9
m
m
Fig. 3. Immunofluorescence analysis of DYRK1A localization in the ad(d–f) show a widespread distribution of DYRK1A (a, d), with the strongf) and weak with mAb 7D10 (small arrow in c) and predominance of nustaining in neurons with mAb 7D10 (b, c and inset in c). Please noteb) than with mAb 8D9 (arrow in e), and the staining of the initial segm(arrowheads in insets). With Triton X-100 and heating of the sectionsregions (thin arrows in h and i), but neuronal nuclei are strongly labboundary between strongly labeled nuclei of the pyramidal neuronshippocampal sector (arrow in h). Scale bars in (a, d, g)�200 �m; in a
xpression of doublecortin starts at the early stage of 2
ifferentiation of newly born neurons in the hippocampalentate gyrus and continues through their axonal and den-ritic targeting (Brown et al., 2003; Kempermann et al.,
100
nin
mAb 7D10
2
CA1
h i
f
c
8D9 mAb 8D9
mAb 8D9
D10
D9e
b
neocortex
neocortex
neocortex
brain. With saponin permeabilization, mAb 7D10 (a–c) and mAb 8D9noreactivity of the astroglial nuclei with mAb 8D9 (small arrows in (e,ining in neurons with mAb 8D9 (e, f and inset in f) and of cytoplasmicer immunostaining of dendritic processes with mAb 7D10 (arrows inaxon of the neocortical large pyramidal neurons with both antibodies
, mAb 8D9 visualizes strongly the nuclei of astroglial cells in all brainin the CA2/CA3 hippocampal sectors (g, h). Please note the sharp
A2 and weakly labeled nuclei of the pyramidal neurons of the CA120 �m.
n X-
apo
CA
mAb
Ab 7
Ab 8
CA1
CA1
ult mouseest immuclear sta
the strongent of theto 80 °C
eled only
003). As shown in Fig. 4d (thick arrows), the nuclei of

mnA1cg
ptoDc1
paz
E. Kida et al. / Neuroscience 195 (2011) 112–127120
doublecortin-positive cells demonstrated a signal to DYRK1ATyr-145/147P�. Only a few doublecortin-positive cells,
ostly those located in deeper granule cell layers, wereegative to DYRK1A Tyr-145/147P� (Fig. 4d, thin arrow).ltogether, these observations indicated that DYRK1A Tyr-45/147P� is expressed in the nuclei of both progenitorells and newly born neurons in the adult dentate sub-
a
d
i
mAb 8D9 G
mAb 8D9DOUBLECORTIN
mAb 8
mAb 8D9 C
Fig. 4. Localization of DYRK1A Tyr-145/147P� in the adult mouseresent in the nuclei of progenitor cells with radial glial-like morpholorrows) except for individual cells showing weak signal (d, thin arrow);one of the hippocampal dentate gyrus. DYRK1A Tyr-145/147P� im
cholinergic axon terminals (j, red) as demonstrated on the merged imaleft corners. Scale bars in (a–e)�20 �m; in (f–k)�10 �m.
ranular zone. s
To evaluate whether DYRK1A Tyr-145/147P� is alsoresent during the division of neuronal progenitor cells inhis region, we employed labeling with BrdU, an analoguef thymidine that incorporates into the newly synthesizedNA of replicating cells during the S phase of the cellycle. As shown in Fig. 4e, a signal to DYRK1A Tyr-145/47P� was demonstrated by BrdU-positive cells in the
b c
e
f
g
h
j k
MERGED
BrdU mAb 8D9 BrdU
mAb 8D9 BrdU
mAb 8D9 BrdU
MERGED
rong immunoreactivity to DYRK1A Tyr-145/147P� with mAb 8D9 isarrows); in the nuclei of doublecortin-positive newly born neurons (d,dU-positive progenitor cells (arrows in e–h) located in the subgranularning also occurred in small varicosities (i, green) located in delicatellow). The dotted rectangles shown in (i–k) are enlarged in the upper
FAP
D9
hAT
brain. Stgy (a–c,and in Brmunostaige (k, ye
ubgranular layer of the hippocampal dentate gyrus. Given

pfiT
c
Ip
cbpsoT
p
FsT
tctp
to
dq
tbDf
Pe
GesopposTppe
E. Kida et al. / Neuroscience 195 (2011) 112–127 121
that the animals were sacrificed 6 h after the first BrdUinjection, and the calculated length of the S-phase in cellscomprising the dentate proliferating zone in adult mousewas estimated at 8.0�0.4 h (Nowakowski et al., 1989), therotocol of BrdU labeling we used could visualize cellsrom the S to the late M phase of the cell cycle. Interest-ngly, in the late M phase, immunoreactivity to DYRK1Ayr-145/147P� was present either in both daughter nuclei
(Fig. 4f, arrows) or only at one pole of the partitioning cell(Fig. 4g, arrow) or in only one daughter nucleus (Fig. 4h,arrow and thin arrow). These data indicate that DYRK1ATyr-145/147P� is involved in both symmmetrical andasymmetrical division of neuronal progenitor cells in theproliferative subgranular zone of the adult dentate gyrus.
DYRK1A Tyr-145/147P� is enriched in someholinergic axon terminals
nterestingly, with Triton X-100 and sporadically with sa-onin, immunoreactivity to DYRK1A Tyr-145/147P� also
was visible within delicate, small varicosities scattered inthe neuropil. These mAb 8D9-positive structures weregenerally rare, being seen most often in the dorsal tha-lamic nuclei, the stratum pyramidale and radiatum of thehippocampus (Fig. 4i), and the stratum moleculare of thehippocampal dentate gyrus (not shown). Double-labelingexperiments by using mAb 8D9 and antibodies specific forvarious neurotransmitter systems allowed us to localizethese varicosities to nerve terminals immunoreactive toChAT (Fig. 4i–k), the acetylcholine (ACh)-synthesizing en-zyme. Semiquantitative analyses of the dorsal thalamicnucleus double-labeled with mAb 8D9 and ChAT, carriedout in the latero-dorsal thalamic nucleus of three disomicand three trisomic animals by using ImageJ software,showed that DYRK1A Tyr-145/147P�-positive varicositiesonstituted from around 5% to 10% of the area occupiedy ChAT-positive structures. Some of these mAb 8D9-ositive axonal varicosities co-localized with synaptophy-in, a marker of synaptic vesicles (not shown). Thesebservations indicate that a small fraction of DYRK1Ayr-145/147P� is transported to cholinergic axons and
axon terminals, where some of them associate with syn-aptic vesicles.
DYRK1A Tyr-145/147P� levels are higher in theostnatal brains of DS subjects than controls
inally, by using immunoblotting, we investigated whether,imilar to in the model animals presented above, DYRK1Ayr-145/147P� levels also are higher in human DS brain in
comparison with the brain of neurologically normal sub-jects. Because the expression levels of DYRK1A are de-velopmentally regulated (Okui et al., 1999), which we alsoobserved at present (Fig. 5a), we studied separately thelevels of DYRK1A in the frontal cortices of human fetusesat midgestation and of children and young adults. Asshown in Fig. 5b, the levels of DYRK1A Tyr-145/147P� inhe brains of DS fetuses represent �139% of the values inontrol fetuses and are slightly lower than the values ob-ained with mAb 7D10 (163%), pAbs H-143 (173%), and
Abs to the N-terminus of DYRK1A (154%). However, inhe frontal cortices of children and young adults, the levelsf DYRK1A Tyr-145/147P� were distinctly higher in DS
subjects in comparison with controls (191%) than the lev-els of DYRK1A detected by mAb 7D10 (156%), pAbsH-143 (164%), and pAbs to the N-terminus of DYRK1A(149%) in this age-group. These data suggest that theextent of phosphorylation of DYRK1A at Tyr-145/147 islower in children and young adults with DS than in thegroup of control subjects of this age-group.
DISCUSSION
Phosphorylation regulates many vital processes in the cellby activating or inhibiting enzyme activity, leading to spe-cific site recognition for other molecules, promoting con-formational changes that facilitate protein association/dis-sociation and subcellular distribution of the target protein(for review see Johnson, 2009). Given the importance ofprotein phosphorylation for the biology of cells, identifica-tion of substrate proteins for DYRK1A and analysis of therole of their phosphorylation have been the subject ofmany published studies. However, only a very few studieshave focused on defining the pattern and the role of phos-phorylation of DYRK1A per se. Previous in vitro studiesemonstrated that autophosphorylation of Ser-520 is re-uired for 14-3-3� binding by DYRK1A and leads to in-
creased catalytic activity of the kinase (Álvarez et al.,2007). Another analysis of DYRK1A in vitro proposed thatautophosphorylation of Tyr-321 in the activation loop isessential for full kinase activity of DYRK1A (Himpel et al.,2001). However, the role of Tyr-321 in the activity ofDYRK1A is still a matter of debate (Adayev et al., 2007).
In the present study, we addressed for the first time,hough indirectly, phosphorylation of DYRK1A in therain tissue by using a monoclonal antibody recognizingYRK1A with Tyr-145/147 only in nonphosphorylated
orm.
hosphorylation of Tyr-145/147 of DYRK1A is morextensive in the control than DS brain
enerally, phosphorylation of tyrosine residues is a rarevent, as it represents only 1.8% of all phosphorylationites, and dynamic changes in tyrosine phosphorylationccur much faster compared with serine/threonine phos-horylation (Olsen et al., 2006). The results of our dephos-horylation experiments as well as densitometry analysesf immunoblots and comparative immunofluorescencetudies with the use of mAb 8D9 specific for DYRK1Ayr-145/147P� indicate that Tyr-145/147 of DYRK1A ap-ears in the brain tissue in both phosphorylated and non-hosphorylated forms. According to our studies, the lev-ls of DYRK1A Tyr-145/147P� species are higher by
�40% in postnatal trisomic brains in comparison withcontrols than those of total DYRK1A, indicating thatDYRK1A species with nonphosphorylated Tyr-145/147is more abundant in the trisomic than disomic brain.These results suggest that apart from increased expres-sion of DYRK1A, differences in phosphorylation levels of
this kinase between DS and control subjects should also
t
Ar
E. Kida et al. / Neuroscience 195 (2011) 112–127122
be considered when analyzing the role of DYRK1A in DSpathogenesis. Interestingly, in contrast to postnatalbrains, the ratios of DYRK1A detected by mAb 8D9 inthe trisomic vs. the disomic human fetal brains we stud-ied at midgestation were similar to those visualized bythe three other antibodies to DYRK1A, suggesting thatthe level of phosphorylation of Tyr-145/147 in prenatal
Fig. 5. Comparison of DYRK1A levels in human brain tissue homodensitometry analysis of this immunoblot after actin adjustment shoc) Diagrams showing the relative levels of DYRK1A in DS and contadults (c) detected by four antibodies tested and calculated by usP�0.05 marked on diagrams by ***, **, and *, respectively. All immusyndrome; Con, controls.
and postnatal trisomic brain varies.
DYRK1A Tyr-145/147P� has specific distribution inhe brain tissue
ccording to our immunofluorescence analyses, immuno-eactivity to DYRK1A Tyr-145/147P� was the strongest in
the nuclei of astroglial cells and was distinctly weaker inneuronal nuclei, except for pyramidal neurons of the CA2/
. (a) Immunoblot (left panel) and diagram (right panel) presentingvelopmental regulation of DYRK1A Tyr-145/147P� expression. (b,n brain tissue homogenates in fetuses (b) and children and young
sitometry analyses of immunoblots with P�0.0005, P�0.005 andere �-actin-adjusted. Abbreviations: w, weeks; y, years; DS, Down
genateswing de
rol humaing den
noblots w
CA3 hippocampal sectors. This pattern contrasts with the

rpata
Th
AlemaSbcsbaditipmocmc(
nbpbtdwtdrt(d
mmblpnreDaswiciaaaTgiTcrtfiD2(
nsc2GebwDTb
Tna
Ttmdpdaopll
E. Kida et al. / Neuroscience 195 (2011) 112–127 123
prominent expression of DYRK1A in neurons and the onlymarginal expression in the astroglial cells in the adultvertebrate brain emerging from both the literature (Häm-merle et al., 2003, 2008; Martí et al., 2003; Wegiel et al.,2004; Ferrer et al., 2005) and our confocal microscopyanalyses with the use of three antibodies to DYRK1A otherthan mAb 8D9. Altogether, these observations indicatethat phosphorylation of Tyr-145/147 is abundant in neu-rons, especially in the neuronal cytoplasm, but it occurssparsely in the astroglial cells, including modified radialglial cells acting as neuronal progenitor cells in the hip-pocampal dentate gyrus. Further studies must elucidatehow phosphorylation/dephosphorylation of Tyr-145/147 af-fects the function of DYRK1A. However, strong immuno-reactivity to DYRK1A Tyr-145/147P� in the nuclei of neu-onal progenitor cells and newly born neurons in the hip-ocampal dentate gyrus as well as in some ChAT-positivexonal terminals suggests the association of phosphoryla-ion/dephosphorylation of Tyr-145/147 of DYRK1A withdult hippocampal neurogenesis and cholinergic system.
he association of DYRK1A Tyr-145/147P� with adultippocampal neurogenesis
compelling line of evidence indicates that increasedevels of DYRK1A may contribute to abnormal neurogen-sis, a common feature of the brain of DS subjects andouse models for DS (Rueda et al., 2005; Chakrabarti etl., 2007; Contestabile et al., 2007; Ishihara et al., 2010).tudies of the neuroepithelial cells of early chicken em-ryos evidenced that Mnb/Dyrk1A mRNA is asymmetri-ally localized during mitosis, being inherited by one of theibling cells after division (Hämmerle et al., 2002). In em-ryonic mouse brain, Mnb/Dyrk1A also is mostly associ-ted with asymmetric cell division, and its expression wasetected in four sequential developmental phases, which
ncluded transient expression in preneurogenic progeni-ors, cell cycle-regulated expression in neurogenic progen-tors, transient expression in recently born neurons andersistent expression in late differentiating neurons (Häm-erle et al., 2008). Functional studies demonstrated thatverexpression of Dyrk1A in the embryonic mouse neo-ortex inhibited neural cell proliferation and promoted pre-ature neuronal differentiation in the developing cerebral
ortex, but without affecting cell fate and layer positioningYabut et al., 2010).
The potential association of DYRK1A also with adulteurogenesis, which continues in the adult mammalianrain in the subventricular zone of the lateral ventricleroducing neural progenitors migrating to the olfactoryulb and in the subgranular zone of the hippocampal den-ate gyrus generating new granule cells, has been ad-ressed only recently by Ferron and colleagues (2010),ho demonstrated the involvement of DYRK1A in subven-
ricular neurogenesis in the mouse brain. Our present studyemonstrates that DYRK1A is also implicated in adult neu-ogenesis in the hippocampal dentate gyrus. Interestingly, inhe subventricular neurogenesis documented by Ferron et al.2010) as well as in the hippocampal adult neurogenesis we
emonstrated, DYRK1A was either symmetrically or asym- retrically distributed during mitosis, in contrast to itsostly asymmetric segregation in mitotic cells in the em-ryonic brain. However, similar to what was observed ear-
ier in mouse embryos in which DYRK1A was widely ex-ressed in most neurogenic progenitors and in newborneurons in the ventricular proliferative zone but was down-egulated as these neurons began to migrate (Hämmerlet al., 2008), our study documented strong expression ofYRK1A Tyr-145/147P� in GFAP-positive progenitorsnd doublecortin-positive newborn neurons located in theubgranular zone of the adult dentate gyrus and distinctlyeaker immunostaining in the newly born neurons located
n the deeper layers of the dentate gyrus and adult granuleells. Furthermore, while predominantly cytoplasmic local-
zation of DYRK1A was demonstrated in the newly gener-ted cell pairs in the adult subventricular zone by Ferron etl. (2010) and by us in the dentate gyrus with the use ofntibodies to DYRK1A other than mAb 8D9, DYRK1Ayr-145/147P� was located in the nuclei of neuronal pro-enitors and newly born hippocampal neurons. These find-
ngs suggest that phosphorylation/dephosphorylation ofyr-145/147 may be involved in complex processes asso-iated with cell division/differentiation underlying adult neu-ogenesis in the mammalian brain. The potential associa-ion of Tyr-145/147 phosphorylation with cell division/dif-erentiation is supported by the results of threendependent phosphoproteomic studies, which identifiedYRK1A phosphopeptides with Tyr-145 (Rush et al.,005; Rikova et al., 2007; Mayya et al., 2009) and Tyr-147Mayya et al., 2009) phosphorylated in tumor cells.
Normal levels of DYRK1A are necessary for self-re-ewal, epidermal growth factor-dependent cell-fate deci-ions, and long-term persistence in vivo of neural stemells isolated from the adult ventricular zone (Ferron et al.,010). The cell-survival-promoting effect of DYRK1A (e.g.uo et al., 2010) and the inhibition of the proliferation ofmbryonic neuronal cells (e.g. Park et al., 2010) have alsoeen ascribed recently to DYRK1A activity. However,hether similar effects on neurogenesis are exerted byYRK1A in the adult hippocampus and what the role ofyr-145/147 phosphorylation is in these processes muste elaborated by further studies.
he association of DYRK1A Tyr-145/147P� witheuronal processes and some cholinergicxon terminals
he presence of DYRK1A in dendritic processes already inhe embryonic brain raised the hypothesis that DYRK1Aay be involved in the signaling mechanisms that regulateendritic differentiation (Hämmerle et al., 2003). In theresent study, we observed DYRK1A signal not only inendrites but also in proximal neurites as well as a focalccumulation of DYRK1A Tyr-145/147P� in the terminalsf cholinergic axons. The presence of DYRK1A in theroximal neurites also was reported by Marti and col-
eagues (2003), but the association of DYRK1A with cho-inergic axon terminals is novel and has not yet been
eported.
Df
aprsr
M
FciigabpifmmccW2
aqgot8wba1tbsnbDni
E. Kida et al. / Neuroscience 195 (2011) 112–127124
Many published in vitro studies have indicated thatYRK1A influences neuritogenesis, mostly promoting it,
or example, by priming sites of GSK3� phosphorylation inMAP1B (Scales et al., 2009), by upregulating the Ras/MAPkinase signaling pathway (Kelly and Rahmani, 2005), byphosphorylating the Notch intracellular domain (NICD) andby attenuating NICD signaling in neural cells, thus attenu-ating the repression of neuritogenesis by NICD (Fernan-dez-Martinez et al., 2009). In this regard, interactions ofDYRK1A with neuronal restricted silencing factors/RE-1silencing transcription factor (NRSF/REST) may be func-tionally important (Canzonetta et al., 2008; Lepagnol-Bes-tel et al., 2009). NRSF/REST plays a strategic role interminal neuronal differentiation, as its downregulation isessential for induction and maintenance of the neural phe-notype (Ballas et al., 2001; Chong et al., 1995; Schoenherret al., 1996). Interestingly, REST/NRSF also regulates thecholinergic gene locus comprising the ChAT agene andthe vesicular ACh transporter gene (for review see Shi-mojo and Hersh, 2004). Degeneration of cholinergic basalforebrain neurons that supply the major cholinergic input toboth the hippocampus and neocortex has been reported inthe brains of young and adult DS subjects (Casanova etal., 1985), and decreases in activities of ChAT and acetyl-cholinesterase levels have been reported in older DS sub-jects (Yates et al., 1980). Also, Ts65Dn mice show markedloss of basal forebrain cholinergic neurons with age (Holtz-man et al., 1996). It is well documented that the cholinergicsystem is a major player in learning and memory pro-cesses. More recent studies suggest that the cholinergicsystem also regulates adult neurogenesis, as its activationpromotes the survival of newborn neurons in the adultdentate gyrus and olfactory bulb (Kaneko et al., 2006),while ablation of the cholinergic system of the basal fore-brain led to decreased adult neurogenesis in these areas(Cooper-Kuhn et al., 2004). Furthermore, functional �7-containing nicotinic ACh receptors that are expressedearly on adult-born neurons are necessary for the normalsurvival, maturation, and integration of these adult-bornneurons in the dentate gyrus (Campbell et al., 2010).
Earlier experiments suggested that DYRK1A may af-fect the splicing machinery in the cholinergic neurons (Toi-ber et al., 2010). Our study demonstrates that DYRK1Amay act not only in the nuclei of cholinergic neurons butalso in cholinergic axon terminals, as its form nonphospho-rylated at Tyr-145/147 is transported in small amountsalong the cholinergic axons in specific brain areas. Inter-estingly, earlier biochemical studies identified synapticvesicle- and endocytotic vesicle-associated proteins suchas dynamin (Chen-Hwang et al., 2002; Huang et al., 2004);amphiphysin 1 (Murakami et al., 2006, 2009); synaptojanin1 (Adayev et al., 2006); and �-synuclein (Kim et al., 2006)s DYRK1A substrates, which led to the hypothesis of aotential role for DYRK1A in controlling synaptic vesicleecycling processes (Murakami et al., 2009). Our studyuggests that some cholinergic nerve terminals may rep-
esent the site of these interactions.ethodological remarks
inally, we would like to make two short methodologicalomments. First, our data underscore the need for caution
n interpreting the results of immunohistochemical studiesnvolving the use of antibodies directed against DYRK1Aiven that the method of tissue processing (e.g. poor visu-lization of neuronal nuclei in formalin-fixed, paraffin-em-edded brain tissues—EK, personal observation) and theretreatment conditions used may significantly affect the
mmunostaining pattern, which also was reported earlieror other proteins showing both the nuclear and cytoplas-ic localization (O’Reilly et al., 2004; Frisch, 2004). Thus,ethodological differences may, at least partially, explain
ontroversies in the published literature regarding the lo-alization of DYRK1A in the brain tissue (Martí et al., 2003;egiel et al., 2004; Ferrer et al., 2005; Hämmerle et al.,
008).Second, a short comment regards our observation that
ntibodies directed against aa 32–51 of the DYRK1A se-uence do not detect the major doublet of DYRK1A mi-rating on SDS-PAGE with Mw �87–94 kDa. The inabilityf the N-terminally directed antibody to detect in brainissue the major DYRK1A polypeptides visualized by mAbD9 and two C-terminally-directed antibodies to DYRK1Ae employed suggests that a portion of DYRK1A in therain is N-terminally truncated between aa 32–51 (thentigen of pAbs anti N-terminus of DYRK1A) and aa 142–47 (the epitope for mAb 8D9). Another possibility implyinghat the major doublet of DYRK1A visualized on immuno-lots represents DYRK1A gene splicing products eithertarting from different translation initiation codon(s) and/orot encompassing aa residues 32–51 is less probable,ecause the two major alternatively spliced transcripts ofyrk1A expressed in the rodent brain differ only by 27ucleotides in codons 69–78 (Kentrup et al., 1996). This
ssue has to be addressed by future studies.
Acknowledgments—This work was supported by the NYS Officefor People with Developmental Disabilities and Tosinvest, Italy.There is no conflict of interest. The authors thank Ms. MaureenStoddard Marlow for copy-editing the manuscript.
REFERENCES
Adayev T, Chen-Hwang MC, Murakami N, Lee E, Bolton DC, HwangYW (2007) Dual-specificity tyrosine phosphorylation-regulated ki-nase 1A does not require tyrosine phosphorylation for activity invitro. Biochemistry 46:7614–7624.
Adayev T, Chen-Hwang MC, Murakami N, Wang R, Hwang YW (2006)MNB/DYRK1A phosphorylation regulates the interactions of syn-aptojanin 1 with endocytic accessory proteins. Biochem BiophysRes Commun 351:1060–1065.
Altafaj X, Dierssen M, Baamonde C, Martí E, Visa J, Guimerà J, OsetM, González JR, Flórez J, Fillat C, Estivill X (2001) Neurodevel-opmental delay, motor abnormalities and cognitive deficits in trans-genic mice overexpressing Dyrk1A (minibrain), a murine model ofDown’s syndrome. Hum Mol Genet 10:1915–1923.
Álvarez M, Altafaj X, Aranda S, de la Luna S (2007) DYRK1A auto-phosphorylation on serine residue 520 modulates its kinase activity
via 14-3-3 binding. Mol Biol Cell 18:1167–1178.
E. Kida et al. / Neuroscience 195 (2011) 112–127 125
Álvarez M, Estivill X, de la Luna S (2003) DYRK1A accumulates insplicing speckles through a novel targeting signal and inducesspeckle disassembly. J Cell Sci 116:3099–3107.
Ballas N, Battaglioli E, Atouf F, Andres ME, Chenoweth J, AndersonME, Burger C, Moniwa M, Davie JR, Bowers WJ, Federoff HJ,Rose DW, Rosenfeld MG, Brehm P, Mandel G (2001) Regulationof neuronal traits by a novel transcriptional complex. Neuron31:353–365.
Becker W, Weber Y, Wetzel K, Eirmbter K, Tejedor FJ, Joost HG(1998) Sequence characteristics, subcellular localization, and sub-strate specificity of DYRK-related kinases, a novel family of dualspecificity protein kinases. J Biol Chem 273:25893–25902.
Brown JP, Couillard-Després S, Cooper-Kuhn CM, Winkler J, AignerL, Kuhn HG (2003) Transient expression of doublecortin duringadult neurogenesis. J Comp Neurol 467:1–10.
Campbell NR, Fernandes CC, Halff AW, Berg DK (2010) Endogenoussignaling through alpha7-containing nicotinic receptors promotesmaturation and integration of adult-born neurons in the hippocam-pus. J Neurosci 30:8734–8744.
Canzonetta C, Mulligan C, Deutsch S, Ruf S, O’Doherty A, Lyle R,Borel C, Lin-Marq N, Delom F, Groet J, Schnappauf F, De Vita S,Averill S, Priestley JV, Martin JE, Shipley J, Denyer G, Epstein CJ,Fillat C, Estivill X, Tybulewicz VL, Fisher EM, Antonarakis SE,Nizetic D (2008) DYRK1A-dosage imbalance perturbs NRSF/REST levels, deregulating pluripotency and embryonic stem cellfate in Down syndrome. Am J Hum Genet 83:388–400.
Casanova MF, Walker LC, Whitehouse PJ, Price DL (1985) Abnor-malities of the nucleus basalis in Down’s syndrome. Ann Neurol18:310–313.
Chakrabarti L, Galdzicki Z, Haydar TF (2007) Defects in embryonicneurogenesis and initial synapse formation in the forebrain of theTs65Dn mouse model of Down syndrome. J Neurosci 27:1483–1495.
Chen-Hwang MC, Chen HR, Elzinga M, Hwang YW (2002) Dynamin isa minibrain kinase/dual specificity Yak1-related kinase 1A sub-strate. J Biol Chem 277:17597–17604.
Chong JA, Tapia-Ramírez J, Kim S, Toledo-Aral JJ, Zheng Y, BoutrosMC, Altshuller YM, Frohman MA, Kraner SD, Mandel G (1995)REST: a mammalian silencer protein that restricts sodium channelgene expression to neurons. Cell 80:949–957.
Contestabile A, Fila T, Ceccarelli C, Bonasoni P, Bonapace L, SantiniD, Bartesaghi R, Ciani E (2007) Cell cycle alteration and de-creased cell proliferation in the hippocampal dentate gyrus and inthe neocortical germinal matrix of fetuses with Down syndrome andin Ts65Dn mice. Hippocampus 17:665–678.
Cooper-Kuhn CM, Winkler J, Kuhn HG (2004) Decreased neurogen-esis after cholinergic forebrain lesion in the adult rat. J NeurosciRes 77:155–165.
Davisson MT, Schmidt C, Akeson EC (1990) Segmental trisomy ofmurine chromosome 16: a new model system for studying Downsyndrome. Prog Clin Biol Res 360:263–280.
Delabar JM, Theophile D, Rahmani Z, Chettouh Z, Blouin JL, Prieur M,Noel B, Sinet PM (1993) Molecular mapping of twenty-four features ofDown syndrome on chromosome 21. Eur J Hum Genet 1:114–124.
Dowjat WK, Adayev T, Kuchna I, Nowicki K, Palminiello S, Hwang YW,Wegiel J (2007) Trisomy-driven overexpression of DYRK1A kinase inthe brain of subjects with Down syndrome. Neurosci Lett 413:77–81.
Epstein CJ (1986) Developmental genetics. Experientia 42:1117–1128.
Fernandez-Martinez J, Vela EM, Tora-Ponsioen M, Ocaña OH, NietoMA, Galceran J (2009) Attenuation of Notch signalling by theDown-syndrome-associated kinase DYRK1A. J Cell Sci 22:1574–1583.
Ferrer I, Barrachina M, Puig B, Martínez de Lagrán M, Martí E, Avila J,Dierssen M (2005) Constitutive Dyrk1A is abnormally expressed inAlzheimer disease, Down syndrome, Pick disease, and related
transgenic models. Neurobiol Dis 20:392–400.Ferron SR, Pozo N, Laguna A, Aranda S, Porlan E, Moreno M, Fillat C,de la Luna S, Sánchez P, Arbonés ML, Fariñas I (2010) Regulatedsegregation of kinase Dyrk1A during asymmetric neural stem celldivision is critical for EGFR-mediated biased signaling. Cell StemCell 7:367–379.
Fotaki V, Dierssen M, Alcántara S, Martínez S, Martí E, Casas C, VisaJ, Soriano E, Estivill X, Arbonés ML (2002) Dyrk1A haploinsuffi-ciency affects viability and causes developmental delay and ab-normal brain morphology in mice. Mol Cell Biol 22:6636–6647.
Frisch S (2004) Nuclear localization of FADD protein. Cell Death Differ11:1361–1362.
Golabek AA, Kida E, Walus M, Wujek P, Mehta P, Wisniewski KE(2003) Biosynthesis, glycosylation, and enzymatic processing invivo of human tripeptidyl-peptidase I. J Biol Chem 278:7135–7145.
Guimera J, Casas C, Estivill X, Pritchard M (1999) Human minibrainhomologue (MNBH/DYRK1): characterization, alternative splicing,differential tissue expression, and overexpression in Down syn-drome. Genomics 57:407–418.
Guimerá J, Casas C, Pucharcòs C, Solans A, Domènech A, PlanasAM, Ashley J, Lovett M, Estivill X, Pritchard MA (1996) A humanhomologue of Drosophila minibrain (MNB) is expressed in theneuronal regions affected in Down syndrome and maps to thecritical region. Hum Mol Genet 5:1305–1310.
Guo X, Williams JG, Schug TT, Li X (2010) DYRK1A and DYRK3promote cell survival through phosphorylation and activation ofSIRT1. J Biol Chem 285:13223–13232.
Hämmerle B, Carnicero A, Elizalde C, Ceron J, Martínez S, Tejedor FJ(2003) Expression patterns and subcellular localization of theDown syndrome candidate protein MNB/DYRK1A suggest a role inlate neuronal differentiation. Eur J Neurosci 17:2277–2286.
Hämmerle B, Elizalde C, Tejedor FJ (2008) The spatio-temporal andsubcellular expression of the candidate Down syndrome geneMnb/Dyrk1A in the developing mouse brain suggests distinct se-quential roles in neuronal development. Eur J Neurosci 27:1061–1074.
Hämmerle B, Vera-Samper E, Speicher S, Arencibia R, Martínez S,Tejedor FJ (2002) Mnb/Dyrk1A is transiently expressed and asym-metrically segregated in neural progenitor cells at the transition toneurogenic divisions. Dev Biol 246:259–273.
Himpel S, Panzer P, Eirmbter K, Czajkowska H, Sayed M, PackmanLC, Blundell T, Kentrup H, Grötzinger J, Joost HG, Becker W(2001) Identification of the autophosphorylation sites and charac-terization of their effects in the protein kinase DYRK1A. Biochem J359:497–505.
Holtzman DM, Santucci D, Kilbridge J, Chua-Couzens J, Fontana DJ,Daniels SE, Johnson RM, Chen K, Sun Y, Carlson E, Alleva E,Epstein CJ, Mobley WC (1996) Developmental abnormalities andage-related neurodegeneration in a mouse model of Down syn-drome. Proc Natl Acad Sci U S A 93:13333–13338.
Huang Y, Chen-Hwang MC, Dolios G, Murakami N, Padovan JC,Wang R, Hwang YW (2004) Mnb/Dyrk1A phosphorylation regu-lates the interaction of dynamin 1 with SH3 domain-containingproteins. Biochemistry 43:10173–10185.
Ishihara K, Amano K, Takaki E, Shimohata A, Sago H, Epstein CJ,Yamakawa K (2010) Enlarged brain ventricles and impaired neu-rogenesis in the Ts1Cje and Ts2Cje mouse models of Downsyndrome. Cereb Cortex 20:1131–1143.
Johnson LN (2009) The regulation of protein phosphorylation.Biochem Soc Trans 37:627–641.
Kaneko N, Okano H, Sawamoto K (2006) Role of the cholinergicsystem in regulating survival of newborn neurons in the adultmouse dentate gyrus and olfactory bulb. Genes Cells 11:1145–1159.
Kelly PA, Rahmani Z (2005) DYRK1A enhances the mitogen-activatedprotein kinase cascade in PC12 cells by forming a complex withRas, B-Raf, and MEK1. Mol Biol Cell 16:3562–3573.
Kempermann G, Gast D, Kronenberg G, Yamaguchi M, Gage FH
(2003) Early determination and long-term persistence of adult-
E. Kida et al. / Neuroscience 195 (2011) 112–127126
generated new neurons in the hippocampus of mice. Development130:39139–39139.
Kentrup H, Becker W, Heukelbach J, Wilmes A, Schürmann A, Hup-pertz C, Kainulainen H, Joost HG (1996) Dyrk, a dual specificityprotein kinase with unique structural features whose activity isdependent on tyrosine residues between subdomains VII and VIII.J Biol Chem 271:3488–3495.
Kim EJ, Sung JY, Lee HJ, Rhim H, Hasegawa M, Iwatsubo T, Min doS, Kim J, Paik SR, Chung KC (2006) Dyrk1A phosphorylates�-synuclein and enhances intracellular inclusion formation. J BiolChem 281:33250–33257.
Korbel JO, Tirosh-Wagner T, Urban AE, Chen XN, Kasowski M, Dai L,Grubert F, Erdman C, Gao MC, Lange K, Sobel EM, Barlow GM,Aylsworth AS, Carpenter NJ, Clark RD, Cohen MY, Doran E,Falik-Zaccai T, Lewin SO, Lott IT, McGillivray BC, Moeschler JB,Pettenati MJ, Pueschel SM, Rao KW, Shaffer LG, Shohat M, VanRiper AJ, Warburton D, Weissman S, Gerstein MB, Snyder M,Korenberg JR (2009) The genetic architecture of Down syndromephenotypes revealed by high-resolution analysis of human seg-mental trisomies. Proc Natl Acad Sci U S A 106:12031–12036.
Korenberg JR, Kawashima H, Pulst S-H, Ikeuchi T, Ogasawara N,Yamamoto K, Schonberg SA, West R, Allen L, Magenis E, IkawaK, Taniguchi N, Epstein CJ (1990) Molecular definition of a regionof chromosome 21 that causes features of the Down syndromephenotype. Am J Hum Genet 47:236–246.
Lepagnol-Bestel AM, Zvara A, Maussion G, Quignon F, Ngimbous B,Ramoz N, Imbeaud S, Loe-Mie Y, Benihoud K, Agier N, Salin PA,Cardona A, Khung-Savatovsky S, Kallunki P, Delabar JM, PuskasLG, Delacroix H, Aggerbeck L, Delezoide AL, Delattre O, GorwoodP, Moalic JM, Simonneau M (2009) DYRK1A interacts with theREST/NRSF-SWI/SNF chromatin remodeling complex to deregu-late gene clusters involved in the neuronal phenotypic traits ofDown syndrome. Hum Mol Genet 18:1405–1414.
Lyle R, Béna F, Gagos S, Gehrig C, Lopez G, Schinzel A, LespinasseJ, Bottani A, Dahoun S, Taine L, Doco-Fenzy M, Cornillet-LefèbvreP, Pelet A, Lyonnet S, Toutain A, Colleaux L, Horst J, Ken-nerknecht I, Wakamatsu N, Descartes M, Franklin JC, Florentin-Arar L, Kitsiou S, Aït Yahya-Graison E, Costantine M, Sinet PM,Delabar JM, Antonarakis SE (2009) Genotype-phenotype correla-tions in Down syndrome identified by array CGH in 30 cases ofpartial trisomy and partial monosomy chromosome 21. Eur J HumGenet 17:454–466.
Ma DK, Bonaguidi MA, Ming GL, Song H (2009) Adult neural stemcells in the mammalian central nervous system. Cell Res 19:672–682.
Martí E, Altafaj X, Dierssen M, de la Luna S, Fotaki V, Alvarez M,Pérez-Riba M, Ferrer I, Estivill X (2003) Dyrk1A expression patternsupports specific roles of this kinase in the adult central nervoussystem. Brain Res 964:250–263.
Mayya V, Lundgren DH, Hwang SI, Rezaul K, Wu L, Eng JK, RodionovV, Han DK (2009) Quantitative phosphoproteomic analysis of T cellreceptor signaling reveals system-wide modulation of protein–pro-tein interactions. Sci Signal 2:ra46.
Miller DL, Currie JR, Mehta PD, Potempska A, Hwang YW, Wegiel J(2003) Humoral immune response to fibrillar beta-amyloid peptide.Biochemistry 42:11682–11692.
Møller RS, Kübart S, Hoeltzenbein M, Heye B, Vogel I, Hansen CP,Menzel C, Ullmann R, Tommerup N, Ropers HH, Tümer Z,Kalscheuer VM (2008) Truncation of the Down syndrome candi-date gene DYRK1A in two unrelated patients with microcephaly.Am J Hum Genet 82:1165–1170.
Murakami N, Bolton D, Hwang YW (2009) Dyrk1A binds to multipleendocytic proteins required for formation of clathrin-coated vesi-cles. Biochemistry 48:9297–9305.
Murakami N, Xie W, Lu RC, Chen-Hwang MC, Wieraszko A, HwangYW (2006) Phosphorylation of amphiphysin I by minibrain kinase/
dual-specificity tyrosine phosphorylation-regulated kinase, a ki-nase implicated in Down syndrome. J Biol Chem 281:23712–23724.
Nowakowski RS, Lewin SB, Miller MW (1989) Bromodeoxyuridineimmunohistochemical determination of the lengths of the cell cycleand the DNA-synthetic phase for an anatomically defined popula-tion. J Neurocytol 18:311–318.
O’Reilly L, Crawford S, Huang DCS, Strasser A (2004) Nuclear locali-sation of FADD—rebuttal. Cell Death Differ 11:1362–1363.
Okui M, Ide T, Morita K, Funakoshi E, Ito F, Ogita K, Yoneda Y, KudohJ, Shimizu N (1999) High-level expression of the Mnb/Dyrk1A genein brain and heart during rat early development. Genomics62:165–171.
Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, MannM (2006) Global, in vivo, and site-specific phosphorylation dynam-ics in signaling networks. Cell 127:635–648.
Palminiello S, Kida E, Kaur K, Walus M, Wisniewski KE, Wierzba-Bobrowicz T, Rabe A, Albertini G, Golabek AA (2008) Increasedlevels of carbonic anhydrase II in the developing Down syndromebrain. Brain Res 1190:193–205.
Park J, Oh Y, Yoo L, Jung MS, Song WJ, Lee SH, Seo H, Chung KC(2010) Dyrk1A phosphorylates p53 and inhibits proliferation ofembryonic neuronal cells. J Biol Chem 285:31895–31906.
Park J, Song WJ, Chung KC (2009) Function and regulation ofDyrk1A: towards understanding Down syndrome. Cell Mol Life Sci66:3235–3240.
Pennington BF, Moon J, Edgin J, Stedron J, Nadel L (2003) Theneuropsychology of Down syndrome: evidence for hippocampaldysfunction. Child Dev 74:75–93.
Pinto L, Götz M (2007) Radial glial cells heterogeneity—the source ofdiverse progeny in the CNS. Prog Neurobiol 83:2–23.
Rikova K, Guo A, Zeng Q, Possemato A, Yu J, Haack H, Nardone J,Lee K, Reeves C, Li Y, Hu Y, Tan Z, Stokes M, Sullivan L, MitchellJ, Wetzel R, Macneill J, Ren JM, Yuan J, Bakalarski CE, Villen J,Kornhauser JM, Smith B, Li D, Zhou X, Gygi SP, Gu TL, Polakie-wicz RD, Rush J, Comb MJ (2007) Global survey of phosphoty-rosine signaling identifies oncogenic kinases in lung cancer. Cell131:1190–1203.
Rueda N, Mostany R, Pazos A, Flórez J, Martínez-Cué C (2005) Cellproliferation is reduced in the dentate gyrus of aged but not youngTs65Dn mice, a model of Down syndrome. Neurosci Lett380:197–201.
Rush J, Moritz A, Lee KA, Guo A, Goss VL, Spek EJ, Zhang H, ZhaXM, Polakiewicz RD, Comb MJ (2005) Immunoaffinity profiling oftyrosine phosphorylation in cancer cells. Nat Biotechnol 23:94–101.
Scales TM, Lin S, Kraus M, Goold RG, Gordon-Weeks PR (2009)Nonprimed and DYRK1A-primed GSK3 beta-phosphorylation siteson MAP1B regulate microtubule dynamics in growing axons. J CellSci 122:2424–2435.
Schoenherr CJ, Paquette AJ, Anderson DJ (1996) Identification ofpotential target genes for the neuron-restrictive silencer factor.Proc Natl Acad Sci U S A 93:9881–9886.
Shimojo M, Hersh LB (2004) Regulation of the cholinergic gene locusby the repressor element-1 silencing transcription factor/neuronrestrictive silencer factor (REST/NRSF). Life Sci 74:2213–2225.
Song WJ, Sternberg LR, Kasten-Sportès C, Keuren ML, Chung SH,Slack AC, Miller DE, Glover TW, Chiang PW, Lou L, Kurnit DM(1996) Isolation of human and murine homologues of the Drosoph-ila minibrain gene: human homologue maps to 21q22.2 in theDown syndrome “critical region.” Genomics 15:331–339.
Tejedor F, Zhu XR, Kaltenbach E, Ackermann A, Baumann A, Canal I,Heisenberg M, Fischbach KF, Pongs O (1995) Minibrain: a newprotein kinase family involved in postembryonic neurogenesis inDrosophila. Neuron 14:287–301.
Toiber D, Azkona G, Ben-Ari S, Torán N, Soreq H, Dierssen M (2010)Engineering DYRK1A overdosage yields Down syndrome-charac-
teristic cortical splicing aberrations. Neurobiol Dis 40:348–359.
E. Kida et al. / Neuroscience 195 (2011) 112–127 127
Wegiel J, Kuchna I, Nowicki K, Frackowiak J, Dowjat K, Silverman WP,Reisberg B, DeLeon M, Wisniewski T, Adayev T, Chen-Hwang MC,Hwang YW (2004) Cell type- and brain structure-specific patterns ofdistribution of minibrain kinase in human brain. Brain Res 1010:69–80.
Wiseman FK, Alford KA, Tybulewicz VLJ, Fisher EMC (2009) Downsyndrome—recent progress and future prospects. Hum Mol Genet18:R75–R83.
Wisniewski KE, Kida E, Golabek AA, Palminiello S, Walus M, Rabe A,
Albertini G (2006) Down syndrome: from pathology to pathogene-sis. In: Down syndrome: neurobehavioral specificity. Downsyndrome series (Rondal JA, Perera J, eds), pp 17–33. London:John Wiley & Sons.
Yabut O, Domogauer J, D’Arcangelo G (2010) Dyrk1A overex-pression inhibits proliferation and induces premature neuronaldifferentiation of neural progenitor cells. J Neurosci 30:4004 – 4014.
Yates CM, Simpson J, Maloney AF, Gordon A, Reid AH (1980) Al-zheimer-like cholinergic deficiency in Down syndrome. Lancet
316:979.(Accepted 12 August 2011)(Available online 19 August 2011)
















![Binder 147, Opecoelidae He-J [Trematoda Taxon Notebooks]](https://img.dokumen.tips/doc/110x75/63290446eedc98f54f00fde8/binder-147-opecoelidae-he-j-trematoda-taxon-notebooks.jpg)












