Embed Size (px)
Citation preview

Phagozytose von nekrotischen Zellen durch gesunde Haut-Fibroblasten und Tumorzellen
Der Naturwissenschaftlichen Fakultät
Der Friedrich-Alexander-Universität Erlangen-Nürnberg
zur
Erlangung des Doktorgrades Dr. rer. nat.
vorgelegt von
Manuela Schwegler
aus München

ii
Als Dissertation genehmigt von der Naturwissenschaftlichen Fakultät der Friedrich-Alexander-Universität
Erlangen-Nürnberg
Tag der mündlichen Prüfung: 27.01.2016
Vorsitzende/r des Promotionsorgans: Prof. Dr. Jörn Wilms
Gutachter/in: Prof. Dr. Falk Nimmerjahn
Prof. Dr. Rainer Fietkau

1
Inhaltsverzeichnis Abkürzungsverzeichnis .......................................................................................... 6
Abbildungsverzeichnis ........................................................................................... 8
Tabellenverzeichnis ............................................................................................... 10
Zusammenfassung ................................................................................................ 11
Abstract .................................................................................................................... 14
1. Einleitung.......................................................................................................... 16
Formen von Zelltod ................................................................................ 16 1.1.
Apoptose ................................................................................................. 17 1.1.1.
Primäre und sekundäre Nekrose .......................................................... 17 1.1.2.
Regulierte Nekrose ................................................................................. 18 1.1.3.
Autophagischer Zelltod .......................................................................... 19 1.1.4.
Immunogenität der Zelltodesformen ..................................................... 19 1.1.5.
Endozytose.............................................................................................. 21 1.2.
Pinozytose ............................................................................................... 21 1.2.1.
Phagozytose ........................................................................................... 22 1.2.2.
Beseitigung sterbender Zellen durch Professionelle Phagozyten ..... 23 1.3.
Phagozytose apoptotischer Zellen ........................................................ 24 1.3.1.
Phagozytose nekrotischer Zellen .......................................................... 25 1.3.2.
Nicht-professionelle Phagozyten und Zell-in-Zell Phänomene .......... 25 1.4.
Nicht-professionelle Phagozyten und Zell-Kannibalismus ................. 26 1.4.1.
Emperipolese .......................................................................................... 27 1.4.2.
Entose ...................................................................................................... 28 1.4.3.
Zielsetzung .............................................................................................. 28 1.5.
2. Material und Methoden .................................................................................. 30
Verwendete Zelllinien, Reagenzien, Geräte und Verbrauchsmaterial2.1.
30

2
Zellkultur .................................................................................................. 36 2.2.
Kultivierungsbedingungen ..................................................................... 36 2.2.1.
Passage und Konservierung von Zellen .............................................. 36 2.2.2.
Transfektion ............................................................................................ 37 2.2.3.
Nukleosid-Inkorporation ......................................................................... 37 2.2.4.
Induktion und Inhibition von nicht-professioneller Phagozytose ........ 38 2.3.
Inaktivierung von Zellen durch Hitzeeinwirkung .................................. 38 2.3.1.
Proliferationspotenzial von Hitze-behandelten Zellen ......................... 38 2.3.2.
Vitalfärbung ............................................................................................. 39 2.3.3.
Phagozytose von hyperthermierten Zellen .......................................... 40 2.3.4.
Inhibition der Phagozytose von Hitze-behandelten Zellen ................. 41 2.3.5.
Funktionale Blockade von E-und P-Cadherin ...................................... 42 2.3.6.
Behandlung mit Annexin V zur Blockade von Phosphatidylserin ...... 42 2.3.7.
Phagozytose von bestrahlten Zellen .................................................... 43 2.3.8.
Beobachtung der Phagozytose-Ereignisse über die Zeit ................... 44 2.3.9.
Erstellung der Objektträger nach der Koinkubation ............................ 45 2.3.10.
Quantifizierung der Phagozytose-Raten .............................................. 45 2.3.11.
Durchflusszytometrie .............................................................................. 46 2.4.
Bestimmung der Zellmorphologie und Zellviabilität............................. 46 2.4.1.
Messung der Zell-Adhäsion ................................................................... 48 2.4.2.
Messung der Konzentration der zellulären Proteine aus Zelllysaten 2.5.
und von freigesetztem HSP70 nach Hyperthermie ........................................... 49
Immunfluoreszenz-Färbung ................................................................... 50 2.6.
Färbe-Verfahren ..................................................................................... 51 2.6.1.
Quantifizierung der Lokalisation von Phagozytose-assoziierten 2.6.2.
Proteinen ............................................................................................................... 51
Immunhistochemie ................................................................................. 52 2.7.

3
Auswahl von Gewebeproben ................................................................ 52 2.7.1.
Färbung von histologischen Schnitten ................................................. 52 2.7.2.
Quantifizierung von Phagozytose-Raten in histologischen Präparaten2.7.3.
53
Mikroskopie ............................................................................................. 53 2.8.
Fluoreszenz- und Phasenkontrastmikroskopie .................................... 53 2.8.1.
Konfokale Mikroskopie ........................................................................... 54 2.8.2.
Lebendzell-Mikroskopie ......................................................................... 54 2.8.3.
Messung der Genexpression ................................................................ 54 2.9.
RNA-Isolation und reverse Transkription ............................................. 54 2.9.1.
Quantitative PCR .................................................................................... 55 2.9.2.
Statistik .................................................................................................... 55 2.10.
3. Ergebnisse ....................................................................................................... 57
Nachweis der Phagozytose ................................................................... 57 3.1.
Induktion von Nekrose und Phagozytose............................................. 59 3.2.
Hyperthermie........................................................................................... 59 3.2.1.
Bestrahlung ............................................................................................. 61 3.2.2.
Phagozytose von hyperthermierten Zellen .......................................... 63 3.3.
Quantifizierung der homotypischen Phagozytose-Raten bei 3.3.1.
verschiedenen Zelltypen ...................................................................................... 63
Quantifizierung der heterotypischen Phagozytose-Raten bei 3.3.2.
verschiedenen Zelltypen ...................................................................................... 65
Protein-Verlust und Freisetzung von HSP70 durch Hyperthermie .... 66 3.3.3.
Einfluss von freigesetzten DAMPs und FBS auf die Phagozytose .... 67 3.3.4.
Aufnahme von nekrotischen Zellen durch angewachsene Zellen ..... 68 3.3.5.
Phagozytose von bestrahlten Zellen und nekrotischer Debris ........... 69 3.4.
Quantifizierung der homotypischen und heterotypischen 3.4.1.
Phagozytose-Raten bei verschiedenen Zelltypen ............................................. 70

4
Aufnahme nekrotischer Debris .............................................................. 71 3.4.2.
Einfluss von Hyperthermie auf die Zelladhäsion und Phagozytose .. 72 3.5.
Mikroskopischer Nachweis der Adhäsion ............................................ 72 3.5.1.
Nachweis der Hyperthermie-induzierten Adhäsion mittels 3.5.2.
Durchflusszytometrie ........................................................................................... 73
Phagozytose-Raten in Abhängigkeit der induzierten Adhäsion ......... 75 3.5.3.
Lokalisation von Zell-in-Zell-assoziierten Proteinen ............................ 75 3.6.
Lokalisation von E-Cadherin.................................................................. 76 3.6.1.
Verdichtung von F-Aktin......................................................................... 76 3.6.2.
Akkumulation von LAMP1 und LC3 ...................................................... 78 3.6.3.
Quantifizierung der Zell-in-Zell-assoziierten Proteine im Zeitverlauf . 79 3.6.4.
Lokalisation der Zell-in-Zell-assoziierten Proteine bei angewachsenen 3.6.5.
Zellen 80
Inhibition der molekularen Maschinerie der nicht-professionellen 3.7.
Phagozyten ........................................................................................................... 81
Einfluss von Bestrahlung, Proteasen und Zytoskelett-Inhibitoren ..... 81 3.7.1.
Blockade des E- und P-Cadherin vermittelten Zellkontakts ............... 83 3.7.2.
Quantifizierung der Dosis-Relation von Annexin V ............................. 87 3.7.3.
Expression von Phagozytose-assoziierten Genen .............................. 88 3.8.
Degradation der inkorporierten Zelle .................................................... 90 3.9.
Zell-in-Zell Strukturen im Gewebe ........................................................ 95 3.10.
4. Diskussion ...................................................................................................... 102
Phagozytose-Induktion durch Hyperthermie ...................................... 102 4.1.
Phagozytose-Induktion durch Röntgenstrahlung .............................. 105 4.2.
Rekrutierung Phagozytose-assoziierter Proteine .............................. 106 4.3.
Zelltod der inkorporierten Zelle ........................................................... 107 4.4.
Nicht-professionelle Phagozytose versus professionelle Phagozytose4.5.
109

5
Nicht-professionelle Phagozytose versus Kannibalismus ................ 111 4.6.
Nicht-professionelle Phagozytose versus Entose ............................. 112 4.7.
Prognostischer Wert von Zell-in-Zell Strukturen bei 4.8.
Krebserkrankungen ............................................................................................ 113
Schlussfolgerung und Ausblick ........................................................... 115 4.9.
5. Literaturverzeichnis ...................................................................................... 118
6. Anhang – Ergänzende Tabellen ................................................................. 128
7. Danksagung ................................................................................................... 132
8. Lebenslauf ...................................................................................................... 133
9. Publikationsliste ............................................................................................ 134

6
Abkürzungsverzeichnis 7-AAD 7-Aminoactinomycin
Aqua dest. Bidestilliertes Wasser
ATP Adenosintriphosphat
AnxA5 Annexin A5
BrdU 5-Brom-2’-desoxyuridin
BSA Rinderserumalbumin
c Konzentration
CD cluster of differentiation
cDNA copy DNA
CIC Zell-in-Zell Struktur (cell-in-cell)
CTFR celltrace far red
CTOG celltrace oregon green
DAMP damage-associated molecular pattern
DAPI 4’,6-Diamidin-2-Phenylindol
DCs Dendritische Zellen (dendritic cells)
DMEM Dulbecco’s modified eagle medium
DMSO Dimethylsulfoxid
DNS Desoxyribonukleinsäure
EDTA Ethylendiamintetraacetat
EdU 5-Ethynyl-2’-Desoxyuridin
F-Aktin filamentöses Aktin, Aktinfilamente
FBS Fötales Rinderserum (fetal bovine serum)
FITC Fluoreszeinisothiocyanat
GFP green fluorescent protein
GTP Guanosintriphosphat
H2B-RFP Histon H2B-red fluorescent protein
HNSCC Kopf-Hals-Plattenepithelkarzinomen (head and neck squa-
mous cell carcinoma)
HSP Hitzeschockprotein
HT Hyperthermie
Ig Immunglobulin
IL Interleukin
LAMP 1 lysosomal-associated membrane protein 1

7
LAP LC3 – assoziierte Phagozytose
LC3 microtubule-associated protein 1 light chain 3
LSM laser scanning microscopy
MeOH Methanol
PAMP pathogen associated molecular patterns
PBS Phosphat-gepufferte Salzlösung (phosphate buffered sa-
line)
PCR Polymerase-Kettenreaktion (polymerase chain reaction)
p-Ezrin Ezrin, phosphoryliert
PS Phosphatidylserin
qRT-PCR quantitative Echtzeit-PCR
RT Raumtemperatur
RAGE receptor for advanced glycation end products
RFP red fluorescent protein
TAM Tumor-assoziierte Makrophagen
TBS Tris-gepufferte Salzlösung (tris buffered saline)
TLR toll like receptor
TMA Tissue-Microarray
TRIS Tris(hydroxymethyl)-aminomethan
β-Me beta-Mercaptoethanol

8
Abbildungsverzeichnis Abbildung 1: Punktewolkendiagramm zur Darstellung von Zellgröße und
Granularität ....................................................................................................... 47
Abbildung 2: Durchflusszytometrische Messung zum Nachweis von Apoptose,
Nekrose und Zellaggregaten ............................................................................. 48
Abbildung 3: Messung der Adhäsion ................................................................ 49
Abbildung 4: Nachweis der Phagozytose von hyperthermierten Zellen mit
konfokaler Laser-Scanning Mikroskopie ........................................................... 58
Abbildung 5: Nachweis der Phagozytose von hyperthermierten Zellen mit
Fluoreszenz- und Lebendzell-Mikroskopie ........................................................ 59
Abbildung 6: Analyse der Induktion der Zelltodesformen Apoptose und Nekrose
nach Hyperthermie ............................................................................................ 60
Abbildung 7: Koloniebildungspotenzial nach Hitze-Behandlung ....................... 61
Abbildung 8: Induktion von Zelltod bei mehreren Zelllinien durch Bestrahlung
und Hitze-Behandlung ...................................................................................... 62
Abbildung 9: Phagozytose von apoptotischen Zellen nach Bestrahlung ........... 63
Abbildung 10: Homotypische Phagozytose-Raten von verschiedenen Zelllinien
.......................................................................................................................... 64
Abbildung 11: Heterotypische Phagozytose-Raten von verschiedenen Zelllinien
.......................................................................................................................... 65
Abbildung 12: Einfluss von Hyperthermie auf die Gesamt-Protein-Konzentration
und die Freisetzung von HSP70 ....................................................................... 67
Abbildung 13: Phagozytose-Raten nach Entfernung von freigesetzten DAMPs
durch Induktion primärer Nekrose und nach FBS-Mangel ................................ 68
Abbildung 14: Phagozytose-Ereignisse bei angewachsenen Zellen ................. 69
Abbildung 15: Vergleich der Phagozytose-Raten von hyperthermierten und
bestrahlten Zellen bei verschiedenen Zelllinien ................................................ 70
Abbildung 16: Phagozytose von nekrotischer Debris ........................................ 72
Abbildung 17: Verstärktes adhäsives Verhalten gegenüber hyperthermierter
Zellen ................................................................................................................ 73
Abbildung 18: Durchflusszytometrische Messung der Adhäsion bei mehreren
Zelllinien ............................................................................................................ 74
Abbildung 19: Hyperthermie-Behandlung induziert eine verstärkte Bindung
zwischen phagozytischen und hyperthermierten Zellen .................................... 74

9
Abbildung 20: Phagozytose-Raten in Abhängigkeit von Hyperthermie-
Behandlung ....................................................................................................... 75
Abbildung 21: Verdichtung von E-Cadherin bei der Phagozytose ..................... 76
Abbildung 22: Verdichtung von F-Aktin und p-Ezrin bei der Phagozytose ........ 77
Abbildung 23: Lokalisation von LAMP1 und LC3 .............................................. 79
Abbildung 24: Quantifizierung der Zell-in-Zell-assoziierten Proteine ................. 80
Abbildung 25: Lokalisation der Phagozytose-assoziierten Proteine bei
angewachsenen Zell-in-Zell Strukturen ............................................................. 81
Abbildung 26: Phagozytose-Raten nach Bestrahlung und Behandlung der
phagozytiertenden Zellen mit Protease- und Zytoskelett-Inhibitoren ................ 82
Abbildung 27: Expression und Lokalisation von E-Cadherin auf vitalen,
hyperthermierten Zellen und Phagozytose-Ereignissen .................................... 84
Abbildung 28: Expression und Lokalisation von P-Cadherin auf vitalen,
hyperthermierten Zellen und Phagozytose-Ereignissen .................................... 85
Abbildung 29: Einfluss funktioneller Blockade von E- und P-Cadherin auf
Adhäsion und Phagozytose .............................................................................. 87
Abbildung 30: Dosis-abhängige Inhibition der Phagozytose von
hyperthermierten Zellen mit Annexin V ............................................................. 88
Abbildung 31: Relative mRNA-Konzentrationen von Phagozytose-relevanten
Genen ............................................................................................................... 89
Abbildung 32: Beobachtung von Degradation nach Phagozytose, Teil 1 .......... 92
Abbildung 33: Beobachtung von Degradation nach Phagozytose, Teil 2 .......... 93
Abbildung 34: Nachweis von lysosomalem Zelltod der internalisierten Zelle .... 94
Abbildung 35: Nachweis von Zell-in-Zell Strukturen in Gewebeschnitten ......... 96
Abbildung 36: Zell-in-Zell Strukturen im Gewebeschnitt ................................... 97
Abbildung 37: Häufigkeit von Zell-in-Zell Strukturen bei verschiedenen
Krebserkrankungen ........................................................................................... 98
Abbildung 38: Häufigkeit von Zell-in-Zell Strukturen in verschiedenen
Krebserkrankungen im Vergleich .................................................................... 100
Abbildung 39: Überlebenskurven nach Kaplan-Meier ..................................... 101
Abbildung 40: Modell zur nicht-professionellen Phagozytose ......................... 116

10
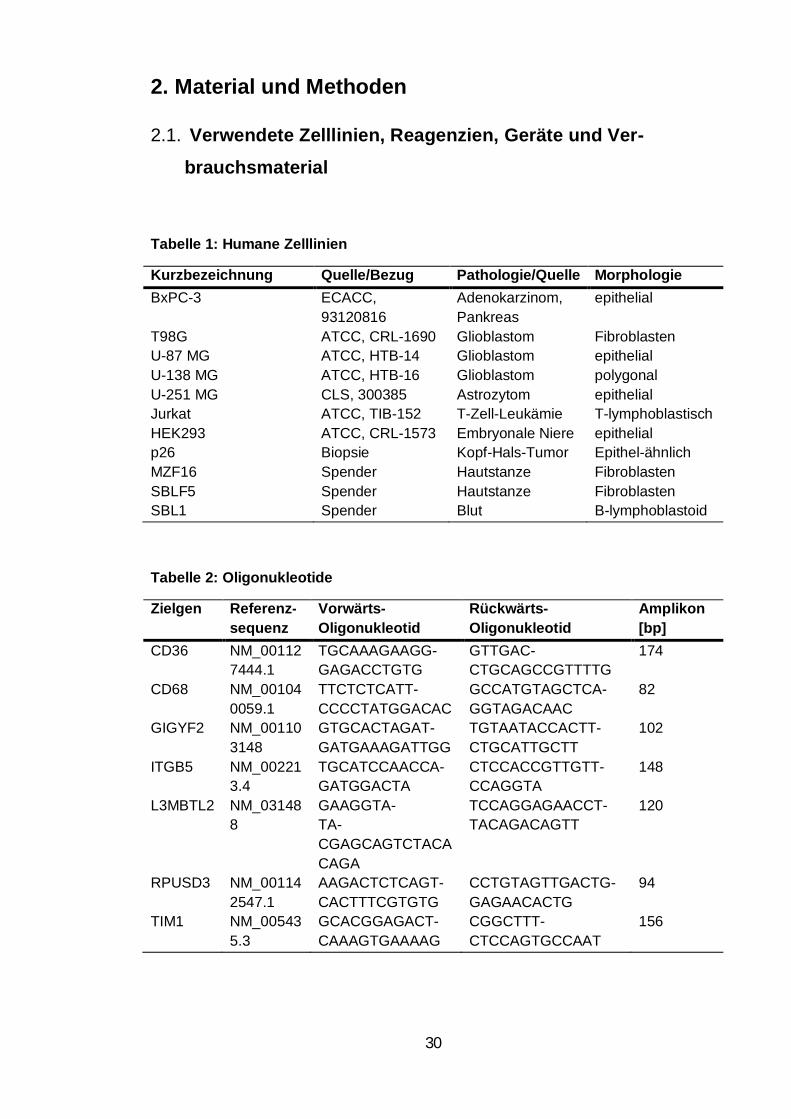
Tabellenverzeichnis Tabelle 1: Humane Zelllinien .................................................................................. 30
Tabelle 2: Oligonukleotide ....................................................................................... 30
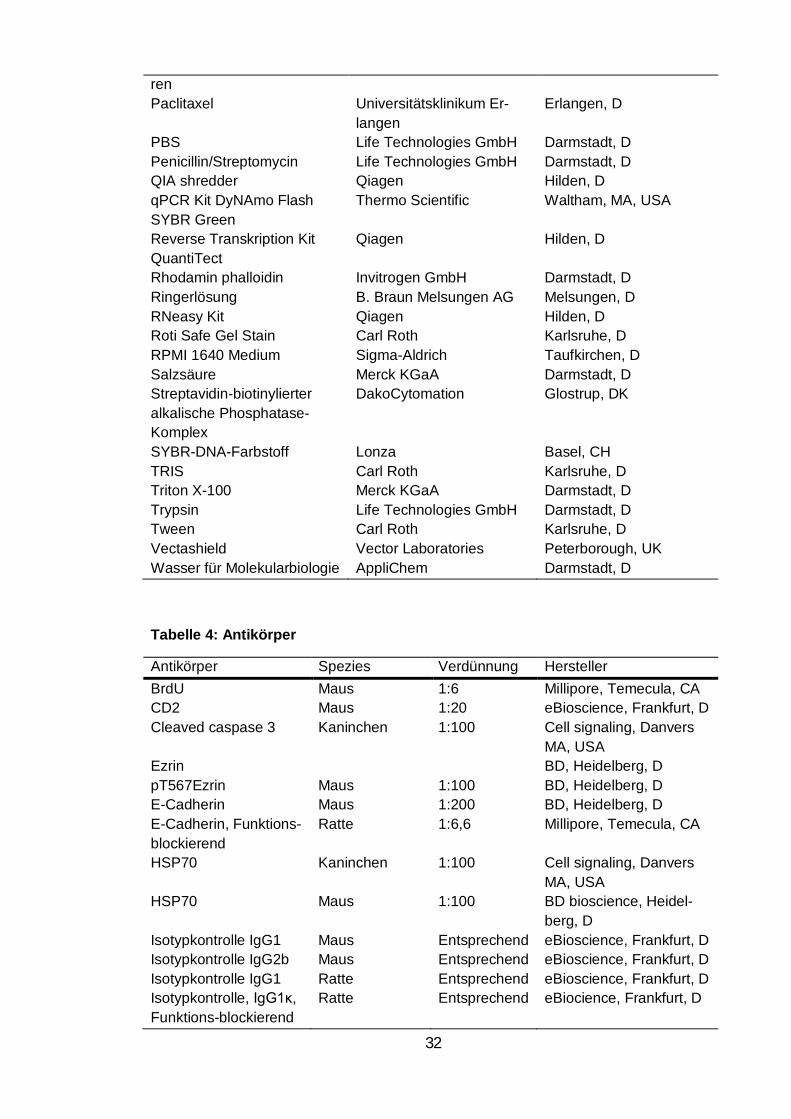
Tabelle 3: Reagenzien ............................................................................................. 31
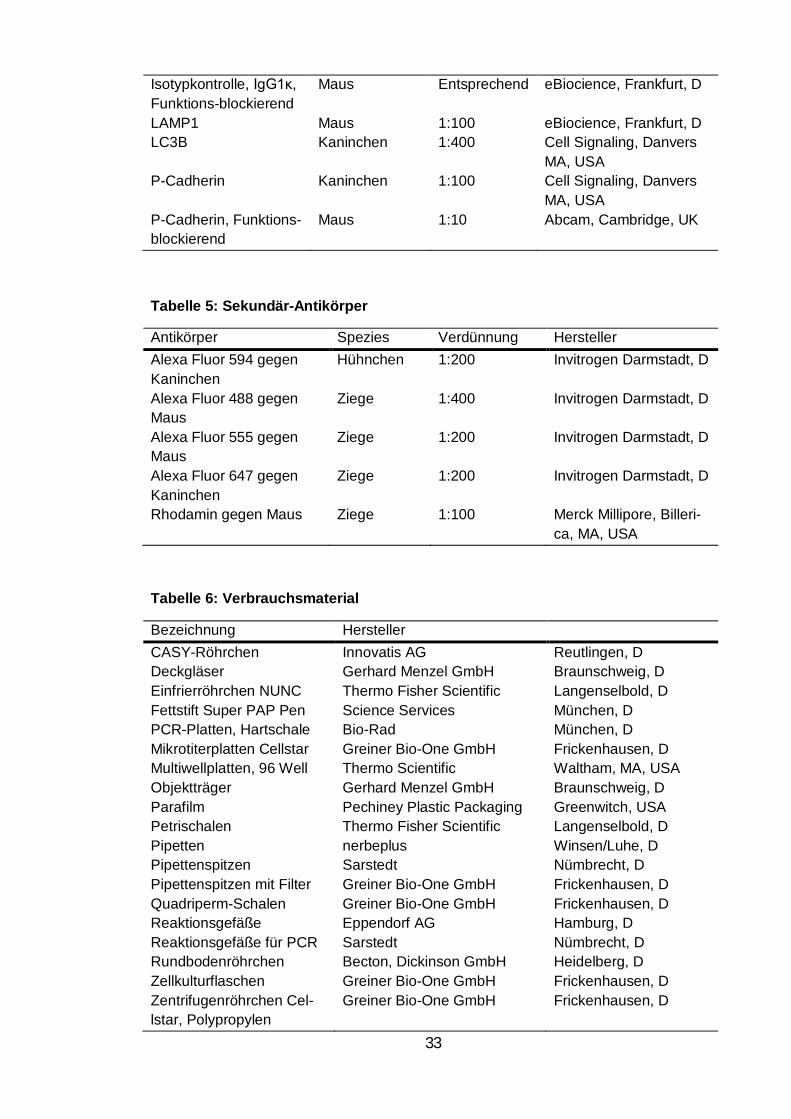
Tabelle 4: Antikörper ................................................................................................ 32
Tabelle 5: Sekundär-Antikörper .............................................................................. 33
Tabelle 6: Verbrauchsmaterial ................................................................................ 33
Tabelle 7: Geräte...................................................................................................... 34
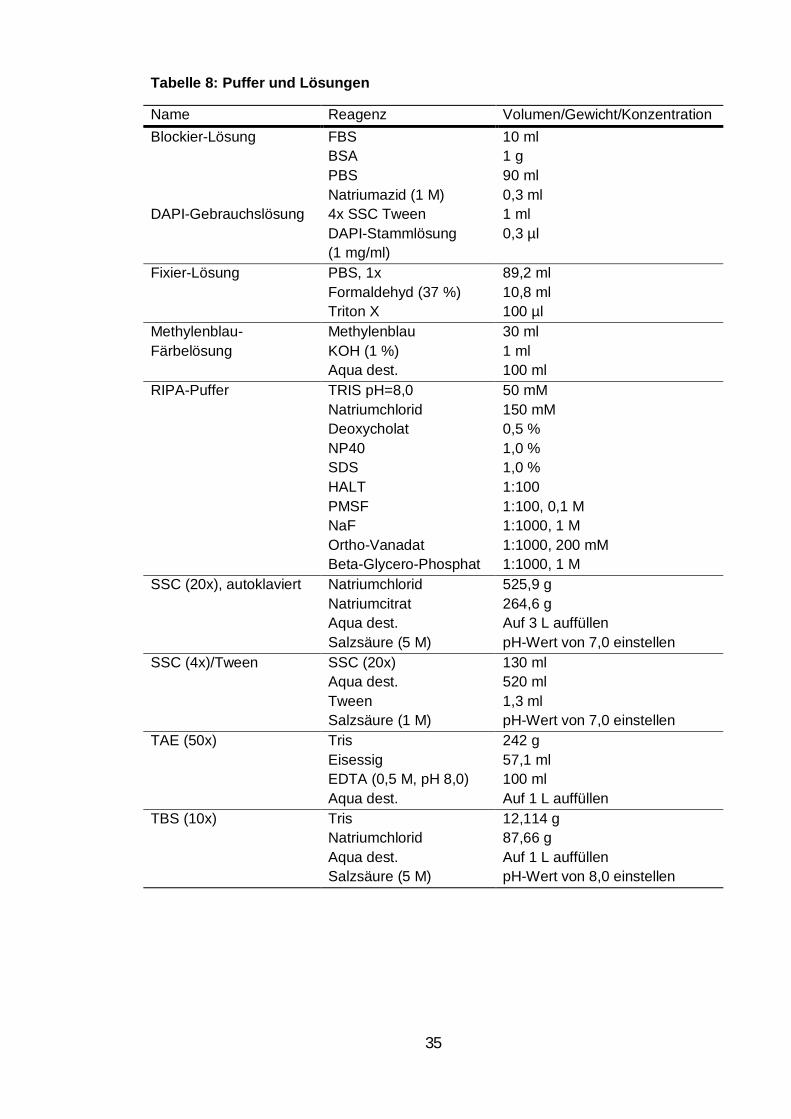
Tabelle 8: Puffer und Lösungen.............................................................................. 35
Tabelle 9: Kulturmedien ........................................................................................... 36
Tabelle 10: Kombination von Zelllinien bei der heterotypischen Phagozytose .. 40
Tabelle 11: Versuchsansätze zur Untersuchung des Zeitverlaufs nach
Phagozytose ............................................................................................................. 44
Tabelle 12: Induktion von Zelltod durch Bestrahlung ........................................... 70
Tabelle 13: Klinische Daten der Kopf-Hals-Plattenepithelkarzinomen, Teil 1 .. 128
Tabelle 14: Klinische Daten der Kopf-Hals-Plattenepithelkarzinomen, Teil 2 .. 129
Tabelle 15: Klinische Daten der Rektum- und Analkarzinom Kohorten ............ 130
Tabelle 16: Überleben der Patienten ................................................................... 131

11
Zusammenfassung Die zeitnahe Phagozytose von apoptotischen und nekrotischen Zellen verhin-
dert den Eintritt von sekundärer Nekrose und die Induktion von Entzündungsre-
aktionen und Autoimmunerkrankungen. Die Phagozytose-Aktivität von benach-
barten Zellen, welche sterbende Zellen aufnehmen, kommt dann zum Einsatz,
wenn die effiziente Phagozytose-Kapazität von professionellen Phagozyten, wie
Makrophagen und dendritischen Zellen, nicht ausreicht bzw. zu wenige dieser
Zellen in kürzester Zeit in das betroffene Gewebe einwandern können.
Die Phagozytose-Fähigkeit verschiedener Zelltypen und die zugrunde liegen-
den molekularen Mechanismen sind noch nicht vollständig untersucht. Insbe-
sondere für die Aufnahme nekrotischer Zellen durch benachbarte Zellen liegen
nicht viele Studien vor. Phagozytose-Ereignisse von Tumorzellen in Tumorge-
webeschnitten, sog. Zell-Kannibalismus, gleichen den Zell-in-Zell Strukturen,
die bei der Beseitigung nekrotischer Zellen durch nicht-professionelle Phagozy-
ten entstehen. Sie weisen ebenfalls einen sichelförmigen und an die Peripherie
gedrängten Zellkern der äußeren Zelle und eine runde Form der aufgenomme-
nen Zelle auf und stellen ein häufig dokumentiertes Ereignis in histologischen
Untersuchungen dar. Studien über eine prognostische Relevanz von Zell-in-Zell
Strukturen sind aber bislang aufgrund kleiner Kohorten-Größen weniger aussa-
gekräftig und im Vergleich zudem widersprüchlich.
In dieser Arbeit sollte die Phagozytose-Kapazität mehrerer Zelltypen getestet,
molekulare Mechanismen sowie eine prognostische Rolle von Zell-in-Zell Struk-
turen in Tumorgewebeschnitten von 83 Patienten mit Rektumkarzinom und von
321 Patienten mit Kopf-Hals-Plattenepithelkarzinom untersucht werden.
Um dies zu erreichen wurde zunächst die Fähigkeit zur homotypischen und he-
terotypischen Aufnahme nekrotischer Zellen bei verschiedenen Zelltypen getes-
tet, inklusive Pankreaskarzinom-Zellen, mehrere morphologisch divergente
Glioblastom-Zelllinien, eine primäre Kopf-Hals-Tumorzelllinie, primäre, gesunde
Hautfibroblasten, semi-adhärente Nierenepithelzellen, T-Zell-Leukämiezellen
sowie lymphoblastoide Zellen. Als Phagozytose-fähig wurden von diesen unter-
suchten Zelllinien alle adhärenten Zelllinien identifiziert. Immunfluoreszenz-
Färbungen zeigten, dass die Phagozytose-Aktivität von einer Akkumulation des
Adhäsionsmoleküls E-Cadherin, von phosphoryliertem Ezrin, dem Autophagie-

12
Marker Mikrotubuli-assoziierten Protein LC3 (light chain 3) sowie des Lysoso-
men-assoziierten Membranproteins LAMP1 begleitet wurde.
Die Hitze-behandelten, nekrotischen Zellen wiesen einen Verlust der Integrität
der Zellmembran sowie eine zytoplasmatische Anhäufung von LC3-positiven
Vesikeln und eine Zunahme der intrazellulären HSP70-Konzentration auf und
wurden von lebenden Zellen in Suspension oder wenn letztere angewachsen
waren, phagozytiert.
Funktionelle Analysen zeigten, dass die Phagozytose-Aktivität der nicht-
professionellen Phagozyten abhängig von einer dynamischen Reorganisation
des Zellskeletts (Aktinfilamente und Mikrotubuli), der GTPase Aktivität von Dy-
namin II und der Erkennung von Phosphatidylserin ist.
Durch histologische Untersuchungen anhand von Tumorgewebeschnitten von
Patienten mit Kopf-Hals-Plattenepithelkarzinom oder Rektumkarzinom wurde
eine prognostische Relevanz von Zell-in-Zell Strukturen nachgewiesen. Niedri-
ge Häufigkeiten von Zell-in-Zell Strukturen korrelierten mit einem längeren
Überleben. Das histologische Material von 23 Patienten mit Analkarzinom be-
stätigte ebenfalls das häufige Vorkommen von Zell-in-Zell Strukturen. Die
Quantifizierung erfolgte nach immunhistologischer Färbung für E-Cadherin
(Membran) und Gegenfärbung mit Hämatoxylin (Zellkerne). Zusätzlich wurde
die Häufigkeit von Zell-in-Zell Strukturen mit apoptotischen Zellen mittels Fär-
bung von aktivierter Caspase 3 analysiert. Die Häufigkeit von Zell-in-Zell Struk-
turen rangierte zwischen 0,7 und 17,3 Ereignissen pro mm2, wohingegen kaum
Zell-in-Zell Strukturen mit apoptotischen Zellen nachgewiesen wurden. Der
prognostischen Relevanz von Zell-in-Zell Ereignissen könnten eine veränderte
Tumorimmunität und Immunzell-Infiltration sowie ein niedrigeres Metastasie-
rungs-Risiko zugrunde liegen. Wenn Zell-in-Zell Strukturen in verschiedenen
Tumorentitäten eine bedeutende Rolle im Tumormilieu spielen, wie es in dieser
Arbeit für Kopf-Hals-Plattenepithelkarzinome und Rektumkarzinome gezeigt
wurde, könnten sie auch bei der Erforschung neuer Behandlungsmodalitäten
berücksichtigt werden.
Zusammenfassend zeigen die Ergebnisse, dass die Phagozytose nekrotischer
Zellen von verschiedenen Tumorzelllinien sowie gesunden Hautfibroblasten
ausgeführt wird und dadurch die Phagozytose-Aktivität von professionellen
Phagozyten unterstützt. Dieser Prozess verhindert die Freisetzung von proin-
flammatorischen Gefahrensignalen und verringert so Entzündungsreaktionen

13
und das Risiko für die Entstehung von Autoimmunerkrankungen. Auch im Tu-
mor werden ähnliche Zell-in-Zell Strukturen gefunden, deren Häufigkeit von
prognostischer Relevanz ist.

14
Abstract Rapid removal of apoptotic and necrotic cells prevents secondary necrosis and
the induction of autoimmune diseases and inflammation. The phagocytic activity
of neighboring cells, which engulf dying cells, may occur if the amount of dying
cells exceeds the phagocytic capacity of professional phagocytes such as den-
dritic cells and macrophages or if the number of immediately recruited phago-
cytes into the tissue is not sufficient.
The scope of this study was the investigation of phagocytosis-capable cell types
as well as the molecular background of the engulfment process of primary ne-
crotic cells by neighboring tumor cells and normal skin fibroblasts. The removal
of necrotic cells by non-professional phagocytes remains poorly characterized.
Phagocytic events of tumor cells have frequently been observed in tumor tissue
sections and were named cell cannibalism. These cell-in-cell structures resem-
ble the phagocytic events that result from the clearance of necrotic cells by non-
professional phagocytes studied herein: The round-shaped inner cell is totally
encircled by the host cell membrane and the semilunar host cell nucleus is
pushed to the periphery. It has been proposed that cell cannibalism could be
used as prognostic factor but these studies are conflicting, postulating a tumor-
promoting and metastasis-protecting role and restricted by the lack of large co-
horts.
In this study, we investigated the phagocytic capacity of several cell types, mo-
lecular mechanisms as well as the prognostic role of tumor cell cannibalism in
tumor tissue sections of 83 patients with rectum carcinoma and 321 patients
with head and neck squamous cell carcinoma.
To achieve this the phagocytic activity regarding the homotypic and heterotypic
engulfment of necrotic cells by several cell types including primary fibroblasts,
pancreatic adenocarcinoma, glioblastoma and head and neck cancer cells,
semi-adherent embryonic kidney cells, leukemic T-cells and lymphoblastoid
cells was studied. Exclusively all adherent cell lines were capable for the en-
gulfment of necrotic cells. Using immunofluorescence staining an accumulation
of adhesion molecule E-cadherin, phosphorylated actin-linker protein ezrin, ly-
sosomal-associated membrane protein 1 (LAMP1) and microtubule-associated
protein 1 light chain 3 (LC3) at the site of engulfment were observed.

15
Heat-treated, necrotic cells exhibited loss of cellular membrane integrity and
increase of the intracellular level of heat-shock protein 70 and LC3-positive ves-
icles and were engulfed by untreated, phagocytic cells in suspension or if the
phagocytic cells were adhered. Functional analyses revealed that nonprofes-
sional phagocytes require dynamic rearrangement of the cytoskeleton (actin
and microtubules), recognition of phosphatidylserine and GTPase activity of
dynamin II, which normally mediates endocytosis.
Histologic examination of tumor tissue sections of patients with head-and-neck
cancer or rectal cancer revealed a prognostic relevance of cell-in-cell structures.
Low frequencies of cell-in-cell structures correlated with better survival rates.
Additionally, analysis of tumor tissues sections of 23 patients with anal carcino-
ma confirmed high abundance of these events in the tissue. Cell-in-cell struc-
tures were observed in all of the cohorts with a frequency ranging from 0.7 to
17.3 cell-in-cell events per mm2. Quantification was performed after immuno-
histochemically staining of the samples for E-cadherin (cell membranes) and
counterstaining with hematoxylin (nuclei). Additionally, analysis of the frequency
of cell-in-cell structures containing apoptotic cells after staining for cleaved
caspase-3 showed that hardly any apoptotic cells were engulfed by tumor cells.
These observations regarding the prognostic value of cell-in-cell structures raise
the possibility that this process has a physiological relevance in tumors regard-
ing tumor immunity, immune cell infiltration and metastatic disease. Future work
will have to address whether cell-in-cell events render the tumor milieu in a way
that affects patients’ outcome and if it might be used to guide treatment strate-
gies.
In conclusion, these study shows that removal of necrotic cells is a feature
common to several types of tumor cells as well as normal skin fibroblasts. It is
likely that the phagocytic activity by non-professional phagocytes contributes to
the efficient dead cell clearance by macrophages and other professional phago-
cytes. This process diminishes the release of proinflammatory damage-
associated molecular pattern molecules and therefore prevents inflammation
and autoimmune disease. Moreover, similar cell-in-cell structures in tumor tis-
sue are prognostic relevant.

16
1. Einleitung
Formen von Zelltod 1.1.Der Zelltod ist ein essentieller Prozess, der zur Erhaltung der zellulären Homö-
ostase in vielzelligen Organismen beiträgt. Es gibt verschiedene Modalitäten
des Zelltodes, die durch extrinsische oder intrinsische Signale bzw. physikali-
sche Einflüsse ausgelöst werden. Die einzelnen Zelltodesformen unterscheiden
sich nicht nur in ihrer Morphologie sondern auch in ihrer Wirkung auf Zellen des
Immunsystems. Nach Definition des Nomenklatur-Komitees für Zelltod gilt für
die Identifizierung von toten Zellen, dass ein irreversibler Verlust der Barriere-
Funktion der Zellmembran vorliegen muss und zusätzlich die Auflösung der Zel-
le in Vesikel (apoptotische Körperchen) oder die Aufnahme von toten Zellen
durch phagozytische Zellen stattfinden muss (Galluzzi, Maiuri et al. 2007,
Kroemer, Galluzzi et al. 2009, Galluzzi, Vitale et al. 2012). Nachdem jedoch
gezeigt werden konnte, dass aufgenommene Zellen unter bestimmten Umstän-
den lebend den Phagosomen entkommen können (Overholtzer, Mailleux et al.
2007), wurde die Definition für Zelltod erweitert. Demzufolge gelten Zellen nur
dann als tot, wenn entweder eine irreversible Permeabilität der Zellmembran
oder eine vollständige Fragmentierung der Zelle vorliegt. Weiterhin werden die
Zelltodes-Modalitäten in zwei Kategorien unterschieden, den passiven, nicht-
regulierten Zelltod (accidental cell death) und den regulierten Zelltod (regulated
cell death). Zur Gruppe des regulierten Zelltods zählt auch der programmierte
Zelltod (programmed cell death), der den eintretenden Zelltod bei der Embryo-
nal-Entwicklung, bei der Erhaltung der Homöostase im Gewebe und der Im-
munantwort beschreibt (Lettre and Hengartner 2006, Taylor, Cullen et al. 2008,
Fuchs and Steller 2011, Delbridge, Valente et al. 2012, Galluzzi, Bravo-San
Pedro et al. 2015). Der passive, nicht-regulierte Zelltod wird durch Einflüsse,
wie physikalische (erhöhte Temperaturen oder Druck), chemische (Detergenzi-
en, Veränderungen des pH-Wertes) und mechanische (Scherkräfte) Stimuli
verursacht und bedient sich keiner molekularen Maschinerie, sodass diese
Form von Zelltod weder durch pharmakologische noch genetische Interventio-
nen verhindert werden kann (Green and Kroemer 2005, Reed 2006,
MacFarlane 2009, Long and Ryan 2012). Der regulierte Zelltod hingegen wird
von einer genetisch kodierten, molekularen Maschinerie ausgeführt (Galluzzi,
Vitale et al. 2012, Galluzzi, Kepp et al. 2014).

17
Apoptose 1.1.1.
Die Apoptose ist eine programmierte Form des Zelltods und stellt den am häu-
figsten eintretenden biologischen Mechanismus zur Erhaltung der Homöostase
in vielzelligen Organismen dar und ist außerdem essentiell für die embryonale
Entwicklung. Sie wird durch extrazelluläre oder intrazelluläre Signale induziert.
Man spricht auch von der extrinsischen und intrinsischen Apoptose. Die extrin-
sische Apoptose wird durch extrazelluläre Stress-Signale ausgelöst, welche die
Zelle via Transmembranrezeptoren empfängt. Beispiele für auslösende Signale
sind die Bindung von Todesliganden (FasL/CD95L, TNFα, TRAIL) an Todesre-
zeptoren (Fas/CD95, TNFα-Rezeptor, TRAILR1-2) (Wajant 2002, Schutze,
Tchikov et al. 2008). Eine andere Art von Rezeptoren (dependence receptors)
leitet die Apoptose ein, wenn die Bindung an ihre spezifischen Liganden unter
eine kritische Konzentration fällt (Mehlen and Bredesen 2011). Die extrinsische
Apoptose resultiert immer in der Aktivierung von Caspase 3 und gilt somit als
Caspase-abhängige Apoptose, wohingegen die intrinsische Apoptose zusätz-
lich auch Caspase-unabhängig vollzogen werden kann (Yuan and Horvitz 1990,
Chautan, Chazal et al. 1999, Daish, Mills et al. 2004). Zu den intrazellulären
Signalen, welche die Apoptose induzieren zählen u.a. DNA-Schäden, oxidativer
Stress und Anhäufung von denaturierten Proteinen im endoplasmatischen Reti-
kulum.
Eine weitere Kondition, welche Apoptose hervorruft, gilt speziell für adhärente
Zellen. Diese gehen in Apoptose, wenn sie von der extrazellulären Matrix abge-
löst werden (Frisch and Francis 1994). Die molekulare Maschinerie gleicht zu-
meist der intrinsischen Apoptose (Reginato, Mills et al. 2003).
Apoptotische Zellen zeichnen sich durch Zell-Schrumpfung und Chromatin-
Kondensation (Pyknose) aus (Kerr, Wyllie et al. 1972). In der späteren Phase
des Zelltodes kommt es zur Bildung intrazellulärer Bläschen (membrane bleb-
bing) und Fragmentierung des Zellkerns (Karyorrhexis). Zuletzt schnürt die Zel-
le apoptotische Körperchen ab (Elmore 2007).
Primäre und sekundäre Nekrose 1.1.2.
Der nekrotische Zelltod wird in die regulierte (siehe 1.1.3) und die nicht-
regulierte oder primäre Nekrose unterschieden. Die primäre Nekrose stellt eine
passive Zelllyse dar, die durch extreme physikalische Einflüsse, welche die

18
Membranintegrität zerstören oder die intrazelluläre ATP-Konzentration reduzie-
ren, hervorgerufen wird (Leist, Single et al. 1997).
Des Weiteren kann nach induzierter Apoptose die sekundäre Nekrose eintreten,
wenn die apoptotischen Körperchen nicht rechtzeitig durch professionelle Pha-
gozyten beseitigt werden (Don, Ablett et al. 1977). Aufgrund der vorangegan-
genen Apoptose weisen sekundär nekrotische Zellen eine degradierte DNA auf,
wohingegen primär nekrotische Zellen eine vollständig intakte DNA enthalten.
Die übrigen morphologischen Charakteristika von nekrotischen Zellen sind eine
permeable Zellmembran, vergrößerte Organellen, die Dissoziation der Riboso-
men vom endoplasmatischen Retikulum und zuletzt der Zerfall der Kernmor-
phologie (Clarke 1990).
Regulierte Nekrose 1.1.3.
Die regulierte Nekrose gleicht morphologisch dem Erscheinungsbild der pri-
mären Nekrose und weist einen Verlust der Membran-Integrität sowie eine
Größenzunahme der Organellen und eine intakte DNA auf. Es wurden inzwi-
schen mehrere Zelltodes-Modalitäten von regulierter Nekrose identifiziert. Dazu
zählen die Nekroptose, Parthanatos, Ferroptose oder Oxytose, MPT-abhängige
Nekrose (mitochondrial permeability transition), Pyroptose und Pyronekrose
und NETose. Letztere Todesform erhielt ihre Bezeichnung, weil sie den Zelltod
beschreibt, der mit der Freisetzung neutrophiler extrazellulärer Fallen (neutro-
phil extracellular traps) aus Chromatin einhergeht.
Bisher am besten erforscht ist die Nekroptose. Sie wird durch die Ligation von
TNFα an den Rezeptor TNFR1 induziert und durch die Interaktion von RIPK3
(receptor interacting protein kinase 3) und seinem Substrat MLKL (mixed line-
age kinase like pseudokinase) vermittelt (Cho, Challa et al. 2009, Zhang, Shao
et al. 2009, Sun, Wang et al. 2012, Zhao, Jitkaew et al. 2012). Ähnlich der In-
duktion von Apoptose (siehe 1.1.1) kann die Nekroptose ebenfalls über die Bin-
dung von anderen Todesliganden, wie FasL oder TRAIL, an Todesrezeptoren
(Fas, TRAILR1, TRAILR2) induziert werden. Weil die Nekroptose ein Caspase-
unabhängiger Zelltod-Mechanismus ist, geschieht dies vor allem dann, wenn
die Apoptose aufgrund von Inhibition (vor allem von Caspase 8) oder Mutation
nicht stattfinden kann (He, Wang et al. 2009, Kaiser, Upton et al. 2011).

19
Autophagischer Zelltod 1.1.4.
Die Autophagie beschreibt eine ausgeprägte Vakuolenbildung im Zytoplasma
sterbender Zellen. Die Konversion von der zytoplasmatischen Form von LC3
(LC3-I, light chain 3) in ihre membranständige Form (LC3-II), die zur Membran
der autophagischen Vesikel (Autophagosomen) rekrutiert wird, kann als Marker
für die Induktion von Autophagie verwendet werden. Autophagosomen enthal-
ten zytoplasmatisches Material oder sogar ganze Organellen. Bei der Ver-
schmelzung dieser LC3-positiven autophagischen Vesikel mit Lysosomen
kommt es zur autophagolysosomalen Degradation und damit schließlich zum
Tod der Zelle. Doch nicht allein die Anzahl von Autophagosomen dient als Indi-
kator für die autophagosomale Aktivität, sondern der dynamische Prozess von
Autophagosomen-Bildung, Fusion mit Lysosomen und Abbau der autophagi-
schen Substrate (autophagic flux). Es wurde gezeigt, dass Tumorzellen, die
Apoptose-inkompetent sind, nach Behandlung mit Chemotherapeutika in den
autophagischen Zelltod gehen (Fazi, Bursch et al. 2008).
Die Funktion der Autophagie wurde aber zuerst als zytoprotektiv beschrieben.
Sie wird in sterbenden Zellen als Stress-Antwort aktiviert und soll den Zelltod
hinauszögern, sodass sich die Zelle regenerieren kann. Insbesondere bei Nähr-
stoffmangel inhibiert die autophagische Maschinerie die Induktion von Apopto-
se, vermutlich durch die Bereitstellung zelleigener Metaboliten. Eine Hemmung
der Autophagie verhindert dann nicht den Zelltod, sondern beschleunigt ihn
stattdessen (Boya, Gonzalez-Polo et al. 2005).
Daher ist der Begriff „autophagischer Zelltod“ nach Empfehlung des Nomenkla-
tur-Komitees für Zelltod nur anzuwenden, wenn bei Inhibition von Autophagie-
assoziierten Proteinen, der Zelltod verhindert wird (Denton, Nicolson et al.
2012, Galluzzi, Vitale et al. 2012).
Immunogenität der Zelltodesformen 1.1.5.
Sterbende Zellen können durch die Freisetzung und Exposition von sog. Gefah-
rensignalen (DAMPs, damage associated molecular patterns) eine Entzün-
dungsreaktion verursachen. DAMPs sind zelluläre Komponenten und Moleküle,
die von sterbenden und gestressten Zellen exponiert oder freigesetzt werden
und das Immunsystem alarmieren. Dazu zählen Zytokine und Alarmine, die vor
allem von sterbenden Zellen produziert werden, wie IL-1α und IL-33, und die

20
S100 Proteine. Beispiele für intrazelluläre Komponenten, die als DAMPs wirken,
sind u.a. Nukleinsäuren, Ribonukleoproteine, Histone, Proteine der HMGB Fa-
milie (high-mobility group box), Hitzeschockproteine, Calretikulin, filamentöses
Aktin (F-Aktin) und ATP (Krysko, Garg et al. 2012). Nach der Bindung von
DAMPs an spezifische Rezeptoren von Immunzellen (pattern recognition recep-
tors), wird die Expression und Sekretion von proinflammatorischen Zytokinen
und Chemokinen in Gang gesetzt.
Die Apoptose gilt dabei meist als nicht-inflammatorisch oder sogar antiinflamm-
atorisch, da bei diesem Prozess keine oder nur sehr wenige intrazelluläre Be-
standteile frei werden (Wickman, Julian et al. 2013). Die effiziente Beseitigung
apoptotisch sterbender Zellen durch Makrophagen verhindert die Induktion von
sekundärer Nekrose. Außerdem wird nach Phagozytose von apoptotischen Zel-
len nicht die Produktion von proinflammatorischen Zytokinen angeregt (Savill
and Fadok 2000). Makrophagen setzen sogar antiinflammatorische Zytokine
wie IL-10 und TGFβ frei (Voll, Herrmann et al. 1997).
Bei der Nekrose hingegen kommt es zur massiven Freisetzung von DAMPs,
die immunstimulatorisch und proinflammatorisch wirken (Kaczmarek,
Vandenabeele et al. 2013). Ein wichtiges Gefahrensignal aus der Proteinfamilie
der Hitzeschockproteine ist HSP70. Hitzeschockproteine sind Chaperone, die
für die korrekte Faltung von Peptidketten durch Stabilisierung sowie für die
Ubiquitinierung von beschädigten Proteinen, was deren Abbau im Proteasom
veranlasst, notwendig sind. Bei massiver Protein-Denaturierung aufgrund von
zellulärem Stress, wie beispielsweise nach Hitzeschock, wird ihre Expression
hochreguliert (Ashburner and Bonner 1979, Slater, Cato et al. 1981). Kommt es
zum nekrotischen Zelltod und damit ihrer Freisetzung sind Hitzeschockproteine
starke Aktivatoren von Antigen-präsentierenden Zellen, da sie durch ihre Funk-
tion als Chaperone, gebundene Proteine an die phagozytischen Antigen-
präsentierenden Zellen, wie dendritische Zellen (DCs), liefern.
DCs, die diese HSP70-Peptidkomplexe phagozytiert haben, präsentieren sie
auf MHC-II-Molekülen bzw. kreuzpräsentieren sie auf MHC-I-Molekülen. Akti-
vierte DCs wandern dann in die Lymphknoten ein und aktivieren in den T-Zell-
Regionen naive, Antigen-spezifische T-Zellen (Suto and Srivastava 1995). Die
Aktivierung der zytotoxischen T-Zellen kann Entzündungsreaktionen und Nek-
rosen (Hodge, Guha et al. 2008) oder sogar autoreaktive Reaktionen hervorru-
fen (Silva, do Vale et al. 2008), da beim nekrotischen Zell-Zerfall viele Autoanti-

21
gene freiwerden. Außerdem können auf diese Weise zytotoxische T-Zellen, die
gegen spezifische Tumorantigene gerichtet sind, aktiviert werden, wenn
HSP70-Tumorantigen-Komplexe durch Rezeptor-vermittelte Endozytose von
Antigen-präsentierenden Zellen aufgenommen und auf MHC-I-Molekülen
kreuzpräsentiert werden (Huang, Golumbek et al. 1994, Arnold-Schild, Hanau
et al. 1999).
Endozytose 1.2.Die Endozytose beinhaltet die Einstülpung und Abschnürung der Zellmembran,
sodass das gebundene Material vollständig von einer Vakuolenmembran um-
hüllt ist. Diese Vesikel werden als Endosomen bezeichnet. Man unterscheidet
die Aufnahme von flüssigem, löslichem Material (Pinozytose) von der Aufnahme
von größerem, festem Material (Phagozytose). Die bei der Pinozytose entste-
henden Endosomen werden auch Makropinosomen genannt, die Endosomen
der Phagozytose Phagosomen. Die Pinozytose wird meist mit der Endozytose
gleichgesetzt und von der Phagozytose abgegrenzt, sodass in diesem Zusam-
menhang die Bezeichnung Endosom statt Makropinosom gebräuchlich ist.
Pinozytose 1.2.1.
Nach Abschnürung des Endosoms von der Zellmembran folgt die sequentielle
Verschmelzung mit anderen Endosomen, sodass es zu einer fortschreitenden
Azidifizierung innerhalb des Vesikels und zur Anreicherung von bestimmten
Membranproteinen kommt. Frühe Endosomen weisen das Membranprotein
RAB-5 auf, späte Endosomen RAB-7. Diese Proteine sind GTPasen und ver-
mitteln die Fusion mit anderen Endosomen (Rink, Ghigo et al. 2005) und
schließlich mit sauren Lysosomen, die positiv für die Membranproteine LAMP1
und LAMP2 sind (Luzio, Rous et al. 2000). Lysosomen enthalten Hydrolasen
und Phosphatasen, die den enzymatischen Abbau des phagozytierten Materials
herbeiführen und dadurch die Abtötung von aufgenommenen Pathogenen, die
Beseitigung sterbender Zellen und die Bereitstellung von Nährstoffen für die
Zelle bewirken. Über den Vorgang der Exozytose werden anschließend die zer-
setzten Partikel durch Fusion der Vesikel (Exosomen) mit der Zellmembran aus
der Zelle ausgeschleust (Johnstone, Adam et al. 1987).

22
Wird die Endozytose durch Oberflächenrezeptoren induziert, handelt es sich um
die Rezeptor-vermittelte Endozytose. Dabei entstehen membranöse Einstül-
pungen, die das Protein Clathrin enthalten (clathrin-coated pits) und zu Clathrin-
haltigen Vesikeln (clathrin-coated vesicles) abgeschnürt werden. Daher wird
diese Arte der Endozytose auch Clathrin-vermittelte Endozytose genannt
(Pearse 1976, Goldstein, Anderson et al. 1979).
Eine wichtige Rolle bei der Endozytose spielt das Protein Dynamin. Die Genva-
riante Dynamin II wird ubiquitär von allen Körperzellen exprimiert (Sontag,
Fykse et al. 1994) und ermöglicht durch GTP-Bindung und -Hydrolyse die Ab-
schnürung der Endosomen, insbesondere der Clathrin-haltigen Vesikel (Damke,
Baba et al. 1994, Hinshaw 2000).
Phagozytose 1.2.2.
Die Erkennung von pro-phagozytischen Antigenen durch Oberflächenrezepto-
ren der Phagozyten induziert die Phagozytose. Ein Beispiel sind Fc-
Rezeptoren, die die Phagozytose von opsonisierten Partikeln stimulieren (siehe
1.3). Im Gegensatz zur Rezeptor-vermittelten Endozytose kommt es bei der
Phagozytose nicht zum „Einsinken“ des aufzunehmenden Partikels in die Zelle,
sondern zur Oberflächenvergrößerung der Zellmembran und zum Umschließen
des aufzunehmenden Materials mit Zellfortsätzen (Lamellipodien). Das Material
wird so in membranumhüllte Phagosomen ins Zytoplasma der Zelle aufgenom-
men. Die Reifung der Phagosomen erfolgt wie bei der Endozytose über RAB-5
und RAB-7 positive Stadien. Ein Unterschied liegt allerdings bei der GTPase
Regulation vor, sodass die Aneignung der Endosomen-Marker weniger durch
Fusion mit anderen Vesikeln zustande kommt, sondern mehr durch eigenstän-
dige Reifung der Phagosomen auftritt. Die zugrunde liegenden Mechanismen
wurden bislang nicht bis ins Detail geklärt, jedoch wurde für die Rekrutierung
von RAB-5 zum Phagosom eine Abhängigkeit von Dynamin identifiziert, das
selbst bereits zum Beginn der Phagozytose zur Aufnahmestelle (phagocytic
cup) transloziert wird (Kinchen, Doukoumetzidis et al. 2008). Zuletzt kommt es
wie auch bei der Endozytose zur Fusion dieser Vesikel mit Lysosomen zu Pha-
golysosomen und das aufgenommene Material wird enzymatisch gespalten.
Handelt es sich bei der phagozytischen Zelle um eine professionelle Antigen-
präsentierende Zelle, insbesondere eine DC, werden anschließend einzelne

23
Antigene der abgebauten Partikel auf MHC-II-Molekülen an der Zelloberfläche
präsentiert bzw. auf MHC-I-Molekülen kreuzpräsentiert, wodurch eine Antigen-
spezifische Immunantwort ausgelöst werden kann (den Haan, Lehar et al.
2000).
Eine Voraussetzung für die Phagozytose ist ein dynamisches Aktin-Zytoskelett,
das die Deformation der Zellmembran und die Formation der Lamellipodien er-
möglicht (Pollard and Cooper 2009). Auch in diesem Zusammenhang wurde
Dynamin eine Rolle als Verbindungsprotein zwischen Zellmembran und Aktin-
Zytoskelett zugeschrieben. Unter Ausbildung helikaler Polymere, die an Aktin-
bindende Proteine binden, sollen sie ein kontraktiles Gerüst zwischen Membran
und Zellskelett bilden und damit einen wichtigen Faktor für die Phagozytose
darstellen (Orth and McNiven 2003). Eine aktuellere Studie zeigte, dass bei In-
hibition von Dynamin die Bildung von Lamellipodien bei Makrophagen mit ein-
hergehender Destabilisierung der Aktinfilamente reduziert wurde (Yamada, Abe
et al. 2009).
Weitere Verbindungsproteine sind Proteine der Ezrin-Radixin-Moesin Familie.
Sie sind an der Regulation der Adhäsion, Membran-Motilität und Zytoskelett-
Reorganisation beteiligt. Die Phosphorylierung von Ezrin an Stelle T567 wird
durch eine aktivierte Rho-Kinase ausgeführt, bevor Fibrillenbündel (stress fi-
bers) und fokale Adhäsionen gebildet werden können (Matsui, Maeda et al.
1998).
Beseitigung sterbender Zellen durch Professionelle Pha-1.3.
gozyten Professionelle Phagozyten werden Phagozytose-fähige Zellen genannt, die mit
hoher Effizienz neben Mikroorganismen und Partikeln auch sterbende oder ver-
änderte Zellen aufnehmen. Zu diesen zählen DCs, Makrophagen, Monozyten,
Neutrophile und Mikroglia. Sie exprimieren verschiedene Rezeptoren
(scavenger receptors), die Pathogen-spezifische Oberflächenmoleküle, opsoni-
sierte Pathogene aber auch kommensale Bakterien und sterbende Zellen an
spezifischen Oberflächenantigenen erkennen (Stuart and Ezekowitz 2005).
Die Beseitigung apoptotischer Zellen verhindert den Eintritt der sekundären
Nekrose (siehe 1.1.2). Die Phagozytose nekrotischer Zellen ist äußerst wichtig,
da die freiwerdenden intrazellulären Moleküle bei der nekrotischen Zelllyse

24
schwere Erkrankungen, wie Autoimmunerkrankungen, neurodegenerative Er-
krankungen und Ischämie, hervorrufen können (Silva, do Vale et al. 2008).
Phagozytose apoptotischer Zellen 1.3.1.
Die Phagozytose von apoptotischen Zellen ist ein sehr häufiges Ereignis, da im
menschlichen Körper täglich ca. 50 – 70 Mio Zellen apoptotisch sterben und
ersetzt werden (Reed 2006). Durch die Ausschüttung von sogenannten “find-
me”-Signalen, wie geringe Konzentrationen von freigesetztem ATP und UTP,
Fraktalkin, Lysophosphatidylcholin und Sphingosin-1-Phosphat induzieren
apoptotische Zellen zunächst die Rekrutierung der professionellen Phagozyten
(Gregory 2009). Danach erkennt der Phagozyt durch die Bindung an spezifi-
sche Rezeptoren pro-phagozytische Signale, die auf der Zellmembran der ster-
benden Zelle exponiert werden, Diese Signale werden auch „eat-me“-Signale
genannt und stimulieren die Phagozytose.
Ein wichtiges pro-phagozytisches Signal, das die Phagozytose von apoptoti-
schen Zellen induziert, ist die Exposition von Phosphatidylserin (PS) auf der
Außenseite der Zellmembran. Die Asymmetrie von Zellmembranen geht bei der
Apoptose aufgrund von fehlender Enzymaktivität der Phospholipid-Flippasen,
welche in gesunden Zellen für die Lokalisation von PS auf der Innenseite der
Membran sorgen, verloren (Fadok, Bratton et al. 1998). Die Erkennung von PS
durch verschiedene PS-Rezeptoren stimuliert die Phagozytose der apoptoti-
schen Zellen bzw. der apoptotischen Körperchen durch Makrophagen (Fadok,
Voelker et al. 1992).
Ein weiteres pro-phagozytisches Signal stellt eine modifizierte Glykosylierung
der Zellmembran dar, die durch Lektine von phagozytischen Zellen erkannt
werden (Dini, Carla et al. 1999).
Die Bindung und Phagozytose von apoptotischen Körperchen durch Makropha-
gen wird außerdem durch den LPS-Rezeptor CD14, Scavenger-Rezeptor der
Klasse A, Thrombospondin-Rezeptor CD36, dem Vitronektin-Rezeptor αvβ3,
dem ABC1 Transporter und dem Makrophagen 61D3 Antigen induziert (Savill,
Hogg et al. 1992, Flora and Gregory 1994, Luciani and Chimini 1996, Platt,
Suzuki et al. 1996, Devitt, Moffatt et al. 1998).

25
Phagozytose nekrotischer Zellen 1.3.2.
Die Aufnahme von nekrotischen Zellen durch professionelle Phagozyten, wie
Makrophagen und Mikroglia-Zellen, wurde ebenfalls durch seine Abhängigkeit
von der Erkennung von exponiertem PS charakterisiert (Hirt and Leist 2003).
Die Tatsache, dass neben apoptotischen auch nekrotische Zellen PS auf der
Zelloberfläche exponieren können, war zuvor unbekannt. Die genannte Studie
lieferte den Beweis, indem die Zellen mit Biotin-gekoppeltem Annexin gefärbt
und anschließend mit Streptavidin-markierten magnetischen Kügelchen inku-
biert wurden. Dabei verhinderte die Größe dieser Kügelchen die Färbung von
PS auf der Innenseite der Zellmembran, selbst bei geschädigter, perforierter
Zellmembran.
Ein weiteres pro-phagozytisches Signal für die Erkennung von nekrotischen
Zellen ist F-Aktin. Dendritische Zellen exprimieren den Rezeptor DNGR-1, der
an F-Aktin bindet und die Induktion der T-Zell-vermittelten Immunität gegen An-
tigene von sterbenden Zellen in Gang setzt (Ahrens, Zelenay et al. 2012).
Als Rezeptor für Hitzeschockproteine, gp96 und Calretikulin gilt das von Makro-
phagen exprimierte Membranprotein CD91, das ebenfalls die Phagozytose von
nekrotischen Zellen stimuliert (Basu, Binder et al. 2001).
Nicht-professionelle Phagozyten und Zell-in-Zell Phänome-1.4.
ne Aber auch der Phagozytose-Aktivität von nicht-professionellen Phagozyten wird
inzwischen eine wichtige Rolle zugeschrieben. Nicht-professionelle Phagozyten
sind benachbarte Zellen, die sterbende Zellen aufnehmen, dabei aber in ihrer
Effizienz und dem exprimierten Rezeptor-Repertoire von den professionellen
Phagozyten abweichen. Im Zusammenhang mit der Aktivität von nicht-
professionellen Phagozyten steht der Mechanismus, der zur Internalisation von
einer Zelle in eine andere Zell führt und den Begriff „Zell-in-Zell Formation“ (cell-
in-cell formation oder CIC formation) definiert (Medicine 2014). Diese Zell-in-
Zell Phänomene wurden in Tumormaterial als Zell-Kannibalismus dokumentiert.
Außerdem kommen diese Zell-in-Zell Strukturen neben der nicht-
professionellen Phagozytose zur Aufnahme sterbender Zellen auch durch Inter-
nalisation von Lymphozyten in andere Zellen (Emperipolese) und der Invasion
von einer Tumorzelle in eine andere Tumorzelle (Entose) zustande.

26
Nicht-professionelle Phagozyten und Zell-Kannibalismus 1.4.1.
Die Fähigkeit zur Beseitigung von apoptotischen, autophagischen oder nekroti-
schen Zellen wurde inzwischen bei verschiedenen Zelltypen wie vaskulären
Myozyten (Kolb, Vranckx et al. 2007), Nierenepithelzellen (Ichimura, Asseldonk
et al. 2008), Mesenchymzellen (Wood, Turmaine et al. 2000) und epithelialen
Brustkarzinom-Zellen (Petrovski, Zahuczky et al. 2007) nachgewiesen. Als mo-
lekularer Mechanismus wurde in den erwähnten Studien die Bindung von expo-
niertem PS an PSRs, die von den nicht-professionellen Phagozyten exprimiert
werden, nachgewiesen. Außerdem wurde die Aufnahme von Mikroorganismen
durch nicht-professionelle Phagozyten auf die Expression von Fibronektin- und
Laminin-Rezeptoren oder der Exposition von Heparan-Sulfaten zurückgeführt
(Herrera, Ming et al. 1994). Ein großer Unterschied zwischen der Phagozytose-
Aktivität von professionellen und nicht-professionellen Phagozyten besteht da-
rin, dass den nicht-professionellen Phagozyten die Expression von Fc- und
Komplement-Rezeptoren fehlt. Die Phagozytose-Aktivität durch benachbarte
Zellen, sogenannte nicht-professionelle Phagozyten, stellt kein Merkmal für be-
stimmte Tumorzelltypen dar, sondern dient vielmehr der Unterstützung profes-
sioneller Phagozyten in verschiedenen Geweben.
Der Begriff “Zell-Kannibalismus” hingegen entstand bei der Beobachtung von
Zell-in-Zell Phänomenen in Tumorgewebeschnitten und Ergüssen. Diese Art
der Phagozytose wurde exklusive malignen Tumorzellen zugeschrieben und
beschreibt die Aufnahme von lebenden und apoptotischen Tumorzellen sowie
Lymphozyten durch andere Tumorzellen (Caruso, Muda et al. 2002, Lugini,
Lozupone et al. 2003, Fais 2007).
In Gewebe-Studien wurde einerseits eine Tumorfördernde Wirkung durch den
Zell-Kannibalismus publiziert, andererseits wurde der Beseitigung von Tumor-
zellen, die sich von der extrazellulären Matrix abgelöst haben, eine Schutzfunk-
tion vor Metastasierung zugesprochen (Kumar, Hosseinzadeh et al. 2001,
Abodief, Dey et al. 2006, Cano, Sandi et al. 2012). Die Tumorfördernde Wir-
kung wurde auf einen möglichen Überlebensvorteil von phagozytierenden Tu-
morzellen bei Nährstoffmangel zurückgeführt. So wurde eine Zunahme der
Phagozytose-Rate von lebenden CD8+-T-Lymphozyten durch metastatische
Melanom-Zellen bei Serummangel festgestellt (Lugini, Matarrese et al. 2006).
Daher wurde Zell-Kannibalismus als Marker für Malignität von Tumoren vorge-
schlagen (Gupta and Dey 2003). Wegen der widersprüchlichen Publikationen,

27
die zum einen eine Tumor-fördernde Wirkung aber auch eine Schutzfunktion
vor Metastasierung vorschlagen, kann die Anzahl von Zell-in-Zell Phänomenen
in Tumorgewebeschnitten nicht als prognostischer Faktor herangezogen wer-
den. Die Ursache für die gegensätzlichen Forschungsergebnisse liegt möglich-
erweise in spezifischen Unterschieden zwischen den betroffenen Geweben und
Krebsarten.
Emperipolese 1.4.2.
Der Begriff „Emperipolese“ benennt die invasive Aktivität von Lymphozyten in
das Zytoplasma größerer Zellen und wurde bereits 1956 von Humble et al. defi-
niert. Dieses Verhalten der Lymphozyten wurde bei Tumorzellen (Humble,
Jayne et al. 1956) und Makrophagen beobachtet (Berman 1966). Der Mecha-
nismus dieses Vorgangs beruht auf einer aktiven Invasion und Aufnahme der
Lymphozyten in Vakuolen innerhalb des Zytoplasmas einer größeren Wirtszelle,
wobei beide Zellen zu Beginn des Prozesses keine Anzeichen von Zelltod zei-
gen.
Emperipolese ist nicht mit der transendothelialen Migration von Leukozyten, die
im Rahmen einer Entzündungsreaktion auftritt, zu verwechseln. Diese findet
zwar hauptsächlich parazellulär an Zell-Zell-Verbindungen des Endothels statt,
kann aber auch transzellulär durch die Bildung von Kanälen aus Vesikeln (vesi-
culo-vacuolar organelles) geschehen (Feng, Nagy et al. 1998, Dvorak and Feng
2001). Der ausschlagegebende Unterschied zur Emperipolese bleibt jedoch die
Tatsache, dass die Lymphozyten lebend die Wirtszelle verlassen.
Eine aktuellere Studie belegte die Internalisation von natürlichen Killerzellen
(NK-Zellen) in verschiedene epitheliale Tumorzelllinien, die entweder im
apoptotischen Zelltod der NK-Zellen oder in der Lyse der Tumorzellen resultier-
te (Wang 1987, Wang, Guo et al. 2009). Ebenso wurde die Tumorzytotoxizität
einer T-Zelllinie auf den Vorgang der Emperipolese zurückgeführt, indem die
invasiven T-Zellen im Zytoplasma der Tumorzelle ihre toxische Granula freisetz-
ten (Takeuchi, Inoue et al. 2010).
Da NK-Zellen und T-Zellen durch ihre Fähigkeit zur Zelltod-Induktion in Tumor-
zellen eine wichtige Rolle im Rahmen der Tumorimmunität spielen (Dunn,
Bruce et al. 2002, Stinchcombe and Griffiths 2007, Vesely, Kershaw et al.

28
2011), kann die Emperipolese eine Immunevasion und Tumorprogression för-
dern.
Eine weitere Studie zeigte das invasive Verhalten von autoreaktiven CD8+-T-
Zellen in Hepatozyten, wobei es bei Inhibition der Emperipolese zur Hepatitis
kommt (Benseler, Warren et al. 2011). In diesem besonderen Fall dient die Em-
peripolese also der Verhinderung von Autoimmunerkrankungen.
Entose 1.4.3.
Als weiterer Mechanismus zur Bildung von Zell-in-Zell Phänomenen beinhaltet
die Entose die invasive Aktivität von Zellen, bei welcher Adhäsionsmoleküle von
der eindringenden Zelle, sowie der sogenannten Wirts-Zelle beteiligt und not-
wendig sind. Als Auslöser für die Entose gilt das Ablösen von lebenden Zellen
von der extrazellulären Matrix. Die Interaktion von E-Cadherinen und P-
Cadherinen sowie die Rho-ROCK Aktivität der aufzunehmenden Zellen spielen
dabei eine essentielle Rolle. Die internalisierten Zellen werden anschließend
entweder durch die Wirtszelle lysosomal abgebaut, durchlaufen eine Zellteilung
oder entkommen der Wirtszelle sogar lebend (Overholtzer, Mailleux et al.
2007). Als mögliche Folge der Entose gilt die Induktion von Aneuploidie in Tu-
morzellen (Krajcovic, Johnson et al. 2011).
Zielsetzung 1.5.Die Beseitigung nekrotisch sterbender Zellen ist essentiell für die Vermeidung
von Entzündungsreaktionen und Autoimmunerkrankungen. Die Beteiligung von
benachbarten, nicht-professionellen Zellen, die professionelle Phagozyten un-
terstützen ist bekannt, aber deren physiologische Rolle und molekulare Hinter-
gründe noch nicht bis ins Detail geklärt.
In dieser Dissertation sollte die Phagozytose von Tumor- und Normalgewebe-
zellen durch nicht-professionelle Phagozyten charakterisiert und deren Effizienz
studiert werden. Dabei stand insbesondere die Beseitigung von primär nekroti-
schen Zellen nach Hitze-Behandlung im Fokus dieser Arbeit. Zunächst sollten
die Effekte von Hyperthermie aber auch von ionisierender Strahlung zur Induk-
tion von Nekrose näher untersucht werden.
Des Weiteren sollte die Frage beantwortet werden, ob die Phagozytose-Aktivität
ein weit verbreitetes Phänomen oder nur wenigen Zelltypen eigen ist. Daher

29
wurden verschiedene Zelltypen verwendet, inklusive Pankreaskarzinom-Zellen,
mehrere morphologisch divergente Glioblastom-Zelllinien, eine primäre Kopf-
Hals-Tumorzelllinie, primäre, gesunde Hautfibroblasten, semi-adhärente Nie-
renepithelzellen, T-Zell-Leukämiezellen sowie lymphoblastoide Zellen. Mittels
Immunfluoreszenz-Färbung sollten Phagozytose-assoziierte Proteine identifi-
ziert und eine mögliche Akkumulation bestimmter Proteine, wie Adhäsionsmo-
leküle, lokalisiert sowie das Schicksal der aufgenommenen Zelle dokumentiert
werden. Funktionelle Analysen zur Beteiligung der Zytoskelett-Reorganisation,
GTPase Aktivität von Dynamin II, der Cadherin-vermittelten Adhäsion und der
Erkennung von PS sollten die molekularen Mechanismen charakterisieren.
Da die resultierenden Phagozytose-Ereignisse als transiente Zell-in-Zell Struk-
turen dem Zell-Kannibalismus in Tumorgewebeschnitten morphologisch ähnel-
ten, sollte außerdem das Vorhandensein von Zell-in-Zell Strukturen in Tumor-
gewebeschnitten von Patienten mit HNSCC, Rektumkarzinom und Analkarzi-
nom quantifiziert werden und eine mögliche prognostische Relevanz überprüft
werden.
30
2. Material und Methoden
Verwendete Zelllinien, Reagenzien, Geräte und Ver-2.1.
brauchsmaterial Tabelle 1: Humane Zelllinien
Kurzbezeichnung Quelle/Bezug Pathologie/Quelle Morphologie BxPC-3 ECACC,
93120816 Adenokarzinom, Pankreas
epithelial
T98G ATCC, CRL-1690 Glioblastom Fibroblasten U-87 MG ATCC, HTB-14 Glioblastom epithelial U-138 MG ATCC, HTB-16 Glioblastom polygonal U-251 MG CLS, 300385 Astrozytom epithelial Jurkat ATCC, TIB-152 T-Zell-Leukämie T-lymphoblastisch HEK293 ATCC, CRL-1573 Embryonale Niere epithelial p26 Biopsie Kopf-Hals-Tumor Epithel-ähnlich MZF16 Spender Hautstanze Fibroblasten SBLF5 Spender Hautstanze Fibroblasten SBL1 Spender Blut B-lymphoblastoid Tabelle 2: Oligonukleotide
Zielgen Referenz-sequenz
Vorwärts-Oligonukleotid
Rückwärts-Oligonukleotid
Amplikon [bp]
CD36 NM_001127444.1
TGCAAAGAAGG-GAGACCTGTG
GTTGAC-CTGCAGCCGTTTTG
174
CD68 NM_001040059.1
TTCTCTCATT-CCCCTATGGACAC
GCCATGTAGCTCA-GGTAGACAAC
82
GIGYF2 NM_001103148
GTGCACTAGAT-GATGAAAGATTGG
TGTAATACCACTT-CTGCATTGCTT
102
ITGB5 NM_002213.4
TGCATCCAACCA-GATGGACTA
CTCCACCGTTGTT-CCAGGTA
148
L3MBTL2 NM_031488
GAAGGTA-TA-CGAGCAGTCTACACAGA
TCCAGGAGAACCT-TACAGACAGTT
120
RPUSD3 NM_001142547.1
AAGACTCTCAGT-CACTTTCGTGTG
CCTGTAGTTGACTG-GAGAACACTG
94
TIM1 NM_005435.3
GCACGGAGACT-CAAAGTGAAAAG
CGGCTTT-CTCCAGTGCCAAT
156

31
Tabelle 3: Reagenzien
Bezeichnung Hersteller 7-AAD BD Becton, Dickinson and
Company Heidelberg, D
Accutase Millipore Darmstadt, D Aceton Carl Roth Karlsruhe, D Agarose Carl Roth Karlsruhe, D Annexin V Invitrogen GmbH Darmstadt, D Annexin V-APC BD Becton, Dickinson and
Company Heidelberg, D
Bafilomycin A1 Millipore Darmstadt, D BCA kit Thermo scientific Rockford, USA β-Mercaptoethanol Merck KGaA Darmstadt, D BrdU Sigma-Aldrich Taufkirchen, D BSA Serva Electrophoresis
GmbH Heidelberg, D
CASY Ton Innovatis AG Reutlingen, D CC1-Puffer (Benchmark UL-TRA)
Roche Grenzach-Wyhlen, D
CellLight Aktin-GFP Invitrogen GmbH Darmstadt, D CellLight H2B-RFP Invitrogen GmbH Darmstadt, D CellTrace Far Red, DDAO-SE
Invitrogen GmbH Darmstadt, D
CellTrace Oregon Green, carboxy-DFFDA SE
Invitrogen GmbH Darmstadt, D
Chloroquin Sigma-Aldrich Taufkirchen, D Colcemid Invitrogen GmbH Darmstadt, D DAPI Roche Grenzach-Wyhlen, D Dulbecco’s DMEM-Medium PAN Biotech Aidenbach, D DMSO Carl Roth Karlsruhe, D Dynasore Merck KGaA Darmstadt, D EdU imaging kit Invitrogen GmbH Darmstadt, D EDTA Merck KGaA Darmstadt, D Eisessig Merck KGaA Darmstadt, D F12-Medium Life Technologies GmbH Darmstadt, D FBS Biochrom AG Berlin, D Formaldehyd Merck KGaA Darmstadt, D Hoechst 33342 Molecular Probes Frankfurt, D HSP70-ELISA Duoset IC R&D Systems Wiesbaden, D Isoton II Beckman Coulter Krefeld, D Latrunculin B Merck KGaA Darmstadt, D L-Glutamin Life Technologies GmbH Darmstadt, D Methanol Merck KGaA Darmstadt, D Methylenblau-Färbelösung Sigma-Aldrich Taufkirchen, D Natriumazid Merck KGaA Darmstadt, D Natriumcitrat Merck KGaA Darmstadt, D Natriumchlorid Carl Roth Karlsruhe, D Nicht-essentielle Aminosäu- Biochrom AG Berlin, D
32
ren Paclitaxel Universitätsklinikum Er-
langen Erlangen, D
PBS Life Technologies GmbH Darmstadt, D Penicillin/Streptomycin Life Technologies GmbH Darmstadt, D QIA shredder Qiagen Hilden, D qPCR Kit DyNAmo Flash SYBR Green
Thermo Scientific Waltham, MA, USA
Reverse Transkription Kit QuantiTect
Qiagen Hilden, D
Rhodamin phalloidin Invitrogen GmbH Darmstadt, D Ringerlösung B. Braun Melsungen AG Melsungen, D RNeasy Kit Qiagen Hilden, D Roti Safe Gel Stain Carl Roth Karlsruhe, D RPMI 1640 Medium Sigma-Aldrich Taufkirchen, D Salzsäure Merck KGaA Darmstadt, D Streptavidin-biotinylierter alkalische Phosphatase-Komplex
DakoCytomation Glostrup, DK
SYBR-DNA-Farbstoff Lonza Basel, CH TRIS Carl Roth Karlsruhe, D Triton X-100 Merck KGaA Darmstadt, D Trypsin Life Technologies GmbH Darmstadt, D Tween Carl Roth Karlsruhe, D Vectashield Vector Laboratories Peterborough, UK Wasser für Molekularbiologie AppliChem Darmstadt, D Tabelle 4: Antikörper
Antikörper Spezies Verdünnung Hersteller BrdU Maus 1:6 Millipore, Temecula, CA CD2 Maus 1:20 eBioscience, Frankfurt, D Cleaved caspase 3 Kaninchen 1:100 Cell signaling, Danvers
MA, USA Ezrin BD, Heidelberg, D pT567Ezrin Maus 1:100 BD, Heidelberg, D E-Cadherin Maus 1:200 BD, Heidelberg, D E-Cadherin, Funktions-blockierend
Ratte 1:6,6 Millipore, Temecula, CA
HSP70 Kaninchen 1:100 Cell signaling, Danvers MA, USA
HSP70 Maus 1:100 BD bioscience, Heidel-berg, D
Isotypkontrolle IgG1 Maus Entsprechend eBioscience, Frankfurt, D Isotypkontrolle IgG2b Maus Entsprechend eBioscience, Frankfurt, D Isotypkontrolle IgG1 Ratte Entsprechend eBioscience, Frankfurt, D Isotypkontrolle, IgG1κ, Funktions-blockierend
Ratte Entsprechend eBiocience, Frankfurt, D
33
Isotypkontrolle, IgG1κ, Funktions-blockierend
Maus Entsprechend eBiocience, Frankfurt, D
LAMP1 Maus 1:100 eBiocience, Frankfurt, D LC3B Kaninchen 1:400 Cell Signaling, Danvers
MA, USA P-Cadherin Kaninchen 1:100 Cell Signaling, Danvers
MA, USA P-Cadherin, Funktions-blockierend
Maus 1:10 Abcam, Cambridge, UK
Tabelle 5: Sekundär-Antikörper
Antikörper Spezies Verdünnung Hersteller Alexa Fluor 594 gegen Kaninchen
Hühnchen 1:200 Invitrogen Darmstadt, D
Alexa Fluor 488 gegen Maus
Ziege 1:400 Invitrogen Darmstadt, D
Alexa Fluor 555 gegen Maus
Ziege 1:200 Invitrogen Darmstadt, D
Alexa Fluor 647 gegen Kaninchen
Ziege 1:200 Invitrogen Darmstadt, D
Rhodamin gegen Maus Ziege 1:100 Merck Millipore, Billeri-ca, MA, USA
Tabelle 6: Verbrauchsmaterial
Bezeichnung Hersteller CASY-Röhrchen Innovatis AG Reutlingen, D Deckgläser Gerhard Menzel GmbH Braunschweig, D Einfrierröhrchen NUNC Thermo Fisher Scientific Langenselbold, D Fettstift Super PAP Pen Science Services München, D PCR-Platten, Hartschale Bio-Rad München, D Mikrotiterplatten Cellstar Greiner Bio-One GmbH Frickenhausen, D Multiwellplatten, 96 Well Thermo Scientific Waltham, MA, USA Objektträger Gerhard Menzel GmbH Braunschweig, D Parafilm Pechiney Plastic Packaging Greenwitch, USA Petrischalen Pipetten
Thermo Fisher Scientific nerbeplus
Langenselbold, D Winsen/Luhe, D
Pipettenspitzen Sarstedt Nümbrecht, D Pipettenspitzen mit Filter Greiner Bio-One GmbH Frickenhausen, D Quadriperm-Schalen Greiner Bio-One GmbH Frickenhausen, D Reaktionsgefäße Eppendorf AG Hamburg, D Reaktionsgefäße für PCR Sarstedt Nümbrecht, D Rundbodenröhrchen Becton, Dickinson GmbH Heidelberg, D Zellkulturflaschen Greiner Bio-One GmbH Frickenhausen, D Zentrifugenröhrchen Cel-lstar, Polypropylen
Greiner Bio-One GmbH Frickenhausen, D

34
Tabelle 7: Geräte
Gerät Hersteller Analysewaage Mettler AC 100
METTLER-TOLEDO Giessen, D
Analysewaage P1200N METTLER-TOLEDO Giessen, D CASY TT Zellzähler Innovatis AG Reutlingen, D CFX96 Real-Time System Bio-Rad München, D CO2-Inkubator HERAEUS Instruments GmbH Hanau, D DNA/RNA UV-cleaner UVC/T-M-AR
Biosan Riga, LV
Durchflusszytometer Galli-os TM
Beckman Coulter Brea, USA
Elektrophoresesystem Power Pac HC
Bio-Rad München, D
Fluoreszenzmikroskop Axi-oplan 2 Imaging
Carl Zeiss Microscopy GmbH Göttingen, D
Fluoreszenzmikroskop Lei-ca DM6000B
Leica Microsystems Ltd.
Konvokales Mikroskop LSM 710
Carl Zeiss Microscopy GmbH Göttingen, D
Lebendzell-Mikroskop Axi-oplan 200
Carl Zeiss Microscopy GmbH Göttingen, D
Lichtmikroskop PrimoVert Carl Zeiss Microscopy GmbH Göttingen, D Magnetrührer MR2002 Heidolph Instruments GmbH Schwabach, D Nanophotometer P330 Implen GmbH München, D pH-Meter C761 Calimatic KNICK Berlin, D Plattformschüttler Rotamax 120
Heidolph Instruments GmbH Schwabach, D
Plattenlesegerät HTS 7000 Perkin Elmer Waltham, MA, USA Röntgenröhre Isovolt Titan GE Inspection Technologies Hürth, D Tischzentrifuge 1-15 Sigma Laborzentrifugen GmbH Osterode am Harz, D Tischzentrifuge Mikro 200 R
Hettich AG Bäch, CH
UV-Tisch UVT-28M Herolab GmbH Wiesloch, D Ventana BenchMark Ultra stainer
Roche Grenzach-Wyhlen, D
Vortex MR2000 Heidolph Instruments GmbH Schwabach, D Wasserbad GFL Gesellschaft für Labortech-
nik GmbH Burgwedel, D
Werkbank Hera Safe HERAEUS Instruments GmbH Hanau, D Werkbank Laminar-Flow HERAEUS Instruments GmbH Hanau, D Zentrifuge Megafuge 1.0RS Thermo Scientific Waltham, MA, USA Zentrifuge Rotina 420 R Hettich AG Bäch, CH Zytofuge 2 Stat Spin StatSpin Inc. Iris Sample Pro-
cessing Westwood, MA, USA
35
Tabelle 8: Puffer und Lösungen
Name Reagenz Volumen/Gewicht/Konzentration Blockier-Lösung FBS 10 ml
BSA 1 g PBS 90 ml Natriumazid (1 M) 0,3 ml
DAPI-Gebrauchslösung 4x SSC Tween 1 ml DAPI-Stammlösung (1 mg/ml)
0,3 µl
Fixier-Lösung PBS, 1x 89,2 ml Formaldehyd (37 %) 10,8 ml Triton X 100 µl
Methylenblau-Färbelösung
Methylenblau 30 ml KOH (1 %) 1 ml Aqua dest. 100 ml
RIPA-Puffer TRIS pH=8,0 50 mM Natriumchlorid 150 mM Deoxycholat 0,5 % NP40 1,0 % SDS 1,0 % HALT 1:100 PMSF 1:100, 0,1 M NaF 1:1000, 1 M Ortho-Vanadat 1:1000, 200 mM Beta-Glycero-Phosphat 1:1000, 1 M SSC (20x), autoklaviert Natriumchlorid 525,9 g
Natriumcitrat 264,6 g Aqua dest. Auf 3 L auffüllen Salzsäure (5 M) pH-Wert von 7,0 einstellen
SSC (4x)/Tween SSC (20x) 130 ml Aqua dest. 520 ml Tween 1,3 ml Salzsäure (1 M) pH-Wert von 7,0 einstellen
TAE (50x) Tris 242 g Eisessig 57,1 ml EDTA (0,5 M, pH 8,0) 100 ml Aqua dest. Auf 1 L auffüllen
TBS (10x) Tris 12,114 g Natriumchlorid 87,66 g Aqua dest. Auf 1 L auffüllen Salzsäure (5 M) pH-Wert von 8,0 einstellen

36
Zellkultur 2.2.Das Arbeiten mit den Zelllinien fand unter sterilen Bedingungen in einer sterilen
Werkbank statt.
Kultivierungsbedingungen 2.2.1.
Die Zellen wurden in einem Brutschrank bei 37 °C, 5 % CO2 und einer relativen
Luftfeuchtigkeit von 95 % inkubiert. Für die Kultivierung wurden Zellkulturfla-
schen aus Polystyrol mit Filterschraubverschluss benutzt. Je nach Zelllinie wur-
den komplementierte Kulturmedien basierend auf F12, RPMI 1640 oder Dulbe-
cco‘s DMEM verwendet. Tabelle 9 zeigt, welche Zusätze zu den Medien gege-
ben wurden. Tabelle 9: Kulturmedien
BxPC-3, HEK293 T98G, U87, U138 U251, HEK293
p26, SBLF5 MZF16
Jurkat, SBL1
RPMI 1640 DMEM F12 RPMI 1640 10 % FBS 10 % FBS 15 % FBS 20 % FBS 100 u/ ml
Penicillin 100 u/ ml
Penicillin 100 u/ ml
Penicillin 100 u/ ml
Penicillin
100 µg/ml
Strepto-mycin
100 µg/ml
Strepto-mycin
100 µg/ml
Strepto-mycin
100 µg/ml
Strepto-mycin
2 % L-Glutamin
2 % L-Glutamin
2 % L-Glutamin
1 % Nicht-essentiel-le Amino-säuren
Passage und Konservierung von Zellen 2.2.2.
Die Suspensionszelllinien (Jurkat und SBL1) wurden mit einer Zelldichte von
0,2 – 1*106 Zellen/ml kultiviert. Um diese Zelldichte für optimale Wachstumsbe-
dingungen zu erhalten, wurden die Zellen ein- bis zweimal wöchentlich gezählt
und entsprechend ein Teil der Zellen entfernt bevor anschließend frisches Me-
dium zugegeben wurde.
Alle adhärent wachsenden Zelllinien wurden ein- bis zweimal wöchentlich nach
Erreichen einer Konfluenz von 90 – 100 % abgelöst. Hautfibroblasten wurden
im Verhältnis 1 zu 1, alle anderen adhärenten Zelllinien im Verhältnis 1 zu 10

37
passagiert. Dazu wurde das Medium komplett entfernt, der Zellrasen mit PBS
gespült und mit Trypsin überschichtet. Nach einer Einwirkzeit von 1 – 2 min
wurde das Trypsin abgenommen und die Zellkulturflasche für weitere 1 – 5 min
in den CO2-Inkubator gegeben. Das Ablösen und gegebenenfalls Vereinzeln
der Zellen wurde im Lichtmikroskop überprüft bevor die Zellen in Medium auf-
genommen und auf die Kulturflaschen verteilt wurden. Zuletzt wurde ausrei-
chend frisches Medium zugegeben.
Für die Konservierung wurden Zellen genutzt, die eine maximale Konfluenz von
80 % erreicht hatten. Nach Trypsin-Behandlung wurden die Zellen in 10 ml Me-
dium aufgenommen und in Zentrifugenröhrchen überführt. Nach Zentrifugation
bei 300 g für 5 min wurde der Überstand verworfen und das Zellsediment in
sterilfiltriertem RPMI 1640 Medium mit 11 % Glycerin resuspendiert. Je Kryo-
röhrchen wurde eine Zellsuspension mit 1 – 5*106 Zellen überführt, die Röhr-
chen in einen mit Isopropanol gefüllten Einfrierbehälter gegeben und bei −80 °C
eingefroren. Nach 24 h wurden die Röhrchen dem Einfrierbehälter entnommen
und in Kisten gelagert.
Transfektion 2.2.3.
BxPC-3 Zellen wurden mit den Transfektions-Reagenzien CellLight Aktin-GFP
und CellLight Histon H2B-RFP transfiziert. Es wurden 0,85*105 Zellen je Näpf-
chen einer Kulturplatte mit zwölf Näpfchen eingesät. Nach 24 Stunden Inkubati-
on im CO2-Brutschrank wurde das Medium restlos entfernt, bevor 720 µl Medi-
um und 80 µl des CellLight-Reagenz‘ (entspricht 95 Partikeln pro Zelle) zuge-
geben wurden. Die Platten wurden kurz geschwenkt und für weitere 26 h inku-
biert. Anschließend wurde das Medium entfernt, der Zellrasen mit PBS gewa-
schen und die Zellen mit Trypsin abgelöst. Nach Aufnahme in frisches Medium
wurden die Zellen entweder zur Analyse der Transfektions-Effizienz auf Objekt-
träger aufgebracht oder im Phagozytose-Versuch verwendet.
Nukleosid-Inkorporation 2.2.4.
Der Einbau von Nukleosiden zur Markierung der Zellkerne vor Hitzebehandlung
und Internalisierung in phagozytische Zellen erfolgte mit EdU bzw. BrdU. Es
wurden 1*105 Zellen in Kulturplatten mit sechs Näpfchen eingesät. Nach 24 h
wurde 30 µM EdU bzw. 100 µM BrdU zugegeben. Nach weiteren 72 h wurden

38
die Zellen abgelöst und im Phagozytose-Versuch verwendet. Inkorporiertes
BrdU wurde mit einem Primärantikörper gegen BrdU und einem Fluorochrom-
markierten Sekundärantikörper nachgewiesen (siehe 2.6). EdU wurde mithilfe
des EdU imaging kit nach Herstellerangaben sichtbar gemacht.
Induktion und Inhibition von nicht-professioneller Phagozy-2.3.
tose Zur Untersuchung der nicht-professionellen Phagozytose wurde ein Standard-
protokoll etabliert. Zwei Tage vor dem Versuch wurden die Zellen in Flaschen
eingesät, wobei die Zellzahl je nach Zelllinie variierte. Entscheidend dabei war,
dass zum Versuchstag eine Konfluenz von maximal 90 % erreicht wurde. Am
Versuchstag wurden die Zellen mit Lebendfarbstoffen grün und rot gefärbt, an-
schließend abgelöst, gezählt und im Verhältnis 1 zu 1 für eine Dauer von 4 –
72 h koinkubiert. Je nach Versuchsbedingung erfolgte die Behandlung zur In-
duktion bzw. Inhibition der Phagozytose vor oder nach dem Ablösen der Zellen.
Zum Beginn und nach 60 min wurde mittels Durchflusszytometrie die Adhäsion
von grünen und roten Zellen gemessen. Nach Ablauf der Koinkubation wurden
die Zellen auf Objektträger zentrifugiert, fixiert und mikroskopiert.
Inaktivierung von Zellen durch Hitzeeinwirkung 2.3.1.
Die Effekte von Hitzeeinwirkung auf Pankreaskarzinom-Zellen wurden unter-
sucht, indem die Zellen 45 min im Wasserbad bei Temperaturen von 46 – 56 °C
(2 °C-Unterschiede) inkubiert wurden und anschließend zu den Zeitpunkten 0 h,
4 h und 24 h für eine Analyse mit dem Durchflusszytometer mit Annexin V-APC
und 7-AAD gefärbt wurden (siehe 2.4.1). Die Inkubation der Zellen für die Zeit-
punkte 4 h und 24 h erfolgte in Suspensionswellplatten im CO2-Inkubator. Für
alle weiteren Versuche zur Phagozytose von Hitze-behandelten Zellen wurde in
dieser Arbeit die Inkubation bei 56 °C für 45 min gewählt.
Proliferationspotenzial von Hitze-behandelten Zellen 2.3.2.
Zur Untersuchung der Auswirkung von Hitze-Behandlung auf die Proliferations-
fähigkeit der Zellen wurde ein Koloniebildungstest durchgeführt. Dabei wurden
Pankreaskarzinom-Zellen abgelöst und mit der Neubauer Zählkammer gezählt.

39
Dabei wurde darauf geachtet, dass die Zellen zuvor vereinzelt wurden. Die Ein-
zelzellsuspension wurde auf vier Zentrifugenröhrchen verteilt. Für die Kontrolle
wurden die Zellen sofort und in definierter Menge (400 Zellen) in Petrischalen
(Ø 6 cm) eingesät. Die übrigen drei Röhrchen wurden für 45 min bei 40, 45 oder
56 °C im Wasserbad inkubiert. Nach der Hyperthermie-Behandlung wurden
ebenfalls jeweils 400 Zellen in Petrischalen eingesät. Es wurden für jede Bedin-
gung und die Kontrolle jeweils Triplikate angefertigt. Beim Einsäen wurde 3 ml
Medium auf die Zellen pipettiert, nach drei und zehn Tagen wurde jeweils 1 ml
Medium zugegeben. Nach 14 Tagen wurden die Kolonien mit einer Methylen-
blau-haltigen Lösung 30 min gefärbt, mit destilliertem Wasser gewaschen, ge-
trocknet und gezählt.
Vitalfärbung 2.3.3.
Die phagozytierenden und die aufzunehmenden Zellen wurden mittels Vitalfär-
bung grün bzw. rot gefärbt. Dafür wurden Celltrace Vitalfarbstoffe verwendet,
die durch die intakte Zellmembran diffundieren und kovalent an Amingruppen
intrazellulärer Proteine binden. Der grüne Farbstoff entwickelt seine Fluores-
zenz erst nach Entfernung der Azetatgruppen durch zelleigene Esterasen, der
rote Farbstoff hingegen muss nicht mehr enzymatisch umgesetzt werden. Die
Vitalfarbstoffe bleiben in den gefärbten Zellen und können nicht auf andere Zel-
len übertragen werden, sodass unterschiedlich behandelte Zellen verlässlich
unterschieden werden können. Zunächst wurde durch Zugabe von 19,8 µl
DMSO zum lyophilisierten Sediment von CTOG (grün) eine 4,255 mM Stocklö-
sung hergestellt. CTFR (rot) wurde mit 16,8 µl DMSO gelöst, um eine 5,88 mM
Stocklösung zu erhalten. Für die Vitalfärbung wurden die Farbstoffe in je nach
Kulturflaschengröße in 5 – 15 ml PBS verdünnt, sodass die Endkonzentration
für CTOG 0,596 µM und für CTFR 1,882 µM betrug. Bei der Färbung von adhä-
renten Zellen wurde das Medium verworfen und der Zellrasen mit PBS gewa-
schen. In eine T-25 Kulturflasche wurden 5 ml Färbelösung pipettiert, in eine T-
75 Kulturflasche 10 ml und in eine T-225 15 ml. Die Zellen wurden mit der Fär-
belösung 20 min bei 37 °C im CO2-Schrank inkubiert. Anschließend wurde die
Färbelösung entfernt, der Zellrasen mit Medium gewaschen und zuletzt mit Me-
dium überschichtet. Für die Umwandlung des grünen Farbstoffs wurden die Zel-
len weitere 10 min im CO2-Schrank inkubiert. Suspensionszellen wurden in ein

40
Zentrifugenröhrchen überführt, mit dem Zellzähler CASY TT gezählt und bei
300 g 5 min zentrifugiert. Die Zellen wurden in 5 ml Färbelösung resuspendiert
und 15 min im CO2-Inkubator inkubiert. Anschließend wurden die Zellen nach
Zentrifugation (5 min, 300 g) in Medium resuspendiert und dabei die Zellkon-
zentration von 106 Zellen pro ml eingestellt. Die grün gefärbten Zellen wurden
für weitere 10 min bei 37 °C inkubiert.
Phagozytose von hyperthermierten Zellen 2.3.4.
Die Phagozytose wurde durch die vierstündige Koinkubation von unbehandel-
ten mit Hitze-behandelten, nekrotischen Zellen in Suspension induziert. Es wur-
de einerseits die homotypische Phagozytose untersucht, indem verschiedene
Tumorzelllinien (Pankreaskarzinom-, Glioblastom-, primäre Kopf-Hals-
Tumorzellen, T-Zell-Leukämiezellen), primäre, gesunde Hautfibroblasten, emb-
ryonale Nierenepithelzellen und lymphoblastoide Zellen mit Hitze-behandelten
Zellen derselben Zelllinie inkubiert wurden. Zum anderen wurden heterotypi-
sche Aufnahmen von Hitze-behandelten Zellen durch andere Zelllinien getestet
(Tabelle 10). Tabelle 10: Kombination von Zelllinien bei der heterotypischen Phagozytose
Vital BxPC-3 SBLF5 T98G Hitze-behandelt SBL1
SBLF5 T98G
SBL1 BxPC-3 T98G
SBL1 SBLF5 BxPC-3
Nach der Vitalfärbung wurden die adhärenten Zellen abgelöst, mit dem automa-
tischen Zellzähler CASY TT gezählt und 5 min bei 300 g zentrifugiert. Die Zell-
konzentration von 106 Zellen pro ml wurde eingestellt, indem das Zellsediment
in einer entsprechenden Menge Medium resuspendiert wurde. Ein Teil der roten
Zellen wurde im Wasserbad bei 56 °C 45 min inkubiert und dadurch nekrotisiert.
Anschließend wurden 300 µl dieser Zellen, ohne zusätzlichen Waschschritt in
1 ml Rundbodenröhrchen mit grünen unbehandelten Zellen im Verhältnis 1 zu 1
bei 37 °C im CO2-Schrank koinkubiert. Als Kontrolle wurden zusätzlich unbe-
handelte rote Zellen mit unbehandelten grünen Zellen koinkubiert.
Zum Beginn und nach 60 min der Koinkubation erfolgte eine Messung der Ad-
häsion am Durchflusszytometer (siehe 2.4.2). Anschließend wurden die Zellsu-

41
spensionen in Suspensionskulturplatten mit 24 Näpfchen überführt, mit 2 µl
Hoechst 33342 versetzt und für die restliche Inkubationsdauer von weiteren 3 h
im CO2-Schrank inkubiert. Erste Phagozytose-Ereignisse entstanden unter den
beschriebenen Bedingungen 2,5 h nach Beginn der Koinkubation. Anschlie-
ßend erfolgte die Zentrifugation der Zellen auf Objektträger und deren Fixie-
rung.
Zur Überprüfung der vollständigen Aufnahme von hyperthermierten durch un-
behandelte Zellen, wurden CTOG-gefärbte vitale Zellen mit ungefärbten Hitze-
behandelten Zellen 4 h in Suspensionskulturplatten koinkubiert. Anschließend
wurden die Zellen bei 300 g für 5 min zentrifugiert und das Sediment in 200 µl
Ringerlösung resuspendiert, sodass die Zellkonzentration 5*106 Zellen/ml ent-
sprach. Die Zellen wurden 30 min auf Eis im Dunkeln mit 4,5 µl 7-AAD gefärbt
und nach erneuter Zentrifugation bei 300 g für 5 min und Suspension in Ringer-
lösung auf Objektträger zentrifugiert und mikroskopiert.
Zur Untersuchung der Phagozytose unter Zelltyp-gerechten Bedingungen der
adhärenten Zellen wurden 1,5*105 Pankreaskarzinom-Zellen auf Objektträger
eingesät, nach 24 h mit 1 ml der Zellsuspension von hyperthermierten Zellen
(c=106/ml) für 4 h bei 37 °C im CO2-Brutschrank überschichtet und anschlie-
ßend fixiert.
Inhibition der Phagozytose von Hitze-behandelten Zellen 2.3.5.
Zur Untersuchung der notwendigen molekularen Mechanismen der nicht-
professionellen Phagozytose wurden Pankreaskarzinom-Zellen unmittelbar vor
der Koinkubation mit Hitze-behandelten Zellen mit unterschiedlichen Noxen be-
handelt.
Der Phagozytose-Versuch zur Aufnahme von Hitze-behandelten Zellen (siehe
2.3.4) wurde zu diesem Zweck modifiziert. Ein Teil der vitalen, grünen Zellen
wurde mit 50 Gy bestrahlt. Des Weiteren wurden grüne Zellen mit den proteoly-
tischen Enzymen Trypsin und Accutase, welche Moleküle von der Zelloberflä-
che abspalten, behandelt. Für diese Behandlung wurde der Zellrasen einer T-
25 Flasche mit je 1 ml 0,05 % Trypsin oder Accutase versetzt und 45 min bei
37 °C im CO2-Inkubator inkubiert. Außerdem wurden grün gefärbte Zellen mit
den Zytoskelett-Inhibitoren Latrunculin B (0,5 µg/ml in Serum-freiem Medium),
Colcemid (90 ng/ml) oder Paclitaxel (0,5 µg/ml) versetzt und 1 h im CO2-

42
Schrank vorbehandelt. Zur Hemmung der GTPase Aktivität wurden grün gefärb-
te Zellen mit Dynasore (26 µg/ml) 1 h bei 37 °C inkubiert. Während der an-
schließenden Koinkubation mit Hitze-behandelten Zellen wurde die Konzentra-
tion der Inhibitoren aufrechterhalten.
Als Kontrollen wurden grüne Zellen mit den Lösungsmitteln DMSO und PBS in
den entsprechenden Konzentrationen versetzt (1 µl bzw. 5 µl DMSO oder 90 µl
PBS auf 5 ml Medium bzw. 1 ml PBS auf eine T-25 Flasche als Kontrolle für
Trypsin/Accutase) und mit roten Hitze-behandelten Zellen inkubiert. Für die Be-
handlung mit Latrunculin B wurde die Kontrolle mit dem Lösungsmittel in Se-
rum-freiem Medium durchgeführt. Des Weiteren wurden unbehandelte grüne
Zellen mit Hitze-behandelten bzw. unbehandelten roten Zellen inkubiert. Nach
Ende der Koinkubationsdauer von 4 h wurden die Zellsuspensionen auf Objekt-
träger aufgebracht, fixiert und mikroskopiert, um die Phagozytose-Raten zu be-
stimmen.
Funktionale Blockade von E-und P-Cadherin 2.3.6.
Als weiterer Ansatz sollte der E- und P-Cadherin-vermittelte Zellkontakt mit
Funktions-blockierenden Antikörpern unterbunden und der Effekt auf Adhäsion-
und Phagozytose-Raten untersucht werden. Für diesen Versuch wurde die
Pankreaskarzinom-Zelllinie BxPC-3 verwendet, die beide Zellmembran-Proteine
exprimiert. Der Phagozytose-Versuch zur Aufnahme von Hitze-behandelten
Zellen (siehe 2.3.4) wurde insofern abgewandelt, als dass zum Beginn der
Koinkubation die blockierenden Antikörper zugegeben wurden. Die verwende-
ten Konzentrationen waren 150 µg/ml für E-Cadherin und 100 µg/ml für P-
Cadherin. Eine Isotypkontrolle mit den entsprechenden Konzentrationen für
beide Antikörper wurde ebenfalls mitgeführt. Außerdem wurden Proben ohne
Hyperthermie-Behandlung angesetzt, um den Effekt der blockierenden Antikör-
per auf die Adhäsion zwischen vitalen Zellen zu untersuchen. Nach Ende der
Koinkubation von 4 h wurden die Zellsuspensionen auf Objektträger aufge-
bracht, fixiert und mikroskopiert, um die Phagozytose-Raten zu quantifizieren.
Behandlung mit Annexin V zur Blockade von Phosphatidylserin 2.3.7.
Für die Blockade von Phosphatidylserin wurde Annexin V (AnxA5) in Konzent-
rationen von 10 – 40 µg/ml (entspricht 280 nM – 1,1 µM) verwendet. Rot-

43
gefärbte Hautfibroblasten, Pankreaskarzinom- und Glioblastom-Zellen wurden
nach der Hitzebehandlung (56 °C, 45 min) zentrifugiert und in Medium mit zu-
gesetztem AnxA5 1 h bei 37 °C inkubiert. Bei der Koinkubation mit unbehandel-
ten grünen Zellen wurden die Konzentrationen von AnxA5 aufrechterhalten. Als
Kontrollen wurden wiederum grüne Zellen mit Hitze-behandelten roten Zellen
ohne AnxA5 und unbehandelte grüne mit unbehandelten roten Zellen inkubiert.
Eine Kontrolle mit dem Lösungsmittel war nicht notwendig, da das AnxA5 in
Medium gelöst worden war.
Nach Ende der Koinkubationszeit von 4 h wurden die Zellsuspensionen auf Ob-
jektträger aufgebracht, fixiert und mikroskopiert, um die Phagozytose-Raten zu
ermitteln.
Phagozytose von bestrahlten Zellen 2.3.8.
Als weitere Methode zur Induktion von Nekrose wurden mehrere der Zelllinien,
die nach Hitze-Behandlung von nicht-professionellen Phagozyten aufgenom-
men wurden, mit unterschiedlichen Dosen Röntgenstrahlung bestrahlt. T-Zell-
Leukämiezellen erhielten eine Dosis von 8 Gy mit anschließender Inkubations-
zeit von drei Tagen im CO2-Inkubator. Pankreaskarzinom-Zellen wurden mit
20 Gy bestrahlt und für weitere sechs Tage inkubiert. Die primäre Kopf-Hals-
Tumorzelllinie wurde mit der Einzeldosis von 2 Gy an fünf aufeinanderfolgenden
Tagen fraktioniert bestrahlt (Gesamtdosis: 10 Gy) und drei Wochen im CO2-
Inkubator inkubiert. Die Induktion von Nekrose und Apoptose wurde durch 7-
AAD/AnxA5-APC (Allophycocyanin) Färbung am Durchflusszytometer gemes-
sen (siehe 2.4.1). Außerdem wurden die homotypischen und heterotypischen
Phagozytose-Raten nach Vitalfärbung und 4 h Koinkubation von unbehandelten
mit bestrahlten Zellen in Suspension bestimmt. Bei der Vitalfärbung der be-
strahlten Zellen wurden das Medium mit den schwimmenden Zellen und der
nekrotischen Debris nicht verworfen, sondern zentrifugiert und wie Suspensi-
onszellen gefärbt (siehe 2.3.3).
Für die mikroskopische Auswertung wurden die Zellsuspensionen auf Objekt-
träger zentrifugiert und fixiert. Sowohl für die durchflusszytometrische Messung
als auch für den Phagozytose-Versuch wurden als Positivkontrolle für die Induk-
tion von Nekrose und Phagozytose hyperthermierte Zellen (56 °C, 45 min) mit-
geführt.

44
Außerdem wurde mittels Immunfluoreszenz-Färbung getestet, ob in bestrahlten
und hyperthermierten Zellen die aktivierte Caspase-3 (cleaved caspase-3)
nachgewiesen werden kann und somit die Induktion von Apoptose. Zu diesem
Zweck wurde die Vitalfärbung auf die CTOG-Färbung der phagozytierenden
Zellen beschränkt, um einer Signalüberlagerung der CTFR-Färbung mit der
Caspase-3 Färbung vorzubeugen. Die Zellsuspensionen wurden 4 h und 15 h
nach Beginn der Koinkubation auf Objektträger zentrifugiert, fixiert und mit ei-
nem Antikörper gegen die aktivierte Caspase-3 gefärbt (siehe 2.6).
Beobachtung der Phagozytose-Ereignisse über die Zeit 2.3.9.
Für die Beobachtung von zellulären Prozessen nach Aufnahme von hyperther-
mierten Zellen wurden vier Ansätze gewählt (Tabelle 11). Tabelle 11: Versuchsansätze zur Untersuchung des Zeitverlaufs nach Phagozy-tose
Unbehandelt BxPC-3 (CTOG, grün) Hitze-behandelt
Jurkat (CD2, rot)
BxPC-3 (H2B-RFP, rot)
BxPC-3 (EdU, rot)
BxPC-3 (HSP70, rot LAMP1, gelb)
Der Versuchsablauf entspricht der Phagozytose von Hitze-behandelten Zellen
(siehe 2.3.4) mit der Modifikation einer verlängerten Inkubationsdauer von bis
zu drei Tagen und einer Kernfärbung mit DAPI statt Hoechst. Außerdem wurde
auf die durchflusszytometrische Messung verzichtet. Die heterotypische Pha-
gozytose wurde anhand von unbehandelten CTOG-gefärbten Pankreaskarzi-
nom-Zellen (BxPC-3), die mit Hitze-behandelten ungefärbten T-Zell-
Leukämiezellen (Jurkat) koinkubiert wurden, untersucht. T-Zell-Leukämiezellen
wurden durch Immunfluoreszenz-Färbung des T-Zell Antigens CD2 sichtbar
gemacht (siehe 2.6). Zur Beobachtung der homotypischen Phagozytose wurden
BxPC-3 Zellen verwendet. Unbehandelte CTOG-gefärbte BxPC-3 Zellen wur-
den mit hyperthermierten, mit Histon H2B-RFP transfizierten Zellen oder mit
Zellen, die das Nukleosid EdU inkorporiert hatten (siehe 2.2.3 – 4), koinkubiert.
Zur Untersuchung der Verteilung von HSP70 und LAMP1 mittels Immunfluores-
zenz wurde zusätzlich ein Ansatz mit vitalen, CTOG-gefärbten und ungefärbten,
hyperthermierten Pankreaskarzinom-Zellen mitgeführt. Nach 4 – 72 h nach Be-
ginn der Koinkubation wurden die Zellsuspensionen auf Objektträger zentrifu-

45
giert und fixiert. Zur Färbung der Nuklei wurden 50 µl der DAPI-
Gebrauchslösung auf die Objektträger pipettiert, diese mit Parafilm bedeckt und
im Dunkeln 1 min inkubiert. Anschließend wurden die Objektträger in bidestil-
liertes Wasser getaucht, getrocknet und mit Vectashield-Medium eingedeckt.
Erstellung der Objektträger nach der Koinkubation 2.3.10.
Nach Beendigung der Koinkubation der Phagozytose-Versuche in Suspension
wurden die Zellsuspensionen in Eppendorf Reaktionsgefäße überführt, in der
Tischzentrifuge bei 300 g für 5 min zentrifugiert und die Sedimente im Aus-
gangsvolumen Ringerlösung resuspendiert, sodass die Zellkonzentration von
106/ml erhalten blieb. Je 50 µl der Suspensionen wurden pro Kammer eines
vorbereiteten Zytozentrifugationsschlittens mit Dreifach-Kammer pipettiert und
auf Objektträger bei 1000 rpm 6 min in einer Zytofuge zentrifugiert. Der Über-
stand wurde entfernt und die Objektträger wurden während des Trocknens mit
Alufolie abgedeckt. Auf den trockenen Objektträgern wurden die Zellbereiche
mit Fettstift umrandet bevor die Objektträger in Quadripermschalen vorsichtig
mit Fixierlösung überschichtet wurden. Die Objektträger mit angewachsenen
Pankreaskarzinom-Zellen wurden vorsichtig mit PBS gespült, um die nicht pha-
gozytierten hyperthermierten Zellen zu entfernen und anschließend fixiert.
Die Fixierung und Permeabilisierung fand jeweils 10 min bei Raumtemperatur
(RT) im Abzug statt. Anschließend wurden die Objektträger dreimal 3 min auf
dem Rüttler bei 50 rpm mit Tris-gepufferter Salzlösung (TBS) gewaschen und
im Dunkeln getrocknet. Die trockenen Objektträger wurden mit Vectashield ein-
gedeckt und im Kühlschrank bei 6 – 8 °C aufbewahrt.
Quantifizierung der Phagozytose-Raten 2.3.11.
Zur vergleichenden Analyse der Effizienz der phagozytischen Aktivität von ver-
schiedenen Zelllinien und der Einflüsse von Inhibitoren wurden die erstellten
Objektträger mikroskopisch ausgewertet. Dafür wurden Rasteraufnahmen mit
einem Fluoreszenzmikroskop aufgenommen, um anschließend den Prozentsatz
an Zell-in-Zell Strukturen mit dem Bild-Analyse-Programm Biomas zu ermitteln.
Diese Prozentangaben geben die Anzahl von Phagozytose-Ereignissen dividiert
durch die Anzahl aller grünen, vitalen Zellen wieder. Es wurden 1000 bis 3000
grüne Zellen ausgewertet, abhängig davon, ob insgesamt eher viele oder nur

46
sehr wenige Zell-in-Zell Strukturen vorhanden waren. Dabei wurden nur voll-
ständig abgeschlossene Phagozytose-Ereignisse, die einen umschließenden
Zellmembransaum um die aufgenommene Zelle zeigten, gewertet.
Durchflusszytometrie 2.4.Die durchflusszytometrischen Messungen wurden mit einem Gallios von Beck-
man Coulter durchgeführt. Die gewonnenen Daten wurden als LMD-Dateien
gespeichert und mit der Software Kaluza 1.2 ausgewertet. Das Gerät misst die
Eigenschaften von einzelnen Zellen anhand der von ihnen erzeugten Licht-
streuung und Fluoreszenzemission. Die Zellen werden durch eine Kapillare ge-
saugt und passieren im sogenannten Probenstrom die Durchflusszelle, welche
umgeben ist von Mantelflüssigkeit. Dadurch kommt es zum Prozess der hydro-
dynamischen Fokussierung. Zusätzlich vibriert die Öffnung der Durchflusszelle,
sodass einzelne Tropfen entstehen, die jeweils nur eine einzelne Zelle enthal-
ten. Die Zellen werden einzeln von einem fokussierten Laserstrahl getroffen und
streuen das Licht im flachen Winkel (Vorwärtsstreuung) und im rechten Winkel
(Seitwärtsstreuung). Das Vorwärtsstreulicht gibt Auskunft über die Größe der
Zelle, das Seitwärtsstreulicht über die Granularität der Zelle. Außerdem können
intrazelluläre und extrazelluläre Moleküle mit Fluorochromen markiert werden,
sodass die Zellen bei Passage des Laserstrahls zusätzlich Fluoreszenzsignale
emittieren. Diese Methode erlaubt eine Bestimmung der Expression von be-
stimmten Molekülen und ebenso eine Aussage über die Integrität der Zell-
membran und den Zellzyklus anhand des DNA-Gehalts.
Bestimmung der Zellmorphologie und Zellviabilität 2.4.1.
Anhand der Vorwärts- und Seitwärtsstreuung können lebende und tote Zellen
einer Zelllinie unterschieden werden. Im Allgemeinen zeichnen sich sterbende
Zellen durch eine starke Zunahme der Granularität und eine Abnahme der Zell-
größe aus. Dadurch erscheinen sie in einem Punktewolkendiagramm, bei dem
die Granularität (Seitwärtsstreuung) gegen die Zellgröße (Vorwärtsstreuung)
aufgetragen wird, als distinkte Zellpopulation, die oberhalb der vitalen Zellpopu-
lation liegt. Nach Hitze-Behandlung (45 min bei 56 °C) nimmt die Zellgranulari-
tät erheblich zu (Abbildung 1). Zur Überprüfung des eingesetzten Mengenver-
hältnisses von 1 zu 1 zwischen vitalen und aufzunehmenden Zellen wurden die
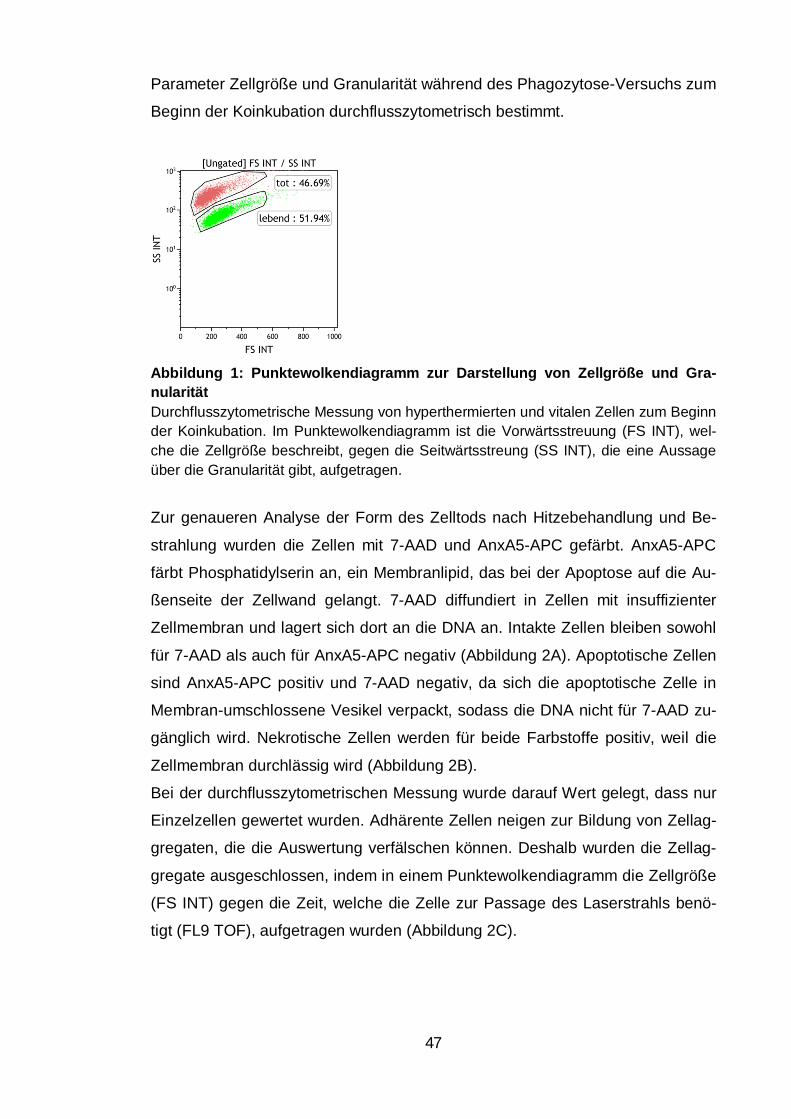
47
Parameter Zellgröße und Granularität während des Phagozytose-Versuchs zum
Beginn der Koinkubation durchflusszytometrisch bestimmt.
Abbildung 1: Punktewolkendiagramm zur Darstellung von Zellgröße und Gra-nularität Durchflusszytometrische Messung von hyperthermierten und vitalen Zellen zum Beginn der Koinkubation. Im Punktewolkendiagramm ist die Vorwärtsstreuung (FS INT), wel-che die Zellgröße beschreibt, gegen die Seitwärtsstreung (SS INT), die eine Aussage über die Granularität gibt, aufgetragen.
Zur genaueren Analyse der Form des Zelltods nach Hitzebehandlung und Be-
strahlung wurden die Zellen mit 7-AAD und AnxA5-APC gefärbt. AnxA5-APC
färbt Phosphatidylserin an, ein Membranlipid, das bei der Apoptose auf die Au-
ßenseite der Zellwand gelangt. 7-AAD diffundiert in Zellen mit insuffizienter
Zellmembran und lagert sich dort an die DNA an. Intakte Zellen bleiben sowohl
für 7-AAD als auch für AnxA5-APC negativ (Abbildung 2A). Apoptotische Zellen
sind AnxA5-APC positiv und 7-AAD negativ, da sich die apoptotische Zelle in
Membran-umschlossene Vesikel verpackt, sodass die DNA nicht für 7-AAD zu-
gänglich wird. Nekrotische Zellen werden für beide Farbstoffe positiv, weil die
Zellmembran durchlässig wird (Abbildung 2B).
Bei der durchflusszytometrischen Messung wurde darauf Wert gelegt, dass nur
Einzelzellen gewertet wurden. Adhärente Zellen neigen zur Bildung von Zellag-
gregaten, die die Auswertung verfälschen können. Deshalb wurden die Zellag-
gregate ausgeschlossen, indem in einem Punktewolkendiagramm die Zellgröße
(FS INT) gegen die Zeit, welche die Zelle zur Passage des Laserstrahls benö-
tigt (FL9 TOF), aufgetragen wurden (Abbildung 2C).

48
Abbildung 2: Durchflusszytometrische Messung zum Nachweis von Apoptose, Nekrose und Zellaggregaten Punktewolkendiagramm zum Nachweis von Apoptose und Nekrose nach AnxA5 und 7-AAD-Färbung von unbehandelten (A) und hyperthermierten (56 °C für 45 min) Pankre-askarzinom-Zellen (B). Zum Ausschluss von Zellaggregaten wurden Punktewolkendia-gramme von Zellgröße (FS INT) gegenüber Passage-Zeit (FL9 TOF) erstellt (C).
Für diese Färbung wurden die Zellen zunächst durch Zugabe von Trypsin abge-
löst. Dabei galt für die bestrahlten Zellen die Änderung, dass zu Beginn das
Medium, sowie anschließend die Waschlösung und das Trypsin in Zentrifugen-
röhrchen überführt werden mussten, damit keine schwimmenden, toten Zellen
verloren gingen. Die Zellen wurden bei 300 g für 5 min zentrifugiert, der Über-
stand verworfen und das Zellsediment in 1 ml Medium resuspendiert. Zur Mes-
sung von Duplikaten je Bedingung wurden je 500 µl in zwei Rundboden-
Röhrchen überführt. Pro Röhrchen wurden 2 µl Höchst pipettiert und diese 1,5 h
im Brutschrank inkubiert. Nach anschließender Zentrifugation (10 min bei
300 g) wurden die Sedimente in je 200 µl Ringerlösung resuspendiert und auf
Eis gestellt. Daraufhin wurden je Röhrchen jeweils 5 µl 7-AAD und AnxA5-APC
zugegeben und nach vorsichtigem Mischen für 30 min im Dunkeln inkubiert.
Anschließend wurden weitere 200 µl Ringerlösung in jedes Röhrchen pipettiert
und die Proben sofort mit dem Durchflusszytometer gemessen.
Messung der Zell-Adhäsion 2.4.2.
Während des Phagozytose-Versuchs wurde nach 60 min Koinkubation die Ad-
häsion zwischen unbehandelten, CTOG- gefärbten und behandelten CTFR-
gefärbten Zellen durchflusszytometrisch quantifiziert. Grüne, CTOG-gefärbte
phagozytische Zellen (FL1 positiv) binden, im Vergleich zur Adhäsions-Rate mit
roten, CTFR-gefärbten unbehandelten Zellen, verstärkt an nekrotische Zellen.
Mit einem Punktewolkendiagramm wurden die Zellpopulationen dargestellt, in-

49
dem die Signale des grünen Fluoreszenzkanals (FL1) gegen die Signale des
roten Fluoreszenzkanals (Fl6) aufgetragen wurden (Abbildung 3). Die Adhäsion
wurde als der Anteil doppelt-positiver Zellen dividiert durch den Anteil aller grün-
gefärbten Zellen kalkuliert.
Abbildung 3: Messung der Adhäsion
Durchflusszytometrische Messung von CTFR-gefärbten, hyperthermierten und CTOG-gefärbten, vitalen Zellen nach 60 min Koinkubation in Suspension. Im Punktewolkendi-agramm sind die Signale des grünen Fluoreszenzkanals (FL1 INT) gegen die Signale des roten Fluoreszenzkanals (FL6 INT) aufgetragen. CTFR: Celltrace Far Red; CTOG: Celltrace Oregon Green.
Mikroskopisch wurde bestätigt, dass zum Messzeitpunkt nach 60 min noch kei-
ne Phagozytose-Ereignisse stattgefunden hatten. Diese Messung stellt also
lediglich die Adhäsion zwischen grünen, vitalen und roten, nekrotischen Zellen
dar und nicht die Phagozytose.
Messung der Konzentration der zellulären Proteine aus Zell-2.5.
lysaten und von freigesetztem HSP70 nach Hyperthermie Zur genaueren Charakterisierung der hyperthermierten Zellen wurde der Effekt
von Hitze-Behandlung (45 min bei 56 °C) in Bezug auf den Gesamt-Protein-
Gehalt von Zellen und die Freisetzung von HSP70 untersucht.
Für die Protein-Isolierung wurden die Zellen trypsiniert und mit kaltem PBS ge-
waschen. Zusätzlich wurde eine Probe mit hyperthermierten Zellen vorbereitet.
Eine definierte Anzahl von Zellen wurde mit Verwendung des RIPA-Puffers (ra-
dioimmunoprecipitation assay) 30 min auf Eis lysiert, wobei 50 µl Puffer pro 106
Zellen pipettiert wurden. Anschließend wurden die Zelllysate 8 min bei 13000 g

50
zentrifugiert und die Überstände gesammelt. Die Protein-Konzentration wurde
mit dem bicinchoninic acid assay kit (BCA kit) bestimmt.
Um die Freisetzung von HSP70 nach Hitzeschock nachzuweisen, wurde ein
Enzyme-linked Immunosorbent assay (ELISA) verwendet. Dazu wurden Über-
stände von einer definierten Anzahl unbehandelter und Hitze-behandelter Zel-
len verwendet, die 4 h und 24 h in Suspensionskulturplatten inkubiert wurden.
Die Zellen wurden durch Zentrifugation bei 300 g für 5 min entfernt und die
Überstände 1 zu 10 verdünnt. Der HSP70 DuoSet IC ELISA (R&D Systems)
wurde nach Herstellerangaben durchgeführt, die Signalstärke nach Farbum-
schlag mit einem Photometer bei 450 nm und 570 nm gemessen und mit der
HTSoft Software ausgewertet.
Immunfluoreszenz-Färbung 2.6.Die Immunfluoreszenz-Färbung ist eine geeignete Methode, um spezifisch int-
razelluläre, Transmembran- oder Oberflächenproteine zu markieren und durch
Fluoreszenz sichtbar zu machen. Bei der in dieser Arbeit verwendeten indirek-
ten Immunzytochemie wird zuerst ein Primär-Antikörper, der gegen das zu fär-
bende Protein gerichtet ist, auf die Probe gebracht und anschließend ein Se-
kundär-Antikörper, der mit einem Fluorochrom gekoppelt ist und an das Fc-
Fragment des Primär-Antikörpers bindet.
Zur Überprüfung einer möglichen Induktion von aktivierter Caspase-3-
abhängiger Apoptose wurde eine Immunfluoreszenz-Färbung nach 4 h und
15 h Koinkubation von vitalen mit hyperthermierten (56 °C, 45 min) Pankreas-
karzinom-Zellen durchgeführt. Des Weiteren wurden mittels Immunfluoreszenz-
Färbung T-Zell-Leukämiezellen anhand des CD2 T-Zell-Antigens im Phagozy-
tose-Versuch sichtbar gemacht. Außerdem wurde die Lokalisation von LC3,
LAMP1, F-Aktin, phosphoryliertem Ezrin und E-Cadherin in Zell-in-Zell Struktu-
ren, ausgebildet von Pankreaskarzinom-Zellen nach Hitzebehandlung, zu den
Zeitpunkten 2,5 h, 3,5 h und 5 h quantifiziert. Diese Lokalisation wurde auch bei
angewachsenen Pankreaskarzinom-Zellen, auf die hyperthermierte Zellen zent-
rifugiert wurden, untersucht. Weiterhin wurde die Akkumulation von HSP70 in
den hyperthermierten Pankreaskarzinom-Zellen nach 4 h, 8 h und 22 h Koinku-
bation und die Expression von P-Cadherin bei unbehandelten BxPC-3-Zellen
nachgewiesen.

51
Färbe-Verfahren 2.6.1.
Die in den Phagozytose-Versuchen hergestellten Objektträger wurden nach
dem dritten Waschschritt nach der Fixierung in Quadripermschalen mit jeweils 7
ml Blockier-Lösung überschichtet und 1 h bei RT in einer Feuchtkammer inku-
biert.
Es folgten drei Waschschritte mit 1x TBS und die Überschichtung mit dem in
1 %-BSA in 1x TBS verdünnten Primär-Antikörper. Zur vollständigen Benetzung
der Zellen wurde Parafilm aufgebracht. Die Objektträger wurden über Nacht in
einer Feuchtkammer im Kühlschrank inkubiert.
Nach Wiederholung der Waschschritte wurde der verdünnte Sekundär-
Antikörper mit Parafilm aufgebracht und die Objektträger wurden 2 h bei RT in
der Feuchtkammer inkubiert. Nach drei weiteren Waschschritten wurden die
Objektträger in der Feuchtkammer mit 50 µl DAPI-Lösung und Parafilm benetzt,
um die Nuklei blau zu färben.
Nach 1 min wurden die Objektträger kurz in bidestilliertes Wasser getaucht und
zum Trocknen im Dunkeln aufgestellt. Nach dem Eindecken mit Vectashield-
Medium und Auflegen eines Deckglases wurden die Objektträger im Kühl-
schrank bei 6 – 8 °C gelagert. Alle verwendeten Antikörper und Lösungen sind
in den Tabellen 4, 5 und 8 aufgeführt.
Quantifizierung der Lokalisation von Phagozytose-assoziierten Pro-2.6.2.
teinen Die Häufigkeit der gerichteten Akkumulation von LC3, LAMP1, F-Aktin, phos-
phoryliertem Ezrin und E-Cadherin an Zell-Zell Kontakten sowie in Zell-in-Zell
Strukturen, wurde im engen Zeitrahmen (2,5 h, 3,5 h und 5 h Koinkubation)
quantifiziert. Dabei wurde untersucht, ob eine sichtbare Verdichtung der unter-
suchten Proteine während des Phagozytose-Prozesses an der Kontaktstelle zur
hyperthermierten Zelle vorzufinden war und, ob die Proteine ringförmig um die
aufgenommen Zelle innerhalb der Zell-in-Zell Struktur vorlagen. Um dies zu er-
reichen, wurden je Bedingung und Färbung sechs Rasterbilder mit dem Fluo-
reszenzmikroskop aufgenommen. Es wurde der Anteil aller gerichteten Lokali-
sationen von allen in jeweils sechs Rasteraufnahmen zu erkennenden Adhä-
sions- und Phagozytose-Ereignissen mit dem Bild-Analyse Programm Biomas
ermittelt.

52
Immunhistochemie 2.7.Neben den Versuchen mit Zelllinien wurden außerdem histologische Schnitte
von Tumorgewebe-Proben auf die Häufigkeit von Zell-in-Zell Strukturen unter-
sucht. Zur Identifizierung von aufgenommenen, apoptotischen Zellen wurde
eine Färbung für die aktivierte Caspase 3 durchgeführt.
Auswahl von Gewebeproben 2.7.1.
Tumorgewebe-Proben von 416 Patienten aus sieben verschiedenen Kohorten
(Tabellen 13 – 15) wurden auf das Vorhandensein von Zell-in-Zell Strukturen
hin untersucht. Die Phagozytose-Ereignisse in fünf Kohorten von Kopf-Hals-
Plattenepithelkarzinomen (head and neck squamous cell carcinoma, HNSCC)
und in je einer Kohorte von Rektum-und Analkarzinomen wurden quantifiziert.
Alle Proben wurden zu Tissue-Microarrays (TMAs) mit Gewebestanzen von 1,6
– 2 mm Durchmesser verarbeitet. Die zugehörigen klinischen Daten wurden
von der Datenbank des Tumorzentrums der Universität Erlangen-Nürnberg zur
Verfügung gestellt.
Alle Patienten hatten eine Einverständniserklärung für die Nutzung der Gewe-
beproben und der klinischen Daten unterschrieben. Die Studie wurde von dem
Ethikkomitee des Universitätsklinikums Erlangen bewilligt.
Färbung von histologischen Schnitten 2.7.2.
Zur Färbung von aktivierter Caspase 3 wurden die Schnitte zunächst über
Nacht mit dem Primär-Antikörper und anschließend zur Amplifikation des Sig-
nals mit einem Biotin-gekoppelten Sekundär-Antikörper inkubiert. Mit einem
Streptavidin-biotinylierten alkalische Phosphatase-Komplex und einem Chro-
mogen (Fast Red) wurde das Caspase 3 Signal sichtbar gemacht.
Die Färbung von E-Cadherin erfolgte mit einem automatischen Ventana
BenchMark Ultra stainer unter Verwendung von CC1-Puffer (Cell Conditioning
Solution 1). Die Zellkerne wurden mit Hämatoxylin oder DAPI gefärbt.

53
Quantifizierung von Phagozytose-Raten in histologischen Präpara-2.7.3.
ten
Die Phagozytose-Raten wurden mithilfe der Bildbearbeitungs-Software Biomas
bestimmt. Pro Patient wurden mindestens zwei TMA Gewebestanzen mit einer
Fläche von 0,75 mm2 ausgewertet. Mit der Software wurden die aufeinanderfol-
genden Schnitte, die mit E-Cadherin bzw. aktivierter Caspase 3 gefärbt worden
waren, zusammengefügt, sodass eine Auswertung der Häufigkeit von Zell-in-
Zell Strukturen und von Apoptose in demselben Gebiet möglich war.
Mikroskopie 2.8.Zur Beobachtung und molekularen Charakterisierung von Phagozytose-
Ereignissen, sowie deren Quantifizierung, wurden die in den Versuchen herge-
stellten Objektträger, sowie die histologischen Präparate, mikroskopisch unter-
sucht. Dabei fanden neben Durchlicht- und Fluoreszenzmikroskopie ebenso
konfokale und Lebendzell-Mikroskopie Anwendung.
Fluoreszenz- und Phasenkontrastmikroskopie 2.8.1.
Für die Quantifizierung der Phagozytose-Raten und der qualitativen sowie
quantitativen Auswertung nach Immunfluoreszenz-Färbung wurde entweder
das Leica DM6000B (Leica Microsystems Ltd.) Fluoreszenzmikroskop, ausge-
stattet mit einer Kamera (DFC350FX, Leica), oder das Zeiss Axioplan 2 Imaging
(Zeiss) Fluoreszenmikroskop, ausgestattet mit einer Kamera (Cool Cube 1,
Zeiss), verwendet. Die Objektive mit 40-, 63- und 100-facher Vergrößerung
wurden genutzt (HCX PL APO, 40x, NA: 1,25, Leica; Plan-Neofluar, 40x, NA:
0,75, Zeiss; Plan-Apochromat, 63x, NA: 1,4, Zeiss).
Mithilfe der Mikroskop-Software Metafer 4 (MetaSystems) und Leica Application
Suite V3 (Leica) wurden Einzelbilder für die qualitative Untersuchung und Ras-
teraufnahmen (8*8 Bilder) für die quantitative Untersuchung aufgenommen.
Die gefärbten TMAs wurden mit einem Hochdurchsatz Scanner (Mirax MIDI
Scan, Zeiss), ausgestattet mit einem Plan-Apochromat Objektiv (20x; NA: 0,8,
Zeiss) und einer Kamera (Stingray F146C, AVT), mikroskopiert.
Die Bildbearbeitung und Auswertung wurde mit den Programmen Biomas und
Photoshop und durchgeführt.

54
Konfokale Mikroskopie 2.8.2.
Mithilfe von konfokaler Mikroskopie wurde der vollständige Membranumschluss
um die aufgenommene, hyperthermierte Zelle verifiziert. Dafür wurden Zyto-
zentrifugation-Präparate aus den Phagozytose-Versuchen (siehe 2.3.4) ver-
wendet. Die Aufnahmen wurden mit einem Zeiss LSM710 (Zeiss) Laser-
Scanning-Mikroskop, ausgestattet mit einem Plan-Apochromat Objektiv (63x;
NA: 1,4; Zeiss), gemacht. Die Bildaufnahme und –Verarbeitung wurde mit der
Mikroskop-Software ZEN (Zeiss) durchgeführt. Alle 3D-Projektionen wurden mit
dem Bildbearbeitungs-Programm ImageJ erstellt.
Lebendzell-Mikroskopie 2.8.3.
Zur Verfolgung des zeitlichen Ablaufs von Phagozytose-Ereignissen wurden
primäre Kopf-Hals-Tumorzellen (p26) mittels Lebendzell-Mikroskopie bis zu
24 h beobachtet. Dafür wurde ein Phagozytose-Versuch mit hyperthermierten
Zellen vorbereitet, wobei auf die Vital- sowie die Kern-Färbung verzichtet wur-
de. Anschließend wurden 3 ml der Zellsuspension in eine Suspensions-
Kulturplatte mit sechs Näpfchen überführt. Zur Beobachtung wurde ein Zeiss
Axioplan 200 Mikroskop, ausgestattet mit einem Plan-Neofluar Objektiv (20x;
NA: 0,5; Zeiss) und einer Digitalkamera (C4742-95, Hamamatsu), verwendet.
Die Zellen befanden sich während der Aufnahmen in einem Inkubator bei 37 °C
und 5 % CO2. Die aufgenommenen Bilder wurden mit den Programmen Voloci-
ty (PerkinElmer) und ImageJ (NIH) zu einem Film zusammengesetzt.
Messung der Genexpression 2.9.Zur Bestimmung der Expression von Phagozytose-assoziierten Genen wurde
aus Phagozytose –kompetenten und –inkompetenten Zellen RNA isoliert. Nach
anschließender Herstellung der copy DNA (cDNA) wurde mittels quantitativer
Echtzeit-PCR (qRT-PCR) die Expression der zu studierenden Gene (CD36,
CD68, ITGB5 und TIM1) untersucht.
RNA-Isolation und reverse Transkription 2.9.1.
Die Isolation von RNA für die qRT-PCR wurde unter Verwendung des Qia
shedder und RNeasy mini kit nach Herstellerangaben ausgeführt. Die Integrität

55
der extrahierten RNA wurde durch Messung der 260 zu 280 nm und 260 zu
230 nm Verhältnisse mit einem Spektrophotometer verifiziert. Mit dem Quanti-
Tect reverse transcription kit wurden jeweils 2 µg RNA in cDNA umgeschrieben
und für die Nicht-Reverse-Transkriptase Kontrolle zusätzlich 0,5 µg RNA mitge-
führt.
Quantitative PCR 2.9.2.
Für die Durchführung der PCR wurden das DyNAmo Flash SYBR Green qPCR
Kit und das Bio-Rad CFX Real-Time System verwendet. Die gemessenen
mRNA Mengen von CD36, CD68, ITGB5 und TIM1 wurden auf den Mittelwert
von drei Referenzgenen (L3MBTL2, GIGYF2 und RPUSD3) normiert, um eine
Aussage über die relative Quantität der untersuchten Gene treffen zu können.
Die Auswahl der am besten geeigneten Referenzgene wurde zusammen mit
der Stabilität von Referenz- und Zielgenen mit Hilfe des Programms qbase und
dem darin verwendeten Algorithmus geNorm bestimmt. Die Genexpressions-
Analysen wurden mit der CFX Manager Software berechnet.
Die verwendeten Oligonukleotide (Tabelle 2) wurden mithilfe von Primer3 input
erstellt. Zunächst wurden einige Test-Reaktionen mit einem cDNA-Pool beste-
hend aus allen verwendeten cDNAs durchgeführt. Zuerst wurden die optimale
Konzentration des jeweiligen Oligonukleotid-Paares (100 – 400 nM) und die
optimale Temperatur der Hybridisierungsphase mithilfe eines Temperaturgradi-
enten ermittelt. Entsprechend der vorangegangenen Tests, wurde die Amplifika-
tionseffizienz eines jeden Oligonukleotid-Paares mit einer Standardkurve, bei
welcher der cDNA-Pool in insgesamt 5 Schritten mit dem Faktor 1 zu 10 ver-
dünnt wurde, getestet. Außerdem wurde das Reaktionsprodukt auf ein Agaro-
segel aufgetragen, um die Länge des gebildeten Amplikons und das Vorhan-
densein von Oligonukleotid-Dimeren zu überprüfen.
Statistik 2.10.Alle Daten wurden als Mittelwert ± Standardabweichung dargestellt. IBM SPSS
Statistics 19 wurde für die Berechnung von signifikanten Unterschieden zwi-
schen unabhängigen Gruppen mithilfe des ungepaarten t-Tests verwendet. Für
die histologischen Daten wurden der Kolmogorov–Smirnov und der Lilliefors
Test angewandt, um die Normalverteilung zu überprüfen. Das Überleben ohne

56
lokales Therapieversagen, das Metastase-freie und das Gesamt-Überleben
wurden nach Kaplan Meier kalkuliert. Mit Hilfe des log-rank Tests wurden die
Überlebenszeiten zwischen Untergruppen von Patienten verglichen. P-Werte <
0,05 wurden als signifikant erachtet.

57
3. Ergebnisse
Nachweis der Phagozytose 3.1.Die Phagozytose-Aktivität von nicht-professionellen Phagozyten wurde anhand
von Tumorzellen und normalen Hautfibroblasten mithilfe eines Phagozytose-
Versuchs in einem Suspensionskultur-System studiert. Nach Induktion der pri-
mären Nekrose durch Hitze-Behandlung oder Bestrahlung mit Röntgenstrah-
lung folgte eine Koinkubation mit unbehandelten, lebenden Zellen, welche die
sterbenden Zellen innerhalb weniger Stunden aufnahmen. Zur Unterscheidung
der nekrotischen und vitalen Zellen wurden die Zellen zu Beginn des Versuchs
mit Vital-Farbstoffen (Celltrace Oregon Green und Celltrace Far Red) grün und
rot markiert. Die verwendeten Farbstoffe reichern sich im gesamten Zytoplasma
an, indem sie an zelleigene Proteine kovalent binden und färben dadurch die
gesamte Zelle mit Ausnahme des Zellkerns homogen an. Die rot-gefärbten Zel-
len wurden abgelöst und im Wasserbad bei 56 °C für 45 min hyperthermiert.
Die Phagozytose-Versuche mit Bestrahlung wichen insofern von diesem Ver-
suchsablauf ab, als dass die Bestrahlung vor der Vital-Färbung durchgeführt
wurde. Die Koinkubation der behandelten, rot-gefärbten Zellen mit vitalen, grün-
gefärbten Zellen in Suspensions-Kulturplatten für 4 h resultierte in der Bildung
von phagozytischen Strukturen. Diese waren durch das Vorhandensein eines
an die Zell-Peripherie gedrängten sichelförmigen Zellkerns der phagozytischen
Zelle, bedingt durch die Aufnahme einer nekrotischen Zelle, gekennzeichnet.
Die vollständige Aufnahme von hyperthermierten Zellen bei einem Phagozyto-
se-Versuch mit den Pankreaskarzinom-Zellen BxPC-3 wurde mit konfokaler
Laser-Scanning Mikroskopie nachgewiesen (Abbildung 4).
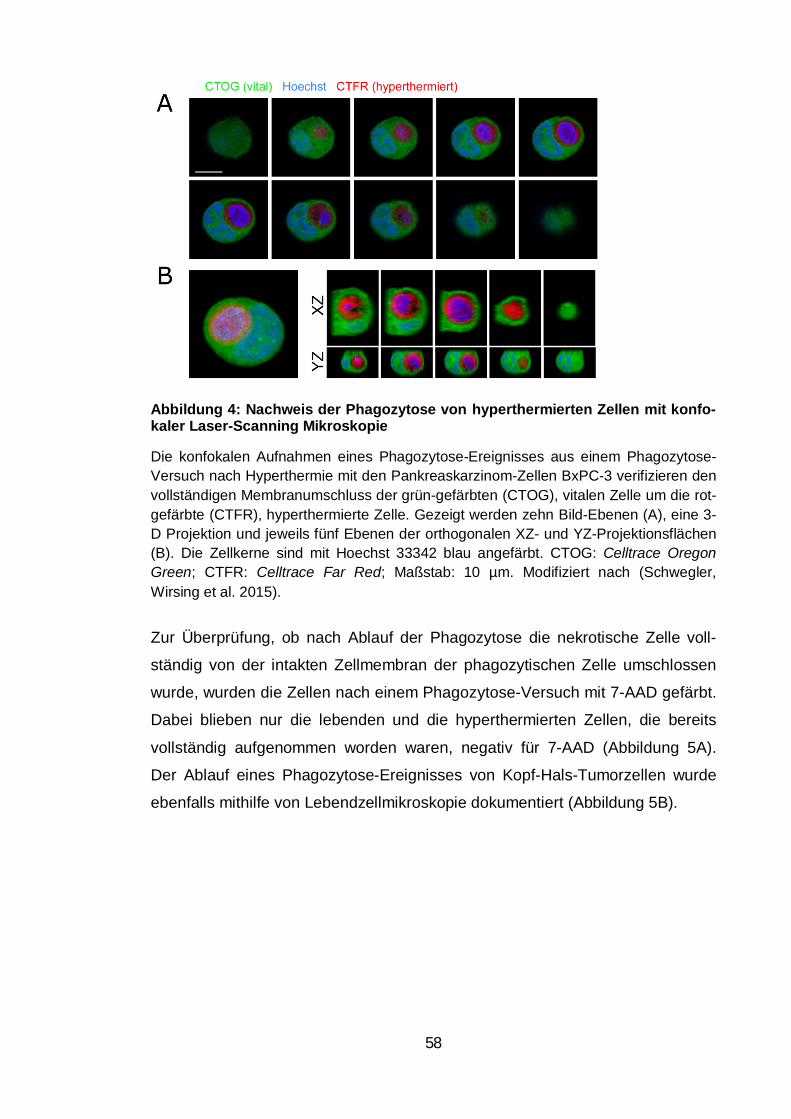
58
Abbildung 4: Nachweis der Phagozytose von hyperthermierten Zellen mit konfo-kaler Laser-Scanning Mikroskopie
Die konfokalen Aufnahmen eines Phagozytose-Ereignisses aus einem Phagozytose-Versuch nach Hyperthermie mit den Pankreaskarzinom-Zellen BxPC-3 verifizieren den vollständigen Membranumschluss der grün-gefärbten (CTOG), vitalen Zelle um die rot-gefärbte (CTFR), hyperthermierte Zelle. Gezeigt werden zehn Bild-Ebenen (A), eine 3-D Projektion und jeweils fünf Ebenen der orthogonalen XZ- und YZ-Projektionsflächen (B). Die Zellkerne sind mit Hoechst 33342 blau angefärbt. CTOG: Celltrace Oregon Green; CTFR: Celltrace Far Red; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Zur Überprüfung, ob nach Ablauf der Phagozytose die nekrotische Zelle voll-
ständig von der intakten Zellmembran der phagozytischen Zelle umschlossen
wurde, wurden die Zellen nach einem Phagozytose-Versuch mit 7-AAD gefärbt.
Dabei blieben nur die lebenden und die hyperthermierten Zellen, die bereits
vollständig aufgenommen worden waren, negativ für 7-AAD (Abbildung 5A).
Der Ablauf eines Phagozytose-Ereignisses von Kopf-Hals-Tumorzellen wurde
ebenfalls mithilfe von Lebendzellmikroskopie dokumentiert (Abbildung 5B).

59
Abbildung 5: Nachweis der Phagozytose von hyperthermierten Zellen mit Fluo-reszenz- und Lebendzell-Mikroskopie
Zur Überprüfung des vollständigen Membranumschlusses durch die phagozytische, grün-gefärbte (CTOG) Zelle um die hyperthermierte, ungefärbte Zelle wurden BxPC-3 Pankreaskarzinom-Zellen nach dem Phagozytose-Versuch mit den DNA-Farbstoffen DAPI (blau) und 7-AAD (rot) angefärbt. Freie, nekrotische Zellen wurden mit dem Membran-impermeablen Farbstoff 7-AAD gefärbt, wohingegen vitale sowie phagozy-tierte Zellen negativ für 7-AAD blieben (A). Ein charakteristisches Phagozytose-Ereignis wurde mittels Lebend-Zell-Mikroskopie aufgenommen. Dargestellt ist der Auf-nahme-Prozess einer hyperthermierten Zelle bei der primären Kopf-Hals-Tumorzelllinie p26 (B). CTOG: Celltrace Oregon Green; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Induktion von Nekrose und Phagozytose 3.2.Nach starker Schädigung der Zellmembran durch physikalische Einflüsse, wie
zum Beispiel Hitze-Behandlung, tritt die primäre Nekrose ein. Als weitere Noxe
wurde ionisierende Bestrahlung verwendet. Der Anteil von nekrotischen Zellen
wurde mithilfe des Zellmembran-impermeablen DNA-Farbstoffs 7-
Aminoactinomycin (7-AAD) durchflusszytometrisch bestimmt. Außerdem wurde
eine Induktion der Caspase-3-abhängigen Apoptose durch Immunfluoreszenz-
Färbung untersucht. Dabei wurde insbesondere der Fragestellung nachgegan-
gen, ob bestrahlte Zellen in Apoptose gehen können, nachdem sie phagozytiert
wurden.
Hyperthermie 3.2.1.
Die Behandlung mit Hyperthermie im Wasserbad bei 56 °C für 45 min induzierte
in 100 % der behandelten Pankreaskarzinom-Zellen BxPC-3 unmittelbar nach
der Behandlung den nekrotischen Zelltod. Wurden die Zellen mit niedrigeren
Temperaturen (48 °C) behandelt, trat die Membranpermeabilität für 7-AAD erst
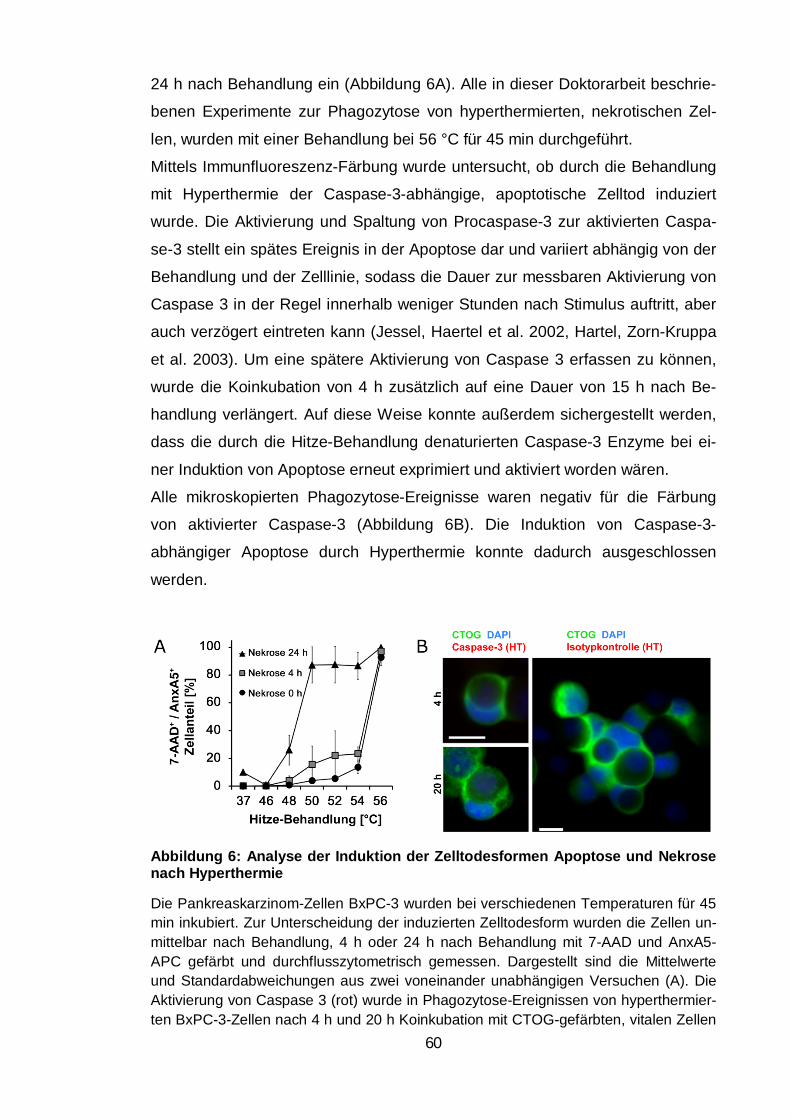
60
24 h nach Behandlung ein (Abbildung 6A). Alle in dieser Doktorarbeit beschrie-
benen Experimente zur Phagozytose von hyperthermierten, nekrotischen Zel-
len, wurden mit einer Behandlung bei 56 °C für 45 min durchgeführt.
Mittels Immunfluoreszenz-Färbung wurde untersucht, ob durch die Behandlung
mit Hyperthermie der Caspase-3-abhängige, apoptotische Zelltod induziert
wurde. Die Aktivierung und Spaltung von Procaspase-3 zur aktivierten Caspa-
se-3 stellt ein spätes Ereignis in der Apoptose dar und variiert abhängig von der
Behandlung und der Zelllinie, sodass die Dauer zur messbaren Aktivierung von
Caspase 3 in der Regel innerhalb weniger Stunden nach Stimulus auftritt, aber
auch verzögert eintreten kann (Jessel, Haertel et al. 2002, Hartel, Zorn-Kruppa
et al. 2003). Um eine spätere Aktivierung von Caspase 3 erfassen zu können,
wurde die Koinkubation von 4 h zusätzlich auf eine Dauer von 15 h nach Be-
handlung verlängert. Auf diese Weise konnte außerdem sichergestellt werden,
dass die durch die Hitze-Behandlung denaturierten Caspase-3 Enzyme bei ei-
ner Induktion von Apoptose erneut exprimiert und aktiviert worden wären.
Alle mikroskopierten Phagozytose-Ereignisse waren negativ für die Färbung
von aktivierter Caspase-3 (Abbildung 6B). Die Induktion von Caspase-3-
abhängiger Apoptose durch Hyperthermie konnte dadurch ausgeschlossen
werden.
Abbildung 6: Analyse der Induktion der Zelltodesformen Apoptose und Nekrose nach Hyperthermie
Die Pankreaskarzinom-Zellen BxPC-3 wurden bei verschiedenen Temperaturen für 45 min inkubiert. Zur Unterscheidung der induzierten Zelltodesform wurden die Zellen un-mittelbar nach Behandlung, 4 h oder 24 h nach Behandlung mit 7-AAD und AnxA5-APC gefärbt und durchflusszytometrisch gemessen. Dargestellt sind die Mittelwerte und Standardabweichungen aus zwei voneinander unabhängigen Versuchen (A). Die Aktivierung von Caspase 3 (rot) wurde in Phagozytose-Ereignissen von hyperthermier-ten BxPC-3-Zellen nach 4 h und 20 h Koinkubation mit CTOG-gefärbten, vitalen Zellen

61
(grün) untersucht (B). Die Zellkerne sind mit DAPI blau angefärbt. CTOG: Celltrace Oregon Green; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Im nächsten Schritt wurden die Überlebensraten nach Hyperthermie-
Behandlung mit verschiedenen Temperaturen mit einem Koloniebildungstest
eruiert (Abbildung 7). Vergleichsweise milder Hitzeschock bei 40 °C resultierte
in Überlebensraten von 75 % bei den Pankreaskarzinom-Zellen BxPC-3, nach
Hitze-Behandlung bei 45 °C hingegen schrumpfte die Überlebensrate auf 6 %.
Bei Behandlung mit 56 °C fand keine Proliferation statt.
Dargestellt sind die relativen Werte in Bezug zur unbehandelten Kontrolle (Kontrol-le=100 %). Mittelwerte und Standardabwei-chungen aus drei unabhängigen, in Triplika-ten durchgeführten, Versuchen. **p < 0,01, ***p ≤ 0,001. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Bestrahlung 3.2.2.
Ionisierende Strahlung wurde als weitere Methode zur Induktion von Zelltod, der
eine Phagozytose stimuliert, gewählt. Verschiedene Dosen und Inkubationszei-
ten nach Bestrahlung wurden mit drei verschiedenen Zelllinien getestet (Abbil-
dung 8). Bei Jurkat T-Zell-Leukämiezellen konnte eine Nekrose-Induktions-Rate
von 41 % nach einer Bestrahlung mit 8 Gy und drei Tagen Inkubationszeit bei
37 °C im CO2-Inkubator erzielt werden. Zugleich wurde ein Anteil apoptotischer
Zellen von 19 % gemessen.
Aufgrund der notwendigen Inkubationsdauer nach Bestrahlung und des Anteils
apoptotischer Zellen ist es möglich, dass es sich bei der hier beobachteten
Membrandurchlässigkeit für 7-AAD um sekundäre Nekrose handelt. Ein Anteil
von 75 % der Pankreaskarzinom-Zellen BxPC-3 wurde durch Bestrahlung mit
20 Gy und anschließender Inkubationsdauer von sechs Tagen nekrotisiert.
Apoptotischer Zelltod wurde dabei in 14 % der bestrahlten Zellen induziert. Die
primäre Kopf-Hals-Tumorzelllinie p26 wurde mit fünf Fraktionen von je 2 Gy im
Abstand von 24 h bestrahlt. Nach der anschließenden Inkubationsdauer von
Abbildung 7: Koloniebildungspotenzial nach Hitze-Behandlung
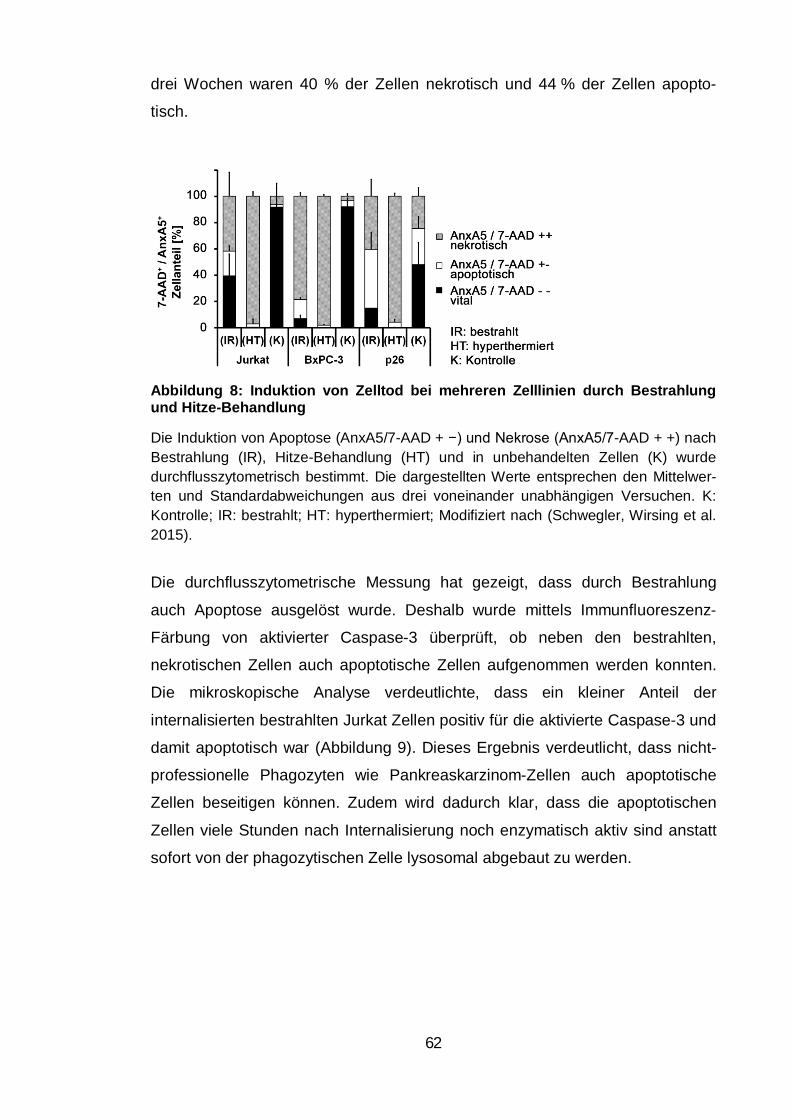
62
drei Wochen waren 40 % der Zellen nekrotisch und 44 % der Zellen apopto-
tisch.
Abbildung 8: Induktion von Zelltod bei mehreren Zelllinien durch Bestrahlung und Hitze-Behandlung
Die Induktion von Apoptose (AnxA5/7-AAD + −) und Nekrose (AnxA5/7-AAD + +) nach Bestrahlung (IR), Hitze-Behandlung (HT) und in unbehandelten Zellen (K) wurde durchflusszytometrisch bestimmt. Die dargestellten Werte entsprechen den Mittelwer-ten und Standardabweichungen aus drei voneinander unabhängigen Versuchen. K: Kontrolle; IR: bestrahlt; HT: hyperthermiert; Modifiziert nach (Schwegler, Wirsing et al. 2015).
Die durchflusszytometrische Messung hat gezeigt, dass durch Bestrahlung
auch Apoptose ausgelöst wurde. Deshalb wurde mittels Immunfluoreszenz-
Färbung von aktivierter Caspase-3 überprüft, ob neben den bestrahlten,
nekrotischen Zellen auch apoptotische Zellen aufgenommen werden konnten.
Die mikroskopische Analyse verdeutlichte, dass ein kleiner Anteil der
internalisierten bestrahlten Jurkat Zellen positiv für die aktivierte Caspase-3 und
damit apoptotisch war (Abbildung 9). Dieses Ergebnis verdeutlicht, dass nicht-
professionelle Phagozyten wie Pankreaskarzinom-Zellen auch apoptotische
Zellen beseitigen können. Zudem wird dadurch klar, dass die apoptotischen
Zellen viele Stunden nach Internalisierung noch enzymatisch aktiv sind anstatt
sofort von der phagozytischen Zelle lysosomal abgebaut zu werden.

63
Abbildung 9: Phagozytose von apoptotischen Zellen nach Bestrahlung
Die Aktivierung von Caspase 3 (rot) wurde in Phagozytose-Ereignissen von bestrahlten Jurkat T-Zellen nach 4 h (A) und 20 h (B) Koinkubation mit CTOG-gefärbten (grün), vitalen Pankreaskarzinom-Zellen BxPC-3 untersucht. Die Isotypkontrolle (rot) bestätigt die Spezifität des verwendeten Antikörpers gegen aktivierte Caspase 3 (C). Die Zell-kerne sind mit DAPI blau angefärbt. CTOG: Celltrace Oregon Green; IR: bestrahlt; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Phagozytose von hyperthermierten Zellen 3.3.Der Versuchsablauf beinhaltete die Vitalfärbung, Hitze-Behandlung einer Probe
und die homotypische (eine Zelllinie) sowie heterotypische (zwei Zelllinien)
Koinkubation mit der unbehandelten Probe in Suspension für 4 h. Anschließend
wurden die Zellen auf Objektträger zentrifugiert, mikroskopiert und die Phagozy-
tose-Ereignisse mit einem Bildanalyse-Programm quantifiziert. Außerdem wur-
den der Hyperthermie-induzierte Protein-Verlust und die HSP70-Freisetzung
nachgewiesen. Zudem wurde der Einfluss von freigesetzten DAMPs durch die
Behandlung mit Hyperthermie sowie die Rolle von FBS bei der Phagozytose
betrachtet. Darüber hinaus wurden angewachsene Zellen mit hyperthermierten
Zellen für 4 h überschichtet, um die Phagozytose-Aktivität im Zellverband, der
für adhärente Zelllinien eine natürlichere Bedingung darstellt als das Suspensi-
onssystem, untersucht.
Quantifizierung der homotypischen Phagozytose-Raten bei ver-3.3.1.
schiedenen Zelltypen Der Phagozytose-Versuch zur Aufnahme von hyperthermierten Zellen wurde
mit elf Zelllinien durchgeführt. Damit sollte untersucht werden, ob es sich um ein
Zelltyp-selektives Phänomen handeln könnte. Zu den verwendeten Zelllinien
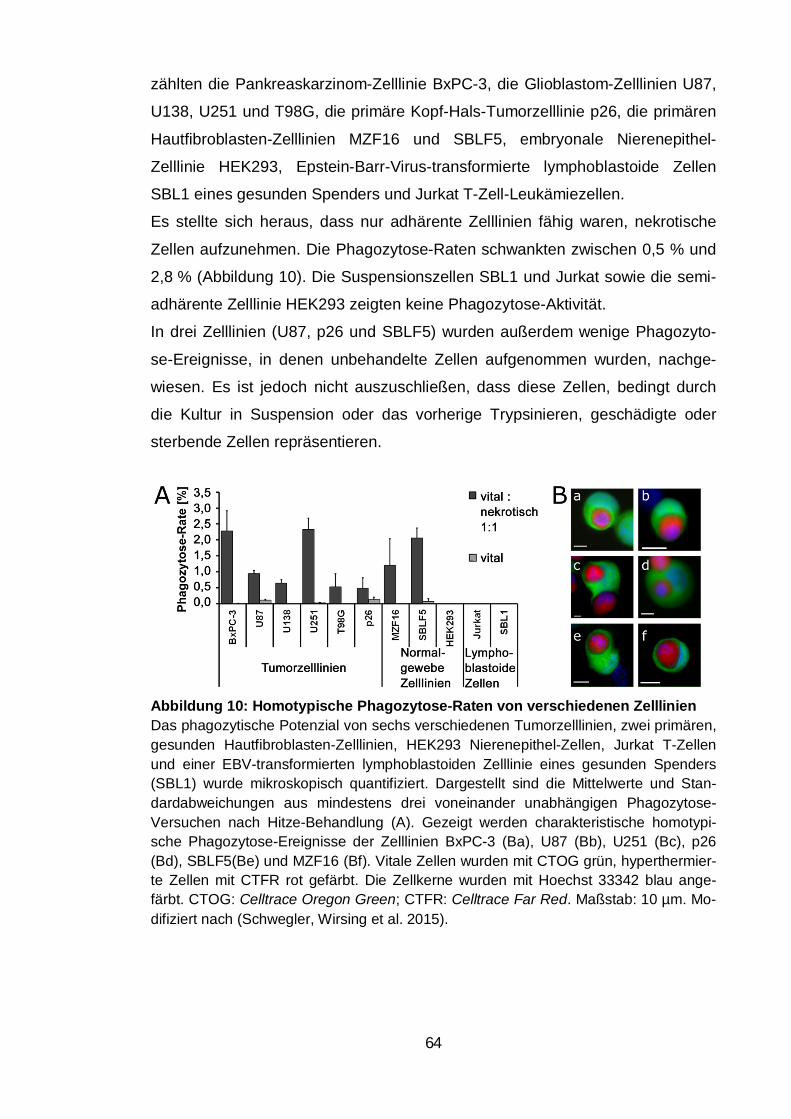
64
zählten die Pankreaskarzinom-Zelllinie BxPC-3, die Glioblastom-Zelllinien U87,
U138, U251 und T98G, die primäre Kopf-Hals-Tumorzelllinie p26, die primären
Hautfibroblasten-Zelllinien MZF16 und SBLF5, embryonale Nierenepithel-
Zelllinie HEK293, Epstein-Barr-Virus-transformierte lymphoblastoide Zellen
SBL1 eines gesunden Spenders und Jurkat T-Zell-Leukämiezellen.
Es stellte sich heraus, dass nur adhärente Zelllinien fähig waren, nekrotische
Zellen aufzunehmen. Die Phagozytose-Raten schwankten zwischen 0,5 % und
2,8 % (Abbildung 10). Die Suspensionszellen SBL1 und Jurkat sowie die semi-
adhärente Zelllinie HEK293 zeigten keine Phagozytose-Aktivität.
In drei Zelllinien (U87, p26 und SBLF5) wurden außerdem wenige Phagozyto-
se-Ereignisse, in denen unbehandelte Zellen aufgenommen wurden, nachge-
wiesen. Es ist jedoch nicht auszuschließen, dass diese Zellen, bedingt durch
die Kultur in Suspension oder das vorherige Trypsinieren, geschädigte oder
sterbende Zellen repräsentieren.
Abbildung 10: Homotypische Phagozytose-Raten von verschiedenen Zelllinien Das phagozytische Potenzial von sechs verschiedenen Tumorzelllinien, zwei primären, gesunden Hautfibroblasten-Zelllinien, HEK293 Nierenepithel-Zellen, Jurkat T-Zellen und einer EBV-transformierten lymphoblastoiden Zelllinie eines gesunden Spenders (SBL1) wurde mikroskopisch quantifiziert. Dargestellt sind die Mittelwerte und Stan-dardabweichungen aus mindestens drei voneinander unabhängigen Phagozytose-Versuchen nach Hitze-Behandlung (A). Gezeigt werden charakteristische homotypi-sche Phagozytose-Ereignisse der Zelllinien BxPC-3 (Ba), U87 (Bb), U251 (Bc), p26 (Bd), SBLF5(Be) und MZF16 (Bf). Vitale Zellen wurden mit CTOG grün, hyperthermier-te Zellen mit CTFR rot gefärbt. Die Zellkerne wurden mit Hoechst 33342 blau ange-färbt. CTOG: Celltrace Oregon Green; CTFR: Celltrace Far Red. Maßstab: 10 µm. Mo-difiziert nach (Schwegler, Wirsing et al. 2015).
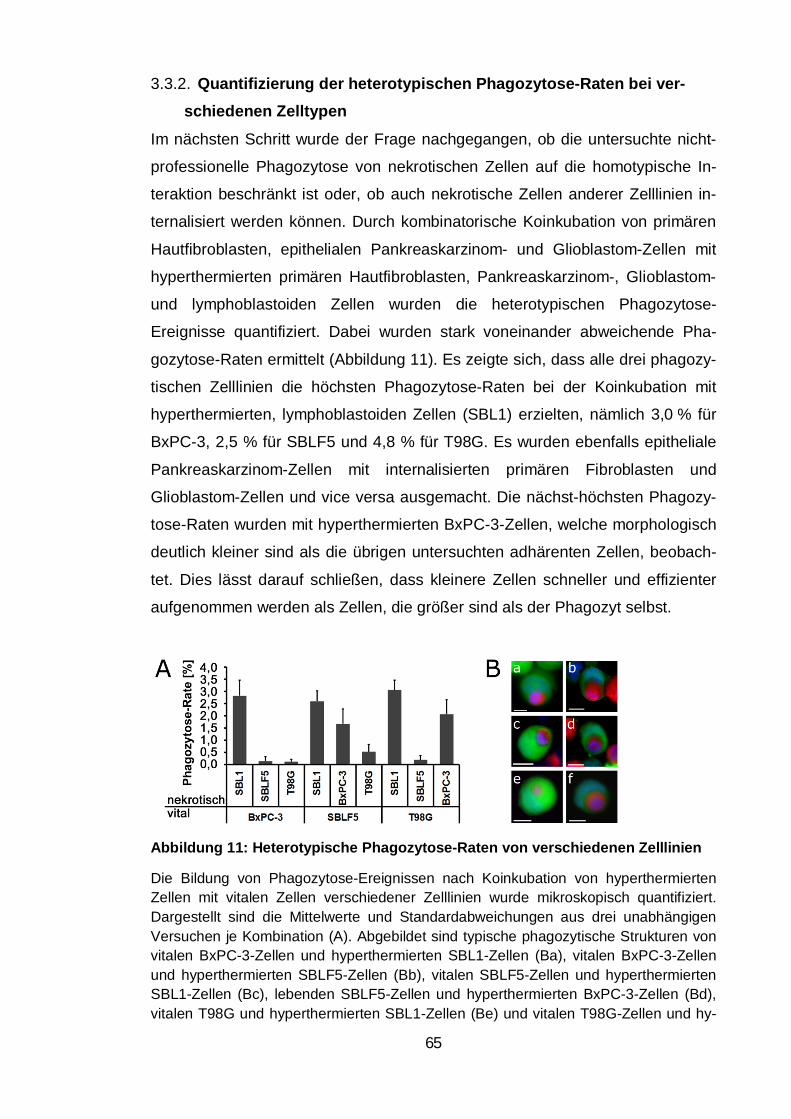
65
Quantifizierung der heterotypischen Phagozytose-Raten bei ver-3.3.2.
schiedenen Zelltypen
Im nächsten Schritt wurde der Frage nachgegangen, ob die untersuchte nicht-
professionelle Phagozytose von nekrotischen Zellen auf die homotypische In-
teraktion beschränkt ist oder, ob auch nekrotische Zellen anderer Zelllinien in-
ternalisiert werden können. Durch kombinatorische Koinkubation von primären
Hautfibroblasten, epithelialen Pankreaskarzinom- und Glioblastom-Zellen mit
hyperthermierten primären Hautfibroblasten, Pankreaskarzinom-, Glioblastom-
und lymphoblastoiden Zellen wurden die heterotypischen Phagozytose-
Ereignisse quantifiziert. Dabei wurden stark voneinander abweichende Pha-
gozytose-Raten ermittelt (Abbildung 11). Es zeigte sich, dass alle drei phagozy-
tischen Zelllinien die höchsten Phagozytose-Raten bei der Koinkubation mit
hyperthermierten, lymphoblastoiden Zellen (SBL1) erzielten, nämlich 3,0 % für
BxPC-3, 2,5 % für SBLF5 und 4,8 % für T98G. Es wurden ebenfalls epitheliale
Pankreaskarzinom-Zellen mit internalisierten primären Fibroblasten und
Glioblastom-Zellen und vice versa ausgemacht. Die nächst-höchsten Phagozy-
tose-Raten wurden mit hyperthermierten BxPC-3-Zellen, welche morphologisch
deutlich kleiner sind als die übrigen untersuchten adhärenten Zellen, beobach-
tet. Dies lässt darauf schließen, dass kleinere Zellen schneller und effizienter
aufgenommen werden als Zellen, die größer sind als der Phagozyt selbst.
Abbildung 11: Heterotypische Phagozytose-Raten von verschiedenen Zelllinien
Die Bildung von Phagozytose-Ereignissen nach Koinkubation von hyperthermierten Zellen mit vitalen Zellen verschiedener Zelllinien wurde mikroskopisch quantifiziert. Dargestellt sind die Mittelwerte und Standardabweichungen aus drei unabhängigen Versuchen je Kombination (A). Abgebildet sind typische phagozytische Strukturen von vitalen BxPC-3-Zellen und hyperthermierten SBL1-Zellen (Ba), vitalen BxPC-3-Zellen und hyperthermierten SBLF5-Zellen (Bb), vitalen SBLF5-Zellen und hyperthermierten SBL1-Zellen (Bc), lebenden SBLF5-Zellen und hyperthermierten BxPC-3-Zellen (Bd), vitalen T98G und hyperthermierten SBL1-Zellen (Be) und vitalen T98G-Zellen und hy-

66
perthermierten BxPC-3-Zellen (Bf); Vitale Zellen wurden mit CTOG grün, hyperther-mierte Zellen mit CTFR rot gefärbt. Die Zellkerne wurden mit Hoechst 33342 blau an-gefärbt. CTOG: Celltrace Oregon Green; CTFR: Celltrace Far Red; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Protein-Verlust und Freisetzung von HSP70 durch Hyperthermie 3.3.3.
Professionelle Antigen-präsentierende Zellen, wie dendritische Zellen, Makro-
phagen und B-Lymphozyten werden durch DAMPs, die von sterbenden Zellen
freigesetzt werden, aktiviert (Matzinger 2002). Dieses intrazelluläre Material
kann aber auch vorerst an der Zellmembran gebunden bleiben und dadurch
Immunzellen aktivieren, wie es für F-Aktin gezeigt wurde (Ahrens, Zelenay et al.
2012). Zur Exposition und Freisetzung von DAMPs in die Umgebung zählen
auch pro-phagozytische Signale, die auf sterbenden Zellen exponiert oder von
ihnen abgeben werden (Grimsley and Ravichandran 2003). Nekrotische Zellen
geben passiv intrazelluläres Material in ihre Umgebung ab. Daher wurde mit
einem bicinchoninic acid assay (BCA) überprüft, ob auch die Hitze-behandelten
Zellen einen Verlust von intrazellulärem Material aufweisen. Dafür wurden
Pankreaskarzinom- Zellen (BxPC-3), Kopf-Hals-Tumorzellen (p26) und
gesunden Hautfibroblasten (SBLF5) trypsiniert und eine Probe je Zelllinie
hyperthermiert (45 min bei 56 °C). Anschließend wurden aus den Zelllysaten
einer definierten Anzahl Zellen der jeweiligen Zelllinie die zellulären Proteine
isoliert. Die photometrische Messung zeigte eine starke Reduktion der Gesamt-
Protein-Konzentration bzw. einen vollständigen Verlust der Proteine nach
Hyperthermie (Abbildung 12A). Dies verdeutlicht, dass die Behandlung primäre
Nekrose induzierte.
Im nächsten Versuch wurde die Freisetzung von HSP70 in den Überstand nach
Hyperthermie bei Pankreaskarzinom-Zellen mit einem ELISA gemessen. Wie
im Phagozytose-Versuch wurden die Zellen nach Behandlung 4 h in
Suspension inkubiert. Zusätzlich wurden Proben für einen 24 h Wert hergestellt.
Zur Kontrolle wurden unbehandelte Zellen mitgeführt. Es wurde keine
signifikante Erhöhung der HSP70-Konzentration nach Hitze-Behandlung
gemessen. Gegensätzlich zur Annahme, dass hyperthermierte Zellen eine
erhöhte HSP70-Freisetzung zeigen würden, wurde eine vermehrte Sekretion
von HSP70 bei den Proben mit vitalen Zellen gemessen. In diesen Proben war
nach 4 h eine Konzentration von ca. 1250 pg/105 festzustellen, nach 24 h stieg

67
die HSP70-Konzentration auf ca. 2450 pg/105 an. Bei den hyperthermierten
Zellen wurde eine vergleichsweise niedrige Konzentration von ca. 500 pg/105
Zellen nach 4 h in Suspension und von ca. 1000 pg/105 nach 24 h in
Suspension nachgewiesen. Unabhängig von der Behandlung wurden in beiden
Proben eine Verdopplung der HSP70-Freisetzung nach 4 h zum 24 h Wert
gemessen (Abbildung 12B).
Abbildung 12: Einfluss von Hyperthermie auf die Gesamt-Protein-Konzentration und die Freisetzung von HSP70
Die Gesamt-Protein-Konzentration der unbehandelten (K) und hyperthermierten (HT) Zelllysate mehrerer Zelllinien wurde nach Durchführung eines BCA-Versuchs photometrisch gemessen (A). Die Konzentration von HSP70 in den Überständen von vitalen und hyperthermierten (HT) Zellen wurde nach 4 h und 24 h in Suspensions-kultur mit einem ELISA bestimmt (B). Dargestellt sind die Mittelwerte und Standabweichungen eines repräsentativen Versuchs von zwei, in Duplikaten durchgeführten Versuchen.
Einfluss von freigesetzten DAMPs und FBS auf die Phagozytose 3.3.4.
Zur Überprüfung, ob nicht-professionelle Phagozyten durch freigesetzte DAMPs
zur Phagozytose angeregt werden, wurde in einem Versuch der Einfluss von
DAMPs auf die Phagozytose untersucht.
Um dies zu erreichen, wurden eine primäre Hautfibroblasten-Zelllinie, eine
Glioblastom-Zelllinie und die Pankreaskarzinom-Zelllinie gewählt und nach Hit-
ze-Behandlung zweimal mit PBS gewaschen bevor sie mit den unbehandelten
Zellen koinkubiert wurden. Die Entfernung des Mediums stellte sicher, dass die
Menge an bei der Hitze-Behandlung freigesetzten, intrazellulären Molekülen
reduziert wurde. Obwohl die Konzentration während der Koinkubation wieder
ansteigen würde, würde dennoch eine verringerte Phagozytose-Rate erwartet,
da die Aktivierung anfangs schwächer ausfallen oder sogar erst später eintreten
würde.
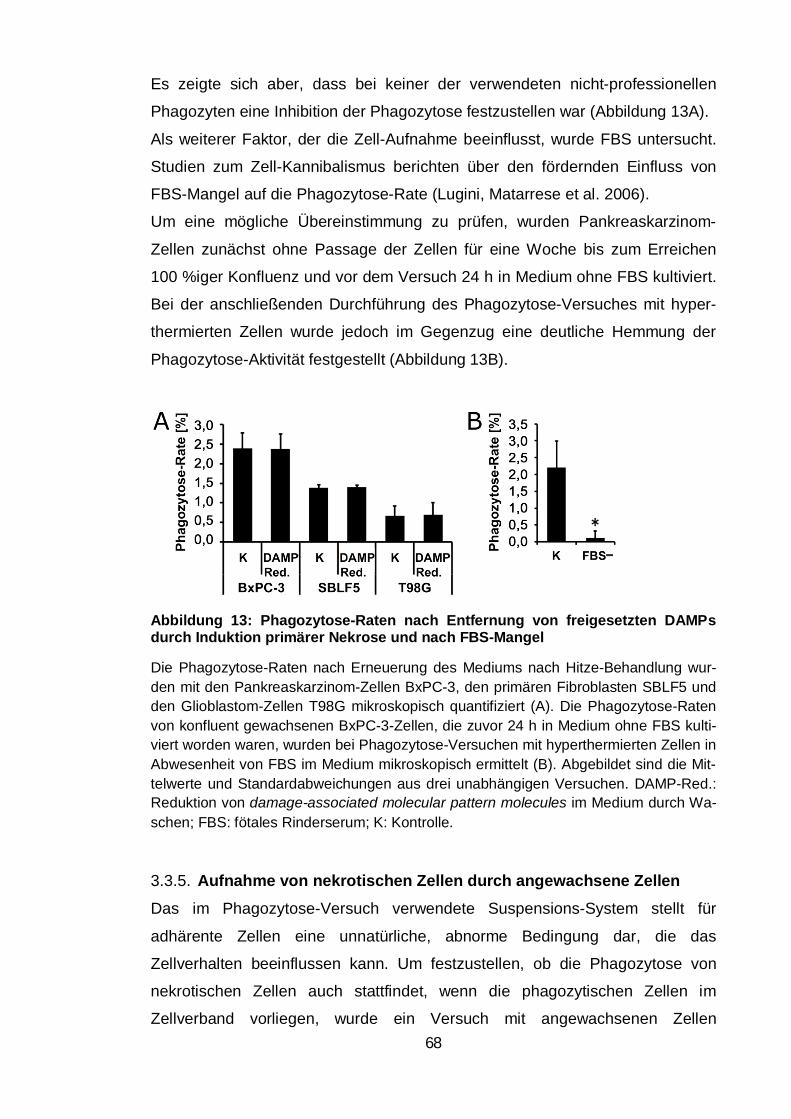
68
Es zeigte sich aber, dass bei keiner der verwendeten nicht-professionellen
Phagozyten eine Inhibition der Phagozytose festzustellen war (Abbildung 13A).
Als weiterer Faktor, der die Zell-Aufnahme beeinflusst, wurde FBS untersucht.
Studien zum Zell-Kannibalismus berichten über den fördernden Einfluss von
FBS-Mangel auf die Phagozytose-Rate (Lugini, Matarrese et al. 2006).
Um eine mögliche Übereinstimmung zu prüfen, wurden Pankreaskarzinom-
Zellen zunächst ohne Passage der Zellen für eine Woche bis zum Erreichen
100 %iger Konfluenz und vor dem Versuch 24 h in Medium ohne FBS kultiviert.
Bei der anschließenden Durchführung des Phagozytose-Versuches mit hyper-
thermierten Zellen wurde jedoch im Gegenzug eine deutliche Hemmung der
Phagozytose-Aktivität festgestellt (Abbildung 13B).
Abbildung 13: Phagozytose-Raten nach Entfernung von freigesetzten DAMPs durch Induktion primärer Nekrose und nach FBS-Mangel
Die Phagozytose-Raten nach Erneuerung des Mediums nach Hitze-Behandlung wur-den mit den Pankreaskarzinom-Zellen BxPC-3, den primären Fibroblasten SBLF5 und den Glioblastom-Zellen T98G mikroskopisch quantifiziert (A). Die Phagozytose-Raten von konfluent gewachsenen BxPC-3-Zellen, die zuvor 24 h in Medium ohne FBS kulti-viert worden waren, wurden bei Phagozytose-Versuchen mit hyperthermierten Zellen in Abwesenheit von FBS im Medium mikroskopisch ermittelt (B). Abgebildet sind die Mit-telwerte und Standardabweichungen aus drei unabhängigen Versuchen. DAMP-Red.: Reduktion von damage-associated molecular pattern molecules im Medium durch Wa-schen; FBS: fötales Rinderserum; K: Kontrolle.
Aufnahme von nekrotischen Zellen durch angewachsene Zellen 3.3.5.
Das im Phagozytose-Versuch verwendete Suspensions-System stellt für
adhärente Zellen eine unnatürliche, abnorme Bedingung dar, die das
Zellverhalten beeinflussen kann. Um festzustellen, ob die Phagozytose von
nekrotischen Zellen auch stattfindet, wenn die phagozytischen Zellen im
Zellverband vorliegen, wurde ein Versuch mit angewachsenen Zellen

69
durchgeführt. Dafür wurden Pankreaskarzinom-Zellen auf Objektträger
eingesät, nach 24 h mit dem Vitalfarbstoff CTOG grün gefärbt und mit CTFR-
gefärbten, hyperthermierten Zellen überschichtet. Nach 4 h bei 37 °C im CO2-
Inkubator wurden die Zellen fixiert und mikroskopiert (Abbildung 14). Die Bilder
beweisen, dass die Aufnahme von hyperthermierten Zellen auch von
angewachsenen Zellen ausgeführt werden kann und die Phagozytose in
Suspensionskultur damit kein künstlich erzeugtes Zellverhalten darstellt.
Abbildung 14: Phagozytose-Ereignisse bei angewachsenen Zellen
Gezeigt sind typische Phagozytose-Ereignisse von auf Objektträger angewachsenen, CTOG-gefärbten BxPC-3 Pankreaskarzinom-Zellen (grün), die CTFR-gefärbte, Hitze-behandelte BxPC-3-Zellen (rot) aufgenommen haben. Zellkerne sind mit DAPI blau gefärbt; CTOG: Celltrace Oregon Green; CTFR: Celltrace Far Red; Maßstab: 10 µm.
Phagozytose von bestrahlten Zellen und nekrotischer De-3.4.
bris Bei den Versuchen zur Induktion von Zelltod durch Bestrahlung wurde der Ein-
tritt von Apoptose und Nekrose nach Bestrahlung und folgender Inkubationszeit
bei BxPC-3 Pankreaskarzinom-Zellen, Jurkat T-Zell-Leukämiezellen und p26
Kopf-Hals-Tumorzellen beobachtet. Des Weiteren wurde die Aufnahme von
wenigen apoptotischen Zellen nachgewiesen (siehe 3.2.2). In nächsten Schritt
sollte die Phagozytose von bestrahlten Zellen quantifiziert und die Effizienz mit
den Phagozytose-Raten von hyperthermierten Zellen verglichen werden. Bei
der mikroskopischen Auswertung dieser Versuche wurde eine große Menge
von nekrotischer Debris nach Bestrahlung ausgemacht.
Die Anhäufung von nekrotischer Debris, welche pro-phagozytische eat-me-
Signale präsentiert und DAMPs freisetzt, gehört zum nekrotischen Zellunter-
gang dazu. Diese zellulären Bestandteile werden im Gewebe von professionel-
len Phagozyten effizient entfernt. Nachdem gezeigt werden konnte, dass nekro-
tische Zellen von nicht-professionellen Phagozyten aufgenommen werden kön-

70
nen, sollte untersucht werden, ob sich diese Phagozytose-Aktivität nur auf gan-
ze Zellen beschränkt oder, ob ebenso Bruchstücke aufgenommen werden.
Quantifizierung der homotypischen und heterotypischen Phagozy-3.4.1.
tose-Raten bei verschiedenen Zelltypen Für die Induktion von Apoptose und Nekrose wurden die Zellen mit unterschied-
lichen Dosen Röntgenstrahlung bestrahlt und anschließend inkubiert (Tabelle
12). Tabelle 12: Induktion von Zelltod durch Bestrahlung
Zelllinie Dosis [Gy] Inkubationszeit [d] Jurkat 8 3 BxPC-3 20 6 P26 5*2 (alle 24 h) 21
Beim Phagozytose-Versuch mit bestrahlten Zellen konnten ebenfalls Phagozy-
tose-Ereignisse nachgewiesen werden (Abbildung 15). Die Häufigkeiten belie-
fen sich auf niedrige Werte für die homotypische Aufnahme von bestrahlten
BxPC-3 (0,5 %) und p26 (0,9 %) Zellen und vergleichsweise hohe Werte für die
heterotypische Aufnahme von bestrahlten Jurkat-Zellen durch BxPC-3-Zellen
(4,2 %) und SBLF5-Zellen (5,7 %). Es wurden keine signifikanten Unterschiede
bei den Phagozytose-Raten mit bestrahlten Zellen im Vergleich mit den Pha-
gozytose-Raten mit hyperthermierten (45 min bei 56 °C) Zellen festgestellt.
Abbildung 15: Vergleich der Phagozytose-Raten von hyperthermierten und be-strahlten Zellen bei verschiedenen Zelllinien
Die phagozytische Effizienz der Pankreaskarzinom-Zellen BxPC-3, der primären, ge-sunden Hautfibroblasten SBLF5 und der p26 Kopf-Hals-Tumorzellen wurde nach ho-motypischer und heterotypischer Koinkubation mit hyperthermierten und bestrahlten Zellen mikroskopisch quantifiziert (A). Abgebildet sind die Mittelwerte und Standardab-

71
weichungen von drei unabhängigen Versuchen. Gezeigt werden zwei charakteristische Phagozytose-Ereignisse nach Koinkubation von vitalen, CTOG-gefärbten BxPC-3-Zellen mit bestrahlten Jurkat T-Zellen (Ba) und vitalen und bestrahlten Kopf-Hals-Tumorzellen P26 (Bb). Die Zellkerne sind mit Hoechst 33342 blau gefärbt. CTOG: Cell-trace Oregon Green; CTFR: Celltrace Far Red; HT: hyperthermiert; IR: bestrahlt; v: vital; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Aufnahme nekrotischer Debris 3.4.2.
Für die Untersuchung der Phagozytose von nekrotischer Debris wurden Versu-
che mit nekrotischen Zellen nach Hyperthermie und Bestrahlung durchgeführt.
Nach Hitze-Behandlung wurden auf den mikroskopischen Präparaten nur weni-
ge freie Zellfragmente von nekrotischen Zellen ausgemacht. Dementsprechend
wurden auch nur sehr wenige phagozytische Strukturen mit nekrotischer Debris
beobachtet und zwar nicht mehr als ein oder zwei Ereignisse pro Experiment
(Abbildung 16A). Wegen dieser niedrigen Häufigkeit wurden diese Phagozyto-
se-Ereignisse von der Quantifizierung ausgeschlossen.
Durch Bestrahlung mit anschließender langer Inkubationszeit hingegen, wurden
große Mengen nekrotischer Debris angehäuft. Zellfragmente unterschiedlicher
Größen sowie intakt erscheinende Zellen wurden auf den mikroskopischen
Präparaten beobachtet.
Die Auswertung erlaubte nicht immer eine verlässliche Aussage darüber, ob
das Zellfragment tatsächlich aufgenommen war oder lediglich auf die Oberflä-
che der phagozytischen Zelle zentrifugiert worden war. Um das Risiko für die
Quantifizierung von falsch-positiven Artefakten zu verringern, wurden per Defi-
nition phagozytische Strukturen auf solche beschränkt, die intakt erscheinende
Zellen mit Zellkern (Abbildung 16Ba) oder große Zellfragmente enthielten, wel-
che eine deutliche Verformung des phagozytischen Nukleus verursachten (Ab-
bildung 16Bb). Alle weiteren ähnlichen Zell-in-Zell Strukturen wurden bei der
Zählung nicht berücksichtigt (Abbildung 16Bc).

72
Abbildung 16: Phagozytose von nekrotischer Debris
Abgebildet sind homotypische, phagozytische Strukturen nach Aufnahme von nekroti-scher Debris nach Hyperthermie von BxPC-3- (Aa) und SBLF5-Zellen (Ab, c) und nach Bestrahlung von p26-Zellen die Phagozytose einer bestrahlten Zelle (Ba) und der nek-rotischen Debris (Bb, c). Vitale Zellen wurden mit CTOG grün gefärbt, hyperthermierte oder bestrahlte Zellen mit CTFR rot. Die Zellkerne wurden mit Hoechst 33342 blau angefärbt. CTOG: Celltrace Oregon Green; CTFR: Celltrace Far Red; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Einfluss von Hyperthermie auf die Zelladhäsion und Pha-3.5.
gozytose Der erste Schritt bei der Phagozytose ist eine starke Adhäsion zwischen pha-
gozytischer und aufzunehmender Zelle. Es wurde gezeigt, dass nekrotische
Zellen, aber nicht lebende Zellen, von nicht-professionellen Phagozyten aufge-
nommen werden können. Daher sollte untersucht werden, ob Unterschiede bei
der Adhäsion zwischen lebenden Zellen im Vergleich zur Adhäsion zwischen
vitalen und hyperthermierten (45 min bei 56 °C) Zellen bestehen. In weiteren
Versuchen wurden sowohl die Adhäsions-Rate als auch die Phagozytose-Rate
unter Verwendung verschiedener Behandlungs-Temperaturen bestimmt.
Mikroskopischer Nachweis der Adhäsion 3.5.1.
Für die Visualisierung der zielgerichteten Membran-Ausstülpungen der pha-
gozytierenden Zellen bei der Bindung an Hitze-behandelte Zellen wurde der

73
Phagozytose-Prozess mittels konfokaler Laser-Scanning-Mikroskopie und Le-
bendzell-Mikroskopie aufgenommen (Abbildung 17).
Abbildung 17: Verstärktes adhäsives Verhalten gegenüber hyperthermierter Zel-len
Gezeigt wird der Zustand verstärkter Adhäsion während des Aufnahme-Prozesses von hyperthermierten, CTFR-gefärbten (rot) Pankreaskarzinom-Zellen von Aktin-GFP-transfizierten (grün) Pankreaskarzinom-Zellen (A, B), oder ungefärbten Zellen, beo-bachtet mit dem Lebendzellmikroskop (C, D). Die Zellkerne wurden mit DAPI blau ge-färbt. CTFR: Celltrace Far Red; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Nachweis der Hyperthermie-induzierten Adhäsion mittels Durch-3.5.2.
flusszytometrie Im nächsten Schritt wurde ermittelt, inwiefern Hyperthermie-induzierte Memb-
ran-Veränderungen die Bindung zwischen phagozytischer und aufzunehmender
Zelle beeinflussen. Um dies zu untersuchen, wurden durchflusszytometrische
Messungen durchgeführt. Die Adhäsion wurde als Anteil der lebenden Zellen,
die an hyperthermierte Zellen gebunden hatten, berechnet. Dabei wurde in allen
verwendeten Zelllinien, mit Ausnahme der HEK293-, der Jurkat- und der SBL1-
Zellen, die keine hyperthermierten Zellen phagozytierten, eine Erhöhung der
Adhäsions-Rate festgestellt, wenn vitale mit Hitze-behandelten Zellen koinku-
biert wurden, im Vergleich zur Koinkubation von nur vitalen Zellen (Abbildung
18).
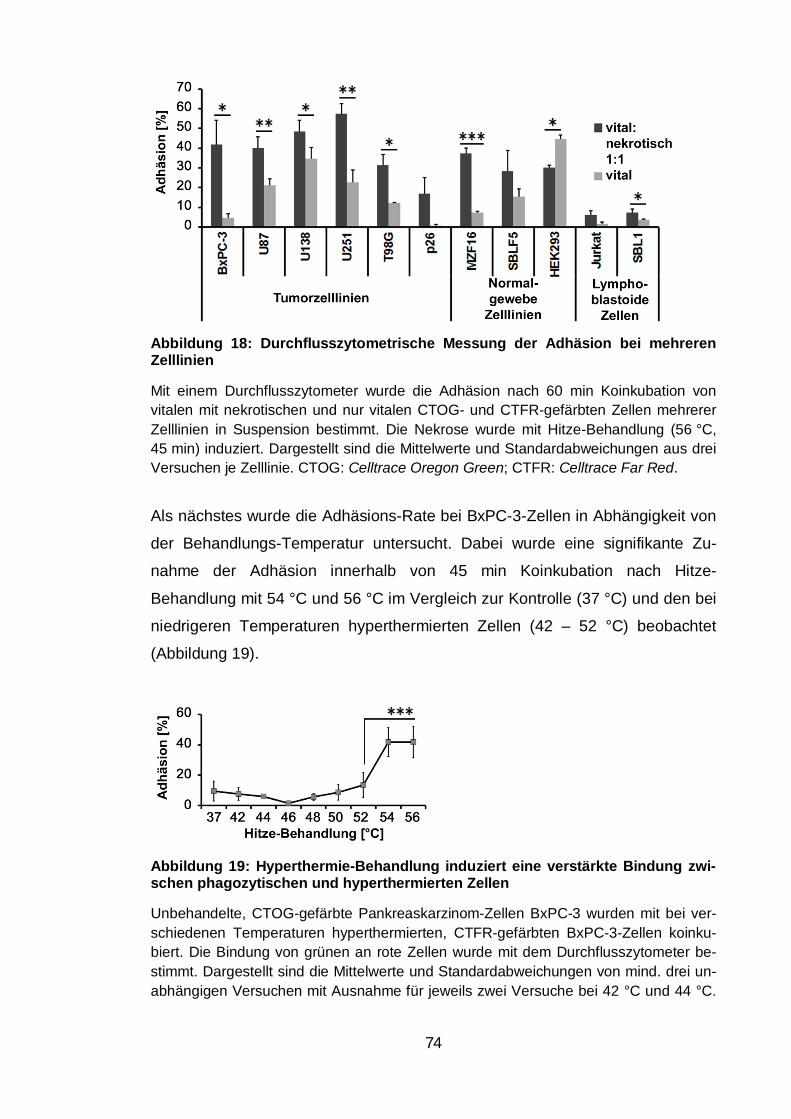
74
Abbildung 18: Durchflusszytometrische Messung der Adhäsion bei mehreren Zelllinien
Mit einem Durchflusszytometer wurde die Adhäsion nach 60 min Koinkubation von vitalen mit nekrotischen und nur vitalen CTOG- und CTFR-gefärbten Zellen mehrerer Zelllinien in Suspension bestimmt. Die Nekrose wurde mit Hitze-Behandlung (56 °C, 45 min) induziert. Dargestellt sind die Mittelwerte und Standardabweichungen aus drei Versuchen je Zelllinie. CTOG: Celltrace Oregon Green; CTFR: Celltrace Far Red.
Als nächstes wurde die Adhäsions-Rate bei BxPC-3-Zellen in Abhängigkeit von
der Behandlungs-Temperatur untersucht. Dabei wurde eine signifikante Zu-
nahme der Adhäsion innerhalb von 45 min Koinkubation nach Hitze-
Behandlung mit 54 °C und 56 °C im Vergleich zur Kontrolle (37 °C) und den bei
niedrigeren Temperaturen hyperthermierten Zellen (42 – 52 °C) beobachtet
(Abbildung 19).
Abbildung 19: Hyperthermie-Behandlung induziert eine verstärkte Bindung zwi-schen phagozytischen und hyperthermierten Zellen
Unbehandelte, CTOG-gefärbte Pankreaskarzinom-Zellen BxPC-3 wurden mit bei ver-schiedenen Temperaturen hyperthermierten, CTFR-gefärbten BxPC-3-Zellen koinku-biert. Die Bindung von grünen an rote Zellen wurde mit dem Durchflusszytometer be-stimmt. Dargestellt sind die Mittelwerte und Standardabweichungen von mind. drei un-abhängigen Versuchen mit Ausnahme für jeweils zwei Versuche bei 42 °C und 44 °C.
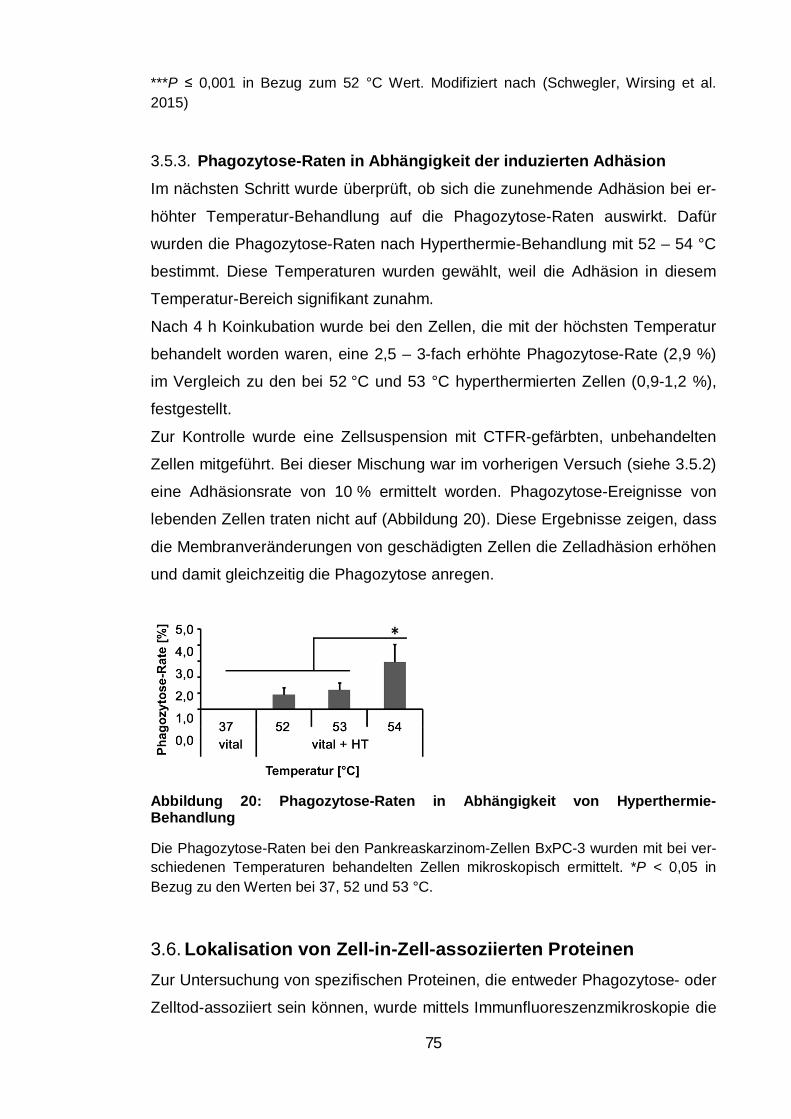
75
***P ≤ 0,001 in Bezug zum 52 °C Wert. Modifiziert nach (Schwegler, Wirsing et al. 2015)
Phagozytose-Raten in Abhängigkeit der induzierten Adhäsion 3.5.3.
Im nächsten Schritt wurde überprüft, ob sich die zunehmende Adhäsion bei er-
höhter Temperatur-Behandlung auf die Phagozytose-Raten auswirkt. Dafür
wurden die Phagozytose-Raten nach Hyperthermie-Behandlung mit 52 – 54 °C
bestimmt. Diese Temperaturen wurden gewählt, weil die Adhäsion in diesem
Temperatur-Bereich signifikant zunahm.
Nach 4 h Koinkubation wurde bei den Zellen, die mit der höchsten Temperatur
behandelt worden waren, eine 2,5 – 3-fach erhöhte Phagozytose-Rate (2,9 %)
im Vergleich zu den bei 52 °C und 53 °C hyperthermierten Zellen (0,9-1,2 %),
festgestellt.
Zur Kontrolle wurde eine Zellsuspension mit CTFR-gefärbten, unbehandelten
Zellen mitgeführt. Bei dieser Mischung war im vorherigen Versuch (siehe 3.5.2)
eine Adhäsionsrate von 10 % ermittelt worden. Phagozytose-Ereignisse von
lebenden Zellen traten nicht auf (Abbildung 20). Diese Ergebnisse zeigen, dass
die Membranveränderungen von geschädigten Zellen die Zelladhäsion erhöhen
und damit gleichzeitig die Phagozytose anregen.
Abbildung 20: Phagozytose-Raten in Abhängigkeit von Hyperthermie-Behandlung
Die Phagozytose-Raten bei den Pankreaskarzinom-Zellen BxPC-3 wurden mit bei ver-schiedenen Temperaturen behandelten Zellen mikroskopisch ermittelt. *P < 0,05 in Bezug zu den Werten bei 37, 52 und 53 °C.
Lokalisation von Zell-in-Zell-assoziierten Proteinen 3.6.Zur Untersuchung von spezifischen Proteinen, die entweder Phagozytose- oder
Zelltod-assoziiert sein können, wurde mittels Immunfluoreszenzmikroskopie die
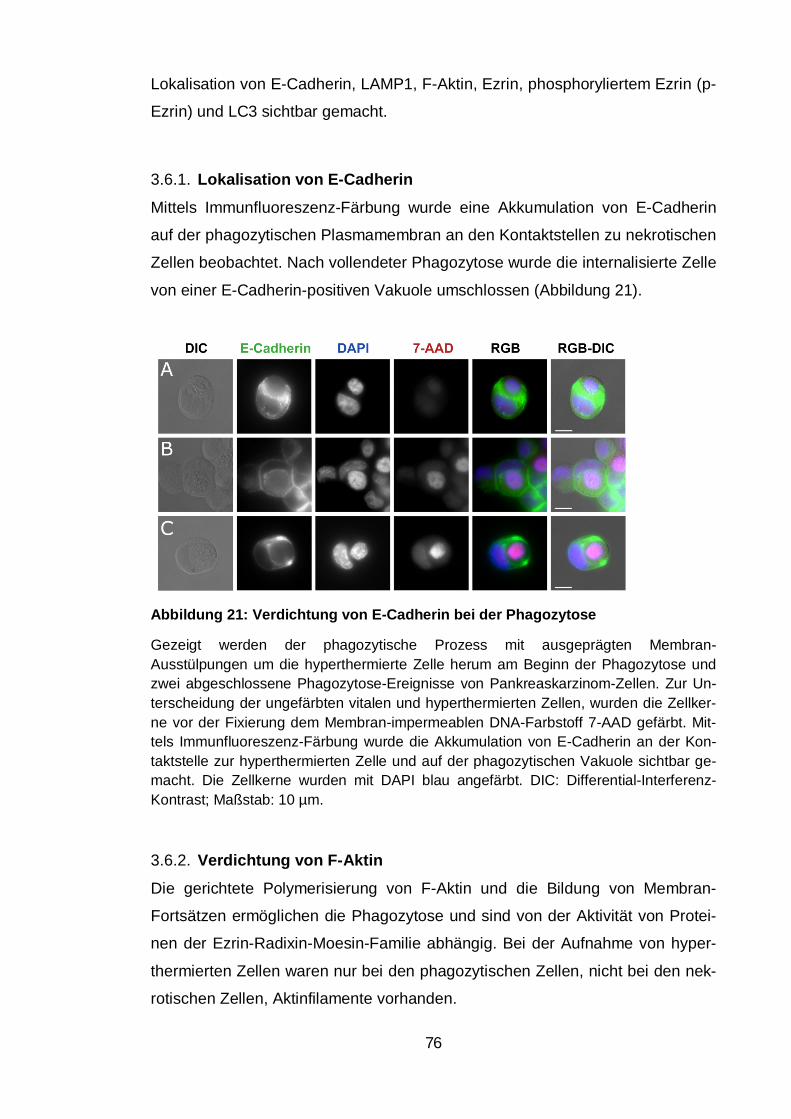
76
Lokalisation von E-Cadherin, LAMP1, F-Aktin, Ezrin, phosphoryliertem Ezrin (p-
Ezrin) und LC3 sichtbar gemacht.
Lokalisation von E-Cadherin 3.6.1.
Mittels Immunfluoreszenz-Färbung wurde eine Akkumulation von E-Cadherin
auf der phagozytischen Plasmamembran an den Kontaktstellen zu nekrotischen
Zellen beobachtet. Nach vollendeter Phagozytose wurde die internalisierte Zelle
von einer E-Cadherin-positiven Vakuole umschlossen (Abbildung 21).
Abbildung 21: Verdichtung von E-Cadherin bei der Phagozytose
Gezeigt werden der phagozytische Prozess mit ausgeprägten Membran-Ausstülpungen um die hyperthermierte Zelle herum am Beginn der Phagozytose und zwei abgeschlossene Phagozytose-Ereignisse von Pankreaskarzinom-Zellen. Zur Un-terscheidung der ungefärbten vitalen und hyperthermierten Zellen, wurden die Zellker-ne vor der Fixierung dem Membran-impermeablen DNA-Farbstoff 7-AAD gefärbt. Mit-tels Immunfluoreszenz-Färbung wurde die Akkumulation von E-Cadherin an der Kon-taktstelle zur hyperthermierten Zelle und auf der phagozytischen Vakuole sichtbar ge-macht. Die Zellkerne wurden mit DAPI blau angefärbt. DIC: Differential-Interferenz-Kontrast; Maßstab: 10 µm.
Verdichtung von F-Aktin 3.6.2.
Die gerichtete Polymerisierung von F-Aktin und die Bildung von Membran-
Fortsätzen ermöglichen die Phagozytose und sind von der Aktivität von Protei-
nen der Ezrin-Radixin-Moesin-Familie abhängig. Bei der Aufnahme von hyper-
thermierten Zellen waren nur bei den phagozytischen Zellen, nicht bei den nek-
rotischen Zellen, Aktinfilamente vorhanden.
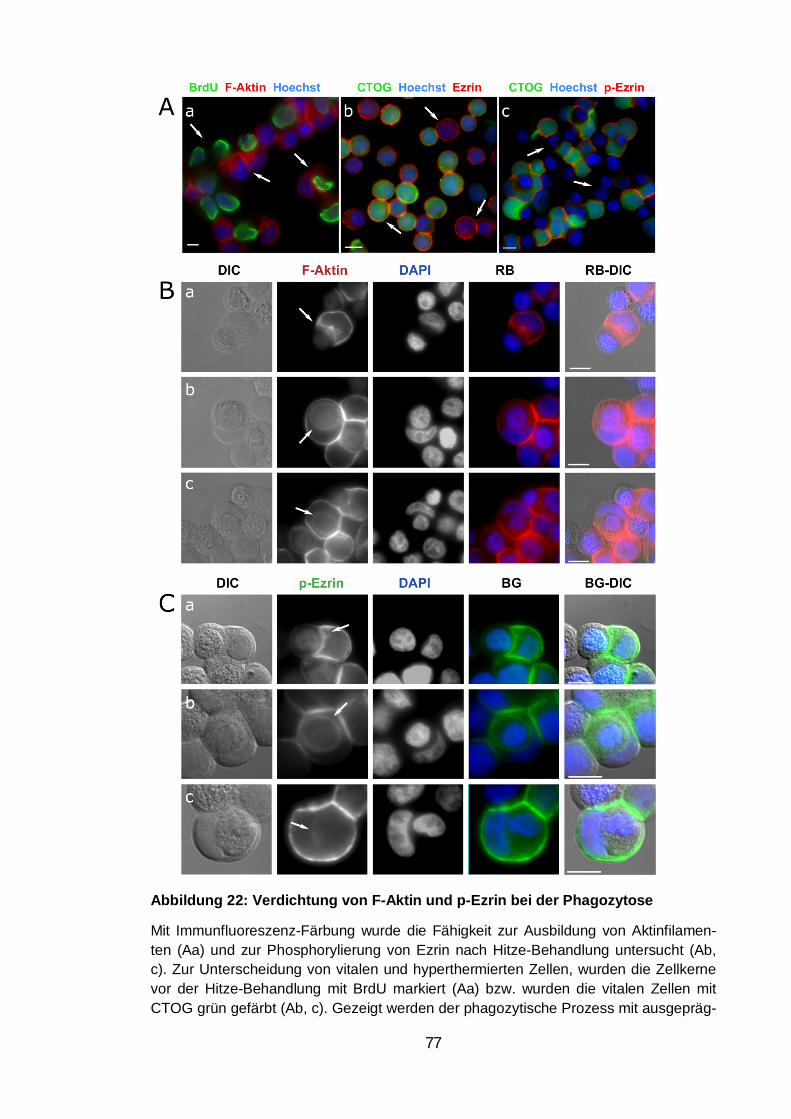
77
Abbildung 22: Verdichtung von F-Aktin und p-Ezrin bei der Phagozytose
Mit Immunfluoreszenz-Färbung wurde die Fähigkeit zur Ausbildung von Aktinfilamen-ten (Aa) und zur Phosphorylierung von Ezrin nach Hitze-Behandlung untersucht (Ab, c). Zur Unterscheidung von vitalen und hyperthermierten Zellen, wurden die Zellkerne vor der Hitze-Behandlung mit BrdU markiert (Aa) bzw. wurden die vitalen Zellen mit CTOG grün gefärbt (Ab, c). Gezeigt werden der phagozytische Prozess mit ausgepräg-

78
ten Membran-Ausstülpungen um die hyperthermierte Zelle herum am Beginn der Pha-gozytose und zwei abgeschlossene Phagozytose-Ereignisse von Pankreaskarzinom-Zellen. Mittels Immunfluoreszenz-Färbung wurde die Akkumulation von F-Aktin (Ba – c) und p-Ezrin (Ca – c) an der Kontaktstelle zur hyperthermierten Zelle und auf der phagozytischen Vakuole sichtbar gemacht. Das p-Ezrin Signal verringert sich auf der Vakuole in kurzer Zeit nach Phagozytose (Cc). Pfeile deuten auf das Vorhandensein bzw. das Fehlen der Proteine (A) und Protein-Akkumulationen (B, C). DIC: Differential-Interferenz-Kontrast; CTOG: Celltrace Oregon Green; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Durch Immunfluoreszenz-Färbung von Ezrin und p-Ezrin wurde erkannt, dass
hyperthermierte Zellen Ezrin in ähnlicher Menge exprimieren wie unbehandelte
Zellen, aber nicht in der Lage sind, es zu phosphorylieren. Das Fehlen von p-
Ezrin und F-Aktin deutet darauf hin, dass Hitze-behandelte Zellen ihre Zell-
membran und ihren Zellkörper nicht aktiv bewegen und verformen können und
stattdessen passiv phagozytiert werden (Abbildung 22A).
Nach der Formation von Zellmembranfortsätzen mit F-Aktin wurden nekrotische
Zellen in F-Aktin positive Vakuolen aufgenommen (Abbildung 22B). Außerdem
wurden verschiedene Stadien der Phagozytose aufgenommen, welche die Bil-
dung von p-Ezrin-positiven Membran-Ausstülpungen, die Aufnahme in p-Ezrin-
positive Vakuolen, gefolgt von einem Verlust von p-Ezrin von den Vakuolen zei-
gen (Abbildung 22C).
Akkumulation von LAMP1 und LC3 3.6.3.
Unmittelbar nach der Phagozytose wurde eine Ringbildung aus LAMP1-
positiven Vesikeln um die nekrotische Zelle herum sichtbar. Im Zytoplasma von
Hitze-behandelten Zellen waren insgesamt viel mehr LAMP1-positive Vesikel zu
erkennen als in den lebenden Zellen (Abbildung 23A).
Ebenso wurden viele LC3-positive Vesikel im Zytoplasma der Hitze-
behandelten Zellen gefunden. Dies könnte auf eine mögliche Induktion von Au-
tophagie hindeuten. In den phagozytierenden, vitalen Zellen waren die LC3-
Signale sehr viel schwächer und akkumulierten vor allem an der Kontaktstelle
zu nekrotischen Zellen. Des Weiteren konnte eine ringförmige Lokalisation von
LC-3 positiven Vesikeln, welche die aufgenommene Zelle umschließen, beo-
bachtet werden (Abbildung 23B).
Mittels Phasenkontrastmikroskopie wurden außerdem granuläre Strukturen in
den hyperthermierten Zellen sichtbar gemacht. Zusammen mit der großen
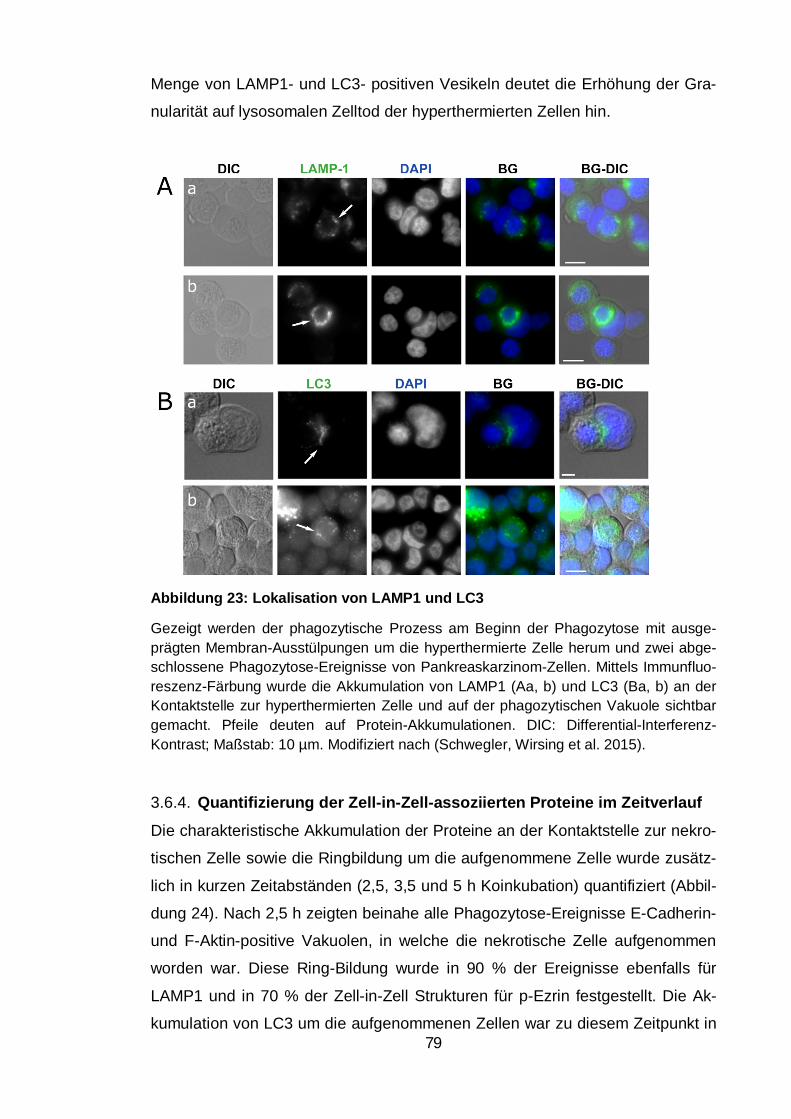
79
Menge von LAMP1- und LC3- positiven Vesikeln deutet die Erhöhung der Gra-
nularität auf lysosomalen Zelltod der hyperthermierten Zellen hin.
Abbildung 23: Lokalisation von LAMP1 und LC3
Gezeigt werden der phagozytische Prozess am Beginn der Phagozytose mit ausge-prägten Membran-Ausstülpungen um die hyperthermierte Zelle herum und zwei abge-schlossene Phagozytose-Ereignisse von Pankreaskarzinom-Zellen. Mittels Immunfluo-reszenz-Färbung wurde die Akkumulation von LAMP1 (Aa, b) und LC3 (Ba, b) an der Kontaktstelle zur hyperthermierten Zelle und auf der phagozytischen Vakuole sichtbar gemacht. Pfeile deuten auf Protein-Akkumulationen. DIC: Differential-Interferenz-Kontrast; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Quantifizierung der Zell-in-Zell-assoziierten Proteine im Zeitverlauf 3.6.4.
Die charakteristische Akkumulation der Proteine an der Kontaktstelle zur nekro-
tischen Zelle sowie die Ringbildung um die aufgenommene Zelle wurde zusätz-
lich in kurzen Zeitabständen (2,5, 3,5 und 5 h Koinkubation) quantifiziert (Abbil-
dung 24). Nach 2,5 h zeigten beinahe alle Phagozytose-Ereignisse E-Cadherin-
und F-Aktin-positive Vakuolen, in welche die nekrotische Zelle aufgenommen
worden war. Diese Ring-Bildung wurde in 90 % der Ereignisse ebenfalls für
LAMP1 und in 70 % der Zell-in-Zell Strukturen für p-Ezrin festgestellt. Die Ak-
kumulation von LC3 um die aufgenommenen Zellen war zu diesem Zeitpunkt in
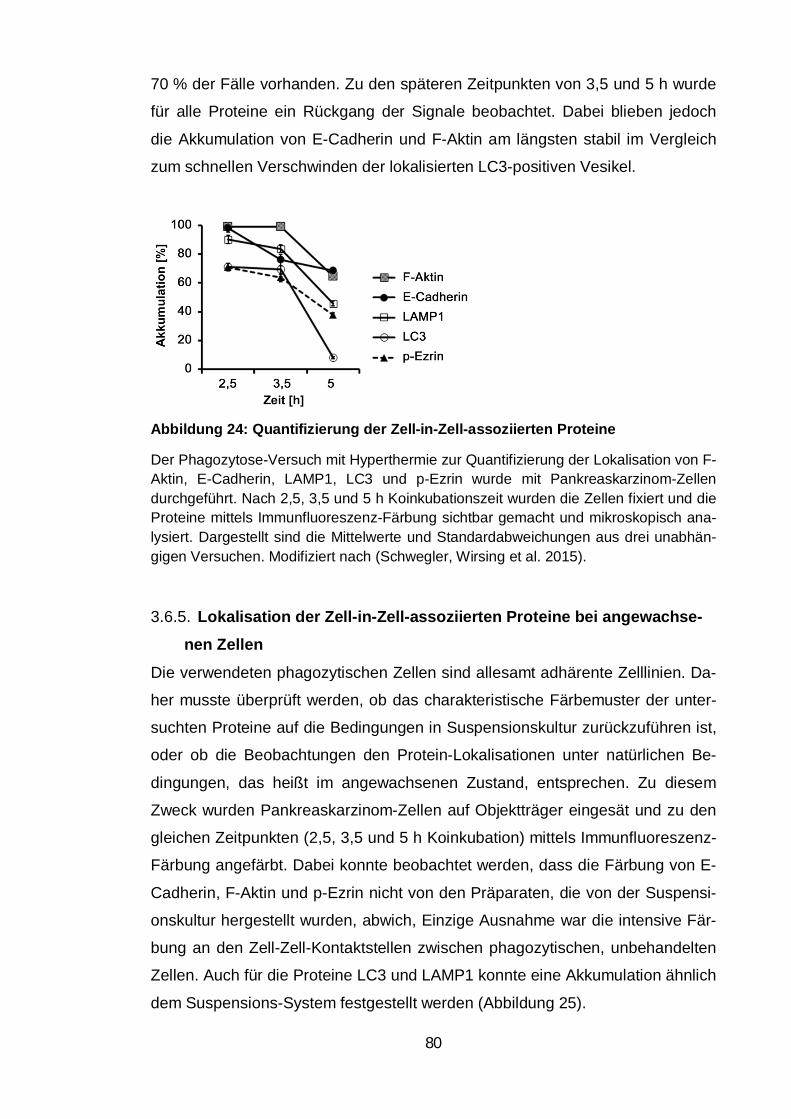
80
70 % der Fälle vorhanden. Zu den späteren Zeitpunkten von 3,5 und 5 h wurde
für alle Proteine ein Rückgang der Signale beobachtet. Dabei blieben jedoch
die Akkumulation von E-Cadherin und F-Aktin am längsten stabil im Vergleich
zum schnellen Verschwinden der lokalisierten LC3-positiven Vesikel.
Abbildung 24: Quantifizierung der Zell-in-Zell-assoziierten Proteine
Der Phagozytose-Versuch mit Hyperthermie zur Quantifizierung der Lokalisation von F-Aktin, E-Cadherin, LAMP1, LC3 und p-Ezrin wurde mit Pankreaskarzinom-Zellen durchgeführt. Nach 2,5, 3,5 und 5 h Koinkubationszeit wurden die Zellen fixiert und die Proteine mittels Immunfluoreszenz-Färbung sichtbar gemacht und mikroskopisch ana-lysiert. Dargestellt sind die Mittelwerte und Standardabweichungen aus drei unabhän-gigen Versuchen. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Lokalisation der Zell-in-Zell-assoziierten Proteine bei angewachse-3.6.5.
nen Zellen
Die verwendeten phagozytischen Zellen sind allesamt adhärente Zelllinien. Da-
her musste überprüft werden, ob das charakteristische Färbemuster der unter-
suchten Proteine auf die Bedingungen in Suspensionskultur zurückzuführen ist,
oder ob die Beobachtungen den Protein-Lokalisationen unter natürlichen Be-
dingungen, das heißt im angewachsenen Zustand, entsprechen. Zu diesem
Zweck wurden Pankreaskarzinom-Zellen auf Objektträger eingesät und zu den
gleichen Zeitpunkten (2,5, 3,5 und 5 h Koinkubation) mittels Immunfluoreszenz-
Färbung angefärbt. Dabei konnte beobachtet werden, dass die Färbung von E-
Cadherin, F-Aktin und p-Ezrin nicht von den Präparaten, die von der Suspensi-
onskultur hergestellt wurden, abwich, Einzige Ausnahme war die intensive Fär-
bung an den Zell-Zell-Kontaktstellen zwischen phagozytischen, unbehandelten
Zellen. Auch für die Proteine LC3 und LAMP1 konnte eine Akkumulation ähnlich
dem Suspensions-System festgestellt werden (Abbildung 25).

81
Abbildung 25: Lokalisation der Phagozytose-assoziierten Proteine bei ange-wachsenen Zell-in-Zell Strukturen
Gezeigt werden der phagozytische Prozess am Beginn der Phagozytose mit ausge-prägten Membran-Ausstülpungen um die hyperthermierte Zelle herum und abge-schlossene Phagozytose-Ereignisse von Pankreaskarzinom-Zellen. Mittels Immunfluo-reszenz-Färbung wurde die Akkumulation von E-Cadherin (A), p-Ezrin (B), LAMP1 (C), LC3 (D) und F-Aktin (E, F) an der Kontaktstelle zur hyperthermierten Zelle und auf der phagozytischen Vakuole sichtbar gemacht. Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Inhibition der molekularen Maschinerie der nicht-3.7.
professionellen Phagozyten Die Untersuchungen zur Phagozytose von nekrotischen Zellen wurden bis auf
wenige Ausnahmen (siehe 3.3.5 und 3.6.5) im Suspensions-System durchge-
führt, wobei gezeigt werden konnte, dass auch angewachsene Zellen zur Auf-
nahme fähig sind. Die Zelltodes-Form Entose findet statt, wenn sich Zellen von
der extrazellulären Matrix ablösen, sich also ebenfalls in Suspension befinden.
Um die Unterschiede zwischen Entose und nicht-professioneller Phagozytose
von nekrotischen Zellen anhand von molekularen Mechanismen beider Ereig-
nisse zu charakterisieren, wurden Versuche zur Inhibition der Phagozytose
durchgeführt.
Einfluss von Bestrahlung, Proteasen und Zytoskelett-Inhibitoren 3.7.1.
Die phagozytischen Zellen wurden vor der Koinkubation mit hyperthermierten
Zellen mit verschiedenen Noxen behandelt. Dazu zählten die Behandlung mit
hohen Dosen von Röntgenstrahlung, Proteasen, Zytoskelett- und GTPase-
Inhibitoren (Abbildung 26).
Die Bestrahlung mit 50 Gy verringerte die Phagozytose-Raten nicht. Die Zellen
wurden solange mit proteolytischen Enzymen (Accutase oder Trypsin) behan-
delt bis die sich die Zellen ohne zusätzliche mechanische Reize (Abklopfen vom
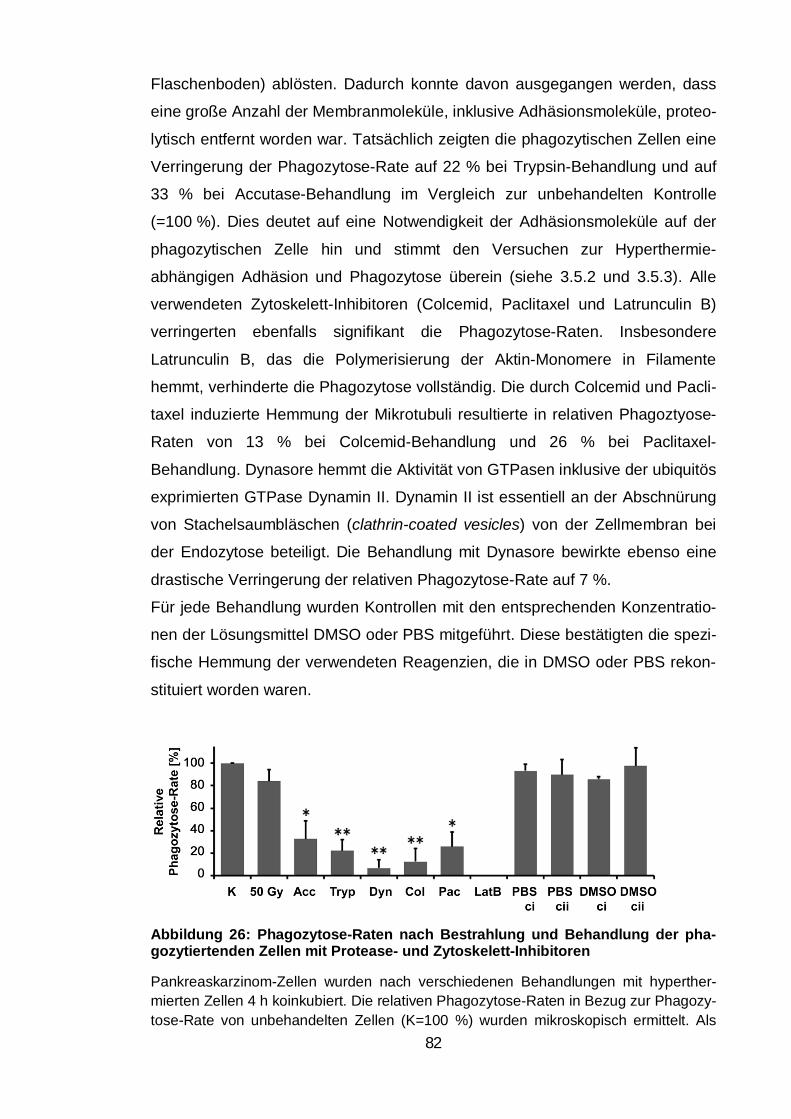
82
Flaschenboden) ablösten. Dadurch konnte davon ausgegangen werden, dass
eine große Anzahl der Membranmoleküle, inklusive Adhäsionsmoleküle, proteo-
lytisch entfernt worden war. Tatsächlich zeigten die phagozytischen Zellen eine
Verringerung der Phagozytose-Rate auf 22 % bei Trypsin-Behandlung und auf
33 % bei Accutase-Behandlung im Vergleich zur unbehandelten Kontrolle
(=100 %). Dies deutet auf eine Notwendigkeit der Adhäsionsmoleküle auf der
phagozytischen Zelle hin und stimmt den Versuchen zur Hyperthermie-
abhängigen Adhäsion und Phagozytose überein (siehe 3.5.2 und 3.5.3). Alle
verwendeten Zytoskelett-Inhibitoren (Colcemid, Paclitaxel und Latrunculin B)
verringerten ebenfalls signifikant die Phagozytose-Raten. Insbesondere
Latrunculin B, das die Polymerisierung der Aktin-Monomere in Filamente
hemmt, verhinderte die Phagozytose vollständig. Die durch Colcemid und Pacli-
taxel induzierte Hemmung der Mikrotubuli resultierte in relativen Phagoztyose-
Raten von 13 % bei Colcemid-Behandlung und 26 % bei Paclitaxel-
Behandlung. Dynasore hemmt die Aktivität von GTPasen inklusive der ubiquitös
exprimierten GTPase Dynamin II. Dynamin II ist essentiell an der Abschnürung
von Stachelsaumbläschen (clathrin-coated vesicles) von der Zellmembran bei
der Endozytose beteiligt. Die Behandlung mit Dynasore bewirkte ebenso eine
drastische Verringerung der relativen Phagozytose-Rate auf 7 %.
Für jede Behandlung wurden Kontrollen mit den entsprechenden Konzentratio-
nen der Lösungsmittel DMSO oder PBS mitgeführt. Diese bestätigten die spezi-
fische Hemmung der verwendeten Reagenzien, die in DMSO oder PBS rekon-
stituiert worden waren.
Abbildung 26: Phagozytose-Raten nach Bestrahlung und Behandlung der pha-gozytiertenden Zellen mit Protease- und Zytoskelett-Inhibitoren
Pankreaskarzinom-Zellen wurden nach verschiedenen Behandlungen mit hyperther-mierten Zellen 4 h koinkubiert. Die relativen Phagozytose-Raten in Bezug zur Phagozy-tose-Rate von unbehandelten Zellen (K=100 %) wurden mikroskopisch ermittelt. Als

83
weitere Kontrollen für die verwendeten Inhibitoren (rekonstituiert in PBS, DMSO oder Medium) wurden entsprechende Konzentrationen (ci und cii) von PBS und DMSO mit-geführt. Dargestellt sind die Mittelwerte und Standardabweichungen von drei unabhän-gigen Experimenten. K: Kontrolle; Gy: Gray; Acc: Accutase; Tryp: Trypsin; Dyn: Dynasore; Col: Colcemid; Pac: Paclitaxel; LatB: Latrunculin B; *p < 0,05, **p ≤ 0,01; Modifiziert nach (Schwegler, Wirsing et al. 2015).
Blockade des E- und P-Cadherin vermittelten Zellkontakts 3.7.2.
Bei der beschriebenen Zelltodes-Form Entose wurde von einer Inhibition der
Entose nach Blockade von E- und P-Cadherin bei MCF-10A nicht-tumorigenen
Brustepithelzellen berichtet (Overholtzer, Mailleux et al. 2007). Nachdem mittels
Immunfluoreszenz-Färbung eine Akkumulation von E-Cadherin an der
Kontaktstelle zur nekrotischen Zelle bei der Phagozytose festgestellt worden
war (siehe 3.6.1), sollte überprüft werden, ob eine Blockade von E-Cadherin zu
einer Reduktion der Phagozytose-Rate führt. Da für Pankreaskarzinom-Zellen
zusätzlich die Expression von P-Cadherin gezeigt wurde (Imai, Hirata et al.
2008), wurde gleichzeitig dieses Transmembranprotein gehemmt. Zunächst
wurde die Expression von E- und P-Cadherin von vitalen und hyperthermierten
Zellen und die Lokalisation in Phagozytose-Ereignissen mittels
Immunfluoreszenz-Färbung untersucht (Abbildungen 27, 28).
E-Cadherin wurde auf den vitalen Zellen stark exprimiert, konnte nach Hitze-
Behandlung allerdings nicht mehr nachgewiesen werden (Abbildung 27A). In
den Zell-in-Zell Strukturen nach Aufnahme von nekrotischen Zellen wurde
wiederum eine Verdichtung des Signals um die aufgenommene Zelle herum,
ausgemacht (Abbildung 27B). Zur Überprüfung der Spezifität des verwendeten
Antikörpers wurde außerdem eine Isotypkontrolle mitgeführt (Abbildung 27C).
P-Cadherin wurde nur sehr schwach angefärbt, was auf eine niedrige
Expression hindeutet. Die Behandlung mit Hyperthermie bewirkte auch hier
einen Verlust von P-Cadherin. Aufgrund des schwachen P-Cadherin Signals
und der Signalüberlagerung von CTFR ist dieser Verlust nur schwer erkennbar
(Abbildung 28A).
Nach Phagozytose von hyperthermierten Zellen zeigen die Zell-in-Zell
Strukturen ebenfalls eine Akkumulation von P-Cadherin um die aufgenommene
Zelle herum (Abbildung 28B). Die mitgeführte Isotypkontrolle bestätigte die
Spezifität des verwendeten Antikörpers (Abbildung 28C).
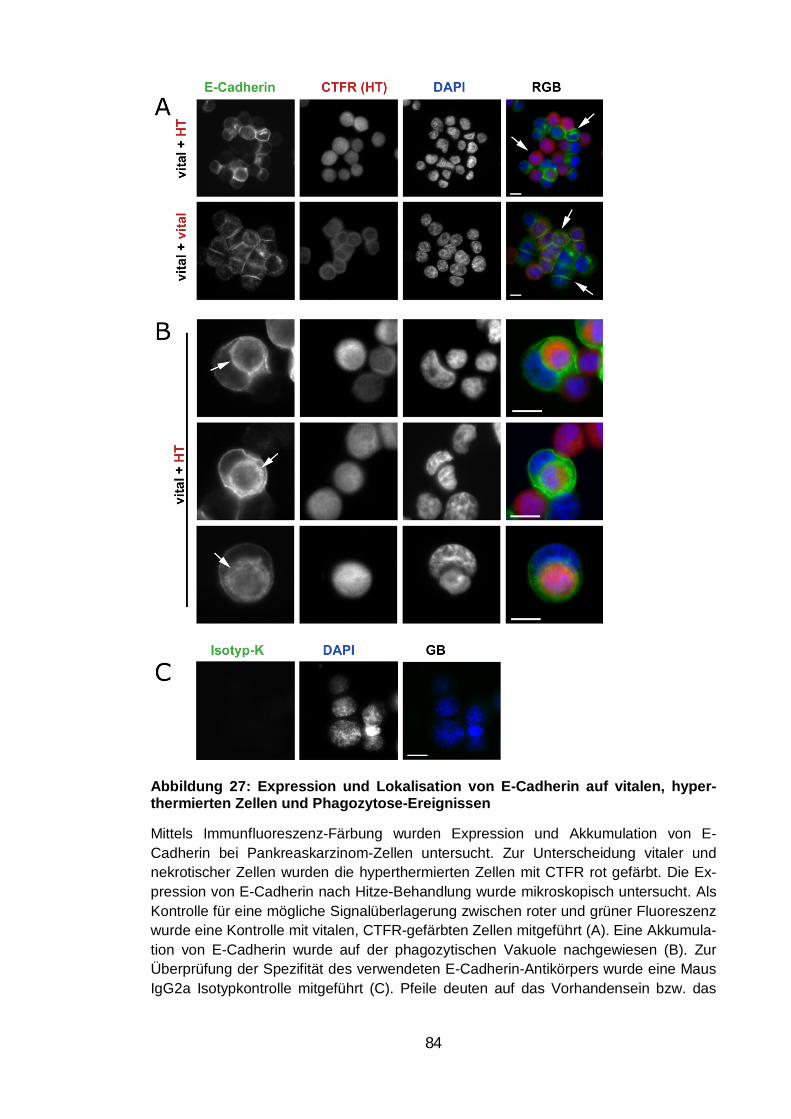
84
Abbildung 27: Expression und Lokalisation von E-Cadherin auf vitalen, hyper-thermierten Zellen und Phagozytose-Ereignissen
Mittels Immunfluoreszenz-Färbung wurden Expression und Akkumulation von E-Cadherin bei Pankreaskarzinom-Zellen untersucht. Zur Unterscheidung vitaler und nekrotischer Zellen wurden die hyperthermierten Zellen mit CTFR rot gefärbt. Die Ex-pression von E-Cadherin nach Hitze-Behandlung wurde mikroskopisch untersucht. Als Kontrolle für eine mögliche Signalüberlagerung zwischen roter und grüner Fluoreszenz wurde eine Kontrolle mit vitalen, CTFR-gefärbten Zellen mitgeführt (A). Eine Akkumula-tion von E-Cadherin wurde auf der phagozytischen Vakuole nachgewiesen (B). Zur Überprüfung der Spezifität des verwendeten E-Cadherin-Antikörpers wurde eine Maus IgG2a Isotypkontrolle mitgeführt (C). Pfeile deuten auf das Vorhandensein bzw. das
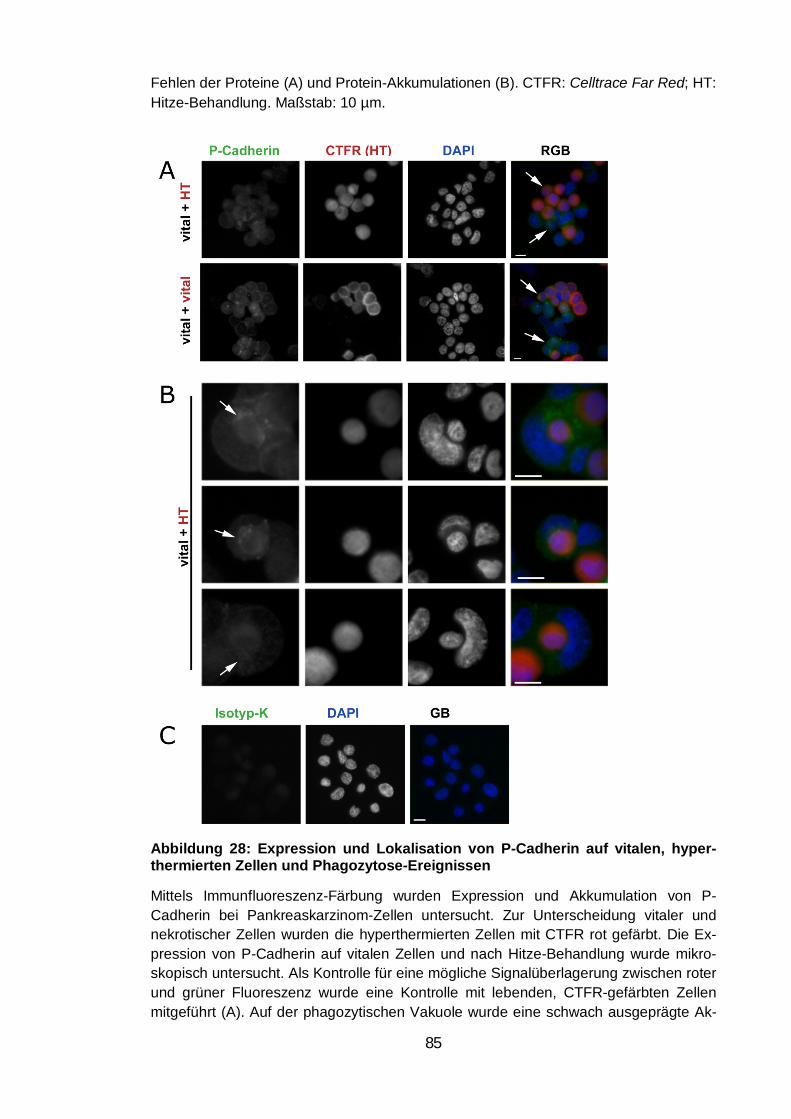
85
Fehlen der Proteine (A) und Protein-Akkumulationen (B). CTFR: Celltrace Far Red; HT: Hitze-Behandlung. Maßstab: 10 µm.
Abbildung 28: Expression und Lokalisation von P-Cadherin auf vitalen, hyper-thermierten Zellen und Phagozytose-Ereignissen
Mittels Immunfluoreszenz-Färbung wurden Expression und Akkumulation von P-Cadherin bei Pankreaskarzinom-Zellen untersucht. Zur Unterscheidung vitaler und nekrotischer Zellen wurden die hyperthermierten Zellen mit CTFR rot gefärbt. Die Ex-pression von P-Cadherin auf vitalen Zellen und nach Hitze-Behandlung wurde mikro-skopisch untersucht. Als Kontrolle für eine mögliche Signalüberlagerung zwischen roter und grüner Fluoreszenz wurde eine Kontrolle mit lebenden, CTFR-gefärbten Zellen mitgeführt (A). Auf der phagozytischen Vakuole wurde eine schwach ausgeprägte Ak-

86
kumulation von P-Cadherin nachgewiesen (B). Zur Überprüfung der Spezifität des ver-wendeten P-Cadherin-Antikörpers wurde eine Kaninchen IgG Isotypkontrolle mitgeführt (C). Pfeile deuten auf das Vorhandensein bzw. das Fehlen der Proteine (A) und Pro-tein-Akkumulationen (B). CTFR: Celltrace Far Red; HT: Hitze-Behandlung. Maßstab: 10 µm.
Der Verlust von E- und P-Cadherin durch Hyperthermie ist insofern brisant, als
dass bisher lediglich homotypische Interaktionen von Cadherinen bzw. hetero-
typische Interaktionen zwischen verschiedenen Cadherin-Typen bekannt sind
(Takeichi 1991, Geiger and Ayalon 1992, Omelchenko, Fetisova et al. 2001,
Niessen and Gumbiner 2002). Demzufolge müssten auf phagozytischer und
nekrotischer Zelle Cadherine vorhanden sein, damit eine Interaktion dieser Pro-
teine bei der Phagozytose von Bedeutung sein könnte. Es wurde aber auch
über mögliche heterotypische Interaktionen spekuliert, wie es für die Bindung
zwischen Cadherinen und Integrinen gezeigt werden konnte (Whittard, Craig et
al. 2002). Demnach wird eine Cadherin-vermittelte Bindung an einen Liganden
auf der hyperthermierten Zelle, trotz deren Cadherin-Verlust, nicht ausge-
schlossen.
Um eine mögliche Funktion dieser Cadherine bei der Phagozytose zu testen
und damit eine Übereinstimmung zur Entose zu prüfen, wurden funktionale,
blockierende Antikörper gegen E- und P-Cadherin verwendet. Diese wurden
während der Koinkubation von 4 h nach Hyperthermie-Behandlung in den
gleichen Konzentrationen, die für die Entose publiziert wurden, mitgeführt. Als
Kontrolle wurden außerdem Proben ohne Hitze-Behandlung mitgeführt. Die
durchflusszytometrische Messung der Adhäsion zeigte keine Reduktion der
Bindung von vitalen an nekrotische Zellen durch die verwendeten Antikörper.
Bei der Quantifizierung der Phagozytose-Raten wurde eine signifikante
Verringerung durch die blockierenden Antikörper festgestellt. Die mitgeführte
Isotypkontrolle zeigte jedoch, dass der inhibierende Effekt unspezifisch war, da
eine ähnlich starke Verringerung der Phagozytose-Rate auch bei diesen Proben
zu beobachten war (Abbildung 29). Die spezifische Hemmung des E- und P-
Cadherin-vermittelten Zellkontakts konnte demnach nicht erzielt werden,
sodass keine Aussage über eine Beteiligung von Cadherin-Interaktionen bei der
Phagozytose gemacht werden kann. Möglicherweise liegt die Ursache in der
unspezifischen Hemmung bei der eingesetzten Konzentration der Antikörper
und Isotypkontrollen. Diese entsprach der Konzentration, die für den Inhibitions-
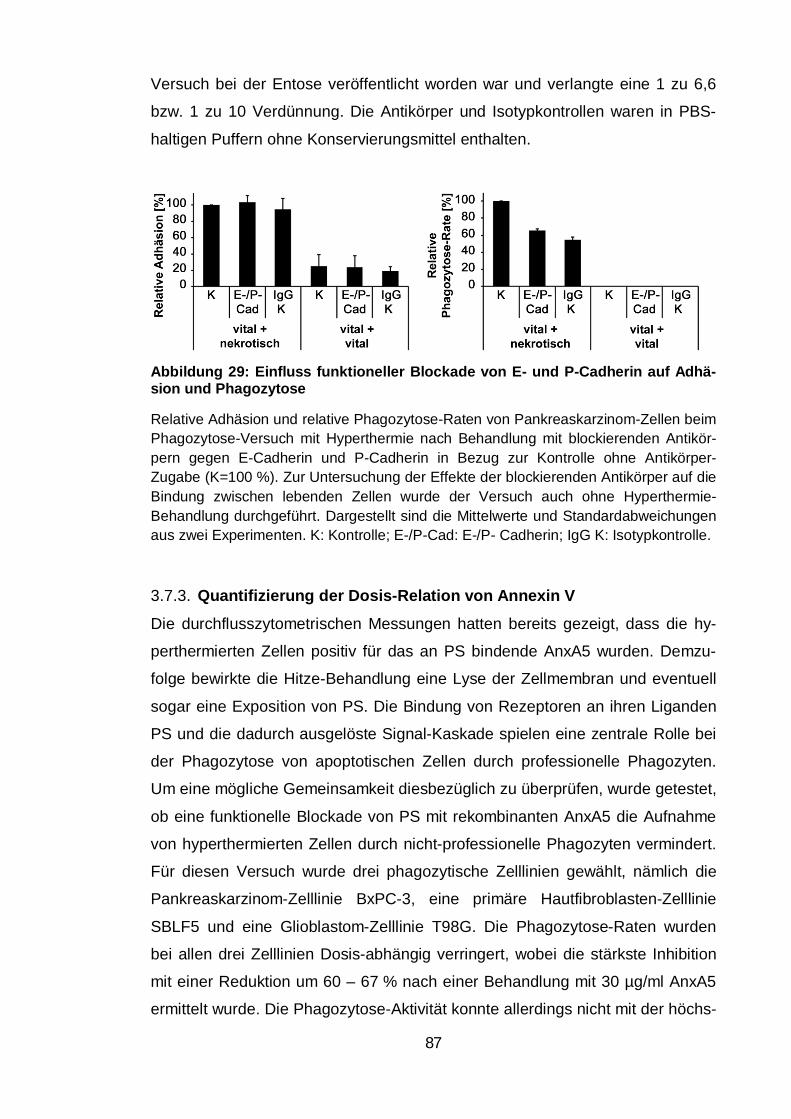
87
Versuch bei der Entose veröffentlicht worden war und verlangte eine 1 zu 6,6
bzw. 1 zu 10 Verdünnung. Die Antikörper und Isotypkontrollen waren in PBS-
haltigen Puffern ohne Konservierungsmittel enthalten.
Abbildung 29: Einfluss funktioneller Blockade von E- und P-Cadherin auf Adhä-sion und Phagozytose
Relative Adhäsion und relative Phagozytose-Raten von Pankreaskarzinom-Zellen beim Phagozytose-Versuch mit Hyperthermie nach Behandlung mit blockierenden Antikör-pern gegen E-Cadherin und P-Cadherin in Bezug zur Kontrolle ohne Antikörper-Zugabe (K=100 %). Zur Untersuchung der Effekte der blockierenden Antikörper auf die Bindung zwischen lebenden Zellen wurde der Versuch auch ohne Hyperthermie-Behandlung durchgeführt. Dargestellt sind die Mittelwerte und Standardabweichungen aus zwei Experimenten. K: Kontrolle; E-/P-Cad: E-/P- Cadherin; IgG K: Isotypkontrolle.
Quantifizierung der Dosis-Relation von Annexin V 3.7.3.
Die durchflusszytometrischen Messungen hatten bereits gezeigt, dass die hy-
perthermierten Zellen positiv für das an PS bindende AnxA5 wurden. Demzu-
folge bewirkte die Hitze-Behandlung eine Lyse der Zellmembran und eventuell
sogar eine Exposition von PS. Die Bindung von Rezeptoren an ihren Liganden
PS und die dadurch ausgelöste Signal-Kaskade spielen eine zentrale Rolle bei
der Phagozytose von apoptotischen Zellen durch professionelle Phagozyten.
Um eine mögliche Gemeinsamkeit diesbezüglich zu überprüfen, wurde getestet,
ob eine funktionelle Blockade von PS mit rekombinanten AnxA5 die Aufnahme
von hyperthermierten Zellen durch nicht-professionelle Phagozyten vermindert.
Für diesen Versuch wurde drei phagozytische Zelllinien gewählt, nämlich die
Pankreaskarzinom-Zelllinie BxPC-3, eine primäre Hautfibroblasten-Zelllinie
SBLF5 und eine Glioblastom-Zelllinie T98G. Die Phagozytose-Raten wurden
bei allen drei Zelllinien Dosis-abhängig verringert, wobei die stärkste Inhibition
mit einer Reduktion um 60 – 67 % nach einer Behandlung mit 30 µg/ml AnxA5
ermittelt wurde. Die Phagozytose-Aktivität konnte allerdings nicht mit der höchs-
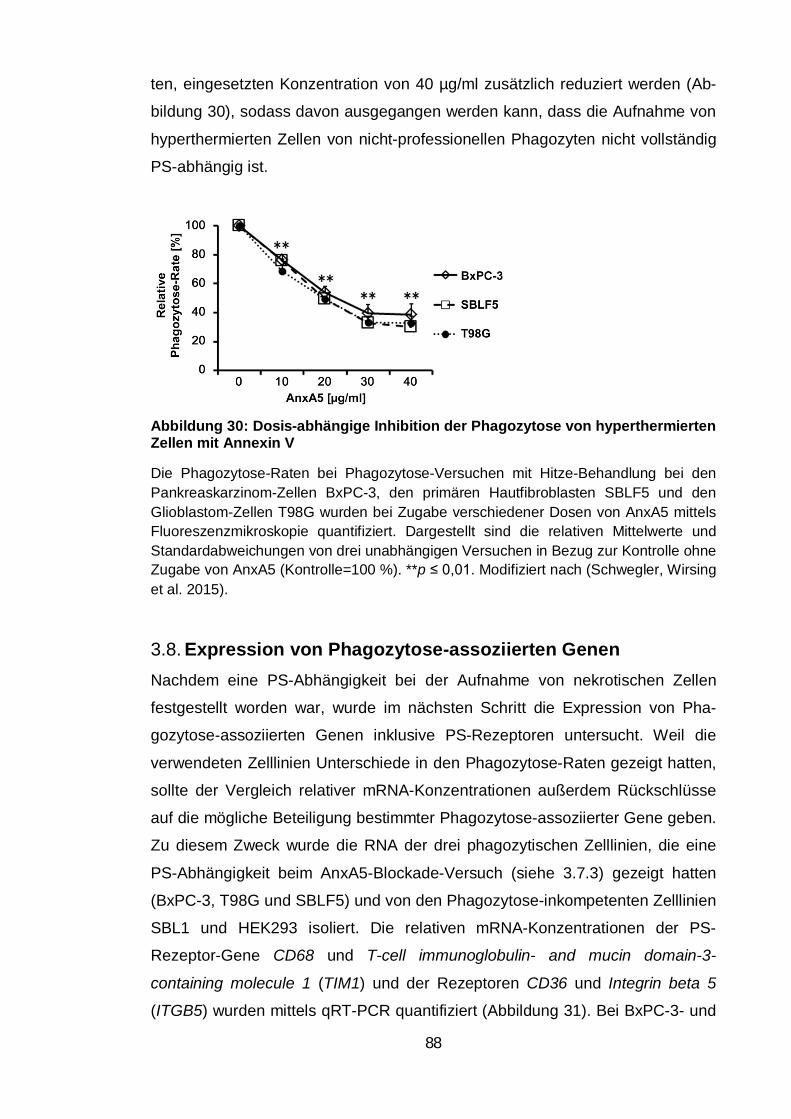
88
ten, eingesetzten Konzentration von 40 µg/ml zusätzlich reduziert werden (Ab-
bildung 30), sodass davon ausgegangen werden kann, dass die Aufnahme von
hyperthermierten Zellen von nicht-professionellen Phagozyten nicht vollständig
PS-abhängig ist.
Abbildung 30: Dosis-abhängige Inhibition der Phagozytose von hyperthermierten Zellen mit Annexin V
Die Phagozytose-Raten bei Phagozytose-Versuchen mit Hitze-Behandlung bei den Pankreaskarzinom-Zellen BxPC-3, den primären Hautfibroblasten SBLF5 und den Glioblastom-Zellen T98G wurden bei Zugabe verschiedener Dosen von AnxA5 mittels Fluoreszenzmikroskopie quantifiziert. Dargestellt sind die relativen Mittelwerte und Standardabweichungen von drei unabhängigen Versuchen in Bezug zur Kontrolle ohne Zugabe von AnxA5 (Kontrolle=100 %). **p ≤ 0,01. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Expression von Phagozytose-assoziierten Genen 3.8.Nachdem eine PS-Abhängigkeit bei der Aufnahme von nekrotischen Zellen
festgestellt worden war, wurde im nächsten Schritt die Expression von Pha-
gozytose-assoziierten Genen inklusive PS-Rezeptoren untersucht. Weil die
verwendeten Zelllinien Unterschiede in den Phagozytose-Raten gezeigt hatten,
sollte der Vergleich relativer mRNA-Konzentrationen außerdem Rückschlüsse
auf die mögliche Beteiligung bestimmter Phagozytose-assoziierter Gene geben.
Zu diesem Zweck wurde die RNA der drei phagozytischen Zelllinien, die eine
PS-Abhängigkeit beim AnxA5-Blockade-Versuch (siehe 3.7.3) gezeigt hatten
(BxPC-3, T98G und SBLF5) und von den Phagozytose-inkompetenten Zelllinien
SBL1 und HEK293 isoliert. Die relativen mRNA-Konzentrationen der PS-
Rezeptor-Gene CD68 und T-cell immunoglobulin- and mucin domain-3-
containing molecule 1 (TIM1) und der Rezeptoren CD36 und Integrin beta 5
(ITGB5) wurden mittels qRT-PCR quantifiziert (Abbildung 31). Bei BxPC-3- und
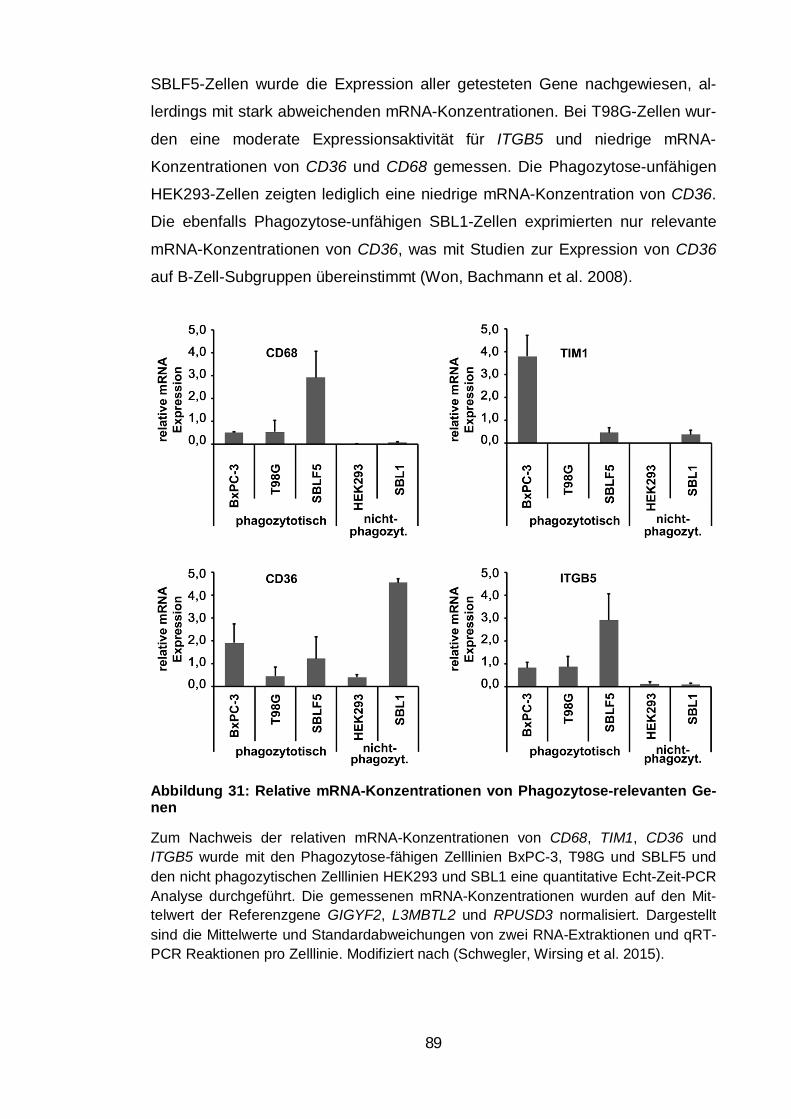
89
SBLF5-Zellen wurde die Expression aller getesteten Gene nachgewiesen, al-
lerdings mit stark abweichenden mRNA-Konzentrationen. Bei T98G-Zellen wur-
den eine moderate Expressionsaktivität für ITGB5 und niedrige mRNA-
Konzentrationen von CD36 und CD68 gemessen. Die Phagozytose-unfähigen
HEK293-Zellen zeigten lediglich eine niedrige mRNA-Konzentration von CD36.
Die ebenfalls Phagozytose-unfähigen SBL1-Zellen exprimierten nur relevante
mRNA-Konzentrationen von CD36, was mit Studien zur Expression von CD36
auf B-Zell-Subgruppen übereinstimmt (Won, Bachmann et al. 2008).
Abbildung 31: Relative mRNA-Konzentrationen von Phagozytose-relevanten Ge-nen
Zum Nachweis der relativen mRNA-Konzentrationen von CD68, TIM1, CD36 und ITGB5 wurde mit den Phagozytose-fähigen Zelllinien BxPC-3, T98G und SBLF5 und den nicht phagozytischen Zelllinien HEK293 und SBL1 eine quantitative Echt-Zeit-PCR Analyse durchgeführt. Die gemessenen mRNA-Konzentrationen wurden auf den Mit-telwert der Referenzgene GIGYF2, L3MBTL2 und RPUSD3 normalisiert. Dargestellt sind die Mittelwerte und Standardabweichungen von zwei RNA-Extraktionen und qRT-PCR Reaktionen pro Zelllinie. Modifiziert nach (Schwegler, Wirsing et al. 2015).

90
Degradation der inkorporierten Zelle 3.9.Zur Beobachtung des Zelltod-Verlaufs nach Phagozytose der Hitze-
behandelten Zelle wurde die Koinkubation in der Suspensions-Kulturplatte auf
bei zu drei Tage verlängert. Zusätzlich zur Präparation nach 4 h, wurden au-
ßerdem nach 24, 48 und 72 h mikroskopische Präparate angefertigt. Dabei soll-
te nicht nur die homotypische Phagozytose sondern auch die heterotypische
Interaktion untersucht werden. Zum einen wurden hyperthermierte Jurkat T-
Zellen mit lebenden, CTOG-gefärbten BxPC-3 Pankreaskarzinom-Zellen inku-
biert. Nach Zytozentrifugation und Fixierung wurden die T-Zellen mittels Immun-
fluoreszenz-Färbung durch Markierung des T-Zell-Antigens CD2 sichtbar ge-
macht. Die Zellkerne wurden mit dem DNA-Farbstoff DAPI gefärbt. Nach 4 h
wurden die typischen phagozytischen Strukturen, welche einen sichelförmigen
Zellkern der phagozytischen Zelle und einen runden Nukleus der aufgenomme-
nen Zelle beinhalten, ausgemacht. Nach 24 h erschien der Zellkern des Pha-
gozyten wieder seine ursprüngliche Form nahezu vollständig zurückerlangt zu
haben, wohingegen der Nukleus der internalisierten Zelle nicht mehr vorhanden
bzw. mit DAPI anzufärben war, was auf einen Verlust der DNA-Integrität hin-
deutet.
Die CD2-positiven phagozytischen Vakuolen blieben bis zu 72 h erhalten, wobei
in manchen Zell-in-Zell Strukturen auch kleine CD2-positive Vesikel im Zyto-
plasma des Phagozyten beobachtet werden konnten (Abbildung 32A).
Der Verlauf von homotypischen Phagozytose-Ereignissen wurde anhand von
hyperthermierten Pankreaskarzinom-Zellen, welche mit Histon H2B-RFP trans-
fiziert worden waren bzw. das Nukleosid EdU inkorporiert hatten, studiert. Diese
Markierung von Histonen bzw. der DNA ermöglichte die Beantwortung der Fra-
ge, ob Kernmaterial der internalisierten Zelle in zytoplasmatischen Vesikeln ab-
gebaut wird oder auch in den phagozytischen Nukleus eingebaut werden könn-
te. Zusätzlich wurden die mikroskopischen Präparate mit DAPI angefärbt,
wodurch die vorherige Beobachtung über den Verlust der DNA-Integrität der
phagozytierten Zelle, innerhalb von 24 h, bestätigt wurde. Des Weiteren wurde
eine Verteilung des H2B-RFP- und EdU-markierten DNA-Materials in der ge-
samten phagozytischen Vakuole nachgewiesen. Dies deutet auf eine Lyse der
Kernmembran und demzufolge den unumkehrbaren Eintritt von Zelltod in der
aufgenommenen Zelle hin. In Übereinstimmung mit dem Verlauf nach heteroty-

91
pischer Phagozytose, wurden phagozytische Strukturen mit einer stabilen Va-
kuole sowie mehreren, kleinen Vesikeln, welche die Bestandteile der hyper-
thermierten Zelle beinhalten, nachgewiesen. Ein Transfer von markiertem DNA-
Material in den Zellkern des Phagozyten wurde zu keinem Zeitpunkt festgestellt
(Abbildung 32B, 33).
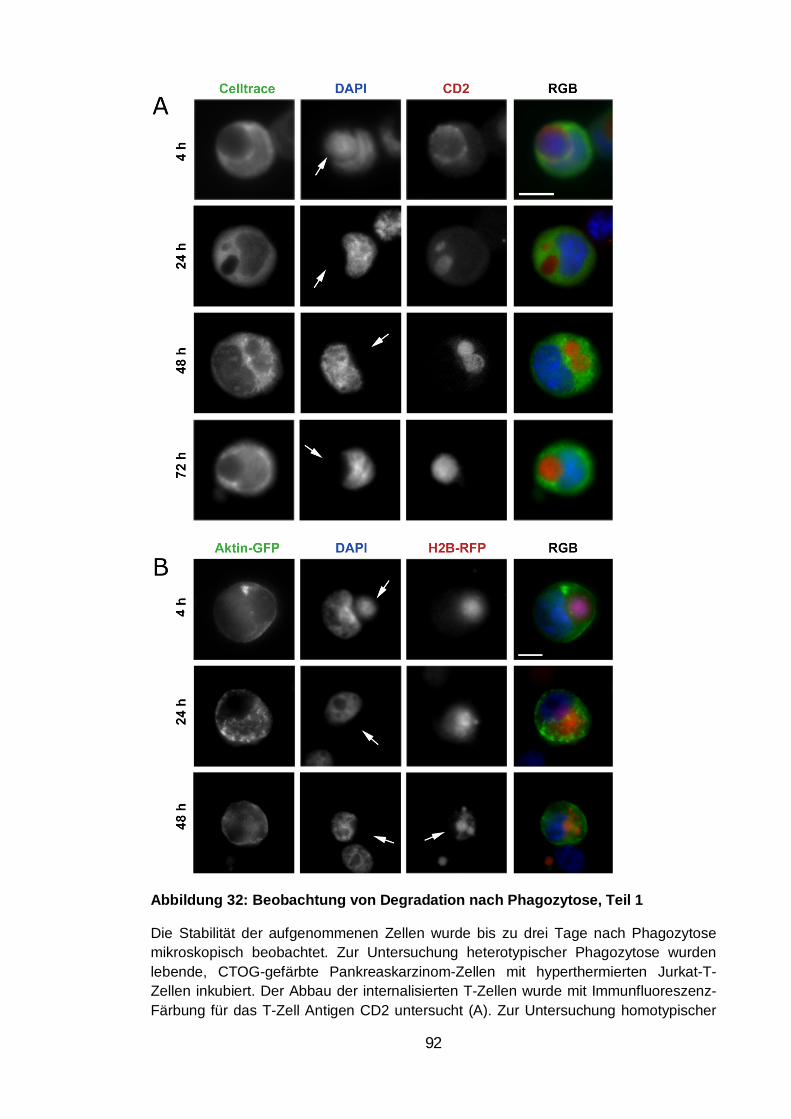
92
Abbildung 32: Beobachtung von Degradation nach Phagozytose, Teil 1
Die Stabilität der aufgenommenen Zellen wurde bis zu drei Tage nach Phagozytose mikroskopisch beobachtet. Zur Untersuchung heterotypischer Phagozytose wurden lebende, CTOG-gefärbte Pankreaskarzinom-Zellen mit hyperthermierten Jurkat-T-Zellen inkubiert. Der Abbau der internalisierten T-Zellen wurde mit Immunfluoreszenz-Färbung für das T-Zell Antigen CD2 untersucht (A). Zur Untersuchung homotypischer
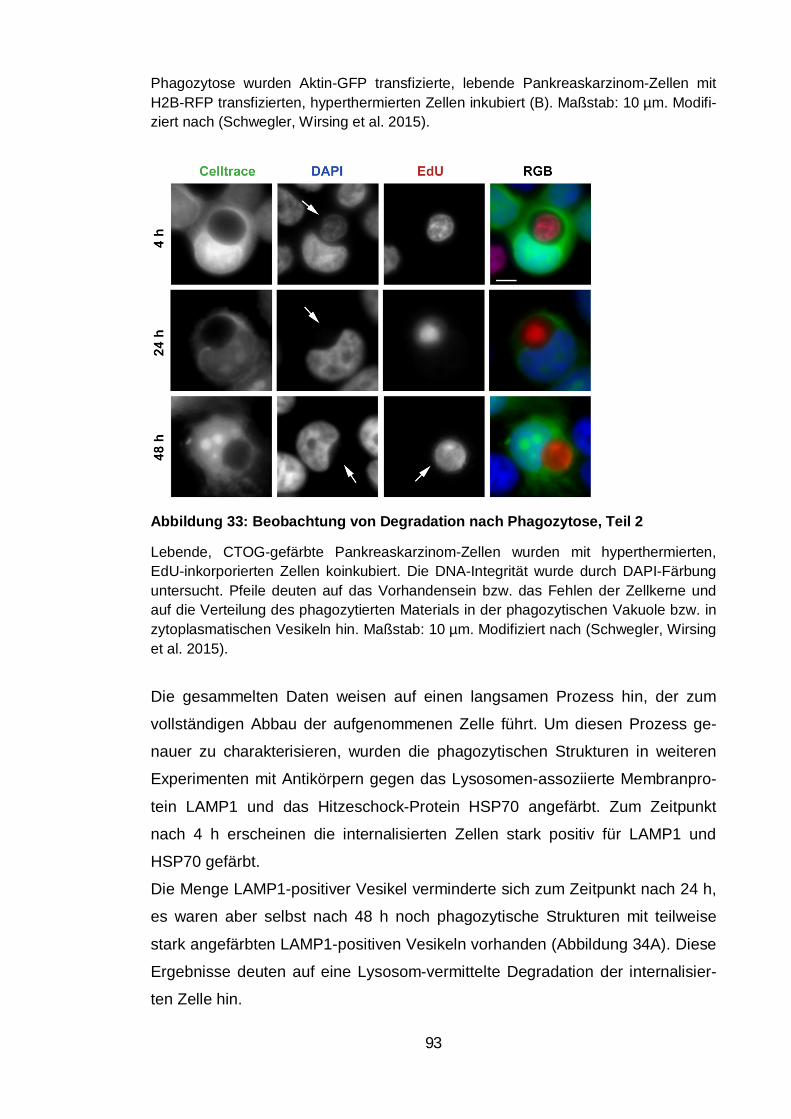
93
Phagozytose wurden Aktin-GFP transfizierte, lebende Pankreaskarzinom-Zellen mit H2B-RFP transfizierten, hyperthermierten Zellen inkubiert (B). Maßstab: 10 µm. Modifi-ziert nach (Schwegler, Wirsing et al. 2015).
Abbildung 33: Beobachtung von Degradation nach Phagozytose, Teil 2
Lebende, CTOG-gefärbte Pankreaskarzinom-Zellen wurden mit hyperthermierten, EdU-inkorporierten Zellen koinkubiert. Die DNA-Integrität wurde durch DAPI-Färbung untersucht. Pfeile deuten auf das Vorhandensein bzw. das Fehlen der Zellkerne und auf die Verteilung des phagozytierten Materials in der phagozytischen Vakuole bzw. in zytoplasmatischen Vesikeln hin. Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Die gesammelten Daten weisen auf einen langsamen Prozess hin, der zum
vollständigen Abbau der aufgenommenen Zelle führt. Um diesen Prozess ge-
nauer zu charakterisieren, wurden die phagozytischen Strukturen in weiteren
Experimenten mit Antikörpern gegen das Lysosomen-assoziierte Membranpro-
tein LAMP1 und das Hitzeschock-Protein HSP70 angefärbt. Zum Zeitpunkt
nach 4 h erscheinen die internalisierten Zellen stark positiv für LAMP1 und
HSP70 gefärbt.
Die Menge LAMP1-positiver Vesikel verminderte sich zum Zeitpunkt nach 24 h,
es waren aber selbst nach 48 h noch phagozytische Strukturen mit teilweise
stark angefärbten LAMP1-positiven Vesikeln vorhanden (Abbildung 34A). Diese
Ergebnisse deuten auf eine Lysosom-vermittelte Degradation der internalisier-
ten Zelle hin.
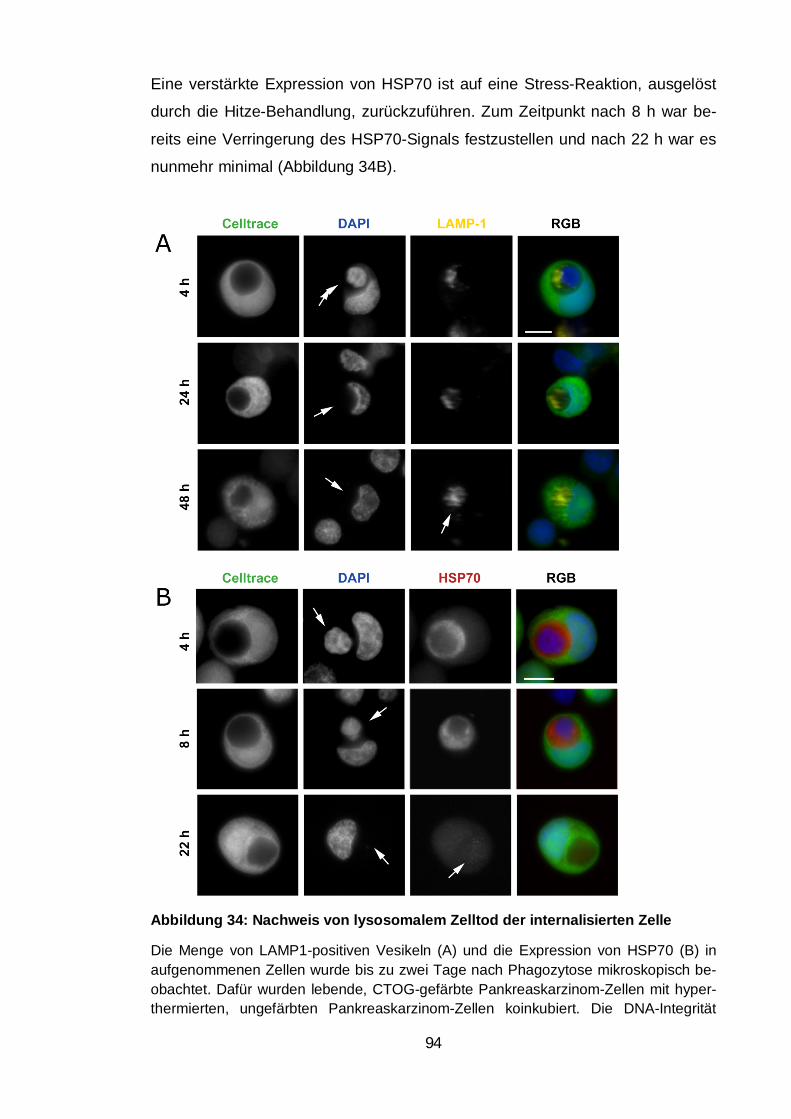
94
Eine verstärkte Expression von HSP70 ist auf eine Stress-Reaktion, ausgelöst
durch die Hitze-Behandlung, zurückzuführen. Zum Zeitpunkt nach 8 h war be-
reits eine Verringerung des HSP70-Signals festzustellen und nach 22 h war es
nunmehr minimal (Abbildung 34B).
Abbildung 34: Nachweis von lysosomalem Zelltod der internalisierten Zelle
Die Menge von LAMP1-positiven Vesikeln (A) und die Expression von HSP70 (B) in aufgenommenen Zellen wurde bis zu zwei Tage nach Phagozytose mikroskopisch be-obachtet. Dafür wurden lebende, CTOG-gefärbte Pankreaskarzinom-Zellen mit hyper-thermierten, ungefärbten Pankreaskarzinom-Zellen koinkubiert. Die DNA-Integrität

95
wurde durch DAPI-Färbung untersucht. Pfeile deuten auf das Vorhandensein bzw. das Fehlen der Zellkerne und auf die Verteilung des phagozytierten Materials in der pha-gozytischen Vakuole hin. CTOG: Celltrace Oregon Green; Maßstab: 10 µm. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Zell-in-Zell Strukturen im Gewebe 3.10.Zur Untersuchung von nicht-professioneller Phagozytose im Gewebe wurden
Tumorgewebeschnitte verwendet. Die Häufigkeit von Zell-in-Zell Strukturen
wurde in 416 Gewebeproben aus sieben verschiedenen Kohorten quantifiziert.
Zu den Kohorten zählten fünf HNSCC Kohorten sowie eine Rektumkarzinom
und eine Analkarzinom Kohorte (Tabellen 13 – 15). Die fünf HNSCC Kohorten
wurden unterschieden als (i) Erkrankung im Frühstadium mit niedrigem Risiko,
behandelt mit adjuvanter RCT, (ii) Fortgeschrittene Erkrankung mit hohem Risi-
ko, behandelt mit definitiver RT, (iii) Metastasierung, behandelt mit adjuvanter
RT oder RCT, (iv) ohne Fernmetastasen, behandelt mit neoadjuvanter RCT und
(v) Tumore, behandelt mit adjuvanter RCT, mit Gewebeproben von Tumorzent-
rum und Invasionsfront.
Das Gesamt Überleben für fünf Jahre entsprach 60,3 % in den HNSCC Kohor-
ten, 75,0 % in der Rektumkarzinom Kohorte und 68,5 % in der Analkarzinom
Kohorte (Tabelle 16).
Um Zell-in-Zell Strukturen in den TMAs identifizieren zu können, wurde das Ad-
häsionsmolekül E-Cadherin angefärbt. Die Zellkerne wurden mit Hämatoxylin
gefärbt (Abbildung 35). Vollständiges Umschließen der internalisierten Zelle mit
der Zellmembran der phagozytischen Zelle, ein sichelförmiger Nukleus der
phagozytischen Zelle und eine runde Form der inneren Zelle waren Vorausset-
zung für die Wertung als Zell-in-Zell Struktur.
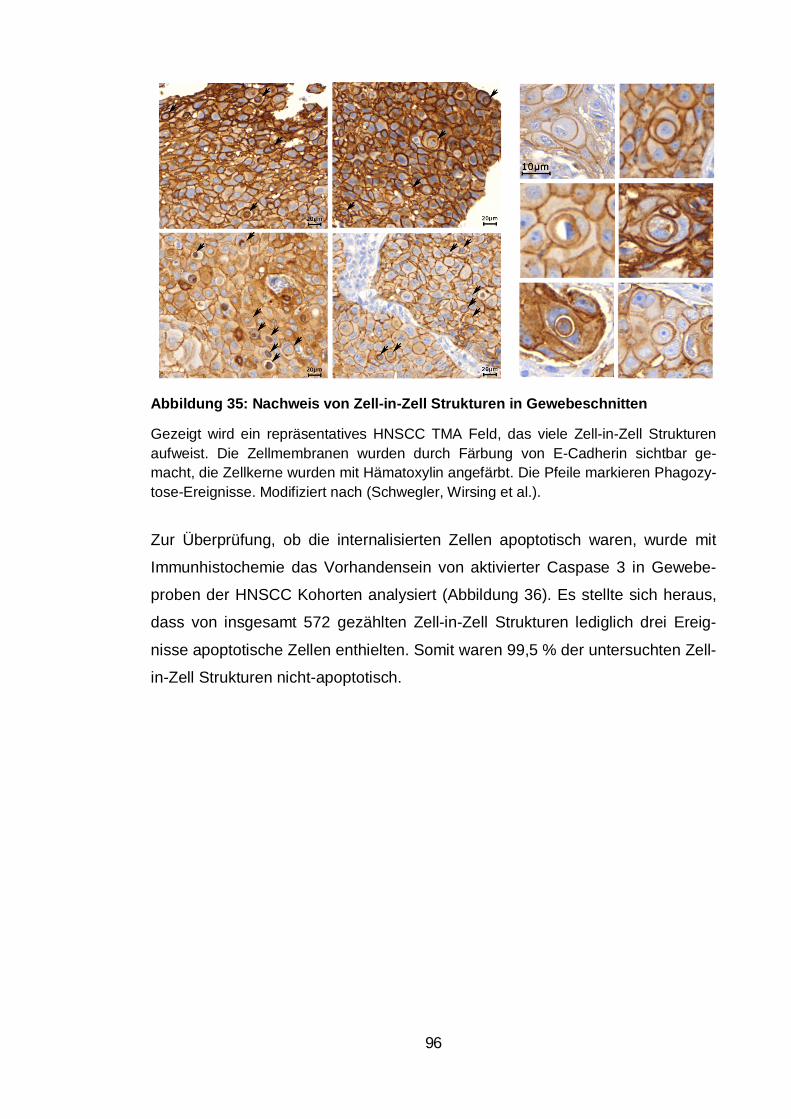
96
Abbildung 35: Nachweis von Zell-in-Zell Strukturen in Gewebeschnitten
Gezeigt wird ein repräsentatives HNSCC TMA Feld, das viele Zell-in-Zell Strukturen aufweist. Die Zellmembranen wurden durch Färbung von E-Cadherin sichtbar ge-macht, die Zellkerne wurden mit Hämatoxylin angefärbt. Die Pfeile markieren Phagozy-tose-Ereignisse. Modifiziert nach (Schwegler, Wirsing et al.).
Zur Überprüfung, ob die internalisierten Zellen apoptotisch waren, wurde mit
Immunhistochemie das Vorhandensein von aktivierter Caspase 3 in Gewebe-
proben der HNSCC Kohorten analysiert (Abbildung 36). Es stellte sich heraus,
dass von insgesamt 572 gezählten Zell-in-Zell Strukturen lediglich drei Ereig-
nisse apoptotische Zellen enthielten. Somit waren 99,5 % der untersuchten Zell-
in-Zell Strukturen nicht-apoptotisch.
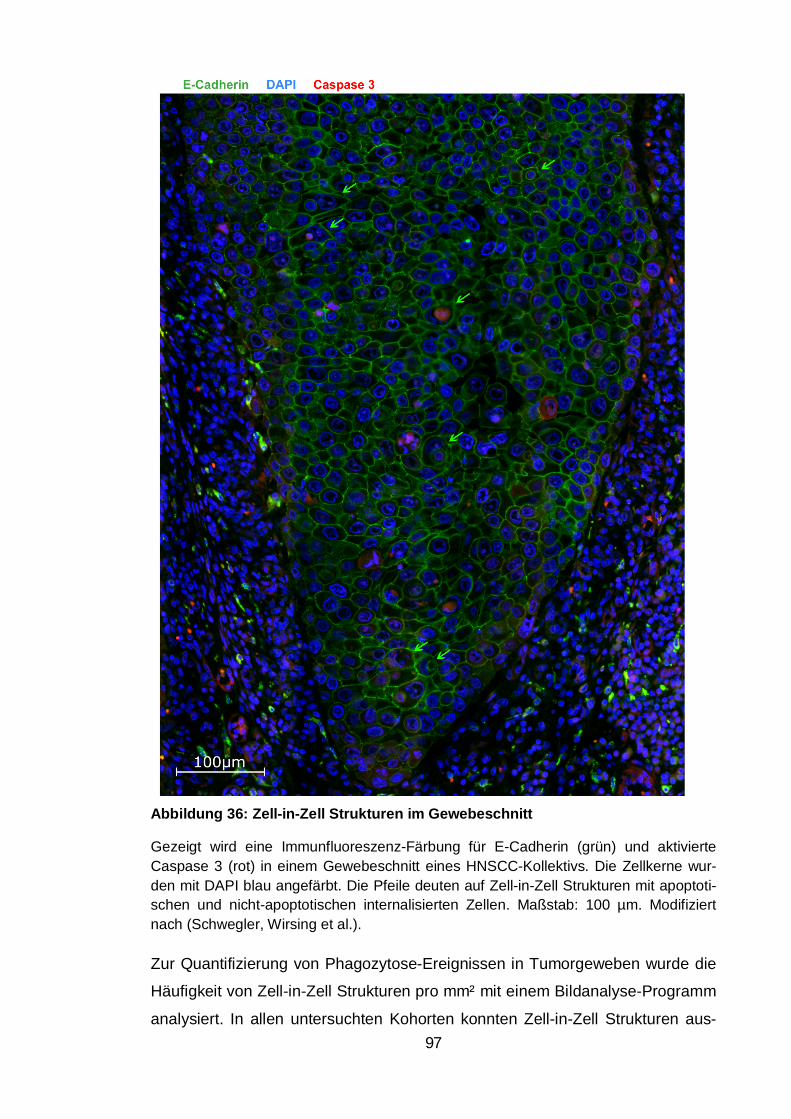
97
Abbildung 36: Zell-in-Zell Strukturen im Gewebeschnitt
Gezeigt wird eine Immunfluoreszenz-Färbung für E-Cadherin (grün) und aktivierte Caspase 3 (rot) in einem Gewebeschnitt eines HNSCC-Kollektivs. Die Zellkerne wur-den mit DAPI blau angefärbt. Die Pfeile deuten auf Zell-in-Zell Strukturen mit apoptoti-schen und nicht-apoptotischen internalisierten Zellen. Maßstab: 100 µm. Modifiziert nach (Schwegler, Wirsing et al.). Zur Quantifizierung von Phagozytose-Ereignissen in Tumorgeweben wurde die
Häufigkeit von Zell-in-Zell Strukturen pro mm² mit einem Bildanalyse-Programm
analysiert. In allen untersuchten Kohorten konnten Zell-in-Zell Strukturen aus-
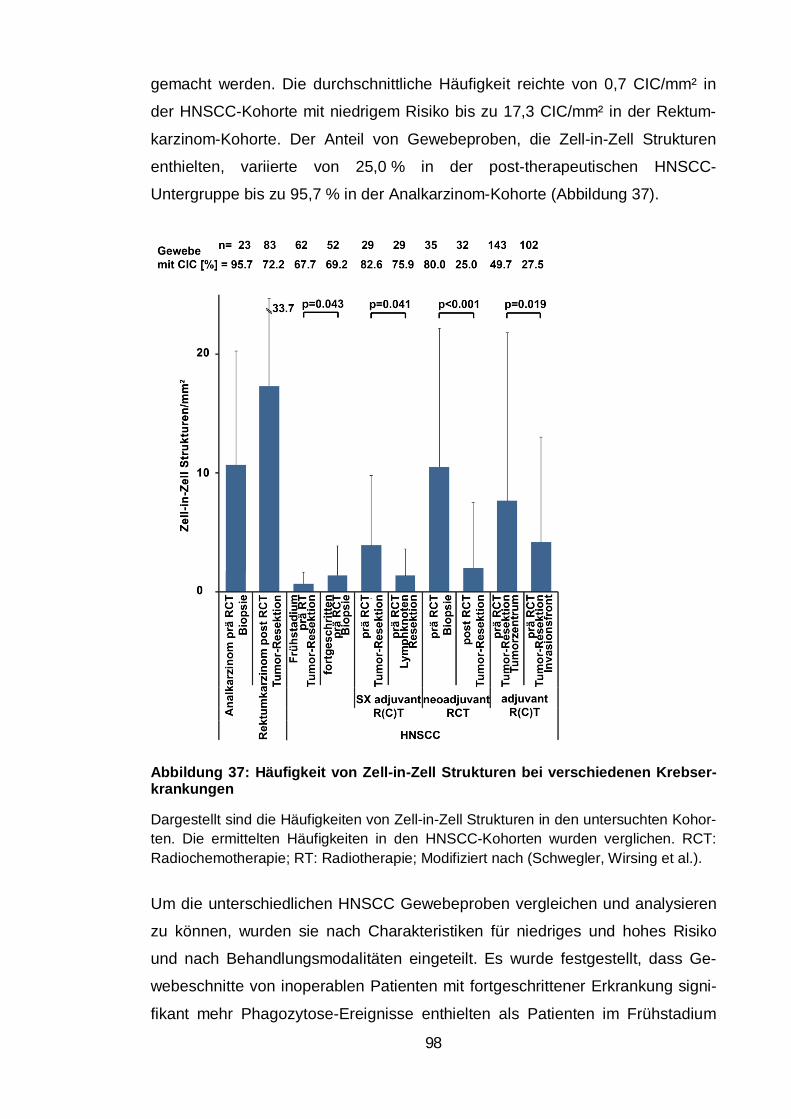
98
gemacht werden. Die durchschnittliche Häufigkeit reichte von 0,7 CIC/mm² in
der HNSCC-Kohorte mit niedrigem Risiko bis zu 17,3 CIC/mm² in der Rektum-
karzinom-Kohorte. Der Anteil von Gewebeproben, die Zell-in-Zell Strukturen
enthielten, variierte von 25,0 % in der post-therapeutischen HNSCC-
Untergruppe bis zu 95,7 % in der Analkarzinom-Kohorte (Abbildung 37).
Abbildung 37: Häufigkeit von Zell-in-Zell Strukturen bei verschiedenen Krebser-krankungen
Dargestellt sind die Häufigkeiten von Zell-in-Zell Strukturen in den untersuchten Kohor-ten. Die ermittelten Häufigkeiten in den HNSCC-Kohorten wurden verglichen. RCT: Radiochemotherapie; RT: Radiotherapie; Modifiziert nach (Schwegler, Wirsing et al.).
Um die unterschiedlichen HNSCC Gewebeproben vergleichen und analysieren
zu können, wurden sie nach Charakteristiken für niedriges und hohes Risiko
und nach Behandlungsmodalitäten eingeteilt. Es wurde festgestellt, dass Ge-
webeschnitte von inoperablen Patienten mit fortgeschrittener Erkrankung signi-
fikant mehr Phagozytose-Ereignisse enthielten als Patienten im Frühstadium

99
der Erkrankung mit niedrigem Risiko (Abbildungen 37 und 38A). Die durch-
schnittliche Anzahl von Zell-in-Zell Strukturen in befallenen Lymphknoten war
signifikant niedriger als in den korrespondierenden Primärtumoren, wohingegen
die Anzahl von Patienten, deren Gewebeproben Phagozytose-Ereignisse ent-
hielten, sehr ähnlich war (Abbildungen 37 und 38B). Der Vergleich von prä-
therapeutischen Biopsien und post-therapeutischen Tumor-Resektionen (sechs
Wochen nach RCT) einer HNSCC Kohorte zeigte eine signifikante Verringerung
der Phagozytose-Ereignisse bei den post-therapeutischen Proben (Abbildungen
37 und 38C). Außerdem wurden die Häufigkeiten von Zell-in-Zell Strukturen im
Tumorzentrum und in der Invasionsfront miteinander verglichen. Dabei wurde
festgestellt, dass sowohl der Anteil von Gewebeproben, die Phagozytose-
Ereignisse enthielten, sowie die durchschnittliche Häufigkeit von Zell-in-Zell
Strukturen im Tumorzentrum höher als in der Invasionsfront waren (Abbildun-
gen 37 und 38D). Die prä-therapeutischen Biopsien der Analkarzinom-Kohorte
wiesen eine hohe durchschnittliche Häufigkeit von 10,7 CIC/mm² auf. Die
höchste Dichte an Zell-in-Zell Strukturen wurde mit einer Häufigkeit von
200 CIC/mm² in einer Gewebeprobe der post-therapeutischen Rektumkarzi-
nom-Kohorte quantifiziert (Abbildungen 37 und 38E). Wegen der geringen Grö-
ße der Kohorte von Patienten mit Analkarzinom (n = 23) wurde die Überlebens-
Analyse als nicht repräsentativ betrachtet.
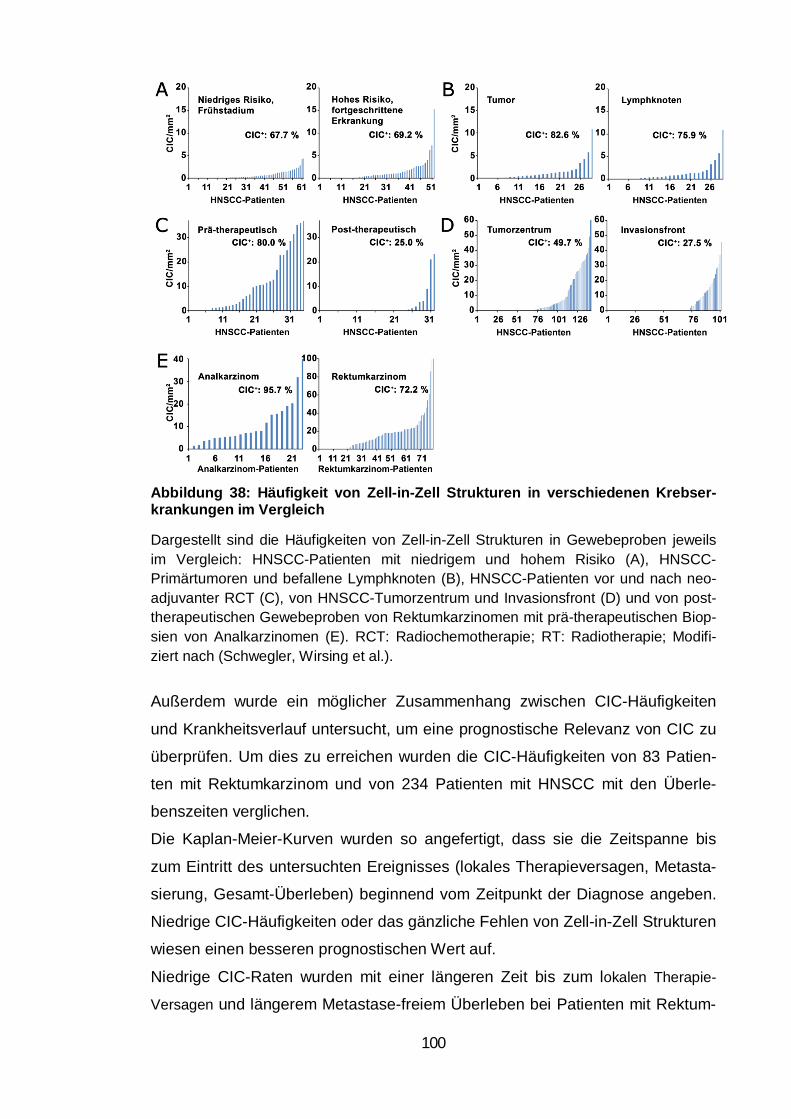
100
Abbildung 38: Häufigkeit von Zell-in-Zell Strukturen in verschiedenen Krebser-krankungen im Vergleich
Dargestellt sind die Häufigkeiten von Zell-in-Zell Strukturen in Gewebeproben jeweils im Vergleich: HNSCC-Patienten mit niedrigem und hohem Risiko (A), HNSCC-Primärtumoren und befallene Lymphknoten (B), HNSCC-Patienten vor und nach neo-adjuvanter RCT (C), von HNSCC-Tumorzentrum und Invasionsfront (D) und von post-therapeutischen Gewebeproben von Rektumkarzinomen mit prä-therapeutischen Biop-sien von Analkarzinomen (E). RCT: Radiochemotherapie; RT: Radiotherapie; Modifi-ziert nach (Schwegler, Wirsing et al.).
Außerdem wurde ein möglicher Zusammenhang zwischen CIC-Häufigkeiten
und Krankheitsverlauf untersucht, um eine prognostische Relevanz von CIC zu
überprüfen. Um dies zu erreichen wurden die CIC-Häufigkeiten von 83 Patien-
ten mit Rektumkarzinom und von 234 Patienten mit HNSCC mit den Überle-
benszeiten verglichen.
Die Kaplan-Meier-Kurven wurden so angefertigt, dass sie die Zeitspanne bis
zum Eintritt des untersuchten Ereignisses (lokales Therapieversagen, Metasta-
sierung, Gesamt-Überleben) beginnend vom Zeitpunkt der Diagnose angeben.
Niedrige CIC-Häufigkeiten oder das gänzliche Fehlen von Zell-in-Zell Strukturen
wiesen einen besseren prognostischen Wert auf.
Niedrige CIC-Raten wurden mit einer längeren Zeit bis zum lokalen Therapie-
Versagen und längerem Metastase-freiem Überleben bei Patienten mit Rektum-

101
karzinom assoziiert (p=0,032). Niedrige CIC-Häufigkeiten oder das Fehlen von
Zell-in-Zell Strukturen korrelierte signifikant mit einem verbesserten Gesamt-
Überleben bei Patienten mit HNSCC (p=0,005) (Abbildung 39).
Abbildung 39: Überlebenskurven nach Kaplan-Meier
Dargestellt sind Kaplan-Meier Kurven für die Kohorten von Patienten mit HNSCC (obe-re Reihe) mit Zell-in-Zell Strukturen (grüne, gepunktete Linie) und ohne Zell-in-Zell Strukturen (blaue Linie). Die Kaplan-Meier Kurven für die Kohorte von Patienten mit Rektumkarzinom (untere Reihe) zeigen Patienten mit mehr oder genau 10 CIC/mm² (grüne, gepunktete Linie) und mit weniger als 10 CIC/mm² (blaue Linie). Modifiziert nach (Schwegler, Wirsing et al.).

102
4. Diskussion
Phagozytose-Induktion durch Hyperthermie 4.1.Die Phagozytose-Aktivität von sog. nicht-professionellen Phagozyten, meist
benachbarten Zellen im Gewebeverband, ist ein bekanntes Phänomen, deren
zugrunde liegenden Mechanismen sowie die dazu fähigen Zelltypen bisher
noch nicht bis ins Detail erforscht sind. Der Fokus dieser Studie wurde auf die
Aufnahme von primär nekrotischen Zellen durch verschiedene Tumorzellen so-
wie primären gesunden Haut-Fibroblasten gelegt. Für die Induktion der pri-
mären Nekrose wurde zum einen die Behandlung mit starker Hitze (45 min bei
56 °C) und zum anderen Bestrahlung gewählt. Da sich die Hyperthermie als
zuverlässigere Methode mit einer Nekrose-Induktionsrate von 100 % bewährte,
wurde diese Behandlung für die meisten in dieser Arbeit durchgeführten Expe-
rimente verwendet.
Auffällig war, dass bei dieser Behandlung, im Gegensatz zur Bestrahlung, kaum
nekrotische Debris entstand. Die Morphologie der hyperthermierten Zellen ent-
spricht dem frühen Stadium der primären Nekrose. Die Zellen enthielten einen
intakten Zellkern bei gleichzeitiger Lyse der Zellmembran. Die durchflusszyto-
metrischen Messungen bestätigten, dass die Behandlung mit Hyperthermie die
Permeabilisierung der Zellmembran und den nekrotischen Zelltod induzierte.
Um den Effekt der Hyperthermie auf die Proteine der behandelten Zellen zu
erfassen, wurde die Protein-Konzentration basierend auf der eingesetzten Zell-
zahl mit einem BCA photometrisch bestimmt. Dabei stellte sich heraus, dass die
Behandlung mit Hitze eine drastische Reduktion der messbaren Proteine her-
vorrief.
Zusätzlich wurde mittels Immunfluoreszenz der Verlust von E-Cadherin, die Un-
fähigkeit zur Phosphorylierung von Ezrin und zur Formation von Aktinfilamenten
nachgewiesen. Insgesamt deuten diese Ergebnisse darauf hin, dass die be-
handelten Zellen ohne eigenes Mitwirken passiv von der phagozytischen Zelle
aufgenommen werden.
Da gezeigt werden konnte, dass unbehandelte, lebende Zellen in demselben
Phagozytose-Versuch im Suspensions-System nicht internalisiert werden, wird
angenommen, dass diese Veränderungen der Zellmembran von nekrotischen
Zellen die Phagozytose stimulieren. Der erste Schritt dabei ist eine verstärkte
Adhäsion an die beschädigten Zellen, wodurch schließlich die Aufnahme der

103
nekrotischen Zellen induziert wird. Die Korrelation zwischen dem Grad der Ad-
häsion und der Phagozytose-Rate wurde mit einem Temperatur-Gradienten
demonstriert. Nach der Behandlung mit Temperaturen zwischen 42 – 56 °C für
45 min wurden die Adhäsions-Rate und die Phagozytose-Rate quantifiziert. Da-
bei wurde eine kritische Temperatur von 54 °C identifiziert, die zu einer signifi-
kanten Erhöhung der Adhäsion und Phagozytose führt. Damit wurde gezeigt,
dass die Phagozytose-Rate vom Grad der Adhäsion abhängt und gleichzeitig,
dass eine Behandlung mit starker Hitze notwendig ist, um die Membran ausrei-
chend zu verändern, wodurch die erhöhte Adhäsion zustande kommt.
Hyperthermie als Methode zur Nekrose-Induktion scheint irrelevant für die Nek-
rose in vivo zu sein, dennoch wird die Behandlung mit Hitzeschock als geeigne-
ter Ansatz für die Untersuchung der Beseitigung sterbender Zellen durch be-
nachbarte, nicht-professionelle Phagozyten angesehen, weil die hyperthermier-
ten Zellen dem charakteristischen Erscheinungsbild nekrotischer Zellen, wie
Lyse der Zellmembran gleichen. Ein weiteres Merkmal nekrotischer Zellen ist
der Verlust des proliferativen Potenzials. Dies wurde mit einem Koloniebil-
dungstest getestet, indem bei verschiedenen Temperaturen behandelte Zellen
verwendet werden. Es wurde beobachtet, dass das proliferative Potenzial nach
Behandlung bei 45 °C noch geringfügig vorhanden war, wohingegen die Be-
handlung bei 56 °C keine Proliferation mehr zuließ. Damit wurde die Induktion
von nekrotischem Zelltod bestätigt.
Hitze-Behandlung induziert Zell-Stress und resultiert in der erhöhten Expression
von Hitzeschockproteinen, die in ihrer Funktion als Chaperone, neu exprimierte
Proteine während deren Faltung stabilisieren und beschädigte Proteine reparie-
ren oder zum Abbau dem Proteasom zuführen (Slater, Cato et al. 1981,
Hightower 1991, Hartl 1996). Der nachgewiesene Protein-Verlust durch die ge-
wählte Temperatur-Behandlung könnte zur erhöhten HSP70-Expression führen.
Wegen der induzierten Disintegrität der Zellmembran wurde eine erhöhte
HSP70-Konzentration im Überstand erwartet. Diese Fragestellung wurde mit
einem ELISA untersucht, bei dem die Überstände basierend auf der eingesetz-
ten Zellzahl von hyperthermierten und unbehandelten Zellen in Suspensionskul-
tur nach 4 h und 24 h bestimmt wurde. Dabei wurden in allen Proben relative
hohe HSP70-Konzentrationen gemessen, wider Erwarten wurden jedoch die
höchsten Werte bei den unbehandelten Zellen festgestellt. Es ist anzunehmen,
dass die hohe HSP70-Konzentration bei den lebenden Zellen auf eine aktive

104
Sekretion zurückzuführen ist und die hohe Menge an HSP70 auf eine Hochre-
gulation der HSP70-Expression aufgrund einer Stressreaktion, die durch die
Kultur in Suspension hervorgerufen wurde. Die Stress-Reaktion von adhärenten
Zellen in Suspensionskultur kann das Eintreten in die Anoikis, einer Form der
Apoptose, induzieren (Meredith, Fazeli et al. 1993, Frisch and Francis 1994).
Die gemessenen, niedrigeren Werte für die HSP70-Konzentration nach Hyper-
thermie sind möglicherweise auf eine Denaturierung von HSP70 und eine Re-
duktion der Genexpression aufgrund der Induktion von primärer Nekrose zu-
rückzuführen und geben die Konzentration von passiv freigesetztem HSP70 an.
Nach Behandlung mit Hyperthermie wurde eine Akkumulation des Autophagie-
Markers LC3 und des Lysosomen-Markers LAMP1 mittels Immunfluoreszenz
beobachtet. LC3-positive Vesikel, anstelle eines diffusen Färbemusters, gelten
als Marker für die Induktion von Autophagie, wobei allein die Anwesenheit die-
ser Vesikel nicht ausreichend ist, um Autophagie nachzuweisen. Eine Rolle der
Autophagie-Maschinerie wurde inzwischen auch für die Pathogenese von neu-
rodegenerativen Erkrankungen impliziert (Shibata, Lu et al. 2006). LC3 und wei-
tere Autophagie-assoziierte Proteine, die bei der Formation von Autophagoso-
men beteiligt sind, wurden auch mit dem nekrotischen Zelltod in Verbindung
gebracht (Yu, Alva et al. 2004, Erlich, Shohami et al. 2006). Diesen Proteinen
wurde sogar eine essentielle Rolle für den Ablauf der Nekrose zugeschrieben,
insbesondere LC3 (Samara, Syntichaki et al. 2008). Aufgrund dieser Studien
und der charakterisierten Eigenschaften der hyperthermierten Zellen wird ange-
nommen, dass die Hitze-Behandlung, trotz der dokumentierten Anhäufung von
LC3-positiven Vesikeln, Nekrose anstelle von Autophagie in den durchgeführten
Versuchen induzierte.
Der Phagozytose-Versuch wurde in einem Suspensions-System durchgeführt,
wobei in einem Experiment mit auf Objektträgern angewachsenen Zellen die
Phagozytose von nekrotischen Zellen durch adhärente Zellen ebenfalls nach-
gewiesen wurde. Daher kann angenommen werden, dass diese Vorgänge auch
im Gewebeverband auftreten können. Die Aufnahme fand gleichermaßen ho-
motypisch wie heterotypisch statt, war also nicht an die Interaktion zwischen
Zellen derselben Zelllinie gebunden. Dies deutet auf die Erkennung ubiquitär
exprimierter Moleküle hin, die Adhäsions-fördernd wirken und schließlich die
Phagozytose stimulieren. Zur Untersuchung beteiligter Proteine wurden die

105
phagozytierenden Zellen vor der Koinkubation mit nekrotischen Zellen mit ver-
schiedenen Agenzien behandelt. Dazu zählte die intensive Behandlung mit Pro-
teasen, welche eine Reduktion von Oberflächenmolekülen wie Adhäsionsmole-
külen bewirken sollten, die Inhibition der Aktinfilamente und Mikrotubuli sowie
die Hemmung der GTPase Aktivität von Dynamin II, das eine essentielle Rolle
bei der Abschnürung von Vesikeln bei der Endozytose spielt und bereits mit der
professionellen Phagozytose assoziiert wurde (siehe 1.2). Alle durchgeführten
Ansätze führten eine erhebliche Reduktion der Phagozytose-Raten herbei, wo-
bei ein vollständiger Verlust der Phagozytose-Fähigkeit bei Hemmung des Ak-
tin-Zytoskeletts verursacht wurde. Aber auch die Hemmung von Dynamin II und
der Mikrotubuli verringerte jeweils drastisch die Phagozytose-Aktivität. Diese
Versuche beweisen die passive Aufnahme der nekrotischen Zellen, die ohne
zelluläre Aktivität der hyperthermierten Zellen zustande kommt, aber durch die
Veränderungen der Zellmembran bei Nekrose-Induktion hervorgerufen wird.
Phagozytose-Induktion durch Röntgenstrahlung 4.2.Neben der Behandlung mit Hyperthermie wurde die Phagozytose durch nicht-
professionelle Phagozyten auch durch Röntgenstrahlung induziert. Die Dosen
und Inkubationszeiten nach Bestrahlung wurden bei den verwendeten Zelllinien
(T-Zell-Leukämiezellen, Pankreaskarzinom- und Kopf-Hals-Tumorzellen) zur
effizienten Induktion von Nekrose optimiert. Gleichzeitig konnte eine Induktion
von Apoptose allerdings nicht vermieden werden, sodass die Phagozytose-
Versuche die Aufnahme von nekrotisch sowie apoptotisch sterbenden Zellen
repräsentieren.
Dabei wurden ähnlich hohe Phagozytose-Raten bei Pankreaskarzinom- und
Kopf-Hals-Tumorzellen sowie gesunden Haut-Fibroblasten im Vergleich mit Hy-
perthermie quantifiziert. Dies deutet darauf hin, dass die untersuchte phagozyti-
sche Aktivität der verwendeten Zelllinien nicht auf die Beseitigung hyperther-
mierter Zellen beschränkt ist, sondern vielmehr einen Prozess zur Aufnahme
beschädigter, sterbender Zellen darstellt. Außerdem konnte gezeigt werden,
dass nicht nur nekrotische, sondern auch bestrahlte, apoptotische Zellen auf-
genommen werden, indem mittels Immunfluoreszenz-Färbung der Apoptose-
Marker aktivierte Caspase 3 in phagozytischen Strukturen nachgewiesen wur-
de. In Bezug auf eine Studie, welche die Phosphatidylserin-Exposition bei Nek-

106
rose beweist (Hirt and Leist 2003), kann angenommen werden, dass sich nek-
rotische und apoptotische Zellen im Verlust der membranösen Phosphatidylse-
rin-Asymmetrie ähneln. Es ist daher möglich, dass diese Gemeinsamkeit die
Phagozytose-Aktivität von nicht-professionellen Phagozyten stimuliert.
Ähnlich der Phagozytose-Versuchen mit Hyperthermie wurden sowohl homoty-
pische als heterotypische Phagozytose-Strukturen beobachtet. Die Aufnahme
von bestrahlten T-Zellen erwies sich dabei am effizientesten, was womöglich
auf deren kleinere Zellgröße zurückzuführen ist.
Durch die Bestrahlung mit folgenden langen Inkubationszeiten (drei Tage bis zu
drei Wochen) häufte sich nekrotische Debris im Überstand an. Um dem An-
spruch gerecht zu werden, dass die Beseitigung nekrotisch sterbender Zellen
untersucht werden soll, muss die Aufnahme von zellulären Bruchstücken eben-
falls berücksichtigt werden. Die Quantifizierung dieser zellulären Bruchstücke
wurde auf eindeutige phagozytische Strukturen beschränkt, bei denen eine si-
chelförmige Deformation des Zellkerns des Phagozyten vorhanden war. Insge-
samt konnte eine große Anzahl derartiger Phagozytose-Ereignisse festgestellt
werden. Dies beweist, dass nicht nur nekrotische Zellen, die sich im frühen Sta-
dium des Zelltods befinden, aufgenommen werden, sondern auch zelluläre
Bruchstücke von nicht-professionellen Phagozyten gebunden und effizient be-
seitigt werden.
Rekrutierung Phagozytose-assoziierter Proteine 4.3.Nachdem die Ergebnisse der Inhibitions-Versuche von Zytoskelett und Memb-
ranoberflächenmolekülen (siehe 3.7.1 – 2) deren essentielle Beteiligung bei der
Durchführung der Phagozytose gezeigt hatten, wurden Immunfluoreszenz-
Färbungen zu verschiedenen Zeitpunkten während des Phagozytose-
Prozesses zur Anfärbung von F-Aktin, E-Cadherin und dem Verbindungsprotein
von Zellmembran und Zellskelett Ezrin unternommen.
Die fluoreszenzmikroskopischen Aufnahmen unterstützen die vorangegange-
nen Ergebnisse und verifizieren eine Verdichtung von F-Aktin sowie von dem
Adhäsionsmolekül E-Cadherin an der Kontaktstelle zur nekrotischen Zelle wäh-
rend des Aufnahmeprozesses. Auch für die phosphorylierte Form von Ezrin, die
für Bewegungen der Zellmembran notwendig ist, konnte ein ähnliches Färbe-
muster nachgewiesen werden.

107
Des Weiteren wurde eine Anhäufung von LC3-positiven Vesikeln an der Kon-
taktstelle zur aufzunehmenden Zelle beobachtet. LC3 wird zusätzlich zur Auto-
phagie auch mit der LC3-assoziierten Phagozytose (LAP) in Verbindung ge-
bracht. Dieser Prozess beschreibt die Rekrutierung von LC3 zu Phagosomen
und anschließender Fusion dieser Vesikel mit Lysosomen.
Der LAP wurde bisher eine Rolle für die Beseitigung von Pathogenen und ster-
benden Zellen durch professionelle Phagozyten, sowie der Aufnahme von au-
tophagischen Zellen (durch Anoikis oder Tamoxifen-Behandlung) durch epithe-
liale Brustkarzinom-Zellen (Petrovski, Zahuczky et al. 2007) und bei der Entose
zugeschrieben (Florey, Kim et al. 2011). In Übereinstimmung mit der LAP zei-
gen die durchgeführten Versuche eine temporäre Rekrutierung von LC3 und
eine Akkumulation von LAMP1-positiven Vesikeln im Zytoplasma des nicht-
professionellen Phagozyten um die aufgenommene Zelle herum. Daher wird ein
ähnlicher Mechanismus, bei dem LAP involviert ist, für die Aufnahme primär
nekrotischer Zellen durch nicht-professionelle Phagozyten vorgeschlagen, wo-
bei weitere zukünftige Experimente dieser Hypothese belegen müssen.
Da es sich bei diesem Phagozytose-Versuch um eine Kombination eines Sus-
pensions-Systems mit adhärent wachsenden Zellen handelt, musste überprüft
werden, ob die beobachteten Färbemuster der untersuchten Proteine auf die für
den Zelltyp unnatürlichen Bedingungen zurückzuführen sind oder aber auch bei
Zellen im angewachsenen Zustand auftreten. Deshalb wurden Versuche mit auf
Objektträgern angewachsenen Zellen durchgeführt und die Immunfluoreszenz-
Färbungen zu den gleichen Zeitpunkten durchgeführt. Dabei wurde eine stark
ähnelnde Lokalisation der angefärbten Proteine in den phagozytischen Struktu-
ren festgestellt. Daher wurde die Interaktion von E-Cadherin, F-Aktin, p-Ezrin,
LC3 und LAMP1 bei der Phagozytose-Aktivität als wichtiger Bestandteil des
Prozesses gewertet.
Zelltod der inkorporierten Zelle 4.4.Die Beobachtungen zur Entwicklung von Phagozytose-Ereignissen über die Zeit
(bis zu drei Tage) ergaben, dass die aufgenommene Zelle innerhalb von 24 h
zugrunde geht. Diese Annahme basiert auf dem beobachteten Zerfall des Nuk-
leus, der mittels DAPI-Färbung verifiziert wurde. Bei Zell-in-Zell Strukturen, die
sich nach 4 h gebildet hatten, waren noch beide Zellkerne sichtbar, wobei der

108
Zellkern der phagozytierenden Zelle sichelförmig an die Peripherie gedrängt
wurde. Nach 24 h war der Zellkern der aufgenommenen Zelle nicht mehr mit
DAPI färbbar. Da dieser DNA-Farbstoff bevorzugt an die kleine Furche der
Doppelhelix bindet (Cavatorta, Masotti et al. 1985), ist anzunehmen, dass die
Integrität der DNA innerhalb von 24 h verloren ging. Zu diesem Zeitpunkt nahm
der Nukleus der phagozytischen Zelle bereits seine ursprüngliche, ovale Form
wieder an.
Des Weiteren wurde eine Verringerung der Immunfluoreszenzsignale für
HSP70 und LAMP1 nachgewiesen wurde. Der Abbau und der Expressions-
stopp dieser angefärbten Proteine beweisen zusammen mit dem Verlust der
Integrität von DNA den unumkehrbaren Zelltod. Jedoch scheint insgesamt der
Abbau der inkorporierten Zelle nur sehr langsam vonstatten zu gehen, da selbst
nach 72 h noch phagozytiertes Material nachgewiesen wurde. Die Frage, ob die
aufgenommene Zelle von der phagozytischen Zelle überhaupt vollständig de-
gradiert wird, konnte nicht beantwortet werden. Es wurde aber auch keine In-
duktion von Zelltod bei der phagozytierenden Zelle beobachtet. Somit deuten
die Ergebnisse doch mehr auf den Abbau der inkorporierten Zelle hin.
In der Folge könnte der Phagozyt das Material mittels Exozytose ausschleusen,
Nährstoffe verwenden und die Moleküle für den Aufbau zellulärer Strukturen
nutzen. In diesem Zusammenhang wurde ein Transfer von DNA-Material bei
der Phagozytose von apoptotische Körperchen durch Fibroblasten, Makropha-
gen und Endothelzellen beschrieben, der eine Übertragung von transkriptions-
fähigen, aktiven Genen ermöglicht (Holmgren, Szeles et al. 1999). Um dies zu
untersuchen wurden die aufzunehmenden Zellen vor der Hitze-Behandlung
entweder mit Histon H2B-RFP transfiziert oder mit dem Nukleosid-Analogon
EdU markiert, um Bestandteile des Chromatins sichtbar zu machen. Innerhalb
von 48 h wurde keine Übertragung des Kernmaterials in den Nukleus des Pha-
gozyten festgestellt. Es ist jedoch nicht vollkommen auszuschließen, dass das
markierte Kernmaterial dennoch in den Nukleus des Phagozyten eingebaut
wurde, sofern es zuvor proteolytisch gespalten wurde und daher nicht mehr
sichtbar war.

109
Nicht-professionelle Phagozytose versus professionelle 4.5.
Phagozytose Es wird angenommen, dass ein Teil der geschätzten 50-70 Mio Zellen, die pro
Tag im menschlichen Körper sterben und ersetzt werden, von benachbarten
Zellen, als nicht-professionelle Phagozyten, entfernt werden (Reed 2006). Da-
bei unterstützen diese Zellen die Immunzellen, welche als professionelle Pha-
gozyten Gewebeschäden erkennen und effizient zelluläre Debris, die pro-
phagozytische „eat-me“ Signale exponiert und DAMPs freisetzt, beseitigen. Zu
diesen endogenen Moleküle zählen F-Aktin (Ahrens, Zelenay et al. 2012), Hit-
zeschockproteine (HSP70, HSP90) und Calretikulin (Obeid, Tesniere et al.
2007), sowie modifizierte Lipide und Proteine der Zellmembran.
Zur Untersuchung, ob diese pro-phagozytischen Signale auch die Phagozytose-
Aktivität bei den verwendeten nicht-professionellen Phagozyten stimulieren,
wurde das Medium nach der Hyperthermie-Behandlung ersetzt. Die gewonne-
nen Phagozytose-Raten wichen nicht von der Kontrolle ab und deuten daher
darauf hin, dass freigesetzte DAMPs, wie das im Überstand nachgewiesene
HSP70 nicht notwendig sind, um die Zellen auf die Phagozytose vorzubereiten.
Die exponierten pro-phagozytischen Signale, vor allem PS, scheinen ausrei-
chend zu sein, um die Phagozytose zu vermitteln. Es kann jedoch nicht ausge-
schlossen werden, dass die Menge von DAMPs, die während der Koinkubation
freigesetzt wird, diesen Effekt verstärkte.
Die Dosis-Wirkungs-Kurve mit AnxA5 demonstrierte, dass PS-Rezeptor Interak-
tionen eine Schlüsselrolle bei der effizienten Aufnahme nekrotischer Zellen
spielen, aber auch, dass die Phagozytose nicht vollständig und limitierend von
der Erkennung von PS abhängig ist. Bei der Expressionsanalyse von verwen-
deten phagozytischen und nicht-phagozytischen Zelllinien stellte sich heraus,
dass die untersuchten pro-phagozytischen Gene von den Zelllinien unterschied-
lich stark exprimiert wurden. Die gewählten Phosphatidylserin- und Tethering-
Rezeptoren (CD68, TIM1, CD36, ITGB5) sind für ihre Funktion bei der Pha-
gozytose apoptotischer Zellen bekannt (Ramprasad, Fischer et al. 1995,
Sambrano and Steinberg 1995, Fadok, Warner et al. 1998, Kobayashi, Karisola
et al. 2007, Singh, D'Mello et al. 2007). Ein Beweis für die direkte Beteiligung
der nachgewiesenen, exprimierten Rezeptoren blieb in dieser Arbeit aus. Auf-
grund der Abhängigkeit von der Bindung an PS auf nekrotischen Zellen, wird

110
jedoch die individuelle Effizienz der verwendeten Zelllinien auf ihr jeweiliges
Rezeptor-Repertoire zurückgeführt. Da gezeigt werden konnte, dass semi-
adhärente Zellen, die also wenig Adhäsionsmoleküle exprimieren, nicht Pha-
gozytose-fähig sind, stellt eine weitere Ursache für die unterschiedlichen Pha-
gozytose-Raten womöglich die Zelltyp-spezifische Expression von Adhäsions-
molekülen dar.
Im Vergleich mit der Phagozytose von kleinen, nekrotischen Partikeln durch
Makrophagen, die mit Raten von bis zu 10 % nach nur 30 min Koinkubation
beschrieben wird, wurden insgesamt eher niedrige Phagozytose-Raten quantifi-
ziert (Brouckaert, Kalai et al. 2004). Erste Phagozytose-Ereignisse wurden frü-
hestens nach 2,5 h beobachtet. Dies deutet darauf hin, dass der phagozytische
Prozess bei nicht-professionellen Phagozyten langsamer abläuft und daher im
Vergleich mit der Phagozytose-Aktivität der professionellen Phagozyten weni-
ger effizient ist. Als Ursache dafür wird das reduzierte Rezeptor-Repertoire für
die Erkennung sterbender Zellen sowie die Tatsache, dass die hyperthermier-
ten Zellen nicht in kleine Partikel zerfallen, sondern als ganze Zellen aufge-
nommen werden müssen, vermutet.
Die Zell-in-Zell Strukturen blieben in Suspensionskultur bis zu drei Tage erhal-
ten, wobei ein Abbau der DNA der aufgenommenen Zelle innerhalb von 24 h
beobachtet wurde (siehe 3.9). Im Gegensatz dazu fördern Makrophagen nach
der Phagozytose von apoptotischen Körperchen sehr effizient den Abbau des
phagozytierten Materials. Dadurch kommt es in der Regel innerhalb von weni-
gen Stunden zum kompletten lysosomalen Abbau des phagozytierten Materials
in die zellulären Grundbausteine wie Peptide, Aminosäuren, Fette, Sterole und
Nukleotide via Phagosomen-Reifung und Fusion mit Lysosomen (siehe 1.2).
Durch die Aktivität der Makrophagen-eigenen lysosomalen DNase II wird die
Aktivität der Caspase-aktivierten DNase der apoptotischen Zelle zur enzymati-
sche Spaltung der DNA unterstützt (McIlroy, Tanaka et al. 2000). Die Akkumu-
lation von LAMP1 positiven Vesikeln um die internalisierte Zelle herum, deutet
auf eine ähnliche Beteiligung von lysosomalen Enzymen der nicht-
professionellen Phagozyten bei der Degradation der nekrotischen Zellen hin.

111
Nicht-professionelle Phagozytose versus Kannibalismus 4.6.Der Zell-Kannibalismus wird als erworbene Fähigkeit maligner Tumorzellen be-
schrieben, die der Nährstoffversorgung kannibalistischer Zellen im Tumormilieu
dienen soll. Dabei unterscheiden die Tumorzellen bei der Aufnahme nicht zwi-
schen lebenden und toten Tumorzellen oder Lymphozyten (Caruso, Muda et al.
2002, Lugini, Lozupone et al. 2003, Fais 2007). Im Gegensatz dazu fand in den
durchgeführten Phagozytose-Versuchen nur die Aufnahme von sterbenden Zel-
len statt. Die selten beobachteten Phagozytose-Ereignisse von lebenden Zellen
wurden eher als Aufnahme von beschädigten Zellen betrachtet, die aufgrund
der Trypsinierung und der langen Inkubation in Suspension vermutlich Memb-
ranveränderungen aufwiesen.
Eine Gemeinsamkeit zum Zell-Kannibalismus stellt die Tatsache dar, dass
nicht-professionellen Phagozyten auch im angewachsenen Zustand phagozy-
tierten, ähnlich dem Zell-Kannibalismus, der im Gewebeverband stattfindet.
Außerdem wurde, aufgrund der Hypothese zur Nährstoffversorgung, der Ein-
fluss von temporärem FBS-Mangel von ein bis drei Tagen auf die kannibalisti-
sche Aktivität untersucht und in Studien mit malignen Melanomzellen als Kanni-
balismus-fördernd nachgewiesen. Dieser Effekt wurde allerdings exklusive bei
metastatischen Melanomzellen gezeigt und für Zellen des Primärtumors ausge-
schlossen (Lugini, Matarrese et al. 2006). Diese Studie impliziert, dass kanniba-
listische Zellen durch die Aufnahme und Degradation anderer Zellen einen Vor-
teil erlangen, um im nährstoffarmen Tumormilieu zu überleben. Eine weitere in
vitro Untersuchung zeigte die Abhängigkeit von Serum auf die Kannibalismus-
Aktivität einer kleinzelligen Lungenkarzinom-Zelllinie. Langanhaltender Serum-
Entzug (vier Wochen) verhinderte die Bildung von Zell-in-Zell Strukturen. Die
kannibalistische Aktivität konnte aber durch Serum-Zugabe erneut aktiviert wer-
den (Brouwer, de Ley et al. 1984). Aufgrund der erheblichen Unterschiede, die
Dauer des Serumentzugs betreffend, können beide Studien nicht direkt mitei-
nander verglichen werden. Es wird jedoch klar, dass der kurzzeitige Serum-
Entzug den Zell-Kannibalismus fördert, sich die Zellen aber in einem gesunden
Grundzustand befinden müssen, um kannibalistisch aktiv sein zu können, was
durch langanhaltenden Serum-Entzug verhindert wird. Bei den in dieser Arbeit
durchgeführten Versuchen wurden die Zellen eine Woche ohne Passage bis
zum konfluenten Wachstum kultiviert. Vor dem Phagozytose-Versuch wurde

112
das Medium mit Serum-freiem Medium für 24 h ersetzt und unter diesen Bedin-
gungen eine erhebliche Verringerung der Phagozytose-Aktivität festgestellt.
Diese Ergebnisse zeigen, dass die Vitalität der phagozytischen Zellen entschei-
den ist, ob Phagozytose stattfindet und die phagozytische Aktivität der unter-
suchten nicht-professionellen Phagozyten nicht als Überlebensmechanismus
angesehen werden kann.
Nicht-professionelle Phagozytose versus Entose 4.7.Die Entose wurde als nicht-apoptotische Zelltodesform beschrieben, die mit der
Bildung morphologisch sehr ähnlicher Zell-in-Zell Strukturen einhergeht (siehe
1.4.3). Diese invasive Aktivität von lebenden Tumorzellen in benachbarte Tu-
morzellen wurde als ursächlicher Mechanismus für einen Teil der in Tumorge-
webeschnitten beobachteten Zell-in-Zell Strukturen vorgeschlagen (Overholtzer,
Mailleux et al. 2007). Weil dieser Prozess durch eine Ablösung von der extra-
zellulären Matrix induziert wird, wurde eine Tumorsuppressor-Funktion vorge-
schlagen, indem Zellen, die sich außerhalb ihrer Nische aufhalten, entfernt wer-
den. Im Gegensatz zu den in dieser Arbeit untersuchten nicht-professionellen
Phagozyten findet Entose nur im Suspensions-System statt, nicht aber, wenn
die Wirtszellen angewachsen sind. In diesem Zusammenhang muss die Hypo-
these über eine Tumorsuppressor-Funktion durch Entose kritisch betrachtet
werden, wenn auch die Wirtszelle von der extrazellulären Matrix abgelöst ist.
Eine aktuellere Studie behauptet eher das Gegenteil, nämlich eine Induktion
von Aneuploidie durch Entose, das die Entstehung maligner Tumorzellen för-
dert (Krajcovic, Johnson et al. 2011). Aneuploide Zellen wurden bei den in die-
ser Arbeit durchgeführten Versuchen nur bei den verwendeten Tumorzelllinien
beobachtet, nicht bei den primären Hautfibroblasten. Daher kann eine Gemein-
samkeit mit der Entose diesbezüglich ausgeschlossen werden. Weiterhin wur-
den Unterschiede bei der Beteiligung von E- und P-Cadherin nachgewiesen.
Die Interaktion dieser Cadherine zwischen Wirtszelle und internalisierender Zel-
le wurde bei der Entose mit Funktions-blockierenden Antikörpern gezeigt
(Overholtzer, Mailleux et al. 2007). Die untersuchte Pankreaskarzinom-Zelllinie
zeigte hingegen keine spezifische Inhibition von Adhäsion und Phagozytose-
Rate bei Zugabe dieser blockierenden Antikörper. Dennoch scheinen auch bei

113
der nicht-professionellen Phagozytose Cadherine beteiligt zu sein, da eine Ak-
kumulation von E-Cadherin an der Kontaktstelle nachgewiesen werden konnte.
Eine Gemeinsamkeit der Phagozytose-Aktivität mit der Entose wurde bei der
Rekrutierung des Autophagiemarkers LC3 beobachtet. Bei der Entose wurde
die Akkumulation von LC3 auf der entotischen Vakuolenmembran beobachtet.
Auch eine Abhängigkeit von den Autophagie-assoziierten Proteinen ATG5,
ATG7 und der Klasse III PI-3-Kinase VPS34 für die Fusion mit Lysosomen und
dem Abbau der internalisierten Zelle wurde nachgewiesen (Florey, Kim et al.
2011). In den in dieser Arbeit durchgeführten Versuchen wurden ebenfalls eine
ringförmige Rekrutierung von LC3, sowie die Akkumulation von Lysosomen um
die phagozytierte Zelle gezeigt. Dies deutet auf eine Involvierung des LC3-
assoziierten Phagozytose-Mechanismus hin, der den lysosomalen Abbau der
entotischen und nekrotischen Zelle auf ähnliche Weise fördert.
Es wurde berichtet, dass die inkorporierten Zellen entweder durch lysosomale
Enzyme der Wirtszelle abgebaut werden, eine Zellteilung durchlaufen oder
noch lebend die Wirtszelle verlassen (Overholtzer, Mailleux et al. 2007). Im Ge-
gensatz dazu trifft für die nicht-professionellen Phagozyten nur der lysosomale
Abbau und Zelltod der aufgenommenen nekrotischen Zelle zu.
Prognostischer Wert von Zell-in-Zell Strukturen bei Krebs-4.8.
erkrankungen Obwohl Zell-in-Zell Strukturen häufig in Tumoren beobachtet werden, wurden
sie bisher nahezu ignoriert und nicht als möglicher prognostischer Faktor in Er-
wägung gezogen (Wang and Wang 2013).
Für die Untersuchung einer möglichen Korrelation zwischen den Häufigkeiten
von CIC-Ereignissen im Tumorgewebe und der Prognose für den Patienten
wurden TMAs verschiedener Kohorten mit Rektumkarzinom und HNSCC her-
angezogen. Die Analyse ergab, dass in allen untersuchten Kohorten CIC-
Strukturen vorhanden waren. Die Anzahl von Patienten mit CIC-Ereignissen
und die CIC-Raten variierten aber beträchtlich und waren zum Teil abhängig
vom Krebs-Typ, dem Stadium, vorangegangener Therapie-Modalität und der
Lokalisation der CIC-Ereignisse innerhalb des Tumors. Zudem wurden niedrige
CIC-Häufigkeiten mit einer besseren Prognose assoziiert. Deshalb wurden CIC-

114
Strukturen in den untersuchten Kohorten von Patienten mit Rektumkarzinom
und HNSCC als prognostisch relevant betrachtet.
In einer Studie von homotypischen CIC-Ereignissen im Tumorgewebe von Pati-
enten mit Pankreasadenokarzinom wurde eine Korrelation zwischen CIC-
Häufigkeiten und niedrigerem Metastasierungs-Risiko festgestellt und eine
Schutzfunktion vor Metastasen durch CIC-Ereignisse vorgeschlagen (Cano,
Sandi et al. 2012). Im Gegensatz dazu belegen andere Studien einen Zusam-
menhang von homotypischem Zell-Kannibalismus in Brustkarzinomen (Abodief,
Dey et al. 2006), Blasenkarzinomen (Kojima, Sekine et al. 1998), Medulloblas-
tomen (Kumar, Hosseinzadeh et al. 2001) sowie von heterotypischem Zell-
Kannibalismus in Magenkarzinomen (Caruso, Muda et al. 2002) mit schlechter
Prognose. In Bezug auf diese widersprüchlichen Studien, deuten die Ergebnis-
se der untersuchten Kohorten darauf hin, dass CIC-Ereignisse auf verschiede-
ne Arten die Tumorerkrankung beeinflussen. Die Ursache für die beschriebenen
Diskrepanzen liegt womöglich in Gewebe- und Zelltyp-spezifischen Unterschie-
den in Bezug auf Tumor-spezifisches Metastasierungs-Potenzial und Immun-
zell-Infiltration begründet. Die Aufnahme von nekrotischem Zellmaterial durch
Tumorzellen könnte zur Immunevasion beitragen, indem die Phagozytose der
nekrotischen Debris durch Antigen-präsentierende Zellen verhindert und
dadurch die Entwicklung einer T-Zell-vermittelten Immunantwort gegen spezifi-
sche Tumorantigene erschwert wird (Arnold-Schild, Hanau et al. 1999).
In den untersuchten Kohorten waren CIC-Strukturen mit internalisierten apopto-
tischen Zellen sehr seltene Ereignisse. Daher wird angenommen, dass die auf-
genommenen Zellen entweder lebende, von der extrazellulären Matrix abgelös-
te Zellen sind oder nekrotische Zellen darstellen. Nach der Phagozytose durch
Tumorzellen erfolgt möglicherweise ein metabolischer Abbau durch die Wirts-
zelle, sodass internalisierte, lebende Zellen keine Möglichkeit zur Apoptose-
Induktion hätten.
Zusammengefasst lassen sich zwei Hypothesen zum Einfluss von CIC-
Strukturen auf Tumorerkrankungen formulieren: Die Phagozytose von Zellen,
die sich von der extrazellulären Matrix abgelöst haben, ähnlich metastasieren-
den Zellen, durch andere Tumorzellen könnte das Risiko für Metastasierung
senken. Die Beseitigung nekrotisch sterbender Zellen im Tumor hingegen ent-
fernt gleichzeitig immunogene Signale, die eine Tumorimmunität generieren
könnten.

115
Schlussfolgerung und Ausblick 4.9.Die Untersuchungen dieser Arbeit zeigen, dass Tumorzellen und gesunde
Haut-Fibroblasten in der Lage sind, nekrotische sowie apoptotische Zellen auf-
zunehmen und dabei sowohl homotypische als auch heterotypische Zell-in-Zell
Strukturen zu bilden. Die Fähigkeit zur Aufnahme von hyperthermierten, primär
nekrotischen Zellen wurde bei mehreren Zelllinien, inklusive Pankreasadeno-
karzinom-, Glioblastom- und Kopf-Hals-Tumorzellen sowie primären Haut-
Fibroblasten demonstriert. Die Phagozytose hyperthermierter Zellen wurde als
PS-abhängig und LC3-assoziiert charakterisiert und konnte nicht von semi-
adhärenten und Suspensions-Zellen ausgeführt werden. Weiterhin wurden die
Aktivität der GTPase Dynamin II sowie das dynamische Rearrangement der
Aktinfilamente und Mikrotubuli als essentielle mechanistische Faktoren dieses
Prozesses identifiziert. Der Phagozytose-Vorgang beinhaltet die Erkennung von
PS an PS-Rezeptoren, die Verdichtung von E-Cadherin, F-Aktin und p-Ezrin an
der Zell-Zell-Kontaktstelle, gefolgt von einer zeitlich begrenzten Ring-ähnlichen
Formation aus E-Cadherin, p-Ezrin, F-Aktin, LC3 und LAMP1 um die aufge-
nommene Zelle herum und resultiert im Zerfall des Nukleus innerhalb von 24 h
und dem lysosomalen Abbau der phagozytierten Zelle (Abbildung 40).

116
Abbildung 40: Modell zur nicht-professionellen Phagozytose
Die Fähigkeit zur Beseitigung von primär nekrotischen Zellen ist vielen verschiedenen Zelltypen, inclusive Tumorzellen und gesunden Hautfibroblasten, zu eigen. Zur Unter-suchung dieses Vorgangs wurde Nekrose durch Hitze-Behandlung in verschiedenen Zelllinien induziert. Die nekrotischen Zellen enthielten zahlreiche LAMP1- und LC3-positive Vesikel sowie zytoplasmatisches HSP70 nach Hyperthermie. Der homotypi-sche sowie heterotyische Phagozytose-Prozess erfordert seitens der phagozytischen Zelle ein dynamisches Rearrangement des Aktin-Zytoskeletts, die Bindung an expo-niertes PS sowie die GTPase Aktivität von Dynamin II. Die Zelllyse mit der Auflösung des Nukleus der aufgenommenen Zelle trat innerhalb von 24 h nach Phagozytose ein. Modifiziert nach (Schwegler, Wirsing et al. 2015).
Weitere funktionelle Untersuchungen, womöglich auch in vivo Experimente,
sind notwendig, um die molekularen Mechanismen der Aktivität von nicht-
professionellen Phagozyten zur Beseitigung nekrotischer, apoptotischer und
autophagischer Zellen, zu identifizieren.
Phagozytische Strukturen mit internalisierten nekrotischen Zellen, wie sie in
dieser Arbeit durch Hyperthermie induziert wurden, könnten womöglich einem
Teil der beobachteten CIC-Ereignisse im Tumorgewebe, die zum Zell-
Kannibalismus zählen, zugrunde liegen. Die histologischen Analysen der Ko-

117
horten von Patienten mit Rektumkarzinom, Analkarzinom und HNSCC zeigten,
dass CIC-Strukturen mit internalisierten nicht-apoptotischen Zellen ein häufiges
Ereignis in Tumorgewebeschnitten darstellen. Außerdem wurde eine Korrelati-
on zwischen CIC-Häufigkeiten und Krankheitsverlauf bei Patienten mit Rektum-
karzinom und HNSCC nachgewiesen, sodass niedrige CIC-Häufigkeiten mit
einer besseren Prognose assoziiert wurden.
Zukünftige Studien sind notwendig, um das mögliche Potenzial von CIC-
Häufigkeiten als prognostischer Marker in weiteren Krebs-Typen zu erfassen. In
diesem Zusammenhang sollten auch Immunzellen sowie spezifische Proteine,
die für den Mechanismus der nicht-professionellen Phagozytose essentiell sind,
in den Gewebeschnitten quantifiziert werden.

118
5. Literaturverzeichnis Abodief, W. T., P. Dey and O. Al-Hattab (2006). "Cell cannibalism in ductal carcinoma of breast." Cytopathology 17(5): 304-305.
Ahrens, S., S. Zelenay, D. Sancho, P. Hanc, S. Kjaer, C. Feest, G. Fletcher, C. Durkin, A. Postigo, M. Skehel, F. Batista, B. Thompson, M. Way, C. Reis e Sousa and O. Schulz (2012). "F-actin is an evolutionarily conserved damage-associated molecular pattern recognized by DNGR-1, a receptor for dead cells." Immunity 36(4): 635-645.
Arnold-Schild, D., D. Hanau, D. Spehner, C. Schmid, H. G. Rammensee, H. de la Salle and H. Schild (1999). "Cutting edge: receptor-mediated endocytosis of heat shock proteins by professional antigen-presenting cells." J Immunol 162(7): 3757-3760.
Ashburner, M. and J. J. Bonner (1979). "The induction of gene activity in drosophilia by heat shock." Cell 17(2): 241-254.
Basu, S., R. J. Binder, T. Ramalingam and P. K. Srivastava (2001). "CD91 is a common receptor for heat shock proteins gp96, hsp90, hsp70, and calreticulin." Immunity 14(3): 303-313.
Benseler, V., A. Warren, M. Vo, L. E. Holz, S. S. Tay, D. G. Le Couteur, E. Breen, A. C. Allison, N. van Rooijen, C. McGuffog, H. J. Schlitt, D. G. Bowen, G. W. McCaughan and P. Bertolino (2011). "Hepatocyte entry leads to degradation of autoreactive CD8 T cells." Proc Natl Acad Sci U S A 108(40): 16735-16740.
Berman, L. (1966). "Lymphocytes and macrophages in vitro. Their activties in relation to functions of small lymphocytes." Lab Invest 15(6): 1084-1099.
Boya, P., R. A. Gonzalez-Polo, N. Casares, J. L. Perfettini, P. Dessen, N. Larochette, D. Metivier, D. Meley, S. Souquere, T. Yoshimori, G. Pierron, P. Codogno and G. Kroemer (2005). "Inhibition of macroautophagy triggers apoptosis." Mol Cell Biol 25(3): 1025-1040.
Brouckaert, G., M. Kalai, D. V. Krysko, X. Saelens, D. Vercammen, M. N. Ndlovu, G. Haegeman, K. D'Herde and P. Vandenabeele (2004). "Phagocytosis of necrotic cells by macrophages is phosphatidylserine dependent and does not induce inflammatory cytokine production." Mol Biol Cell 15(3): 1089-1100.
Brouwer, M., L. de Ley, C. A. Feltkamp, J. Elema and A. P. Jongsma (1984). "Serum-dependent "cannibalism" and autodestruction in cultures of human small cell carcinoma of the lung." Cancer Res 44(7): 2947-2951.
Cano, C. E., M. J. Sandi, T. Hamidi, E. L. Calvo, O. Turrini, L. Bartholin, C. Loncle, V. Secq, S. Garcia, G. Lomberk, G. Kroemer, R. Urrutia and J. L. Iovanna (2012). "Homotypic cell cannibalism, a cell-death process regulated by the nuclear protein 1, opposes to metastasis in pancreatic cancer." EMBO Mol Med 4(9): 964-979.
Caruso, R. A., A. O. Muda, A. Bersiga, L. Rigoli and C. Inferrera (2002). "Morphological evidence of neutrophil-tumor cell phagocytosis (cannibalism) in human gastric adenocarcinomas." Ultrastruct Pathol 26(5): 315-321.
Cavatorta, P., L. Masotti and A. G. Szabo (1985). "A time-resolved fluorescence study of 4',6'-diamidine-2-phenylindole dihydrochloride binding to polynucleotides." Biophys Chem 22(1-2): 11-16.

119
Chautan, M., G. Chazal, F. Cecconi, P. Gruss and P. Golstein (1999). "Interdigital cell death can occur through a necrotic and caspase-independent pathway." Curr Biol 9(17): 967-970.
Cho, Y. S., S. Challa, D. Moquin, R. Genga, T. D. Ray, M. Guildford and F. K. Chan (2009). "Phosphorylation-driven assembly of the RIP1-RIP3 complex regulates programmed necrosis and virus-induced inflammation." Cell 137(6): 1112-1123.
Clarke, P. G. (1990). "Developmental cell death: morphological diversity and multiple mechanisms." Anat Embryol (Berl) 181(3): 195-213.
Daish, T. J., K. Mills and S. Kumar (2004). "Drosophila caspase DRONC is required for specific developmental cell death pathways and stress-induced apoptosis." Dev Cell 7(6): 909-915.
Damke, H., T. Baba, D. E. Warnock and S. L. Schmid (1994). "Induction of mutant dynamin specifically blocks endocytic coated vesicle formation." J Cell Biol 127(4): 915-934.
Delbridge, A. R., L. J. Valente and A. Strasser (2012). "The role of the apoptotic machinery in tumor suppression." Cold Spring Harb Perspect Biol 4(11).
den Haan, J. M., S. M. Lehar and M. J. Bevan (2000). "CD8(+) but not CD8(-) dendritic cells cross-prime cytotoxic T cells in vivo." J Exp Med 192(12): 1685-1696.
Denton, D., S. Nicolson and S. Kumar (2012). "Cell death by autophagy: facts and apparent artefacts." Cell Death Differ 19(1): 87-95.
Devitt, A., O. D. Moffatt, C. Raykundalia, J. D. Capra, D. L. Simmons and C. D. Gregory (1998). "Human CD14 mediates recognition and phagocytosis of apoptotic cells." Nature 392(6675): 505-509.
Dini, L., E. C. Carla, M. Deluca, G. Faraldi and G. Tagliafierro (1999). "Phagocytosis of apoptotic cells: liver recognition and molecular mechanisms." Ital. J. Zool. 66: 317-322.
Distel, L. V., R. Fickenscher, K. Dietel, A. Hung, H. Iro, J. Zenk, E. Nkenke, M. Buttner, G. Niedobitek and G. G. Grabenbauer (2009). "Tumour infiltrating lymphocytes in squamous cell carcinoma of the oro- and hypopharynx: prognostic impact may depend on type of treatment and stage of disease." Oral Oncol 45(10): e167-174.
Don, M. M., G. Ablett, C. J. Bishop, P. G. Bundesen, K. J. Donald, J. Searle and J. F. Kerr (1977). "Death of cells by apoptosis following attachment of specifically allergized lymphocytes in vitro." Aust J Exp Biol Med Sci 55(4): 407-417.
Dunn, G. P., A. T. Bruce, H. Ikeda, L. J. Old and R. D. Schreiber (2002). "Cancer immunoediting: from immunosurveillance to tumor escape." Nat Immunol 3(11): 991-998.
Dvorak, A. M. and D. Feng (2001). "The vesiculo-vacuolar organelle (VVO). A new endothelial cell permeability organelle." J Histochem Cytochem 49(4): 419-432.
Elmore, S. (2007). "Apoptosis: a review of programmed cell death." Toxicol Pathol 35(4): 495-516.
Erlich, S., E. Shohami and R. Pinkas-Kramarski (2006). "Neurodegeneration induces upregulation of Beclin 1." Autophagy 2(1): 49-51.

120
Fadok, V. A., D. L. Bratton, S. C. Frasch, M. L. Warner and P. M. Henson (1998). "The role of phosphatidylserine in recognition of apoptotic cells by phagocytes." Cell Death Differ 5(7): 551-562.
Fadok, V. A., D. R. Voelker, P. A. Campbell, J. J. Cohen, D. L. Bratton and P. M. Henson (1992). "Exposure of phosphatidylserine on the surface of apoptotic lymphocytes triggers specific recognition and removal by macrophages." J Immunol 148(7): 2207-2216.
Fadok, V. A., M. L. Warner, D. L. Bratton and P. M. Henson (1998). "CD36 is required for phagocytosis of apoptotic cells by human macrophages that use either a phosphatidylserine receptor or the vitronectin receptor (alpha v beta 3)." J Immunol 161(11): 6250-6257.
Fais, S. (2007). "Cannibalism: a way to feed on metastatic tumors." Cancer Lett 258(2): 155-164.
Fazi, B., W. Bursch, G. M. Fimia, R. Nardacci, M. Piacentini, F. Di Sano and L. Piredda (2008). "Fenretinide induces autophagic cell death in caspase-defective breast cancer cells." Autophagy 4(4): 435-441.
Feng, D., J. A. Nagy, K. Pyne, H. F. Dvorak and A. M. Dvorak (1998). "Neutrophils emigrate from venules by a transendothelial cell pathway in response to FMLP." J Exp Med 187(6): 903-915.
Flora, P. K. and C. D. Gregory (1994). "Recognition of apoptotic cells by human macrophages: inhibition by a monocyte/macrophage-specific monoclonal antibody." Eur J Immunol 24(11): 2625-2632.
Florey, O., S. E. Kim, C. P. Sandoval, C. M. Haynes and M. Overholtzer (2011). "Autophagy machinery mediates macroendocytic processing and entotic cell death by targeting single membranes." Nat Cell Biol 13(11): 1335-1343.
Frisch, S. M. and H. Francis (1994). "Disruption of epithelial cell-matrix interactions induces apoptosis." J Cell Biol 124(4): 619-626.
Fuchs, Y. and H. Steller (2011). "Programmed cell death in animal development and disease." Cell 147(4): 742-758.
Galluzzi, L., J. M. Bravo-San Pedro, I. Vitale, S. A. Aaronson, J. M. Abrams, D. Adam, E. S. Alnemri, L. Altucci, D. Andrews, M. Annicchiarico-Petruzzelli, E. H. Baehrecke, N. G. Bazan, M. J. Bertrand, K. Bianchi, M. V. Blagosklonny, K. Blomgren, C. Borner, D. E. Bredesen, C. Brenner, M. Campanella, E. Candi, F. Cecconi, F. K. Chan, N. S. Chandel, E. H. Cheng, J. E. Chipuk, J. A. Cidlowski, A. Ciechanover, T. M. Dawson, V. L. Dawson, V. De Laurenzi, R. De Maria, K. M. Debatin, N. Di Daniele, V. M. Dixit, B. D. Dynlacht, W. S. El-Deiry, G. M. Fimia, R. A. Flavell, S. Fulda, C. Garrido, M. L. Gougeon, D. R. Green, H. Gronemeyer, G. Hajnoczky, J. M. Hardwick, M. O. Hengartner, H. Ichijo, B. Joseph, P. J. Jost, T. Kaufmann, O. Kepp, D. J. Klionsky, R. A. Knight, S. Kumar, J. J. Lemasters, B. Levine, A. Linkermann, S. A. Lipton, R. A. Lockshin, C. Lopez-Otin, E. Lugli, F. Madeo, W. Malorni, J. C. Marine, S. J. Martin, J. C. Martinou, J. P. Medema, P. Meier, S. Melino, N. Mizushima, U. Moll, C. Munoz-Pinedo, G. Nunez, A. Oberst, T. Panaretakis, J. M. Penninger, M. E. Peter, M. Piacentini, P. Pinton, J. H. Prehn, H. Puthalakath, G. A. Rabinovich, K. S. Ravichandran, R. Rizzuto, C. M. Rodrigues, D. C. Rubinsztein, T. Rudel, Y. Shi, H. U. Simon, B. R. Stockwell, G. Szabadkai, S. W. Tait, H. L. Tang, N. Tavernarakis, Y. Tsujimoto, T. Vanden Berghe, P. Vandenabeele, A. Villunger, E. F. Wagner, H. Walczak, E. White, W. G. Wood, J. Yuan, Z. Zakeri, B. Zhivotovsky, G. Melino and G.

121
Kroemer (2015). "Essential versus accessory aspects of cell death: recommendations of the NCCD 2015." Cell Death Differ 22(1): 58-73.
Galluzzi, L., O. Kepp, S. Krautwald, G. Kroemer and A. Linkermann (2014). "Molecular mechanisms of regulated necrosis." Semin Cell Dev Biol 35: 24-32.
Galluzzi, L., M. C. Maiuri, I. Vitale, H. Zischka, M. Castedo, L. Zitvogel and G. Kroemer (2007). "Cell death modalities: classification and pathophysiological implications." Cell Death Differ 14(7): 1237-1243.
Galluzzi, L., I. Vitale, J. M. Abrams, E. S. Alnemri, E. H. Baehrecke, M. V. Blagosklonny, T. M. Dawson, V. L. Dawson, W. S. El-Deiry, S. Fulda, E. Gottlieb, D. R. Green, M. O. Hengartner, O. Kepp, R. A. Knight, S. Kumar, S. A. Lipton, X. Lu, F. Madeo, W. Malorni, P. Mehlen, G. Nunez, M. E. Peter, M. Piacentini, D. C. Rubinsztein, Y. Shi, H. U. Simon, P. Vandenabeele, E. White, J. Yuan, B. Zhivotovsky, G. Melino and G. Kroemer (2012). "Molecular definitions of cell death subroutines: recommendations of the Nomenclature Committee on Cell Death 2012." Cell Death Differ 19(1): 107-120.
Geiger, B. and O. Ayalon (1992). "Cadherins." Annu Rev Cell Biol 8: 307-332.
Goldstein, J. L., R. G. Anderson and M. S. Brown (1979). "Coated pits, coated vesicles, and receptor-mediated endocytosis." Nature 279(5715): 679-685.
Grabenbauer, G. G., G. Lahmer, L. Distel and G. Niedobitek (2006). "Tumor-infiltrating cytotoxic T cells but not regulatory T cells predict outcome in anal squamous cell carcinoma." Clin Cancer Res 12(11 Pt 1): 3355-3360.
Green, D. R. and G. Kroemer (2005). "Pharmacological manipulation of cell death: clinical applications in sight?" J Clin Invest 115(10): 2610-2617.
Gregory, C. (2009). "Cell biology: Sent by the scent of death." Nature 461(7261): 181-182.
Grimsley, C. and K. S. Ravichandran (2003). "Cues for apoptotic cell engulfment: eat-me, don't eat-me and come-get-me signals." Trends Cell Biol 13(12): 648-656.
Gupta, K. and P. Dey (2003). "Cell cannibalism: diagnostic marker of malignancy." Diagn Cytopathol 28(2): 86-87.
Hartel, S., M. Zorn-Kruppa, S. Tykhonova, P. Alajuuma, M. Engelke and H. A. Diehl (2003). "Staurosporine-induced apoptosis in human cornea epithelial cells in vitro." Cytometry A 55(1): 15-23.
Hartl, F. U. (1996). "Molecular chaperones in cellular protein folding." Nature 381(6583): 571-579.
He, S., L. Wang, L. Miao, T. Wang, F. Du, L. Zhao and X. Wang (2009). "Receptor interacting protein kinase-3 determines cellular necrotic response to TNF-alpha." Cell 137(6): 1100-1111.
Herrera, E. M., M. Ming, E. Ortega-Barria and M. E. Pereira (1994). "Mediation of Trypanosoma cruzi invasion by heparan sulfate receptors on host cells and penetrin counter-receptors on the trypanosomes." Mol Biochem Parasitol 65(1): 73-83.
Hightower, L. E. (1991). "Heat shock, stress proteins, chaperones, and proteotoxicity." Cell 66(2): 191-197.

122
Hinshaw, J. E. (2000). "Dynamin and its role in membrane fission." Annu Rev Cell Dev Biol 16: 483-519.
Hirt, U. A. and M. Leist (2003). "Rapid, noninflammatory and PS-dependent phagocytic clearance of necrotic cells." Cell Death Differ 10(10): 1156-1164.
Hodge, J. W., C. Guha, J. Neefjes and J. L. Gulley (2008). "Synergizing radiation therapy and immunotherapy for curing incurable cancers. Opportunities and challenges." Oncology (Williston Park) 22(9): 1064-1070; discussion 1075, 1080-1061, 1084.
Holmgren, L., A. Szeles, E. Rajnavolgyi, J. Folkman, G. Klein, I. Ernberg and K. I. Falk (1999). "Horizontal transfer of DNA by the uptake of apoptotic bodies." Blood 93(11): 3956-3963.
Huang, A. Y., P. Golumbek, M. Ahmadzadeh, E. Jaffee, D. Pardoll and H. Levitsky (1994). "Role of bone marrow-derived cells in presenting MHC class I-restricted tumor antigens." Science 264(5161): 961-965.
Humble, J. G., W. H. Jayne and R. J. Pulvertaft (1956). "Biological interaction between lymphocytes and other cells." Br J Haematol 2(3): 283-294.
Ichimura, T., E. J. Asseldonk, B. D. Humphreys, L. Gunaratnam, J. S. Duffield and J. V. Bonventre (2008). "Kidney injury molecule-1 is a phosphatidylserine receptor that confers a phagocytic phenotype on epithelial cells." J Clin Invest 118(5): 1657-1668.
Imai, K., S. Hirata, A. Irie, S. Senju, Y. Ikuta, K. Yokomine, M. Harao, M. Inoue, T. Tsunoda, S. Nakatsuru, H. Nakagawa, Y. Nakamura, H. Baba and Y. Nishimura (2008). "Identification of a novel tumor-associated antigen, cadherin 3/P-cadherin, as a possible target for immunotherapy of pancreatic, gastric, and colorectal cancers." Clin Cancer Res 14(20): 6487-6495.
Jessel, R., S. Haertel, C. Socaciu, S. Tykhonova and H. A. Diehl (2002). "Kinetics of apoptotic markers in exogeneously induced apoptosis of EL4 cells." J Cell Mol Med 6(1): 82-92.
Johnstone, R. M., M. Adam, J. R. Hammond, L. Orr and C. Turbide (1987). "Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes)." J Biol Chem 262(19): 9412-9420.
Kaczmarek, A., P. Vandenabeele and D. V. Krysko (2013). "Necroptosis: the release of damage-associated molecular patterns and its physiological relevance." Immunity 38(2): 209-223.
Kaiser, W. J., J. W. Upton, A. B. Long, D. Livingston-Rosanoff, L. P. Daley-Bauer, R. Hakem, T. Caspary and E. S. Mocarski (2011). "RIP3 mediates the embryonic lethality of caspase-8-deficient mice." Nature 471(7338): 368-372.
Kerr, J. F., A. H. Wyllie and A. R. Currie (1972). "Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics." Br J Cancer 26(4): 239-257.
Kinchen, J. M., K. Doukoumetzidis, J. Almendinger, L. Stergiou, A. Tosello-Trampont, C. D. Sifri, M. O. Hengartner and K. S. Ravichandran (2008). "A pathway for phagosome maturation during engulfment of apoptotic cells." Nat Cell Biol 10(5): 556-566.

123
Kobayashi, N., P. Karisola, V. Pena-Cruz, D. M. Dorfman, M. Jinushi, S. E. Umetsu, M. J. Butte, H. Nagumo, I. Chernova, B. Zhu, A. H. Sharpe, S. Ito, G. Dranoff, G. G. Kaplan, J. M. Casasnovas, D. T. Umetsu, R. H. Dekruyff and G. J. Freeman (2007). "TIM-1 and TIM-4 glycoproteins bind phosphatidylserine and mediate uptake of apoptotic cells." Immunity 27(6): 927-940.
Kojima, S., H. Sekine, I. Fukui and H. Ohshima (1998). "Clinical significance of "cannibalism" in urinary cytology of bladder cancer." Acta Cytol 42(6): 1365-1369.
Kolb, S., R. Vranckx, M. G. Huisse, J. B. Michel and O. Meilhac (2007). "The phosphatidylserine receptor mediates phagocytosis by vascular smooth muscle cells." J Pathol 212(3): 249-259.
Krajcovic, M., N. B. Johnson, Q. Sun, G. Normand, N. Hoover, E. Yao, A. L. Richardson, R. W. King, E. S. Cibas, S. J. Schnitt, J. S. Brugge and M. Overholtzer (2011). "A non-genetic route to aneuploidy in human cancers." Nat Cell Biol 13(3): 324-330.
Kroemer, G., L. Galluzzi, P. Vandenabeele, J. Abrams, E. S. Alnemri, E. H. Baehrecke, M. V. Blagosklonny, W. S. El-Deiry, P. Golstein, D. R. Green, M. Hengartner, R. A. Knight, S. Kumar, S. A. Lipton, W. Malorni, G. Nunez, M. E. Peter, J. Tschopp, J. Yuan, M. Piacentini, B. Zhivotovsky, G. Melino and D. Nomenclature Committee on Cell (2009). "Classification of cell death: recommendations of the Nomenclature Committee on Cell Death 2009." Cell Death Differ 16(1): 3-11.
Krysko, D. V., A. D. Garg, A. Kaczmarek, O. Krysko, P. Agostinis and P. Vandenabeele (2012). "Immunogenic cell death and DAMPs in cancer therapy." Nat Rev Cancer 12(12): 860-875.
Kumar, P. V., M. Hosseinzadeh and G. R. Bedayat (2001). "Cytologic findings of medulloblastoma in crush smears." Acta Cytol 45(4): 542-546.
Leist, M., B. Single, A. F. Castoldi, S. Kuhnle and P. Nicotera (1997). "Intracellular adenosine triphosphate (ATP) concentration: a switch in the decision between apoptosis and necrosis." J Exp Med 185(8): 1481-1486.
Lettre, G. and M. O. Hengartner (2006). "Developmental apoptosis in C. elegans: a complex CEDnario." Nat Rev Mol Cell Biol 7(2): 97-108.
Long, J. S. and K. M. Ryan (2012). "New frontiers in promoting tumour cell death: targeting apoptosis, necroptosis and autophagy." Oncogene 31(49): 5045-5060.
Luciani, M. F. and G. Chimini (1996). "The ATP binding cassette transporter ABC1, is required for the engulfment of corpses generated by apoptotic cell death." EMBO J 15(2): 226-235.
Lugini, L., F. Lozupone, P. Matarrese, C. Funaro, F. Luciani, W. Malorni, L. Rivoltini, C. Castelli, A. Tinari, A. Piris, G. Parmiani and S. Fais (2003). "Potent phagocytic activity discriminates metastatic and primary human malignant melanomas: a key role of ezrin." Lab Invest 83(11): 1555-1567.
Lugini, L., P. Matarrese, A. Tinari, F. Lozupone, C. Federici, E. Iessi, M. Gentile, F. Luciani, G. Parmiani, L. Rivoltini, W. Malorni and S. Fais (2006). "Cannibalism of live lymphocytes by human metastatic but not primary melanoma cells." Cancer Res 66(7): 3629-3638.

124
Luzio, J. P., B. A. Rous, N. A. Bright, P. R. Pryor, B. M. Mullock and R. C. Piper (2000). "Lysosome-endosome fusion and lysosome biogenesis." J Cell Sci 113 ( Pt 9): 1515-1524.
MacFarlane, M. (2009). "Cell death pathways--potential therapeutic targets." Xenobiotica 39(8): 616-624.
Matsui, T., M. Maeda, Y. Doi, S. Yonemura, M. Amano, K. Kaibuchi, S. Tsukita and S. Tsukita (1998). "Rho-kinase phosphorylates COOH-terminal threonines of ezrin/radixin/moesin (ERM) proteins and regulates their head-to-tail association." J Cell Biol 140(3): 647-657.
Matzinger, P. (2002). "The danger model: a renewed sense of self." Science 296(5566): 301-305.
McIlroy, D., M. Tanaka, H. Sakahira, H. Fukuyama, M. Suzuki, K. Yamamura, Y. Ohsawa, Y. Uchiyama and S. Nagata (2000). "An auxiliary mode of apoptotic DNA fragmentation provided by phagocytes." Genes Dev 14(5): 549-558.
Medicine, U. S. N. L. o. (2014). " http://www.nlm.nih.gov/cgi/mesh/2014/MB_cgi?mode=&index=25547&field=all&HM=&II=&PA=&form=&input." Medical Subject Headings Bethesda (MD) National Library of Medicine (US)(Accessed 2014 Sept 20).
Mehlen, P. and D. E. Bredesen (2011). "Dependence receptors: from basic research to drug development." Sci Signal 4(157): mr2.
Meredith, J. E., Jr., B. Fazeli and M. A. Schwartz (1993). "The extracellular matrix as a cell survival factor." Mol Biol Cell 4(9): 953-961.
Niessen, C. M. and B. M. Gumbiner (2002). "Cadherin-mediated cell sorting not determined by binding or adhesion specificity." J Cell Biol 156(2): 389-399.
Obeid, M., A. Tesniere, F. Ghiringhelli, G. M. Fimia, L. Apetoh, J. L. Perfettini, M. Castedo, G. Mignot, T. Panaretakis, N. Casares, D. Metivier, N. Larochette, P. van Endert, F. Ciccosanti, M. Piacentini, L. Zitvogel and G. Kroemer (2007). "Calreticulin exposure dictates the immunogenicity of cancer cell death." Nat Med 13(1): 54-61.
Omelchenko, T., E. Fetisova, O. Ivanova, E. M. Bonder, H. Feder, J. M. Vasiliev and I. M. Gelfand (2001). "Contact interactions between epitheliocytes and fibroblasts: formation of heterotypic cadherin-containing adhesion sites is accompanied by local cytoskeletal reorganization." Proc Natl Acad Sci U S A 98(15): 8632-8637.
Orth, J. D. and M. A. McNiven (2003). "Dynamin at the actin-membrane interface." Curr Opin Cell Biol 15(1): 31-39.
Overholtzer, M., A. A. Mailleux, G. Mouneimne, G. Normand, S. J. Schnitt, R. W. King, E. S. Cibas and J. S. Brugge (2007). "A nonapoptotic cell death process, entosis, that occurs by cell-in-cell invasion." Cell 131(5): 966-979.
Pearse, B. M. (1976). "Clathrin: a unique protein associated with intracellular transfer of membrane by coated vesicles." Proc Natl Acad Sci U S A 73(4): 1255-1259.
Petrovski, G., G. Zahuczky, K. Katona, G. Vereb, W. Martinet, Z. Nemes, W. Bursch and L. Fesus (2007). "Clearance of dying autophagic cells of different origin by professional and non-professional phagocytes." Cell Death Differ 14(6): 1117-1128.

125
Platt, N., H. Suzuki, Y. Kurihara, T. Kodama and S. Gordon (1996). "Role for the class A macrophage scavenger receptor in the phagocytosis of apoptotic thymocytes in vitro." Proc Natl Acad Sci U S A 93(22): 12456-12460.
Pollard, T. D. and J. A. Cooper (2009). "Actin, a central player in cell shape and movement." Science 326(5957): 1208-1212.
Pretscher, D., L. V. Distel, G. G. Grabenbauer, M. Wittlinger, M. Buettner and G. Niedobitek (2009). "Distribution of immune cells in head and neck cancer: CD8+ T-cells and CD20+ B-cells in metastatic lymph nodes are associated with favourable outcome in patients with oro- and hypopharyngeal carcinoma." BMC Cancer 9: 292.
Ramprasad, M. P., W. Fischer, J. L. Witztum, G. R. Sambrano, O. Quehenberger and D. Steinberg (1995). "The 94- to 97-kDa mouse macrophage membrane protein that recognizes oxidized low density lipoprotein and phosphatidylserine-rich liposomes is identical to macrosialin, the mouse homologue of human CD68." Proc Natl Acad Sci U S A 92(21): 9580-9584.
Reed, J. C. (2006). "Drug insight: cancer therapy strategies based on restoration of endogenous cell death mechanisms." Nat Clin Pract Oncol 3(7): 388-398.
Reginato, M. J., K. R. Mills, J. K. Paulus, D. K. Lynch, D. C. Sgroi, J. Debnath, S. K. Muthuswamy and J. S. Brugge (2003). "Integrins and EGFR coordinately regulate the pro-apoptotic protein Bim to prevent anoikis." Nat Cell Biol 5(8): 733-740.
Rink, J., E. Ghigo, Y. Kalaidzidis and M. Zerial (2005). "Rab conversion as a mechanism of progression from early to late endosomes." Cell 122(5): 735-749.
Samara, C., P. Syntichaki and N. Tavernarakis (2008). "Autophagy is required for necrotic cell death in Caenorhabditis elegans." Cell Death Differ 15(1): 105-112.
Sambrano, G. R. and D. Steinberg (1995). "Recognition of oxidatively damaged and apoptotic cells by an oxidized low density lipoprotein receptor on mouse peritoneal macrophages: role of membrane phosphatidylserine." Proc Natl Acad Sci U S A 92(5): 1396-1400.
Savill, J. and V. Fadok (2000). "Corpse clearance defines the meaning of cell death." Nature 407(6805): 784-788.
Savill, J., N. Hogg, Y. Ren and C. Haslett (1992). "Thrombospondin cooperates with CD36 and the vitronectin receptor in macrophage recognition of neutrophils undergoing apoptosis." J Clin Invest 90(4): 1513-1522.
Schutze, S., V. Tchikov and W. Schneider-Brachert (2008). "Regulation of TNFR1 and CD95 signalling by receptor compartmentalization." Nat Rev Mol Cell Biol 9(8): 655-662.
Schwegler, M., A. M. Wirsing, A. J. Dollinger, B. Abendroth, F. Putz, R. Fietkau and L. V. Distel (2015). "Clearance of primary necrotic cells by non-professional phagocytes." Biol Cell 107(10): 372-387.
Schwegler, M., A. M. Wirsing, H. M. Schenker, L. Ott, J. M. Ries, M. Büttner-Herold, R. Fietkau, F. Putz and L. V. Distel "Prognostic Value of Homotypic Cell Internalization by Nonprofessional Phagocytic Cancer Cells." BioMed Research International(Article ID 359392, in press).

126
Shibata, M., T. Lu, T. Furuya, A. Degterev, N. Mizushima, T. Yoshimori, M. MacDonald, B. Yankner and J. Yuan (2006). "Regulation of intracellular accumulation of mutant Huntingtin by Beclin 1." J Biol Chem 281(20): 14474-14485.
Silva, M. T., A. do Vale and N. M. dos Santos (2008). "Secondary necrosis in multicellular animals: an outcome of apoptosis with pathogenic implications." Apoptosis 13(4): 463-482.
Singh, S., V. D'Mello, P. van Bergen en Henegouwen and R. B. Birge (2007). "A NPxY-independent beta5 integrin activation signal regulates phagocytosis of apoptotic cells." Biochem Biophys Res Commun 364(3): 540-548.
Slater, A., A. C. Cato, G. M. Sillar, J. Kioussis and R. H. Burdon (1981). "The pattern of protein synthesis induced by heat shock of HeLa cells." Eur J Biochem 117(2): 341-346.
Sontag, J. M., E. M. Fykse, Y. Ushkaryov, J. P. Liu, P. J. Robinson and T. C. Sudhof (1994). "Differential expression and regulation of multiple dynamins." J Biol Chem 269(6): 4547-4554.
Stinchcombe, J. C. and G. M. Griffiths (2007). "Secretory mechanisms in cell-mediated cytotoxicity." Annu Rev Cell Dev Biol 23: 495-517.
Stuart, L. M. and R. A. Ezekowitz (2005). "Phagocytosis: elegant complexity." Immunity 22(5): 539-550.
Sun, L., H. Wang, Z. Wang, S. He, S. Chen, D. Liao, L. Wang, J. Yan, W. Liu, X. Lei and X. Wang (2012). "Mixed lineage kinase domain-like protein mediates necrosis signaling downstream of RIP3 kinase." Cell 148(1-2): 213-227.
Suto, R. and P. K. Srivastava (1995). "A mechanism for the specific immunogenicity of heat shock protein-chaperoned peptides." Science 269(5230): 1585-1588.
Tabachnyk, M., L. V. Distel, M. Buttner, G. G. Grabenbauer, E. Nkenke, R. Fietkau and D. Lubgan (2012). "Radiochemotherapy induces a favourable tumour infiltrating inflammatory cell profile in head and neck cancer." Oral Oncol 48(7): 594-601.
Takeichi, M. (1991). "Cadherin cell adhesion receptors as a morphogenetic regulator." Science 251(5000): 1451-1455.
Takeuchi, M., T. Inoue, T. Otani, F. Yamasaki, S. Nakamura and M. Kibata (2010). "Cell-in-cell structures formed between human cancer cell lines and the cytotoxic regulatory T-cell line HOZOT." J Mol Cell Biol 2(3): 139-151.
Taylor, R. C., S. P. Cullen and S. J. Martin (2008). "Apoptosis: controlled demolition at the cellular level." Nat Rev Mol Cell Biol 9(3): 231-241.
Vesely, M. D., M. H. Kershaw, R. D. Schreiber and M. J. Smyth (2011). "Natural innate and adaptive immunity to cancer." Annu Rev Immunol 29: 235-271.
Voll, R. E., M. Herrmann, E. A. Roth, C. Stach, J. R. Kalden and I. Girkontaite (1997). "Immunosuppressive effects of apoptotic cells." Nature 390(6658): 350-351.
Wajant, H. (2002). "The Fas signaling pathway: more than a paradigm." Science 296(5573): 1635-1636.

127
Wang, S., Z. Guo, P. Xia, T. Liu, J. Wang, S. Li, L. Sun, J. Lu, Q. Wen, M. Zhou, L. Ma, X. Ding, X. Wang and X. Yao (2009). "Internalization of NK cells into tumor cells requires ezrin and leads to programmed cell-in-cell death." Cell Res 19(12): 1350-1362.
Wang, X. L., W (1987). "Mechanisms of natural killer cell-mediated tumor cell cytolysis at a single cell level." J Med Coll PLA 2: 107-117.
Wang, Y. and X. N. Wang (2013). "Cell-in-cell: A virgin land of cell biology." Oncoimmunology 2(10): e25988.
Whittard, J. D., S. E. Craig, A. P. Mould, A. Koch, O. Pertz, J. Engel and M. J. Humphries (2002). "E-cadherin is a ligand for integrin alpha2beta1." Matrix Biol 21(6): 525-532.
Wickman, G. R., L. Julian, K. Mardilovich, S. Schumacher, J. Munro, N. Rath, S. A. Zander, A. Mleczak, D. Sumpton, N. Morrice, W. V. Bienvenut and M. F. Olson (2013). "Blebs produced by actin-myosin contraction during apoptosis release damage-associated molecular pattern proteins before secondary necrosis occurs." Cell Death Differ 20(10): 1293-1305.
Won, W. J., M. F. Bachmann and J. F. Kearney (2008). "CD36 is differentially expressed on B cell subsets during development and in responses to antigen." J Immunol 180(1): 230-237.
Wood, W., M. Turmaine, R. Weber, V. Camp, R. A. Maki, S. R. McKercher and P. Martin (2000). "Mesenchymal cells engulf and clear apoptotic footplate cells in macrophageless PU.1 null mouse embryos." Development 127(24): 5245-5252.
Yamada, H., T. Abe, S. A. Li, Y. Masuoka, M. Isoda, M. Watanabe, Y. Nasu, H. Kumon, A. Asai and K. Takei (2009). "Dynasore, a dynamin inhibitor, suppresses lamellipodia formation and cancer cell invasion by destabilizing actin filaments." Biochem Biophys Res Commun 390(4): 1142-1148.
Yu, L., A. Alva, H. Su, P. Dutt, E. Freundt, S. Welsh, E. H. Baehrecke and M. J. Lenardo (2004). "Regulation of an ATG7-beclin 1 program of autophagic cell death by caspase-8." Science 304(5676): 1500-1502.
Yuan, J. Y. and H. R. Horvitz (1990). "The Caenorhabditis elegans genes ced-3 and ced-4 act cell autonomously to cause programmed cell death." Dev Biol 138(1): 33-41.
Zhang, D. W., J. Shao, J. Lin, N. Zhang, B. J. Lu, S. C. Lin, M. Q. Dong and J. Han (2009). "RIP3, an energy metabolism regulator that switches TNF-induced cell death from apoptosis to necrosis." Science 325(5938): 332-336.
Zhao, J., S. Jitkaew, Z. Cai, S. Choksi, Q. Li, J. Luo and Z. G. Liu (2012). "Mixed lineage kinase domain-like is a key receptor interacting protein 3 downstream component of TNF-induced necrosis." Proc Natl Acad Sci U S A 109(14): 5322-5327.
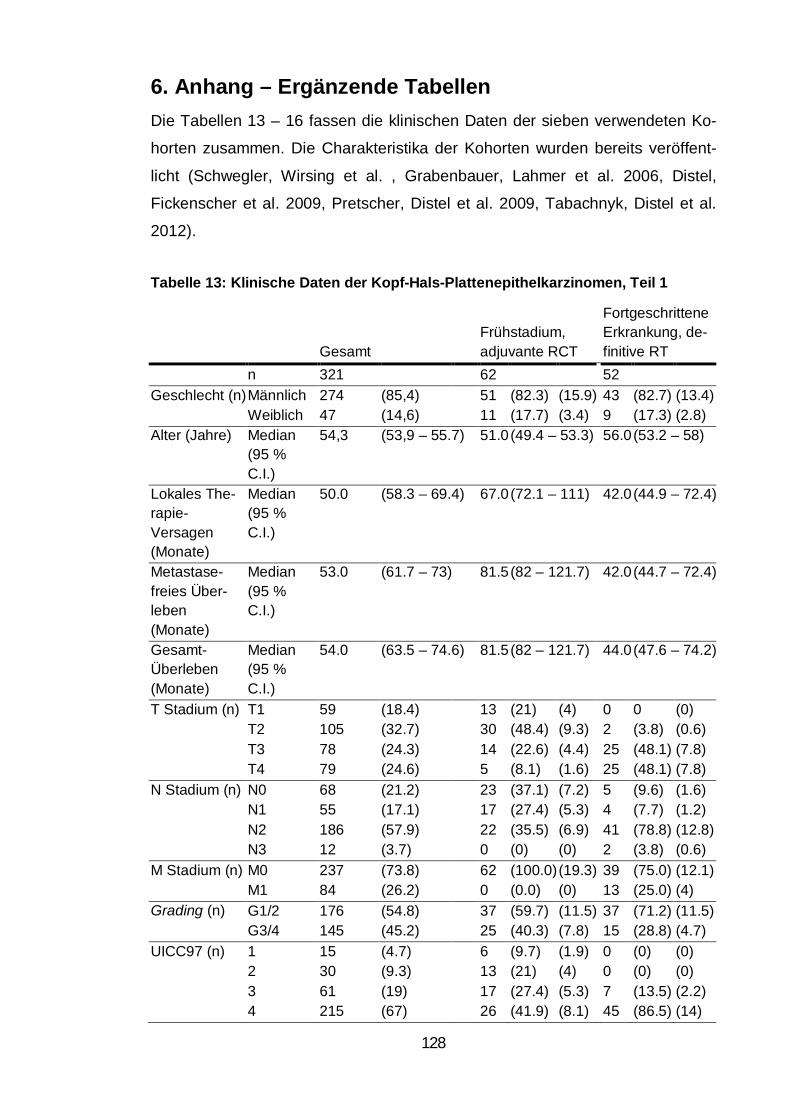
128
6. Anhang – Ergänzende Tabellen Die Tabellen 13 – 16 fassen die klinischen Daten der sieben verwendeten Ko-
horten zusammen. Die Charakteristika der Kohorten wurden bereits veröffent-
licht (Schwegler, Wirsing et al. , Grabenbauer, Lahmer et al. 2006, Distel,
Fickenscher et al. 2009, Pretscher, Distel et al. 2009, Tabachnyk, Distel et al.
2012). Tabelle 13: Klinische Daten der Kopf-Hals-Plattenepithelkarzinomen, Teil 1
Gesamt Frühstadium, adjuvante RCT
Fortgeschrittene Erkrankung, de-finitive RT
n 321 62 52 Geschlecht (n) Männlich 274 (85,4) 51 (82.3) (15.9) 43 (82.7) (13.4)
Weiblich 47 (14,6) 11 (17.7) (3.4) 9 (17.3) (2.8) Alter (Jahre) Median
(95 % C.I.)
54,3 (53,9 – 55.7) 51.0(49.4 – 53.3) 56.0(53.2 – 58)
Lokales The-rapie-Versagen (Monate)
Median (95 % C.I.)
50.0 (58.3 – 69.4) 67.0(72.1 – 111) 42.0(44.9 – 72.4)
Metastase-freies Über-leben (Monate)
Median (95 % C.I.)
53.0 (61.7 – 73) 81.5(82 – 121.7) 42.0(44.7 – 72.4)
Gesamt-Überleben (Monate)
Median (95 % C.I.)
54.0 (63.5 – 74.6) 81.5(82 – 121.7) 44.0(47.6 – 74.2)
T Stadium (n) T1 59 (18.4) 13 (21) (4) 0 0 (0) T2 105 (32.7) 30 (48.4) (9.3) 2 (3.8) (0.6) T3 78 (24.3) 14 (22.6) (4.4) 25 (48.1) (7.8) T4 79 (24.6) 5 (8.1) (1.6) 25 (48.1) (7.8)
N Stadium (n) N0 68 (21.2) 23 (37.1) (7.2) 5 (9.6) (1.6) N1 55 (17.1) 17 (27.4) (5.3) 4 (7.7) (1.2) N2 186 (57.9) 22 (35.5) (6.9) 41 (78.8) (12.8) N3 12 (3.7) 0 (0) (0) 2 (3.8) (0.6)
M Stadium (n) M0 237 (73.8) 62 (100.0)(19.3) 39 (75.0) (12.1) M1 84 (26.2) 0 (0.0) (0) 13 (25.0) (4)
Grading (n) G1/2 176 (54.8) 37 (59.7) (11.5) 37 (71.2) (11.5) G3/4 145 (45.2) 25 (40.3) (7.8) 15 (28.8) (4.7)
UICC97 (n) 1 15 (4.7) 6 (9.7) (1.9) 0 (0) (0) 2 30 (9.3) 13 (21) (4) 0 (0) (0) 3 61 (19) 17 (27.4) (5.3) 7 (13.5) (2.2) 4 215 (67) 26 (41.9) (8.1) 45 (86.5) (14)
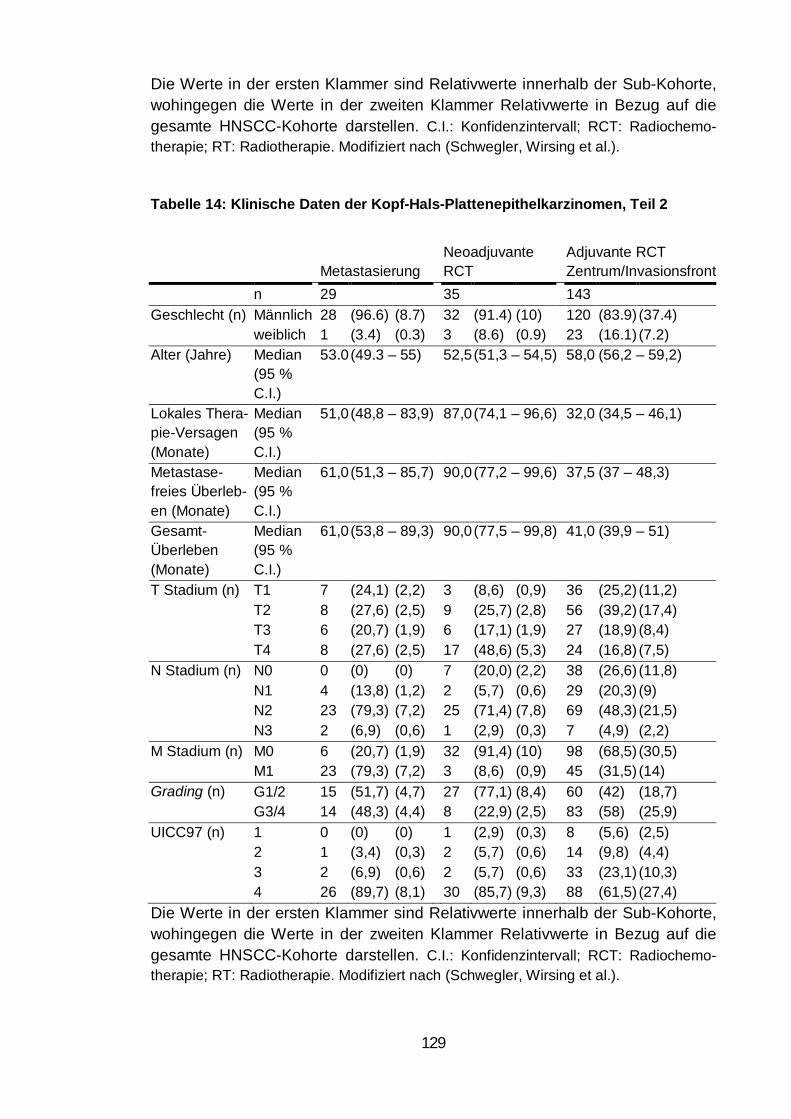
129
Die Werte in der ersten Klammer sind Relativwerte innerhalb der Sub-Kohorte, wohingegen die Werte in der zweiten Klammer Relativwerte in Bezug auf die gesamte HNSCC-Kohorte darstellen. C.I.: Konfidenzintervall; RCT: Radiochemo-therapie; RT: Radiotherapie. Modifiziert nach (Schwegler, Wirsing et al.). Tabelle 14: Klinische Daten der Kopf-Hals-Plattenepithelkarzinomen, Teil 2
Metastasierung Neoadjuvante RCT
Adjuvante RCT Zentrum/Invasionsfront
n 29 35 143 Geschlecht (n) Männlich 28 (96.6) (8.7) 32 (91.4) (10) 120 (83.9) (37.4)
weiblich 1 (3.4) (0.3) 3 (8.6) (0.9) 23 (16.1) (7.2) Alter (Jahre) Median
(95 % C.I.)
53.0(49.3 – 55) 52,5(51,3 – 54,5) 58,0 (56,2 – 59,2)
Lokales Thera-pie-Versagen (Monate)
Median (95 % C.I.)
51,0(48,8 – 83,9) 87,0(74,1 – 96,6) 32,0 (34,5 – 46,1)
Metastase-freies Überleb-en (Monate)
Median (95 % C.I.)
61,0(51,3 – 85,7) 90,0(77,2 – 99,6) 37,5 (37 – 48,3)
Gesamt-Überleben (Monate)
Median (95 % C.I.)
61,0(53,8 – 89,3) 90,0(77,5 – 99,8) 41,0 (39,9 – 51)
T Stadium (n) T1 7 (24,1) (2,2) 3 (8,6) (0,9) 36 (25,2) (11,2) T2 8 (27,6) (2,5) 9 (25,7) (2,8) 56 (39,2) (17,4) T3 6 (20,7) (1,9) 6 (17,1) (1,9) 27 (18,9) (8,4)
T4 8 (27,6) (2,5) 17 (48,6) (5,3) 24 (16,8) (7,5) N Stadium (n) N0 0 (0) (0) 7 (20,0) (2,2) 38 (26,6) (11,8)
N1 4 (13,8) (1,2) 2 (5,7) (0,6) 29 (20,3) (9) N2 23 (79,3) (7,2) 25 (71,4) (7,8) 69 (48,3) (21,5) N3 2 (6,9) (0,6) 1 (2,9) (0,3) 7 (4,9) (2,2)
M Stadium (n) M0 6 (20,7) (1,9) 32 (91,4) (10) 98 (68,5) (30,5) M1 23 (79,3) (7,2) 3 (8,6) (0,9) 45 (31,5) (14) Grading (n) G1/2 15 (51,7) (4,7) 27 (77,1) (8,4) 60 (42) (18,7)
G3/4 14 (48,3) (4,4) 8 (22,9) (2,5) 83 (58) (25,9) UICC97 (n) 1 0 (0) (0) 1 (2,9) (0,3) 8 (5,6) (2,5)
2 1 (3,4) (0,3) 2 (5,7) (0,6) 14 (9,8) (4,4) 3 2 (6,9) (0,6) 2 (5,7) (0,6) 33 (23,1) (10,3)
4 26 (89,7) (8,1) 30 (85,7) (9,3) 88 (61,5) (27,4) Die Werte in der ersten Klammer sind Relativwerte innerhalb der Sub-Kohorte, wohingegen die Werte in der zweiten Klammer Relativwerte in Bezug auf die gesamte HNSCC-Kohorte darstellen. C.I.: Konfidenzintervall; RCT: Radiochemo-therapie; RT: Radiotherapie. Modifiziert nach (Schwegler, Wirsing et al.).
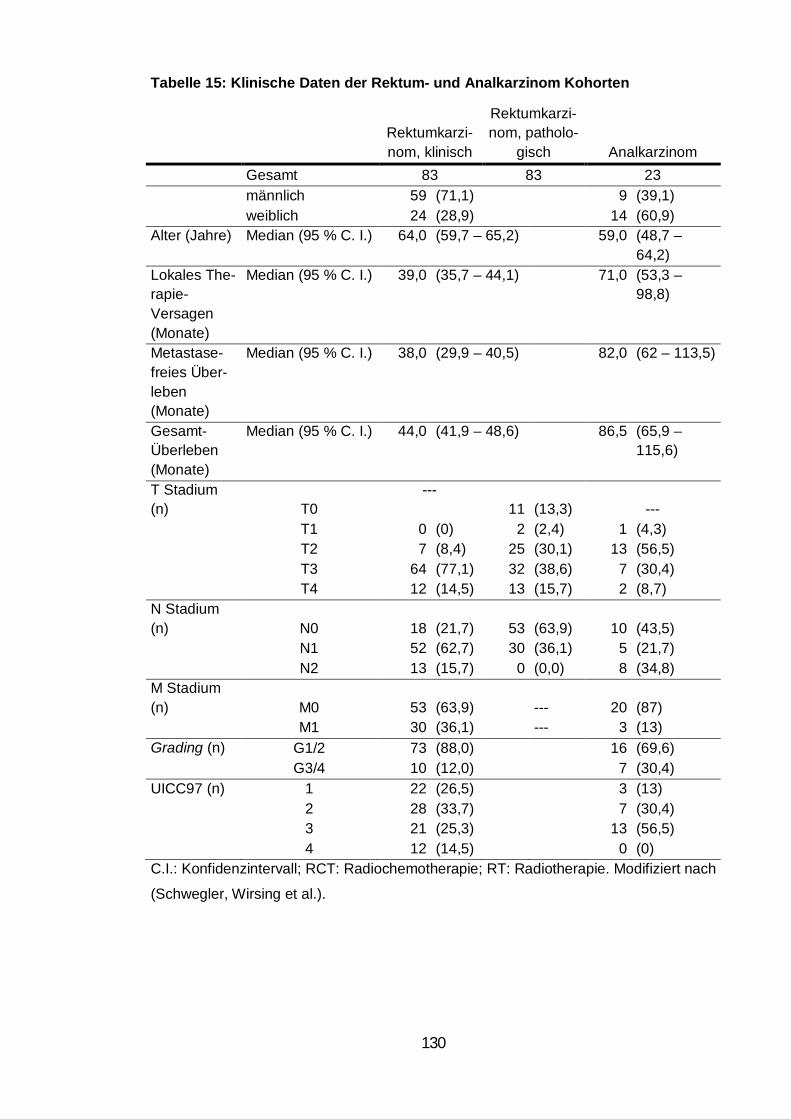
130
Tabelle 15: Klinische Daten der Rektum- und Analkarzinom Kohorten
Rektumkarzi-nom, klinisch
Rektumkarzi-nom, patholo-
gisch Analkarzinom Gesamt 83 83 23
männlich 59 (71,1) 9 (39,1) weiblich 24 (28,9) 14 (60,9)
Alter (Jahre) Median (95 % C. I.) 64,0 (59,7 – 65,2)
59,0 (48,7 – 64,2)
Lokales The-rapie-Versagen (Monate)
Median (95 % C. I.) 39,0 (35,7 – 44,1) 71,0 (53,3 – 98,8)
Metastase-freies Über-leben (Monate)
Median (95 % C. I.) 38,0 (29,9 – 40,5)
82,0 (62 – 113,5)
Gesamt-Überleben (Monate)
Median (95 % C. I.) 44,0 (41,9 – 48,6) 86,5 (65,9 – 115,6)
T Stadium (n) T0
--- 11 (13,3) ---
T1 0 (0) 2 (2,4) 1 (4,3) T2 7 (8,4) 25 (30,1) 13 (56,5) T3 64 (77,1) 32 (38,6) 7 (30,4)
T4 12 (14,5) 13 (15,7) 2 (8,7) N Stadium (n) N0 18 (21,7) 53 (63,9) 10 (43,5)
N1 52 (62,7) 30 (36,1) 5 (21,7) N2 13 (15,7) 0 (0,0) 8 (34,8)
M Stadium (n) M0 53 (63,9) --- 20 (87) M1 30 (36,1) --- 3 (13) Grading (n) G1/2 73 (88,0) 16 (69,6)
G3/4 10 (12,0) 7 (30,4) UICC97 (n) 1 22 (26,5) 3 (13)
2 28 (33,7) 7 (30,4) 3 21 (25,3) 13 (56,5)
4 12 (14,5) 0 (0) C.I.: Konfidenzintervall; RCT: Radiochemotherapie; RT: Radiotherapie. Modifiziert nach
(Schwegler, Wirsing et al.).
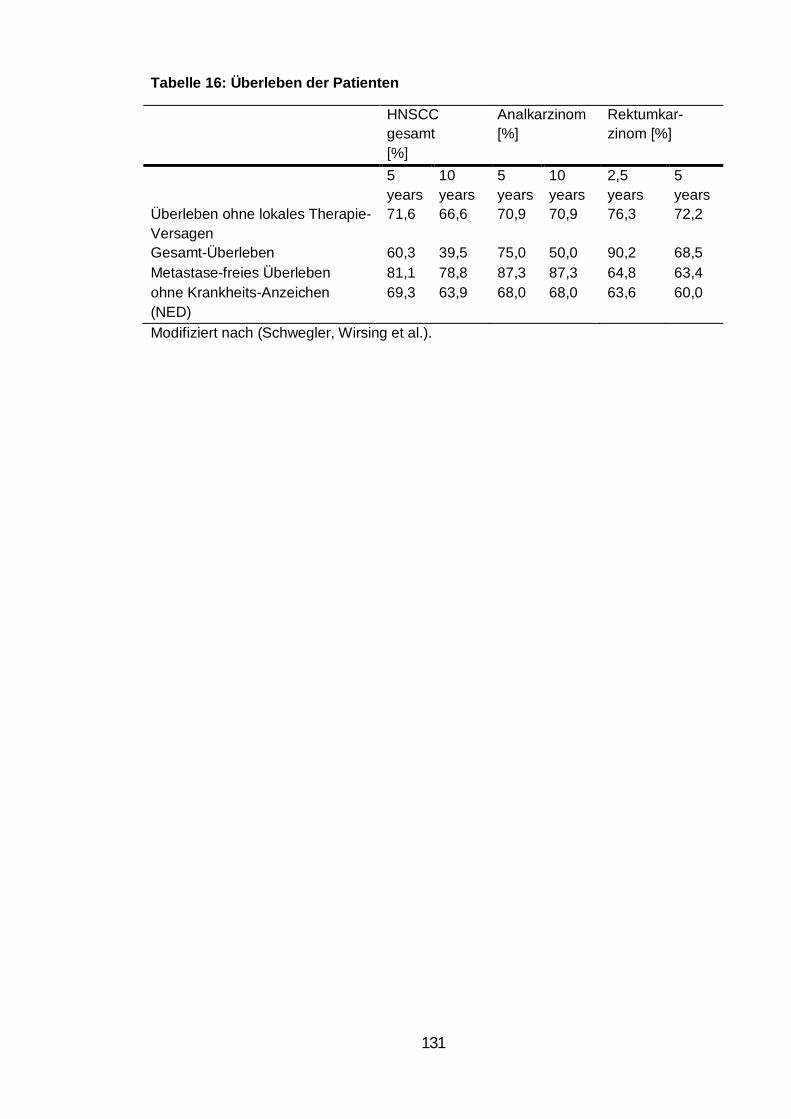
131
Tabelle 16: Überleben der Patienten
HNSCC gesamt [%]
Analkarzinom [%]
Rektumkar-zinom [%]
5 years
10 years
5 years
10 years
2,5 years
5 years
Überleben ohne lokales Therapie-Versagen
71,6 66,6 70,9 70,9 76,3 72,2
Gesamt-Überleben 60,3 39,5 75,0 50,0 90,2 68,5 Metastase-freies Überleben 81,1 78,8 87,3 87,3 64,8 63,4 ohne Krankheits-Anzeichen (NED)
69,3 63,9 68,0 68,0 63,6 60,0
Modifiziert nach (Schwegler, Wirsing et al.).

132
7. Danksagung Ich bedanke mich bei Prof. Dr. Rainer Fietkau (Direktor der Strahlenklinik) für
die finanzielle Unterstützung und die Tätigkeit als Gutachter.
Mein großer Dank gilt PD Dr. Luitpold Distel für die Bereitstellung des Themas
und die Betreuung, insbesondere der histologischen Untersuchungen, sowie die
Tätigkeit als Prüfer.
Großer Dank geht auch an Prof. Dr. Falk Nimmerjahn, der sich als Betreuer an
der Naturwissenschaftlichen Fakultät, als Gutachter sowie als Prüfer zur Verfü-
gung gestellt hat.
Bei Prof. Dr. Benedikt Kost bedanke ich mich für die freundliche Bereitschaft als
Zweitprüfer.

133
8. Lebenslauf
PERSÖNLICHE DATEN Name: Manuela Schwegler Geburtsdatum: 23.12.1984 Geburtsort: München Staatsangehörigkeit: Deutsch AUSBILDUNG 12/2010 – 05/2015 Promotion an der Strahlenklinik Erlangen
Titel der Dissertation: Phagozytose von nekrotischen Zellen durch gesunde Haut-Fibroblasten und Tu-morzellen
10/2004 – 03/2010 Studium Biologie (Diplom) Ludwig-Maximilians-Universität München, Ab-schlussnote: 1,3; Titel der Diplomarbeit: Untersu-chungen des Einflusses von MCAM auf die Migrati-on von Endothelzellen in der Angiogenese
1995 – 2004 Pestalozzi-Gymnasium München, Abitur

134
9. Publikationsliste
Schwegler M, Wirsing AM, Dollinger AJ, Abendroth B, Putz F, Fietkau R, Distel
LV. Clearance of primary necrotic cells by nonprofessional phagocytes. Biology
of the Cell, 2015; 107(10): 372-387.
Schwegler M, Wirsing AM, Schenker HM, Ott L, Fietkau R, Putz F, Distel LV.
Prognostic Value of Homotypic Cell Internalization by Non-Professional Phago-
cytic Cancer Cells. BioMed Research International, 2015; Article ID 359392
(elektronische Veröffentlichung vor dem Druck 04.10.2015).
Kroeber J, Wenger B, Schwegler M, Daniel C, Schmidt M, Djuzenova CS, Polat
B, Flentje M, Fietkau R, Distel LV. Distinct increased outliers among 136 rectal
cancer patients assessed by γH2AX. Radiation Oncology, 2015; 10:36.
Wenger B, Schwegler M, Brunner M, Daniel C, Schmidt M, Fietkau R, Distel LV.
PML-nuclear bodies decrease with age and their stress response is impaired in
aged individuals. BMC Geriatr., 2014; 14: 42.
Hecht M, Harrer T, Büttner M, Schwegler M, Erber S, Fietkau R, Distel LV. Cy-
totoxic effect of efavirenz is selective against cancer cells and associated with
the cannabinoid system. AIDS, 2013; 27: 2031-2040.





























