Embed Size (px)
Citation preview

Peroxisome Proliferator-Activated Receptor-�-MediatedPositive Energy Balance in the Rat Is Associated withReduced Sympathetic Drive to Adipose Tissues andThyroid Status
William T. Festuccia, Serdar Oztezcan, Mathieu Laplante, Magalie Berthiaume, Chantal Michel,Shinya Dohgu, Raphael G. Denis, Marcia N. Brito, Nilton A. Brito, David S. Miller, William A. Banks,Timothy J. Bartness, Denis Richard, and Yves Deshaies
Laval Hospital Research Center (W.T.F., M.L., M.B., C.M., R.G.D., D.R., Y.D.), Faculty of Medicine, Laval University,Quebec, Canada G1V 4G5; Department of Biochemistry (S.O.), Medical Faculty of Istanbul, Istanbul University, Istanbul34452, Turkey; Geriatrics Research (S.O., S.D., W.A.B.), Education, and Clinical Center, Veterans Affairs Medical Center,St. Louis, St. Louis, Missouri 63106; Division of Geriatrics (W.A.B.), Department of Internal Medicine, St. Louis UniversitySchool of Medicine, St. Louis, Missouri 63104; Laboratory of Pharmacology and Chemistry (D.S.M.), National Institute ofEnvironmental Health Science, National Institutes of Health, Research Triangle Park, North Carolina 27709; and BiologyDepartment (M.N.B., N.A.B., T.J.B.), Georgia State University, Atlanta, Georgia 30302
Peroxisome proliferator-activated receptor-� (PPAR�) acti-vation up-regulates thermogenesis-related genes in rodentwhite and brown adipose tissues (WAT and BAT) without in-creasing whole-body energy expenditure. We tested herewhether such dissociation is the result of a negative modula-tion of sympathetic activity to WAT and BAT and thyroid axiscomponents by PPAR� activation. Administration of thePPAR� agonist rosiglitazone (15 mg/kg�d) for 7 d to maleSprague Dawley rats increased food intake (10%), feed effi-ciency (31%), weight gain (45%), spontaneous motor activity(60%), and BAT and WAT mass and reduced whole-body oxy-gen consumption. Consistent with an anabolic setting, ros-iglitazone markedly reduced sympathetic activity to BAT andWAT (>50%) and thyroid status as evidenced by reduced levelsof plasma thyroid hormones (T4 and T3) and mRNA levels ofBAT and liver T3-generating enzymes iodothyronine type 2
(�40%) and type 1 (�32%) deiodinases, respectively. Rosigli-tazone also decreased mRNA levels of the thyroid hormonereceptor (THR) isoforms �1 (�34%) and � (�66%) in BAT andisoforms �1 (�20%) and �2 (�47%) in retroperitoneal WAT.These metabolic effects were associated with a reduction inmRNA levels of the pro-energy expenditure peptides CRH andCART in specific hypothalamic nuclei. A direct central actionof rosiglitazone is, however, unlikely based on its low brainuptake and lack of metabolic effects of intracerebroventric-ular administration. In conclusion, a reduction in BAT sym-pathetic activity and thyroid status appears to, at least partly,explain the PPAR�-induced reduction in energy expenditureand the fact that up-regulation of thermogenic gene expres-sion does not translate into functional stimulation of whole-body thermogenesis in vivo. (Endocrinology 149: 2121–2130,2008)
PEROXISOME PROLIFERATOR-activated receptor (PPAR)-�is a ligand-activated nuclear receptor that is mainly ex-
pressed in white adipose tissue (WAT) and rodent brown adiposetissue (BAT) where it controls the expression of several proteinsinvolved in lipid metabolism. Although the identity of functionalendogenous PPAR� ligands remains uncertain, the receptor isspecifically activated by thiazolidinediones, a class of syntheticagonists that, because of their beneficial effect on whole-body in-
sulin sensitivity, are widely used in the treatment of insulin-resis-tant states such as type 2 diabetes and metabolic syndrome (1, 2).
WAT and BAT, the major sites in rodents of energy storageand nonshivering thermogenesis, respectively, are the maintargets of PPAR� activation. In these tissues, PPAR� activa-tion is associated with an increase in the number of differ-entiated adipocytes with enhanced capacity to take up andstore lipids (3, 4); such an effect partially accounts for theconcomitant reduction in circulating nonesterified fatty acidsand triacylglycerol (TAG) levels. Paradoxically to enhancedfat storage, PPAR� activation is also associated with in-creases not only in WAT and BAT mitochondrial biogenesisand levels of the thermogenic uncoupling protein 1 (UCP1)but also in those of key proteins involved in fatty acid oxi-dation and lipolysis (5–8). In accordance with its positiveeffects on the lipolytic, oxidative, and thermogenic machin-eries, in vitro PPAR� activation in fetal primary brown adi-pocytes, 3T3-L1 adipocytes, and primary white adipocytesincreases rates of lipolysis, fatty acid oxidation, O2 consump-tion, and uncoupled respiration (5–7).
Despite the potent catabolic profile found in vitro, in vivo
First Published Online January 24, 2008Abbreviations: ARC, Arcuate region; BAT, brown adipose tissue;
CART, cocaine- and amphetamine-regulated transcript; CsA, cyclo-sporin A; D1 and D2, type 1 and type 2 iodothyronine deiodinase;HBMEC, human brain microvascular endothelial cells; icv, intracere-broventricular; ING, inguinal; NE, norepinephrine; NETO, NE turnover;P-gp, p-glycoprotein; PPAR, peroxisome proliferator-activated receptor;pPVN, parvocellular subregion of the hypothalamic paraventricularnucleus; RETRO, retroperitoneal; SMA, spontaneous locomotor activity;TAG, triacylglycerol; THR, thyroid hormone receptor; UCP1, uncou-pling protein 1; WAT, white adipose tissue.Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
0013-7227/08/$15.00/0 Endocrinology 149(5):2121–2130Printed in U.S.A. Copyright © 2008 by The Endocrine Society
doi: 10.1210/en.2007-1553
2121

administration of PPAR� agonists brings about opposite ef-fects. Indeed, in the face of a robust increase in the expressionof lipolytic, oxidative, and thermogenic genes in WAT andBAT, the net result of PPAR� activation in vivo is an increasein energy storage rather than in energy expenditure. In fact,administration of PPAR� agonists to rodents and humansincreases body weight and fat mass gains as well as foodintake and efficiency, all indicative of an anabolic condition(9–12). Notably, the high thermogenic, oxidative, and lipo-lytic potential (i.e. mRNA and protein content and concom-itant increase in related processes when assessed in vitro)induced by PPAR� activation are actuated at the functionallevel in vivo only under pharmacological �-adrenergic stim-ulation (9, 13, 14). This strongly suggests that the PPAR�-induced up-regulation of BAT and WAT lipolytic, oxidative,and thermogenic machineries are silenced by the in vivoneural and hormonal milieu such that energy storage, ratherthan expenditure, is favored.
Release of norepinephrine (NE) by the sympathetic ner-vous system innervating fat pads, along with the thyroidhormone T3, constitute the major activators of WAT and BATlipolysis and thermogenesis (15, 16). In the present study, wetested the hypothesis that the dissociation between geneexpression and biological processes found in vivo underPPAR� activation is caused by a concomitant reduction insympathetic activity and thyroid status. To this end, weassessed in rats the effects of the PPAR� agonist rosiglitazoneon WAT and BAT NE turnover rates (NETO), a measure ofsympathetic activity, and on the hypothalamic-pituitary-thy-roid axis, including BAT and liver type 2 and 1 iodothyroninedeiodinase (D2 and D1, respectively) and thyroid hormonereceptor (THR) mRNA levels.
Because PPAR� is expressed at low levels in some hypo-thalamic regions (17), we further tested the hypothesis thatrosiglitazone exerts its effects on energy balance centrally.This was addressed by assessing the effects of rosiglitazoneon the expression of hypothalamic neuropeptides involvedin the control of energy expenditure, the ability of rosigli-tazone to cross the blood-brain barrier, and the impact ofcentral administration of rosiglitazone on energy balance.
Materials and MethodsAnimals and treatment
Animal care and handling were performed in accordance with theCanadian Guide for the Care and Use of Laboratory Animals. All ex-perimental procedures received prior approval of the Laval Universityanimal care committee. Male Sprague Dawley rats (Charles River Lab-oratories, St. Constant, Quebec, Canada) initially weighing 175–200 gwere individually housed in stainless steel cages in a room kept at 23 �1 C with a 12-h light, 12-h dark cycle (lights on at 0800 h). After a 4-dadaptation period, rats were matched by weight and divided into controland rosiglitazone-treated groups that received a nonpurified powderedrodent diet (Charles River Rodent Diet no. 5075; Woodstock, Ontario,Canada; digestible energy content, 12.9 kJ/g) alone (control) or supple-mented with the PPAR� agonist rosiglitazone (purchased as AVANDIAat a local pharmacy) at a dose of 15 mg/kg�d for 7 d. This dose waschosen based on preliminary studies that showed its effectiveness toincrease both visceral and sc adipose depot mass within a short periodof treatment (e.g. 7 d). Ground rosiglitazone was mixed with the pow-dered chow diet, and the desired dose was achieved by adjusting theamount of drug to the average food consumption and body weight ofrats every other day.
Energy expenditure and spontaneous motor activity
O2 consumption, CO2 production, and respiratory quotient weredetermined over 1 wk in an open-circuit system with an O2 (AppliedElectrochemistry, Naperville, IL; S-3A1) and a CO2 analyzer (AppliedElectrochemistry, CD-3A). Data are presented as ml/min�kg0.75 bodyweight. Locomotor activity was measured with the AccuScan DigiscanActivity Monitor (AccuScan Instruments, Columbus, OH), with the aidof the VersaMax software (version 1.30; AccuScan Instruments). Ratswere placed individually in acrylic chambers (40 � 40 � 30 cm), andmotor activity was measured for 48 h after a 24-h adaptation period.Cages contained an array of beams (every 2.5 cm) in all three dimensions(16 in x-axis left-right, 16 in y-axis front-back, and 16 in z-axis vertical).Motor activity was determined by breaks in photobeams in the hori-zontal plane and converted into spontaneous locomotor activity (SMA,meters per day).
Adipocyte morphology by light microscopy, DNA, andTAG content
Portions of retroperitoneal (RETRO) WAT and interscapular BATwere fixed in 0.1 mm PBS (pH 7.4) containing 4% paraformaldehyde andembedded in paraffin. Thin sections were mounted on glass slides anddyed with hematoxylin/eosin. Digital images of tissue slices were cap-tured and analyzed as previously described (4). Tissue DNA content wasdetermined using the DNeasy tissue kit (QIAGEN, Mississauga, On-tario, Canada) following manufacturer’s instructions. Tissue TAG con-tent was measured after lipid extraction with chloroform-methanol (2:1)with an enzymatic kit (Roche Diagnostics, Montreal, Canada).
NETO
NETO rate, a reliable index of sympathetic activity in a given tissue,was estimated from the decline in tissue NE content after inhibition ofcatecholamine synthesis with dl-�-methyl-tyrosine ester (Sigma Chem-ical Co., St. Louis, MO). Rats were killed by anesthetic (ketamine/xylazine) overdose before or 4 h after ip injection of dl-�-methyl-ty-rosine ester (350 mg/kg body weight), exactly as previously described(18–20). These time points are based on previous studies establishing thelinearity of the logarithmic decline in NE content in rat tissues afterinhibition of catecholamine synthesis (21, 22). The inguinal (ING) andRETRO fat depots and interscapular BAT were rapidly removed,weighed, frozen in liquid nitrogen, and stored at �80 C for later de-termination of NE content. NETO experiments were repeated threetimes with high reproducibility. Rates of NETO were calculated as theproduct of the fractional turnover rate (k) and the endogenous NEcontent at time 0 as previously described (23). Fractional turnover rate(k) was calculated by the formula: k � (log [NE]0� log [NE]4)/(0.434 �4), where [NE]0 and [NE]4 are the NE content at times 0 and 4 h,respectively.
Tissue NE content
Tissue NE content was measured as previously described (22).Briefly, tissues were homogenized in 0.2 n perchloric acid, 1 mm EDTA,1% sodium metabisulfite, with dihydroxybenzylamide as an internalstandard, and centrifuged, and the supernatant destined for catechol-amine quantification was extracted with alumina. Catecholamines wereeluted from alumina with the above homogenization solution withoutinternal standard and assayed by HPLC.
Intracerebroventricular (icv) rosiglitazone infusion
Cannulae were implanted into the third ventricle of isoflurane-anes-thetized rats as previously described (24). Cannulae were connectedwith silicon catheters to mini-osmotic pumps (model 2002; Alzet, Cu-pertino, CA) that delivered a solution of rosiglitazone maleate(BRL49653; GlaxoSmithKline, Mississauga, Ontario, Canada) at doses of50 and 500 �g/ml dissolved in sterile saline containing 12.5% dimeth-ylsulfoxide (pH 7.4). Pumps implanted in the interscapular region de-livered the solution at a rate of 0.5 �l/h for 7 d. Control rats were infusedwith vehicle. Before and 2 d after surgery, rats were given analgesics
2122 Endocrinology, May 2008, 149(5):2121–2130 Festuccia et al. • PPAR� Activation and Energy Balance

(ketoprophene, 5 mg/kg, twice a day). Cannula positioning was eval-uated microscopically a posteriori in all rats.
Hypothalamic dissection
Hypothalami were rapidly dissected using as landmarks the opticchiasma rostrally and the mammillary bodies caudally and frozen inliquid nitrogen for later RNA extraction and quantification of neuropep-tide expression levels by PCR.
In situ hybridization for CRH and cocaine- andamphetamine-related transcript (CART)
Hypothalamic mRNA levels of CRH and CART, two neuropeptidesinvolved in the central regulation of sympathetic activity, were mea-sured by in situ hybridization essentially as previously described (25).Briefly, hypothalamic sections were mounted onto poly-l-lysine-coatedslides, dehydrated in ethanol, fixed in paraformaldehyde, digested withproteinase K (10 �g/ml), acetylated with 0.25% acetic anhydride, anddehydrated in ethanol gradient. Sections were incubated overnight withantisense 35S-labeled cRNA probe (107 cpm/ml) for CRH or CART at 60C. Slides were rinsed with sodium chloride/sodium citrate solution,digested with RNase-A, washed in descending concentrations of sodiumchloride/sodium citrate solution, and dehydrated in ethanol gradient.Slides were defatted in toluene, dipped in NTB2 nuclear emulsion (East-man Kodak, Rochester, NY), and exposed for 7 d before being devel-oped. Slides were examined by dark-field microscopy using an OlympusBX51 microscope (Olympus America, Melville, NY). Images were ac-quired with an Evolution QEi camera and analyzed with ImagePro plusversion 5.0.1.11 (MediaCybernetics, Silver Spring, MD). The system wascalibrated for each set of analyses to prevent saturation of the integratedsignal. Mean pixel densities were obtained by taking measurementsfrom both hemispheres of one to four brain sections and subtractingbackground readings taken from areas immediately surrounding theregion analyzed.
RNA isolation and quantification
RNA was isolated from adipose depots and hypothalamus usingQIAzol and the RNeasy lipid tissue kit (QIAGEN). For cDNA synthesis,expand reverse transcriptase (Invitrogen) was used following the man-ufacturer’s instructions, and cDNA was diluted in DNase-free water(1:25) before quantification by real-time PCR. mRNA transcript levelswere measured in duplicate samples using a Rotor Gene 3000 system(Montreal Biotech, Montreal, Quebec, Canada). The primers used for thePCR are available upon request to the corresponding author. Chemicaldetection of the PCR products was achieved with SYBR Green I (Mo-lecular Probes, Willamette Valley, OR). At the end of each run, melt-curve analyses were performed, and a few samples representative ofeach experimental group were run on agarose gel to ensure the speci-ficity of the amplification. Results are expressed as the ratio between theexpression of the target gene and the housekeeping genes 36B4(NM_022402) for adipose tissues and liver or GAPDH (NM_017008) forhypothalamus, which were selected because no significant variation intheir expression was observed between control and rosiglitazone-treated rats.
Rosiglitazone transport through the blood-brain barrier
All in vivo experiments investigating rosiglitazone transport throughthe blood-brain barrier were performed in male CD-1 mice (35–40 g)from our in-house colony (W.A.B., Veterans Affairs Medical Center, St.Louis, MO). Mice were used because the methodology involves intra-cranial injection procedures that have been optimized for this speciesand are not readily adaptable to the rat. At least in terms of blood-to-brain rosiglitazone transport, there appears to be no difference betweenthe two species based on the present findings in mice and previousevidence in rats (26). In addition, to investigate the applicability of thefindings obtained with mice to other species, rosiglitazone transport inand out of the brain was also investigated in human brain microvascularendothelial cells, a human model of the blood-brain barrier (see below).
Blood-to-brain uptake
Mice had free access to food and water and were maintained on a 12-hdark, 12-h light cycle in a room with controlled temperature (24 � 1 C)and humidity (55 � 5%). Mice were anesthetized with an ip injection of0.2 ml of a 40% urethane solution. The left jugular vein and the rightcarotid artery were isolated (27). Approximately 1.2 � 106 cpm [3H]ro-siglitazone (American Radiolabeled Chemical, St. Louis, MO) with orwithout 1 �g unlabeled rosiglitazone was injected into the jugular veinin 200 �l lactate-buffered Ringer’s solution containing 1% BSA. At 1, 2,3, 4, 5, 7.5, 10, 20, and 30 min after injection, blood was freely collectedfor about 15 sec from a cut in the carotid artery, and serum was isolatedby centrifugation. Mice were then immediately decapitated, blood wascollected, and the whole brain minus pineal and pituitary glands wasremoved and weighed. Brain and serum samples were incubated withtissue solubilizer (1 ml/200 mg tissue and 500 �l/50 �l serum) in a waterbath at 40 C for 24 h. Scintillation cocktail (10 ml) was added, andsamples were allowed to quench for 24 h in the dark before determi-nation of radioactivity content.
The percentage of the iv dose injected per milliliter of serum wascalculated with equation 1: % Inj/ml � 100 � (cpm/ml serum)/ (cpm/injection). The brain/serum ratio (microliters per gram) was obtained byequation 2: brain/serum ratio � (cpm/brain)/[(cpm/�l serum) � (brainweight)]. The percentage of injected dose taken up per gram of brain (%Inj/g) was calculated from equation 3: % Inj � (R � 10) (% S), where Ris brain-to-blood ratio expressed in microliters per gram calculated fromequation 2 above, 10 �l is taken as the volume of blood in 1 g of brain,and % S is the % Inj/ml as calculated in equation 1 above.
Brain-to-blood transport
The icv injection method was used to study brain-to-blood transport(28). Mice (n � 7–9 per group) were anesthetized with an ip injection ofurethane. After the scalp was removed, a hole was made through thecranium 1.0 mm lateral and 0.5 mm posterior to the bregma with a26-gauge needle. The entire needle except 2.5 mm of the tip was coveredwith PE-10 tubing. This ensured that the tip of the needle penetrated theskull and brain tissue forming the roof of the lateral ventricle but did notpenetrate the ventricular floor. One microliter of lactated Ringer’s so-lution with 1% BSA containing 5000 cpm of [3H]rosiglitazone with orwithout unlabeled rosiglitazone (1.5 �g/mouse) or the P-glycoprotein(P-gp) inhibitor SDZ PSC833 (100 ng/mouse) was injected into the
TABLE 1. Final body weight, cumulative food intake, feedefficiency, O2 consumption (VO2), CO2 production (VCO2),respiratory quotient (RQ), SMA, mass of WAT and BAT, andserum levels of hormones and metabolites in rats treated or notwith rosiglitazone (RSG) for 7 d
Control RSG
Final body weight (g) 303 � 7 328 � 5a
Body weight gain (g) 37 � 4 54 � 4a
Food intake (g) 144 � 5 159 � 5a
Feed efficiency (%)b 25 � 2 33 � 2a
VO2 (ml/min�kg bw0.75) 14.8 � 0.2 13.9 � 0.2a
VCO2 (ml/min�kg bw0.75) 11.8 � 0.2 11.1 � 0.2a
RQ 0.798 � 0.006 0.796 � 0.007SMA (m/d) 66 � 7 106 � 12a
BAT (g) 0.31 � 0.01 0.67 � 0.02a
ING WAT (g) 3.2 � 0.2 4.3 � 0.2a
RETRO WAT (g) 2.0 � 0.2 2.7 � 0.1a
Insulin (pM) 186 � 22 122 � 18a
Glucose (mM) 7.9 � 0.3 7.8 � 0.1NEFA (mM) 0.47 � 0.05 0.20 � 0.07a
TAG (mM) 2.0 � 0.2 1.1 � 0.1a
ACTH (�M) 28 � 3 16 � 1a
Leptin (ng/ml) 4.6 � 0.4 5.2 � 0.5Adiponectin (�g/ml) 3.8 � 0.4 16.8 � 0.8a
Data are means � SEM of six to 12 rats. bw, Body weight; NEFA,nonesterified fatty acids.
a P � 0.05 vs. control.b Calculated as grams body weight gain per 100 g food ingested.
Festuccia et al. • PPAR� Activation and Energy Balance Endocrinology, May 2008, 149(5):2121–2130 2123

lateral ventricle with a 1-�l Hamilton syringe. The amount of rosigli-tazone available for transport was determined in mice that were killedwith an overdose of urethane 10–20 min before the icv injection. Themice were decapitated and brains were harvested 10 min after the icvinjection. The whole brains were removed and incubated with tissuesolubilizer and scintillation cocktail as above and counted. The amountof [3H]rosiglitazone transported was expressed as a percentage of thecounts available for transport (A): % transported � 100 � (A � 10-minmean)/(A).
In vitro human blood-brain barrier model
Human brain microvascular endothelial cells (HBMECs) were pur-chased from Applied Cell Biology Research Institute (Kirkland, WA).HBMECs were cultured in CS-C Complete Medium (Cell Systems Corp.,Kirkland, WA) supplemented with 50 �g/ml gentamicin (Life Tech-nologies, Inc., Invitrogen, Grand Island, NY) at 37 C with a humidifiedatmosphere of 5% CO2/95% O2 air. HBMECs were treated with PassageReagent Group (Cell Systems) and then seeded on the inside of thefibronectin/collagen IV-coated (0.1 and 0.5 mg/ml, respectively) poly-ester membrane (0.33 cm2, 0.4-�m pore size) of a Transwell-Clear insert(Costar, Corning, NY) (4 � 104 cells per well) placed in the well of a24-well culture plate. Four to five days after seeding, HBMECs at pas-sages 4–6 were used for the transport experiments.
[3H]Rosiglitazone transport
For the transport experiments, the medium was removed and HB-MECs were washed with serum-free CS-C medium (Cell Systems). HB-MECs were preincubated with or without PSC833 (10 �m) and cyclo-sporin A (CsA; 10 �m) (Sigma) for 30 min. To initiate the transportexperiments, [3H]rosiglitazone and [14C]sucrose (2 �Ci/ml, approxi-mately 1.8 � 106 cpm/ml) with or without the P-gp inhibitors PSC833(10 �m) and CsA (10 �m) were loaded on the luminal (100 �l) orabluminal chamber (600 �l). The side opposite to that to which thelabeled materials were loaded is the collecting chamber. Samples wereremoved from the collecting chamber at 5, 10, 20, and 60 min andimmediately replaced with an equal volume of fresh serum-free me-dium. The sampling volume from the luminal and abluminal chamberwas 50 and 300 �l, respectively. Samples were mixed with 6 ml scin-tillation cocktail (Bio-Safe II; Research Products International Corp.,Mount Prospect, IL), and radioactivity was determined in a liquid scin-tillation counter. The permeability coefficient and clearance of [3H]ro-siglitazone and [14C]sucrose was calculated as previously described (29).Clearance was expressed as microliters of radioactive tracer diffusingfrom the luminal to abluminal (influx) chamber or from the abluminalto luminal (efflux) chamber and was calculated from the initial level ofradioactivity in the loading chamber and final level of radioactivity inthe collecting chamber: clearance (microliters) � [C]C � VC/[C]L, where[C]L is the initial radioactivity in 1 �l of loading chamber (in cpm permicroliter), [C]C is the radioactivity in 1 �l of collecting chamber (in cpmper microliter), and VC is the volume of collecting chamber (in micro-liters). During a 60-min period of the experiment, the clearance volumeincreased linearly with time. The volume cleared was plotted vs. time,and the slope was estimated by linear regression analysis. The slope of
clearance curves for the HBMEC monolayer plus Transwell membranewas denoted by PSapp, where PS is the permeability � surface areaproduct (in microliters per minute). The slope of the clearance curve witha Transwell membrane without HBMECs was denoted by PSmembrane.The real PS value for the HBMEC monolayer (PSe) was calculated from1/PSapp � 1/PSmembrane � 1/PSe. The PSe values were divided by thesurface area of the Transwell inserts (0.33 cm2) to generate the endo-thelial permeability coefficient (Pe, in centimeters per minute).
Serum determinations
Plasma glucose concentrations were measured by the glucose oxidasemethod with the YSI 2300 STAT plus glucose analyzer. Plasma insulin,leptin, adiponectin (Linco Research, St. Charles, MO), total and free T3and T4 (Coat-A-Count; Diagnostic Products Corp., Los Angeles, CA),TSH (Biotrak rat TSH; Amersham Biosciences, Oakville, Ontario, Can-ada) and ACTH (Somagen Diagnostic, Edmonton, Alberta, Canada)were determined by RIA. Plasma TAG and nonesterified fatty acid levelswere measured by enzymatic methods (Roche Diagnostics, Montreal,Quebec, Canada, and Wako Chemicals, Richmond, VA, respectively).
Statistical analysis
Results are expressed as means � sem. Simple effects of rosiglitazonetreatment were analyzed by Student’s unpaired t test. When appropri-ate, factorial ANOVA followed by either Newman-Keuls’ multiplerange test or Dunnett’s test or Tukey-Kramer’s was used for multiplecomparisons. P � 0.05 was taken as the threshold of significance.
Results
Final body weight and body weight gain were signifi-cantly increased (8 and 45%, respectively) by 7 d of rosigli-tazone treatment (Table 1). The higher body weight of ros-iglitazone-treated rats was associated with increases in foodintake (10%), feed efficiency (31%), and mass of brown andwhite adipose depots (BAT, 116%; ING, 25%; RETRO, 25%).In addition, rosiglitazone induced a small, but significantdecrease in the rates of O2 consumption and CO2 production(�7%), despite markedly increasing SMA (60%). Confirmingits beneficial effects on insulin sensitivity and lipemia, ros-iglitazone significantly reduced fasting plasma levels of in-sulin (�35%), nonesterified fatty acids (�60%), and TAG(�45%) but did not alter fasting glycemia. Rosiglitazone alsosignificantly decreased plasma ACTH levels (�42%), leftleptin levels unchanged, and increased those of adiponectin4-fold.
The effects of rosiglitazone on BAT and WAT NE content,fractional turnover rates, and NETO are presented in Table2. One week of rosiglitazone markedly reduced sympathetic
TABLE 2. Effect of rosiglitazone (RSG) treatment on NE content, fractional turnover rates (k), and NETO rates in rat BAT, RETRO, andING adipose depots
NE content (ng)a k (%/h) NETO (ng NE/tissue�h)
BATControl 314 � 21 9.2 � 0.8 29 � 3RSG 296 � 20 4.0 � 0.6b 12 � 2b
RETROControl 75 � 2 11.5 � 1.2 8.6 � 1.0RSG 72 � 3 4.6 � 0.5b 3.3 � 0.3b
INGControl 124 � 10 6.6 � 0.7 8.2 � 1.6RSG 140 � 16.3 2.8 � 0.4b 3.9 � 0.5b
Data are means � SEM of 12 rats.a BAT, ng/tissue; WAT, ng/pad.b P � 0.05 vs. control.
2124 Endocrinology, May 2008, 149(5):2121–2130 Festuccia et al. • PPAR� Activation and Energy Balance

activity to BAT and WAT as evidenced by the lower frac-tional turnover rates and NETO to interscapular BAT (�56and �58%, respectively), RETRO (�60 and �62%), and ING(�57 and �53%) compared with untreated rats.
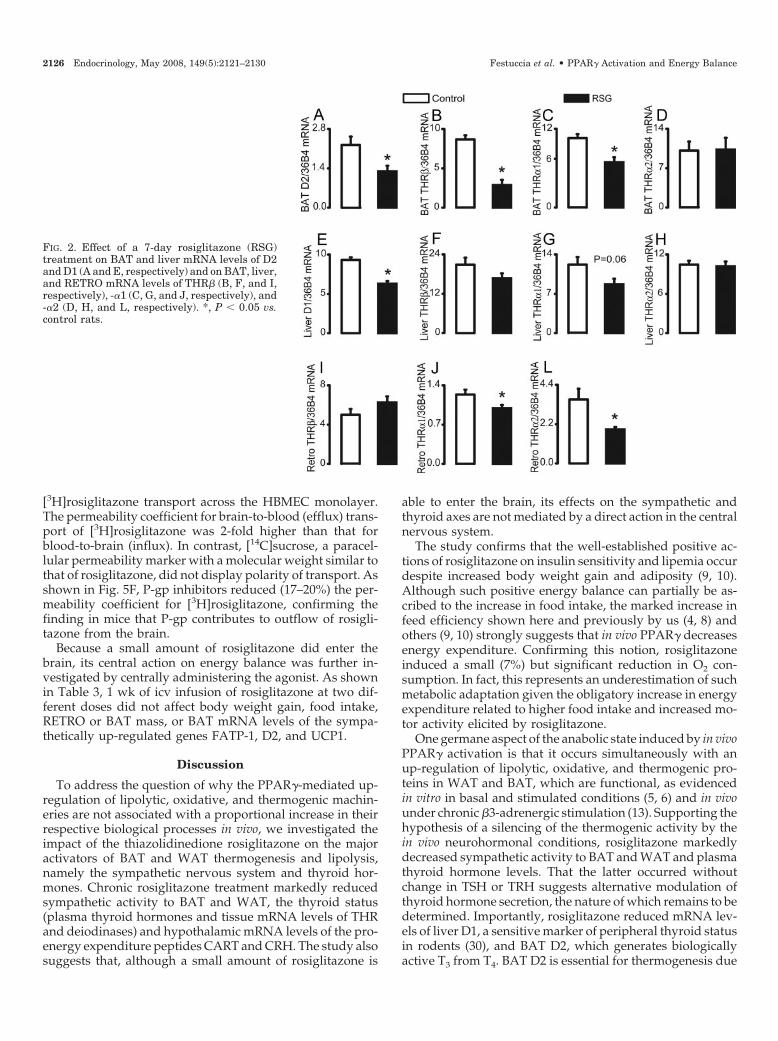
Because thyroid hormones exert an important role in theamplification of sympathetically mediated activation of BATand WAT lipolysis and thermogenesis, we next investigatedthe effects of rosiglitazone on the hypothalamic-pituitary-thyroid axis and on the mRNA levels of the deiodinases andTHR isoforms. Rosiglitazone did not affect hypothalamicTRH mRNA (Fig. 1A) or plasma TSH levels (Fig. 1B) butsignificantly reduced plasma total and free T4 (�26 and�32%, respectively; Fig. 1, C and D) and total T3 (�15%, Fig.1E), whereas free T3 remained unaffected (Fig. 1F). In BAT,rosiglitazone reduced the potential for peripheral conversionof T4 to the biologically active T3 by decreasing BAT D2(�40%, Fig. 2A). Rosiglitazone also significantly reducedBAT THR� (�66%) and -�1 (�34%) mRNA levels, withoutaffecting those of THR�2. In the liver, where rosiglitazonesignificantly reduced D1 mRNA levels (�32%, Fig. 2E), noeffect of the agonist was found on any of the three thyroidhormone isoforms (Fig. 2, F–H). In RETRO, rosiglitazonesignificantly reduced the mRNA levels of THR�1 (�20%, Fig.2J) and -�2 (�47%, Fig. 2L), without affecting those of THR�(Fig. 2I).
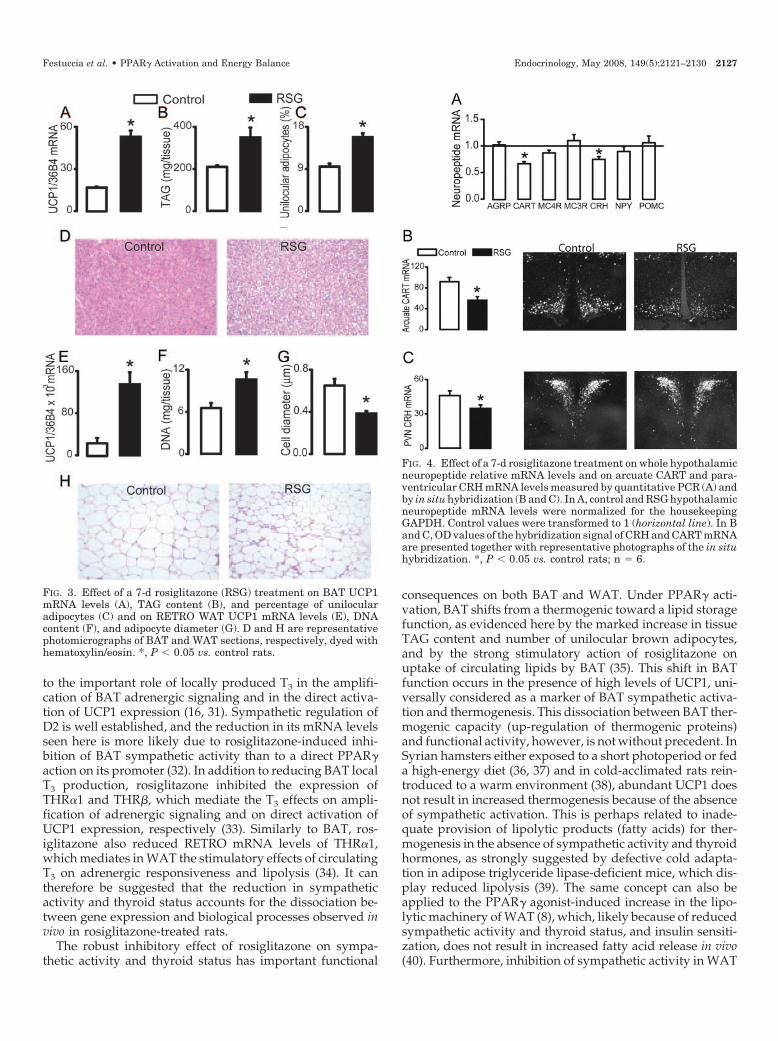
The effects of rosiglitazone on BAT and WAT morphologyand UCP1 expression are depicted in Fig. 3. Noteworthy, therosiglitazone-induced increase in BAT UCP1 mRNA levels(Fig. 3A) was associated with a marked increase in tissue
TAG content (66%, Fig. 3B) and the percentage of unilocularadipocytes (66%, Fig. 3, C and D). In RETRO, the rosiglita-zone-induced increase in UCP1 mRNA levels (Fig. 3E) wasassociated with an increase in DNA content (Fig. 3F) and areduction in adipocyte cell diameter (Fig. 3, G and H).
To investigate the central mechanisms by which PPAR�activation reduces sympathetic activity to WAT and BAT andthyroid status, we assessed the effect of 1 wk of rosiglitazonetreatment on the hypothalamic mRNA levels of several neu-ropeptides involved in the control of energy homeostasis. Asdepicted in Fig. 4A, rosiglitazone significantly reducedwhole hypothalamic mRNA levels of CART (�40%) andCRH (�25%), without altering the mRNA levels of agouti-related protein (AGRP), proopiomelanocortin (POMC),melanocortin receptors 3 and 4 (MC3/4R), or neuropeptideY (NPY). By using in situ hybridization, we confirmed thereductions in CART and CRH mRNA levels found in wholehypothalamic extracts and localized them to specific hypo-thalamic nuclei. CART mRNA levels were significantly re-duced in the arcuate region (ARC, Fig. 4B), but not in theparaventricular, dorsomedial or lateral hypothalamus (datanot shown). CRH mRNA levels were more modestly, butsignificantly reduced in the parvocellular subregion of thehypothalamic paraventricular nucleus (pPVN, Fig. 4C).
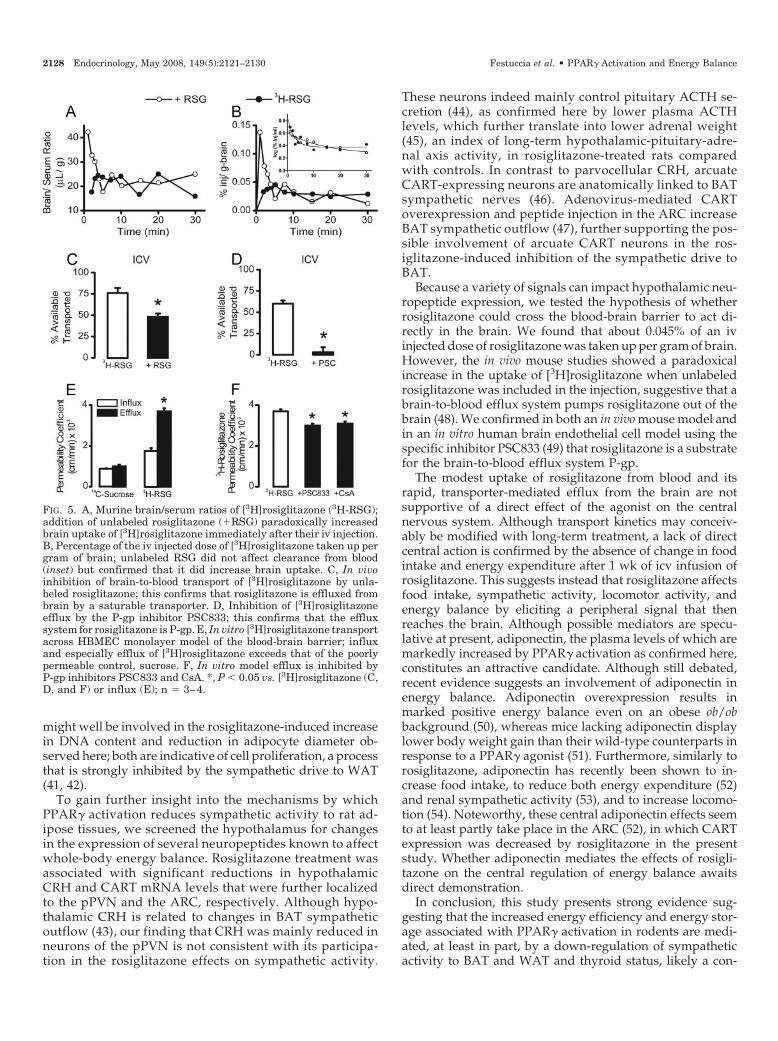
To test the hypothesis that rosiglitazone affects sympa-thetic activity to BAT and WAT and energy balance by actingdirectly in the brain, we first investigated the ability of ros-iglitazone to cross the blood-brain barrier. Figure 5A showsthe relationship between the brain/serum ratio and timeafter iv injection of [3H]rosiglitazone with or without unla-beled rosiglitazone. The brain/serum ratio exceeded the typ-ical 8- to 12-�l vascular space of the brain, indicating uptakeof [3H]rosiglitazone. There was no increase in the brain/serum ratio over time, however, indicating that an equilib-rium between brain and blood was rapidly reached. Unla-beled rosiglitazone paradoxically increased early brainuptake. Figure 5B shows the percentage of the iv-injecteddose that was taken up per unit weight of brain (% Inj/g)after correction for brain vascular space. The peak value for[3H]rosiglitazone was 0.045% Inj/g and occurred 4 min afterinjection. Coinjection of unlabeled rosiglitazone produced anearlier and higher peak (0.134% Inj/g at 1 min) than labeledrosiglitazone alone. This again indicated a paradoxical in-crease in blood-to-brain uptake of [3H]rosiglitazone whencoadministered with unlabeled rosiglitazone. Clearance of[3H]rosiglitazone from blood was minimally affected by un-labeled rosiglitazone (Fig. 5B, inset). Because the presence ofa saturable brain-to-blood transporter is a classic explanationfor the paradoxical increase in tracer uptake in the presenceof unlabeled tracer observed here, the ability of the blood-brain barrier to transport rosiglitazone out of the brain wasalso assessed. Figure 5C shows that 77% of icv-injected[3H]rosiglitazone was transported out of the brain within 10min. Unlabeled rosiglitazone significantly inhibited trans-port. Figure 5D shows that the P-gp inhibitor PSC833 nearlyabolished brain-to-blood transport of [3H]rosiglitazone, thusidentifying P-gp as the brain-to-blood transporter of rosigli-tazone. The presence of a brain-to-blood transport of ros-iglitazone was also evaluated in a human endothelial modelof blood-brain barrier. Figure 5E shows the polarity of
FIG. 1. Effect of a 7-d rosiglitazone (RSG) treatment on hypothalam-ic-pituitary-thyroid axis: hypothalamic TRH mRNA levels (A),plasma TSH (B), total (C) and free T4 (D), and total (E) and free T3(F). *, P � 0.05 vs. control rats.
Festuccia et al. • PPAR� Activation and Energy Balance Endocrinology, May 2008, 149(5):2121–2130 2125
[3H]rosiglitazone transport across the HBMEC monolayer.The permeability coefficient for brain-to-blood (efflux) trans-port of [3H]rosiglitazone was 2-fold higher than that forblood-to-brain (influx). In contrast, [14C]sucrose, a paracel-lular permeability marker with a molecular weight similar tothat of rosiglitazone, did not display polarity of transport. Asshown in Fig. 5F, P-gp inhibitors reduced (17–20%) the per-meability coefficient for [3H]rosiglitazone, confirming thefinding in mice that P-gp contributes to outflow of rosigli-tazone from the brain.
Because a small amount of rosiglitazone did enter thebrain, its central action on energy balance was further in-vestigated by centrally administering the agonist. As shownin Table 3, 1 wk of icv infusion of rosiglitazone at two dif-ferent doses did not affect body weight gain, food intake,RETRO or BAT mass, or BAT mRNA levels of the sympa-thetically up-regulated genes FATP-1, D2, and UCP1.
Discussion
To address the question of why the PPAR�-mediated up-regulation of lipolytic, oxidative, and thermogenic machin-eries are not associated with a proportional increase in theirrespective biological processes in vivo, we investigated theimpact of the thiazolidinedione rosiglitazone on the majoractivators of BAT and WAT thermogenesis and lipolysis,namely the sympathetic nervous system and thyroid hor-mones. Chronic rosiglitazone treatment markedly reducedsympathetic activity to BAT and WAT, the thyroid status(plasma thyroid hormones and tissue mRNA levels of THRand deiodinases) and hypothalamic mRNA levels of the pro-energy expenditure peptides CART and CRH. The study alsosuggests that, although a small amount of rosiglitazone is
able to enter the brain, its effects on the sympathetic andthyroid axes are not mediated by a direct action in the centralnervous system.
The study confirms that the well-established positive ac-tions of rosiglitazone on insulin sensitivity and lipemia occurdespite increased body weight gain and adiposity (9, 10).Although such positive energy balance can partially be as-cribed to the increase in food intake, the marked increase infeed efficiency shown here and previously by us (4, 8) andothers (9, 10) strongly suggests that in vivo PPAR� decreasesenergy expenditure. Confirming this notion, rosiglitazoneinduced a small (7%) but significant reduction in O2 con-sumption. In fact, this represents an underestimation of suchmetabolic adaptation given the obligatory increase in energyexpenditure related to higher food intake and increased mo-tor activity elicited by rosiglitazone.
One germane aspect of the anabolic state induced by in vivoPPAR� activation is that it occurs simultaneously with anup-regulation of lipolytic, oxidative, and thermogenic pro-teins in WAT and BAT, which are functional, as evidencedin vitro in basal and stimulated conditions (5, 6) and in vivounder chronic �3-adrenergic stimulation (13). Supporting thehypothesis of a silencing of the thermogenic activity by thein vivo neurohormonal conditions, rosiglitazone markedlydecreased sympathetic activity to BAT and WAT and plasmathyroid hormone levels. That the latter occurred withoutchange in TSH or TRH suggests alternative modulation ofthyroid hormone secretion, the nature of which remains to bedetermined. Importantly, rosiglitazone reduced mRNA lev-els of liver D1, a sensitive marker of peripheral thyroid statusin rodents (30), and BAT D2, which generates biologicallyactive T3 from T4. BAT D2 is essential for thermogenesis due
FIG. 2. Effect of a 7-day rosiglitazone (RSG)treatment on BAT and liver mRNA levels of D2and D1 (A and E, respectively) and on BAT, liver,and RETRO mRNA levels of THR� (B, F, and I,respectively), -�1 (C, G, and J, respectively), and-�2 (D, H, and L, respectively). *, P � 0.05 vs.control rats.
2126 Endocrinology, May 2008, 149(5):2121–2130 Festuccia et al. • PPAR� Activation and Energy Balance
to the important role of locally produced T3 in the amplifi-cation of BAT adrenergic signaling and in the direct activa-tion of UCP1 expression (16, 31). Sympathetic regulation ofD2 is well established, and the reduction in its mRNA levelsseen here is more likely due to rosiglitazone-induced inhi-bition of BAT sympathetic activity than to a direct PPAR�action on its promoter (32). In addition to reducing BAT localT3 production, rosiglitazone inhibited the expression ofTHR�1 and THR�, which mediate the T3 effects on ampli-fication of adrenergic signaling and on direct activation ofUCP1 expression, respectively (33). Similarly to BAT, ros-iglitazone also reduced RETRO mRNA levels of THR�1,which mediates in WAT the stimulatory effects of circulatingT3 on adrenergic responsiveness and lipolysis (34). It cantherefore be suggested that the reduction in sympatheticactivity and thyroid status accounts for the dissociation be-tween gene expression and biological processes observed invivo in rosiglitazone-treated rats.
The robust inhibitory effect of rosiglitazone on sympa-thetic activity and thyroid status has important functional
consequences on both BAT and WAT. Under PPAR� acti-vation, BAT shifts from a thermogenic toward a lipid storagefunction, as evidenced here by the marked increase in tissueTAG content and number of unilocular brown adipocytes,and by the strong stimulatory action of rosiglitazone onuptake of circulating lipids by BAT (35). This shift in BATfunction occurs in the presence of high levels of UCP1, uni-versally considered as a marker of BAT sympathetic activa-tion and thermogenesis. This dissociation between BAT ther-mogenic capacity (up-regulation of thermogenic proteins)and functional activity, however, is not without precedent. InSyrian hamsters either exposed to a short photoperiod or feda high-energy diet (36, 37) and in cold-acclimated rats rein-troduced to a warm environment (38), abundant UCP1 doesnot result in increased thermogenesis because of the absenceof sympathetic activation. This is perhaps related to inade-quate provision of lipolytic products (fatty acids) for ther-mogenesis in the absence of sympathetic activity and thyroidhormones, as strongly suggested by defective cold adapta-tion in adipose triglyceride lipase-deficient mice, which dis-play reduced lipolysis (39). The same concept can also beapplied to the PPAR� agonist-induced increase in the lipo-lytic machinery of WAT (8), which, likely because of reducedsympathetic activity and thyroid status, and insulin sensiti-zation, does not result in increased fatty acid release in vivo(40). Furthermore, inhibition of sympathetic activity in WAT
FIG. 3. Effect of a 7-d rosiglitazone (RSG) treatment on BAT UCP1mRNA levels (A), TAG content (B), and percentage of unilocularadipocytes (C) and on RETRO WAT UCP1 mRNA levels (E), DNAcontent (F), and adipocyte diameter (G). D and H are representativephotomicrographs of BAT and WAT sections, respectively, dyed withhematoxylin/eosin. *, P � 0.05 vs. control rats.
FIG. 4. Effect of a 7-d rosiglitazone treatment on whole hypothalamicneuropeptide relative mRNA levels and on arcuate CART and para-ventricular CRH mRNA levels measured by quantitative PCR (A) andby in situ hybridization (B and C). In A, control and RSG hypothalamicneuropeptide mRNA levels were normalized for the housekeepingGAPDH. Control values were transformed to 1 (horizontal line). In Band C, OD values of the hybridization signal of CRH and CART mRNAare presented together with representative photographs of the in situhybridization. *, P � 0.05 vs. control rats; n � 6.
Festuccia et al. • PPAR� Activation and Energy Balance Endocrinology, May 2008, 149(5):2121–2130 2127
might well be involved in the rosiglitazone-induced increasein DNA content and reduction in adipocyte diameter ob-served here; both are indicative of cell proliferation, a processthat is strongly inhibited by the sympathetic drive to WAT(41, 42).
To gain further insight into the mechanisms by whichPPAR� activation reduces sympathetic activity to rat ad-ipose tissues, we screened the hypothalamus for changesin the expression of several neuropeptides known to affectwhole-body energy balance. Rosiglitazone treatment wasassociated with significant reductions in hypothalamicCRH and CART mRNA levels that were further localizedto the pPVN and the ARC, respectively. Although hypo-thalamic CRH is related to changes in BAT sympatheticoutflow (43), our finding that CRH was mainly reduced inneurons of the pPVN is not consistent with its participa-tion in the rosiglitazone effects on sympathetic activity.
These neurons indeed mainly control pituitary ACTH se-cretion (44), as confirmed here by lower plasma ACTHlevels, which further translate into lower adrenal weight(45), an index of long-term hypothalamic-pituitary-adre-nal axis activity, in rosiglitazone-treated rats comparedwith controls. In contrast to parvocellular CRH, arcuateCART-expressing neurons are anatomically linked to BATsympathetic nerves (46). Adenovirus-mediated CARToverexpression and peptide injection in the ARC increaseBAT sympathetic outflow (47), further supporting the pos-sible involvement of arcuate CART neurons in the ros-iglitazone-induced inhibition of the sympathetic drive toBAT.
Because a variety of signals can impact hypothalamic neu-ropeptide expression, we tested the hypothesis of whetherrosiglitazone could cross the blood-brain barrier to act di-rectly in the brain. We found that about 0.045% of an ivinjected dose of rosiglitazone was taken up per gram of brain.However, the in vivo mouse studies showed a paradoxicalincrease in the uptake of [3H]rosiglitazone when unlabeledrosiglitazone was included in the injection, suggestive that abrain-to-blood efflux system pumps rosiglitazone out of thebrain (48). We confirmed in both an in vivo mouse model andin an in vitro human brain endothelial cell model using thespecific inhibitor PSC833 (49) that rosiglitazone is a substratefor the brain-to-blood efflux system P-gp.
The modest uptake of rosiglitazone from blood and itsrapid, transporter-mediated efflux from the brain are notsupportive of a direct effect of the agonist on the centralnervous system. Although transport kinetics may conceiv-ably be modified with long-term treatment, a lack of directcentral action is confirmed by the absence of change in foodintake and energy expenditure after 1 wk of icv infusion ofrosiglitazone. This suggests instead that rosiglitazone affectsfood intake, sympathetic activity, locomotor activity, andenergy balance by eliciting a peripheral signal that thenreaches the brain. Although possible mediators are specu-lative at present, adiponectin, the plasma levels of which aremarkedly increased by PPAR� activation as confirmed here,constitutes an attractive candidate. Although still debated,recent evidence suggests an involvement of adiponectin inenergy balance. Adiponectin overexpression results inmarked positive energy balance even on an obese ob/obbackground (50), whereas mice lacking adiponectin displaylower body weight gain than their wild-type counterparts inresponse to a PPAR� agonist (51). Furthermore, similarly torosiglitazone, adiponectin has recently been shown to in-crease food intake, to reduce both energy expenditure (52)and renal sympathetic activity (53), and to increase locomo-tion (54). Noteworthy, these central adiponectin effects seemto at least partly take place in the ARC (52), in which CARTexpression was decreased by rosiglitazone in the presentstudy. Whether adiponectin mediates the effects of rosigli-tazone on the central regulation of energy balance awaitsdirect demonstration.
In conclusion, this study presents strong evidence sug-gesting that the increased energy efficiency and energy stor-age associated with PPAR� activation in rodents are medi-ated, at least in part, by a down-regulation of sympatheticactivity to BAT and WAT and thyroid status, likely a con-
FIG. 5. A, Murine brain/serum ratios of [3H]rosiglitazone (3H-RSG);addition of unlabeled rosiglitazone (�RSG) paradoxically increasedbrain uptake of [3H]rosiglitazone immediately after their iv injection.B, Percentage of the iv injected dose of [3H]rosiglitazone taken up pergram of brain; unlabeled RSG did not affect clearance from blood(inset) but confirmed that it did increase brain uptake. C, In vivoinhibition of brain-to-blood transport of [3H]rosiglitazone by unla-beled rosiglitazone; this confirms that rosiglitazone is effluxed frombrain by a saturable transporter. D, Inhibition of [3H]rosiglitazoneefflux by the P-gp inhibitor PSC833; this confirms that the effluxsystem for rosiglitazone is P-gp. E, In vitro [3H]rosiglitazone transportacross HBMEC monolayer model of the blood-brain barrier; influxand especially efflux of [3H]rosiglitazone exceeds that of the poorlypermeable control, sucrose. F, In vitro model efflux is inhibited byP-gp inhibitors PSC833 and CsA. *, P � 0.05 vs. [3H]rosiglitazone (C,D, and F) or influx (E); n � 3–4.
2128 Endocrinology, May 2008, 149(5):2121–2130 Festuccia et al. • PPAR� Activation and Energy Balance

sequence of an indirect (peripheral) action of PPAR� acti-vation on central modulators of energy balance.
Acknowledgments
We are very grateful for the invaluable professional assistance ofMelanie Alain, Sebastien Poulin, Josee Lalonde, Yves Gelinas, PierreSamson, and Emily Kelso.
Received November 12, 2007. Accepted January 14, 2008.Address all correspondence and requests for reprints to: Dr. Yves Deshaies,
Faculty of Medicine, Laval University, Laval Hospital Research Centre, LavalHospital–d’Youville Y3110, 2725 Chemin Sainte-Foy, Quebec, Canada G1V4G5. E-mail: [email protected].
This work was supported by a grant from the Canadian Institutes ofHealth Research (CIHR) to Y.D. and Veterans Affairs Merit Review andR010513334 to W.A.B. W.T.F. and R.G.D. were recipients of a Postdoc-toral Fellowship from the CIHR-funded Obesity Research Training Pro-gram led by the Laval Hospital Research Center. M.L. and M.B. heldstudentships from the Natural Sciences and Engineering Research Coun-cil of Canada and the CIHR-funded Obesity Research Training Program,respectively.
This manuscript is dedicated to the memory of Dr. Renato HéliosMigliorini.
Disclosure Statement: The authors have nothing to disclose.
References
1. Semple RK, Chatterjee VK, O’Rahilly S 2006 PPAR � and human metabolicdisease. J Clin Invest 116:581–589
2. Olefsky JM 2000 Treatment of insulin resistance with peroxisome proliferator-activated receptor � agonists. J Clin Invest 106:467–472
3. Yamauchi T, Kamon J, Waki H, Murakami K, Motojima K, Komeda K, IdeT, Kubota N, Terauchi Y, Tobe K, Miki H, Tsuchida A, Akanuma Y, NagaiR, Kimura S, Kadowaki T 2001 The mechanisms by which both heterozygousperoxisome proliferator-activated receptor � (PPAR�) deficiency and PPAR�agonist improve insulin resistance. J Biol Chem 276:41245–41254
4. Berthiaume M, Sell H, Lalonde J, Gelinas Y, Tchernof A, Richard D, De-shaies Y 2004 Actions of PPAR� agonism on adipose tissue remodeling, insulinsensitivity, and lipemia in absence of glucocorticoids. Am J Physiol RegulIntegr Comp Physiol 287:R1116–R1123
5. Wilson-Fritch L, Nicoloro S, Chouinard M, Lazar MA, Chui PC, Leszyk J,Straubhaar J, Czech MP, Corvera S 2004 Mitochondrial remodeling in adiposetissue associated with obesity and treatment with rosiglitazone. J Clin Invest114:1281–1289
6. Teruel T, Hernandez R, Rial E, Martin-Hidalgo A, Lorenzo M 2005 Rosigli-tazone up-regulates lipoprotein lipase, hormone-sensitive lipase and uncou-pling protein-1, and down-regulates insulin-induced fatty acid synthase geneexpression in brown adipocytes of Wistar rats. Diabetologia 48:1180–1188
7. van Harmelen V, Dicker A, Ryden M, Hauner H, Lonnqvist F, Naslund E,Arner P 2002 Increased lipolysis and decreased leptin production by humanomental as compared with subcutaneous preadipocytes. Diabetes 51:2029–2036
8. Festuccia WT, Laplante M, Berthiaume M, Gelinas Y, Deshaies Y 2006PPAR� agonism increases rat adipose tissue lipolysis, expression of glyceridelipases, and the response of lipolysis to hormonal control. Diabetologia 49:2427–2436
9. Burkey BF, Dong M, Gagen K, Eckhardt M, Dragonas N, Chen W, Grosen-stein P, Argentieri G, de Souza CJ 2000 Effects of pioglitazone on promotingenergy storage, not expenditure, in brown adipose tissue of obese fa/fa Zuckerrats: comparison to CL 316,243. Metabolism 49:1301–1308
10. Larsen PJ, Jensen PB, Sorensen RV, Larsen LK, Vrang N, Wulff EM, Was-
sermann K 2003 Differential influences of peroxisome proliferator-activatedreceptors-� and -� on food intake and energy homeostasis. Diabetes 52:2249–2259
11. Akazawa S, Sun F, Ito M, Kawasaki E, Eguchi K 2000 Efficacy of troglitazoneon body fat distribution in type 2 diabetes. Diabetes Care 23:1067–1071
12. Joosen AM, Bakker AH, Gering MJ, Westerterp KR 2006 The effect of thePPAR� ligand rosiglitazone on energy balance regulation. Diabetes Metab ResRev 22:204–210
13. Sell H, Berger JP, Samson P, Castriota G, Lalonde J, Deshaies Y, Richard D2004 Peroxisome proliferator-activated receptor � agonism increases the ca-pacity for sympathetically mediated thermogenesis in lean and ob/ob mice.Endocrinology 145:3925–3934
14. Thurlby PL, Wilson S, Arch JR 1987 Ciglitazone is not itself thermogenic butincreases the potential for thermogenesis in lean mice. Biosci Rep 7:573–577
15. Cannon B, Nedergaard J 2004 Brown adipose tissue: function and physio-logical significance. Physiol Rev 84:277–359
16. Silva JE 2006 Thermogenic mechanisms and their hormonal regulation.Physiol Rev 86:435–464
17. Moreno S, Farioli-Vecchioli S, Ceru MP 2004 Immunolocalization of perox-isome proliferator-activated receptors and retinoid X receptors in the adult ratCNS. Neuroscience 123:131–145
18. Shi H, Bowers RR, Bartness TJ 2004 Norepinephrine turnover in brown andwhite adipose tissue after partial lipectomy. Physiol Behav 81:535–542
19. Collins S, Kuhn CM, Petro AE, Swick AG, Chrunyk BA, Surwit RS 1996 Roleof leptin in fat regulation. Nature 380:677
20. Penn DM, Jordan LC, Kelso EW, Davenport JE, Harris RB 2006 Effects ofcentral or peripheral leptin administration on norepinephrine turnover indefined fat depots. Am J Physiol Regul Integr Comp Physiol 291:R1613–R1621
21. Brito MN, Brito NA, Garofalo MA, Kettelhut IC, Migliorini RH 1998 Sym-pathetic activity in brown adipose tissue from rats adapted to a high protein,carbohydrate-free diet. J Auton Nerv Syst 69:1–5
22. Migliorini RH, Garofalo MA, Kettelhut IC 1997 Increased sympathetic ac-tivity in rat white adipose tissue during prolonged fasting. Am J Physiol272:R656–R661
23. Brodie BB, Costa E, Dlabac A, Neff NH, Smookler HH 1966 Application ofsteady state kinetics to the estimation of synthesis rate and turnover time oftissue catecholamines. J Pharmacol Exp Ther 154:493–498
24. Arvaniti K, Richard D, Picard F, Deshaies Y 2001 Lipid deposition in ratscentrally infused with leptin in the presence or absence of corticosterone. Am JPhysiol Endocrinol Metab 281:E809–E816
25. Timofeeva E, Richard D 1997 Functional activation of CRH neurons andexpression of the genes encoding CRH and its receptors in food-deprived lean(Fa/?) and obese (fa/fa) Zucker rats. Neuroendocrinology 66:327–340
26. Wang Q, Dryden S, Frankish HM, Bing C, Pickavance L, Hopkins D, Buck-ingham R, Williams G 1997 Increased feeding in fatty Zucker rats by thethiazolidinedione BRL 49653 (rosiglitazone) and the possible involvement ofleptin and hypothalamic neuropeptide Y. Br J Pharmacol 122:1405–1410
27. Banks WA, Kastin AJ 1993 Measurement of transport of cytokines across theblood-brain barrier. In: Souza EB, ed. Methods in neurosciences. Vol 16, SanDiego: Academic Press; 67–77
28. Banks WA, Kastin AJ 1989 Quantifying carrier-mediated transport of peptidesfrom the brain to the blood. Methods Enzymol 168:652–660
29. Dehouck MP, Jolliet-Riant P, Bree F, Fruchart JC, Cecchelli R, Tillement JP1992 Drug transfer across the blood-brain barrier: correlation between in vitroand in vivo models. J Neurochem 58:1790–1797
30. Zavacki AM, Ying H, Christoffolete MA, Aerts G, So E, Harney JW, ChengSY, Larsen PR, Bianco AC 2005 Type 1 iodothyronine deiodinase is a sensitivemarker of peripheral thyroid status in the mouse. Endocrinology 146:1568–1575
31. de Jesus LA, Carvalho SD, Ribeiro MO, Schneider M, Kim SW, Harney JW,Larsen PR, Bianco AC 2001 The type 2 iodothyronine deiodinase is essentialfor adaptive thermogenesis in brown adipose tissue. J Clin Invest 108:1379–1385
32. Silva JE, Larsen PR 1983 Adrenergic activation of triiodothyronine productionin brown adipose tissue. Nature 305:712–713
33. Ribeiro MO, Carvalho SD, Schultz JJ, Chiellini G, Scanlan TS, Bianco AC,
TABLE 3. Effect of a 7-d icv vehicle or rosiglitazone (RSG) infusion on body weight gain, daily food intake, RETRO adipose tissue andBAT mass, and BAT FATP-1, D2, and UCP1 mRNA levels
Vehicle RSG (50 �g/ml) RSG (500 �g/ml)
Body weight gain (g) 54 � 1 56 � 2 52 � 0.3Food intake (g/d) 25.0 � 0.8 25.8 � 1.9 25.4 � 0.8RETRO mass (g) 1.37 � 0.04 1.35 � 0.12 1.32 � 0.02BAT mass (g) 0.23 � 0.03 0.24 � 0.02 0.25 � 0.02BAT FATP-1/L27 mRNA 2.2 � 0.3 2.5 � 0.3 2.2 � 0.3BAT D2/L27 mRNA 2.0 � 0.3 1.9 � 0.1 2.3 � 0.2BAT UCP1/L27 mRNA 4.0 � 0.6 4.0 � 0.4 4.0 � 0.6
Data are means � SEM of four rats.
Festuccia et al. • PPAR� Activation and Energy Balance Endocrinology, May 2008, 149(5):2121–2130 2129

Brent GA 2001 Thyroid hormone-sympathetic interaction and adaptive ther-mogenesis are thyroid hormone receptor isoform-specific. J Clin Invest 108:97–105
34. Liu YY, Schultz JJ, Brent GA 2003 A thyroid hormone receptor � gene mu-tation (P398H) is associated with visceral adiposity and impaired catechol-amine-stimulated lipolysis in mice. J Biol Chem 278:38913–38920
35. Laplante M, Festuccia WT, Soucy G, Gelinas Y, Lalonde J, Deshaies Y 2007Involvement of adipose tissues in the early hypolipidemic action of PPAR�agonism in the rat. Am J Physiol Regul Integr Comp Physiol 292:R1408–R1417
36. Sigurdson SL, Himms-Hagen J 1988 Control of norepinephrine turnover inbrown adipose tissue of Syrian hamsters. Am J Physiol 254:R960–R968
37. McElroy JF, Wade GN 1986 Short photoperiod stimulates brown adiposetissue growth and thermogenesis but not norepinephrine turnover in Syrianhamsters. Physiol Behav 37:307–311
38. Griggio MA 1982 The participation of shivering and nonshivering thermo-genesis in warm and cold-acclimated rats. Comp Biochem Physiol A 73:481–484
39. Haemmerle G, Lass A, Zimmermann R, Gorkiewicz G, Meyer C, Rozman J,Heldmaier G, Maier R, Theussl C, Eder S, Kratky D, Wagner EF, KlingensporM, Hoefler G, Zechner R 2006 Defective lipolysis and altered energy metab-olism in mice lacking adipose triglyceride lipase. Science 312:734–737
40. Oakes ND, Thalen PG, Jacinto SM, Ljung B 2001 Thiazolidinediones increaseplasma-adipose tissue FFA exchange capacity and enhance insulin-mediatedcontrol of systemic FFA availability. Diabetes 50:1158–1165
41. Bowers RR, Festuccia WT, Song CK, Shi H, Migliorini RH, Bartness TJ 2004Sympathetic innervation of white adipose tissue and its regulation of fat cellnumber. Am J Physiol Regul Integr Comp Physiol 286:R1167–R1175
42. Foster MT, Bartness TJ 2006 Sympathetic but not sensory denervation stim-ulates white adipocyte proliferation. Am J Physiol Regul Integr Comp Physiol291:R1630–R1637
43. Egawa M, Yoshimatsu H, Bray GA 1990 Effect of corticotropin releasinghormone and neuropeptide Y on electrophysiological activity of sympatheticnerves to interscapular brown adipose tissue. Neuroscience 34:771–775
44. Swanson LW, Sawchenko PE, Lind RW 1986 Regulation of multiple peptidesin CRF parvocellular neurosecretory neurons: implications for the stress re-sponse.Prog Brain Res 68:169–190
45. Berthiaume M, Laplante M, Tchernof A, Deshaies Y 2007 Metabolic action of
peroxisome proliferator-activated receptor � agonism in rats with exogenoushypercorticosteronemia. Int J Obes (Lond) 31:1660–1670
46. Elias CF, Lee C, Kelly J, Aschkenasi C, Ahima RS, Couceyro PR, Kuhar MJ,Saper CB, Elmquist JK 1998 Leptin activates hypothalamic CART neuronsprojecting to the spinal cord. Neuron 21:1375–1385
47. Kong WM, Stanley S, Gardiner J, Abbott C, Murphy K, Seth A, ConnoleyI, Ghatei M, Stephens D, Bloom S 2003 A role for arcuate cocaine andamphetamine-regulated transcript in hyperphagia, thermogenesis, and coldadaptation. FASEB J 17:1688–1690
48. Banks WA 2005 Critical roles of efflux systems in health and disease. In: TaylorEM, ed. Efflux transporters and the blood-brain barrier. Hauppauge, NY: NovaScience Publishers; 21–53
49. Drion N, Lemaire M, Lefauconnier JM, Scherrmann JM 1996 Role of P-glycoprotein in the blood-brain transport of colchicine and vinblastine. J Neu-rochem 67:1688–1693
50. Kim JY, van de Wall E, Laplante M, Azzara A, Trujillo ME, Hofmann SM,Schraw T, Durand JL, Li H, Li G, Jelicks LA, Mehler MF, Hui DY, DeshaiesY, Shulman GI, Schwartz GJ, Scherer PE 2007 Obesity-associated improve-ments in metabolic profile through expansion of adipose tissue. J Clin Invest117:2621–2637
51. Nawrocki AR, Rajala MW, Tomas E, Pajvani UB, Saha AK, Trumbauer ME,Pang Z, Chen AS, Ruderman NB, Chen H, Rossetti L, Scherer PE 2006 Micelacking adiponectin show decreased hepatic insulin sensitivity and reducedresponsiveness to peroxisome proliferator-activated receptor � agonists. J BiolChem 281:2654–2660
52. Kubota N, Yano W, Kubota T, Yamauchi T, Itoh S, Kumagai H, Kozono H,Takamoto I, Okamoto S, Shiuchi T, Suzuki R, Satoh H, Tsuchida A, MoroiM, Sugi K, Noda T, Ebinuma H, Ueta Y, Kondo T, Araki E, Ezaki O, NagaiR, Tobe K, Terauchi Y, Ueki K, Minokoshi Y, Kadowaki T 2007 Adiponectinstimulates AMP-activated protein kinase in the hypothalamus and increasesfood intake. Cell Metab 6:55–68
53. Tanida M, Shen J, Horii Y, Matsuda M, Kihara S, Funahashi T, ShimomuraI, Sawai H, Fukuda Y, Matsuzawa Y, Nagai K 2007 Effects of adiponectin onthe renal sympathetic nerve activity and blood pressure in rats. Exp Biol Med(Maywood) 232:390–397
54. Saito K, Arata S, Hosono T, Sano Y, Takahashi K, Choi-Miura NH, NakanoY, Tobe T, Tomita M 2006 Adiponectin plays an important role in efficientenergy usage under energy shortage. Biochim Biophys Acta 1761:709–716
Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
2130 Endocrinology, May 2008, 149(5):2121–2130 Festuccia et al. • PPAR� Activation and Energy Balance





























