Embed Size (px)
Citation preview

655
ISSN 0031-0301, Paleontological Journal, 2008, Vol. 42, No. 6, pp. 655–664. © Pleiades Publishing, Ltd., 2008.Original Russian Text © T.N. Hermann, V.N. Podkovyrov, 2008, published in Paleontologicheskii Zhurnal, 2008, No. 6, pp. 81–88.
INTRODUCTION
The main problem of the paleontology of Precam-brian organisms is the determination of their systematicposition, nature, and the chronology of their emergenceand further evolution. Most of the Neoproterozoicmicrofossils are difficult to define taxonomically basedon their morphology. The evolutionary history ofmicroorganisms is still to be reconstructed, predomi-nantly by comparison with those modern forms oforganic life which are close to the fossil forms in mor-phology, mode of life, and (wherever possible) devel-opmental cycles.
This paper deals with the relationships of two mor-phologically distinct fossil microorganisms that belongto the form genera
Arctacellularia
Hermann, 1976 and
Glomovertella
Reitlinger, 1948. These microfossilswere revealed in the microbiota of the Miroedikha For-mation (Miroedikha River, Turukhansk region, north-western Siberia), the isotopic age is estimated to lie inthe range from 850 to 950 Ma (Knoll et al., 1995;Petrov and Semikhatov, 1997). Organic remains wereobtained by the standard palynological method of mac-eration in HF from thinly laminated clayey shales ofgray and dark gray color.
The Miroedikha microbiota is one of the richest bio-tas of the Late Riphean in terms of the diversity ofunmineralized organic-walled microfossils in a goodstate of preservation (Timofeev, 1966, 1969; Timofeevet al., 1976; Hermann, 1974, 1981, 1986, 1990; Veis,1988; Veis and Vorob’eva, 1998).
In the geological history of the Riphean of theTurukhansk region, the Miroedikha Formation corre-sponds to the eventful time interval that separated theperiods of the development of stable carbonate plat-forms. During this time interval the shallow-water set-tings of the inner shelf gave way to the deepwater set-ting of the outer shelf, thus making it possible to con-sider the deposits of the Miroedikha paleobasin as atransgressive complex (Petrov and Semikhatov, 1997).The facial-ecological distribution of microfossils andchange in the assemblages of organic remains of theentire section of the Late Riphean of the TurukhanskUplift is discussed in a number of publications (Veisand Petrov, 1994; Veis et al., 1998, 1999).
This microbiota is characterized by the extremeabundance of filamentous remains of cyanobacterialnature (Hermann, 1974, 1981), against the backgroundof which large fragments of chainlike aggregates con-sisting of cells shaped like ellipsoids, short barrels, andcylinders clearly stand out. These microfossils wererecognized as a separate genus
Arctacellularia
withtwo species:
A. ellipsoidea
Hermann, 1976 and
A. doli-iformis
Hermann, 1976 (Timofeev et al., 1976, pp. 38–39,pl. X, figs. 1–3, 5; pl. XIII, fig. 9). More recently twomore species of this genus (represented by single cells)were described:
A. kelleri
Hermann et Jankauskas and
A. varia
Yankauskas et Hermann; the latter is a syn-onym of the species
A. tetragonale
Maithy.
Chains and isolated cells of
Arctacellularia
are areadily identifiable Precambrian morphotype. Numer-ous finds of this genus revealed in clayey rocks of the
On the Nature of the Precambrian Microfossils
Arctacellularia
and
Glomovertella
T. N. Hermann and V. N. Podkovyrov
Institute of Precambrian Geology and Geochronology, Russian Academy of Sciences, nab. Makarova 2, St. Petersburg, 199034 Russia
e-mail: [email protected]
Received April 13, 2007
Abstract
—Possible phylogenetic relationships between the organic-walled microfossils of the form genus
Arctacellularia
Hermann, 1976, representing chainlike aggregates of cells, and the filaments of the genus
Glo-movertella
Reitlinger, 1948, which comprises variously coiled filaments, are discussed. The possible relation-ship of these microfossils with different morphologies to lower ascomycetes is discussed. The chains of
Arcta-cellularia
are interpreted as exogenously produced spores, and the filaments that are associated with them aretreated as vegetative hyphae that have a characteristic coiling during their transition phase, perhaps before theformation of sporophores. An emended diagnosis of the genus
Arctacellularia
and emended descriptions of thespecies
A. ellipsoidea
Hermann, 1976,
A. kelleri
Hermann et Jankauskas, 1989 are provided; and a new species,
Glomovertella miroedikhia
sp. nov., is described.
DOI:
10.1134/S0031030108060117
Key words
: microfossils, fungi, ascomycetes, Miroedikha Group, Upper Riphean, Siberia.
656
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 6
2008
HERMANN, PODKOVYROV
stratotype of the Upper Riphean of the Southern Urals(Bederyshinskaya Member of the Zilmerdak Formationand the Uk Formation; Jankauskas, 1982, pl. XXXV,figs. 4, 5, 9, 13; pl. XXXIV, figs. 6, 11; pl. XIII, figs. 9, 12).The Vychegda Formation of the Upper Riphean ofTiman yielded microfossils in association with theannelidomorphous Parmia (Gnilovskaya et al., 2000).
Arctacellularia
specimens were also recorded from theclayey facies of the Middle Riphean of Canada (BylotGroup, Baffin Island; Hofmann and Jackson, 1994,pl. 12, figs. 16–20) and the Late Precambrian of Zaire(Maithy, 1975).
Uniserial chains of ellipsoidal cells that are in closecontact with one another were assigned to the species
Arctacellularia ellipsoidea
Hermann, 1976. The cellshave characteristic thickenings, lenticular in the contactzone and spheroidal at the center. Narrow longitudinalcords can be observed along a chain of cells. In thedescription of the holotype of this species no mentionwas made of the fact (regarded then as merely coinci-dental) that a chain of cells was located in the vicinityof narrow straight and aseptate filaments slightly over-lapping one another (Pl. 8, fig. 1). More recently, thismicrobiota yielded specimens which show such a directassociation of chainlike aggregates with filaments,which are usually looped in the form of an elongateskein (Pl. 8, figs. 4, 5, 11). Filaments twisted into askein are located in close proximity and parallel to each
other, slightly overlapping one another (Pl. 8, figs. 9,13, 15). Repeated records of two interrelated organismswith different morphologies were the reason for a moredetailed study of such organic remains.
The species
Arctacellularia doliiformis
Hermann,1976 was also characterized by a uniserial chain of cellsthat varies in shape from ellipsoidal to short-barrel andcylindrical. The dividing walls of the cell are thickenedand lenticular. Narrow cords run along a chain of cells(Pl. 8, fig. 7). Hofmann and Jackson (1994) believe thatthis species is identical to
A. ellipsoidea
, since in thechain of the holotype of
A. doliiformis
the cells in theinitial developmental phase are ellipsoidal in shape(Pl. 8, fig. 7, a narrow portion of the chain). Accordingto Hofmann and Jackson, the cells of this species,which are of equal width, but of different lengths, maybe heterocysts or akinetes of cyanobacteria
Gloeotri-chia
. Although we concur that the two species are iden-tical, we propose our own, different, interpretation ofthe nature of these microfossils.
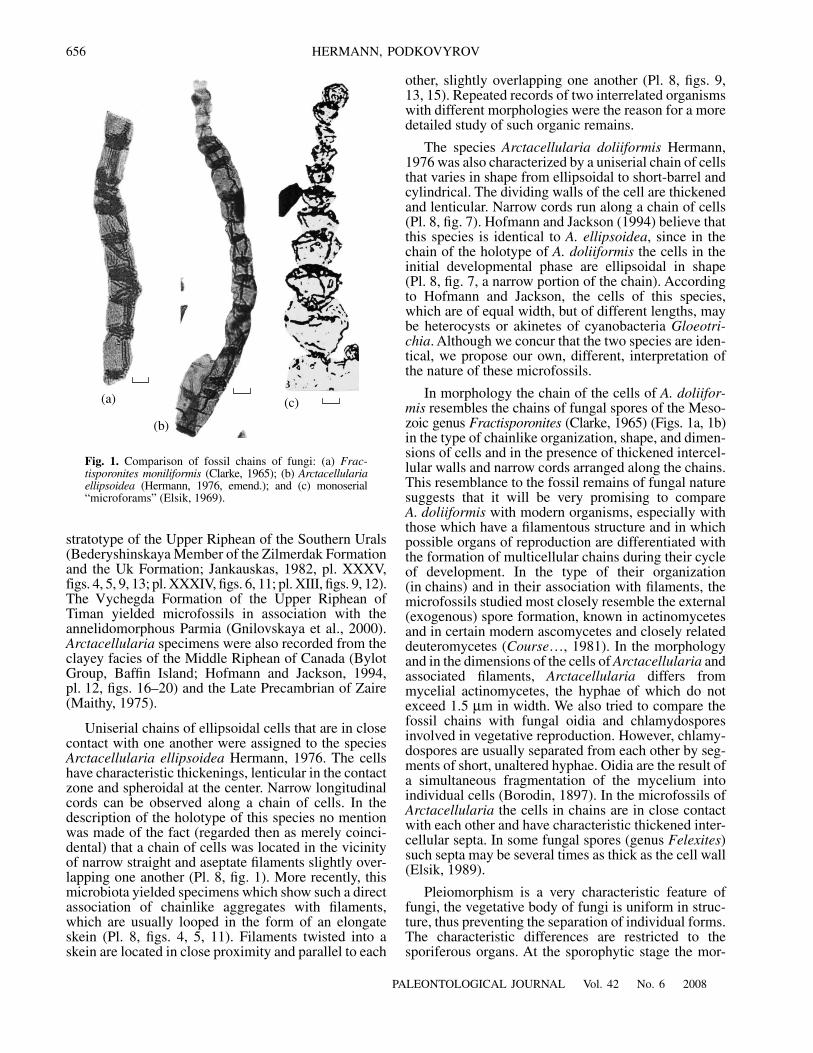
In morphology the chain of the cells of
A. doliifor-mis
resembles the chains of fungal spores of the Meso-zoic genus
Fractisporonites
(Clarke, 1965) (Figs. 1a, 1b)in the type of chainlike organization, shape, and dimen-sions of cells and in the presence of thickened intercel-lular walls and narrow cords arranged along the chains.This resemblance to the fossil remains of fungal naturesuggests that it will be very promising to compare
A. doliiformis
with modern organisms, especially withthose which have a filamentous structure and in whichpossible organs of reproduction are differentiated withthe formation of multicellular chains during their cycleof development. In the type of their organization(in chains) and in their association with filaments, themicrofossils studied most closely resemble the external(exogenous) spore formation, known in actinomycetesand in certain modern ascomycetes and closely relateddeuteromycetes (
Course
…, 1981). In the morphologyand in the dimensions of the cells of
Arctacellularia
andassociated filaments,
Arctacellularia
differs frommycelial actinomycetes, the hyphae of which do notexceed 1.5
µ
m in width. We also tried to compare thefossil chains with fungal oidia and chlamydosporesinvolved in vegetative reproduction. However, chlamy-dospores are usually separated from each other by seg-ments of short, unaltered hyphae. Oidia are the result ofa simultaneous fragmentation of the mycelium intoindividual cells (Borodin, 1897). In the microfossils of
Arctacellularia
the cells in chains are in close contactwith each other and have characteristic thickened inter-cellular septa. In some fungal spores (genus
Felexites
)such septa may be several times as thick as the cell wall(Elsik, 1989).
Pleiomorphism is a very characteristic feature offungi, the vegetative body of fungi is uniform in struc-ture, thus preventing the separation of individual forms.The characteristic differences are restricted to thesporiferous organs. At the sporophytic stage the mor-
(a)
(b)
(c)
Fig. 1.
Comparison of fossil chains of fungi: (a)
Frac-tisporonites moniliformis
(Clarke, 1965); (b)
Arctacellulariaellipsoidea
(Hermann, 1976, emend.); and (c) monoserial“microforams” (Elsik, 1969).

PALEONTOLOGICAL JOURNAL
Vol. 42
No. 6
2008
ON THE NATURE OF THE PRECAMBRIAN MICROFOSSILS 657
phology of a fungus may change to the extent that inde-pendent phases of the development of fungi aredescribed under different specific and generic names.
In the case of asexual reproduction, as in some mod-ern ascomycetes, the chains of spores (conidia) formexogenously. Usually conidia are originally unicellular;however, quite often they become secondary multicel-lular because of the septa that they evolve. The ascalstage is rare in some species of these fungi. This stageis difficult both to observe in nature and to produce arti-ficially. Many ascomycetes more frequently occur in theconidial stage. The ascospores are of diverse shape: fromgloboid or ellipsoidal to cylindrical. Morphologically theconidia markedly differ from the vegetative hyphae, whichhave a tendency to a tight interlacing when they form thesporiferous organs (
Course…
,
1981).
Hypothetically, the chains of cells of the species
A. ellipsoidea
may represent the chains of exosporesthat formed during the fragmentation of the sporiferoushyphae by breaking off its short portions capable ofrapid rounding and developing a covering of its own(
Course
…, 1981). In certain modern ascomycetes thespecialized hyphae (conidiophores) bear the chains ofspores (arthrospores) and may be associated withimmature perithecia. After reaching their maximummaturity, the arthrospores assume the shape of a cubewith slightly rounded vertices and serve as “reinfectionagents” that reinfect the spores (Taylor et al., 2004).
Another process of the formation of exosporescould proceed by the segmentation of sporiferous fila-ments, when at a certain stage of their development thefilaments are partitioned by transversely running divid-ing walls into separate cells with walls that also becomethicker, slightly more rounded, and transform into iso-lated spores. During the formation of exospores in thesporiferous zone, the septa appear simultaneously orwithin short time intervals in the direction from theupper to the lower portion of the hyphae or, occasion-ally, in the reverse direction (
Course
…, 1981). Thedevelopment of exospores of the species
A. doliiformis
[=
A. ellipsoidea
] (Pl. 8, fig. 7) could follow this type ofspore formation. In the narrowed part of this specimen,there is a division into small oval-shaped cells. Towardsthe end of the filament the shape and dimensions of thecells markedly change, possibly reflecting stages in thegradual development of arthrospores (Taylor et al.,2004). The chains of exospores (arthrospores) couldbreak into fragments with different numbers of cells(Pl. 8, figs. 2, 3, 10, 17), as well as into single largercells (mature arthrospores) (
Arctacellularia kelleri
Hermann et Jankauskas, 1989; Pl. 8, figs. 6, 8, 14, 16;Pl. 9, figs. 13–16).
Fossils of variously coiled filaments have beenrepeatedly recorded by different authors during thestudies of Precambrian microbiota. The records of fila-ments coiled in spiral and annular patterns are espe-cially numerous in the Vendian (Timofeev, 1973; Ase-eva, 1974; Golovenok and Belova, 1983; Kolosov,
1984; Burzin, 1995; etc.). Reitlinger (1948) was thefirst to provide the description of calcified filamentsthat are coiled in the spiral and skein patterns, hedescribed them from the deposits of the Sinian Forma-tion of the Lower Cambrian on the northern slope of theAldan Shield in Siberia under the generic names
Obruchevella
and
Glomovertella.
The genus
Obruchev-ella
was placed in the family Girvaneliaceae of the classHormogoniophyceae of cyanobacteria, whereas thegenus
Glomovertella
cannot be placed in the system ofcalcareous algae (
Fossil
…, 1987). The tightly coiledfilaments of
Glomovertella
apparently reflect a differ-ent type of behavior at a certain phase of the life cycle.In this instance the new species
G. miroedikhia
sp. nov.is interpreted as a temporary phase of the transition ofthe vegetative hyphae to the exogenous spore formationwith a tendency to the loop-shaped interlacing of them.
The liability of fossil filaments to bending, curling,and twisting does not necessarily result from tapho-nomical changes. The type of their behavior, in partic-ular variations in coiling, and the possible associationwith other organisms of different morphology, arereflections of specific biological properties in the pro-cess of the development of such organisms. The degreeand character of aggregation is among the most impor-tant properties of microorganisms associated with theunity of the rhythm of life, for example, with periods ofreproduction. The temporary association of microor-ganisms with each other is due to the temporality of thecontinuous progress of the processes of growth anddevelopment of cells, their division, aggregation, anddifferentiation (Kondrat’eva, 1989).
It is believed that any species of living organisms,irrespective of its systematic position, represents adynamic structurally multidimensional self-replicatingsystem consisting of individuals with varying qualita-tive characteristics and different ranks (
BiologicalCybernetics
, 1977), among which the local populationserves as an evolutionary unit. The study of the popula-tion structure of species and the structure of local pop-ulations ranks among the most important problems ofpopulation biology, systematics, and biogeography.
The microfossils of
Arctacellularia
and
Glomover-tella miroedikhia
sp. nov. are assumed to be organismsof fungal nature at different stages of their life cyclethat are closely related to each other and are probablyclose to ascomycetes of filamentous structure. Thechains are interpreted as the conidial stage of spore for-mation that is temporarily associated with the remainsof hyphae in the ontogenetic development. Both organ-isms at some time could be stable or labile, developing,capable of transformation, and at a certain phase of theirdevelopment could exist independently of each other.
Among the lower organisms, fungi are the mostactively metabolizing eukaryotes that possess quitediverse methods of reproduction, high reproductiveperformance, and are capable of developing in all envi-ronmental conditions. Fungi have no need of light, con-

658
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 6
2008
HERMANN, PODKOVYROV
Plate 8
20
µ
m 20
µ
m
20
µ
m 10
µ
m
10
µ
m
10
µ
m
1‡ 1b
1c
4
2
17
765
3
9
10
161513 1412
811

PALEONTOLOGICAL JOURNAL
Vol. 42
No. 6
2008
ON THE NATURE OF THE PRECAMBRIAN MICROFOSSILS 659
tain no chlorophyll, and can grow in all places wherethere are sources of organic matter. They are extremelypersistent and perform an important biochemical role inthe Earth’s biosphere.
The study of fossil fungi started at the end of the19th century (Felix, 1894). This subject received moreconsiderable study after the publication of the series ofpapers on the taxonomic and ecological diversity offossil fungi (Edwards, 1922; Hammen, 1955; Benesh,1960; Varma and Rawat, 1963; Clarke, 1965; Dilcher,1965).
In our country the study of fossil fungi is associatedwith the study of coals macerated by the method devel-oped by Grichuk (1940). Chiguryaeva (1956) publishedfigures of Tertiary fungi in “Atlas of Microspores fromthe Tertiary Deposits of the USSR.” Slightly laterPopov (1959, 1962, 1967, etc.) reported the records offossil fungi and pointed to the importance of the studyof these organisms and their role in the study of stratig-raphy and lithogenesis and in the reconstruction ofpaleoclimate, phylogenies, and the physical geographicenvironment of the past.
More or less as algae, fossil fungal remains arewidespread in time and space. The greatest number offungi has been described from the Jurassic and youngerdeposits. They were also recorded from the Precam-brian (Timofeev, 1970; Tiffney and Barghoorn, 1974).Fungal filaments were separated during the studies ofmicrofossils from the deposits of the Bitter SpringsFormation of central Australia (age about 850 Ma),from where Schopf (1968) described the narrow fila-ments of
Eomycetopsis
and gave them a name that sug-gests a fungal nature of these fossils. Timofeev et al.(1976) discovered Phycomycetes, large (200–500
µ
min diameter) cells with “tails,” in the Lakhanda Forma-tion of the Upper Riphean Uchur–Maya region in Sibe-ria (age 1020–1030 Ma, Semikhatov et al., 2000).
Subsequently the same deposits yielded a richassemblage of organic remains of unique preservationthat were very diverse morphologically (Timofeev andHermann, 1979). Some microfossils were interpretedas remains of Myxomycetes, Zygomycetes, and Asco-mycetes (Hermann, 1979; Hermann and Podkovyrov,2006). Burzin (1993) described the earliest Chytridio-mycetes from the deposits of the Upper Vendian. In the
Early Neoproterozoic Wynniatt Formation, VictoriaIsland, Canada (age 723 Ma) acanthomorphic acri-tarchs of two form genera
Tappania
and
Germino-sphaera
were described as a single natural taxon
Tap-pania
, which is probably closely related to one of thebranches of ascomycetes (Butterfield, 2005). The mostancient specimens of
Tappania
were discovered in theRoper Group of Australia, in deposits with an age of1430 million years (Yin, 1997, Yin et. al., 2005). Belovaand Akhmedov (2006) drew attention to the presence ofthe remains of fungi in even more ancient deposits withan age of 1.9 billion years.
The modern advancements in the use of the molec-ular clock allow more accurate age determinations ofthe most important evolutionary events in the Precam-brian. At present there are slightly different estimates ofthe degree of divergence between different groups ofeukaryotic organisms. Molecular phylogeny supportsthe concept of a common ancestor of animals and fungi.It is suggested that the group Opisthokonta (animals +choanozoa + fungi) diverged from their sister groupAmoebozoa at least 750 Ma ago (Porter et al., 2003).The molecular clock shows that the divergence of fungiand metazoa occurred 1500 million years ago (Hegeset al., 2004), and green algae and main branches offungi participated in the early colonization of theEarth’s land surface (1 billion years ago) (Heckmanet al., 2001). The group of researchers from 35 scien-tific institutions of different countries carried out theproject “Reconstruction of the Early Evolution of Fungiusing a Six-Gene Phylogeny.” According to their esti-mate, the line shared by fungi and animals divergedfrom the other eukaryotes about 1 billion years ago, andfungi and plants diverged about 600 million years ago(James et al., 2006). The most intriguing suggestionconsists of the revision of the eukaryotic tree, in whichthe ancestral stem of fungi is placed in the Archean(Martin et al., 2003).
Systematic Paleontology provides an emendeddiagnosis of the genus
Arctacellularia
and the descrip-tion of the species
A. ellipsoidea
Hermann, 1976;
A. kelleri
Hermann et Jankauskas, 1989; and
Glomov-ertella miroedikhia
sp. nov.
E x p l a n a t i o n o f P l a t e 8
Figs. 1–5, 7, 10–12, and 17.
Arctacellularia ellipsoidea
Hermann, 1979: (1) chain of cells is associated with the remains? of asporiferous filament (hypha); (1) specimen no. 49/24-XI.72: (1a) general appearance, (1b, 1c) magnified fragments; (2, 17) frag-ments of isolated chains in which some cells divided into two cells: (2) specimen no. 49/8-XI.77; (17) specimen no. 49/8-XI.72;(3, 7, 10) fragments of isolated chains with narrow longitudinal cords: (3) specimen no. 49/8-XI.77; (7) specimen no. 49/45-XI.72;(10) specimen no. 49/57-XI.72; (4, 5, 11, 12) chains of cells associated with filaments coiled in loop-shaped pattern: (4) specimenno. 49/59-XI.72, (5) specimen no. 49/22-XI.72, (11) specimen no. 49/22-XI.72, (12) specimen no. 49/3-XI.72.
Figs. 6, 8, 14, and 16.
Arctacellularia kelleri
Jankauskas et Hermann, 1989: (6) specimen no. 49/25-XI.72; (8) specimen no. 49/46-XI.72;(14) specimen no. 49/3-XI.72; (16) specimen no. 49/7-XI.72.
Figs. 9, 13, and 15.
Glomovertella miroedikhia
sp. nov.: (9) specimen no. 49/3-XI.72, (13) holotype no. 49/35-XI.72, (15) specimenno. 49/5-XI.72.Scale bar 10
µ
m, except for otherwise indicated.
660
PALEONTOLOGICAL JOURNAL
Vol. 42
No. 6
2008
HERMANN, PODKOVYROV
Plate 9
1 2 3
654
7 8 9
1011 12
13 14 15 16 17 18

PALEONTOLOGICAL JOURNAL
Vol. 42
No. 6
2008
ON THE NATURE OF THE PRECAMBRIAN MICROFOSSILS 661
MATERIAL
The collection of microfossils is housed in the Insti-tute of Precambrian Geology and Geochronology of theRussian Academy of Sciences (IGGD), St. Petersburg(coll. 49).
SYSTEMATIC PALEONTOLOGY
Genus
Arctacellularia Hermann, 1976
Arctacellularia Hermann: Timofeev et al., 1976, p. 37.
Ty p e s p e c i e s. A. ellipsoidea Hermann, 1976;Upper Riphean, Miroedikha Formation; MiroedikhaRiver, Turukhansk region, northwestern Siberia, Russia.
D i a g n o s i s. Uniserial, unbranched multicellularchains, devoid of external covering, formed by cells inclose contact with each other, varying in shape formellipsoidal to barrel-shaped or cylindrical. Cells in con-tact zone with characteristic thickenings of lenticularform. Chains temporarily associated with filaments,coiled in loop-shaped pattern, shaped like elongatedskein. Chains and filaments may occur separately fromeach other.
S p e c i e s c o m p o s i t i o n. A. ellipsoidea Her-mann, 1976; A. kelleri Hermann et Jankauskas 1989;A. tetragonalum (Maithy, 1975) Hofmann et Jackson,1994.
O c c u r r e n c e. Upper Riphean: Russia (TheSouthern Urals, northwestern Siberia), Zaire, China;Middle Riphean of Canada.
Arctacellularia ellipsoidea Hermann, 1976
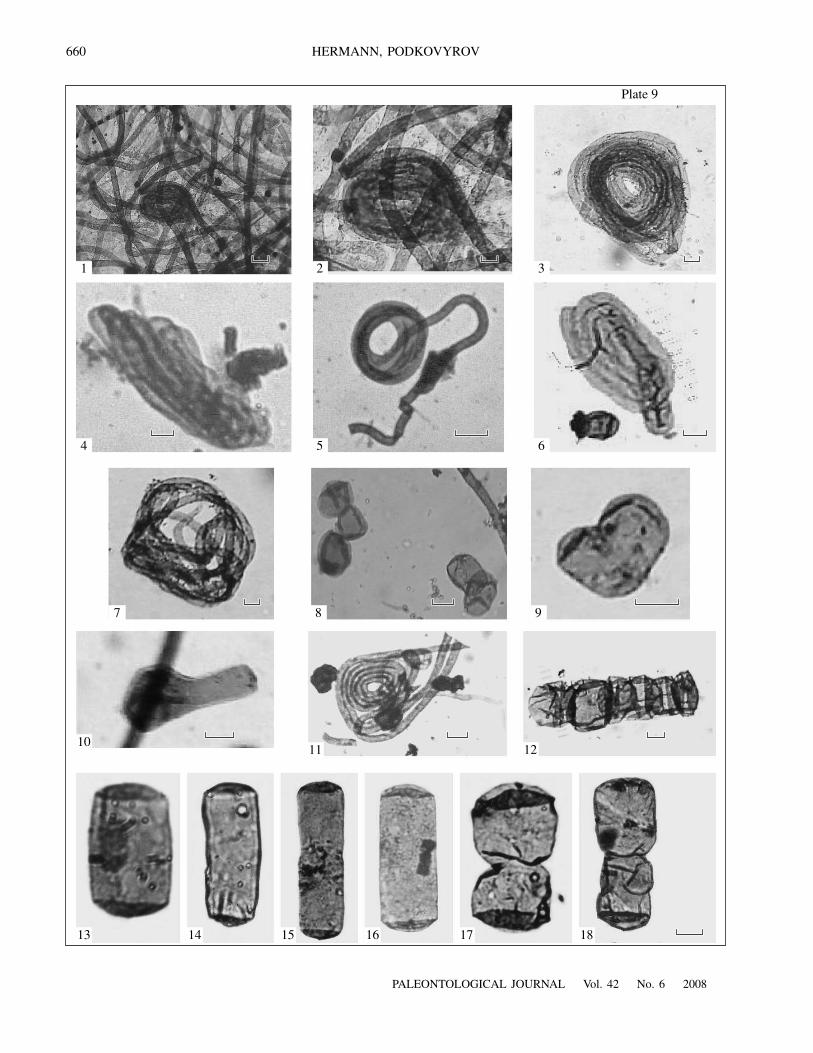
Plate 8, figs. 1–5, 7, 10, 11; Plate 9, fig. 12
Arctacellularia ellipsoidea Hermann: Timofeev et al., 1976,p. 39, pl. 11, figs. 1, 2; Jankauskas, 1982, p. 117, pl. 35, figs. 4, 5, 9,13; pl. 38, fig. 6, 11; pl. 48, figs. 9, 12; Xing et al., 1985, p. 63, pl. 7,fig. 13; Jankauskas et al. 1989, p. 136, pl. XXXVIII, figs. 1a, 1b, 6,9, 14; pl. 13, fig. 9; Hofmann and Jackson, 1994, p. 16, pl. 12,figs. 16–18.
Arctacellularia doliiformis Hermann: Timofeev et al., 1976,p. 38, pl. X, figs. 1–5.
? Glenobotridium kanshiensis Maithy: Maithy, 1975, p. 139,pl. 3, fig. 20.
H o l o t y p e. IGGD, no. 49/24-X1.72; Russia,northwestern Siberia, Turukhansk region, MiroedikhaRiver, Miroedikha Formation; Upper Riphean.
D e s c r i p t i o n. The chains are multicellular,unbranched, consisting of cells predominantly ellipsoi-dal in shape, more rarely barrel-shaped to cylindrical,devoid of an external covering, isolated or in contactwith filaments coiled into looping pattern (hyphae).The ellipsoidal cells are 20–25 µm wide, 10–15 µmhigh, the cells occasionally retain a spot at the center(condensed cytoplasm?). The barrel-shaped and cylin-drical cells vary in width only slightly (18–25 µm)whereas in length they vary from 30–35 to 60 µm. Inthe zone where the cells are in contact with each other,they have thickenings of lenticular form. Narrow cordsrun along the chains of cells. The number of cells inchains varies; in the holotype, this number is 20. Themaximum length of chains in fragments is 300 µm.
R e m a r k s. The presence of different types of theformation of chains (segmentation or fragmentation ofsporiferous hyphae) and the aberration of growthobserved in certain cells in chains suggest the polymor-phism characteristic of fungal organisms. Some of thecells in chains are quite rounded (Pl. 8, fig. 2, the low-ermost cell with a pair of cells formed above it). Thechain in Pl. 8, fig. 17 shows the same pattern: therounded cell (third from the bottom) with two pairs ofseparated cells. The process of cell division was alsoobserved in the chains associated with hyphae (see Pl. 8,figs. 5, 11). These facts may suggest the possibility offurther multiserial formation of fungal spores.
O c c u r r e n c e. The Middle Riphean (Bylot Group)of Canada; Upper Riphean: Miroedikha Formation ofnorthwestern Siberia; Zilmerdak and Uk formations ofthe Karatavian of the Southern Urals; Southern Timan,Dzhezhinskaya Parma; Bushimay Supergroup, Zaire.
M a t e r i a l. Five specimens of interconnectedmicrofossils and numerous finds of isolated chains.
Arctacellularia kelleri Hermann et Jankauskas, 1989
Plate 8, figs. 6, 8, 14, and 16; Plate 9, figs. 13–16
Arctacellularia kelleri Hermann et Jankauskas: Jankauskaset al., 1989, p. 136, pl. XXXVIII, figs. 3–5.
Arctacellularia varia Jankauskas et Hermann: Jankauskas et al.,1989, p. 136, pl. XXXVIII, figs. 6–8, 10–13.
Arctacellularia sp.: Jankauskas, 1982, p. 117. pl. XXXVII,figs. 1–3, 5–8, 14; pl. XXXVIII, figs. 1, 2.
E x p l a n a t i o n o f P l a t e 9Figs. 1–3, 5, 7, and 11. Glomovertella eniseica Hermann, 1989: concentric coiling of filaments: (1, 2) specimen no. 49/t-16.1.71;(3, 11) specimen no. 49/5-6.XII.73; (5) specimen no. 49/3-23.II.74; (7, 11) specimen no. 49/5-6.XII.73.Figs. 4 and 6. Glomovertella miroedikhia sp. nov.: (4) specimen no. 49/46-XI.72; (6) specimen no. 49/3-23.II.74.Figs. 8–10. Arctacellularia sp.: (8) specimen no. 49a/1, fragmentation of chains into single cells; (9, 10) budding of cells: (9) spec-imen no. 49/46-XI.72, (10) specimen no. 49/2-11.XII.73.Fig. 12. Arctacellularia ellipsoidea Hermann, 1976, chain with a markedly hypertrophied cell, specimen no. 49/57-XI.72.Figs. 13–16. Arctacellularia kelleri Jankauskas et Hermann, 1989: (13) specimen no. 49/7-XI.72, (14) specimen no. 49/43,(15) specimen no. 49/3-XI.72, (16) specimen no. 49/3-XI.76.Figs. 17 and 18. Arctacellularia tetragonale (Meithy, 1975): (17) specimen no. 49/60-XI.72; (18) specimen no. 49/46-XI.72.Scale bar 10 µm.

662
PALEONTOLOGICAL JOURNAL Vol. 42 No. 6 2008
HERMANN, PODKOVYROV
H o l o t y p e. IGGD, no. 49/6-76; Russia, north-western Siberia, Turukhansk Region, Miroedikha River,Miroedikha Formation, Upper Riphean.
D e s c r i p t i o n. The cells are single or arranged inpairs, smooth-walled, barrel-shaped and cylindrical.The lateral walls are straight, smooth, the front and rearwalls are slightly convex (rounded) with characteristicnarrow lens-shaped thickenings. The cells are 10–25 µm wide and 40 to 100 µm long. Occasionally thereis a thickening (distinct or indistinct) inside the cellsthat follows the contours of the covering enclosing it.
C o m p a r i s o n. The single cells of the genus Arc-tacellularia, which are cylindrical in shape and have asize comparable with the size of single cells of thegenus Archaeoellipsoides, differ from the latter in thecharacteristic lens-shaped thickenings located on the shortfront and rear walls, thus forming a slight convexity.
As previously noted, the single cells of A. kelleri areconsidered as possible mature arthrospores that belongto representatives of the quite extensive group of asco-mycetes. At the terminal stage of development theshape of arthrospores changed from ellipsoidal and bar-rel-shaped to cylindrical. The development of singlecells shows some degree of variability: they show thebeginning of centripileous division (Pl. 8, fig. 16) andbudding (Pl. 9, figs. 9, 10).
R e m a r k s. Different deviations from the regularoval or cylindrical shapes of cells are due to differentstates of preservation of microfossils.
O c c u r r e n c e. The Upper Riphean of northwest-ern Siberia (Miroedikha Formation) in the SouthernUrals (Zilmerdak and Uk formations).
M a t e r i a l. Numerous specimens in varying statesof preservation from the Miroedikha Formation ofnorthwestern Siberia.
Genus Glomovertella Reitlinger, 1948Glomovertella miroedikhia Hermann, sp. nov.
Plate 8, figs. 9, 13, 15; Plate 9, figs. 4, 6
Glomovertella firma Reitlinger: Reitlinger, 1948, p. 80. pl. 1,fig. 5; 1959, p. 23, pl. VIII, figs. 4–8.
Glomovertella eniseica (Hermann): Jankauskas et al., 1989,p. 108, pl. XXIX, figs. 5, 7, 8.
E t y m o l o g y. From the Miroedikha River.H o l o t y p e. IGGD, no. 49/35-XI.72; Russia,
northwestern Siberia, Turukhansk Region, MiroedikhaRiver, Miroedikha Formation; Upper Riphean.
D i a g n o s i s. Individual aseptate filaments coiledin loop-shaped pattern in the form of skein, beingclosely spaced and arranged parallel to each other. Fil-aments (hyphae) 8–10 µm wide and remain constant inwidth in each specimen. Ends of filaments invisible andmay be hidden inside. Clumps of hyphae range from100 to 150 µm in length and 80–100 µm in width.
D e s c r i p t i o n. The single aseptate filaments arecoiled in loop-shaped pattern in the form of an elongateskein, in which the filaments are arranged parallel to
one another and overlap one another. The filaments are8–10 µm wide, each specimen has a constant width, thetips of the filaments penetrate deep into the skein. Theskeins are 100–150 µm long and 80–100 µm wide. Thefilaments may be temporarily associated with thechains of the cells of Arctacellularia.
C o m p a r i s o n. G. miroedikhia sp. nov. differsfrom the species G. eniseica in the parallel arrangementof filaments in the skein instead of a tangle of filamentsin the latter .
R e m a r k s. In addition to the above reasons, thepredisposition of filaments to coiling is characteristic ofmicrosclerotia of fungi.
M a t e r i a l. Numerous specimens in a good state ofpreservation from the type locality.
ACKNOWLEDGMENTSThis work was supported by Program 18 of the
Presidium of the Russian Academy of Sciences and bythe Russian Foundation for Basic Research (projectno. 07-05-00906).
REFERENCES1. E. A. Aseeva, “On Spiral- and Ring-Shaped Structures in
the Upper Precambrian Deposits of Podolia,” Paleontol.Sb., No. 11, Issue 2, 95–98 (1974).
2. M. Yu. Belova and A. M. Akhmedov, “Petsamomyces, aNew Genus of Organic-Walled Microfossils from theCoal-Bearing Deposits of the Early Proterozoic, KolaPeninsula,” Paleontol. Zh., No. 5, 3–12 (2006) [Paleon-tol. J. 40 (5), 465–475 (2006)].
3. L. Benesh, “Paleomycology: A New Line of Micro-scopic Investigations of Coal,” Izv. Akad. Nauk SSSR,Ser. Geol., No. 11, 47–52 (1960).
4. Biological Cybernetics, Ed. by A. B. Kogan (VysshayaShkola, Moscow, 1977) [in Russian].
5. I. P. Borodin, Sketch of Mycology (Izd. A.N. Petrova,St. Petersburg, 1897) [in Russian].
6. M. B. Burzin, “The Earliest Chytridiomycetes (Mycota,Chytridiomycetes incertae sedis) from the Upper Ven-dian of the East European Platform,” in Fauna and Eco-systems of the Geological Past (Nauka, Moscow, 1993),pp. 21–33 [in Russian].
7. M. B. Burzin, “Late Vendian Helicoid FilamentousMicrofossils,” J. Paleontol. 29 (1A), 1–34 (1995).
8. N. J. Butterfield, “Probable Proterozoic Fungi,” Paleobi-ology 31 (1), 165–182 (2005).
9. A. A. Chiguryaeva, Atlas of Microspores from the Ter-tiary Deposits of the USSR (Khar’kovsk. Gos. Univ.,Khar’kov, 1956) [in Russian].
10. R. T. Clarke, “Fungal Spores from Vermejo FormationCoal Beds (Upper Cretaceous) of Central Colorado,”Mountain Geol. 3 (2), 85–93 (1965).
11. Course in Lower Plants, Ed. by M. V. Gorlenko(Vysshaya Shkola, Moscow, 1981) [in Russian].
12. D. L. Dilcher, “Epihyllous Fungi from Eocene Depositsin Western Tennessee, USA,” Palaeontogr. B 116 (1–4),1–54 (1965).

PALEONTOLOGICAL JOURNAL Vol. 42 No. 6 2008
ON THE NATURE OF THE PRECAMBRIAN MICROFOSSILS 663
13. W. N. Edwards, “An Eocene Microthyreaceous Fungusfrom Vull, Scotland,” Trans. Brit. Mycol. Soc. 8, 66–72(1922).
14. W. C. Elsik, “The Fungal Morphotype Felixites n. gen.,”Pollen et Spores 31 (1–2), 155–159 (1989a).
15. W. C. Elsik, “Late Neogene Palynomorph Diagrams,Northern Gulf of Mexico,” Trans. Gulf Coast Assoc.Geol. Soc. 19, 509–528 (1989b).
16. W. C. Elsik, Fungal Palynomorphs (Texas University,Houston, 1991), pp. 1–6.
17. J. Felix, “Studien über fossile Pilze,” Z. Dtsch. Geol.Ges. 46, 269–280 (1894).
18. Fossil Calcareous Algae: Proceedings of the Institute ofGeology and Geophysics of the Siberian Division of theAcademy of Sciences of the USSR, Issue 674, Ed. byV. N. Dubatolov (Nauka, Novosibirsk, 1987) [in Russian].
19. M. B. Gnilovskaya, A. F. Veis, Yu. R. Bekker, et al., “Pre-Ediacarian Fauna from Timan (Annelidomorphs of theLate Riphean),” Stratigr. Geol. Korrelyatsiya 8 (4), 11–39(2000) [Stratigr. Geol. Correlation 8 (4), 327–352 (2000)].
20. V. K. Golovenok and M. Yu. Belova, “Records ofObruchevella from the Riphean of the Patom Plateauand from the Vendian of Southern Kazakhstan,” Dokl.Akad. Nauk SSSR 272 (6), 1462–1464 (1983).
21. V. P. Grichuk, “A Method of the Preparation of Sedimen-tary Rocks Low in Organic Remains for the Purposes ofPollen Analysis,” Probl. Fiz. Geogr. 8, 5–15 (1940).
22. T. van der Hammen, “Principios para la nomenclaturapalinológica sistemática,” Bol. Geol. 2 (2), 1–21 (1955).
23. D. S. Heckman, D. M. Geiser, B. R. Eidell, et al.,“Molecular Evidence for the Early Colonization of Landby Fungi and Plants,” Science 293 (5532), 1129–1133(2001).
24. S. B. Hedges, J. E. Blair, M. L. Venturi, and J. L. Shoe,“A Molecular Timescale of Eukaryote Evolution and theRise of Complex Multicellular Life,” BMC Evol. Biol. 4(2), 1–9 (2004).
25. T. N. Hermann, “On the Experience of the Extraction ofLarge Plant Remains and Microfossils using ChemicalDissolution of Rocks,” in Microfossils of the USSR(Nauka, Novosibirsk, 1974), pp. 97–99 [in Russian].
26. T. N. Hermann, “Records of Fungi from the Riphean,” inPaleontology of the Precambrian and Early Cambrian(Nauka, Leningrad, 1979), pp. 129–136 [in Russian].
27. T. N. Hermann, “Records of Filamentous Algae in theMiroedikha Formation of the Upper Precambrian,” Pale-ontol. Zh., No. 4, 118–121 (1981).
28. T. N. Hermann, “On Records of Blue-Green Filamen-tous Algae from the Upper Precambrian (MiroedikhaFormation),” in Current Problems in Modern Paleoal-gology (Naukova Dumka, Kiev, 1986), pp. 37–40 [inRussian].
29. T. N. Hermann, The Organic World One Billion YearsAgo (Nauka, Leningrad, 1990) [in Russian].
30. T. N. Hermann and V. N. Podkovyrov, “Fungal Remainsfrom the Late Riphean,” Paleontol. Zh., No. 2, 89–95(2006) [Paleontol. J. 40 (2), 207–214 (2006)].
31. H. J. Hofmann and G. D. Jackson, “Shale-Facies Micro-fossils from the Bylot Supergroup, Baffin Island, Can-ada,” J. Paleontol. 68 (4, Part 3), 1–32 (1994).
32. T. Y. James, F. Kauff, C. L. Schoch, et al., “Reconstruc-tion of the Early Evolution of Fungi using a Six-GenePhylogeny,” Nature 443 (7113), 818–822 (2006).
33. T. V. Jankauskas, Stratotype of the Riphean (Paleontol-ogy and Paleomagnetism) (Nauka, Moscow, 1982) [inRussian].
34. T. V. Jankauskas, N. S. Mikhailova, T. N. Hermann,et al., Microfossils of the Precambrian of the USSR(Nauka, Leningrad, 1989) [in Russian].
35. A. H. Knoll, A. J. Kaufman, and M. A. Semikhatov, “TheCarbon-Isotopic Composition of Proterozoic Carbon-ates: Riphean Successions from Northwestern Siberia(Anabar Massif, Turukhansk Uplift),” Amer. J. Sci. 295(7), 823–850 (1995).
36. P. N. Kolosov, Late Precambrian Microorganisms of theEast Siberian Platform (Yakut. Fil. Sib. Otd. Akad. NaukSSSR, Yakutsk, 1984) [in Russian].
37. N. V. Kondrat’eva, Morphology of Prokaryotic Algae(Naukova Dumka, Kiev, 1989) [in Russian].
38. P. K. Maithy, “Microorganisms from the Bushimay Sys-tem (Late Precambrian) of Kanshi, Zaire,” Palaeobot. 22,133–149 (1975).
39. W. C. Martin, M. Rotte, U. Hoffmeister, et al., “EarlyCell Evolution, Eukaryotes, Anoxia, Sulfide, Oxygen,Fungi First (?), and a Tree of Genomes Revised,”IUBMB Life 55, 193–204 (2003).
40. Microfossils of the Precambrian of the USSR, Ed. byB. S. Sokolov (Nauka, Leningrad, 1989) [in Russian].
41. P. Yu. Petrov and M. A. Semikhatov, “Structure andEnvironmental Conditions of a Transgressive UpperRiphean Complex: Miroedikha Formation of theTurukhansk Uplift, Siberia,” Litol. Polezn. Iskop., No. 1,14–35 (1997) [Lithol. Miner. Resour., No. 1, 11–29(1997)].
42. P. A. Popov, “Fossil Fungi in the Tertiary Deposits of theYenisey Ridge,” Dokl. Akad. Nauk SSSR 128 (4), 827–829 (1959).
43. P. A. Popov, “Fossil Fungi of the West Siberian Plain andYenisey Ridge,” Botan. Zh. 47, 1596–1610 (1962).
44. P. A. Popov, “Microscopic Fungi As a Subject of Palyno-logical Investigations,” Mikol. Fitopatol. 1, 158–163(1967).
45. S. M. Porter, R. Meisterfield, and A. H. Knoll, “Vase-Shaped Microfossils from the Neoproterozoic ChuarGroup, Grand Canyon: A Classification Guided byModem testae amoebae,” J. Paleontol. 77 (3), 409–429(2003).
46. E. A. Reitlinger, “Cambrian Foraminifers of Yakutia,”Byull. Mosk. O–va Ispyt. Prir., Otd. Biol., No. 2, 23(1948).
47. E. A. Reitlinger, Atlas of Microscopic Remains andProblematical Forms from the Ancient Strata of Siberia(Akad. Nauk SSSR, Moscow, 1959) [in Russian].
48. J. W. Schopf, “Microflora of the Bitter Springs Forma-tion, Late Precambrian, Central Australia,” J. Paleontol.42, 651–688 (1968).
49. M. A. Semikhatov, “Methodological Principles of Riph-ean Stratigraphy,” Stratigr. Geol. Korrelyatsiya 3 (6),33–50 (1995).
50. M. A. Semikhatov, G. V. Ovchinnikova, I. M. Gorokhov,et al., “Isotope Age of the Middle–Upper Riphean

664
PALEONTOLOGICAL JOURNAL Vol. 42 No. 6 2008
HERMANN, PODKOVYROV
Boundary: Pb–Pb Geochronology of the LakhandaGroup Carbonates, Eastern Siberia,” Dokl. Ross. Akad.Nauk 372 (2), 216–221 (2000) [Dokl. Earth Sci. 372 (4),625–629 (2000)].
51. T. N. Taylor, S. D. Klavins, M. Krigs, et al., “Fungi fromthe Rhynie Chert: A View from the Dark Side,” Trans.Roy. Soc. Edinburg. Earth Sci. 94 (Part 4), 457–473(2004).
52. B. H. Tiffney and E. S. Baghoorn, “The Fossil Record ofthe Fungi,” Occ. Pap. Farlow Herbarium, No. 7, 1–42(1974).
53. B. V. Timofeev, A Micropaleophytologic Investigation ofAncient Formations (Nauka, Leningrad, 1966) [in Rus-sian].
54. B. V. Timofeev, Spheromorphids of the Proterozoic(Nauka, Leningrad, 1969) [in Russian].
55. B. V. Timofeev, “Une découverte de Phycomycetes dansle Précambrien,” Rev. Palaeobot. 10, 79–81 (1970).
56. B. V. Timofeev, Microphytofossils of the Precambrian ofUkraine (Nauka, Leningrad, 1973) [in Russian].
57. B. V. Timofeev and T. N. Hermann, “PrecambrianMicrobiota of the Lakhanda Formation,” in Paleontol-ogy of the Precambrian and Early Cambrian (Nauka,Leningrad, 1979), pp. 137–147 [in Russian].
58. B. V. Timofeev, T. N. Hermann, and N. S. Mikhailova,Microphytofossils of the Precambrian, Cambrian, andOrdovician (Nauka, Leningrad, 1976) [in Russian].
59. C. R. Varma and M. S. Rawat, “A Note on Some Dipo-rate Grains Recovered from Tertiary Horizons of India
and Their Potential Marker Value,” Grana Palinol. 4,130–139 (1963).
60. A. F. Veis, “Microfossils of the Riphean and Vendian ofthe Uchur–Maya and Turukhan Regions of Siberia,” Izv.Akad. Nauk SSSR, Ser. Geol., No. 5, 47–64 (1988).
61. A. F. Veis and P. Yu. Petrov, “Major Features of theFacial-Ecological Distribution of Microfossils in theRiphean Basins of Siberia,” Stratigr. Geol. Korrelyatsiya2 (5), 44–64 (1994).
62. A. F. Veis and N. G. Vorob’eva, “New Data on the Com-position and Stratigraphic Significance of the Miro-edikha Microbiota from the Turukhansk Area, Siberia,”Dokl. Ross. Akad. Nauk 358 (6), 794–798 (1998) [Dokl.Earth Sci. 358 (2), 174–178 (1998)].
63. A. F. Veis, P. Yu. Petrov, and N. G. Vorob’eva, “The LateRiphean Miroedikha Microbiota from Siberia. Part 1:Composition and Facial-Ecological Distribution ofOrganic-Walled Microfossils,” Stratigr. Geol. Korre-lyatsiya 6 (5), 15–38 (1998) [Stratigr. Geol. Correlation6 (5), 440–461 (1998)].
64. A. F. Veis, P. Yu. Petrov, and N. G. Vorob’eva, “The LateRiphean Miroedikha Microbiota from Siberia. Commu-nication 2: Interpretation in Terms of Biotic Paleosuc-cession,” Stratigr. Geol. Korrelyatsiya 7 (1), 18–40(1999) [Stratigr. Geol. Correlation 7 (1), 15–34 (1999)].
65. L. Yin, “Acanthomorphic Acritarchs from Meso-Neoprot-erozoic Shales of the Ruyang Group, Shanxi, China,”Rev. Paleobot. Palynol. 98, 15–25 (1997).
66. L. Yin, X. Yuan, F. Meng, and J. Hu, “Protists of theUpper Mesoproterozoic Ruyang Group in Shanxi Prov-ince, China,” Precambr. Res. 141, 49–66 (2005).





























