Embed Size (px)
Citation preview

Neural Circuitry of Spatial Orientation and Episodic Memory
Processing: Investigation in Neurodegenerative Diseases
Sicong Tu
Prince of Wales Clinical School
Faculty of Medicine
University of New South Wales
Dissertation submitted for the degree of Doctor of Philosophy
June, 2016.

PLEASE TYPE THE UNIVERSITY OF NEW SOUTH WALES
Thesis/Dissertation Sheet
Surname or Family name: Tu
First name: Slcong
Abbreviation for degree as given in the University calendar: PhD
School: Prince of Wales Clinical School
Title: Neural Circuitry of Spatial Orientation and Episodic Memory Processing: Investigation in Neurodegenerative Diseases
Other name/s:
Faculty: Medicine
Abstract 350 words maximum: (PLEASE TYPE)
Research into the neuroanatomical bases of memory has, for a long time, focused on medial temporal lobe brain structures, namely the
hippocampus, which plays a key role in episodic memory (memory of specific events) and spatial memory processes. The work described in this
thesis investigates the behavioural impact to episodic and spatial memory, resulting from neural changes beyond the hippocampus, in
neurodegenerative conditions, such as Alzheimer's disease (AD), frontotemporal dementia (FTD), and thalamic stroke. Novel cognitive tasks were
employed in combination with advanced neuroimaging to examine dissociable episodic and spatial memory performance in these patient
populations as a result of contrasting atrophy in a neuroanatomical circuit of memory, the circuit of Papez. Focus was placed on identifying specific
memory functions associated with atrophy in the Papez memory circuit that could be targeted to improve differential clinical diagnosis of AD and
FTD.
Two clinically feasible behavioural tasks were developed to objectively assess spatial orientation and long-term contextual memory, respectively.
Findings Indicated spatial orientation is a key discriminating feature for AD and FTD patients, associated with integrity of the retrosplenial cortex, a
region affected by early AD pathology, but not FTD. The long-term contextual memory task demonstrated focal damage to the thalamus, a region
commonly affected in AD and FTD, can result in accelerated forgetting of newly learnt material over a 4-week period. Longitudinal neuroimaging
identified divergent patterns of atrophy affecting brain structures in the Papez memory circuit in AD and FTD, specifically Involving the posterior
cingulate gyrus and anterior thalamus, consistent with observed pattern of behavioural performance on experimental tasks. Furthermore,
additional longitudinal neuroimaging examining whole-brain white matter degeneration highlighted the potential application of diffusion weighted
magnetic resonance imaging in detecting underlying disease pathology, in-vivo, in FTD.
The findings from this thesis demonstrate unique changes are present in Papez memory structures beyond the hippocampus across dementia
syndromes, which have a significant impact on episodic and spatial memory deficits, and can be used to develop sensitive clinical measures to
improve differential diagnosis.
Declaration relating to disposition of project thesis/dissertation
I hereby grant to the University of New South Wales or its agents the right to archive and to make available my thesis or dissertation in whole or in part in the University libraries in all forms of media, now or here after known, subjectto the provisions of the Copyright Act 1968. I retain all property rights, such as patent rights. I also retain the right to use in future works (such as articles or books) all or part of this thesis or dissertation.
I also authorise University Microfilms to U$e the 350 word abstract of my thesis in Dissertation Abstracts International (this is applicable to doctoral theses only).
The University recognises that there may be exceptional circumstances requiring restrictions on copying or conditions on use. Requests for restriction for a period of up to 2 years must be made in writing. Requests for a longer period of restriction may be considered in exceptional circumstances and require the approval of the Dean of Graduate Research.
FOR OFFICE USE ONLY Date of completion of requirements for Award:
THIS SHEET IS TO BE GLUED TO THE INSIDE FRONT COVER OF THE THESIS

iv
Originality Statement
‘I hereby declare that this submission is my own work and to the best of my knowledge it
contains no materials previously published or written by another person, or substantial
proportions of materials which have been accepted for the award of any other degree or
diploma at UNSW or any other educational institution, except where due acknowledgement
is made in the thesis. Any contribution made to the research by others, with whom I have
worked at UNSW or elsewhere, is explicitly acknowledged in the thesis. I also declare that
the intellectual content of this thesis is the product of my own work, except to the extent that
assistance from others in the project’s design and conception or in style, presentation and
linguistic expression is acknowledged.’
Signed:
Sicong Tu
Date: 30 May 2016

COPYRIGHT STATEMENT
‘I hereby grant the University of New South Wales or its agents the right to archive and to make available my thesis or dissertation in whole or part in the University libraries in all forms of media, now or here after known, subject to the provisions of the Copyright Act 1968. I retain all proprietary rights, such as patent rights. I also retain the right to use in future works (such as articles or books) all or part of this thesis or dissertation. I also authorise University Microfilms to use the 350 word abstract of my thesis in Dissertation Abstract International (this is applicable to doctoral theses only). I have either used no substantial portions of copyright material in my thesis or I have obtained permission to use copyright material; where permission has not been granted I have applied/will apply for a partial restriction of the digital copy of my thesis or dissertation.'
Signed ……………………………………………...........................
Date ……………………………………………...........................
AUTHENTICITY STATEMENT
‘I certify that the Library deposit digital copy is a direct equivalent of the final officially approved version of my thesis. No emendation of content has occurred and if there are any minor variations in formatting, they are the result of the conversion to digital format.’
Signed ……………………………………………...........................
Date ……………………………………………...........................

i
Table of Contents
Page No.
Table of Contents i
Acknowledgements iii
Originality Statement iv
Supervisor Statement v
List of First Author Publications Arising from PhD vi
List of Conference Proceedings Arising from PhD vii
List of Tables ix
List of Figures xi
Abbreviations xiii
Chapter 1. Introduction 1
The Papez Circuit and its Role in Memory 3
Clinical Features of Alzheimer’s Disease and Frontotemporal Dementia 7
Episodic Memory in Alzheimer’s Disease and Frontotemporal Dementia 11
Spatial Memory in Alzheimer’s Disease and Frontotemporal Dementia 17
Neuroimaging Principles 20
Structural Brain Imaging in FSL 23
Diffusion Brain Imaging in FSL 24
Aims and Hypotheses 27

ii
Chapter 2. Spatial Orientation in Dementia 29
Publication I – “Lost in spatial translation – A novel tool to objectively assess spatial
disorientation in Alzheimer’s disease and frontotemporal dementia 30
Publication II – “Egocentric vs. allocentric spatial memory in behavioural variant
frontotemporal dementia and Alzheimer’s disease 42
Chapter 3. Long-term Contextual Memory in Thalamic Stroke 73
Publication III – “Accelerated forgetting of contextual details due to focal medio-dorsal
thalamic lesion” 74
Chapter 4. Longitudinal Papez Memory Circuit Integrity in Dementia 83
Publication IV – “Longitudinal Papez circuit integrity in Alzheimer’s disease and
frontotemporal dementia” 84
Chapter 5. Longitudinal White Matter Degradation in Primary Progressive
Aphasia 114
Publication V – “Divergent longitudinal propagation of white matter degradation in
logopenic and semantic variants of primary progressive aphasia 115
Chapter 6. Conclusion 124
Reference List for Introduction and Conclusion Chapters 129
Appendices 152
Supplemental Material for Publications in Chapters 152
Declaration Supporting Inclusion of Publications in the Thesis 166

iii
Acknowledgements
First and foremost, I am deeply grateful to my family and friends who have supported me
throughout my studies. Whether it was patiently listening to the achievements and
frustrations of my research or offering encouragement and advice, you have all been a great
source of comfort and support to me. To my supervisor Michael Hornberger, thank you so
much for sharing your passion for research and passing down your knowledge and expertise.
I will always cherish our first meeting years ago in a quiet corner of the kitchen at NeuRA
when you first introduced me to the exciting field of memory research in dementia which
made all of this possible. I am also extremely grateful to my supervisor Olivier Piguet who
has always kept his door open for when I needed advice and made sure I felt included in the
FRONTIER research group from my first day. This PhD has been a long and challenging
journey, but I couldn’t have asked for better supervisors to guide me through it.
I would like to thank everyone in the FRONTIER group. It has been a pleasure
working together with such a friendly, open, and supportive group of researchers. The project
was much more enjoyable and interesting because of you all. I will always hold fond
memories of my time at FRONTIER and hope the team continues to grow and prosper. I am
also deeply grateful to the volunteers from the community as well as the patients and their
carers who made the time and effort in their everyday lives to contribute to my research.
My gratitude goes out to the Alzheimer’s Australia Dementia Research Foundation
and Australian National Health and Medical Research Council for their financial support
throughout my PhD candidature. Finally, I would like to thank the University of New South
Wales and the Prince of Wales Clinical School for providing the financial support to present
the findings of my research at international conferences, providing the opportunity to
appreciate the breadth of research being undertaken in my field globally.

v
Supervisor Statement
I hereby certify that all co-authors of the published or submitted papers agree to Sicong Tu
submitting those papers as part of his Doctoral Thesis.
Signed:
Michael Hornberger
Date: 31 May 2016
Olivier Piguet
Date: 30 May 2016

vi
List of First Author Publications Arising from PhD
Tu, S., Leyton, C. E., Hodges, J. R., Piguet, O., & Hornberger, M. (2015). Divergent
longitudinal propagation of white matter degradation in logopenic and semantic variants of
primary progressive aphasia. Journal of Alzheimer's Disease : JAD, 49(3), 853-861.
doi:10.3233/JAD-150626 [doi]
Tu, S., Miller, L., Piguet, O., & Hornberger, M. (2014). Accelerated forgetting of contextual
details due to focal medio-dorsal thalamic lesion. Frontiers in Behavioral Neuroscience, 8
Tu, S., Piguet, O., Hodges, J. R., Hornberger, M. Longitudinal Papez circuit integrity in
Alzheimer’s disease and frontotemporal dementia. (in submission)
Tu, S., Spiers, H. J., Hodges, J. R., Piguet, O., Hornberger, M. Egocentric vs. allocentric
spatial memory in behavioural variant frontotemporal dementia and Alzheimer’s disease.
(in submission)
Tu, S., Wong, S., Hodges, J. R., Irish, M., Piguet, O., & Hornberger, M. (2015). Lost in
spatial translation - A novel tool to objectively assess spatial disorientation in alzheimer's
disease and frontotemporal dementia. Cortex; a Journal Devoted to the Study of the Nervous
System and Behavior, 67, 83-94. doi:10.1016/j.cortex.2015.03.016 [doi]

vii
List of Conference Proceedings Arising from PhD
Tu, S., Miller, L., Piguet, O., Hornberger, M. Long-term anterograde memory in thalamic
stroke patients. Prince of Wales Clinical School Research Symposium, 11th October, 2013,
Sydney, Australia.
Tu, S., Miller, L., Hornberger, M. Anterior thalamus contributions to long-term consolidation
of contextual memory. 43rd Annual Meeting of the Society for Neuroscience, 9-13th
November, 2013, San Diego, USA.
Tu, S., Miller, L., Piguet, O., Hornberger, M. Impact of thalamic lesions on long-term
memory consolidation. 41st Annual Coast Association TOW Research Awards, 22nd
November, 2013, Sydney, Australia.
[ORAL] Tu, S. Thalamic contributions to remote memory in healthy participants and stroke
patients. 11th Annual World Congress of the Society for Brain Mapping and Therapeutics,
17-19th March, 2014, Sydney, Australia.
[ORAL] Tu, S. Impact of thalamic lesions on long-term memory. 4th Forefront Scientific
Meeting, 26th May, 2014, Sydney, Australia.
[ORAL] Tu, S. Accelerated forgetting of contextual details due to focal medio-dorsal
thalamic lesion. Australian Research Council Centre of Excellence in Cognition and its
Disorders Annual Workshop, 21st August, 2014, Sydney, Australia.
[ORAL] Tu, S. Spatial orientation discriminates frontotemporal dementia from Alzheimer’s
disease. Brain Sciences Symposium, 17th October, 2014, Sydney, Australia.

viii
Tu, S., Wong, S., Piguet, O., Hodges, J., Hornberger, M. Spatial orientation performance
differs across dementia subtypes. Inter-university Neuroscience and Mental Health
Conference, 29-30th September, 2014, Sydney, Australia.
Tu, S., Wong, S., Piguet, O., Hodges, J., Hornberger, M. Spatial orientation discriminates
behavioural variant FTD from Alzheimer’s disease. 9th International Conference on
Frontotemporal Dementias. 23-25th October, 2014, Vancouver, Canada.
[ORAL] Tu, S. Orientation dysfunction in frontotemporal dementia and Alzheimer’s disease.
International Neuropsychological Society Mid-Year Meeting, 3rd July, 2015, Sydney,
Australia.
Tu, S., Piguet, O., Hornberger, M. Longitudinal memory circuit integrity in AD and FTD. 5th
European Society for Neuroscience Conference, 9-11th September, 2015, Tampere, Finland.
[ORAL] Tu, S. Orientation discriminates frontotemporal dementia and Alzheimer’s disease.
Prince of Wales Clinical School Research Seminar, 16th October, 2015, Sydney, Australia.
[ORAL] Tu, S. Thalamic contributions to long-term memory retrieval. International
Conference On Memory, 17-22nd July, 2016, Budapest, Hungary.
[ORAL] Tu, S. Spatial orientation in Alzheimer’s disease and frontotemporal dementia.
Australian Research Council Centre of Excellence in Cognition and its Disorders Annual
Workshop, 24th August, 2016, Sydney, Australia.

ix
List of Tables
Page No.
Publication I
Table 1 Participant demographic characteristics and performance on
standardized neuropsychological assessments 32
Table 2 Voxel-based morphometry differences in patient groups 36
Publication II
Table 1 Participant demographic characteristics and performance on
standardized neuropsychological assessments 53
Publication III
Table 1 Participant demographic characteristics, lesion localization and
performance on standardized neuropsychological assessments 75
Table 2 Mean FA, mean diffusivity and tract volume of the mammillothalamic
tract in thalamic patients and controls 79
Publication IV
Table 1 Participant demographic characteristics and performance on
standardized neuropsychological assessments 89

x
Publication V
Table 1 Participant demographic characteristics, global cognition and single-
word task performance 117
Table 2 Mean fractional anisotropy values of white matter tracts 121

xi
List of Figures
Page No.
Chapter 1
Figure 1.1 Core components of the Papez circuit studied in-vivo 4
Figure 1.2 Clinical dichotomy of Alzheimer’s disease and frontotemporal
dementia syndromes 8
Figure 1.3 Neuropathology in Alzheimer’s disease and frontotemporal
dementia syndromes 11
Figure 1.4 Overview of the principal components of imaging analyses in FSL 23
Figure 1.5 Overview of the longitudinal DTI processing pipeline 26
Publication I
Figure 1 Example screenshots of the virtual supermarket task 33
Figure 2 Participant spatial orientation performance 35
Figure 3 ROC curve for memory and orientation performance in diagnosing
dementia patients 35
Figure 4 Voxel-based morphometry analysis of structural grey matter in patients 37
Publication II
Figure 1 Example of egocentric and allocentric components of the virtual
supermarket task 49
Figure 2 Egocentric and allocentric heading direction performance 55

xii
Figure 3 Participant performance on the spatial layout component of the
virtual supermarket task 57
Figure 4 Mean Euclidean distance of participant’s spatial representations from the
centre of the map 58
Figure 5 Mean left and right hippocampal volume in participant groups 60
Publication III
Figure 1 Long-term recognition and recall memory performance in thalamic
patients and healthy controls 78
Figure 2 Axial slices of thalamic patient lesions 79
Publication IV
Figure 1 Longitudinal change in participant performance across standard
cognitive measures 97
Figure 2 Longitudinal grey matter integrity in Papez structures 99
Figure 3 Longitudinal white matter integrity in Papez structures 100
Publication V
Figure 1 Cross-sectional white matter tract degeneration in patients and controls 119
Figure 2 Longitudinal white matter tract degeneration in patients and controls 120

xiii
Abbreviations
Aβ Beta-amyloid
ACE-R Addenbrooke’s Cognitive Examination Revised
AD Alzheimer's Disease
ALF Accelerated Long-term Forgetting
ANOVA Analysis of Variance
AT Anterior Thalamic Nuclei
BET Brain Extraction Tool
BvFTD Behavioural Variant Frontotemporal Dementia
CDR Clinical Dementia Rating
CSF Cerebrospinal Fluid
D&PT Doors and People Test
DLPFC Dorsal Lateral Prefrontal Cortex
DTI Diffusion Tensor Image
DWI Diffusion Weighted Imaging
FA Fractional Anisotropy
FAST FMRIB Automatic Segmentation Tool
FLIRT FMRIB Linear Image Registration Tool
FSL FMRIB Software Library
FTD Frontotemporal Dementia
FUS RNA-binding Protein Fused in Sarcoma
FWE Family-wise Error

xiv
Hb Hemoglobin
LvPPA Logopenic Variant Primary Progressive Aphasia
MD Medio-dorsal Thalamic Nuclei
MNI Montreal Neurological Institute
MRI Magnetic Resonance Imaging
MTT Mammillothalamic Tract
NavPPA Nonfluent/Agrammatic Primary Progressive Aphasia
PPA Primary Progressive Aphasia
RAVLT Rey Auditory Verbal Learning Test
RCFT Rey Complex Figure Test
RF Radio Frequency
ROC Receiver Operating Characteristic
ROI Regions of Interest
SD Semantic Dementia
SvPPA Semantic Variant Primary Progressive Aphasia
TBSS Tract-based Spatial Statistics
TDP-43 TAR-DNA-binding Protein 43
TE Echo Time
TFCE Threshold-free Cluster Enhancement
TR Repetition Time
VBM Voxel-based Morphometry

1
Chapter 1 – Introduction
In essence, memory is stored information gained from past experiences in our lives that can be
retained and retrieved. This knowledge shapes our everyday behaviour and contributes to how
we respond to everyday events and social interaction. Our current understanding of the neural
organization of memory has been progressively refined over centuries of research from
conceptual musings to modern day neuroimaging. Despite thousands of studies, however, the
neural substrates underlying memory processing are still not fully understood. A key component
in understanding how memory works is by examining what happens when it fails.
Amnesic patients have provided the most compelling evidence for specific neural
structures underlying memory processes, the most notable being patient HM (Scoville & Milner,
1957). Patient HM underwent bilateral resection of the medial temporal lobe, which structural
MRI has confirmed affected the entorhinal cortex and hippocampal complex (dentate gyrus,
hippocampus, subiculum) (Corkin, Amaral, Gonzalez, Johnson, & Hyman, 1997), to alleviate
untreatable epileptic seizures. An unforeseen consequence of this surgery was development of
severe anterograde amnesia, namely a loss of ability to create new memories and a partial or
complete inability to recall previously experienced events. It is now well-established that the
hippocampus, a subcortical brain structure located in the medial temporal lobe, plays a critical
role in memory processes, and damage to this structure is a strong predictor of patients
developing amnesic symptoms (Squire & Alvarez, 1995; Winocur & Moscovitch, 2011).
Damage to the hippocampus, however, does not account for all reported cases of amnesia in
patients. In particular, damage to extra-hippocampal grey and white matter subcortical structures

2
including the fornix, mammillary bodies, mammillothalamic tract, thalamus, and posterior
cingulate, also result in varying degrees of amnesia in patients (Carlesimo, Lombardi, &
Caltagirone, 2011; Gaffan & Gaffan, 1991; Jung, Chanraud, & Sullivan, 2012; Valenstein et al.,
1987). These structures are either directly or indirectly connected to the hippocampus, and
together have been proposed to constitute a neural memory circuit otherwise known as the circuit
of Papez (Papez, 1995). This is of particular interest in neurodegenerative brain conditions, such
as Alzheimer’s disease (AD) and frontotemporal dementia (FTD), whereby patients demonstrate
a range of memory related impairments and pathological change involving multiple Papez circuit
structures, beyond the hippocampus (Hornberger et al., 2012; Tan et al., 2014).
This thesis explores unique patterns of cognitive change in memory related processes,
driven by divergent disease pathology beyond the hippocampus in two neurodegenerative
conditions, AD and FTD. Focus is placed on identifying changes to key Papez brain structures
underlying memory processing to target cognitive functions most sensitive in clinical diagnosis
of these patient populations. To investigate these brain-behaviour relationships, this thesis
reports novel experiments as well as established memory measures, in combination with multi-
modal grey and white matter magnetic resonance imaging (MRI) to assess regional change in-
vivo at clinic presentation and over time.
In this introduction, an overview is provided of brain structures constituting the circuit of
Papez (Papez, 1995) and their increasingly recognised relationship to memory function. The
introduction will then outline clinical features of episodic and spatial memory in AD and FTD
conditions, and their underlying neural correlates. This is followed by an overview of structural
and diffusion neuroimaging. The experimental chapters then discuss improving the diagnostic
accuracy of discriminating between AD and FTD through the use of a novel assessment of

3
spatial memory, proof of concept of longitudinal anterograde memory testing, and characterizing
longitudinal memory circuitry change in AD and FTD. Papers included in these chapters either
have been published in peer-reviewed journals or are in submission. Together, the studies offer
novel insights on how changes in neural substrates of memory beyond the hippocampus in AD
and FTD contribute to cognitive symptoms and inform the development of better diagnostic
protocols. Broader conclusions and implications are then discussed in the conclusions chapter.
The Papez Circuit and its Role in Memory
The Papez circuit was initially introduced into the literature in 1937 as an anatomical basis for
emotion perception and expression (Papez, 1995). Since then, the neural components comprising
the circuit have been widely associated with memory function, in particular, episodic (i.e.,
specific events or episodes linked to a specific temporal-spatial context) and spatial memory
across lesion and functional neuroimaging studies (Carlesimo et al., 2007; Carlesimo et al., 2011;
Maddock, Garrett, & Buonocore, 2001; Rudebeck et al., 2009; Tsivilis et al., 2008). The Papez
circuit (Papez, 1995) comprises a series of limbic brain structures, and their white matter
pathways, critical for memory processing. The anatomy of the circuit has previously been
delineated using fiber dissection at post-mortem and involves, in order, the hippocampus, fornix,
mammillary body, mammillothalamic tract, anterior nucleus of the thalamus, anterior thalamic
radiation, cingulate gyrus, parahippocampal gyrus, and entorhinal cortex, returning to the
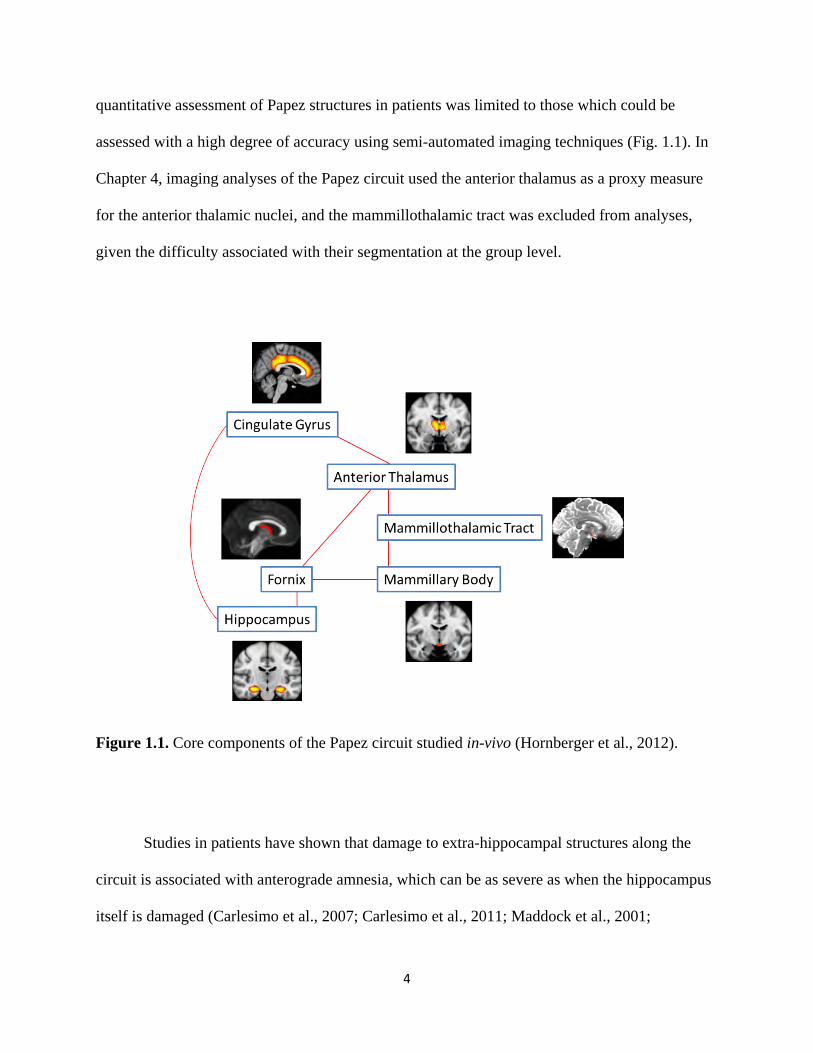
hippocampus to complete the circuit (Shah, Jhawar, & Goel, 2012) (Fig 1.1). Importantly, the
integrity of each component of the circuit can be assessed in-vivo using volumetric or diffusion
MRI techniques (Granziera et al., 2011; Hornberger et al., 2012). In the experimental chapters,
4
quantitative assessment of Papez structures in patients was limited to those which could be
assessed with a high degree of accuracy using semi-automated imaging techniques (Fig. 1.1). In
Chapter 4, imaging analyses of the Papez circuit used the anterior thalamus as a proxy measure
for the anterior thalamic nuclei, and the mammillothalamic tract was excluded from analyses,
given the difficulty associated with their segmentation at the group level.
Figure 1.1. Core components of the Papez circuit studied in-vivo (Hornberger et al., 2012).
Studies in patients have shown that damage to extra-hippocampal structures along the
circuit is associated with anterograde amnesia, which can be as severe as when the hippocampus
itself is damaged (Carlesimo et al., 2007; Carlesimo et al., 2011; Maddock et al., 2001;

5
Rudebeck et al., 2009; Tsivilis et al., 2008). Specifically, damage to the circuit appears to impair
recollection of episodic memory (i.e., retrieving or ‘remembering’ specific details of an event)
rather than familiarity (i.e., feeling or ‘knowing’ that an event was previously experienced).
Within the literature, extra-hippocampal associated amnesia resulting from focal lesions
affecting the anterior thalamic or medio-dorsal nuclei of the thalamus, following stroke, is the
most consistently reported among Papez structures (Carlesimo et al., 2011). The severity of
amnesic symptoms in thalamic stroke patients, however, varies greatly according to lesion size,
location, and whether white matter pathways (i.e., mammillothalamic tract) are affected
(Carlesimo et al., 2007; Cipolotti et al., 2008; Edelstyn, Hunter, & Ellis, 2006; Hampstead &
Koffler, 2009; Kishiyama et al., 2005; Van der Werf et al., 2003).
The hippocampus and posterior components of the circuit (posterior cingulate gyrus,
parahippocampal gyrus) have also been critically tied to spatial navigation ability in healthy
individuals (Baumann & Mattingley, 2010; Maguire et al., 2000; Maguire, Nannery, & Spiers,
2006; Marchette, Vass, Ryan, & Epstein, 2014; Moffat, 2009), and in patients with focal lesion
resulting from stroke (Ino et al., 2007; Serino, Cipresso, Morganti, & Riva, 2014; Takahashi,
Kawamura, Shiota, Kasahata, & Hirayama, 1997). A recent study in a small cohort of epileptic
patients has even demonstrated that direct stimulation of the fornix and head/body of the
hippocampus, via surgically implanted electrodes, improves visual-spatial memory performance
(Miller et al., 2015). There appears, however, to be dissociation in the role of Papez structures, in
particular between hippocampal and posterior cingulate, during spatial memory processing.
Spatial coding of direction is relative to the location of landmarks within the environment and
can be determined using two conceptually different frameworks: egocentric (i.e., location of
objects relative to yourself) or allocentric (i.e., location of objects relative to other objects).

6
While both processes require a working internal representation of the environment, egocentric
processes draws upon body-oriented information including visual and body movement dependent
on medial parietal lobe regions (Land, 2014; Vann, Aggleton, & Maguire, 2009; Zaehle et al.,
2007). In contrast, allocentric processes requires the formation of an internal map of the
environment, which has been tied to hippocampal function (O'Keefe & Nadel, 1978).
Nevertheless, egocentric and allocentric processes are not mutually exclusive. Rather, they work
in conjunction to support everyday spatial navigation. A meta-analysis of functional
neuroimaging studies suggests they share overlapping brain networks extending across occipital,
parietal, frontal, and temporal lobes (Boccia, Nemmi, & Guariglia, 2014). Critically,
involvement of medial parietal lobe regions and hippocampus differs in egocentric and
allocentric processing, respectively (Baumann & Mattingley, 2010; Ino et al., 2007; Takahashi et
al., 1997; Vann et al., 2009; Zaehle et al., 2007). Focal lesions affecting medial parietal lobe
structures, such as the retrosplenial region of the posterior cingulate, have also been shown to
significantly impair egocentric rather than allocentric spatial memory processing (Ino et al.,
2007; Takahashi et al., 1997).
A prevailing view in the literature is that the hippocampus is the core structure
underpinning memory impairments in patients. The aforementioned studies, however, highlight
that episodic and spatial memory impairment resulting from damage to extra-hippocampal Papez
structures is a common occurrence. The question that arises from these observations is, are the
impairments due to hippocampal disconnection, such that additional Papez structures act as
efferent/afferent relays of hippocampal information to higher-order cortical brain regions, or do
extra-hippocampal Papez structures facilitate memory processing independent of the
hippocampus? In particular, does damage to different regions of the Papez circuit translate into

7
dissociable patterns of cognitive impairment and can this be detected at the behavioural level?
This question has important clinical implications in the context of diagnosis and disease
progression in neurodegenerative diseases, such as AD and FTD, where recent in-vivo and post-
mortem studies highlight extensive, but divergent, patterns of change along the Papez circuit
(Hornberger et al., 2012; Tan et al., 2014).
Clinical Features of Alzheimer’s Disease and Frontotemporal Dementia
Alzheimer’s disease (AD) is the most common form of younger onset dementia (< 65 years of
age), followed by frontotemporal dementia (FTD). FTD consists of 3 distinct clinical variants
each characterized by distinct profiles of cognitive impairment and atrophy to brain regions
(Gorno-Tempini et al., 2011; Rascovsky et al., 2011). Within the FTD spectrum, there is a
behavioural variant (bvFTD), and 2 language related variants that have recently been collectively
categorized as primary progressive aphasias (PPA) (Gorno-Tempini et al., 2011): semantic
(svPPA) and nonfluent/agrammatic (navPPA). In addition, PPA comprises a third syndrome
labelled as logopenic variant (lvPPA) (Fig. 1.2). This syndrome, however, is most consistently
associated with Alzheimer’s pathology rather than frontotemporal lobar degeneration pathology.
Within the literature the semantic variant (svPPA) of PPA has been and, in some cases, continues
to be reported using its original nomenclature of semantic dementia (SD). Both terms of
classification have been used across different results chapters in this thesis.
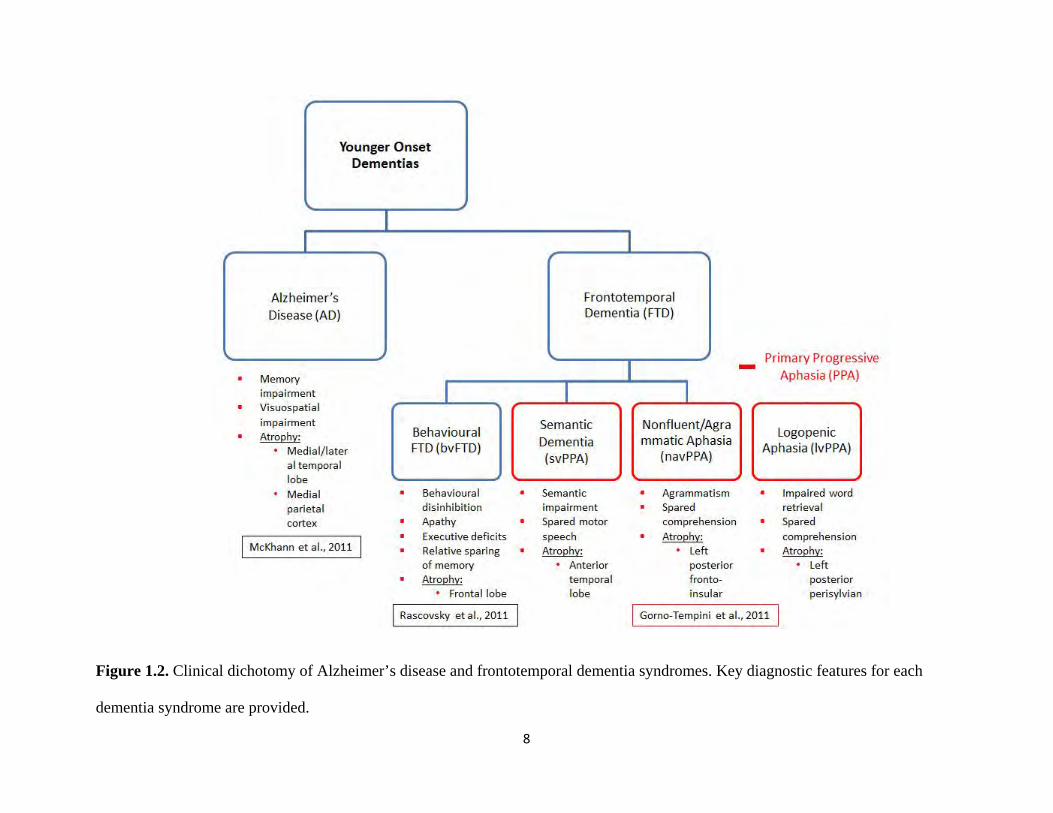
8
Figure 1.2. Clinical dichotomy of Alzheimer’s disease and frontotemporal dementia syndromes. Key diagnostic features for each
dementia syndrome are provided.

9
At clinical presentation, a number of cognitive features and patterns of brain atrophy are
used to distinguish between AD and FTD dementia syndromes for initial diagnosis. AD patients
present predominantly with significant episodic memory impairment across standardized delayed
(~ 15 min) visual/verbal recall measures with atrophy affecting medial temporal lobe regions, in
particular the hippocampus, extending posteriorly to include medial and lateral parietal lobe
brain regions bilaterally (McKhann et al., 2011). BvFTD patients present predominantly with
changes in social behaviour and personality, including disinhibition and apathy, with relative
sparing of memory, and frontal lobe atrophy, in particular affecting the orbitofrontal cortex,
anterior cingulate gyrus, and insula (Rascovsky et al., 2011; Seeley et al., 2008). PPA variants
are distinguished according to divergent patterns of speech and language features, and associated
cortical atrophy (Gorno-Tempini et al., 2011). SvPPA patients, otherwise known as semantic
dementia, demonstrate a conceptual loss of knowledge and lateralized (most commonly, left)
atrophy of the anterior temporal lobe. NavPPA and lvPPA patients present with impairments to
speech resulting from agrammatism and impaired word retrieval, and left lateralized atrophy in
the posterior fronto-insular and perisylvian regions, respectively. In contrast to svPPA, however,
navPPA and lvPPA patients show intact comprehension on standardized measures of language
(Gorno-Tempini et al., 2011).
Despite these established clinical features for clinical diagnosis for each dementia
syndrome, patients commonly present with a myriad of concomitant cognitive deficits across
multiple domains (i.e., memory, behaviour, language), making correct diagnosis during life
difficult. Consequently, pathological confirmation at post-mortem remains the gold standard for
confirming diagnosis (Fig. 1.3). AD pathology is defined by the abnormal accumulation of
insoluble formations of extracellular beta-amyloid (Aβ) (i.e., senile plaques) and progressive

10
intracellular tau accumulation (i.e., neurofibrillary tangles) (Braak & Braak, 1991; Braak &
Braak, 1998). In FTD, pathology is associated with a number of proteinopathies (Mackenzie et
al., 2010), with the most prominent ones being tau, TAR-DNA-binding protein 43 (TDP-43) and
RNA-binding protein fused in sarcoma (FUS) (Chare et al., 2014). Clinico-pathological studies
have shown, using current diagnostic guidelines, underlying pathology within clinical variants of
FTD patients is overlapping and heterogeneous, with a proportion of patients also having
underlying AD pathology (Chare et al., 2014; Rohrer, Rossor, & Warren, 2012; Seelaar, Rohrer,
Pijnenburg, Fox, & van Swieten, 2011). Pathology is relatively homogenous in some clinical
variants in the FTD spectrum, such as svPPA (TDP-43) and lvPPA (AD pathology). In contrast,
cases of bvFTD show the greatest degree of pathological heterogeneity (Chare et al., 2014),
which reflects the difficulty of accurate diagnoses in the early stages of the disease, particularly
when episodic memory is impaired (Hornberger, Piguet, Graham, Nestor, & Hodges, 2010).
Memory performance is regarded as one of the key discriminants between AD and FTD, and
should be preserved relative to behaviour and language deficits for respective FTD variants
(Gorno-Tempini et al., 2011; Rascovsky et al., 2011). Patients across all variants of FTD do,
however, present with varying degrees of episodic memory impairment on standardized
neuropsychological screening (Hodges & Graham, 2001; Hornberger et al., 2010; Hornberger &
Piguet, 2012; Piguet, Leyton, Gleeson, Hoon, & Hodges, 2015).

11
Figure 1.3. Neuropathology in Alzheimer’s disease and frontotemporal dementia syndromes.
Episodic Memory in Alzheimer’s Disease and Frontotemporal Dementia
Episodic memory impairment is a hallmark feature of AD attributed to underlying hippocampal
pathology present in the earliest stages of the disease (Braak & Braak, 1998; McKhann et al.,
2011). In contrast, impairment in ‘frontal’ related features, including behavioural, language and
executive dysfunction, are associated with clinical variants of FTD (Gorno-Tempini et al., 2011;
Rascovsky et al., 2011). Given that episodic memory impairments in AD, the most prominent
symptom, must reach a level such that there is interference with everyday life, to advance from
the prodromal diagnosis of mild cognitive impairment (Albert et al., 2011; McKhann et al.,
2011), memory performance is regarded as a key diagnostic discriminant between AD and FTD
(Gorno-Tempini et al., 2011; Rascovsky et al., 2011). Furthermore, for a long time, memory

12
impairment has been regarded as an exclusion criterion for a diagnosis of FTD (Neary et al.,
1998). There is now, however, consistent evidence across pathologically confirmed cases of FTD
that not only can patients present with episodic memory impairments (Binetti, Locascio, Corkin,
Vonsattel, & Growdon, 2000; Caine, Patterson, Hodges, Heard, & Halliday, 2001; A. Graham et
al., 2005; Hodges et al., 2004; Papageorgiou et al., 2016), but is also an initial symptom that
appears alongside changes in behaviour and language (Binetti et al., 2000), and closely
associated with tau pathology (Papageorgiou et al., 2016). In FTD, episodic memory impairment
has long been documented in the semantic variant (svPPA) (Adlam, Patterson, & Hodges, 2009;
K. S. Graham, Simons, Pratt, Patterson, & Hodges, 2000; Hodges & Graham, 2001). More
recently, increasing evidence suggests significant episodic memory impairment is also present in
autopsy confirmed behavioural variant (bvFTD) patients (Caine et al., 2001; A. Graham et al.,
2005), and in some cases have been reported to reach the same level of impairment as AD
(Bertoux et al., 2014; Hornberger et al., 2010; Pennington, Hodges, & Hornberger, 2011). In one
study, Bertoux and colleagues (2014) showed that in bvFTD patients that had been screened for
the absence of AD pathology in-vivo via positron emission tomography of Aβ deposition,
episodic memory demonstrated chance level of diagnostic sensitivity compared to AD. This has
significant implications for differential diagnosis of bvFTD from AD in the early stages whereby
there may be a similar level of concomitant clinical features, including executive dysfunction,
disinhibition, and memory deficits. Particularly given that executive dysfunction and behavioural
features can also be observed in AD patients (Binetti et al., 1996; McKhann et al., 2011).
Episodic memory can be broadly conceptualized as a 3 stage process, whereby new
information is first encoded, then stored or consolidated, and later retrieved. Clinical measures of
episodic memory (i.e., Rey Auditory Verbal Learning Test; California Verbal Learning Test;

13
Rey Complex Figure Test; Doors and People Memory Test) typically involve a single or multiple
encoding trials whereby patients are explicitly shown a set of visual or verbal stimuli which they
are asked to immediately recall. This is then followed by a delayed (~ 10 - 15 min) recall and
recognition condition (i.e., participants shown or read stimuli and asked to indicate if they are
new or previously seen). In AD, recall memory performance on tasks of this nature have
consistently been reported to be impaired during encoding and delayed retrieval conditions
(Salmon, 2000; Salmon & Bondi, 2009; Schoemaker, Gauthier, & Pruessner, 2014; Weintraub,
Wicklund, & Salmon, 2012), such that all 3 stages of episodic memory processing appears to be
affected by AD pathology. While the same pattern of deficit emerges for recognition memory,
performance appears to show greater variability compared to recall memory, and in some studies
did not differ significantly compared to matched healthy elderly controls (Dalla Barba, 1997;
Rauchs et al., 2007). It should be noted that this may result from neglecting the rate of false
positive recognition responses, which have been shown to be elevated in AD (Budson, Daffner,
Desikan, & Schacter, 2000).
Longitudinal studies of episodic memory in AD have demonstrated that there is a long
preclinical stage, at least 7 years, of stable annual decline prior to symptom onset (Backman,
Small, & Fratiglioni, 2001; Linn et al., 1995; Small, Fratiglioni, Viitanen, Winblad, & Backman,
2000). Interestingly, memory and general cognitive performance appears to follow a pattern of
gradual decline during the prolonged preclinical stage of AD (Backman et al., 2001; Linn et al.,
1995; Small et al., 2000) and after disease onset (Hsieh, Hodges, Leyton, & Mioshi, 2012).
Furthermore, there appears to be an abrupt period, immediately preceding diagnosis of dementia,
whereby memory declines rapidly (Backman et al., 2001; Small et al., 2000). These longitudinal
findings are, however, based on annual or biennial neuropsychological screening. Development

14
of novel measures able to monitor change in memory performance across a more sensitive time-
frame (i.e., days, weeks, months) may help to further characterise the pattern of decline
immediately preceding onset of dementia.
Relative to AD, studies of memory in FTD are not as common and have gained attention
only in recent years. Of particular interest are dissociations in episodic memory performance
between AD and the semantic (svPPA) and behavioural (bvFTD) variants of FTD (Hornberger &
Piguet, 2012). SvPPA patients are characterised by a loss of semantic information (i.e.,
conceptual knowledge of the world) that has a specific adverse impact on verbal memory. On
clinical measures of verbal memory, svPPA are significantly impaired, often to the same level as
AD (Perry & Hodges, 2000). In stark contrast, svPPA performance on visual recall and
recognition is intact and similar to healthy controls (Graham, Becker, & Hodges, 1997). Their
semantic deficits, however, become apparent when perceptually different learning and test items
(i.e., different colored telephones) are used to assess visual recall and recognition (Graham et al.,
2000). This has been suggested to reflect a deficit during the encoding of new episodic memory,
such that the lack of semantic information is being compensated by perceptual input (Hornberger
& Piguet, 2012).
In regard to AD and bvFTD, recent studies have increasingly shown that the profile of
episodic memory performance in these two patient populations are overlapping, which can
present a significant challenge to early clinical diagnosis (Bertoux et al., 2014; Hornberger et al.,
2010; Pennington et al., 2011). In particular, episodic memory performance on standard visual
and verbal recall and recognition measures show significant variability across bvFTD patients
(Bertoux et al., 2014; Hornberger et al., 2010; Irish, Piguet, Hodges, & Hornberger, 2014;
Pennington et al., 2011; Ranjith, Mathuranath, Sharma, & Alexander, 2010), which holds

15
significant difficulty for interpreting performance on standardized clinical memory measures. As
a whole, however, bvFTD patients do show significantly better episodic memory function than
AD, particularly on measures of delayed recall (Perry & Hodges, 2000; Wicklund, Johnson,
Rademaker, Weitner, & Weintraub, 2006). Notably, AD and bvFTD patients appear to show
dissociation on paired associative recall tasks in the early stages of disease (Blackwell et al.,
2004; Lee, Rahman, Hodges, Sahakian, & Graham, 2003) rather than traditional free recall
measures. Paired associative recall tasks, such as the Paired Associate Learning Test from the
Cambridge Neuropsychological Testing Battery, in which visual test stimuli are encoded with a
specific location, appears to be selectively impaired in the early stages of AD (Blackwell et al.,
2004), but intact in bvFTD (Lee et al., 2003). Recognition memory, while variable, can also be
intact in bvFTD, with some studies reporting no significant difference in performance to healthy
controls (Hornberger et al., 2010; Hutchinson & Mathias, 2007). This suggests that, in contrast to
the encoding deficits seen in svPPA, the episodic memory impairments reported in bvFTD may
be largely due to retrieval deficits, given their improved performance when cues are provided.
Collectively, the aforementioned studies suggest traditional clinical recall and recognition
measures can be improved for differential diagnosis of AD and bvFTD, particularly in the early
stage of disease. Development of a long-term memory assessment (i.e., days, weeks, months), in
particular, examining contextual memory retrieval (i.e., stimuli and location association), which
remains unexplored, may show greater sensitivity in differentiating between early disease cases
of AD and bvFTD (Blackwell et al., 2004; Lee et al., 2003; Perry & Hodges, 2000; Wicklund et
al., 2006). The application of long-term prospective memory assessment in diagnosis of AD and
bvFTD has a neuroanatomical basis. Recent studies of in-vivo and post-mortem memory circuit
integrity in AD and bvFTD suggests hippocampal volume is a poor structural marker, as

16
extensive hippocampal atrophy during the disease course of both conditions has been reported
(de Souza et al., 2013; Hornberger et al., 2012). There does, however, appear to be dissociation
in the integrity of extra-hippocampal memory structures along the Papez circuit, namely the
anterior thalamus, which appears to be selectively affected in FTD, but not AD (Hornberger et
al., 2012). The anterior thalamus is an important hub for hippocampal and frontal lobe
connectivity, with anatomical pathways connecting the hippocampus and prefrontal cortex
(Aggleton, Dumont, & Warburton, 2011). Notably, the hippocampus and prefrontal cortex are
suggested to play an interactive role in selective retrieval of long-term contextual information
(Preston & Eichenbaum, 2013). Preston and Eichenbaum (2013) posit a temporal relationship
whereby hippocampal involvement decreases while prefrontal involvement increases as the
degree of contextual details fades and the target memory becomes more generalized. Differences
in the integrity of hippocampal-prefrontal pathways may result in marked differences in rate of
memory retention, after controlling for level of initial memory encoding. As a first step, patients
with neurodegenerative conditions such as thalamic stroke are ideal to assess the assumption that
selective damage to the anterior thalamus results in accelerated long-term forgetting of newly
learnt contextual information. As evidenced by previous investigations of episodic memory in
thalamic stroke patients, focal lesions to the anterior thalamic and medio-dorsal nuclei of the
thalamus results in anterograde amnesia similar to when the hippocampus is damaged (Carlesimo
et al., 2011).

17
Spatial Memory in Alzheimer’s Disease and Frontotemporal Dementia
Apart from episodic memory, spatial memory deficits are another key early cognitive feature of
AD, functionally resulting in impaired navigational ability and topographical disorientation
(Kwok, Yuen, Ho, & Chan, 2010; Pai & Jacobs, 2004). While spatial memory processing can be
conceptualized into the same 3 stages as episodic memory (encoding, storage, retrieval), two
fundamentally different reference frames can be used to form an internal working representation
of the environment, egocentric (i.e., location of objects relative to yourself) or allocentric (i.e.,
location of objects relative to other objects). As mentioned earlier, there is significant overlap in
neural substrates underlying episodic and spatial memory, in particular, along the Papez circuit.
A key distinction is that egocentric representations tend to be supported predominantly by medial
parietal lobe structures, while allocentric representations are largely dependent on hippocampal
function (Baumann & Mattingley, 2010; Epstein, Parker, & Feiler, 2007; Ino et al., 2007;
Takahashi et al., 1997; Vann et al., 2009; Zaehle et al., 2007). Notably, both regions are affected
in the earliest stages of AD pathology (Braak & Braak, 1998; Nestor, Fryer, Ikeda, & Hodges,
2003; Whitwell et al., 2011), which is consistent with findings that both egocentric and
allocentric spatial processes can be impaired during spatial navigation in AD (Bellassen, Igloi, de
Souza, Dubois, & Rondi-Reig, 2012; Burgess, Trinkler, King, Kennedy, & Cipolotti, 2006;
Kalova, Vlcek, Jarolimova, & Bures, 2005; Laczo et al., 2009; Laczo et al., 2012; Morganti,
Stefanini, & Riva, 2013; Serino et al., 2014). In contrast, this does not appear to be the case in
FTD, with spatial memory reported to be relatively intact (Bellassen et al., 2012; Pengas et al.,
2010; Yew, Alladi, Shailaja, Hodges, & Hornberger, 2013). Spatial memory performance may,
therefore, be a useful diagnostic discriminant for early differential diagnosis in AD and FTD.

18
Spatial memory has been studied across a number of real and virtual experimental tasks
in AD with the most widely used being the Hidden Goal Task (Kalova et al., 2005; Serino et al.,
2014). This task is analogous to the classical Morris Water Maze used in rodents (Morris,
Garrud, Rawlins, & O'Keefe, 1982). The core feature of the task involves navigating to an
invisible, but progressively learnt, location within a confined environment containing a number
of external landmarks along the perimeter (Kalova et al., 2005). Findings using this task
consistently demonstrate AD as well as amnestic mild cognitive impairment patients perform
significantly worse than healthy controls on egocentric and allocentric conditions (Hort et al.,
2007; Kalova et al., 2005; Laczo et al., 2012; Serino et al., 2014). Performance on the Hidden
Goal Task, however, appears to primarily require participants to adopt an allocentric strategy and
has been shown to be heavily dependent on the hippocampus (Laczo et al., 2012; Nedelska et al.,
2012). During training, initial spatial locations are encoded using an allocentric framework.
Consequently, performance during the egocentric condition is reliant on translating spatial
locations from an allocentric to egocentric framework. Indeed, increasing evidence suggests that
impaired ability to integrate egocentric and allocentric spatial information underlies spatial
navigation impairments in AD (Burgess et al., 2006; Morganti et al., 2013; Pai & Yang, 2013;
Serino et al., 2014).
Studies of spatial memory in FTD are few and have primarily focused on comparing AD
and svPPA (Luzzi et al., 2015; Pengas et al., 2010), or included multiple clinical FTD variants
(Bellassen et al., 2012). Across all studies, FTD patients consistently outperformed AD patients
on spatial memory performance (Bellassen et al., 2012; Luzzi et al., 2015; Pengas et al., 2010).
Interestingly, while not stated by the authors, findings by Bellassen and colleagues (2012)
indirectly suggests that translation of spatial information between egocentric and allocentric

19
frameworks is intact in FTD, but impaired in AD. The study (Bellassen et al., 2012) employed a
novel virtual environment whereby participants were tested on sequential navigation and route-
tracing that assessed their capacity to translate egocentric sequences of body turns while
navigating onto an allocentric spatial map. This procedure follows a similar design to the
experimental task carried out by Morganti and colleagues (2013) demonstrating impaired
egocentric-allocentric translation in AD.
As a whole, the aforementioned studies highlight a significant dissociation in spatial
memory performance in the early stages of AD and FTD. Nevertheless, objective assessment of
spatial memory is often overlooked during routine clinical assessment in these patient cohorts.
There appears to be a selective deficit in the translation between different frameworks used to
determine location within an environment that is present in AD, but not FTD. A structure located
at the tail of the posterior cingulate, the retrosplenial cortex, has been proposed to serve as a hub
that is anatomically suited to integrate multi-modal spatial information from egocentric and
allocentric processes (Vann et al., 2009), and has been shown through functional neuroimaging
in healthy individuals to be critical for judgements of heading direction (Baumann & Mattingley,
2010; Iaria, Chen, Guariglia, Ptito, & Petrides, 2007; Marchette et al., 2014). Notably, the
posterior cingulate and, in particular, the retrosplenial region (Brodmann Areas 29/30) is
significantly affected during the early or prodromal stages of AD (Hornberger et al., 2012;
Nestor et al., 2003; Pengas, Hodges, Watson, & Nestor, 2010; Whitwell et al., 2011), while in
the early stages of FTD atrophy is typically found in the anterior cingulate (Hornberger et al.,
2012; Seelaar et al., 2011; Seeley et al., 2008). Development of clinically feasible measures
assessing the cognitive functions of spatial memory (i.e., judgement of heading direction) may
prove to be a sensitive diagnostic discriminant of early AD and FTD, in particular bvFTD.

20
Neuroimaging Principles
Over the past decade, neuroimaging has become a powerful tool that has become deeply
integrated into cognitive research in healthy individuals as well as in patient populations.
Different MRI modalities allow the study of structural change in grey and white matter, as well
as functional activity and connectivity of different brain regions in-vivo. A core feature of MRI is
the deployment of a powerful, and uniform, magnetic field in combination with the ability to
transmit, and receive, radio frequency (RF) energy to disrupt the alignment of protons contained
in nuclei within water molecules located in body tissue (Bitar et al., 2006; Pooley, 2005). By
plotting and decomposing the returning RF signal, released when the nuclei return to their resting
alignment, as corresponding levels of intensity (i.e., shades of grey) in a matrix arrangement of
pixels, in-vivo images can be generated providing an accurate and location specific visualization
delineating the brain’s intrinsic tissue properties (i.e., cerebrospinal fluid [CSF], white matter,
grey matter). Furthermore, by varying RF pulses based on repetition time (i.e., time between
successive pulse sequences) and echo time (i.e., time between transmitting and receiving the
echo signal), MRI is able to adjust the contrast and brightness of brain tissue. Clinically, 3
different pulse sequences (T1, T2, Flair) are commonly used for structural MRI (Bitar et al.,
2006). T1-weighted pulse sequence provides the greatest balance in contrast between different
types of brain tissue and, in particular, creates the greatest contrast between grey and white
matter, making it ideal for automated segmentation of grey matter for structural neuroimaging
analyses. In contrast, T2-weighted and Flair pulse sequences are more sensitive to CSF and
demyelination, respectively.
Advanced applications of MRI are also widely available to detect changes in metabolic
function, such as change in the magnetic state of hemoglobin (Hb) during oxygen saturation

21
(Gore, 2003), which forms the basis of functional neuroimaging research when the brain is
monitored either at rest or paired with cognitively demanding tasks. Diffusion weighted imaging
(DWI) is another application of MRI used to detect proton displacement in water molecules
(Acosta-Cabronero & Nestor, 2014; Hagmann et al., 2006) to make inferences on the integrity of
white matter tracts in the brain. The physical basis of DWI lies in Brownian motion, otherwise
known as ‘self-diffusion’, whereby molecules will undergo pseudo-random kinetic fluctuations
as a result of intrinsic thermal energy (Einstein, 1905). While diffusion of water molecules
follows a random trajectory in an unrestricted environment, in a restricted environment, such as
the brain, diffusion is hindered by cellular membranes or other microstructural components. In
particular, diffusion of water molecules will occur in a principal direction along myelinated
bundles of axons composing white matter tracts. In neurodegenerative conditions, interpretations
of white matter integrity is based on the assumption that changes resulting from disease, such as
permeability of cellular membranes, axonal loss, or demyelination, will be reflected by metrics
derived from diffusion imaging.
In the following experimental chapters, structural and diffusion imaging analyses were
carried out in combination with novel behavioural tasks to determine neural substrates
underlying cognitive processes, or independently to characterise in-vivo neural changes resulting
from disease pathology. Analyses were performed using the comprehensive neuroimaging
package, FMRIB Software Library (FSL).
FSL is an all-inclusive software package providing a range of toolboxes to analyze
imaging data across different MRI modalities. While the processing pipeline utilized across
different toolboxes will vary, all imaging analyses follow 3 principal stages: i) preprocessing, ii)
normalization, and iii) localization (Fig. 1.4). Each individual’s scan are initially processed

22
individually, however, statistical comparisons are always performed at the group level. During
the normalization stage, individual scans are normalized and registered to a standard brain
template whereby statistical comparisons of regional change across patient groups occur. Current
neuroimaging techniques provide a good representation of differences at a population level
between groups, but poorly at an individual level, limiting its clinical utility. Concerns have been
raised regarding the reliability of reported imaging findings in clinical research due to gross
anatomical differences when aligning healthy and atrophied brains, resulting in artificial results
(Bookstein, 2001; Davatzikos, 2004). Rather than being a limitation, however, these concerns
highlight the need for imaging processing pipelines specific to the disease population, such as
ensuring appropriate thresholding, registration algorithms, and attentiveness to misalignment of
data are being implemented (Ashburner & Friston, 2000; Pereira et al., 2010; Ridgway et al.,
2009). In the experimental chapters, pulse sequences used during acquisition of MRI data and
consequent imaging processing pipelines are based on established parameters previously
reported in the literature (Bitar et al., 2006; Hagmann et al., 2006). Typically, imaging findings
in the dementia literature appear to be robust and show reasonably consistent findings in disease
related regional change while employing fundamentally different techniques (Du et al., 2007;
Irish et al., 2014), including convergence between in-vivo and post-mortem data (Hornberger et
al., 2012).

23
Figure 1.4. Overview of the principal components of imaging analyses in FSL.
Structural Brain Imaging in FSL
Group-wise structural brain imaging analyses in FSL are conducted using a technique called
voxel-based morphometry (VBM). VBM is a method that provides an unbiased assessment of
grey matter volume in-vivo (Ashburner & Friston, 2000). As mentioned previously, T1-weighted
MRI pulse sequence generates excellent contrast between grey and white matter, and CSF, which
makes this type of data ideal for automated segmentation in structural analyses. In the
experimental chapters, a semi-automated processing pipeline was employed in FSL to perform
key components of VBM analysis. First, brain extraction was carried on each individual’s T1-
weighted scan to remove the skull and non-brain matter using FSL’s brain extraction algorithm
(Smith, 2002), followed by manual inspection of each processed scan for quality. The brain is
then parcellated into individual voxels, and grey matter is segmented, resulting in each voxel

24
containing an intensity value for grey matter corresponding to a specific location (Zhang, Brady,
& Smith, 2001). Each individual’s scan is normalized and averaged to generate a study specific
brain template. Template scans are then registered to a standard brain template (i.e., the Montreal
Neurological Institute standard brain), such that scans are now represented according to a
standardized x, y, z coordinate space, and reported regional differences can be corroborated and
compared across studies. Parametric and non-parametric voxel-wise statistical tests are then
performed either across the whole brain or masked for regions of interest to determine significant
grey matter change. A unique aspect of FSL involves the manner in which statistically
significant clusters are formed. FSL employs threshold-free cluster enhancement (TFCE) which
does not require the setting of an arbitrary cluster forming threshold (Smith & Nichols, 2009). In
contrast to traditional cluster-based thresholding, TFCE has been shown to improve sensitivity
over a wide range of MRI signal strengths (Smith & Nichols, 2009).
Diffusion Brain Imaging in FSL
Group-wise diffusion analyses in FSL are conducted using a technique called tract-based spatial
statistics (TBSS). TBSS is a method for in-vivo assessment of diffusion MRI data to provide a
quantitative assessment of white matter integrity (Smith et al., 2006). As mentioned previously,
diffusion imaging is in essence a measure of the degree in which water molecules follow a
restricted or unrestricted trajectory of movement along myelinated axons comprising white
matter tracts. A DWI represents the best estimate of the rate of water diffusion at each voxel of
the brain. To provide a representation of the degree and principal direction of diffusion within
each voxel, a tensor model is applied, resulting in a diffusion tensor image (DTI). Previous
studies have demonstrated that, during acquisition, the minimum number of directions, in which
diffusion is measured, must range between 20 – 30 for robust reconstruction of tensor models

25
(Batchelor, Atkinson, Hill, Calamante, & Connelly, 2003; Jones, 2004; Papadakis, Murrills, Hall,
Huang, & Carpenter, 2000). In the experimental chapters, DWI was acquired using 32 directions.
Using DTI data, scalar metrics, representing degree of free and restricted diffusion, can then be
generated, such as fractional anisotropy (FA) and mean diffusivity, as well as axial diffusivity
and radial diffusivity. FA is the most commonly reported DTI metric in the literature, acting as a
proxy measure of white matter integrity. In TBSS, whole-brain FA maps from each individual’s
DTI image are normalized to a standard brain template (i.e., the Illinois Institute of Technology
standard brain) and represented in a standard coordinate space. Individual FA maps are then
skeletonized, using FSL’s thinning algorithm, to define the lines of maximum FA, which
correspond to the centers of white matter tracts (Smith et al., 2006). Voxel-wise statistical tests
are then run on skeletal FA maps in a manner similar to VBM.
The strong point of the TBSS processing pipeline is the unique skeletonization
algorithm employed for DTI analysis, which significantly improves the inter-subject alignment
for group analysis (Smith et al., 2006). While the TBSS processing pipeline provides good inter-
subject registration, the registration process is performed using FA maps, a scalar metric, rather
than the full tensor model. Critically, tensor based registration maintains voxel orientation,
information which is lost when converting to scalar DTI metrics. Recent studies have shown
tensor based registration results in superior construction of unbiased spatially normalized group
templates of the brain using diffusion MRI data (Bach et al., 2014; Hui Zhang et al., 2007;
Keihaninejad et al., 2013), with good reproducibility of DTI metrics (Keihaninejad et al., 2013).
This is of particular importance in longitudinal diffusion imaging (Mahoney et al., 2015),
whereby multiple registration and normalization steps are necessary (Fig. 1.5). In chapters 4 and
5, tensor based registration was performed using DTI-TK, an independent tool, prior to
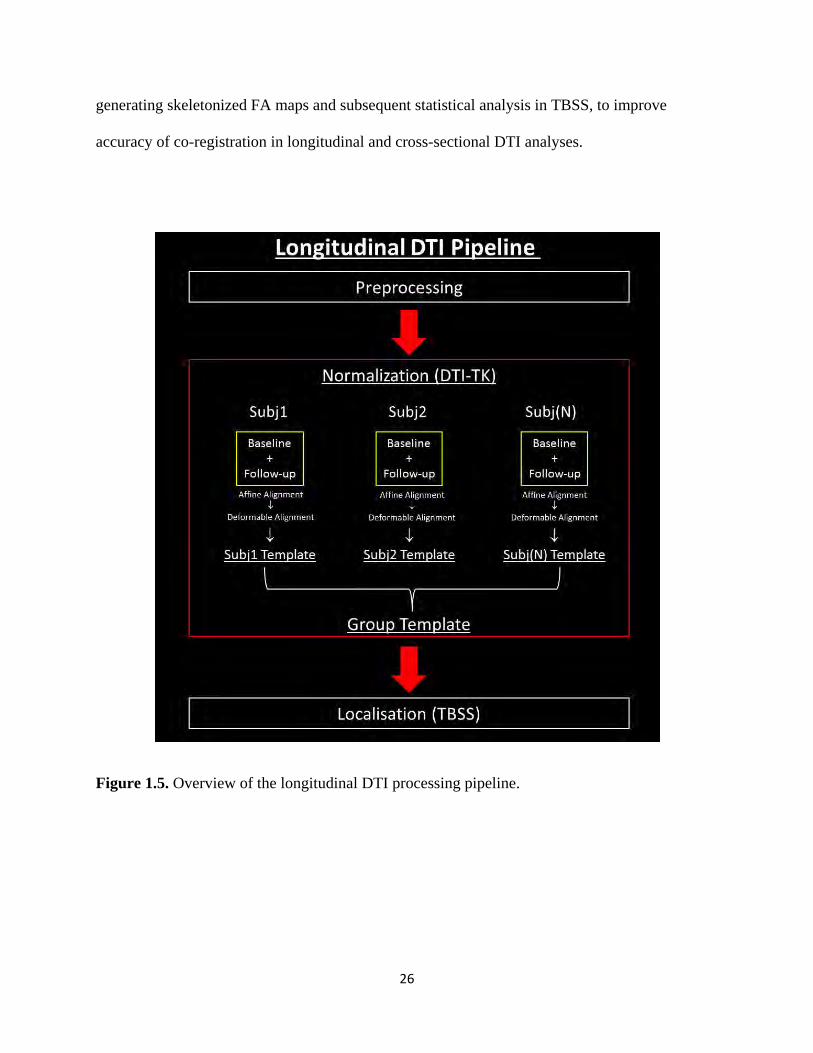
26
generating skeletonized FA maps and subsequent statistical analysis in TBSS, to improve
accuracy of co-registration in longitudinal and cross-sectional DTI analyses.
Figure 1.5. Overview of the longitudinal DTI processing pipeline.

27
Aims and Hypotheses
As stated at the start of the chapter, the primary focus of this thesis is to investigate cognitive
change beyond the hippocampus in AD and FTD in the context of episodic and spatial memory.
Specifically the aims are:
1. Assess the diagnostic utility of heading orientation in differential AD and bvFTD
diagnosis. There is a distinctive difference in spatial memory performance between AD
and FTD. A direct comparison between AD and bvFTD remains unexplored, but holds
significant diagnostic utility as an additional clinical feature to episodic memory, which
holds a variable level of diagnostic sensitivity. Atrophy in posterior regions of the brain,
including the posterior cingulate and retrosplenial region, which are critical for
judgements of direction, is also selectively affected in the early stages of AD, but not
bvFTD. This neuroanatomical dissociation is hypothesized to translate into a selective
behavioural deficit in the ability to maintain heading orientation when adopting an
egocentric strategy, rather than allocentric.
2. Assess the impact of thalamic integrity on long-term contextual memory. The
thalamus is an important relay structure mediating hippocampal-prefrontal connectivity,
which is suggested to play a key role in long-term contextual memory retrieval. Lesions
focal to the thalamus is hypothesized to result in different rates of long-term retention of
contextual memory in patients with thalamic stroke. Establishing the feasibility and
sensitivity of long-term contextual memory assessment in thalamic stroke patients will
inform future assessment in more densely amnesic dementia conditions, particularly
given the thalamus is proposed to be more selectively affected in FTD, but not AD.

28
3. Characterise longitudinal in-vivo integrity of the Papez memory circuit in AD and
FTD. Distinct differences in the integrity of structures comprising the Papez circuit have
been shown in-vivo and at post-mortem in AD and FTD. The pattern of focal change
along the Papez circuit longitudinally in-vivo remains unexplored. Divergent patterns of
grey matter atrophy and integrity of white matter tracts is hypothesized to underlie
progressive decline on episodic memory in AD and FTD.
4. Assess the diagnostic utility of longitudinal diffusion MRI in svPPA and lvPPA.
Employing tensor-based registration to enhance standard DTI processing in TBSS
provides a significant increase in co-registration accuracy, resulting in increased
statistical power that becomes particularly evident in longitudinal analyses. Longitudinal
characterisation of in-vivo white matter change has significant diagnostic implications in
FTD, where underlying disease pathology is heterogeneous. Primary pathology in svPPA
(TDP-43) and lvPPA (AD pathology) are different, but relatively homogenous within
each respective clinical variant. It is hypothesized that pathological differences in these
patient cohorts will drive different patterns of white matter degradation in well-defined
clinical cases.

29
Chapter 2 – Spatial Orientation in Dementia
Publication I – “Lost in spatial translation – A novel tool to objectively assess spatial
disorientation in Alzheimer’s disease and frontotemporal dementia”
Publication II – “Egocentric vs. allocentric spatial memory in behavioural variant
frontotemporal dementia and Alzheimer’s disease” (in submission)

30
Available online at www.sciencedirect.com
ScienceDired
ELSEVIER Journal homepage: www.elsevier.com / locate/cortex
Research report
Lost in spatial translation - A novel tool to objectively assess spatial disorientation in Alzheimer's disease and frontotemporal dementia
Cross Mark
Sicong Tu a,b,c, Stephanie Wong a,b,c, john R. Hodges a,b,c,
Muireann Irish a,b,d, Olivier Piguet a,b,c and Michael Hornberger b,c,e, *
• Neuroscience Research Australia, Randwick, Sydney, Australia b Australian Research Council Centre of Excellence in Cognition and its Disorders, Sydney, Australia c School of Medical Sciences, University of New South Wales, Sydney, Australia d School of Psychology, University of New Scuth Wales, Sydney, Australia c Department of Clinical Neurosciences, University of Cambridge, Cambridge, United Kingdom
ART ICL E INFO
Article history: Received 27 November 2014 Reviewed 5 March 2015 Revised 19 March 2015 Accepted 23 March 2015 Action editor Stefano Cappa Published online 2 April 2015
Keywords: Orientation
Retrosplenial cortex Alzheimer's disease Frontotemporal dementia
ABSTRACT
Spatial disorientation is a prominent feature of early Alzheimer's disease {AD) attributed to degeneration of medial temporal and parietal brain regions, including the retrosplenial cortex {RSC). By contrast, frontotemporal dementia (FTD) syndromes show generally intact spatial orientation at presentation. However, currently no clinical tasks are routinely administered to objectively assess spatial orientation in these neurodegenerative conditions. In this study we investigated spatial orientation in 58 dementia patients and 23 healthy controls using a novel virtual supermarket task as well as voxel-based morphometry (VBM). We compared performance on this task with visual and verbal memory function, which has traditionally been used to discriminate between AD and FTD. Participants viewed a series of videos from a first person perspective travelling through a virtual supermarket and were required to maintain orientation to a starting location. Analyses revealed significantly impaired spatial orientation in AD, compared to FTD patient groups. Spatial orientation performance was found to discriminate AD and FTD patient groups to a very high degree at presentation. More importantly, integrity of the RSC was identified as a key neural correlate of orientation performance. These findings confirm the notion that i) it is feasible to assess spatial orientation objectively via our novel Super· m arket task; ii) impaired orientation is a prominent feature that can be applied clinically to discriminate between AD and FTD and iii) the RSC emerges as a critical biomarker to assess spatial orientation deficits in these neurodegenerative conditions.
© 2015 Elsevier Ltd. All rights reserved.
• Corresponding author. Departmen t of Clinical Neurosciences, University of Cambridge, Cambridge, CB2 OSZ, United Kingdom. E·mail address: [email protected] .ac.uk (M. Hornberger).
http:// dx.doi.org/1 0.1016/j .cortex. 2015.03.016 0010·9452/© 2015 Elsevier Ltd. All rights reserved.

31
84 CO RT EX 6 7 (2015) 83 - 94-
1. Introduction
Spatial and temporal disorientation is a well-documented early symptom of Alzheimer's disease (AD) (Hornberger, Piguet, Graham, Nestor, & Hodges, 2010; Pai & Jacobs, 2004; Pengas, Hodges, Watson, & Nestor, 2010, Pengas, Patterson, et a!., 2010; Yew, Alladi, Sha ilaja, Hodges, & Hornberger, 2013). For patients diagnosed with one of the frontotemporal dem entia (FTD) syndromes, however, orientation is reported to be relatively intact (Bellassen, Igloi, de Souza, Dubois, &
Rondi-Reig, 2012 ; Pengas, Hodges, et a1., 2010, Pengas, Pa tter· son, et a!., 2010; Yew eta!., 2013). This raises the question of whether orientation can be used as a discriminant of AD and FTD, in particular, between AD and the behavioural variant of FTD (bvFTD), where significant memory impairment in a subset of bvFTD patien ts can lead to diagnostic uncertainty (Hornberger eta!., 2010).
Spatial navigation in general has been well studied in de· mentia patients including mild cognitive impairmen t (MCI), the prodromal stage of AD (for a review see Serino, Cipresso, Morganti, & Riva, 2014). Investigations of orientation in dement ia patients, however, h ave been limited, given the lack of suitable, and practical, tasks t hat can be easily utilised in a clinical setting. Orientation can be characterised as being either egocentric or allocentric; cognitive processes which are subserved by different brain regions. Egocentric spatial orientation (i.e., location of objects in relation to the self) has been suggested to be dependent on parietal cortices while a llocentric spatial orientation (i.e., location of objects in relation to other objects) is critically dependent on medial tern· poral lobe structures, including the hippocampus (Burgess, Becker, King, & O'Keefe, 2001). Significant structural and metabolic changes are present in the parietal lobe and retro· splenial region (Brodm ann Areas 29 and 30) in AD (Nestor, Fryer, Ikeda, & Hodges, 2003; Pengas, Hodges, et a!., 2010;
Tan, Wong, Hodges, Halliday, & Hornberger, 2013), but not bvFTD (Irish, Piguet, Hodges, & Hornberger, 2014; Tan eta!., 2013). Egocentric spatial orientation may be, therefore, a suitable measure to discriminate between the two conditions. The importance of the retros plenial region for spatial orien · tation has been highlighted in a case report of a taxi driver who suffered foca l left retrosplenial haemorrhage and immediately presented with selective egocentric spatial disorientation (lno eta!., 2007). Evidence from functional imaging studies further suggests that egocentric navigation is subserved by the parietal cortex and, in particular, the retro· splenial cortex (RSC) for heading direction (for a review see, Boccia, Nemmi, & Guariglia , 2014).
The specialised role of the RSC in orientation during spatial navigation has been consistently demonstrated across functional neuroimaging studies (Baumann & Matt ingley, 2010;
Epstein, Parker, & Feiler, 2007; !aria, Chen, Guariglia, Ptito, &
Petrides, 2007; Marchette, Vass, Ryan, & Epstein, 2014). The RSC is the gateway to key occipital, temporal, and parietal lobe structures responsible for processing visual information, constructing an internal model of the environment (allocentric framework) and updating directional information based on movement from the motor system, respectively (Vann, Aggleton, & Maguire, 2009). Consequently, the RSC acts as a
neural hub for the integration and processing of egocentric, allocentric and visual information necessary to orientate oneself within an environment (Epstein & Vass, 2013; Vann et a!., 2009). Functional imaging studies have consistently shown activity in the RSC in healthy young participants dur· ing tasks involving orientation within a learnt virtual envi· ronment, when making judgements of relative direction (Baumann & Mattingley, 2010; Epstein et a!., 2007; Marchette e t a!., 2014), and a lso during active navigation using land· m arks as reference (!aria et a!. , 2007). Multi-voxel pattern analysis carried out by Marchette eta!. (2014) indicated that the location of environmental fea tures, in addition to directional information, is encoded within the ne ural activity elicited by the RSC.
While the aforementioned studies have implemented behavioural tasks that excel in evoking RSC involvement, assessment of orientation is predicated on the accurate acquisition and formation of an internal representation of a new experimental environment and landmarks (with the exception of Epstein eta!., 2007), a process which is critically dependent on the hippocampus (Boccia eta!., 2014; Ekstrom et a!., 2003; Hirshhom, Grady, Rosenbaum, Winocur, &
Moscovitch , 2012; !aria et a!., 2007). In patients with episodic memory deficits (i.e., com promised hippocampal func tion) both the t ime requ ired, and demands of the initial learning phase would be significantly increased, reducing efficacy in a clinical setting. To our knowledge, the current most ecologi· cally valid assessment of orientation in memory impaired patients involve topographical map assessments of land· m arks within a patient's local city or surrounding locale (Campbell, 1 Iepner, & Miller, 2014; Pai & Yang, 2013), similar to that implemented by E?stein eta!. (2007). These tasks, how· ever, are limited to participants familiar with specific envi· ronments (i.e., downtown Sydney), but can be overcome as in the case of the personalised versions used by Pai and Yang (2013), where they targeted unique landmarks near each participan t's residence. Therefore, a spatial orientation task that does not require prior training and widely applicable to objectively assess memory impaired patients is necessary.
In the current study, we utilised a virtual supermarket environment that does not require prior learning of a spatial layout to assess spatial orientation in AD and FTD. Partici· pan ts viewed the environment from a first person perspective and maintained spatial orientation using an egocentric frame of reference. Spatial orientation performance was, therefore, dependent on two variables: i) incidental formation of a working egocentric representation of the environment, and ii) updating egocentric memory in response to movement through the environment (Land, 2014). AD, and fTD patients diagnosed with the behavioural (bvFTD) or semantic (SD) variants were tested - both have shown to have hippocampal but not RSC atrophy. We aimed to assess: i) the clinical applicability of the virtual supermarket task in these patient cohorts, ii) sensitivity of spatial orientation as a diagnostic discriminan t between AD and bvFTD, and iii) neural correlates of spatial orientation in AD. We hypothesized th at while orientation is dependent on memory processes, the retro· s plenial region would be critical for egocentric spatial orientation, such that spatial orientation would be associated with reduced structural integrity of the RSC.

32
2. Meth ods
2 .1. Participa nts
Fifty eight dementia patients (20 AD; 24 bvFTD; 14 SD) and 23
age- and education-matched healthy controls were recruited from t he Sydney frontotemporal dementia research group (FRONTIER) database. All participants were assessed at the FRONTIER clinic located at Neuroscience Research Australia, Sydney. Study approval w as provided by the South Eastern Sydney Local Health District Human Research Ethics Committee. All participants provided signed consent for neuropsychological assessment a nd neuroimaging prior to testing. Patient cohorts were matched for disease duration and clinical disease severity. All dementia patients fulfilled internat ional consensus criteria for AD (McKhann et a!., 2011) , bvFTD (Rascovsky eta!., 2011), and SD (Gorno-Tempini eta!. , 2011).
Clinical diagnoses were established by consensus among sen ior neurologist, occupational therapist and neuropsychologist, based on a clinical interview , comprehensive neuropsychological assessment, and evidence of brain at ro phy on structural neuroimaging. All bvFTD patients showed disease progression as well as atrophy on scans to exclude any phenocopy cases (Kipps, Hodges, & Hornberger , 2010). Participant demographics and clinical characteristics are p rovided in Table 1 .
Briefly, AD patients presen ted predominantly with significant episodic memory impairment with preserved social behaviour. BvFTD patients demonstrated changes in social funct ioning, loss of insight , disinhibition an d increase d
85
apathy. SD patients were predomin antly left lateralised (3 right) and showed loss of general conceptual knowledge in t he form of significant naming and comprehension impairment. Exclusion criteria for all participants in clu ded prior history of mental illness, h ead injury, movement disorders, alcohol and drug abuse, limited English proficiency, and, for controls , presence of abnormality on MRI.
Participant s were administered a battery of cognitive tests to assess overall cognitive func tion, verbal and visual mem ory, and working memory. This assessment included: Addenbrooke's Cognitive Examinat ion-Revised (ACE-R), Rey Auditory Verbal Learning Test (RA VL T), Rey Com plex rigure Test (RCIT), and Digit Span. ror a brief description of cognitive tasks see Supplementary Table 1.
2.2 . Virtual supermarket task
Spatial orientation was assessed using an ecologica l virtual supermarket environment. The layout of the virtual environment d id no t include any notable landm arks and any spatial representation was acquired through incidental encoding during test trials. A total of 14 video t rials (2 sections of 7
v ideos) were created from an English version of the 'Virtual Supermarket' (Water lander, Scarpa, Lentz, & Steenhuis, 2011) based on Australian a nd New Zealan d supermarkets. Videos were presented from a fi rs t person perspective and partic ipants were taken to set locations t hroughout t he supermarket, which involved moving while making a series of 90" turns (Fig. 1) . Participants were a sked to imagine that they were standing behind a trol'ey and pushing it to diffe rent locations of the supermarket. At the end of e ach trial, participant s h ad
Table 1 - Participant demographic ch aracteristics a nd p erforman ce on s ta ndardised neuropsychological assessm ents .
AD (n ~ 20)
Sex (M/F) 10 M, 10 F Handedness (UR/B) 4 L, 16 R Age (y.o) 66.7 (8) Education (yrs) 12.1 (3.3) Disease durat ion (yrs} 5.8(4.8) CDR (SOB) 3.5 (2.1) ACE-R Total (/100) 70.2 (9.1) Memory (126) 12.4 (4.5) Orientation (110) 7.3 (2.1) RAVLT Tl-5 (175) 25 (6.9) 30 min delay (115) 1.4 (1.6) Recognition (115) 10.5 (2.8) RCFT Copy (136) 25.3 (7.7) Delayed (/36) 3.5 (3.5) Digit sp an Forward (116) 8.9 (1.9) Backward (114) 4.3 (1.4)
bvFTD (n ~ 24)
19M, 5 F 1L, 22 R,1 B 64.7 (9.3) 11.8 (3.1) 6.6 (4.3) 4.6 (3.7)
82.1 (10.2) 19.6 (5.2) 9 (1.3)
36.2 (11.3) 6.2 (4.1) 11.9 (2.5)
29.6 (5.6) 11.3 (7.8)
9.5 (1.6) 5.3 (1.7)
SD (n ~ 14)
7 M, 7 F 3 L, 11 R
64.5 (8)
13.7 (1.9) 5.9 (1.9) 3.9 (3.2)
63.4 (20.9) 14.9 (5.8) 8.3 (2.7)
31.5 (2. 6) 12.9 (7.3)
9.5 (2.6) 6.7 (2.1)
Controls (n ~ 23)
11 M,12 F 1 L, 22 R
68 (3.4) 13.3 (3.1)
96.3 (2.4) 24.7 (1.4) 9.9 (.3)
54.4 (8.6) 11.3 (2.4) 14 (1.3)
32.8 (3.4) 19.9 (8.2)
11.5 (2.3) 8 (2.8)
Group effect
n!s n!s n!s n!s
AD vs. bvFTD
n!s n!s n!s n!s
n!s
n!s
n!s n!s
Note: Clinical deme ntia rating (CDR) sum o f boxes; Adde nbrooke's cognitive exam ination revised {ACE-R); Rey auditory verbal learning test {RAVLT); Rey complex figure test {RCFT). Beh avioural da ta on the RA VL T, RCFT and Digit Spa n was available for 16 control p a rtic ipa n ts. RAVL T
scores were available f rom 16 AD and 17 bvFTD patie nts; SD patients were no t adminis ter ed the RAVLT due to the ir language impairmen t . RCFT scores were ava ilable from 16 AD, 21 bv iTD, a nd 13 SO patie nts. Digit Spa n scores were available from 18 AD, 24 bvFTD a nd 13 SO patie n ts . n/s - not s ignificant. "p < .01. '"p < .005.

33
86 CORTEX 6 7 (201 5) 83 9 4
Finish
'
Section 1
II II
• e =Starting Location
Section 2
+
x = Section 1 Final Locations
+=Section 2 Final locations
Fig. 1 - Screens hots from an example trial from Section 1 (left) and spatial layout of the virtual supermarket (right). The video begins at the starting location and involves 3 x 90" turns to arrive at the final location. Participants were asked to respond with the direction to the starting location from the final location.
to indicate the direction of the starting location. All t rials began at the same location, but followed different routes to reach a different end point in each trial. Each trial within each section was standardised for length and number of turns (Section 1: 20 sec, 3 turns; Section 2: 40 sec, 5 turns). For all participants, Section 1 was administered first, followed by Section 2. No feedback was provided during test trials.
Prior to testing, participants were instructed they would be viewing a number of short videos that involved moving to different locations of a supermarket. After arriving at the new location, they would be required to make a decision about the direction of the original starting location. Participants were explicitly told they would start from the same starting location across trials and asked to keep track of the direction of the starting location throughout the videos. At the end of each trial, participants are shown a snapshot of the final location and cued by the onscreen text ("[n which direction is the starting location?'') to provide a response (Fig. 1). Critically, correct directional responses could not be made from only viewing the final screenshot. The task itself does not require any training component to successfully complete test trials and limits prior participant exposure of the supermarket layout to a brief practice trial. A practice video trial (10 sec, 2 turns), was given at the start of testing to introduce participants to the virtual supermarket environment and make sure task instructions were well understood. [n particular, the practice trial aimed to make clear that the direction, not path taken, of the starting location from the final location was requested.
Participants were made aware that only a general direction that involved a d istinction on two principal components (i.e.,
left/right and front/behind) was required. [n most cases, participants spontaneously pointed to a particular d irection. Some patients, h owever, required direct prompts by the task administrator (i.e., ' [s the starting location to the left or right of where you are now?'; ' [s the starting location in front of or behind where you are now?'). Segregating responses in this manner allowed for better comprehension and accurate responding from patients with greater generalised cognitive impairment. Previous versions of the task attempted using a circular illustration representing a 360" field of view segmented into 4 quartered sections (i.e ., left/front; right/ fr ont; right/behind; left/behind) for respondin g. While elderly control participants had no difficulty responding in this manner, a number of patients showed confusion leading to inaccurate responding. Spatial orientation performance in the current version of the task was scored on individual directional components (UR; F/B) as well as on an overall score, which required a correct response on both directional components. Each directional component, and overall performance, in Sections 1 and 2 were analysed independently. Overall performance was, however, the key variable of interest.
2.3. Statistical analyses
Differences in participant group demographics, performance on standard cognitive tests were assessed using one-way analysis of variance (ANOVA). Orientation performance on the experimental task were assessed u sing multivariate analysis of covariance (MANCOVA) and two-tailed post hoc multiple comparisons to compare spatial orientation

34
performance between groups while taking into account degree of memory impairment on standard cognitive tests in SPSS 21.0 (IBM Corp., Armonk, NY).
A composite memory score was created by averaging performance on the memory component of the ACE-R and delayed recall components on the RAVLT and RCFT as a percentage of the total score. For participants with missing assessments, a composite score was calculated if performance on at least 2 of the 3 memory components w ere available. Composite memory performance was compared with averaged overall spatial o rientation performance on Sections 1 and 2 of the experimental task using logistic regression. Receiver operating characteristic (ROC) curves of sensitivity and specificity were also calculated using the method by DeLong, DeLong, and Clarke-Pearson (1988) in MedCalc fo r Windows, version 14.8.1 (MedCalc Software, Ostend, Belgium). In all analyses, p values< .OS were considerec. stat istically significant.
2.4. Imaging acquisition
Whole-brain s tructural T1 images were acquired for all par ticipants using a 3T Philips MRI scanner with standard quadrature head coil (eight channels). Structural T1 scan s were acquired as follows: coronal orientation, matrix 256 x 256, 200 slices, 1 mm isotropic, TE/TR ~ 2.5/5.4 ms, flip angle a ~ 8°. Prior to analyses, all participant scans were visua lly inspected for significant head movements and artefacts, and excluded from imaging ana lyses. Scans were missing from 7 control participants. Imaging analyses included MRI data from 16 AD, 18 bvFTD, 12 SD and 15 control participants. All scans were examined by a rad iologist for s tructural abnormalities .
2.5. Imaging analyses
Voxel-based morphometry (VBM) w a.s conducted on wholebrain T1-weighted scans, using the VBM toolbox in FMRIB's Software Lib rary (FSL; http://www.fmrib.ox.ac.uk/fsV). First, t he brain was extracted from each scan using FSL's BET algorithm with a fractional intensity threshold of .22 (Smith, 2002). Each scan was visually checked following brain extraction to ensure no brain ma tter was excluded, a nd no non-brain matter was included. A study specific template of grey matt er w as genera ted from 12 scans from each participant cohort. An equivalent number of scans from each cohort were used to create the template, avoiding potential bias towards any single group's topography during registration. Te mplate sca ns were then registered to the Montreal Neurological Institu te (MNI) s tandard brain (MNI 152) us;ng non-lin ear b-spline representation of the registration warp field, resulting in a st udy-specific grey matter template at 2 mm3 resolution in MNI standard space. Simultaneously, participant brainextracted sca ns were processed with the FMRIB's Automatic Segmentation Tool (FAST) (Zhang, Brady, & Smith, 2001), via a hidden Markov random field model and an associated Expectation-Maximization a lgorithm, segmen ting brain tissue into CSF, grey matter and white matter. The FAST algorithm a lso corrected scans for spatial intensity variations such as bias field or radio-frequency inhomogeneities, resulting in partial volume maps. The following step saw grey ma tter partial volume maps th en non -linearly registered to the
87
s tudy-specific template via b-spline representa tion of the registration warp. These maps were then modulated by dividing by the Jacobian of the warp field , to correct for any contraction/enlargement caused by t he non-linear component of the transformation. After n ormalisation and modulation, grey matter maps were smoothed using an isotropic Gaussian kernel (sigma~ 3 mm).
Statistical analysis was performed w ith a voxel-wise gene ral linear model. Significant clusters w ere formed by employing the threshold-free clus ter enhancement (TFCE) method (Smith & Nichols, 2009). TFCE is a cluster-based thresholding method which does not require t he setting of an a rbitrary cluste r forming threshold. Instead , it takes a raw statist ics image and produces an output image in w hich the voxel-wise values represent the amount of cluster-like local spatial support . The TFCE image is t hen turned into voxelwise p-values via ?ermutation testing. We employed permutat ion-based non-parametric testing with 5000 permutations (Nichols & Holmes, 2002).
Comparisons of whole-brain grey matter in tegrity were carried out between each patient group a nd controls, as w ell as between AD a nd bvFTD cohorts. Report ed clusters a re corrected for multiple comparisons via Family-wise Error (FWE) and tested for significa nce at p < .005. Talairach and Harvard-Oxford CorticaVSubcortical Atlases were used as references to identify brain s tructures comprising significant clust ers. A mask of the RSC (Brodmann areas 29, 30) was manually traced on the MNI 152 standard brain and used to calculate each participant 's grey matter vo lume in this region. Whole-brain and RSC grey matter were correlated w ith avera ged overall o rientation performance across Sections 1 and 2.
3. Results
3.1. Demographics and cognitive testing
Participant cohorts were well ma tched for demograph ic variables, an d patient groups were ma tched for disease duration a nd disease severity (Table 1; all p values> .1). ANOVA of participan t groups' performance across standard cognitive test s revealed significant group d ifferen ces for all components (all p values < .003). In the two groups of in terest, bvFTD showed a be tter cognit ive profile than AD on the ACE-R screening of general cognition (all p values < .01), verbal memory (RAVLT: T1-5, 30 min delay; all p values < .003) , and v isual memory (RCFT: Delayed; p ~ .009). The two patient groups, however, did not differ on workin g memory as indicated by the Digit Span forwards (p > .7) and backw ards (p > .4). Import antly, all aspects of episod ic memory in bvFTD patients were significantly impa ired compared to controls (Supplementary Table 2; a ll p values < .02).
3.2. Spatial orientation performance
Spatial orien tation was scored for correc t response on the two direct ional components (front/back and left/righ t). Overall performa nce required correct judgem ent of orien tation on both directional components (Fig. 2). MANCOVA was performed using m emory performance on th e ACE-R as a
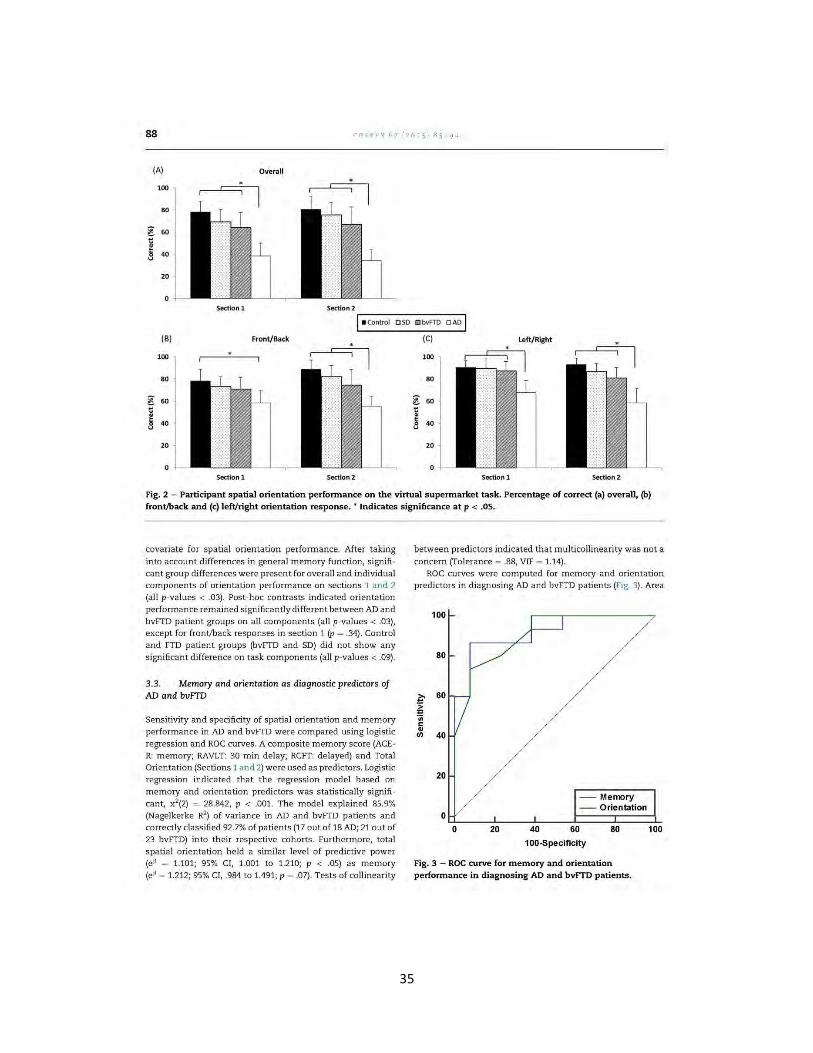
35
88 COR TEX 6 7 ( 201 5) 83 94
(A) Overall
100
80
g 60 t: ~ s v
40
20
0
Section 1 Section 2
!•control O SD O bvFTD DAD I (B) Front/Back (C) Left/ Right
100 100
80 80
~ 60 l 60 t:
~ 40
t:
~ 40
20 20
0 0
Section 1 Section 2 Section 1 Section 2
Fig. 2 - Participant spatial orientation performance on the virtual supermarket task. Pe rcentage of correct (a) overall, (b) front/back and (c) left/right orientation response. • Indicates significance at p < .OS.
covariate for spatial orientation performance. After taking into account differences in general memory function, signifi · cant group differences were present for overall and individual components of orientation performance on sections 1 and 2 (all p-values < .03). Post-hoc contrasts indicated orientation performance remained significantly different between AD and bviTD patient groups on all components (all p-values < .03), except for front/back responses in section 1 (p = .34). Control and ITD patient groups (bviTD and SD) did not show any significant difference on task components (all p-values < .09).
3.3. Memory and orientation as diagnostic predictors of AD and bvFTD
Sensitivity and specificity of spatial orientation and memory performance in AD and bviTD were compared using logistic regression and ROC curves. A composite memory score (ACER: memory; RAVLT: 30 min delay; RCIT: delayed) and Total Orientation (Sections 1 and 2) were used as predictors. Logistic regression indicated that the regression model based on memory and orientation predictors was statistically significant, x2(2) = 28.842, p < .001. The model explained 85.9% (Nagelkerke R2
) of variance in AD and bviTD patients and correctly classified 92.7% of patients {17 out of 18 AD; 21 out of 23 bviTD) into their respective cohorts. Furthermore, total spatial orientation held a similar level of predictive power (eP = 1.101; 95% CI, 1.001 to 1.210; p < .05) as memory (eP = 1.212; 95% CI, .984 to 1.491; p = .07). Tests of collinearity
between predictors indicated that multicollinearity was not a concern (Tolerance = .88, VIF = 1.14).
ROC curves were computed for memory and orientation predictors in diagnosing AD and bviTD patients (Fig. 3). Area
z. ~ ·;;;
" .. rn 40
20
-- Memory -- Orientation
0~--~--~--L===~==l 0 20 40 60 80 100
100.Specificlty
Fig. 3 - ROC curve for memory and orientation performance in diagnosing AD and bvFTD patients.

36
under t he curve (AUC) values indicated memory (AUC ~ .918, SE ~ .052; 95% CI, .751-.988) and total orientation (AUC ~ .905, SE ~ .054; 95% CI, .734- .983) had a similar leve l of diagnostic accuracy. Pairwise comparison of memory and orientation ROC curves revealed no significant difference between the two pre dictors (p ~ .87).
3 .4. Structu ral imaging results
Whole-brain grey matter integrity was examined using VBM to compare patient cohorts with healthy controls (Supplementary Table 3; Supplementary Fig. 1). The pattern of alrophy presen l in each paLie nL gro•.1p was consisle nl wiLh previous reports in the literature (Irish eta!., 2014; Rohrer et a!., 2008). Briefly, AD patients showed temporal and parietal lobe atrophy. In particular, grey matter integrity was reduced in the retrosplenial region as well as bilateral hippocam pi. In bvFTD patients, only clusters in the medial prefrontal cortex was found to significantly differ, compared to controls, after t hresholding. In SD patients, atrophy was foun d in the left medial prefrontal cortex and t emporal lobes. Notably, SD patients also showed significant bilateral atrophy in the h ippocampus, with greater atrophy in the left hippocampus, due to the inclusion of both left and right latera!ised SD cases .
VBM analyses were a lso conducted between AD and FTD patient groups (Table 2). Findings indicated AD patients showed significantly greater atrophy in medial parietal and retrosplenial regions, compared to bvFTD patients (Fig. 4A). Similarly, compared to SD, AD pa tie nts showe d greater atrophy in media l pariet al and right lateral parietal lobe regions. ReporLed c:luslers were conecLed fot n1ulliple con1parisons using FWE correction a nd significant at p < .005 .
In AD, total orie ntation (Section s 1 and 2) performance was correlated with whole-brain grey m atter integr ity to determine the neural correla tes of their impaired performance (Fig. 4B). Orientation performance was found to correlate wit h t he ret rosplenial region (Brodmann areas 23, 29, 30; MNI coordinates: 6, - 46, 24) as well as the left lingual gyrus (MNI co-ordinates: - 14, - 66, - 6) . Whole brain volume did not show a significant correlat ion with orientation performance.
4. Discussion
The current study demonstrated that spatial orientation ca n be u sed to discriminate between AD and bvFTD beyond their memory impairment. The virtual supermarket task was
89
successfully used to assess spatial orient ation in amnesic dementia patient populations with hippocampal atrophy. Notably, orientat ion was impaired in AD, but relatively intact in FTD patient groups, even after accounting for differences in performance on episodic memory tasks. Orien tation performance showed the same level of diagnostic sensitivity as standa rdised measures o f episod ic memory. Th is finding is cons istent with surrogate reports of temporal and spatial disorientation in everyday life during t he early stages of AD (Kw ok, Yuen, Ho, & Chan, 2010; Pai & Jacobs, 2004), but not in FTD, and formally a ddressed orientation performance beyond the context of a general screening of cognition or clin ical interview.
The ability to orient ourselves to topographical features within our immediate environment requires an internal working representation (egocent ric memory) of objects relative to head and body orientation (Land, 2014). A key featu re of this internal model of the outside world is t he ability to continually update the directional relationship between external objects and the self Our experimental ta sk aimed to mimic this p rocess by engaging participants within t he context of a novel, but familiar, supermarket shoppin g scenario whereby they were taken to various locations within the s tore, while having to main tain and update the directional relationship to the starting location. The task aimed to engage egocentric memory with a re latively low allocentric spatial map contribution of the supermarket environment, which is suggested to be formed and stored in the hippocampus and medial tem poral cortices (Burgess , 2006; Burgess eta!., 2001 ; Byrne , Becker, & Burgess, 2007). Egocentric and allocentric representations are complementary processes for navigating the rea l world a nd information from each framework freely updates the other (Burge ss, 2006; Land, 2014; Vann et a!., 2009). Here , however, reduced integrity of parietal, rather than temporal lobe, struc:ures w as associated with impaired orientation performance in AD patien ts, which would support the view that t he experimental task is assessing egocentric memory.
A number of virtual reality tasks based on route learning a nd hidden goal parad;gms have previously been developed to assess egocentric a nd allocentric spatial processing in AD and MCI (Serino et a!., 2014). Findings indicate deficits in allocentric and egocentric spatial representations (Bellassen et a!., 2012; )heng & Pai, 2009; Laczo et a!. , 2012; Morgan ti, Stefanin i, & Riva, 2013; Plan cher, Tirard, Gyselinck, Nicolas , & Piolino, 2012; Weniger, Ruhleder, La nge, Wolf, & Ir!e, 2011; a lthough see Burgess, Trinkler, Kin g, Kennedy, & Cipolotti ,
Table 2 - Voxel-based morphometry results showing regions of significant grey matter intensity differences between AD and FTD patien t groups.
Contrast Regions Hemisphere MNI co-ordinates Number ofvoxels
X y z
bvFTD > AD Precuneus/ Posterior cingulate!BA23/BA30 Bilateral -70 34 1212 Lateral occipital cortex/Superior parietal lobe Right 46 - 66 42 556
SD > AD Precuneus/Posterior cingulate!BA23/BA30/BA31 Bilateral - 60 20 2260 Lateral occipital cortex/Superior parietal lobe Right 46 - 62 26 1706 Middle temporal gyrus Right 44 - 56 96
"BA - Brodmann area.
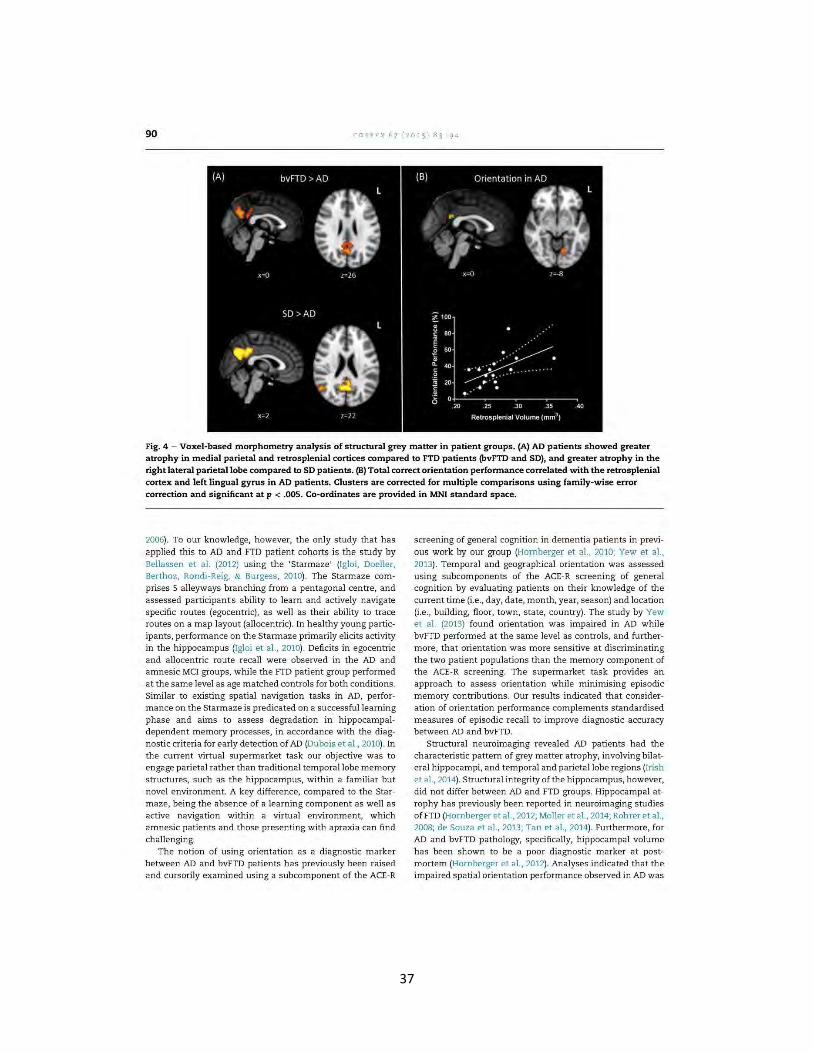
37
90 CORTEX 6 7 ( 201 5) 83 9 4
Fig. 4 - Voxel-based morphometry analysis of structural grey matter in patient groups. (A) AD patients showed greater atrophy in medial parietal and retrosplenial cortices compared to FTD patients (bvFTD and SO), and greater atrophy in the right lateral parietal lobe compared to SO patients. (B) Total correct orientation performance correlated with the retrosplenial cortex and left lingual gyrus in AD patients. Clusters are corrected for multiple comparison s using family-wise error correction and significant at p < .005. Co-ordinates are provided in MNI standard space.
2006). To our knowledge, however, the only study that has applied this to AD and FTD patient cohorts is the study by Bellassen et al. (2012) using the 'Starmaze' (Igloi, Doeller, Berthoz, Rondi-Reig, & Burgess, 2010). The Starmaze comprises 5 alleyways branching from a pentagonal centre, and assessed participants ability to learn and actively navigate specific routes (egocentric), as well as their ability to trace routes on a map layout (allocentric). In healthy young participants, performance on the Starmaze primarily elicits activity in the hippocampus (Igloi et al., 2010). Deficits in egocentric and allocentric route recall were observed in the AD and amnesic MCI groups, while the ITD patient group performed at the same level as age matched controls for both conditions. Similar to existing spatial navigation tasks in AD, performance on the Starmaze is predicated on a successful learning phase and aims to assess degradation in hippocampaldependent memory processes, in accordance with the diagnostic criteria for early detection of AD (Dubois et al., 2010). In the current virtual supermarket task our objective was to engage parietal rather than traditional temporal lobe memory structures, such as the hippocampus, within a familiar but novel environment. A key difference, compared to the Starmaze, being the absence of a learning component as well as active navigation within a virtual environment, which amnesic patients and those presenting with apraxia can find challenging.
The notion of using orientation as a diagnostic marker between AD and bvFTD patients has previously been raised and cursorily examined using a subcomponent of the ACE-R
screening of general cognition in dementia patients in previous work by our group (Hornberger et al., 2010; Yew et al. , 2013). Temporal and geographical orientation was assessed using subcomponents of the ACE-R screening of general cognition by evaluating patients on their knowledge of the current time (i.e., day, date, month, year, season) and location (i.e ., building, floor, town, state, country). The study by Yew et al. (2013) found orientation was impaired in AD while bviTD performed at the same level as controls, and furthermore, that orientation was more sensitive at discriminating the two patient populations than the memory component of the ACE-R screening. The supermarket task provides an approach to assess orientation while minimising episodic memory contributions. Our results indicated that consideration of orientation performance complements standardised measures of episodic recall to improve diagnostic accuracy between AD and bviTD.
Structural neuroimaging revealed AD patients had the characteristic pattern of grey matter atrophy, involving bilateral hippocampi, and temporal and parietal lobe regions (Irish et al., 2014). Structural integrity of the hippocampus, however, did not differ between AD and FTD groups. Hippocampal atrophy has previously been reported in neuroimaging studies ofiTD (Hornberger et al., 2012; Moller et al., 2014; Rohrer et al., 2008; de Souza et al., 2013; Tan et al., 2014). Furthermore, for AD and bvFTD pathology, specifically, hippocampal volume has been shown to be a poor diagnostic marker at postmortem (Hornberger et al., 2012). Analyses indicated that the impaired spatial orientation performance observed in AD was

38
related to redu ced grey matter volume in the left lingual gyrus and retrosplenial region of t he posterior cin gulate. This finding is consistent w ith the view that the RSC plays a central role in spatial navigation (for a review see Vann eta!., 2009). The RSC is suggested to act as a hub for the integration and t ranslat ion of different frameworks (i.e., visual information from the occipital cortex; body orientat ion from the pariet al cortex [egocentric]; spatial map of the environment from the hippocampus [allocentric)) and holds reciprocal anatomical connections w ith the occipital and parietal cortices, and the hippocam pal forma tion (Burgess, 2006; Burgess et a!., 2001; Byrne et a!., 2007; Vann et a!., 2009). Functional imaging studies in humans consistently elicit strong activation in the RSC when navigating through familiar environments (Vann et a!. , 2009). In particular, stud ies by Spiers and Maguire (2006) , and Baumann and Mattingley (2010), both observed strong activation of the RSC d uring retrieval of directional informat ion from topographical representations during spatial navigation tasks. Notably , the study by Baumann and Mattingley (2010) utilised a v irtual environment s tripped of all environmental cues creat ing an immediate sense of d isorientation. Pa rticipan ts were extensively trained to loca te a nd navigate to specific stimuli and la ter exposed to paired stim uli images represe nting either the same or differen t heading direct ions at test Retrieval of heading direction was found to activate the retrosplenial region fo r bot h conditions, but significa ntly higher wh en paired stimuli re presented different heading directions.
Human lesion studies also highligh t selective topographica l disorientation as a result of dam age to the retrosplenia! region (Ina et al., 2007; Osawa, Maeshima, &
Kunishio, 2008; Taka hashi, Kawamura, Shiota, Kasahata , &
Hirayama, 1997; a lt hough see Maeshima et a!., 2014). Patients with hippoca mpa l lesions, however , demonstrate impa ire d spatia l navigation, but a preserved sense of direction w ithin a familiar environment (Spiers & Maguire, 2007). In the curren t study, SD patients with confirmed h ippocam pal atrophy showed well preserved orientation on t he exper ime ntal task, wh ile AD patient s with at rophy in t he medial parietal lobe a nd re trosplen ia l region were severely im paired. Th ese behavioura l findings in AD a nd SD a re consistent with previous findings by Pengas, Hodges, et a!. (2010), Pengas, Patterson, et a!. (2010) us ing a virtu al route learning paradigm w ith active naviga tion in combin ation with a hea ding orientation test. AD patients prove d t o be sign ifican tly im paired on route learning as well as heading orien tation while SD patients showed no signific ant differences in performance to con trols. This same pattern of d issociation between AD, SD and a ge-matched control cohorts w as observed for o rientation performance in the curren t virtual supermarket task. Although Pengas, Hodges, eta!. (2010), Pengas, Patterson, et a !. (2010) discuss st ud ies in SD t ha t have demonstrated atrophy in medial t em poral lobe structures (Chan et a!., 2001; Davies, Graham, Xuereb, Williams, & Hodges, 2004), t he s tate of hippocam pal atrophy in their patien t cohorts is unclear. In t he current st udy, AD and SD pa tient groups sh owed bila tera l h ippocam pal a trophy compared to controls, but a direct con trast between AD a nd SD did not find any significant diffe re nces in the structure . This further suggests atrophy in the parietal lobe,
91
namely the re tros plenial region of the posterio r cingula te underlies observed orientation deficits in AD.
Behavioural and s:ruc tura l imaging analyses confirmed th at t he virtual supermarket task is a suitable me asure of spatial orientation, specific to expected AD pat hology and accompanying disorientation . More importantly, in contrast to other tasks it is clinically feasib le to use , as t he tot al time taken for each sect ion is only -7 min in t he dementia patients . Thus, inclusion of the Supermarket task in a clin ical se ttin g would allow more objective assessment of spat ial orientation deficits instead of only relying on th e generic orien tation com ponent in general cognitive screening tests . Some caveats, however, m us t be acknowledged. The supermarket environment (Water lander eta!., 2011) was designed to reflect a n accurate representation of real-life supermarkets and in t he curren t task was not s tripped of these natu ralist ic features to inc rease understanding and engage dementia patients . Therefore, compared to other tasks, such as the 'tunnel task ' whereby parti cipants a re also required to ma intain orientation to a starting location w ith in a topographica lly featureless tunnel environment (Schonebeck, Thanhauser, & Debus, 2001), the supermarket paradigm may no t be seen as a "pure" cognitive as sessment of spat ial orien tation . The task does , however, disc rimin ate between AD and bvFTD patients w ithin a clinical se tting. An other issue is the extent to which o rien tation performance is dependent on memory function. We add ressed this by including differences in general memory func tion as a covariate in our be havioural analyses, but the RSC which we identified as the key structure resulting in impaired orien tation perform ance in AD is also involve d in various m em ory processes , su ch as a utobiographical memory retrieval (Vann et a!. , 2009). Another pote n tial limitation is the lack of pathologica l confirmation in pa tie nts. Pat ien ts with AD and bvFTD can presen t with varying levels of memory impairment a nd th e current findings will need to be replica ted to
confirm t he efficacy of the supermarket t ask. In conclusion, d isorien tation is a significant impairment
p resen t in AD, but relatively in tact in FTD patien ts, w hich can be teased apart by assessing egocentric orientation. The neura l correlates associated with impaired o rien tat ion in AD include occipital and parie tal cortices, in particular the RSC.
Acknowledgements
We thank Dr. Waterlander for p rovid ing the virtual supermarket environment for the current study. This work was supported by fund ing to Forefront, a collaborative research group dedicated to the study of frontotemporal dementia and motor neurone disease, from th e National Health an d Medical research Council (NHMRC) of Au stralia program gran t (#1037746) , the Australian Resea rch Council (ARC) Cen tre of Excellence in Cognition and its Disorde rs Memory Node (#C£110001021) and an ARC Discovery Project grant (DP1093279) ; ST is supported by Alzheimer's Australia De me ntia Resea rch Foundation a nd NHMRC of Australia aw ards. OP is su pported by a 1\HMRC Career Development Fellowsh ip (APP1022684). MH is supported by Alzheimer Research UK and

39
92
the Isaac Newton Trust . These sources had no role in the s tudy design, collection, analyses and interpretation of data, writing
of the manuscript , or in the decis ion to submit the paper for publication. The authors declare no competing financial in terests. We are grateful to the research participants involved with t he ForeFront research studies.
Supplementary data
Supplement ary data related to this article can be found at h ttp://dx.doi .org/10.1016/j.cortex. 2015.03.016.
REFEREN C ES
Baumann, 0., & Mattingley,]. B. (2010). Medial parietal cortex encodes perceived heading direction in humans. The journal of Neuroscience, 30(39), 12897- 12901. http://dx.doi.orfi10.1523/ )NEUROSCI.3077-10.2010.
Bellassen, V., Igloi, K., de Souza, L. C., Dubois, B., & Rondi-Reig, L. (2012) . Temporal order memory assessed during spat iotemporal navigation as a behavioral cognitive marker fo r differential alzheimer's disease diagnosis. The journal of Neuroscience, 32(6), 1942- 1952. http://dx.doi.orfi10.1523/ )NEUROSCI.4556-11.2012.
Boccia, M., Nemmi, F., & Guariglia, C. (2014). Neuropsychology of environm ental navigation in humans: review and metaanalysis of FMRI studies in healthy participants. Neuropsychology Review, 24(2), 236- 251. http:// dx .doi .orfi 10.1007/ s11065-014-9247 -8.
Burgess, N. (2006) . Spatial memory: how egocentric and allocentric combine. Trends in Cognitive Sciences, 10(12), 551- 557. S1364-6613(06)00271-3 [pii]
Burgess, N., Becker, S., King, ]. A., & O'Keefe, ]. (2001). Memory for events and their spatial context: models and experiments . Philosophical Transactions of the Royal Society of London Series B, Biological Sciences, 356(1413), 1493-1503. http://dx.doi.orfi 10.1098/rstb.2001.0948.
Bu rgess, N., Trinkler, 1., King,]., Ken nedy, A., & Cipolotti, L. (2006). Impaired allocentric spat ial memory underlyin g topographical disorientation. Reviews in the Neurosciences, 17(1- 2), 239- 251.
Byrne, P., Becker, S., & Burgess, N. (2007). Rem embering the past and imagining the future: a neural model of spatial memory and imagery. Psychological Review, 114(2), 340- 375, 2007-05396-005 [pii] .
Campbell,]. I., Hepner, I. ] ., & Miller, L. A. (2014). The influence of age and sex on m emory for a familiar environment. Journal of Environmenta l Psychology, 40(0), 1-8. http://dx.doi.orfi10.1016/j. jenvp.2014 .04.007.
Chan, D., Fox, N. C., Scahill, R. 1., Crum, W. R. , Whitwell, ]. L., Leschzine r, G., et al. (2001). Patterns of tem poral lobe atrophy in semantic dementia and alzheimer's disease. Annals of Neurology, 49 (4), 433- 442.
Davies, R. R., Graham, K. S., Xuereb, ]. H., Williams, G. B., & Hodges, J. R. (2004) . The human perirhinal cortex and semantic memory. The European journal of Neuroscience, 20(9), 2441- 2446. EjN3710 [pii] .
DeLong, E. R., DeLong, D. M., & Clarke-Pearson, D. L. (1988). Comparing the areas under two o r more correlate d receiver operating characteristic curves: a nonparametric approach. Biometrics, 44(3), 837- 845. h ttp://dx.d oi.orfi10.2307/2531595.
Dubois, B., Feldman, H. H., )acova, C., Cummings,]. L., Dekosky, S. T., Barberger-Gateau, P., et aL (2010). Revising the definition of alzheimer's disease: a new lexicon. The lancet Neurology, 9(11), 1118-1127. h ttp://dx .doi.orfi10.1016/S1474-4422(10)70223-4.
Ekstrom, A. D., Kahana, M. )., Caplan, ]. B., Fields, T. A., Isham, E. A., Newman, E. L., et aL (2003). Cellular networks underlying h uman spatial navigation. Nature, 425(6954), 184-188. http://dx.do' .org/10.1038/ nature01964.
Epstein, R. A., Parker, W. E., & Feiler, A. M. (2007). Where am I now> distinct roles for parahippocam pal and retrosplenial cortices in place recognition . The journal of Neuroscience, 27(23), 6141- 6149, 27/23/6141 [pii].
Epstein, R. A., & Vass, L. K. (2013). Neural systems for landmark based wayfinding in humans. Philosophical T ransactions of the Royal Society of London. Series B, Biological Sciences, 369(1635), 20120533. http:/ /dx.doi.orfi10.1098/rstb .2012.0533.
Gorno-Tempini, M. L., Hillis, A. E., Weintraub, S., Kertesz, A, Mendez, M., Cappa, S. F., et aL (2011). Classification of primary progressive aphasia and its variants. Neurology, 76(11), 1006-1014. http://dx.doi.org/10.1212/WNL.Ob013e31821103e6.
Hirshhom, M., Grady, C., Rosenbaum, R S., Winocur, G., & Moscovitch, M. (2012). The h ippocampus is involved in mental navigation for a recently learned, but not a h ighly familiar environment: a longitudinal fMRI study. Hipp ocampus, 22(4), 842- 852. ht tp://dx.do '.org/10.1002/hipo.20944.
Hornberger, M., Piguet, 0. , Graham, A.]., Nestor, P. ]., & Hodges, ]. R (2010) . How preserved is episodic memory in behavioral variant frontotemporal dementia? Neurology, 74(6), 472- 479. http://dx.do '.org/10.1212/WNLOb013e3181cef85d.
Hornberger, M., Wong, S., Tan, R., Irish, M., Piguet, 0 ., Kril, ]., et aL (2012) . In vivo and post-mortem memory circuit integrity in frontotemporal dem entia and alzheimer's disease. Brain: A journal of Neurology, 135(Pt 10), 3015- 3025. http //dx.doi.orfi 10.1093/brain/aws239.
!aria, G., Chen,]., Guariglia, C., Ptito, A, & Petrides, M. (2007). Retrosplenial and hippocampal brain regions in human navigat ion: complemen tary func tional contributions to the formation and use of cognitive maps. European Journal of Neuroscience, 25(3), 890- 899. h ttp://dx.doi.orfi10.1111/j.1460-9568.2007 .05371.x.
Jglo i, K., Doeller, C. F., Berthoz, A., Rondi-Reig, L., & Burgess, N. (2010) . Lateralized hu:nan hippocampal activity predicts navigat ion based on sequence or place memory. Proceedings of the National Academy of Sciences of the United States of America, 107(32), 14466- 14471. http ://dx.doi.orfi10.1073/ pnas.1004243107.
!no, T. , Doi, T., Hirose, S., Kimura, T. , Ito,]. , & Fukuyama, H. (2007). Directional disorien tation following left retrosplenial hemorrhage: a case report with fMRI studies. Cortex, 43(2), 248- 254.
Irish, M., Piguet, 0 ., Hodges, ]. R., & Hornberger, M . (2014). Common and unique gray matter correlates of episodic memory dysfunction in frontotemporal dementia and alzheimer's disease. Human Brain Mapping, 35(4), 1422- 1435. h ttp://dx.doi.orgl10.1002/hbm.22263.
)heng, S. S., & Pai, M. C. (2009). Cognitive map in patients with mild alzheimer's disease: a computer-generated arena study. Behavioural Brain Research, 200(1), 42- 47. http://dx.doi.orfi 10.1016/ j .bbr.2008.12.029.
Kipps, C. M., Hodges, ]. R., & Hornberger, M. (2010) . Nonprogressive behavioural frontotemporal dementia: recen t developments and clin ical implications of the 'bvFTD phenocopy syndrome'. Current Opinion in Neurology, 23(6), 628- 632. h ttp://dx .doi .or fi10.1097/WCO. Ob013e3283404309.

40
Kwok, T. C., Yuen, K. S., Ho, F. K, & Chan, W. M. (2010). Getting lost in the community: a phone survey on the communitydwellin g demented people in h ong kong. lnternational}ournal of Geriatric Psychiatry, 25(4), 427-432. h rtp:// dx.doi .o rg/10.1002/ gps.2361.
Laczo, ]., Andel, R., Vyhnalek, M., Vlcek, K., Magerova, H., Varjassyova, A, et al. (2012). From morris water maze to computer tests in the prediction of alzheimer's disease. NeuroDegenerative Diseases, 10(1-4), 153- 157. h ttp://dx.doi.orgl 10.1159/000333121.
Land, M. F. (2014). Do we have an internal model of the outs ide world' Philosophical Transactions of the Royal Society B: Biological Sciences, 369(1636) h ttp://dx.doi.org/10.1098/rstb.2013.0045.
Maeshima, S., Osawa, A, Yamane, F., Yoshihara, T., Kanazawa, R., & Ishihara, s. (2014). Retrosplenial amnesia without topographic disorientation caused by a lesion in the nondominant hemisphere. journal of Stroke and Cerebrovascular Diseases, 23(3), 441-445. http://dx.doi.org/10.1016/ j .js trokecerebrovasdis .2013.03.026.
Marchette, S. A , Vass, L. K., Ryan, ]., & Epstein, R A (2014). Anchoring the neural compass : coding of local spatial reference frames in h uman medial parietal lobe. Nature Neuroscience, 17(11), 1598- 1606. h ttp//dx.doi.org/10.1038/ nn.3834 .
McKhann, G. M., Knopman, D. S., Chertkow, H., Hyman, B. T., Jack, C. R., Jr., Kawas, C. H., et al. (2011) . The diagnosis of dementia due to alzheimer's disease: recommendations from the national institute on aging-alzheimer's association workgroups on diagnostic guidelines for alzheimer's disease. Alzheimer's & Dementia, 7(3), 263- 269. http://dx.doi.orgl 10.1016/j.jalz.2011.03 .005.
Moller, C., Dieleman, N., van der Flie r, W. M., Vers teeg, A , Pijnenburg, Y , Scheltens , P., et al. (2014). More a trophy of deep gray matter structures in frontotemporal dementia com pared to alzheimer's disease. journal of Alzileimer's Disease, 200014591423TOPL (pii].
Morganti, F., Stefanini, S., & Riva, G. (2013). From allo- to egocentric spatial ability in early alzheimer's disease: a study with virtual reality spat ial tasks. Cognitive Neuroscience, 4(3- 4), 171-180. ht tp :!/dx.doi.org/10.1080/17588928 2013.854762.
Nestor, P. )., Frye r, T. D., Ikeda, M., & Hodges, ]. R (2003). Retrosplenial cortex (BA 29/ 30) h ypometabolism in mild cognitive impairment (prodromal alzheimer's disease) . The European Journal of Neuroscience, 18(9), 2663- 2667, 2999 [pii].
Nichols, T. E., & Holmes, A P. (2002). Nonparam etric permutation tests for functional neuroimaging: a primer with examples. Human Brain Mapping, 15(1), 1-25. http://dx.doi .org/10.1002/ hbm1058.
Osawa, A , Maeshima, S., & Kunishio, K (2008). Topographic disorientation and amnes ia due to cerebral hemorrhage in the left retrosplenial region. European Neurology, 59(1- 2), 79- 82, 000109572 [pii] .
Pai, M. C., & Jacobs, W. J. (2004). Topograph ical disorientation in community-residing patients with alzheimer's disease. 1nternational}ournal of Geriatric Psychiatry, 19(3), 250- 255. http:// dx .doi.org/10.1002/gps.1081.
Pai, M. C., & Yang, Y C. (2013). Impaired t ranslat ion of spat ial representation in young onset alzheimer's disease patients. Current Alzheimer Research, 10(1), 95- 103. CAR-EPUB-20121002-1 [pii].
Pengas, G., Hodges,]. R., Watson, P., & ;-.Jes tor, P. ]. (2010a). Focal posterior cin gulate a troph y in incipient alzheimer's disease. Neurobiology of Aging, 31(1), 25- 33. http://dx.doi.org/10.1016/ j.neurobiolaging.2008.03.014.
Pen gas, G., Patte rson, K., Arnold, R J., Bird, C. M., Bu rgess, N., & Nestor, P. J. (2010b). Lost and fo un d: bespoke memory tes ting
for alzheimer's disease and semantic dementia. Journal of Alzheimer's Disease, 21(4), 1347- 1365.
93
Plancher, G. , Tirard, A., Gyselinck, V., Nicolas, S., & Piolino, P. (2012). Using virtual reality to ch aracterize episodic memory profiles in amnestic mild cognitive impairment and alzheimer's disease: Influence of active and passive encoding. Neuropsychologia, 50(3), 592- 602. http://dx.doi.org/10.1016/ j. neuro psychologia .2011 .12. 013.
Rascovsky, K., Hodges, ; . R., Knopman, D., Mendez, M. F., Kramer, ]. H., Neuhaus,]., et al. (2011). Sensitivity of revised diagnostic criteria for the behavioural varian t of frontotemporal dementia. Brain, 134(Pt 9), 2456- 2477. h t tp:// dx.doi.orgl10.1093/brain/awr179.
Rohrer, ]. D., McNaught, E., Foster, ]., Clegg, S. L., Barnes, ]., Omar, R., et al. (2008). Tracking progression in frontotemporal lobar degeneration: serial MRI in semantic dementia. Neurology, 71(18), 1445- 1451. h ttp://dx.doi.org/10.1212/ 01.wnl.0000327889.13734.cd.
Schone beck, B., Thanhauser, )., & Debus, G. (2001). The "tun nel task": a method for exam ina tion of cognitive processes in spatial orientation performance (Die Tunnelaufgabe: Eine Methode zur Un ters uchung kognitiver Teilprozesse rau mlicher Orient ierungsle is tungen]. Zeitschrift Fur Experimentelle Psychologie : Organ Der Deutschen Gesellschaft Fur Psychologie, 48(4), 339- 364.
Serino, S., Cipresso, P., Morganti, F., & Riva, G. (2014). The role of egocentric and allocentric abilities in alzheimer's disease: a systematic review. Ageing Research Reviews, 16, 32- 44. h ttp:// dx.doi. org/10 .1016/j .arr .2014. 04.004.
Smith , S. M. (2002). Fast robust automated brain extraction. Human Brain Mapping, 17(3), 143- 155. http://dx.doi.orgl 10.1002/hbm.10062.
Smith , S.M., & Nichols, T. E. (2009). Threshold-free cluster enhancemen t: addressing problems of smoothing, threshold dependence and localisation in cluster inference. Neuro1mage, 44(1), 83- 98. http://dx .doi.org/10.1016/ j .neuroimage.2008.03.061.
de Souza, L. C., Ch upin, M., Bertoux, M., Lehericy, S., Dubois, B., Lamari, F., et al . (2013). Is hippocampal volume a good m arker to differentiate alzheimer's disease from frontotemporal dementia? journal of Alzheimer's Disease, 36(1), 57- 66. http:// dx.doi.org/10.3233/JAD-122293.
Spiers , H.). , & Maguire, E. A. (2006). Th oughts, behaviour, and brain dynamics during navigation in the real world . Neuroimage, 31(4), 1826- 1840. S1053-8119(06)00101-7 [pii].
Spiers, H.). , & Maguire, E. A. (2007). Th e neuroscience of remote spatial memory: a tale of two cities. Neuroscience, 149(1), 7- 27 . S0306-4522(07)00800-7 [pii].
Takah ashi, N., Kawamura, M., Shiota, )., Kasahata, N., &
Hirayama, K. (1997). Pure topographic disorien tation due to right retrosplenial lesion. Neurology, 49(2) , 464- 469.
Tan, R. H., Wong, S., Hodges,). R., Halliday, G. M., &
Hornberger, M. (2013). Retrosplenial cortex (BA 29) volumes in behavioral variant frontotemporal dementia and alzheimer's disease. Dementia and Geriatric Cognitive Disorders, 35(3- 4) , 177-182. h ttp!/dx.doi.org/10.1159/000346392.
Tan, R. H., Wong, S., Kril, ). J., Piguet, 0 ., Hornberger, M., Hodges,). R., et al. (2014) . Beyond the temporal pole: limbic memory circuit in the semantic variant of primary progressive aphasia. Brain, 137(Pt 7), 2065- 2076. ht tp ://dx.doi.org/10.1093/ brain/ awu118.
Van n, S. D., Aggleton, J. P., & Maguire, E. A. (2009). What does the retrosplenial cortex do? Nature Reviews Neuroscience, 10(11), 792- 802. h ttp//dx.doi.orgl10.1038/nm2733.
Waterlander, W. E., Scarpa, M., Len tz, D., & Steenhuis, I. H. (2011). The virtual supermarket: an innovat ive research tool to study

41
94
consumer food purchasing behaviour. BMC Public Health, 11. http:/ /dx.doi.org/10.1186/1471-2458-11-589, 589- 2458-11-589.
Weniger, G., Ruhleder, M., Lange, C., Wolf, S., & lrle, E. (2011). Egocentric and allocentric memory as assessed by virtual reality in individuals with amnestic mild cognitive impairment. Neuropsychologia, 49(3), 518- 527. h ttp:// dx.doi.o rg/10 .1016/j . neuro psycho logia.201 0.12.031.
Yew, B., Alladi, S., Shailaja, M., Hodges,}. R., & Hornberger, M. (2013) . Lost and forgotten? orientat ion versus memory in
alzheimer's disease and frontotemporal dementia . journal of Alzheimer 's Disease, 33(2), 473-481. h ttp://dx .doi.org/10.3233/ )AD-2012-120769.
Zhang, Y., Brady, M., & Smith, S. (2001) . Segmentation of brain MR images through a h idden markov random field model and the expectation-maximization algorithm. IEEE Transactions on Medical Imaging, 20(1), 45- 57. h ttp:!/dx.doi.org/10.1109/ 42.906424.

42
Egocentric vs. allocentric spatial memory in behavioural variant
frontotemporal dementia and Alzheimer’s disease
Sicong Tu1,2,3, Hugo J. Spiers4, John R. Hodges1,2,3, Olivier Piguet1,2,3, Michael Hornberger2,3,5
1 Neuroscience Research Australia, Randwick, Sydney, Australia.
2 Australian Research Council Centre of Excellence in Cognition and its Disorders, Sydney,
Australia.
3 School of Medical Sciences, University of New South Wales, Sydney, Australia.
4 Institute of Behavioural Neuroscience, Department of Experimental Psychology, University
College London, London, United Kingdom.
5Norwich Medical School, University of East Anglia, Norwich, United Kingdom.
Running Title: Spatial Representation in bvFTD/AD
Corresponding author:
Prof. Michael Hornberger
Norwich Medical School, University of East Anglia, Norwich, NR47UQ, United Kingdom
Tel: +44 (0)1603 597139

43
Abstract
Background: Diagnosis of behavioural variant frontotemporal dementia (bvFTD) can be
challenging, in particular when patients present with significant memory problems, which can
increase the chance of a misdiagnosis of Alzheimer’s disease (AD). Growing evidence suggests
spatial orientation is a reliable cognitive marker able to differentiate these two clinical
syndromes.
Objective: Assess the integrity of egocentric and allocentric heading orientation and memory in
bvFTD and AD, and their clinical implications.
Method: A cohort of 22 patient with dementia (11 bvFTD; 11 AD) and 14 healthy controls were
assessed on the virtual supermarket task of spatial orientation and a battery of standardized
neuropsychological measures of visual and verbal memory performance.
Results: Judgements of egocentric and allocentric heading direction were differentially impaired
in bvFTD and AD, with AD performing significantly worse on egocentric heading judgements
than bvFTD. Both patient cohorts, however, showed similar degree of impaired allocentric
spatial representation, and associated hippocampal pathology.
Conclusions: The findings suggest egocentric heading judgements offer a more sensitive
discriminant of bvFTD and AD than allocentric map-based measures of spatial memory.
Keywords: orientation, hippocampus, frontotemporal dementia, Alzheimer’s disease

44
Introduction
Alzheimer’s disease (AD) and the behavioural variant of frontotemporal dementia (bvFTD) are
two neurodegenerative dementia conditions with distinct and overlapping cognitive and
pathological features [1,2]. For a long time differences in memory performance has been
proposed to be a key clinical feature in the early differential diagnosis of bvFTD and AD, with
AD patients expressing greater memory deficits than bvFTD patients [3,4]. Growing evidence,
however, shows that bvFTD patients exhibit considerable variability in memory function,
resulting in a misdiagnosis of AD [5,6]. This may be a result of significant hippocampal
pathology, which has also been reported in bvFTD [7,8]. Nevertheless, posterior regions of the
brain, in particular, the posterior cingulate and parietal lobe remain relatively intact in sporadic
bvFTD, but are affected in the early stages of AD [9,10]. This has important implications for the
neural basis of spatial memory and the development of neuro-anatomically targeted cognitive
tools to aid in differential diagnosis of dementia syndromes.
Spatial disorientation is a prominent feature in the early stages of Alzheimer’s disease
(AD), but tends to be preserved in frontotemporal dementia (FTD) [6,11-15]. Increasing
evidence shows that spatial orientation is a sensitive diagnostic discriminant of AD and bvFTD
[11,12], while episodic memory performance is less helpful [5,6,16,17]. While the diagnostic
accuracy of AD and bvFTD can be significantly improved when considering spatial orientation
in conjunction with established memory measures [11,12], orientation is rarely assessed
objectively as part of routine cognitive screenings in dementia. An outstanding issue is how can
orientation best be used to differentially diagnose AD and bvFTD in a generalized clinical
setting?

45
Spatial orientation requires the representation of the spatial relationships among separate
entities in the world. Information can be referenced with respect to the body (egocentric
representations) or with respect to invariant landmarks in the environment (allocentric
representations) [18]. The posterior parietal cortex has been implicated in coding egocentric
information and a circuit involving the hippocampus and parahippocampal structures has been
argued to support allocentric representations, with the retrosplenial cortex thought to mediate
both frameworks of spatial information processing [18,19]. In particular the hippocampus has
been argued to form an internal map of space to support long-term memory for space and events
occurring in them [18]. While spatial navigation has been studied in AD and its prodromal stage
[20], with a variety of tasks adapted for patient testing [21-23], bvFTD patients’ spatial abilities
have rarely been examined [11]. This is likely due to the lack of observed spatial memory
impairment in bvFTD reported in the clinic. When considered in the context of differential
diagnosis with AD, however, this feature becomes highly relevant.
Assessments of spatial navigation tapping into both egocentric and allocentric processing
have been extensively studied in AD, with tasks drawing upon newly acquired [21,24] or
existing spatial information [23]. Patients with an AD diagnosis have been consistently reported
impaired on spatial orientation task [20], with a selective deficit in the translation of information
between egocentric and allocentric information [23], resulting in an impaired ability to make
accurate judgement of heading direction [11]. This finding corroborates with reports of
significant topographical disorientation in community dwelling AD patients, placing a significant
level of burden on caregivers [13]. In bvFTD, however, this does not appear to be the case
[11,14,21,25]. Previous studies of spatial navigation in FTD involved the semantic language
variant, who performed at a comparable level to healthy controls on tasks requiring egocentric

46
and allocentric based spatial memory [14,21,25]. Of greater clinical interest is comparative
spatial processing ability between AD and bvFTD, given the difficulty of diagnosis in the earliest
stages when behavioural symptoms remain mild.
To our knowledge, previous work by our group provides the only characterisation of
egocentric spatial processing in bvFTD patients [11]. Whether bvFTD patients also show
hippocampal-dependent allocentric spatial memory deficits remains unknown. To address this
question we used an ecological virtual supermarket environment, which does not require prior
learning or training, to assess judgements of egocentric heading direction in AD and bvFTD.
Spatial orientation performance using the virtual supermarket task has been shown to hold the
same level of sensitivity as episodic memory in differentiating between AD and bvFTD patients,
with significantly increased diagnostic accuracy when the two variables were considered in
combination [11]. In the current study, the virtual supermarket task was employed with a novel
spatial layout component to compare egocentric and allocentric spatial processing in AD and
bvFTD. Structural neuroimaging was carried out in combination to assess the impact of
hippocampal pathology on spatial memory in these two patient cohorts.
Method
Participants
Twenty two dementia patients (11 AD; 11 bvFTD) and 14 age-matched healthy controls were
recruited from the Sydney frontotemporal dementia research group (FRONTIER) database. All
participants were assessed at the FRONTIER clinic located at Neuroscience Research Australia,
Sydney. Study approval was provided by the South Eastern Sydney Local Health District and the
University of New South Wales human research ethics committees. All participants provided

47
signed consent for neuropsychological assessment and neuroimaging prior to testing. All
dementia patients fulfilled international consensus criteria for AD [26] and bvFTD [2]. Clinical
diagnoses were established by consensus among senior neurologist, occupational therapist and
neuropsychologist, based on a clinical interview, comprehensive neuropsychological assessment,
and evidence of brain atrophy on structural neuroimaging. All bvFTD patients showed disease
progression as well as atrophy on scans to exclude any phenocopy cases [27]. Participant
demographics and clinical characteristics are provided in Table 1.
Briefly, AD patients presented with predominantly episodic memory impairment with
preserved social behaviour. BvFTD patients demonstrated changes in social functioning, loss of
insight, disinhibition and increased apathy. Exclusion criteria for all participants included prior
history of mental illness, head injury, movement disorders, alcohol and drug abuse, limited
English proficiency, and, for controls, presence of abnormality on MRI. Participants were
administered a battery of cognitive tests to assess overall cognitive function, verbal and visual
memory, and working memory. This assessment included: Addenbrooke’s Cognitive
Examination-Revised (ACE-R), Rey Auditory Verbal Learning Test (RAVLT), Rey Complex
Figure Test (RCFT), and Digit Span. For a brief description of cognitive tasks see
Supplementary Table 1.
Virtual supermarket task
Spatial orientation was assessed using an ecological virtual supermarket task consisting of 14
video trials [11], based on a small-scale supermarket environment [28] (Fig. 1A). Videos were
presented from a first person perspective and involved travelling to set locations within the
supermarket while making a series of 90 degree turns. All trials began at the same location, but
followed different routes to reach their respective end locations. Video trials were standardized

48
for length and number of turns, such that half of the trials lasted 20s with 3 turns, while the other
half lasted 40s with 5 turns. All video trials were presented in a randomized order across
participants. At the end of each trial, participants were prompted to make a judgement of heading
direction from the new location relative to the starting location based on egocentric body turns
(Fig. 1A). Critically, correct judgements of direction cannot be made from simply viewing the
image of the new location at the end of each video trial. Building upon a previous study using
this task [11], participants are then presented with a spatial map of the supermarket environment,
with starting location marked, and asked to indicate current location and heading direction. This
requires participants to translate their current view to a map coordinate and orientation, thus
drawing upon allocentric spatial representations (Fig. 1B).

49
Figure 1. Example of egocentric and allocentric components of the virtual supermarket task. (a) Participants view videos travelling to
a new location within the supermarket and asked for heading direction to starting location. (b) Participants are presented with a spatial
map and asked to mark current location and heading direction.

50
Prior to testing, participants were explicitly instructed that they would be viewing a
number of short videos that involved moving to new locations within the supermarket and were
required to make a judgement of heading direction and location relative to the original starting
location. A single practice video trial (10 s, 2 turns) was provided at the start of testing to
introduce participants to the virtual supermarket environment and make clear task instructions
were understood. No further training is provided. Consequently, the formulation of a working
internal spatial representation of the immediate environment for making judgements of direction
and location [29] reflects incidental acquisition present in everyday spatial navigation. Correct
judgements of heading direction required participants to accurately distinguish whether the
relative starting location was in front/behind and to the left/right of their current location using
an egocentric, followed by an allocentric framework. For spatial location, locations marked
within a 4 mm radius of the correct location were considered to be correct. In addition, the
Euclidean distance between the participants’ response and the correct location was measured, as
was the mean Euclidean distance between all marked locations and the centre of the spatial map.
The mean Euclidean distance to the centre was used to examine any bias in the responses, such
as would occur if responses were clustered in the middle or edges of the map.
Statistical Analyses
Differences in participant group demographics, performance on standard cognitive tests, and
spatial performance on the experimental task, were assessed using analysis of variance
(ANOVA) and two-tailed post hoc multiple comparisons between groups in SPSS 21.0 (IBM
Corp., Armonk, NY). In all analyses, p values < .05 were considered statistically significant.
Imaging Acquisition

51
Whole-brain structural T1 images were acquired for all participants using a 3T Philips MRI
scanner with standard quadrature head coil (eight channels). Structural T1 scans were acquired as
follows: coronal orientation, matrix 256 x 256, 200 slices, 1mm isotropic, TE/TR = 2.5/5.4 ms,
flip angle α = 8°. Prior to analyses, all participant scans were visually inspected for significant
head movements and artefacts, and excluded from imaging analyses. All scans were examined
by a radiologist for structural abnormalities.
Imaging Analyses
Voxel-based morphometry (VBM) was conducted on whole-brain T1-weighted scans, using the
VBM toolbox in FMRIB’s Software Library (FSL; http://www.fmrib.ox.ac.uk/fsl/). First, the
brain was extracted from each scan using FSL’s BET algorithm with a fractional intensity
threshold of 0.22 [30]. Each scan was visually checked following brain extraction to ensure no
brain matter was excluded, and no non-brain matter was included. A study specific template of
grey matter was generated from 11 scans for each participant cohort. An equivalent number of
scans from each cohort were used to create the template, avoiding potential bias towards any
single group’s topography during registration. Template scans were then registered to the
Montreal Neurological Institute (MNI) standard brain (MNI 152), resulting in a study-specific
grey matter template at 2 mm3 resolution in MNI standard space. Simultaneously, participant
brain-extracted scans were segmented into CSF, grey matter and white matter using FMRIB's
Automatic Segmentation Tool (FAST) [31]. The FAST algorithm corrected scans for spatial
intensity variations such as bias field or radio-frequency inhomogeneity, resulting in partial
volume maps. Grey matter partial volume maps were then non-linearly registered to the study-
specific template. After normalization and modulation, grey matter maps were smoothed using
an isotropic Gaussian kernel (sigma = 3 mm).

52
Region of interest analyses of the hippocampus and retrosplenial cortex (Brodmann areas
29/30) were carried out between each patient group and controls, as well as between AD and
bvFTD cohorts. These two regions were examined based on a priori evidence of spatial memory
deficits linked to these regions [32]. Grey matter volumes were extracted with reference to the
Harvard-Oxford subcortical structural atlas and correlated with spatial orientation performance.
Scans were spatially normalized to a common group template during processing and corrected
for total intracranial volume.
Results
Demographics
AD and bvFTD patient cohorts were well matched on demographic variables, including age,
education, and disease duration and functional severity (Table 1; all p values > 0.2). The healthy
control cohort was matched for age (p values > 0.7), but demonstrated higher mean years of
education compared to bvFTD patients (p value = 0.013). ANOVA of participant groups’
performance across standard cognitive tests revealed significant group differences for all
components (all p values < 0.03). Between patient groups, bvFTD showed a better cognitive
profile on the ACE-R compared to AD (all p values < 0.01), and verbal memory recall on the
RAVLT (all p values < 0.02). The two patient groups, however, did not differ on visual memory
and working memory as indicated by the RCFT (all p values > 0.1) and digit span (all p values >
0.6), respectively. Compared to controls, AD performed significantly worse on all cognitive test
components (all p values < 0.05). BvFTD showed a similar pattern of impairment. Performance
was, however, not significantly worse for orientation on the ACE-R and delayed
recall/recognition on the RAVLT, compared to controls.
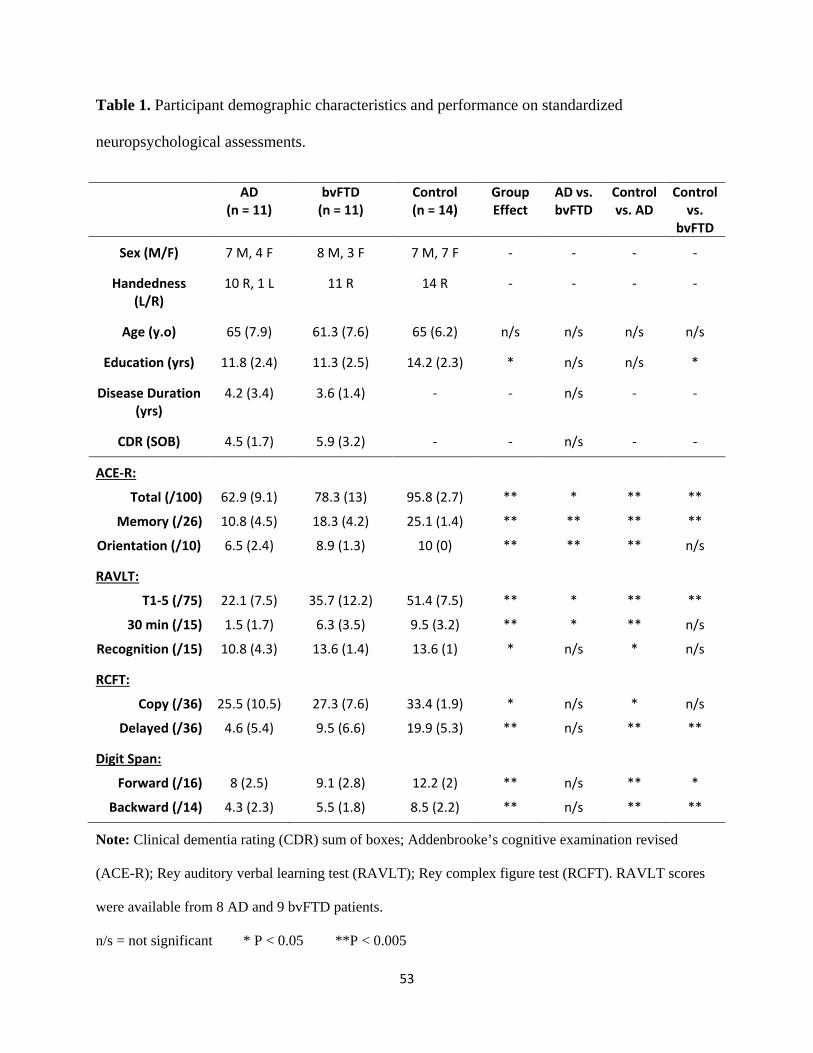
53
Table 1. Participant demographic characteristics and performance on standardized
neuropsychological assessments.
AD (n = 11)
bvFTD (n = 11)
Control (n = 14)
Group Effect
AD vs. bvFTD
Control vs. AD
Control vs.
bvFTD
Sex (M/F) 7 M, 4 F 8 M, 3 F 7 M, 7 F - - - -
Handedness (L/R)
10 R, 1 L 11 R 14 R - - - -
Age (y.o) 65 (7.9) 61.3 (7.6) 65 (6.2) n/s n/s n/s n/s
Education (yrs) 11.8 (2.4) 11.3 (2.5) 14.2 (2.3) * n/s n/s *
Disease Duration (yrs)
4.2 (3.4) 3.6 (1.4) - - n/s - -
CDR (SOB) 4.5 (1.7) 5.9 (3.2) - - n/s - -
ACE-R:
Total (/100)
Memory (/26)
Orientation (/10)
62.9 (9.1)
10.8 (4.5)
6.5 (2.4)
78.3 (13)
18.3 (4.2)
8.9 (1.3)
95.8 (2.7)
25.1 (1.4)
10 (0)
**
**
**
*
**
**
**
**
**
**
**
n/s
RAVLT:
T1-5 (/75)
30 min (/15)
Recognition (/15)
22.1 (7.5)
1.5 (1.7)
10.8 (4.3)
35.7 (12.2)
6.3 (3.5)
13.6 (1.4)
51.4 (7.5)
9.5 (3.2)
13.6 (1)
**
**
*
*
*
n/s
**
**
*
**
n/s
n/s
RCFT:
Copy (/36)
Delayed (/36)
25.5 (10.5)
4.6 (5.4)
27.3 (7.6)
9.5 (6.6)
33.4 (1.9)
19.9 (5.3)
*
**
n/s
n/s
*
**
n/s
**
Digit Span:
Forward (/16)
Backward (/14)
8 (2.5)
4.3 (2.3)
9.1 (2.8)
5.5 (1.8)
12.2 (2)
8.5 (2.2)
**
**
n/s
n/s
**
**
*
**
Note: Clinical dementia rating (CDR) sum of boxes; Addenbrooke’s cognitive examination revised
(ACE-R); Rey auditory verbal learning test (RAVLT); Rey complex figure test (RCFT). RAVLT scores
were available from 8 AD and 9 bvFTD patients.
n/s = not significant * P < 0.05 **P < 0.005

54
Spatial Orientation Performance
Heading orientation was scored for a correct judgement of direction to the starting location at the
end of each trial using egocentric and allocentric frameworks on the first-person and spatial map
components of the virtual supermarket task, respectively (Fig. 2). ANOVA indicated significant
group differences across both conditions (all p values < 0.001). Compared to controls, AD
performed significantly worse on both conditions (all p values < 0.001), however, bvFTD were
impaired only in the allocentric condition (p value = 0.01). Within patient groups, AD performed
significantly worse than bvFTD in both conditions of heading orientation (all p values < 0.04).
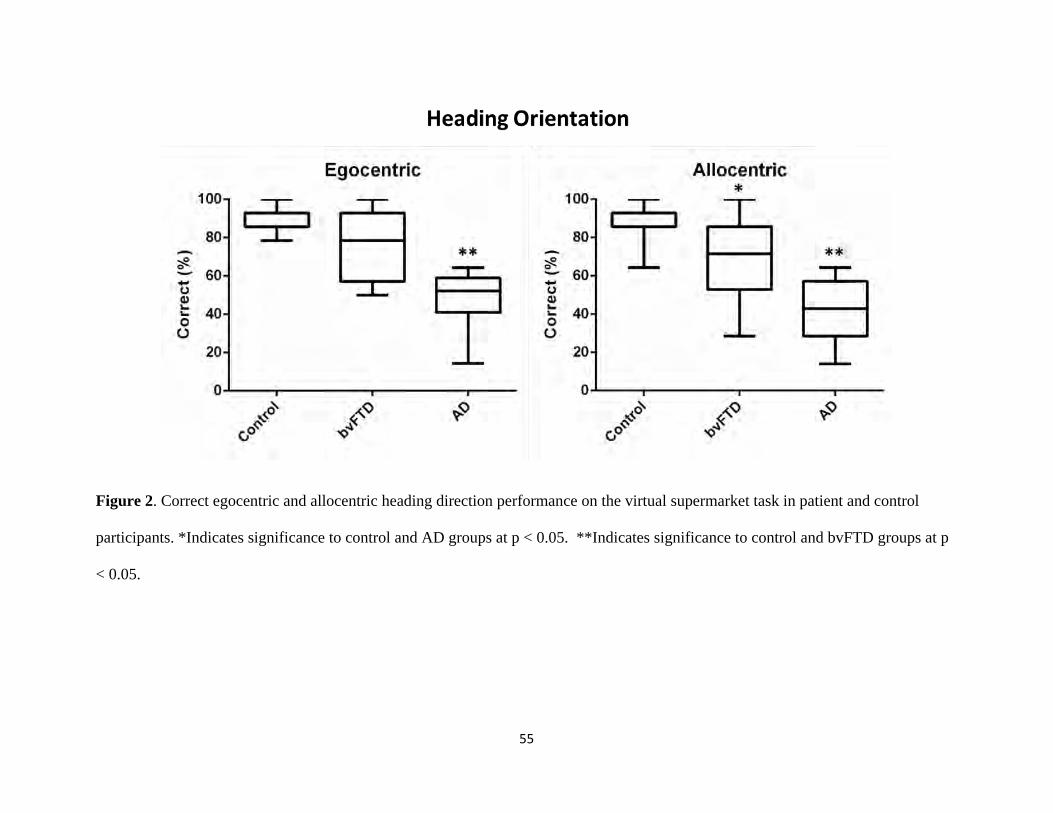
55
Figure 2. Correct egocentric and allocentric heading direction performance on the virtual supermarket task in patient and control
participants. *Indicates significance to control and AD groups at p < 0.05. **Indicates significance to control and bvFTD groups at p
< 0.05.

56
Spatial representation was scored for correct indication of location and distance from
correct location for each trial on the supermarket layout (Fig. 3). ANOVA indicated significant
group differences across both conditions (all p values < 0.001). Patient groups were impaired in
locating the correct location on each trial, and distance from the correct location was
significantly greater compared to controls (all p values < 0.001). While there was no significant
difference in performance between patients groups in regard to forming an accurate spatial
representation of the supermarket layout, the measure of distal relationships between trial
locations found AD were significantly more impaired than bvFTD and control (Fig. 4; all p
values < 0.01). Furthermore, this dissociation is reflected through qualitative differences in the
pattern of responses on the spatial map component (Supplementary Fig. 1). Specifically, AD
patients demonstrated little ability in integrating navigational information from an egocentric
framework to form an allocentric spatial representation of location and distance. When asked for
a location at the end of each trial, patients often mention they are “at the back of the
supermarket” resulting in a clustered spatial representation often located near the outer edges of
the spatial map. In contrast, while AD and bvFTD performance did not significantly differ in
terms of accuracy, bvFTD patient’s demonstrated evidence of ability to incorporate egocentric
information, resulting in a spatial representation that was evenly distributed similar to that of
controls.
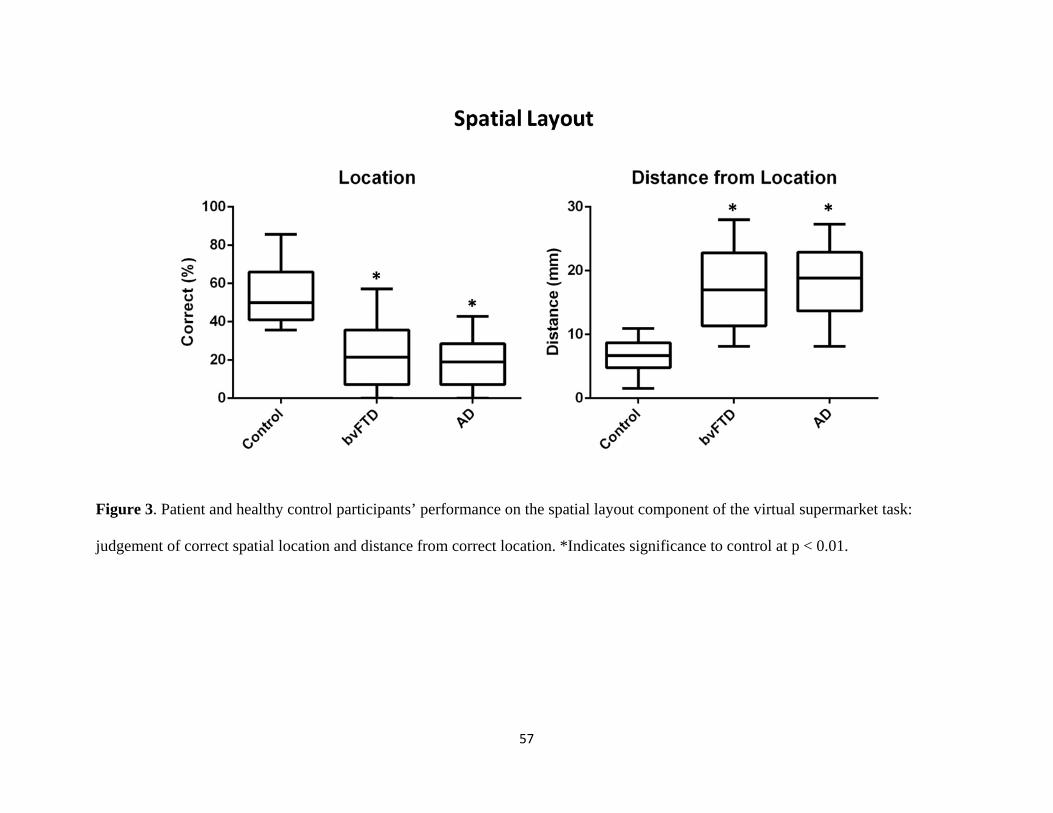
57
Figure 3. Patient and healthy control participants’ performance on the spatial layout component of the virtual supermarket task:
judgement of correct spatial location and distance from correct location. *Indicates significance to control at p < 0.01.
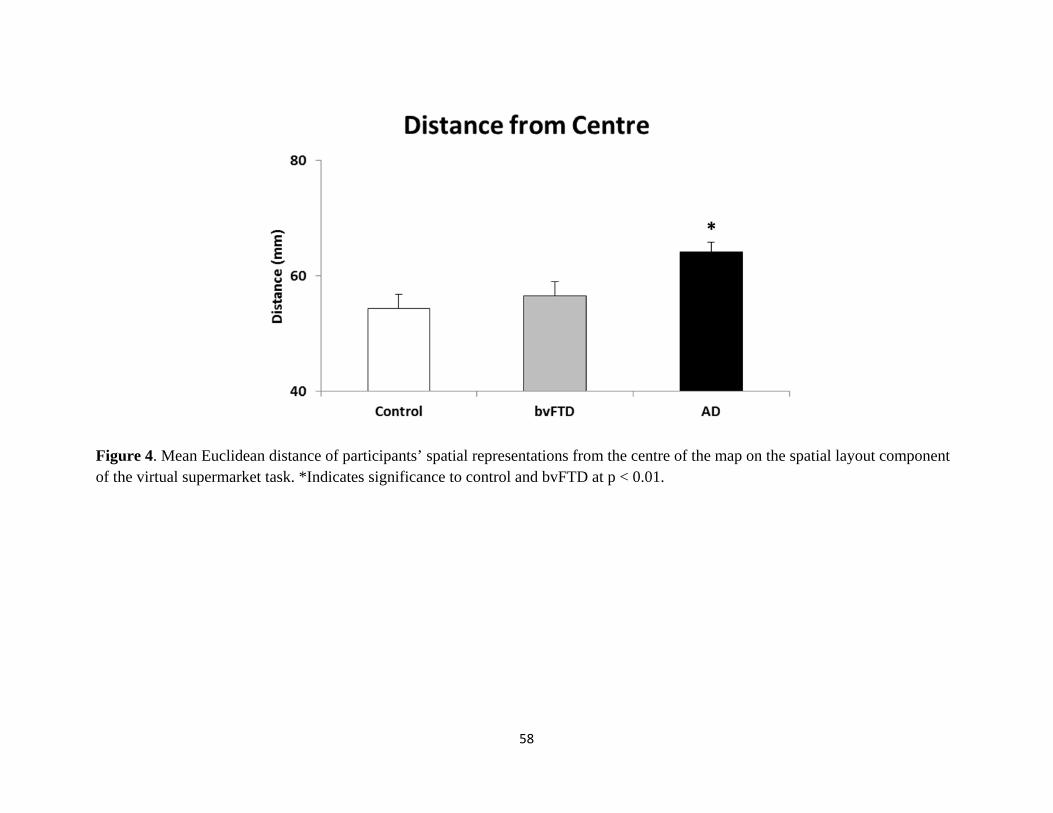
58
Figure 4. Mean Euclidean distance of participants’ spatial representations from the centre of the map on the spatial layout component of the virtual supermarket task. *Indicates significance to control and bvFTD at p < 0.01.

59
Hippocampal and Retrosplenial Volume
Mean left and right whole hippocampal volume was compared across participant groups (Fig. 5).
Bilateral hippocampal volume was significantly reduced compared to control in bvFTD and AD
patient groups (all p values < 0.05). Hippocampal volume did not, however, differ between
patient groups (all p values > 0.1). Volume of the retrosplenial cortex (Brodmann Areas 29, 30)
was also compared between control, bvFTD, and AD (Supplementary Fig. 2). No significant
differences were found between participant groups (all p values > 0.4). Hippocampal and
retrosplenial volume did not show a significant correlation with egocentric and allocentric
orientation performance in each participant group (all p values > 0.1).
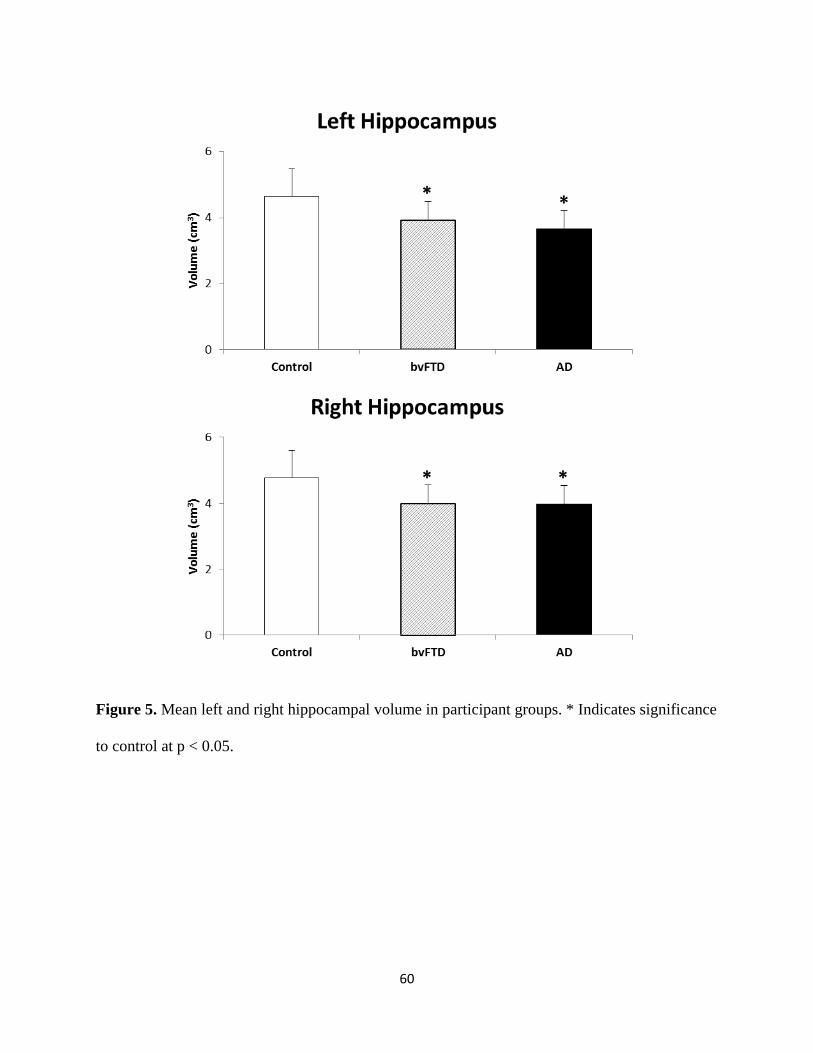
60
Figure 5. Mean left and right hippocampal volume in participant groups. * Indicates significance
to control at p < 0.05.

61
Discussion
Using a novel spatial memory task set in a virtual reality supermarket we reveal differences in
the spatial orientation and memory performance of bvFTD and AD patients. While AD patients
were impaired at both judging the egocentric direction back to a starting location and estimating
distances and locations on a map, bvFTD patients were only impaired on estimating distances
and locations on a map. This is important because, while FTD and AD have been compared
before on spatial tasks [21], prior studies have not directly compared bvFTD and AD [21], their
differential performance on egocentric and allocentric spatial processing, or the association with
underlying hippocampal and retrosplenial neural substrates of spatial memory. Notably, it is the
differential discrimination of bvFTD and AD that poses the greatest challenge during clinical
diagnosis of dementia. Structural neuroimaging uncovered a similar degree of hippocampal
atrophy in both patient cohorts, compared to controls. Given the strong links between the
hippocampus and allocentric memory [18], and past evidence of AD patients spatial impairments
[11,13], it is plausible that the hippocampal damage in both patient groups relates to the impaired
allocentric map-based memory observed. By contrast, the spared performance of the bvFTD
patients in the context of their extensive hippocampal damage suggests that other brain regions
may mediate the ability to estimate egocentric heading direction.
Recent years has seen an influx of advanced virtual paradigms being used to address
spatial navigation deficits in AD [21,22,24,33-35], as well as pre-symptomatic carriers with a
genetic mutation linked to the disease [36]. A concern, however, is the extensive training
necessary to form a working representation of the environment, to allow subsequent testing, in a
patient population characterised by memory impairment. In this regard measures of heading
direction, relying on existing environmental knowledge [13,37] or a familiar construct (i.e. novel

62
supermarket), provides a purer measure of disorientation with minimal overlap of existing
memory impairment, as we have previously shown using a variation of the current experimental
task [11]. This is of particular interest in differentiating different patient populations with
underlying hippocampal pathology, such as bvFTD and AD, as our current findings demonstrate
similar levels of impaired ability to form an accurate spatial representation of the environment
but a clear preservation of heading orientation performance in bvFTD.
Spatial representation of the virtual supermarket was acquired incidentally across trials in
the current experimental task. Without explicit training, the accuracy of identifying correct trial
locations was expected to be low. Nevertheless, average bvFTD and AD performance in
identifying the correct location and erroneous distance from correct location was impaired by
more than twofold, compared to control participants. This is consistent with previous spatial
navigation studies that have reported allocentric hippocampal dependent spatial memory is
impaired in AD [20,38]. The hippocampus is suggested to be of particular importance in accurate
representation of distance between environmental objects [39,40]. Our findings, reveal bvFTD
have a similar level of impairment to allocentric spatial memory as AD patients. While bvFTD is
characterised by frontal and anterior temporal lobe atrophy, recent findings have implicated
significant hippocampal pathology during disease progression to the same degree as AD [7,8].
Similarly, in the current study, bvFTD and AD showed the same level of bilateral hippocampal
atrophy, which may account for the similar degree of inaccuracy in judging location and distance
for spatial layout.
While bvFTD and AD patient groups showed the same level of accuracy in forming a
correct spatial representation of the virtual supermarket, there were inherent quantitative and
qualitative differences in estimating distance between the two patient groups that may hold

63
clinical value. Across trials, bvFTD patients showed some semblance of being able to translate
egocentric information obtained from viewing the video trials to an allocentric representation of
spatial location. Despite being inaccurate, bvFTD patients were able to indicate a more logical
position on the provided supermarket layout maps, compared to AD who would often indicate
they had travelled to the same location across trials but were facing a different direction. This
inability to translate spatial information from different frames of reference has been noted in
previous studies in AD [13,21] and believed to represent topographical impairment resulting
from dysfunction in the retrosplenial region of the posterior cingulate [19,41,42].
Our finding that bvFTD patients’ show persevered ability to estimate heading direction to
a point of origin has implications for the brain regions supporting path integration. Path
integration is the ability to use self-motion cues to estimate the distance and direction to a point
of origin, which is required in our heading orientation test. There has been disagreement in past
neuropsychological studies about the extent to which the hippocampus is required for path
integration [43-45] . Our data provide further evidence that, in the context of significant
hippocampal damage and impaired allocentric spatial memory, the ability to estimate the heading
orientation to a starting location can be relatively spared. Thus, in AD it may be damage to
subcortical circuits and possibly the retrosplenial cortex that disrupts this ability. More research
with larger patient cohorts will be useful to determine this.
In the current study we limited structural neuroimaging to hippocampal and retrosplenial
volume given the focus on egocentric and allocentric spatial representation in bvFTD and AD.
The volume of the retrosplenial cortex is, however, small and prone to partial volume effects
reducing the power to detect structural change. In combination with our patient sample size, this
may have resulted in the absence of a significant correlation between neural structures and

64
behavioural performance, despite evidence of a functional dissociation between patient groups,
on the virtual supermarket task. Future studies with a larger sample size will allow a more
comprehensive approach for investigating underlying neural mechanisms of spatial memory
processing in dementia. FTD and AD, in particular, provide a unique opportunity to further
elucidate the functional interaction of regions beyond the medial temporal lobe in topographical
memory processes, given the similar degree of hippocampal pathology present across these
clinical syndromes.
In conclusion, allocentric spatial representations are impaired in both bvFTD and AD,
which share hippocampal pathology during the course of disease. In contrast, heading orientation
is preserved in bvFTD and should be the targeted by clinical tasks of spatial memory
performance to aid differential diagnosis.

65
Acknowledgements
This work was supported by funding to Forefront, a collaborative research group dedicated to the
study of frontotemporal dementia and motor neurone disease, from the National Health and
Medical research Council (NHMRC) of Australia program grant (#1037746), the Australian
Research Council (ARC) Centre of Excellence in Cognition and its Disorders Memory Node
(#CE110001021) and an ARC Discovery Project grant (DP1093279). ST is supported by
Alzheimer’s Australia Dementia Research Foundation and NHMRC of Australia awards. OP is
supported by a NHMRC Career Development Fellowship (APP1022684). MH is supported by
Alzheimer Research UK and the Isaac Newton Trust. These funding sources had no involvement
in the study design, collection, analysis and interpretation of data, writing the manuscript, and in
the decision to submit the manuscript for publication. The authors report no conflict of interest.
We are grateful to the research participants and carers involved with ForeFront research studies.

66
References
[1] Chare L, Hodges JR, Leyton CE, McGinley C, Tan RH, Kril JJ, Halliday GM (2014) New
criteria for frontotemporal dementia syndromes: clinical and pathological diagnostic
implications. J Neurol Neurosurg Psychiatry 85, 865-870.
[2] Rascovsky K, Hodges JR, Knopman D, Mendez MF, Kramer JH, Neuhaus J, van Swieten JC,
Seelaar H, Dopper EG, Onyike CU, Hillis AE, Josephs KA, Boeve BF, Kertesz A, Seeley WW,
Rankin KP, Johnson JK, Gorno-Tempini ML, Rosen H, Prioleau-Latham CE, Lee A, Kipps CM,
Lillo P, Piguet O, Rohrer JD, Rossor MN, Warren JD, Fox NC, Galasko D, Salmon DP, Black
SE, Mesulam M, Weintraub S, Dickerson BC, Diehl-Schmid J, Pasquier F, Deramecourt V,
Lebert F, Pijnenburg Y, Chow TW, Manes F, Grafman J, Cappa SF, Freedman M, Grossman M,
Miller BL (2011) Sensitivity of revised diagnostic criteria for the behavioural variant of
frontotemporal dementia. Brain 134, 2456-2477.
[3] Perry RJ, Hodges JR (2000) Differentiating frontal and temporal variant frontotemporal
dementia from Alzheimer's disease. Neurology 54, 2277-2284.
[4] Glosser G, Gallo JL, Clark CM, Grossman M (2002) Memory encoding and retrieval in
frontotemporal dementia and Alzheimer's disease. Neuropsychology 16, 190-196.
[5] Bertoux M, de Souza LC, Corlier F, Lamari F, Bottlaender M, Dubois B, Sarazin M (2014)
Two distinct amnesic profiles in behavioral variant frontotemporal dementia. Biol Psychiatry 75,
582-588.

67
[6] Hornberger M, Piguet O, Graham AJ, Nestor PJ, Hodges JR (2010) How preserved is
episodic memory in behavioral variant frontotemporal dementia? Neurology 74, 472-479.
[7] de Souza LC, Chupin M, Bertoux M, Lehericy S, Dubois B, Lamari F, Le Ber I, Bottlaender
M, Colliot O, Sarazin M (2013) Is hippocampal volume a good marker to differentiate
Alzheimer's disease from frontotemporal dementia? J Alzheimers Dis 36, 57-66.
[8] Hornberger M, Wong S, Tan R, Irish M, Piguet O, Kril J, Hodges JR, Halliday G (2012) In
vivo and post-mortem memory circuit integrity in frontotemporal dementia and Alzheimer's
disease. Brain 135, 3015-3025.
[9] Bruner E, Jacobs HI (2013) Alzheimer's disease: the downside of a highly evolved parietal
lobe? J Alzheimers Dis 35, 227-240.
[10] Nestor PJ, Fryer TD, Ikeda M, Hodges JR (2003) Retrosplenial cortex (BA 29/30)
hypometabolism in mild cognitive impairment (prodromal Alzheimer's disease). Eur J Neurosci
18, 2663-2667.
[11] Tu S, Wong S, Hodges JR, Irish M, Piguet O, Hornberger M (2015) Lost in spatial
translation - A novel tool to objectively assess spatial disorientation in Alzheimer's disease and
frontotemporal dementia. Cortex 67, 83-94.
[12] Yew B, Alladi S, Shailaja M, Hodges JR, Hornberger M (2013) Lost and forgotten?
Orientation versus memory in Alzheimer's disease and frontotemporal dementia. J Alzheimers
Dis 33, 473-481.

68
[13] Pai MC, Jacobs WJ (2004) Topographical disorientation in community-residing patients
with Alzheimer's disease. Int J Geriatr Psychiatry 19, 250-255.
[14] Pengas G, Patterson K, Arnold RJ, Bird CM, Burgess N, Nestor PJ (2010) Lost and found:
bespoke memory testing for Alzheimer's disease and semantic dementia. J Alzheimers Dis 21,
1347-1365.
[15] Pengas G, Hodges JR, Watson P, Nestor PJ (2010) Focal posterior cingulate atrophy in
incipient Alzheimer's disease. Neurobiol Aging 31, 25-33.
[16] Pennington C, Hodges JR, Hornberger M (2011) Neural correlates of episodic memory in
behavioral variant frontotemporal dementia. J Alzheimers Dis 24, 261-268.
[17] Irish M, Piguet O, Hodges JR, Hornberger M (2014) Common and unique gray matter
correlates of episodic memory dysfunction in frontotemporal dementia and Alzheimer's disease.
Hum Brain Mapp 35, 1422-1435.
[18] O'Keefe J, Nadel L (1978) The Hippocampus as a Cognitive Map, Oxford University Press.
[19] Vann SD, Aggleton JP, Maguire EA (2009) What does the retrosplenial cortex do? Nat Rev
Neurosci 10, 792-802.
[20] Serino S, Cipresso P, Morganti F, Riva G (2014) The role of egocentric and allocentric
abilities in Alzheimer's disease: a systematic review. Ageing Res Rev 16, 32-44.

69
[21] Bellassen V, Igloi K, de Souza LC, Dubois B, Rondi-Reig L (2012) Temporal order
memory assessed during spatiotemporal navigation as a behavioral cognitive marker for
differential Alzheimer's disease diagnosis. J Neurosci 32, 1942-1952.
[22] Laczo J, Vlcek K, Vyhnalek M, Vajnerova O, Ort M, Holmerova I, Tolar M, Andel R, Bojar
M, Hort J (2009) Spatial navigation testing discriminates two types of amnestic mild cognitive
impairment. Behav Brain Res 202, 252-259.
[23] Pai MC, Yang YC (2013) Impaired translation of spatial representation in young onset
Alzheimer's disease patients. Curr Alzheimer Res 10, 95-103.
[24] Laczo J, Andel R, Vyhnalek M, Vlcek K, Magerova H, Varjassyova A, Nedelska Z, Gazova
I, Bojar M, Sheardova K, Hort J (2012) From Morris Water Maze to computer tests in the
prediction of Alzheimer's disease. Neurodegener Dis 10, 153-157.
[25] Luzzi S, Cafazzo V, Damora A, Fabi K, Fringuelli FM, Ascoli G, Silvestrini M, Provinciali
L, Reverberi C (2015) The neural correlates of road sign knowledge and route learning in
semantic dementia and Alzheimer's disease. J Neurol Neurosurg Psychiatry 86, 595-602.
[26] McKhann GM, Knopman DS, Chertkow H, Hyman BT, Jack CR,Jr, Kawas CH, Klunk WE,
Koroshetz WJ, Manly JJ, Mayeux R, Mohs RC, Morris JC, Rossor MN, Scheltens P, Carrillo
MC, Thies B, Weintraub S, Phelps CH (2011) The diagnosis of dementia due to Alzheimer's
disease: recommendations from the National Institute on Aging-Alzheimer's Association
workgroups on diagnostic guidelines for Alzheimer's disease. Alzheimers Dement 7, 263-269.

70
[27] Kipps CM, Hodges JR, Hornberger M (2010) Nonprogressive behavioural frontotemporal
dementia: recent developments and clinical implications of the 'bvFTD phenocopy syndrome'.
Curr Opin Neurol 23, 628-632.
[28] Waterlander WE, Scarpa M, Lentz D, Steenhuis IH (2011) The virtual supermarket: an
innovative research tool to study consumer food purchasing behaviour. BMC Public Health 11,
589-2458-11-589.
[29] Land MF (2014) Do we have an internal model of the outside world? Philosophical
Transactions of the Royal Society B: Biological Sciences 369.
[30] Smith SM (2002) Fast robust automated brain extraction. Hum Brain Mapp 17, 143-155.
[31] Zhang Y, Brady M, Smith S (2001) Segmentation of brain MR images through a hidden
Markov random field model and the expectation-maximization algorithm. IEEE Trans Med
Imaging 20, 45-57.
[32] Spiers HJ, Maguire EA (2007) The neuroscience of remote spatial memory: a tale of two
cities. Neuroscience 149, 7-27.
[33] Morganti F, Stefanini S, Riva G (2013) From allo- to egocentric spatial ability in early
Alzheimer's disease: a study with virtual reality spatial tasks. Cogn Neurosci 4, 171-180.
[34] Weniger G, Ruhleder M, Lange C, Wolf S, Irle E (2011) Egocentric and allocentric memory
as assessed by virtual reality in individuals with amnestic mild cognitive impairment.
Neuropsychologia 49, 518-527.

71
[35] Plancher G, Tirard A, Gyselinck V, Nicolas S, Piolino P (2012) Using virtual reality to
characterize episodic memory profiles in amnestic mild cognitive impairment and Alzheimer's
disease: influence of active and passive encoding. Neuropsychologia 50, 592-602.
[36] Kunz L, Schroder TN, Lee H, Montag C, Lachmann B, Sariyska R, Reuter M, Stirnberg R,
Stocker T, Messing-Floeter PC, Fell J, Doeller CF, Axmacher N (2015) Reduced grid-cell-like
representations in adults at genetic risk for Alzheimer's disease. Science 350, 430-433.
[37] Campbell JI, Hepner IJ, Miller LA (2014) The influence of age and sex on memory for a
familiar environment. J Environ Psychol 40, 1-8.
[38] Burgess N, Trinkler I, King J, Kennedy A, Cipolotti L (2006) Impaired allocentric spatial
memory underlying topographical disorientation. Rev Neurosci 17, 239-251.
[39] Morgan LK, Macevoy SP, Aguirre GK, Epstein RA (2011) Distances between real-world
locations are represented in the human hippocampus. The Journal of neuroscience : the official
journal of the Society for Neuroscience 31, 1238-45.
[40] Howard LR, Javadi AH, Yu Y, Mill RD, Morrison LC, Knight R, Loftus MM, Staskute L,
Spiers HJ (2014) The hippocampus and entorhinal cortex encode the path and Euclidean
distances to goals during navigation. Curr Biol 24, 1331-1340.
[41] Ino T, Doi T, Hirose S, Kimura T, Ito J, Fukuyama H (2007) Directional disorientation
following left retrosplenial hemorrhage: a case report with fMRI studies. Cortex 43, 248-254.
[42] Takahashi N, Kawamura M, Shiota J, Kasahata N, Hirayama K (1997) Pure topographic
disorientation due to right retrosplenial lesion. Neurology 49, 464-469.

72
[43] Worsley CL, Recce M, Spiers HJ, Marley J, Polkey CE, Morris RG (2001) Path integration
following temporal lobectomy in humans. Neuropsychologia 39, 452-464.
[44] Philbeck JW, Behrmann M, Levy L, Potolicchio SJ, Caputy AJ (2004) Path integration
deficits during linear locomotion after human medial temporal lobectomy. J Cogn Neurosci 16,
510-520.
[45] Kim S, Sapiurka M, Clark RE, Squire LR (2013) Contrasting effects on path integration
after hippocampal damage in humans and rats. Proc Natl Acad Sci U S A 110, 4732-4737.

73
Chapter 3 – Long-term Contextual Memory in Thalamic Stroke
Publication III – “Accelerated forgetting of contextual details due to focal medio-dorsal
thalamic lesion”

74
l!U'@~o~ D[lj] BEHAVIORAL NEUROSCIENCE
ORIGINAL RESEARCH ARTICLE published: 15 September 2014 do1 10 3389finbeh 2014 00320
Accelerated forgetting of contextual details due to focal medio-dorsal thalamic lesion Sicong Tu'-2, Laurie Miller2
•3
, Olivier Piguet1•2 and Michael Hornberger'.2-"*
1 Neuroscience Research Australia, S chool o f Medical Sciences , University o f Ne w South Wales , Sydney, NSW Australia 2 Australian Research Council Centre of Exce!!ence in Cognition and its Disorders, Sydney, NSW Austra!ia 1 Central Clinical Schoof, Neuropsychology Unit, Royal Prince Alfred Hospital, University of Sydney, Sydney, NSW, Australia 4 Department of Clinical Neurosciences, Universi!y of Cambridge, Cambridge, UK
Edited by: Regina M. Sui! ivan, Nathan Kline Institute and New York University School o f Medicine , USA
Reviewed by: Jane Pia iffy, Centre National de fa
Recherche Scientifique, France M ichael A. Burman, University of
New England, USA
*Correspondence.-Michaef Hornberger, Department of Clinical Neurosciences, University of Cambndge, Cambndge, C8 2 OSZ, UK
Effects of thalamic nuclei damage and related w hite matter tracts on memory performance are still debated, This is particularly evident for the medio-dorsal thalamus w hich has been less clear in predicting amnesia than anterior thalamus changes, The current study addresses this issue by as sessing 7 thalamic stroke patients w ith cons istent unilateral lesions focal to the left medio-dorsal nuclei for immediate and delayed memory performance on standard visual and verbal tests of anterograde memory, and over the long-term (> 24 h) on an object-location associative memory task, Thalamic patients showed selective impairment to delayed reca ll, but intact recognition memory, Pat ients also showed accelerated forgetting of contextual details afte r a 24 h delay, compared to controls, Importantly, the mammillothalamic tract was intact in all patients, w hich suggests a role for the medio-dorsal nuclei in recall and early consolidation memory processes,
e-mail: [email protected] Keywords: thalamus, anterograde memory, stroke, mammillothalamic tract, MRI
INTRODUCTION The thalamus is one of the major relay centers of the brain, and part of the limbic memory circuit, comprising hippocampus, fornix, mammillary bodies, mammillothalamic tract, thalamus, and cingulate cortex (Aggleton and Brown, 1999), Numerous direct and indirect projections connect mesial temporal lob e structures central for memory with the thalamus (Aggleton et aL, 2011; Carlesimo et al,, 20 11 ; Preston and Eich enbaum, 2013), A well-established connection is between the hippocampus and anterior thalamic nucleus (AT), via the mammillothalamic tract (MTI), Another connection exists between the perirhinal cortex and the medio-dorsal nucleus (MD) via the inferior thalamic peduncle (Saunders eta] ,, 2005 ), While amnesia is often observed in patients who have sustained a st roke involving AT or MD and the MTI (for a review see, Carlesimo et aL, 20 ll), the unique contribution of thalamic nuclei to amnesia is still not very well understood,
With a couple of exceptions (Van der Werf et aL, 2003; Perren et aL, 2005), reports of memory impairment in patients with focal thalamic lesions have been mostly confined to case studies (Kishiyama et al,, 2005; Edelstyn et aL, 2006; Carlesimo et aL, 2007; Cipolotti et a],, 2008; Hampstead and Koffler, 2009), This is partly attributed to the low patient incidence, and variability in the size and location of lesions (Le, uni- vs bi-lateral nuclei involved) , Recruitment of a homogen eous sample is therefore difficult given that many patients exhibit lesions that affect multiple th alamic nuclei, namely AT and MD (Van der Werf et al,, 2003; Perren et al,, 2005; Cipolotti et al,, 2008), Studies investigating the involvement of a single nucleus are rare and have been mostly confined to patients with damage to the AT, with a strong evidence of memory disturbance in these patients (Carlesimo et aL,
2011), The impact of focal MD damage on memory is, however, virtually unexplored at the group leveL
The MD is part of the "extended hippocampal system" (Aggleton and Brown, 1999), The contribution of MD damage to anterograde memory impairment following thalamic stroke is, however, less clear, A meta-analysis by Carlesimo and colleagues (20 ll) h ighlighted that only half the patients with MD lesions sh owed clinically diagnosed memory impairment, In contrast, all patients with lesions involving th e AT were affected, Importantly, the patients with MD lesions who were amnesic sh owed additional damage to the MTI, Lesion to th e MTI has been long known to be a strong predictor of anterograde amnesia (Vann, 2010; Carlesimo et aL, 20 11 ), While a strong relation exists between MTT damage and anterograde memory impairment (Cipolotti et al,, 2008), no study has directly contrasted MTI and thalamic integrity with regard to episodic memory performance, This is of particular relevance, given the debate surrounding the contribution of the MD to anterograde memory impairment,
Characterization of the memory deficits shown in p atients with focal thalamic lesions is also limited, Indeed, most studies have employed standard memory tests, which generally measure retrieval of info rmation post encoding after a short p eriod of time (30- 60 min), In these patients, performance across memory measures indicated a prevalent deficit to delayed memory retrieval components, which has been suggested to reflect long-term anterograde memory impairment with intact short-term mem ory acquisition and retrieval (Carlesimo et aL, 201 1) , Anterograde memory over th e long-term (Le,, > 24 h) has, however, been virtually unexplored, As such, the rate oflong-term forgetting for newly acquired information has yet to be examined in th alamic
Frontiers in Behavioral Neuroscience www.frontiersin.org Septerrber 2014 1 Volume 8 1 A"1c le 320 11
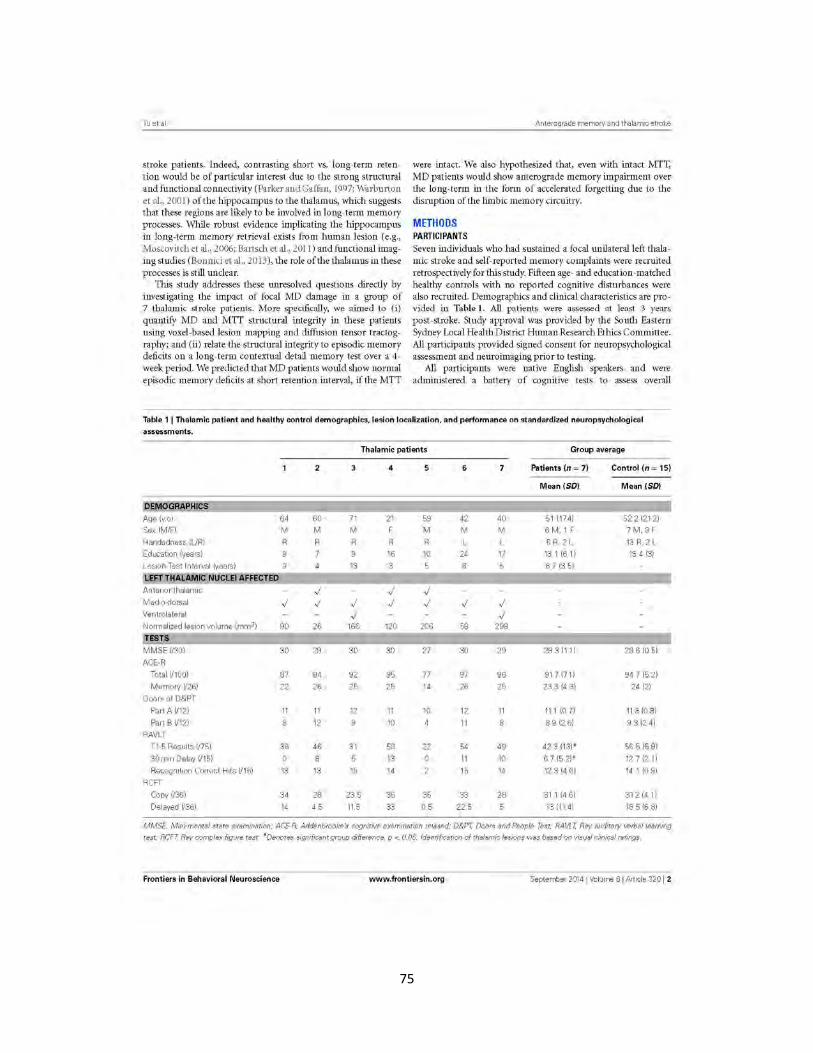
75
Tu et al
stroke patients. Indeed, contrasting short vs. long-term retention would be of particular interest due to the st rong structu ral and functional connect ivity (Parker and Gaffan, 1997; Warburton et al., 2001 ) of the hippocampus to the thalamus, which suggests that these regions are likely to be involved in long-term memory processes. While robust evidence implicating the hippocampus in long-term memory retrieval exists from human lesion (e.g., Moscovitch et al., 2006; Bartsch et al ., 2011 ) and functional imaging studies (Bonnici et al., 2013), the role of the thalamus in these processes is still unclear.
This study addresses these unresolved questions directly by investigating the impact of focal MD damage in a group of 7 thalamic stroke patients. More specifically, we aimed to (i) quantify MD and MTT structural integrity in these pat ients using voxel-based lesion mapping and diffusion tensor tractography; and (ii) relate the structural integrity to episodic memory deficits on a long-term contextual detail memory test over a 4-week period. We predicted that MD patients would show normal episodic memory deficits at short retention interval, if the MTT
Ante rograde memory and t halamic stroke
were intact. We also hypothesized that, even with intact MTT, MD patients would show anterograde memory impairment over the long-term in the form of accelerated forgetting due to the disruption of the limbic memory circuitry.
METHODS PARTICIPANTS Seven individuals who had sustained a focal unilateral left thalamic stroke and self-reported memory complaints were recruited retrospectively for this study. Fifteen age- and education-matched healthy controls with no reported cognitive disturbances were also recruited. Demographics and clinical characteristics are pro vided in Table 1. All patients were assessed at least 3 years post-stroke. Study approval was provided by the South Eastern Sydney Local Health District Human Research Ethics Committee. All participants provided signed consent for neuropsychological assessment and neuroimaging prior to testing.
All participants were native English speakers and were admin istered a battery of cognitive tests to assess overall
Table 1 I Thalamic patient and healthy control demographics, lesion localization, and performan ce on standardized neuropsychological
assessments.
Thalamic patients Group average
2 4 5 6 7 Patients (n = 7) Control (n = 15)
Mean (SD) Mean (SD)
AQe lv.ol 64 60 71 21 59 42 40 51 11741 52.2 12121 Sex IM/FI M M M F M M M 6 M. 1 F 7 M. 8 F
Handedness IURI R R R R R 5 R. 2 L 13 R. 2 L Education (years) 9 16 10 24 17 13.1 (611 13.4 (3)
Lesron-Test Interval (years) 9 13 3 5 8 5 6 7 13 51
LEFT THALAMIC NUCLEI AFFECTED
Anter~or thalamrc ..; ..; ..; Medro-dorsal ..; ..; ..; ..; ..; ..; ..; Ventrolateral ..; ..; Normalized les ion vo lume (mm3J 90 26 166 120 206 58 298
TESTS
MMSE 1/301 30 29 30 30 27 30 29 29 311 11 29 .6 (0 51
ACE-R
Total 1/1001 87 94 92 95 77 97 96 91.7 171) 94.7 (5 21
Memory 1/261 22 26 25 25 14 26 25 233 14 31 24 121
Doors of D&PT
Part AI/121 11 11 12 11 10 12 11 111 (0.7) 113 (0 8)
Part B 1/121 8 12 10 11 8.9 12.61 9.3 124)
RAVLT
T1-5 Results 1/751 36 46 3 1 58 22 54 49 42 3 (13)' 56 5 (5 91
30 mrn Delay 1/151 0 8 13 11 10 6.7 (5.21' 12.712.11
Recognit ion Correct Hrts 1/151 13 13 15 14 15 14 12.3 14.61 14.1 (0.91
RCFT
Copy 1/361 34 28 23 5 36 35 33 28 3 11 (4 6) 3 1214 1)
Delayed 1/361 14 4 5 11 5 33 0 5 22 5 13 11141 18 5 16 81
MMSE, Mini--mental state examination; ACFR, Addenbrooke's cognitive examination revised; D&PT, Doors and People Test; RAVLT, Rey auditory verbal learning
test; RCFT, Rey complex f1gure test. *Denotes signif1cant group difference, p < 0.05. Identification of thalamic lesions was based on visual cliniccl ratings.
Frontiers in Behavioral Neuroscience www. frontiersin . org Septerrber 2014 1 Volume 8 1 Artrcle 320 I 2

76
Tu et al
cognitive function as well as verbal and visual memory. This assessment included: Mini-Mental State Examination (MMSE), Addenbrooke's Cognitive Examination-Revised (ACE-R), doors subtest of the Doors & People Test (D&PT), Rey Auditory Verbal Learning Test (RAVLT), and Rey Complex Figure Test (RCFT). For a brief description of cognitive tasks see Supplementary Table 1. Differences in patient and control demographics and performance on standard cognitive tests were assessed using two -tailed independent samples t-test in SPSS 21.0 (IBM Corp.). P-value < 0.05 was considered statistically significant.
Each participant also underwent a brain MRI scan. All scans were examined by a radiologist for structural abnormalities. None were reported for control participants. All thalamic st roke patients showed infarctions isolated to the thalamus. Assessment of affected thalamic nuclei was performed by LM based on clinical visual rating (Table 1). In two cases additional atrophy was present in the cerebellum (patient 2) and brain stem (patient 3).
LONG-TERM MEMORYTASK Episodic long-term memory was assessed using an object-based visual recognition and recall memory retrieval task. A database of 325 images depicting everyday objects on a white background was created. The word frequency of each object was calculated using the MRC Psycholinguistic Database (http:/ /www.psych.rl. ac.uk) and images were allocated into 13 stimuli sets, each containing 25 images. Stimulus sets were matched for total word frequency, as a proxy measure of object familiarity, so that each set had a similar level of difficulty. Each participant was assigned one stimulus set to encode as target stimuli during training, with remaining sets used as novel clistractors across assessments. Sets assigned as target stimuli were counterbalanced across participants.
During training, 25 target objects were p resented one at a time on either the left or right hand side of a computer monitor. Participants were explicitly asked to remember the object shown as well as the side of presentation. Each target stimulus was shown for 3-s, followed by a 1-s fixation cross. This was followed immediately by a test phase. At test, stimuli consisting of the previously encoded items randomly intermixed with 25 novel items were presented centrally on the computer screen. Participants were asked to make an old/new recognition decision followed by a left/right recall decision (i.e., "Was this object shown on the left or right side of the screen?"). Responses did not have a time limit and no feedback was provided. This procedure was repeated until participants achieved at least 90% correct recognition and recall memory retrieval of the target stimuli on two consecutive trainin~ runs. All participants reached criterion within 3 trainin~ runs. For each new test run, target stimuli images were mixed with a previously unseen set of novel images, such that each set of novel stimuli was only used once throughout all assessments. Training was carried out using E-Prime 2.0 software (Psychology Software Tools, Pittsburgh, PA).
Post -training assessments were carried out via the internet by adapting the task for online assessment using WebExp (http:// groups.inf.ed.ac. uk/webexp/). The test component of the task was reproduced as a java-based application accessible through
Anterograde memory and thalamic stroke
any internet browser. Assessments were carried out after the fol lowing encoding delays: I h, 24 h , I week, 2 weeks, and 4 weeks. The user interface was designed to be simple and clear and each participant was provided a list of unique URL links corresponding to each online assessment. After completing each assessment a number corresponding to that session's result was generated, which participants were required to provide the examiner for data classification. The first post-training assessment was completed I h after the final test phase of training and under the experimenter's supervision to ensure the correct delay was fol lowed and instructions lor completing online testing was clearly understood. Scores of interest were item recognition (i.e., correct identification of whether the image was a target/novel object) and recall of contextual detail (i.e., whether the item was originally presented on the left or the right of the screen). Recognition performance was scored by the "two-high threshold model"; performance index: P, (hit rate - false alarm rate); b ias index: B, (false alarm rate/[ 1-(hit rate - false alarm rate)]) , values greater than 0 indicate conservative bias, values less than 0 indicate liberal bias (Snodgrass and Corwin, 1988; Soei et al., 2008; Pergola et a!., 2012). Separate repeated-measures ANOVAs were carried out across all post -encoding assessments for (i) recognition (P, and B, independently) and (ii) recall across all participants to test for significant within and between group effects, as well as interaction effect with increasing memory retrieval delay. Post-hoc ANOVA contrast examined the change between each subsequent pair of time-points. Independent means comparison t-tests were also conducted between patients and controls for all delayed assessments. Analyses were performed using SPSS 2 1.0 (IBM Corp.). P-value < 0.05 was considered statistically significant.
IMAGING ACOUlSITlON Whole-brain Tl and diffusion weighted images were acquired for all participants using a 3T Philips MRI scanner with standard quadrature head coil (eight channels). Structural Tl scans were acquired as follows: coronal orientat ion, matrix 256 x 256, ISO slices, I mm isotropic, TE/TR = 2.5/5.4 ms, flip angle a =
8°. DTI-weighted sequences were acquired as follows: 32 gradient direction DTI sequence (repetition time/echo time/inversion time: 8400/68/90 ms; b-value = 1000 s/mm2; 55 2.5-mm horizontal slices, end resolution: 2 .5 x 2.5 x 2.5 mm\ field of view 240 x 240 mm, 96 x 96 matrix; repeated twice) (Kwon et al., 2010). Prior to analyses, all participant scans were visually inspected for significant head movements and artifacts; none were found.
THALAMIC LESION LOCALIZATION Lesions within the thalamus were manually traced on each participant's st ructural scan using the Harvard -Oxford Subcortical Structural Atlas in MRJcron. Individual tracings were then normalized using FSL's linear registration tool FLIRT to the MNI standard brain. Patient 's normalized scans were then overlaid to provide a group profile of thalamic lesion location. Lesion vol umes were calculated and correlations with patients' memory performance on standard cognitive memory tests and the long-term memory task were investigated using Pearson's correlation.
Frontiers in Behavioral Neuroscience www.frontiersin.org Septerrber 2014 1 Volume 8 1 A"rcle 320 13

77
Tu et al
TRACTOGRAPHY Probabilistic tractography was carried out to reconstruct the mammillothalamic tract using FMRIB's diffusion toolbox (FDT). DTI-weighted images were eddy-corrected by linearly registering each diffusion weighted volume to the reference T-weighted nondiffusion bO image. All images were brain-extracted and a binary brain mask was created. Diffusion tensor models were fit ted at each voxel, resulting in maps of three eigenvalues (Ll, L2, L3 ), which allowed calculation of fractional anisotropy (FA) and mean diffusivity maps for each subject.
A high resolution DTI data set [Subject 100307; Q2 data set obtained from the MGH-UCLA Human Connectome Project (HCP) database ] was used as a reference for the tractography in our participants.
Manual fiber tractography was carried out for each subject using the probabilistic cross-fiber tracking tool in FDT, PROBTRACKX. Prior to running fiber tracking, Markov Chain Monte Carlo sampling was conducted on eddy corrected DTI images to generate distributions of diffusion paramete rs and model crossing fibers at each voxel. Fiber tracking was then initiated with 5000 streamline samples, 0.5 mm step lengths and 0.2 curvature threshold. MTT were determined using three regions of interest (ROI): two seed regions located at the i) anterior thalamus ii) mammillary body, and a waypoint placed at the level between the mammillary body and bicommissural plane (as previously described by Kwon et al., 2010). A connectivity distribution from all voxels within the seed regions, passing through the waypoint ROI, was generated and thresholded to 30% of the maximum connectivity value to remove image noise. Thresholded fiber connectivity maps were binarized to create a mask of the MTT. Mean FA and mean diffusivity values were then calculated within each participant's fib er tract mask.
RESULTS BEHAVIORAl RESUlTS Thalamic patients and controls were well matched for age and education (p-values > 0.9). Thalamic patients performed in the normal range on assessments of overall cognitive funct ion (MMSE, ACE-R: total and memory subscore) and did not differ significantly from the control group (Table 1). On standard cognitive visual memory tests, patients showed no significant differences compared to controls on the doors subtest of the D&PT and RCFT (all p-values > 0.1) . Patient performance on the 3-min delayed recall of the RCFT varied considerably, with 5 patients performing below the lOth percentile normative score. Performance on the verbal based RAVLT, however, was significantly lower in patients than controls for immediate (Tl -5), and delayed recall (p-values < 0.05), but not on the recognition component (p > 0.3).
On the long-term memory task, 12/15 controls and 5/7 patients reached the successful encoding criterion of correct item recognition and recall of contextual detail for at least 90% of stimuli, on two consecutive training trials, in the first two trials. Two patients (patient I and 4) and 3 controls required one additional training trial to meet criterion. Performances on successful encoding trials were compared between patients and controls. Results indicated both groups of participants learnt target
Ante rograde memory and thalamic stroke
stimuli to the same level for both recognition (old/new) and recall (left/right) retrieval (all p-values > 0.1 ) (Figure 1).
Repeated-measures ANOVA was carried out across all postencoding assessments for both recognition and contextuallocation separately. For recognition performance index (P, ), ANOVA indicated a significant effect of time [F(2.4S7, 49.733) = 14.906, p < 0.001] but no group effect [F( l , 20) = 3.4, p = 0.08] and no interaction [F(2.4S7, 49.733) = 0.948, p = 0.411]. Bias index for recognition responses in patients and controls (0.26 and 0.15, respectively) showed no significant time or group effect across the 4-week delay (all p-values > 0.2). For contextual location, analyses revealed significant main effects of group [F(l, zo) = 8.49, p = 0.009], time [F(2 .589 st.ns) = 62.71,p < 0.001], and a group x time interaction [F (2.589, 51.778) = 3.797, p = 0.02]. Post-hoc contrasts, comparing change in contextual location retrieval between each subsequent assessment and the previous assessment (i.e., 1- 24 h), indicated significant within-group effects across each pair of time-points (all p-values < 0.02 ). Significant between-group effects were also found across each pair of timepoints (all p-values < 0.04) except for the final 2 week delay (2-4 weeks; p = 0.62) . T-tests revealed significant group differences at all delays (p < 0.05), except at 1 h post-encoding (p = 0.38).
STRUCTURAl IMAGING RESUlTS Lesions in thalamic patients were localized in the left thalamus, with patient 3 showing a small additional lesion in the right anterior thalamus (IS mm3 ) . Manual lesion tracing from patients' structural scans showed lesions affect ing the left MD in all patients (Figure 2A). When we created a group lesion representation, after normalizing patient scans onto a standard brain, left MD was the only regwn to show overlap on the group lesion map (Figure 2B).
The MD lesion in 3 patients (2, 4, and 5) partially extended into the AT. Overall, total thalamic lesion volumes did not correlate with memory performance on standard memory tests or the long-term memory task (all p -values > 0.1). Patient 5, however, who performed the lowest across all standardized assessments of memory, was the individual with the largest lesion involving AT (though not the largest total thalamic lesion). We performed the same behavioral analyses between patients and controls on pat ients with and without AT involvement. No significant differences in performance were detected on any of the standard cognitive tests or across delayed assessments on the long-terrn memory task (all p-values > 0.1; Supplementary Figure !).
PROBABiliSTIC TRACTOGRAPHY RESUlTS The MTT was individually reconstructed in both hemispheres in patients and cont rols (Supplementary Figure 2). The tract was first reconstructed using HCP data to obtain the anatomical pathway and a guide for reconstruction in our data. The tract runs posterior to the column of the fornix and lateral from the mammillary bodies to the anterior thalamus. Group FA, mean diffusivity, and tract volume of the MTT were extracted from both hemispheres (Table 2). Diffusion measures (FA, mean diffusivity) of the MTT were consistent across patients, including those with additional lesion involvement of the AT. Independent samples 1-
test revealed no significant differences in FA, mean diffusivity or
Frontiers in Behavioral Neuroscience www. frontiersin. org Septerrber 2014 1 Volume 8 1 Artrcle 320 I 4
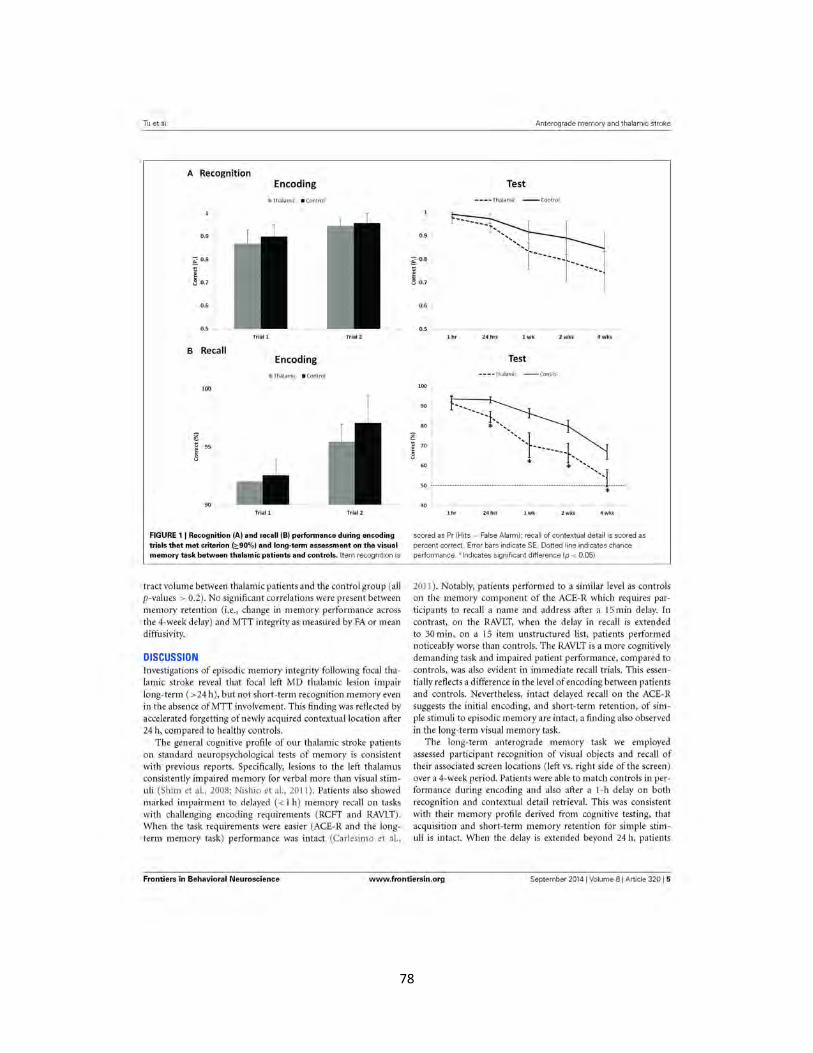
78
Tu et al.
A Recognition
~ 0 .8
t: ~
8 0 .7
0.6
0.5
B Recall
100
~ t: 95 ~
8
90
Encoding
~Thalamic • Control
Tria l l Trial 2
Encoding
@.Thalamic • Control
Triall Trial 2
FIGURE 1 I Recognition (A) and recall (B) performance during encoding trials that met criterion (:::90%) and long-term assessment on the visual memory task between thalamic patients and controls. Item recognition is
tract volume between thalamic patients and the control group (all p-values > 0.2). No significant correlations were present between memory retention (i.e., change in memory performance across the 4-week delay) and MTT integrity as measured by FA or mean diffusivity.
DISCUSSION Investigations of episodic memory integrity following focal thalamic stroke reveal that focal left MD thalamic lesion impair long-term (>24h), but not short-term recognition memory even in the absence ofMTT involvement. This finding was reflected by accelerated forgetting of newly acquired contextual location after 24 h, compared to healthy controls.
The general cognitive profile of our thalamic stroke patients on standard neuropsychological tests of memory is consistent with previous reports. Specifically, lesions to the left thalamus consistently impaired memory for verbal more than visual stimuli (Shim et al., 2008; Nishio et al., 2011 ). Patients also showed marked impairment to delayed ( < 1 h) memory recall on tasks with challenging encoding requirements (RCFT and RAVLT). When the task requirements were easier (ACE-Rand the longterm memory task) performance was intact (Carlesimo et al.,
~ 0 .8
t:
~ u 0.7
Anterograde memory and thalamic stroke
Test
---· Thalamic --contro l
...... ... ................. ... .... ............ ....
lhr 24 hrs lwk 2wks 4wks
Test - - - - Thalamic - control
~~~~~~~~I
• ----------1------.J ' * f .. ..
so ...... ...... .... ........ ... .... .. ............. .... ...... ............. . .. ~ .. .. .. .. -.~~~.J .. . . oL *
scored as Pr (Hits - False Alarm); recall of contextual detail is scored as
percent correct. Error bars indicate SE. Dotted line indicates chance performance. *Indicates significant difference (p < 0.05).
2011 ). Notably, patients performed to a similar level as controls on the memory component of the ACE-R which requires participants to recall a name and address after a 15 min delay. In contrast, on the RAVLT, when the delay in recall is extended to 30 min, on a 15 item unstructured list, patients performed noticeably worse than controls. The RAVLT is a more cognitively demanding task and impaired patient performance, compared to controls, was also evident in immediate recall trials. This essentially reflects a difference in the level of encoding between patients and controls. Nevertheless, intact delayed recall on the ACE-R suggests the initial encoding, and short-term retention, of simple stimuli to episodic memory are intact, a finding also observed in the long-term visual memory task.
The long-term anterograde memory task we employed assessed participant recognition of visual objects and recall of their associated screen locations (left vs. right side of the screen) over a 4-week period. Patients were able to match controls in performance during encoding and also after a 1-h delay on both recognition and contextual detail retrieval. This was consistent with their memory profile derived from cognitive testing, that acquisition and short-term memory retention for simple stimuli is intact. When the delay is extended beyond 24 h, patients
Frontiers in Behavioral Neuroscience www.frontiersin.org September 20141 Volume 81 Article 320 I 5
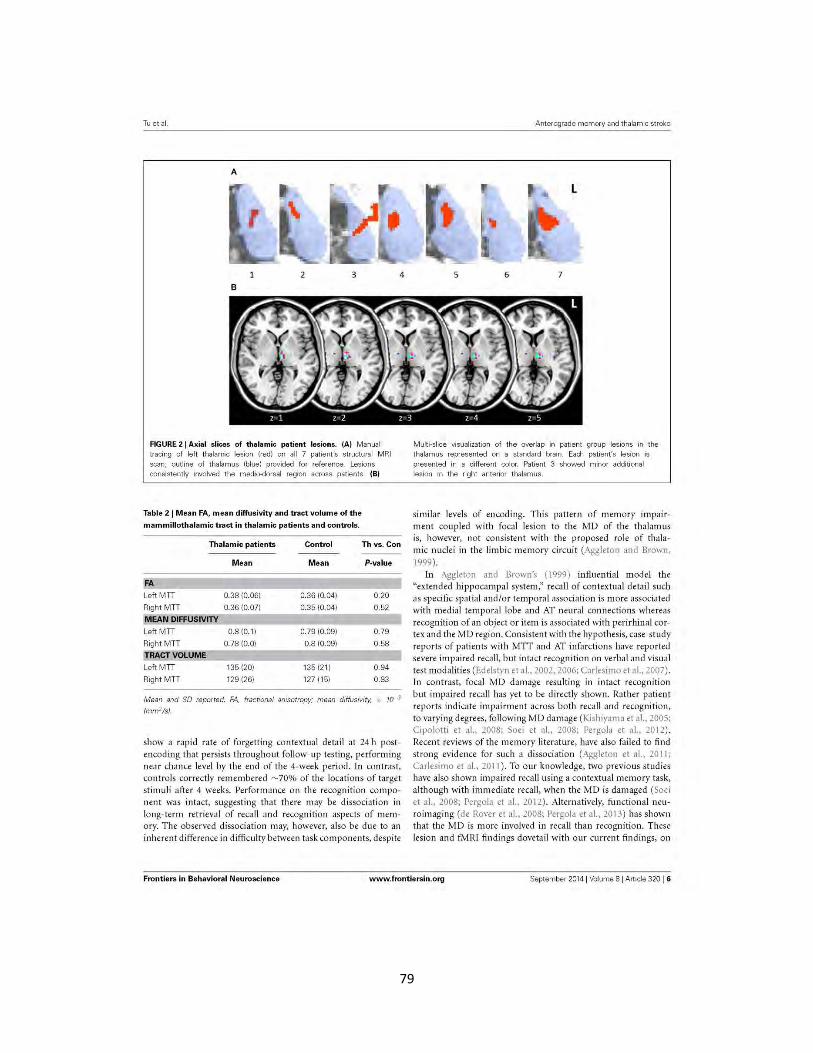
79
Tuetal.
A
2 3
B
FIGURE 21 Axial slices of thalamic patient lesions. {A) Manual tracing of left thalamic lesion (red) on all 7 patient's structural MRI scan; outline of thalamus (blue) provided for reference. Lesions consistently involved the medio-dorsal region across patients. (B)
Table 21 Mean FA, mean diffusivity and tract volume of t he mammillothalamic tract in thalamic patients and controls .
4
Thalamic patients Cont rol Th vs. Con
Mean Mean P-value
FA
LeftMTI 0.38 (0.06) 0.36 (0.041 0.20
Right MTT 0.36 (0.07) 0.35 (0.041 0.52
MEAN DIFFUSIVITY LeftMTI 0.8 (0.1) 0.79 (0.09) 0.79
Right MTT 0.78 (0.0) 0.8 (0.09) 0.58
TRACT VOLUME LeftMTI 135 (20) 135 (21) 0.94
Right MTT 129 (26) 127 (15) 0.83
Mean and SO reported. FA fractional anisotropy; mean diffusivity, x JCI3
lmni'/s).
show a rapid rate of forgetting contextual detail at 24 h postencoding that persists throughout follow-up testing, performing near chance level by the end of the 4-week period. In contrast, controls correctly remembered ~70% of the locations of target stimuli after 4 weeks. Performance on the recognition component was intact, suggesting that there may be dissociation in long-term retrieval of recall and recogn ition aspects of memory. T he observed dissociation may, however, also be due to an inherent difference in difficulty between task components, despite
Anterograde memory and thalamic stroke
5 6 7
Multi-slice visualization of the overlap in patient group lesions in the thalamus represented on a standard brain. Each patient's lesion is presented in a different color. Patient 3 showed minor additional lesion in the right anterior thalamus.
similar levels of encoding. This pattern o f memory impairment coupled with focal lesion to the MD of the thalam us is, however, not consistent with the proposed role of thalamic nuclei i11 t he lim bic memory circuit (Aggleton and Brown, 1999).
In Aggleton and Brown's (1999) influential model the "extended hippocampal system," recall of contextual detail such as specific spatial and/or temporal association is more associated with medial tem poral lobe and AT neural connections whereas recognition of an object or item is associated with perirhinal cortex and the MD region. Consistent with the hypothesis, case-study reports of patients with MTT and AT infarctions have reported severe impaired recall, but intact recognition on verbal and visual test modalities (Edelstyn et al., 2002, 2006; Carlesimo et al., 2007). In contrast, focal MD damage resulting in intact recognition but impaired recall has yet to be directly shown. Rat her patient reports indicate im pairment across both recaU and recognit ion, to varying degrees, following MD damage (Kishiyama et al., 2005; Cipolotti et al., 2008; Soei et al., 2008; Pergola et al., 2012). Recent reviews of the memory literature, have also failed to find strong evidence for such a dissociation (Aggleton et al., 201 I; Carlesimo et al., 2011). To our knowledge, two previous studies have also shown impaired recall using a contextual memory task, a lthough with immediate recall, when the MD is damaged (Soei et al., 2008; Pergola et al., 201 2). Alternatively, functional neuroimaging (de Rover et al., 2008; Pergola et al., 2013) has shown that the MD is more involved in recall than recognition. These lesion and fMRJ findings dovetail with our current fi ndings, on
Frontiers in Behavioral Ne uroscience www.frontiersin.org September 2014 ( Volume 8 I Anicle 320 I 6

80
Tu et al
a group level, showing impaired recall of contextual detail but relatively intact object recognition memory performance. More importantly, lesion volume in patients was not correlated with memory performance in our study, although extension into the AT region, in one patient, resulted in greater memory impairment across cognitive assessments. Therefore, the current findings suggest that the memory functions attributed to sub -thalamic nuclei (AT: recall processes; MD: recognition processes) may be oversimplified. A graded involvement of multiple thalamic nuclei in the recall and recognition dichotomy (Aggleton et a!., 2011; Pergola et a!., 2012) is more consistent with the current findings.
In addition, we were able to directly rule out MTT involve ment for the memory impairment observed in our cohort of patients. Indeed, diffusion measures and tract volume were similar in patients and controls, and showed consistency within groups across hemispheres. Integrity of the MTT in the three patients with additional AT involvement also did not differ from remaining patients. To our knowledge, this is the first study to quantify the integrity of the mammillothalamic tract consistently in thalamic stroke patients on a group level. Diffusion imaging of the MTT provided an objective method of characterizing the extent of lesions in thalamic stroke and their disruption to the limbic memory circuit. In particular, it allowed us to dissociate the extent of thalamic nuclei and MTT contributions to memory impairment.
Importantly, the MD has strong connections to the dorsal lateral prefrontal cortex (DLPFC) (Aggleton et a!., 2011 ). The specific role of the DLPFC in episodic memory processes is not fully established; however, recent functional neuro imaging evidence has shown that the DLPFC is particularly involved in contextual encoding, especially for the association of between-items compared to associations of within-item features (Blumenfeld et a!., 2011). Further, Pergola and colleagues (2013) have shown that the DLPFC and MD are both activated during associative memory retrieval. The contribution of DLPFC and MD connectivity to long-term episodic memory, however, remains unclear. One potential avenue to investigate this in the future would be to contrast thalamic lesion with DLPFC lesion patients directly to elucidate the contributions of each region.
The observed change in retrieval of contextual detail between our thalamic patients and controls in the first week, despite reaching the same level of encoding, behaviorally resembles the accelerated long-term forgetting (ALF) phenomenon identified in some patients with temporal lobe epilepsy (for a review see, Fitzgerald et a!., 2013). Notably, some patients show rapid forgetting fro m as early as 24 h despite normal learning and initial retention, and intact hippocampus (Mayes et a!., 2003; Muhlert et a!., 20 10) . ALF is, therefore, believed to represent a consolidation deficit rather than impaired acquisition (Hoefeijzers et al., 2013). No prior studies have implicated the MD in ALF, although the AT has been shown to be a crucial structure for seizure propagation in temporallobe epilepsy (Mueller eta!., 20 10). Interestingly, Vilberg and Davachi (2013) have recently demonst rated, using a verbal word and object/scene memory task, increased connectivity between the perirhinal cortex and hippocampus fo r selective consolidation of object based memory. Our findings also suggest
Anterograde memory and thalamic stroke
that the MD plays a significant role in long-te rm memory pro cesses, and might be relevant for early stages of consolidation. Similar to the role multiple thalamic nuclei play in declarative memory (Aggleton et a!., 2011), they may also serve roles in memory as a function of time in the HC-cortical consolidation process. The function of the thalamus as a relay structure in the consolidation process, however, has yet to be included in theoret ical models, which remain focused on the hippocampus (Squire and Wixted, 2011; Winocur and Moscovitch, 2011 ). This may be due to a number of factors, (i) when the MTT is not involved, patients with focal damage to the thalami show a more varied and milder anterograde memory impairment ( Carlesimo et a!., 20 II ) that is not detected across a number of standard cogni tive tests and not investigated over the long-term, (ii) anatomical connections leading to thalamic nuclei from medial temporal lobe structures, and from thalamic nuclei to specific cortical regions remains unclear in humans, and (iii) absence of functional imaging studies implicating the thalamus in consolidation processes.
Despite these promising findings, some methodological limitations warrant attention. In particular, despite very careful lesion mapping, we cannot exclude further thalamic damage on a cellular level which could have affected our results. Clearly, intrathalamic connect ivity is complex and thus our results may have been influenced by changes which were not picked up macroscopically. The MTT tractography analysis was conducted according to published protocols (Kwon et al., 2010), however the tract is very small and therefore voxel size of DTI acquisition and partial volume effects might have affected our findings. We addressed accuracy issues by first reconstructing th e tract in a high resolution Human Connectome Project data set to visually assess overall shape and anatomical location of the t ract in our data. Tract volumes of the MTT were bigger than those reported in previous studies (Cipolotti et al., 2008; Kwon et al., 2010), likely due to a larger voxel size acquisition of our DTI data. Obtained FA and mean diffusivity values were, however, consistent across hemispheres and comparable to those reported by Kwon and colleagues (2010), who reliably reconstructed the tract in 25 healthy young controls.
In conclusion, unilateral lesion focal to the left medio-dorsal nuclei of the thalamus, in the absence of damage to the mammillothalamic tract, impairs anterograde memory recall. The findings support the notion that the medio -dorsal nuclei play a role in long-term delayed retrieval of recall type memory processes.
ACKNOWLEDGMENTS The HCP data set was provided by the MGH-UCLA Human Connectome Project. HCP data are disseminated by the Laboratory of Neuro Imaging at the University of Califo rnia, Los Angeles. This work was supported in part by an Australian Research Council (ARC) Discovery Project (DP1093279); the ARC Centre of Excellence in Cognition and its Disorders (CE11000102 1); Sicong Tu is supported by Alzheimer's Australia Dementia Research Foundation and National Health and Medical Research Council (NH&MRC) of Australia awards. MH is supported by an ARC Research Fellowship (DPI IO 104202 );
Frontiers in Behavioral Neuroscience www.frontiersin.org Septerrber 2014 1 Volume 8 1 A"rcle 320 I 7

81
Tu et al
Olivier Pigue! is supported by a NH&MRC Career Development Fellowship (APP1022684) . These sources had no role in the study design, collection, analyses and inte1pretation of data, writing of the manuscript, or in the decision to submit the paper for publication. The authors declare no competing financial interests.
SUPPLEMENTARY MATERIAL The Supplementary Material for this article can be found online at: http:/ /www.frontiersin.org/journal! 10.3389/fnbeh.ZO 14. 00320/abstract
REFERENCES Aggleton, ). P., and Brown, M. W. (1999). Episodic memory, amnesia, and the
hippocampal-anterior thalamic axis. Behav. BraiTI Sci. 22, 425-444. discussion: 444-489. doi: 10.1017/SOI40525X99002034
Aggleton, J.P., Dumont, J. R., and Warburton, E. C. (2011). Unraveling the contributions of the diencephalon to recognition memory: a review. Learn. Mem. 18, 384-400. doi: 10.1101/lm.l884611
Bartsch, T., Dohring, )., Rohr, A., jansen, 0 ., and Deuschl, G. (2011) . CAl neurons in the human hippocampus are cr itical for autobiographical memory, mental time t ravel_. and autonoetic consciousness. Proc. Nat/. Acad. Sci. U.S.A. 108, 17562-17567. doi: 10.1073/pnas.11 10266108
Blumenfeld, R. S., Parks, C. M., Yonelinas, A. P., and Ranganath, C. (2011). Putting Lhe pieces together: the role of dorsolateral prefrontal cortex in relalional memory encodin~; ]. Ccgn. Neurosci. 23, 257-265. doi: 10.1162/jom.2010. 21459
Bonnid, H. M., Chadwick, M. )., and Magu ire, E. A. (2013). Representations of recent and remote autobiographicaJ memories in hippocampal subtields . Hippocampus23, 849-854. doi: 10.! 002/hipo.22155
Carlesimo, G. A., Lombardi, M. G., and Caltagirone, C. (2011). Vascular thalamic amnesia: a reappraisal. Neuropsjchologia 49, 777- 789. doi: 10.1016/j.neuropsychologia.20 11.01.026
Carlesimo_. G. A, Serra. L., Padda, L., Cherubini, A., Bozzali, M., and Caltagirone, C. (2007). Bilateral damage to the mammillo-thalamic tract impairs recollection bu t not familiarity in the recognition process: a single case investigation. Neuropsyd10logia 45, 2467-2479. doi: 10.1016/j.neuropsychologia.2007. 03.025
Cipolotti, L., Husain, M., Crinion, )., Bird, C. M., Khan, S. S., Losseff, N., et al. (2008). The role of the thalamus in amnesia: a tractography, highresolution MRI and neu ropsychological study. Neuropsychologia 46,2745- 2758. doi: 10.1016/j.neuropsychologia.2008.05.009
de Rover, M., Petersson, K. M., van der Werf, S. P., Cools, A. R., Berger, H. J,, and Fernandez, G. (2008). Neural correlates of strategic memory retrieval: differentiating between spatial-associative and temporal-associative strategies. Hum. BrainMapp. 29,1068-1079. doi: 10.!002/hbm.20445
Edelstyn, N. M., Elli~ S. )., Jenkinson, P., and Sawyer, A. (2002). Contribution of the left dorsomedial thalamus to recognition memory: a neuropsychologicaJ case study. Neurocase 8, 442- 452. doi: 10.1076/neur.8.5.442.16180
Edelstyn, N. M .• Hunter, B .• and Elli~ S. ) . (2006). Bilateral dorsolateral thalamic lesions disrupts conscious recollection. Neuropsychologia 44, 931-938. doi: 10.1016/j.neuropsychologia,2005.08.012
Fitzgerald, Z., Mohamed, A., Ricci, M., Thayer, Z., and Miller, L. (2013). Accelerated long-term forgetting: a newly identified memory impairment in epilepsy. /. Oin. Neurosci. 20, 1486-1491. doi: 10.1016/j.jocn.2013.04.037
Hampstead, B. M., and Koffler, S. P. (2009). Thalamic contr ibutions to anterograde, retrograde, and implicit memory: a case study. C!in Neuropsychol. 23, 1232- 1249. doi: 10.1080/ 13854040902936679
Hoefeijzers, S., Dewar, M., Della Sala, S., Zeman, A., and Butler, C. (2013). Accelerated long-term forgetting in transient epileptic amnesia: an acqu isition or consolidation defidt? Neuropsychologia 51, 1549- 1555. doi: 10.10 16/j.neuropsychologia.20 13.04.017
Kishiyama, M. M., Yonelinas, A. P .• Kroll, N. E., Lazzara, M. M., Nolan, E. C., jones, E. G., et al. (2005). Bilateral thalamic lesionsaffoct recollection- and familiaritybased recognition memory judgments. Cortex 41 , 778-788. doi: 10.1016/SOOl 0-9452(08)70296-X
Anterograde merrmy and t halamc stroke
Kwon, H. G., Hong, ). H., and )ang, S. H. (2010). Mammillothalamic tract in human brain: diffusion tensor tractography study. Neurosci. Lett. 481, 51-53. doi: 10.1016/j.neulet.2010.06.052
Mayes, A. R., Isaac, C. L., Hold stock, ). S., Cariga, P., Gummer, A., and Robert~ N. (2003). Long-term amnesia: a review and detailed illustrative case study. Cortex 39, 567-603. doi: 10.1016/SOO I0-9452(08)70855-4
Moscovitch, M., Nadel, L., Winocur, G., Gilboa, A., and Rosenbaum, R. S. (2006). The cognitive neuroscience of remote episodic, semantic and spatial memory. Cu ... Opin. Neurobiol. 16, 179-190. doi: 10.1016/j.conb.2006.03.013
Mueller, S. G., Laxer, K. D., Barako~ )., Cheong, I., Finlay, D., Garcia, P., et al. (2010). Involvement of the thalamocorticaJ network in TLE with and without mesiotemporal sclerosis.Epilepsia 51, 1436-1445. doi: 10.1 111/j. l528-1 167. 2009.0241 3.x
Muhlert. N., Milton, F., Butler, C. R., Kapur, N., and Zeman, A. z. (2010). Accelerated forgetting of real-life events in Transient Epileptic Amnesia Neuropsymologia 48, 3235-3244. doi: 10.1016/j.neuropsychologia.2010. 07.001
Nishio, Y., Hashimoto, M., Ishii, K., and Mori, E. (2011). Neuroanatomy of a neurobehavioral disturbance in the left anterior thalamic infarction. f. Nwrol. Neuro""K· Psychiatry 82, 1195-1200. doi: 10.1136/jn np.20!0.236463
Parker, A., and Gaffan, D. (1997). Marnillary body lesions in monkeys impair object-in-place memory: functional unity of the fornix-mamillary system. ]. Cogn. Neurosci. 9, 512-521. doi: 10.1162/j ocn.l 997.9.4.512
Pergola, G., Gunturkun, 0 ., Koch, B.,Schwarz, M., Daum, 1., and Suchan, B. (2012). Recall deficits in stroke patients with thalamic lesions covary with damage to the parvocellular mediodorsal nucleus of the thalamus. Neuropsychologia 50, 2477-2491. doi: 10.1016/j.neuropsychologia.2012.06.019
Pergola, G., Ranft, A., Mathias, K., and Suchan, B. (2013) . The role of the thalamic nudei in recognition memory accompanied by recall during encoding and relrievaJ: an tMRI study. Neuroimage 74, 195- 208. doi: I 0.1 0!6/j.neuroimage.20 13.02.017
Perren, F., Clarke, S., and Bogousslavsk")', ). (2005). The syndrome of combined polar and paramedian thalamic infarction. Ardr. NwroL 62~ 1212-1216. doi: 10.1001/archneur.62.8.1212
Preston, A. R., and Eichenbaum, H. (2013). Interplay of hippocampus and prefrontal cortex in memory. Curr. Bioi. 23, R764-R773. doi: 10.1016/j.cub.2013.05.011
Saunders, R. C., Mishkin, M., and Aggleton, ). P. (2005). Projections from the entorhinal cortex, per irhinal cortex, presubiculum, and parasubiculum to the medial thalamus in macaque monkeys: identifying different pathways using disconnection techniques. Exp. Brain Res. 167, 1-16. doi: 10.1007/s00221-005-2361-3
Shim, Y. S., Kim, ). S., Shon, Y. M., Chung, Y. A., Ahn, K. )., and Yang, D. W. (2008). A serial study of regional cerebral blood flow deficits in patients with lett anterior thalamic infarction: anatomical and neuropsychological correlates. ]. Neurol. Sci. 266, 84-91. doi: 10.1016/j.jns.2007.09.016
Snodgrass,). G., and Corwin, ). ( 1988). Pragmatics of measuring recognition mem· ory: applications to dementia and amnesia f. Exp. Psycho!. Gtn. 117, 34-50. doi: 10.1037/0096-3445.117.1.34
Soei, E., Koch, B., Schwarz, M., and Daum, I. (2008). Involvement of the human thalamus in relational and non-relational memory. Eur./. Neurosci. 28, 2533-2541. doi: 10.1111/j.1460-9568.2008.06536.x
Squire. L. R., and Wixted, J, T. (2011). The cognitive neuroscience of human memory since H.M. Amw. Rev. Neurosci. 34, 259- 288. doi: 10.1146/ annurevneuro-061010- 113720
Van der Werf, Y. D., Schelten~ P., Undeboom, )., Witter, M. P., Uyling~ H. B., and Jolles, J. (2003). Deficits of memory, executive functioning and attention following infarction in the thalamus; a study of 22 cases with localised lesions. Ne<,opsychologia 41, 1330-1344. doi: 10.1016/50028-3932(03) 00059-9
Vann, S. D. (2010). Re-evaluating the role of the mammillary bodies in memory. Neuropsymologia 48,2316-2327. doi: 10.1016/j.neuropsychologia.2009.1 0.019
Vilberg, K. L., and Davachi, L. (2013). Perirhinal-hippocampal connectivity during reactivation is a marker for object-based memory consolidation. Nwron 79, 1232-1242. doi: 10.1016/j.neuron .2013.07.013
Warburton, E. C ., Baird, A., Morgan, A., Muir, ). L., and Aggleton, ). P. (2001). The conjoint importance of the hippocampus and anterior thalamic nuclei for allocentric spatial learning: evidence from a disconnection study in the rat. ]. Neurosci. 21, 7323-7330.
Frontiers in Behavioral Neuroscience www. frontiersin.org Septerrber 2014 I Volurre 81 Article 320 I 8

82
Tu et al
Winocur, G .• and Moscovitch, M. (2011). Memory transformation and systems consolidatio n. f. Int. Nemoprychol. Soc. 17, 766- 780. doi: 10.1017/S!3SS6177 11000683
ConOkt of Interest Staterrent: The authors declare that the research was conducted in the absence of any commercial or finandal relationships that could be construed as a potential conflict of interest.
Received: 06 June 2014; accepted: 28 August 2014; published online: 15 September 21!14.
Anterograde memory and thalamic stroke
Citation: Tu S, Miller L Piguet 0 and Hornberger M (2014) Accelerated forgetting of contextual details due to focal medio- dorsal thalamic lesion. Front. Behav. Neurosci.
8:320. doi: 10.3389/fnbeh.2014.00320 This article was submitted to the journal Frontiers in Behavioral Neuroscience. Copyright e 2014 Tu, Miller, Piguet and Hornberger. This is an open-access article
distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or repro1luction in other forums is permitted, provided the
original author(s) or licensor are credited and that the original publication in this
journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
Frontiers in Behavioral Neuroscience www.frontiersin.org Septerrber 2014 1 Volume 8 1 A"rcle 320 19

83
Chapter 4 – Longitudinal Papez Memory Circuit Integrity in
Dementia
Publication IV – “Longitudinal Papez circuit integrity in Alzheimer’s disease and
frontotemporal dementia” (in submission)

84
Longitudinal Papez circuit integrity in Alzheimer’s disease and
frontotemporal dementia
Sicong Tu1,2,3, Olivier Piguet1,2,3, John R. Hodges1,2,3, Michael Hornberger2,4
1 Neuroscience Research Australia, Randwick, Sydney, Australia.
2 Australian Research Council Centre of Excellence in Cognition and its Disorders, Sydney,
Australia.
3 School of Medical Sciences, University of New South Wales, Sydney, Australia.
4Norwich Medical School, University of East Anglia, Norwich, United Kingdom.
Running Title: Papez Circuit in AD/FTD
Corresponding author:
Prof. Michael Hornberger
Norwich Medical School, University of East Anglia, Norwich, NR47UQ, United Kingdom
Tel: +44 (0)1603 597139

85
Abstract
Background: Damage to extra-hippocampal structures along the circuit of Papez can result in
episodic memory impairment as severe as when the hippocampus itself is damaged. Recent
evidence suggests Papez circuit structures are differentially affected in dementia syndromes with
memory deficits, such as Alzheimer’s disease (AD), and the behavioural (bvFTD) and semantic
variants (svPPA) of frontotemporal dementia.
Objective: Characterise longitudinal in-vivo Papez circuit integrity in AD, bvFTD, and svPPA,
and the impact on progressive memory decline.
Method: Longitudinal change over 1 year was assessed in a cohort of 88 patients with dementia
(35 AD; 34 bvFTD; 19 svPPA) and 15 healthy controls on a battery of standardized
neuropsychological measures (general cognition, visual and verbal episodic memory), and
integrity of Papez circuit structures.
Results: Dissociable patterns of baseline and longitudinal change were observed in Papez
structures between AD, bvFTD, and svPPA, in particular along the cingulate gyrus. Decline on
measures of general cognition showed greater sensitivity than specific measures of verbal and
visual memory.
Conclusions: Disease pathology in AD, bvFTD, and svPPA differentially affects the integrity of
Papez structures associated with episodic memory performance. These neural changes were not
detectable using standard episodic memory measures and more sensitive tasks are proposed to
monitor longitudinal disease progression.
Keywords: Papez circuit, episodic memory, Alzheimer’s disease, frontotemporal dementia

86
Introduction
Alzheimer’s disease (AD) and frontotemporal dementia (FTD) are the two most common
younger onset dementias. Clinically, AD is defined by the predominant episodic memory
deficits, whereas FTD subtypes are defined by primary language and behavioural problems.
Increasing evidence, however, indicates that some FTD patients can show significant episodic
memory problems, in particular for the behavioural (bvFTD) and semantic language (svPPA)
variants of FTD [1-3]. Deficits in episodic memory processing in these neurodegenerative
conditions has been attributed mostly to hippocampal dysfunction, with findings from in-vivo
and post-mortem studies highlighting significant hippocampal atrophy during the course of
disease [4-6]. While the hippocampus is well recognised as a crucial hub for episodic memory,
with damage resulting in mild to severe amnesic symptoms [7], difference in clinical memory
profiles (i.e., visual memory recall intact in svPPA, impaired in AD and bvFTD) highlight the
contribution of extra-hippocampal structures comprising an extended whole brain memory
network [8,9]. Indeed, even within anatomically well-established circuits of memory function,
such as the circuit of Papez [10,11], multiple extra-hippocampal relay structures are affected in
both AD and FTD.
The Papez circuit comprises: the hippocampus, fornix, mammillary body,
mammillothalamic tract, anterior nucleus of the thalamus, anterior thalamic radiation, cingulate
gyrus, parahippocampal gyrus, and entorhinal cortex. Key components of the circuit
(hippocampus, fornix, anterior thalamus, cingulate gyrus) have been consistently shown to be
associated with episodic memory in a variety of patient populations and healthy individuals [12-
15]. In AD, the hippocampus, fornix and cingulate gyrus show structural and/or metabolic
change in the early stages of disease pathology and have been proposed to be indicators of

87
conversion from mild cognitive impairment, the prodromal stage [16-18], and underlie observed
memory impairments [16,19,20]. In contrast to AD, early disease pathology in FTD is relatively
focal and typically found in cortical regions, including the orbitofrontal and anterior cingulate
cortices in bvFTD [3,21], and anterior temporal lobe in svPPA [22]. While hippocampal atrophy
in svPPA has long been recognised, primarily in the anterior hippocampus [23], only recently has
similar dysfunction been recognised in bvFTD [4,24,25]. Recent studies of pathologically
confirmed patients have also shed light on significant extra-hippocampal atrophy along the Papez
circuit in bvFTD and svPPA, namely in the anterior thalamus and fornix [4,24]. Despite these
changes seen in Papez circuit regions, the focus of most neurodegenerative memory diagnosis
and research has been solely on the hippocampus. One reason for this is that it has been argued
that the hippocampus is the main memory processing component of the Papez circuit, while the
remaining parts lie ‘downstream’ from the hippocampus and are therefore likely involved only in
the modulation of episodic memory rather than its actual encoding and retrieval. From a disease
staging point of view, this would suggest that the hippocampus is first involved with other Papez
regions being only affected after, due to the downstream spread of the neurodegenerative
pathology.
The current study explores this hypothesis by characterising longitudinal in-vivo Papez
circuit integrity in clinically well-defined cases of AD, bvFTD and svPPA patients. In particular,
whether there are dissociable patterns of longitudinal change between patient groups and to what
extent this impacts episodic memory performance.

88
Methods
Participants
Eighty eight dementia patients (35 AD; 34 bvFTD; 19 svPPA) and 15 healthy controls were
recruited from the Sydney frontotemporal dementia research group (FRONTIER) database. All
participants were assessed at the FRONTIER clinic located at Neuroscience Research Australia,
Sydney. Study approval was provided by the South Eastern Sydney Local Health District and
University of New South Wales Human Research Ethics Committees. All participants provided
signed consent for neuropsychological assessment and neuroimaging prior to testing. Patient
cohorts were matched for age, education, and disease duration. Patients were assessed at two
time-points: baseline and 1-year follow-up. All dementia patients fulfilled international
consensus criteria for AD [1], bvFTD [3], and svPPA [2]. Clinical diagnoses were established by
consensus among senior neurologist, occupational therapist and neuropsychologist, based on a
clinical interview, comprehensive neuropsychological assessment, and evidence of brain atrophy
on structural neuroimaging. All bvFTD patients showed disease progression as well as atrophy
on scans to exclude any phenocopy cases [26]. Participant demographics and clinical
characteristics are provided in Table 1.
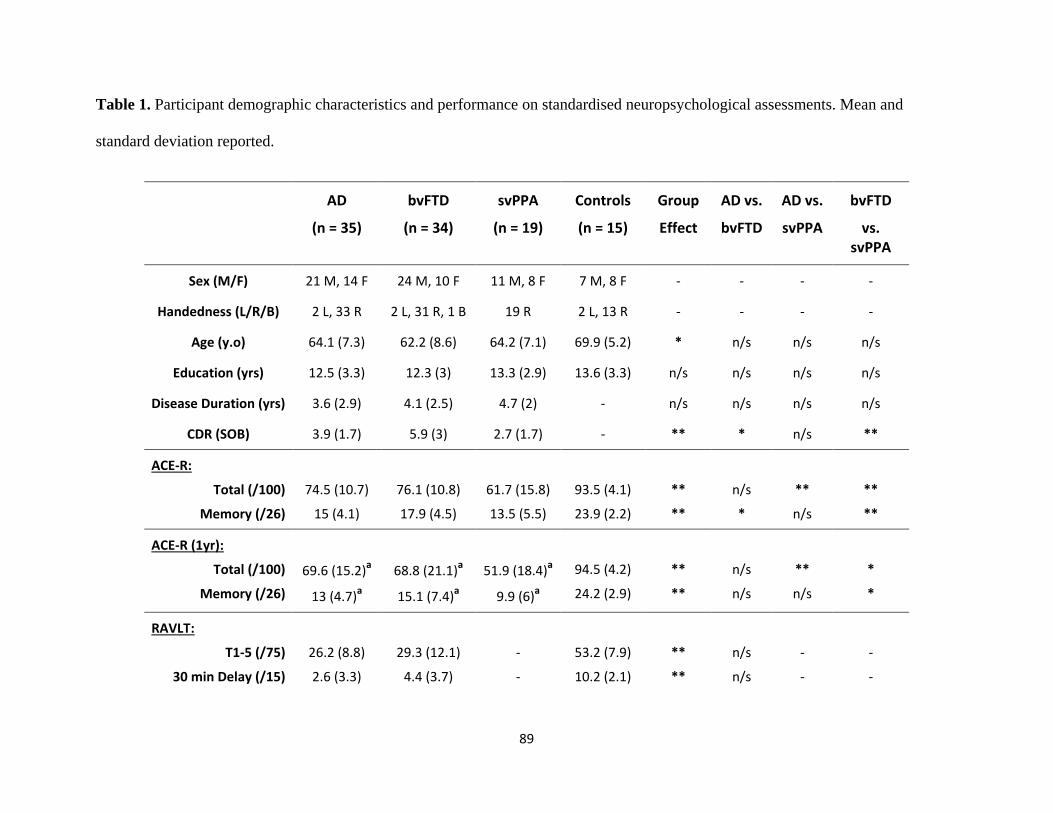
89
Table 1. Participant demographic characteristics and performance on standardised neuropsychological assessments. Mean and
standard deviation reported.
AD
(n = 35)
bvFTD
(n = 34)
svPPA
(n = 19)
Controls
(n = 15)
Group
Effect
AD vs.
bvFTD
AD vs.
svPPA
bvFTD
vs. svPPA
Sex (M/F) 21 M, 14 F 24 M, 10 F 11 M, 8 F 7 M, 8 F - - - -
Handedness (L/R/B) 2 L, 33 R 2 L, 31 R, 1 B 19 R 2 L, 13 R - - - -
Age (y.o) 64.1 (7.3) 62.2 (8.6) 64.2 (7.1) 69.9 (5.2) * n/s n/s n/s
Education (yrs) 12.5 (3.3) 12.3 (3) 13.3 (2.9) 13.6 (3.3) n/s n/s n/s n/s
Disease Duration (yrs) 3.6 (2.9) 4.1 (2.5) 4.7 (2) - n/s n/s n/s n/s
CDR (SOB) 3.9 (1.7) 5.9 (3) 2.7 (1.7) - ** * n/s **
ACE-R:
Total (/100)
Memory (/26)
74.5 (10.7)
15 (4.1)
76.1 (10.8)
17.9 (4.5)
61.7 (15.8)
13.5 (5.5)
93.5 (4.1)
23.9 (2.2)
**
**
n/s
*
**
n/s
**
**
ACE-R (1yr):
Total (/100)
Memory (/26)
69.6 (15.2)a
13 (4.7)a
68.8 (21.1)a
15.1 (7.4)a
51.9 (18.4)a
9.9 (6)a
94.5 (4.2)
24.2 (2.9)
**
**
n/s
n/s
**
n/s
*
*
RAVLT:
T1-5 (/75)
30 min Delay (/15)
26.2 (8.8)
2.6 (3.3)
29.3 (12.1)
4.4 (3.7)
-
-
53.2 (7.9)
10.2 (2.1)
**
**
n/s
n/s
-
-
-
-
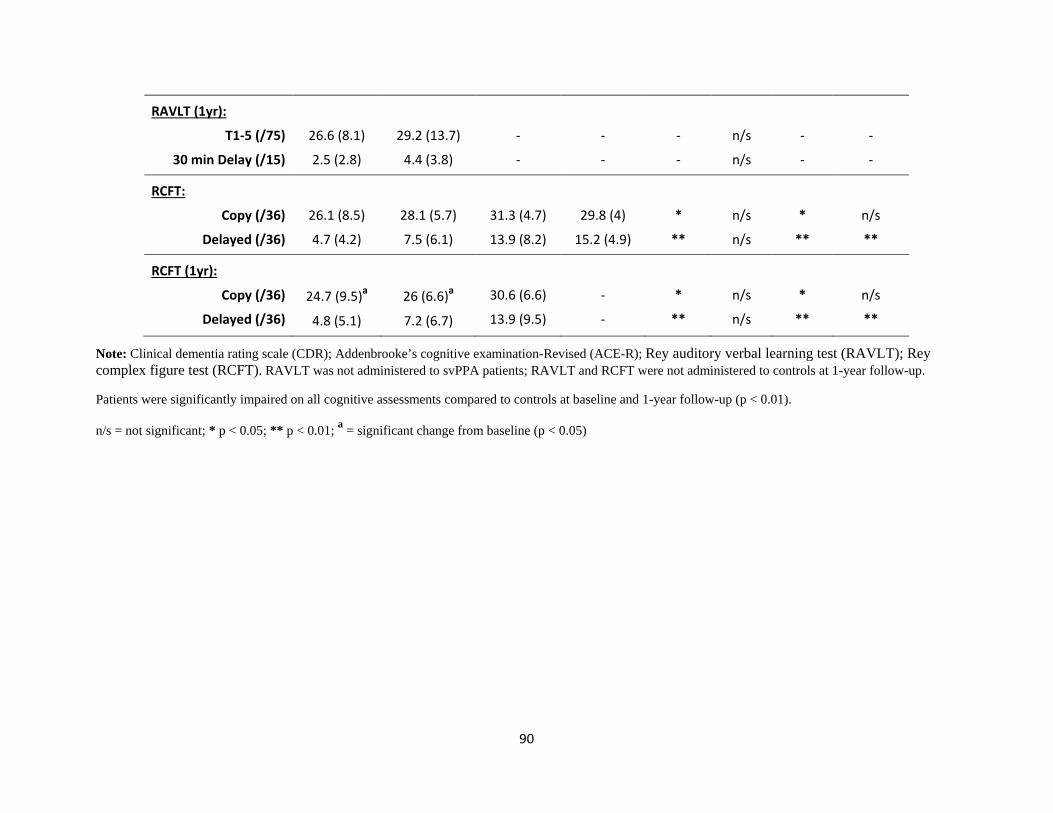
90
RAVLT (1yr):
T1-5 (/75)
30 min Delay (/15)
26.6 (8.1)
2.5 (2.8)
29.2 (13.7)
4.4 (3.8)
-
-
-
-
-
-
n/s
n/s
-
-
-
-
RCFT:
Copy (/36)
Delayed (/36)
26.1 (8.5)
4.7 (4.2)
28.1 (5.7)
7.5 (6.1)
31.3 (4.7)
13.9 (8.2)
29.8 (4)
15.2 (4.9)
*
**
n/s
n/s
*
**
n/s
**
RCFT (1yr):
Copy (/36)
Delayed (/36)
24.7 (9.5)a
4.8 (5.1)
26 (6.6)a
7.2 (6.7)
30.6 (6.6)
13.9 (9.5)
-
-
*
**
n/s
n/s
*
**
n/s
**
Note: Clinical dementia rating scale (CDR); Addenbrooke’s cognitive examination-Revised (ACE-R); Rey auditory verbal learning test (RAVLT); Rey complex figure test (RCFT). RAVLT was not administered to svPPA patients; RAVLT and RCFT were not administered to controls at 1-year follow-up.
Patients were significantly impaired on all cognitive assessments compared to controls at baseline and 1-year follow-up (p < 0.01).
n/s = not significant; * p < 0.05; ** p < 0.01; a = significant change from baseline (p < 0.05)

91
Briefly, AD patients presented predominantly with significant episodic memory
impairment with preserved social behaviour. BvFTD patients demonstrated changes in social
functioning, loss of insight, disinhibition and increased apathy. SvPPA patients showed loss of
general conceptual knowledge in the form of significant naming and comprehension impairment.
Exclusion criteria for all participants included prior history of mental illness, head injury,
movement disorders, alcohol and drug abuse, limited English proficiency, and, for controls,
presence of abnormality on MRI. Participants were administered a battery of cognitive tests to
assess general cognitive function, and verbal and visual memory. This assessment included:
Addenbrooke’s Cognitive Examination-Revised (ACE-R), Rey Auditory Verbal Learning Test
(RAVLT), and Rey Complex Figure Test (RCFT). For a brief description of cognitive tasks see
Supplementary Table 1.
Imaging Acquisition
All participants underwent whole-brain T1 and serial diffusion weighted imaging using a 3T
Philips MRI scanner with standard quadrature head coil (eight channels) at initial and follow-up
assessments. Structural T1-weighted images were acquired using the following sequences:
coronal orientation, matrix 256 x 256, 200 slices, 1 x 1 mm in-plane resolution, slice thickness 1
mm, echo time/repetition time = 2.6/5.8 ms, flip angle α = 8°. Thirty-two direction diffusion
weighted images were acquired using the following sequence: repetition time/echo
time/inversion time = 8400/68/90 ms, b-value = 1000s/mm2, 55 slices, end resolution: 2.5 x 2.5 x
2.5 mm, field of view: 240 mm x 240 mm, 96 x 96 matrix, repeated twice. All scans were
examined by a radiologist to rule out structural abnormalities; none were reported for control
participants. Prior to analyses, all participant scans were visually inspected for significant head
movements and artefacts.

92
Statistical Analyses
Behavioural data were analysed using IBM SPSS statistics (version 21.0; IBM Corp., Armonk,
NY). Kolmogorov-Smirnov tests were run to determine the suitability of variables for parametric
analyses. Analyses of variance (ANOVA), followed by Tukey post-hoc tests, were used to
explore main effects between participant groups on demographic variables (age; education;
disease duration; clinical dementia rating) and visual memory (RCFT). ACE-R performance was
analysed using Kruskal-Wallis tests, followed by post-hoc Mann-Whitney tests. Verbal memory
performance on the RAVLT was analysed using two-tailed independent samples t-test.
Longitudinal change within participant groups were assessed using two-tailed paired samples t-
test and Wilcoxon tests, for parametric and non-parametric data, respectively. Longitudinal
change between participant groups was assessed using linear regression with baseline
performance included as a covariate. In all analyses p values < 0.05 were considered to be
significant.
Imaging Analyses
Grey Matter: Voxel-based morphometry (VBM) was conducted on whole-brain T1-weighted
scans, using the VBM toolbox in FMRIB’s Software Library (FSL;
http://www.fmrib.ox.ac.uk/fsl/) using the same optimal processing pipeline employed in previous
studies by our group [4,6]. Briefly, brain extraction was performed using FSL BET algorithm
[27] with an optimal fractional intensity threshold of 0.22. Each scan was visually checked
following brain extraction to ensure no brain matter was excluded, and no non-brain matter was
included. A study specific template of grey matter was generated from 15 scans for each
participant cohort, to avoid topographical bias during registration. For the longitudinal analysis,
each individual’s initial and follow-up scan was co-registered to generate a within-subject

93
template representing a half-way transformation between the two time-points, prior to
registration on the study-specific template [28]. Template scans were then registered to the
Montreal Neurological Institute (MNI) standard brain (MNI 152), resulting in a study-specific
grey matter template at 2 mm3 resolution in MNI standard space. Participant brain-extracted
scans were segmented into CSF, grey matter and white matter, and non-linearly registered to the
study-specific template and smoothed using an isotropic Gaussian kernel (sigma = 3 mm).
White Matter: White matter integrity was assessed using the TBSS toolbox in FSL [29], and
complemented by DTI-TK for improved tensor based registration in longitudinal analyses
[30,31]. Briefly, serial diffusion-weighted sequences from each participant were averaged to
improve signal-to-noise ratio before being corrected for eddy current distortions. A binary brain
mask was generated from the non-diffusion volume (b0) and used to fit a tensor model onto
diffusion-weighted images. DTI-TK was used to generate study-specific brain atlases, using each
participant’s diffusion tensor image, through an iterative process of initial rigid registration,
followed by non-linear registration, to determine the best mapping with each individual’s scan
using the least amount of deformation [32,33]. For the longitudinal analysis, each individual’s
initial and follow-up scan was co-registered to generate a within-subject template representing a
half-way transformation between the two time-points, prior to construction of a group atlas [32].
Group atlases were registered to the Illinois Institute of Technology standard brain atlas, version
4.0 [34]. Transformation matrices were generated for each stage of registration to create a
deformation field that defines the mapping directly from each individual’s native scan space to
standard space in a single interpolation [32]. Individual fractional anisotropy (FA) maps are then
generated, concatenated and skeletonized [35] to define the lines of maximum FA, which
correspond to the centres of white matter tracts.

94
Comparison of whole-brain grey and white matter integrity was carried out between
patient groups and control at baseline to establish overall atrophy profile in each patient cohort.
Region of interest analyses were then conducted to specifically examine the integrity of Papez
grey (hippocampus; mammillary bodies; anterior thalamus; posterior cingulate) and white
(fornix; cingulum) matter structures at baseline between patient groups and longitudinal change
within group over the 1-year period. Statistical analyses were performed using a voxel-wise
general linear model and threshold-free cluster enhancement [36] to detect significant clusters of
change. We employed permutation-based non-parametric testing with 5000 permutations per
contrast [37]. Age was included as a nuisance variable in these analyses. Reported clusters are
corrected for multiple comparisons via family-wise error (FWE) and tested for significance at p
< .05. Papez structures were delineated using the Harvard-Oxford subcortical atlas, John
Hopkins University tractography atlas, and manual segmentation, to allow grey and white
volume extraction for further analyses using repeated measures ANOVA in SPSS.
Results
Demographics
Patient cohorts were well matched for age, education and disease duration (Table 1). Clinical
rating of functional impairment (CDR), however, did differ between patient cohorts (all p values
< 0.05), reflecting different degrees of inherent social disturbance across conditions. Healthy
control participants were overall well matched with all patient cohorts for age and years of
education, but were significantly older than the bvFTD group (p value = 0.007).

95
Longitudinal Memory Profile
ANOVAs indicated significant differences between groups on performance across standard
cognitive measures at baseline and 1-year follow-up (all p values < 0.05), with the exception of
RAVLT follow-up performance due to the absence of svPPA and control testing (Table 1).
Patients were significantly impaired on all cognitive assessments compared to controls at
baseline and 1-year follow-up (all p values < 0.01).
Patient Baseline Performance: BvFTD patients showed the least impairment on general cognition
(ACE-R: Total) and verbal memory (ACE-R: Memory; RAVLT), compared to AD and svPPA.
BvFTD performance, however, was not significantly better than AD, except on the memory
component of the ACE-R (p value = 0.032). SvPPA performed significantly worse than bvFTD
on the ACE-R (all p values < 0.003), and AD on ACE-R total score (p value = 0.001). For visual
memory recall (RCFT: Delayed), however, svPPA performed significantly better than AD and
bvFTD patient groups (all p values < 0.003). AD and bvFTD showed the same level of
performance for visual memory recall on the RCFT (p value = 0.372).
Patient 1-Year Follow-up Performance: Pattern of verbal and visual memory impairment
remained the same at follow-up, compared to baseline, between patient cohorts. AD and bvFTD
showed the same level of verbal and visual memory impairment (ACE-R; RAVLT; RCFT; all p
values > 0.9). Compared to svPPA, bvFTD showed significantly better performance on all
components of the ACE-R (p values < 0.04), while AD only demonstrated better performance for
total ACE-R score (p value = 0.01). For visual memory recall on the RCFT, svPPA performance
remained significantly higher than AD (p value < 0.001) and bvFTD (p value = 0.003). Paired
samples t-test indicated a significant decline in performance over 1 year on the ACE-R within all

96
patient cohorts (all p values < 0.02), but not verbal or visual memory recall on the RAVLT and
RCFT, respectively. Regression analysis comparing follow-up performance across all cognitive
measures between patient groups, using baseline performance as a covariate, however, did not
detect any significant differences (all p values < 0.1). Percentage of decline across standard
cognitive measures indicated very mild visual and verbal memory decline over a period of 1 year
for all participant cohorts (Fig. 1). ANOVA indicated total score on the ACE-R measure of
general cognition was the only task that showed significant group differences, specifically,
patient cohorts all showed significantly greater decline relative to control (all p values < 0.05),
but not between patient groups. Similarly, specific measures of verbal and visual memory recall
showed no significant differences between patient groups. BvFTD showed the greatest degree of
variability in longitudinal change across cognitive measures, followed by AD.
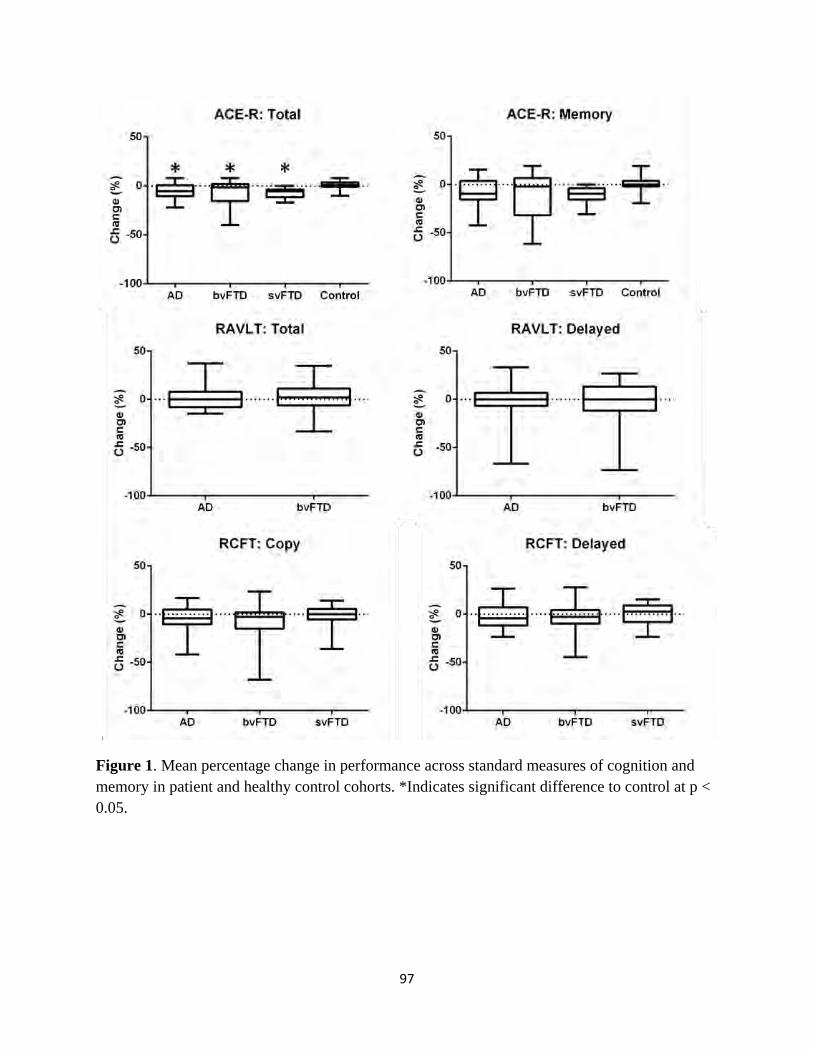
97
Figure 1. Mean percentage change in performance across standard measures of cognition and memory in patient and healthy control cohorts. *Indicates significant difference to control at p < 0.05.

98
Papez Circuit Integrity
Baseline comparisons of grey matter in the Papez structures in patient cohorts, relative to control,
indicated significantly reduced integrity in the hippocampus, anterior thalamus, and cingulate
gyrus (Fig. 2). A similar pattern of atrophy was detected bilaterally along the entire length of the
hippocampus and anterior thalamus in AD and FTD patient groups. Patient groups, however,
demonstrated dissociable patterns of atrophy in the cingulate gyrus. Specifically, atrophy
primarily involved the posterior cingulate in AD, anterior cingulate in bvFTD, and both regions
of the cingulate being intact in svPPA. Longitudinal comparison of change over 1 year within
patient cohorts showed divergent patterns of progressive atrophy between all patient groups. In
AD, progressive atrophy was observed in the posterior cingulate and anterior thalamus, but not
hippocampus. In contrast, progressive atrophy was present in the head and body of the right
hippocampus in bvFTD and svPPA. Progressive atrophy was detected along the entire length of
the left hippocampus in svPPA only. Similar to baseline, dissociable patterns of longitudinal
cingulate gyrus change was detected across patient groups. Specifically, atrophy remained
confined to the posterior cingulate in AD, anterior cingulate in svPPA, and both regions of the
cingulate affected in bvFTD.
Baseline comparison of Papez white matter structures across patient cohorts indicated
significantly reduced integrity in the fornix and left cingulum for all patient groups, relative to
control (Fig. 3). In contrast, longitudinal comparison of change over 1 year within patient cohorts
was dissociable between patient groups. Both AD and bvFTD showed progressive decline in
fornix integrity, with bvFTD also showing additional decline in the right cingulum. In contrast,
svPPA showed no significant decline to the fornix and cingulum over the 1-year period.
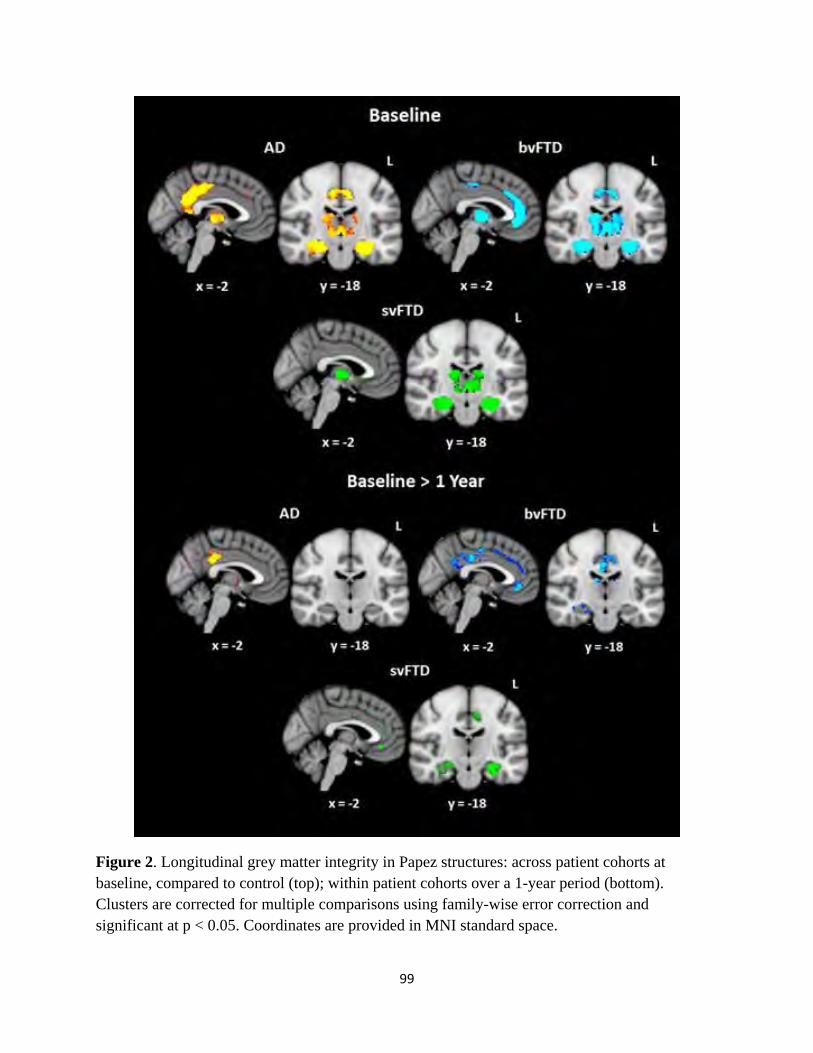
99
Figure 2. Longitudinal grey matter integrity in Papez structures: across patient cohorts at baseline, compared to control (top); within patient cohorts over a 1-year period (bottom). Clusters are corrected for multiple comparisons using family-wise error correction and significant at p < 0.05. Coordinates are provided in MNI standard space.
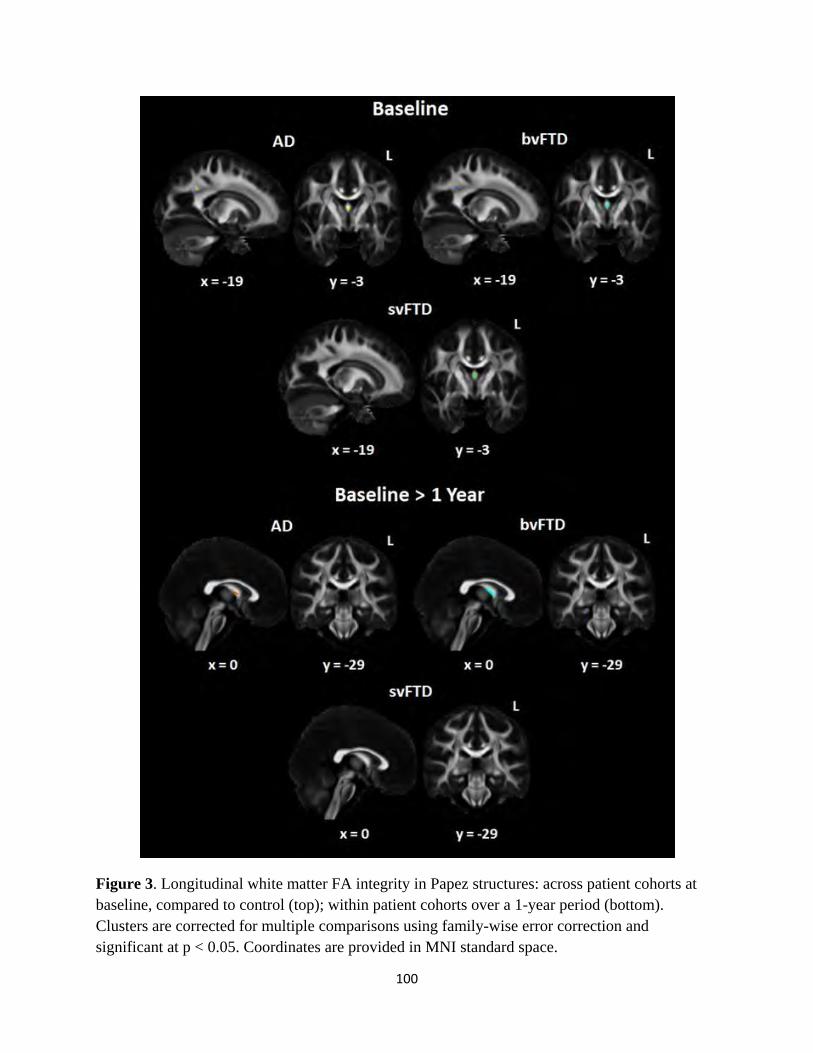
100
Figure 3. Longitudinal white matter FA integrity in Papez structures: across patient cohorts at baseline, compared to control (top); within patient cohorts over a 1-year period (bottom). Clusters are corrected for multiple comparisons using family-wise error correction and significant at p < 0.05. Coordinates are provided in MNI standard space.

101
Discussion
Using VBM and tensor based TBSS, we characterised longitudinal grey and white matter change
along the Papez circuit across AD and FTD patient groups in combination with their clinical
episodic memory profile. Structural imaging revealed dissociable patterns of atrophy to
structures tied critically to memory between patient groups, in particular, the anterior thalamus
and cingulate gyrus. Cognitive measures of verbal and visual episodic recall, however, did not
show significant differences across patient cohorts, with 1-year performance maintained relative
to baseline performance. Rather, decline in general cognition was the only measure to show a
significant decline. These findings highlight the presence of significant subcortical changes,
beyond the hippocampus, present in the early stages of neurodegenerative diseases, and hold
clinical and theoretical implications for episodic memory processing.
The anterior thalamus, a key component of the Papez memory circuit, was found to be
commonly affected in both AD and FTD groups. While the effects of focal damage to the
thalamus, in particular, the anterior and medio-dorsal thalamic nuclei, resulting in delayed
episodic recall is well documented [38,39], this region is often overlooked in regard to memory
performance in dementia. This finding is, however, consistent with recent studies highlighting
the prominence of subcortical atrophy in FTD cases with confirmed pathology or associated
genetic abnormality [4,24,40]. Interestingly, evidence from post-mortem studies suggest that by
the end stage, the anterior thalamus is a prominent marker of disease, correlating with increased
severity of episodic memory deficits and dementia severity in bvFTD and svPPA, respectively
[4,24]. The current in-vivo findings, however, suggest that while a prominent anterior thalamic
involvement is present in the early stage of AD, bvFTD and svPPA, faster deterioration appears
to take place in AD than in the other groups over the 1-year period. Given that the thalamus

102
holds important pathways connecting the hippocampus and prefrontal cortex [41,42], these
findings hold significant implications for long-term contextual memory retrieval, which is
believed to be dependent on hippocampal-prefrontal cortex interaction [39,43]. Specifically,
memory consolidation deficits may be more sensitive than existing clinical measures of delayed
episodic recall in assessing early memory decline.
Another key Papez structure found to show dissociable disease pathology in AD and FTD
is anterior and posterior cingulate gyrus. The cingulate gyrus has been linked to a number of
cognitive processes, in particular the anterior portion with attention and set-shifting [44], and the
posterior portion with spatial memory [6,45]. Hypometabolism in the posterior cingulate is an
early indicator of AD pathology [16,46], while pathological studies in FTD show greater anterior
cingulate involvement in bvFTD [4], and selective neuronal loss in the anterior cingulate in
svPPA [24]. Our findings of divergent pattern of atrophy in the cingulate gyrus mirror previous
metabolic and post-mortem findings. Notably, svPPA did not show significant change in either
anterior or posterior cingulate regions at baseline, relative to controls. Rather, deterioration of the
anterior cingulate occurs after disease onset in svPPA. In contrast, AD and bvFTD show
significant grey matter reduction in the cingulate gyrus at onset and further deterioration with
disease progression. As we demonstrated in a previous study [6], the graded dissociation in
posterior cingulate integrity in AD, bvFTD, and svPPA, can be assessed cognitively through
spatial orientation and improve accuracy of differential dementia diagnoses. This neural
dissociation, however, does not appear to be reflected through standard clinical measures of
memory.
The visual and verbal measures of episodic memory employed in the current study reflect
typical screening procedures for memory performance currently employed in dementia clinics.

103
The lack of significant memory decline in AD and bvFTD over a 1-year period, despite
progressive degeneration of the Papez memory circuit, highlights the need for sensitive measures
targeting specific memory processes rather than general visual and verbal recall. Evidence from
focal lesion studies of the anterior thalamus [38] and posterior cingulate [47] suggests there is
limited, if any, distinction with the amnesic profile resulting from hippocampal damage on
standard measures of episodic recall. Functional neuroimaging, however, suggests that the
anterior thalamus and posterior cingulate play a key role in more specialized memory processes,
namely contextual memory recall and judgements of heading direction, respectively [48,49].
Both structures are anatomically well placed to facilitate cognitive processing dependent on
integrating multi-modal information, and hold dense reciprocal connections with the
hippocampus and virtually all cortical regions of the brain within the Papez circuit [50,51].
The current study provides new insight of the longitudinal impact of early stage AD and
FTD disease pathology on the Papez memory circuit. All efforts were made in the careful
selection of well-defined AD, bvFTD and svPPA cases fulfilling international diagnostic criteria
[1-3] with the aid of experienced neurologists. Despite these exciting findings, some limitations
to our study exist, such as the lack of genetic testing and pathological confirmation of underlying
disease pathology. Further, we could not measure the mammillary bodies, one of the keys
regions for memory processing in the Papez circuit. This was mainly due to the resolution of the
scanning, which does not allow measuring such a small structure.Our findings, however, are
consistent with previous pathological studies [4,24] and employ imaging techniques shown to be
robust for longitudinal analyses [28,33].

104
In conclusion, dissociable patterns to the Papez memory circuit are present in the early
stages of AD and FTD, but are not reflected by longitudinal assessment on conventional
measures of episodic memory. Specific measures of long-term contextual recall and spatial
orientation may offer better tracking of longitudinal memory decline.

105
Acknowledgements
This work was supported by funding to Forefront, a collaborative research group dedicated to the
study of frontotemporal dementia and motor neurone disease, from the National Health and
Medical research Council (NHMRC) of Australia program grant (#1037746), the Australian
Research Council (ARC) Centre of Excellence in Cognition and its Disorders Memory Node
(#CE110001021) and an ARC Discovery Project grant (DP1093279). ST is supported by
Alzheimer’s Australia Dementia Research Foundation and NHMRC of Australia awards. OP is
supported by a NHMRC Career Development Fellowship (APP1022684). MH is supported by
Alzheimer Research UK and the Isaac Newton Trust. These funding sources had no involvement
in the study design, collection, analysis and interpretation of data, writing the manuscript, and in
the decision to submit the manuscript for publication. The authors report no conflict of interest.
We are grateful to the research participants and carers involved with ForeFront research studies.

106
References
[1] McKhann GM, Knopman DS, Chertkow H, Hyman BT, Jack CR,Jr, Kawas CH, Klunk WE,
Koroshetz WJ, Manly JJ, Mayeux R, Mohs RC, Morris JC, Rossor MN, Scheltens P, Carrillo
MC, Thies B, Weintraub S, Phelps CH (2011) The diagnosis of dementia due to Alzheimer's
disease: recommendations from the National Institute on Aging-Alzheimer's Association
workgroups on diagnostic guidelines for Alzheimer's disease. Alzheimers Dement 7, 263-269.
[2] Gorno-Tempini ML, Hillis AE, Weintraub S, Kertesz A, Mendez M, Cappa SF, Ogar JM,
Rohrer JD, Black S, Boeve BF, Manes F, Dronkers NF, Vandenberghe R, Rascovsky K,
Patterson K, Miller BL, Knopman DS, Hodges JR, Mesulam MM, Grossman M (2011)
Classification of primary progressive aphasia and its variants. Neurology 76, 1006-1014.
[3] Rascovsky K, Hodges JR, Knopman D, Mendez MF, Kramer JH, Neuhaus J, van Swieten JC,
Seelaar H, Dopper EG, Onyike CU, Hillis AE, Josephs KA, Boeve BF, Kertesz A, Seeley WW,
Rankin KP, Johnson JK, Gorno-Tempini ML, Rosen H, Prioleau-Latham CE, Lee A, Kipps CM,
Lillo P, Piguet O, Rohrer JD, Rossor MN, Warren JD, Fox NC, Galasko D, Salmon DP, Black
SE, Mesulam M, Weintraub S, Dickerson BC, Diehl-Schmid J, Pasquier F, Deramecourt V,
Lebert F, Pijnenburg Y, Chow TW, Manes F, Grafman J, Cappa SF, Freedman M, Grossman M,
Miller BL (2011) Sensitivity of revised diagnostic criteria for the behavioural variant of
frontotemporal dementia. Brain 134, 2456-2477.
[4] Hornberger M, Wong S, Tan R, Irish M, Piguet O, Kril J, Hodges JR, Halliday G (2012) In
vivo and post-mortem memory circuit integrity in frontotemporal dementia and Alzheimer's
disease. Brain 135, 3015-3025.

107
[5] Davies RR, Hodges JR, Kril JJ, Patterson K, Halliday GM, Xuereb JH (2005) The
pathological basis of semantic dementia. Brain 128, 1984-1995.
[6] Tu S, Wong S, Hodges JR, Irish M, Piguet O, Hornberger M (2015) Lost in spatial translation
- A novel tool to objectively assess spatial disorientation in Alzheimer's disease and
frontotemporal dementia. Cortex 67, 83-94.
[7] Aggleton JP (2014) Looking beyond the hippocampus: old and new neurological targets for
understanding memory disorders. Proc Biol Sci 281, 10.1098/rspb.2014.0565.
[8] King DR, de Chastelaine M, Elward RL, Wang TH, Rugg MD (2015) Recollection-related
increases in functional connectivity predict individual differences in memory accuracy. J
Neurosci 35, 1763-1772.
[9] Irish M, Piguet O, Hodges JR, Hornberger M (2014) Common and unique gray matter
correlates of episodic memory dysfunction in frontotemporal dementia and Alzheimer's disease.
Hum Brain Mapp 35, 1422-1435.
[10] Papez JW (1995) A proposed mechanism of emotion. 1937. J Neuropsychiatry Clin
Neurosci 7, 103-112.
[11] Shah A, Jhawar SS, Goel A (2012) Analysis of the anatomy of the Papez circuit and
adjoining limbic system by fiber dissection techniques. J Clin Neurosci 19, 289-298.
[12] Rudebeck SR, Scholz J, Millington R, Rohenkohl G, Johansen-Berg H, Lee AC (2009)
Fornix microstructure correlates with recollection but not familiarity memory. J Neurosci 29,
14987-14992.

108
[13] Maddock RJ, Garrett AS, Buonocore MH (2001) Remembering familiar people: the
posterior cingulate cortex and autobiographical memory retrieval. Neuroscience 104, 667-76.
[14] Carlesimo GA, Lombardi MG, Caltagirone C (2011) Vascular thalamic amnesia: a
reappraisal. Neuropsychologia 49, 777-789.
[15] Irish M, Piguet O, Hodges JR, Hornberger M (2014) Common and unique gray matter
correlates of episodic memory dysfunction in frontotemporal dementia and Alzheimer's disease.
Hum Brain Mapp 35, 1422-1435.
[16] Nestor PJ, Fryer TD, Ikeda M, Hodges JR (2003) Retrosplenial cortex (BA 29/30)
hypometabolism in mild cognitive impairment (prodromal Alzheimer's disease). Eur J Neurosci
18, 2663-2667.
[17] Lupton MK, Strike L, Hansell NK, Wen W, Mather KA, Armstrong NJ, Thalamuthu A,
McMahon KL, de Zubicaray GI, Assareh AA, Simmons A, Proitsi P, Powell JF, Montgomery
GW, Hibar DP, Westman E, Tsolaki M, Kloszewska I, Soininen H, Mecocci P, Velas B,
Lovestone S, Alzheimer's Disease Neuroimaging Initiative, Brodaty H, Ames D, Trollor JN,
Martin NG, Thompson PM, Sachdev PS, Wright MJ (2016) The effect of increased genetic risk
for Alzheimer's disease on hippocampal and amygdala volume. Neurobiol Aging 40, 68-77.
[18] Wisse LE, Reijmer YD, ter Telgte A, Kuijf HJ, Leemans A, Luijten PR, Koek HL,
Geerlings MI, Biessels GJ, Utrecht Vascular Cognitive Impairment (VCI) Study Group (2015)
Hippocampal disconnection in early Alzheimer's disease: a 7 tesla MRI study. J Alzheimers Dis
45, 1247-1256.

109
[19] Yew B, Alladi S, Shailaja M, Hodges JR, Hornberger M (2013) Lost and forgotten?
Orientation versus memory in Alzheimer's disease and frontotemporal dementia. J Alzheimers
Dis 33, 473-481.
[20] Sexton CE, Mackay CE, Lonie JA, Bastin ME, Terriere E, O'Carroll RE, Ebmeier KP
(2010) MRI correlates of episodic memory in Alzheimer's disease, mild cognitive impairment,
and healthy aging. Psychiatry Res 184, 57-62.
[21] Seeley WW, Crawford R, Rascovsky K, Kramer JH, Weiner M, Miller BL, Gorno-Tempini
ML (2008) Frontal paralimbic network atrophy in very mild behavioral variant frontotemporal
dementia. Arch Neurol 65, 249-255.
[22] Gorno-Tempini ML, Hillis AE, Weintraub S, Kertesz A, Mendez M, Cappa SF, Ogar JM,
Rohrer JD, Black S, Boeve BF, Manes F, Dronkers NF, Vandenberghe R, Rascovsky K,
Patterson K, Miller BL, Knopman DS, Hodges JR, Mesulam MM, Grossman M (2011)
Classification of primary progressive aphasia and its variants. Neurology 76, 1006-1014.
[23] Hodges JR, Graham KS (2001) Episodic memory: insights from semantic dementia. Philos
Trans R Soc Lond B Biol Sci 356, 1423-1434.
[24] Tan RH, Wong S, Kril JJ, Piguet O, Hornberger M, Hodges JR, Halliday GM (2014)
Beyond the temporal pole: limbic memory circuit in the semantic variant of primary progressive
aphasia. Brain 137, 2065-2076.

110
[25] de Souza LC, Chupin M, Bertoux M, Lehericy S, Dubois B, Lamari F, Le Ber I, Bottlaender
M, Colliot O, Sarazin M (2013) Is hippocampal volume a good marker to differentiate
Alzheimer's disease from frontotemporal dementia? J Alzheimers Dis 36, 57-66.
[26] Kipps CM, Hodges JR, Hornberger M (2010) Nonprogressive behavioural frontotemporal
dementia: recent developments and clinical implications of the 'bvFTD phenocopy syndrome'.
Curr Opin Neurol 23, 628-632.
[27] Smith SM (2002) Fast robust automated brain extraction. Hum Brain Mapp 17, 143-155.
[28] Douaud G, Mackay C, Andersson J, James S, Quested D, Ray MK, Connell J, Roberts N,
Crow TJ, Matthews PM, Smith S, James A (2009) Schizophrenia delays and alters maturation of
the brain in adolescence. Brain 132, 2437-2448.
[29] Smith SM, Jenkinson M, Johansen-Berg H, Rueckert D, Nichols TE, Mackay CE, Watkins
KE, Ciccarelli O, Cader MZ, Matthews PM, Behrens TE (2006) Tract-based spatial statistics:
voxelwise analysis of multi-subject diffusion data. Neuroimage 31, 1487-1505.
[30] Mahoney CJ, Simpson IJ, Nicholas JM, Fletcher PD, Downey LE, Golden HL, Clark CN,
Schmitz N, Rohrer JD, Schott JM, Zhang H, Ourselin S, Warren JD, Fox NC (2015)
Longitudinal diffusion tensor imaging in frontotemporal dementia. Ann Neurol 77, 33-46.
[31] Tu S, Leyton CE, Hodges JR, Piguet O, Hornberger M (2015) Divergent Longitudinal
Propagation of White Matter Degradation in Logopenic and Semantic Variants of Primary
Progressive Aphasia. J Alzheimers Dis 49, 853-861.

111
[32] Keihaninejad S, Zhang H, Ryan NS, Malone IB, Modat M, Cardoso MJ, Cash DM, Fox NC,
Ourselin S (2013) An unbiased longitudinal analysis framework for tracking white matter
changes using diffusion tensor imaging with application to Alzheimer's disease. Neuroimage 72,
153-163.
[33] Hui Zhang, Avants BB, Yushkevich PA, Woo JH, Sumei Wang, McCluskey LF, Elman LB,
Melhem ER, Gee JC (2007) High-Dimensional Spatial Normalization of Diffusion Tensor
Images Improves the Detection of White Matter Differences: An Example Study Using
Amyotrophic Lateral Sclerosis. Medical Imaging, IEEE Transactions on 26, 1585-1597.
[34] Varentsova A, Zhang S, Arfanakis K (2014) Development of a high angular resolution
diffusion imaging human brain template. Neuroimage 91, 177-186.
[35] Smith SM, Jenkinson M, Johansen-Berg H, Rueckert D, Nichols TE, Mackay CE, Watkins
KE, Ciccarelli O, Cader MZ, Matthews PM, Behrens TE (2006) Tract-based spatial statistics:
voxelwise analysis of multi-subject diffusion data. Neuroimage 31, 1487-1505.
[36] Smith SM, Nichols TE (2009) Threshold-free cluster enhancement: addressing problems of
smoothing, threshold dependence and localisation in cluster inference. Neuroimage 44, 83-98.
[37] Nichols TE, Holmes AP (2002) Nonparametric permutation tests for functional
neuroimaging: a primer with examples. Hum Brain Mapp 15, 1-25.
[38] Carlesimo GA, Lombardi MG, Caltagirone C (2011) Vascular thalamic amnesia: a
reappraisal. Neuropsychologia 49, 777-789.

112
[39] Tu S, Miller L, Piguet O, Hornberger M (2014) Accelerated forgetting of contextual details
due to focal medio-dorsal thalamic lesion. Frontiers in Behavioral Neuroscience 8.
[40] Cistaro A, Pagani M, Montuschi A, Calvo A, Moglia C, Canosa A, Restagno G, Brunetti M,
Traynor BJ, Nobili F, Carrara G, Fania P, Lopiano L, Valentini MC, Chio A (2014) The
metabolic signature of C9ORF72-related ALS: FDG PET comparison with nonmutated patients.
Eur J Nucl Med Mol Imaging 41, 844-852.
[41] Aggleton JP, Dumont JR, Warburton EC (2011) Unraveling the contributions of the
diencephalon to recognition memory: a review. Learn Mem 18, 384-400.
[42] Xu W, Sudhof TC (2013) A neural circuit for memory specificity and generalization.
Science 339, 1290-1295.
[43] Preston AR, Eichenbaum H (2013) Interplay of hippocampus and prefrontal cortex in
memory. Curr Biol 23, R764-73.
[44] Bissonette GB, Powell EM, Roesch MR (2013) Neural structures underlying set-shifting:
roles of medial prefrontal cortex and anterior cingulate cortex. Behav Brain Res 250, 91-101.
[45] Burgess N (2006) Spatial memory: how egocentric and allocentric combine. Trends Cogn
Sci 10, 551-557.
[46] Minoshima S, Giordani B, Berent S, Frey KA, Foster NL, Kuhl DE (1997) Metabolic
reduction in the posterior cingulate cortex in very early Alzheimer's disease. Ann Neurol 42, 85-
94.

113
[47] Rudge P, Warrington EK (1991) Selective impairment of memory and visual perception in
splenial tumours. Brain 114 ( Pt 1B), 349-360.
[48] Epstein RA, Vass LK (2013) Neural systems for landmark-based wayfinding in humans.
Philos Trans R Soc Lond B Biol Sci 369, 20120533.
[49] Pergola G, Ranft A, Mathias K, Suchan B (2013) The role of the thalamic nuclei in
recognition memory accompanied by recall during encoding and retrieval: an fMRI study.
Neuroimage 74, 195-208.
[50] Wolff M, Alcaraz F, Marchand AR, Coutureau E (2015) Functional heterogeneity of the
limbic thalamus: From hippocampal to cortical functions. Neurosci Biobehav Rev 54, 120-130.
[51] Vann SD, Aggleton JP, Maguire EA (2009) What does the retrosplenial cortex do? Nat Rev
Neurosci 10, 792-802.

114
Chapter 5 – Longitudinal White Matter Degradation in Primary
Progressive Aphasia
Publication V – “Divergent longitudinal propagation of white matter degradation in
logopenic and semantic variants of primary progressive aphasia”

115
Journal of Alzheimer's Disease 49 (201 6) 853-861 DO! 10.3233/JAD-150626 IOS Press
Divergent Longitudinal Propagation of White Matter Degradation in Logopenic and Semantic Variants of Primary Progressive Aphasia
Sicong Tua,b,c, Cristian E. Leytona,b,d, John R. Hodgesa,b,c, Olivier Piguet"·b,c and Michael Hornberger"·c,e,* a Neuroscience Research Australia, Randwick, Sydney, Australia b Australian Research Council Centre of Excellence in Cognition and its Disorders, Sydney, Australia 0 School of Medical Sciences, University of New South Wales, Sydney, Australia d Faculty of Health Sciences, University of Sydney, Sydney, Australia eDepartment of Clinical Neurosciences, University of Cambridge, Cambridge, UK
Handling Associate Editor: Sharon Naismith
Accepted 15 September 2015
Abstract.
853
Background: Clinico-pathological distinction of primary progressive aphasia (PPA) can be challenging at clinic presentation. In particular, cross-sectional neuroimaging signatures across the logopenic (lvPPA) and semantic (svPPA) variants are difficult to establish, with longitudinal profiles showing greater divergence. Objective: Assess longitudinal propagation of white matter degradation in lvPPA and svPPA to determine disease progression over time, and whether this refiects distinct underlying pathology. Method: A cohort of27 patients with dementia (12lvPPA; IS svPPA) and 12 healthy controls were assessed at baseline and !-year follow-up on the Addenbrooke's Cognitive Examination-Revised and Sydney Language Battery. Diffusion weighted images were collected at both time-points and analyzed for longitudinal wltite matter change using DTI-TK and TBSS. Results: LvPPA patients showed a significant decline in naming and repetition, over I year, wltile svPPA patients declined in naming and comprehension. Longitudinal imaging revealed widespread bilateral degradation of wbite matter tracts in lvPPA over a !-year period with early involvement of the left posterior inferior longitudinal fasciculus (ILF). SvPPA demonstrated focal left lateralized wltite matter degradation involving the uncinate fasciculus (UF) and anterior ILF, propagating to the right UF with disease progression. Conclusions: LvPPA and svPPA cohorts showed distinct longitudinal cognitive and wltite matter profiles. We propose differences in multi-centricand focal white matter dysfunction in lvPPA and svPPA, respectively, refiect underlying pathological differences. Tbe clinical relevance of white matter degradation and mechanisms underlying disease propagation are discussed.
Keywords: Diffusion tensor imaging, frontotemporal dementia, primary progressive aphasia, white matter
*Correspondence to: Dr. Michael Hornberger, Department of Clinical Neurosciences, University of Cambridge, Cambridge CB2 OSZ, UK. Tel.: +44 1223 760694; E-mail: mh486@medschl. cam.ac.uk.
INTRODUCTION
Primary progressive aphasias (PPA) are a clinicopathological heterogeneous group of neurodegenerative conditions characterized by the progressive breakdown of language skills and associated brain networks [1]. PPA comprises three distinct clinical
ISSN 1387-2877116/$35.00 © 2016- !OS Press and the authors. All rights reserved

116
854 S. Tu et al. I Divergent White Matter Degradation in PPA
syndromes: logopenic (lvPPA), semantic (svPPA), and nonfluent (nfvPPA) variants [1, 2]. Clinical classification into PPA variants occurs at three levels: an initial clinical diagnosis determined by neuropsychological testing, imaging supported structural change, and finally pathological diagnosis either at postmortem or via in vivo PET ligands. Cognitively, lvPPA, svPPA, and nfvPPA variants are characterized by core language deficits involving repetition, comprehension, and agrammatism, respectively [1]. Structural neuroimaging studies also highlight distinct, primarily left hemispheric, patterns of atrophy in temporal and frontal lobe structures, across PPA variants [1-6].
Pathological diagnosis across all PPAs is less clear with the presence of tau and TAR-DNAbinding protein 43 (TDP-43) pathological subtypes of frontotemporal lobar degeneration (FTLD) [7], and Alzheimer's disease (AD) pathology reported in each variant [8-10]. Of all PPA subtypes, lvPPA and svPPA appear to show the most homogenous clinico-pathological mapping, attributed to either AD or FTLD-TDP, respectively [8]. The impact of this distinction in pathology can be clearly seen in the localization and degree of structural abnormality in lvPPA and svPPA. Notably, lvPPA show widespread cortical atrophy to posterior regions of the brain, predominantly affecting the left temporo-parietal junction region, while svPPA show focal change associated with the anterior temporal lobes [2, 3]. However, crosssectional studies comparing white matter integrity in lvPPA and svPPA cohorts often fail to show such a distinction [11, 12], likely due to wide distributed white matter changes seen in lvPPA.
The current study investigates whether longitudinal white matter assessment allows a better distinction of clinicallvPPA and svPPA. Importantly, a subgroup of patients had confirmed amyloid in vivo biomarkers to allow pathological confirmation. We hypothesized that differences in disease pathology can be detected using a longitudinal analysis approach, and, in particular, patients with lvPPA will show white matter changes across multiple white matter tracts, relative to svPPA, which will show highly localized white matter changes in the temporal lobes only.
METHOD
Participants
Twenty-seven dementia patients (15 svPPA; 12 lvPPA) and 12 healthy controls were recruited from the Sydney frontotemporal dementia research group
(FRONTIER) clinical database. All participants underwent an initial assessment and a 12 ± 2-month follow-up assessment. At each visit, patients underwent a comprehensive assessment, which included a clinical interview, neurological examination, cognitive assessment, and whole-brain MRI. All patients met current clinical diagnostic criteria for probable lvPPA and svPPA [1], and diagnosis was established by consensus among the neurologist, neuropsychologist, and occupational therapist. All participants' scans were visually inspected to rule out abnormalities other than atrophy. Participants showing evidence of cerebrovascular disease, including marked small vessel disease, were excluded. Amyloid-13 deposition using Pittsburgh compound B (PiB) PET was assessed in a subset of PPA patients (9 lvPPA, 6 svPPA). AlllvPPA patients scanned were found to be PiE-positive, while svPPA were all negative with the exception of 1 case, which was excluded from analyses. Patient cohorts were matched for age, education, disease duration, and functional disease severity using the Frontotemporal Dementia Rating Scale (FRS) [13] (Table 1). Patients with a change in diagnosis, atypical features, or absence of progressive language impairment were excluded from the study. Healthy controls were matched for education and all scored above 881100 on the Addenbrooke's Cognitive Examination-Revised (ACE-R) screening of general cognition [14]. Wholebrain MRI scans were visually assessed for structural and white matter abnormalities; none were reported in this group.
Cognitive assessment
All participants were administered the ACE-R as a general screening of cognition assessing: attention, verbal fluency, visuospatial ability, memory, and language [14]. Patients' language and speech impairment were comprehensively examined using the Sydney Language Battery (SYD-BAT) [15]. The SYD-BAT consists of 30 test words, each representing an imageable noun, 3 or more syllables in length. Each word was assessed for comprehension, naming, repetition, and semantic association, to provide a comprehensive assessment of language and speech impairment. Performance on the ACE-Rand SYD-BAT were determined at initial and follow-up assessments.
Imaging acquisition
All participants underwent whole-brain T1 and serial diffusion weighted imaging using a 3T Philips
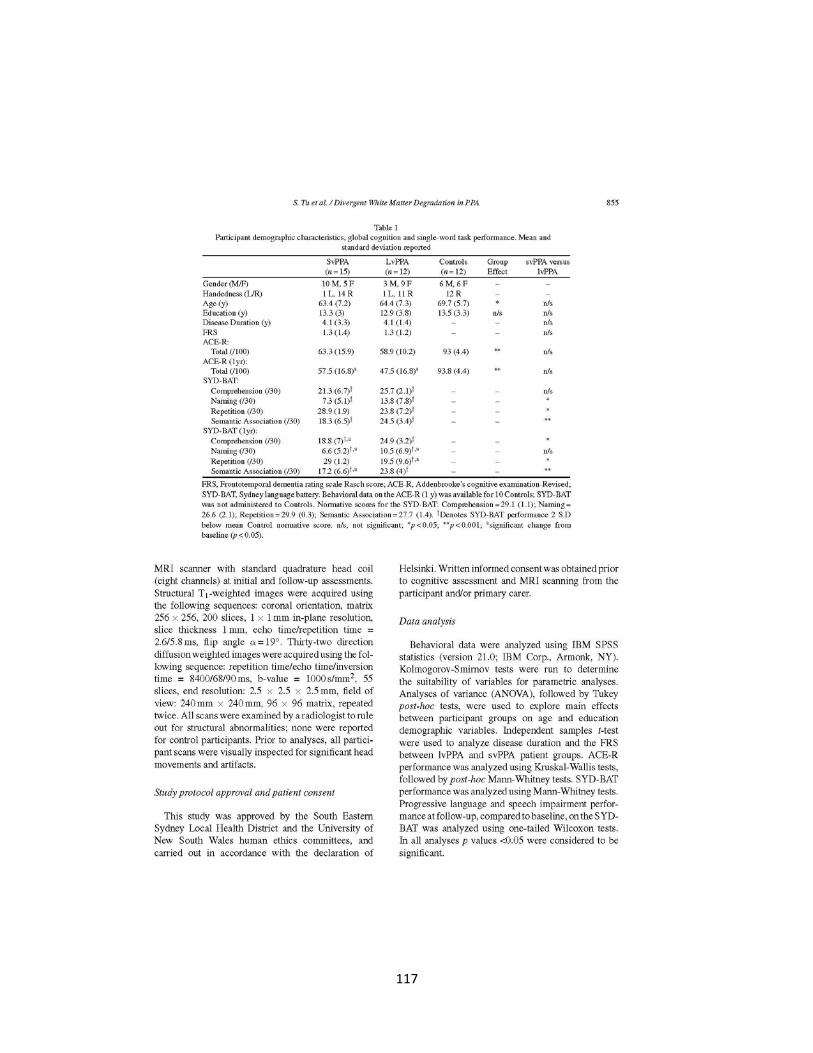
117
S. Tu et al. I Divergent White Matter Degradation inPPA 855
Table I Participant demographic characteristics, global cognition and single-word task performance. Mean and
standard deviation reported
SvPPA (n~ 15)
Gender (MIF) 10M,5F Handedness (LIR) IL, 14R Age(y) 63 .4 (7.2) Ed ucation (y) 13.3 (3) Disease Duration (y) 4.1 (3.3) FRS 1.3 (1.4) ACE-R:
Total (/100) 63.3 (15.9) ACE-R (l yr):
Total (/1 00) 57.5 (16.8)' SYD-BAT:
Comprehension (/30) 21.3 (6 7)1 Naming (/30) 7.3 (5.1)1 Repetition (/30) 28.9(1.9) Semantic Association (/30) 18.3 (6 5)1
SYD-BAT (lyr): Comprehension (/30) 18.8 (7)1·· Naming (/30) 6.6 (5.2)1·•
Repetition (/30) 29 (1.2) Semantic Association (/30) 17.2 (6.6)1·•
LvPPA (n~ 12)
3M,9F IL, l!R
64.4 (7.3) 12.9 (3.8) 4 .1 (1.4) 1.3 (1.2)
58.9 (10.2)
47.5 (16.8)'
25.7 (2.1)1 13.8 (7 .8)1 23.8 (7.2)1 24.5 (3.4)1
24.9 (3 .2)1 10.5 (6.9)1·• 19.5 (9.6)1·• 23.8 (4)1
Controls (n~ 12)
6M,6F 12 R
69.7 (5.7) 13.5 (3.3)
93 (4.4)
93.8 (4.4)
Group svPPA versus Effect I vPPA
n/s n/s n/s
n/s n/s
n/s
n/s
n/s
n/s
FRS, Frontotemporal dementia rating scale Rasch score; ACE-R, Addenbrooke 's cognitive examination-Revised; SYD-BAT, Sydneylanguagebattery. Behavioral data on the ACE-R (l y) was available for 10 Controls; SYD-BAT was not administered to Controls . Normative scores for the SYD-BAT: Comprehension =29 .1 ( 1.1); Naming = 26.6 (2.1); Repetition~29.9 (0.3); Semantic Association~27.7 (1.4). !Denotes SYD-BAT performance 2 S.D below mean Control normative score. n/s, not significant; *p <0.05; **p < 0.001; asignificant change from baseline (p <0.05).
MRI scanner with standard quadrature head coil (eight channels) at initial and follow-up assessments. Structural T 1-weighted images were acquired using the following sequences: coronal orientation, matrix 256 x 256, 200 slices, 1 x 1 mm in-plane resolution, slice thickness 1 mm, echo time/repetition time = 2.6/5.8 ms, flip angle a = 19°. Thirty-two direction diffusion weighted images were acquired using the following sequence: repetition time/echo time/inversion time = 8400/68/90ms, b-value = 1000 s/mm2, 55 slices, end resolution: 2.5 x 2.5 x 2.5 mm, field of view: 240 mm x 240 mm, 96 x 96 matrix, repeated twice. All scans were examined by a radiologist to rule out for structural abnormalities; none were reported for control participants. Prior to analyses, all participant scans were visually inspected for significant head movements and artifacts.
Study protocol approval and patient consent
This study was approved by the South Eastern Sydney Local Health District and the University of New South Wales human ethics committees, and carried out in accordance with the declaration of
Helsinki. Written informed consent was obtained prior to cognitive assessment and MRI scanning from the participant and/or primary carer.
Data analysis
Behavioral data were analyzed using IBM SPSS statistics (version 21.0; IBM Corp., Armonk, NY). Kolmogorov-Smirnov tests were run to determine the suitability of variables for parametric analyses. Analyses of variance (ANOVA), followed by Tukey post-hoc tests, were used to explore main effects between participant groups on age and education demographic variables. Independent samples t-test were used to analyze disease duration and the FRS between lvPPA and svPPA patient groups. ACE-R performance was analyzed using Kruskal-Wallis tests, followed by post-hoc Mann-Whitney tests. SYD-BAT performance was analyzed using Mann-Whitney tests. Progressive language and speech impairment performance at follow-up, compared to baseline, on the S YOBAT was analyzed using one-tailed Wilcoxon tests. In all analyses p values <0.05 were considered to be significant.

118
856 S. Tu etal. I Divergent White Mauer Degradation ill PPA
Imaging analysis
Differences in whole-brain fractional anisotropy (FA) were examined for mean group changes at baseline and follow-up, and longitudinally for interindividual change across time. TBSS [17] was used to perform a skeleton-based analysis of white matter FA, complemented by DTI-TK to provide a tensorbased registration for analyses [ 16]. Tensor based registration of diffusion weighted data is superior to traditional scalar based registration methods in constructing an unbiased spatially normalized brain atlas [ 18], with good reproducibility of diffusion metrics [19]. Serial diffusion-weighted sequences from each participant were averaged to improve signal-to-noise ratio before being COJTected for eddy current distortions. A binary brain mask was generated [20] from the non-diffusion volume (bo) and used to fit a tensor model onto diffusion-weighted images [21] . A wholebrain diffusion tensor image from each participant was used to generate study-specific brain atlases in DTI-TK. Group atlases were constructed through an iterative process of initial rigid registration, followed by non-linear registration, to determine the best mapping with each individual's scan using the least amount of deformation [16, 19]. For the longitudinal analysis, each individual's initial and follow-up scan was co-registered to generate a within-subject template representing a halfway transformation between the two timepoints, prior to construction of a group atlas [19]. Group atlases were registered to the Illinois Institute of Technology standard brain atlas, version 4.0 [22]. Transformation matrices were generated for each stage of registration to create a deformation field that defines the mapping directly from each individual's native scan space to standard space in a s ingle interpolation [1 9]. Individual FA maps are then generated, concatenated, and skeletonized [ 17] to define the lines of maximum FA, which correspond to the centers of white matter tracts. Group FA skeletons were tested for significant differences using voxel-wise general linear modeling via permutation-based non-parametric testing [23] with 5000 permutations per contrast. Axial and radial diffusivity were also assessed in the same manner. Age was included as a nuisance variable in these analyses. Reported clusters were threshold-free cluster enhancement (TFCE) corrected for multiple comparisons at p < 0.05. Delineation of white matter tracts implicated in significant clusters was performed with reference to Johns Hopkins University tractography atlas. Anatomically specific masks of the uncinate fasciculus (UF), inferior longitudinal fasciculus (ILF), and superior
longitudinal fasciculus (SLF) were generated from the atlas using a threshold of 30%, and visually inspected to ensure appropriate coverage. Using these masks, DTI metrics were extracted across both hemispheres, at baseline and follow-up, for atlas-based voxel-wise region of interest (ROI) analyses in patient and control cohorts, using repeated measures ANOVA in SPSS.
RESULTS
Denwgraphics and cognitive testing
PPA patient groups were well matched for age, education, disease duration, and functional severity (FRS) (Table 1; p values >0.87). The control cohort was older than both patients groups (both p values <0.05). Screening of general cognition (ACE-R: Total) indicated significant group differences (p < 0.001) with controls performing the highest. PPA patient groups did not show a significant difference in overall cognitive performance at baseline (p = 0.51) or 1-year follow-up (p = 0. 14 ), but did show a significant decline over a 1-year period (p values <0.02). Control participants did not show a decline in general cognition in the same period (p = 0.37).
PPA patient groups showed dissociable patterns of performance on language specific tasks, namely, lvPPA scored higher than svPPA on single-word comprehension, naming, and semantic association, but not repetition (Table 1). Overall pattern of impairment for single-word language tasks and performance on the SYD-BAT is consistent with the literature, and all patients performed 2 standard deviations below normative control performance, except for svPPA on single-word repetition [15]. At baseline, significant differences in language impairment was observed for naming, repetition, and semantic association (p values <0.05), but not comprehension (p = 0.13). At followup, however, significant differences were found for comprehension, as well as repetition and semantic association (p values <0.05), but not naming (p = 0.14 ). This change can be explained by the distinct pattern of progressive impairment on the SYD-BAT subscales between lvPPA and svPPA patients over the 1-year period. PPA patient groups showed dissociable pattern of longitudinal change on SYD-BAT subscales, with a significant decline in repetition for lvPPA (p<0.001), while comprehension and semantic association both declined in svPPA (p values <0.05). In contrast, naming performance on the SYD-BAT worsened significantly in both patient groups over the 1-year period (p values <0.05).
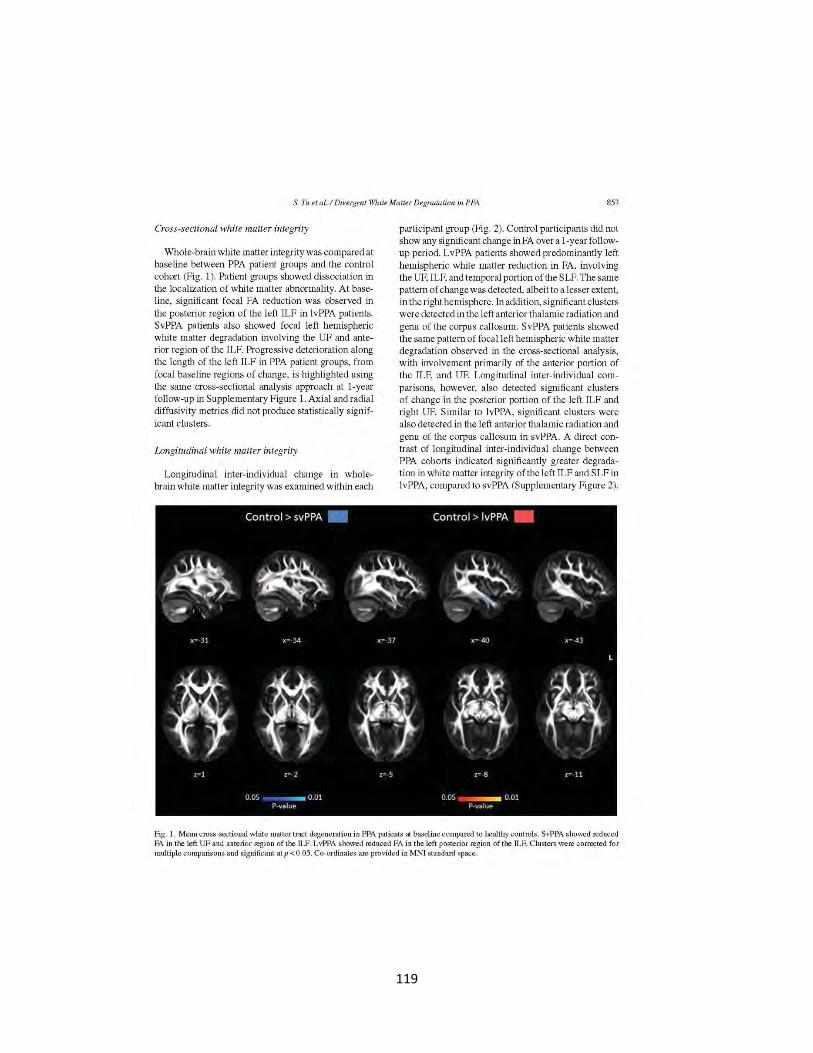
119
S. Tu eta!. I Divergent White Matter Degradation in PPA 857
Cross-sectional while matter integrity
Whole-brain white matter integrity was compared at baseline between PPA patient groups and the control cohort (Fig. 1). Patient groups showed dissociation in the localization of white matter abnormality. At baseline, significant focal FA reduction was observed in the posterior region of the left ILF in lvPPA patients. SvPPA patients also showed focal left hemispheric white matter degradation involving the UF and anterior region of the ILF. Progressive deterioration along the length of the left ILF in PPA patient groups, from focal baseline regions of change, is highlighted using the same cross-sectional analysis approach at 1-year follow-up in Supplementary Figure 1. Axial and radial diffusivity metrics did not produce statistically significant clusters.
Longitudinal white mntter integrity
Longitudinal inter-individual change in wholebrain white matter integrity was examined within each
participant group (Fig. 2). Control participants did not show any significant change in FA over a 1-year followup period. LvPPA patients showed predominantly left hemispheric white matter reduction in FA, involving the UF, ILF, and temporal portion of the S LF. The same pattern of change was detected, albeit to a lesser extent, in the right hemisphere. In addition, significant clusters were detected in the left anterior thalamic radiation and genu of the corpus callosum. SvPPA patients showed the same pattern of focal left hemispheric white matter degradation observed in the cross-sectional analysis, with involvement primarily of the anterior portion of the ILF, and UF. Longitudinal inter-individual comparisons, however, also detected significant clusters of change in the posterior portion of the left ILF and right UF. Similar to lvPPA, s ignificant clusters were also detected in the left anterior thalamic radiation and genu of the corpus callosum in svPPA. A direct contrast of longitudinal inter-individual change between PPA cohorts indicated significantly greater degradation in white matter integrity of the left !LF and SLF in lvPPA, compared to svPPA (Supplementary Figure 2).
Fig. 1. Mean cross-sectional white matter tract degeneration i n PPA patients at baseline compared to healthy controls. SvPPA s howed reduced FA in the left UF and anterior region of the ILF. LvPPA showed reduced FA in the left posterior region of the ILF. Clusters were corrected for multiple comparisons and s ignificant at p < 0.05. Co-ordinates are provided in MNI standard space.
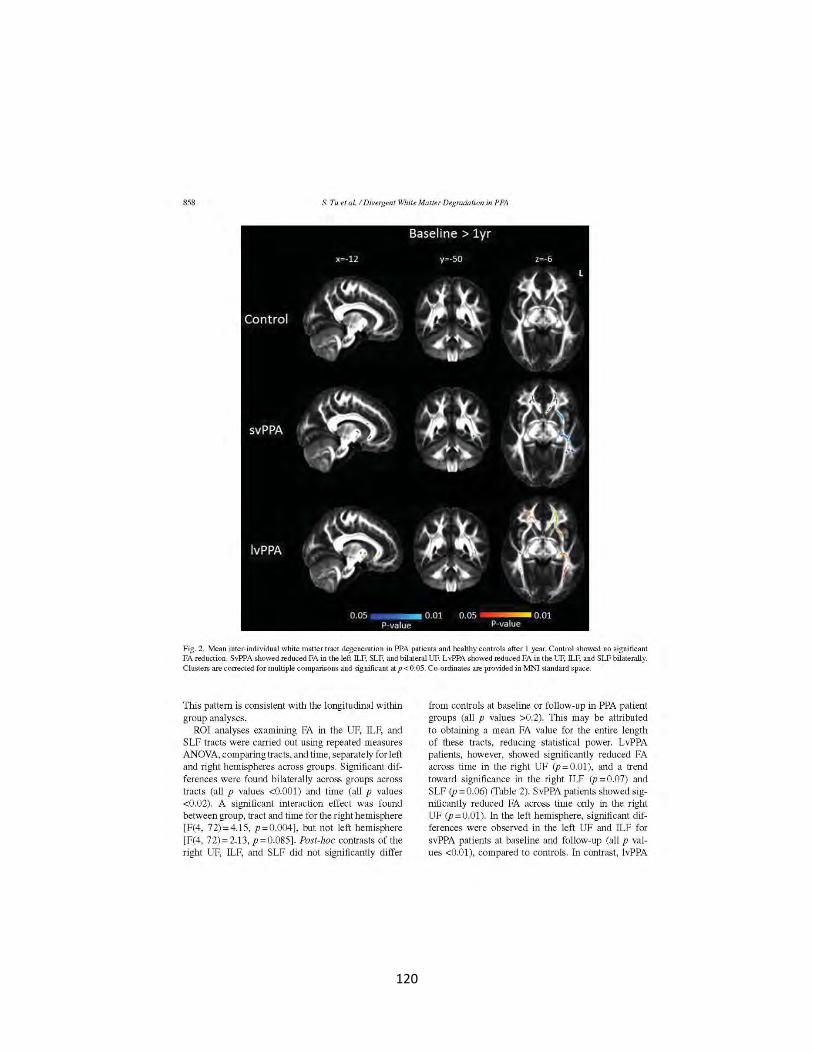
120
858 S. Tu et al.l Divergent White Matter Degradation in PPA
Fig . 2. Mean inter· individual white matter tract degeneration in PPA patients and healthy controls after 1 year. Control showed no significant FA reduction. SvPPA showed reduced FA io the left ILF, SLF, and bilateral UF. L vPPA showed reduced FA in the UF, ILF, and SLF bilaterally. Clusters are corrected for multiple comparisons and significant at p < 0.05. Co-ordinates are provided in MNl standard space.
This pattern is consistent with the longitudinal within group analyses.
ROI analyses examining FA in the UF, ILF, and SLF tracts were carried out using repeated measures AN OVA, comparing tracts, and time, separately for left and right hemispheres across groups. Significant differences were found bilaterally across groups across tracts (all p values <0.001) and time (all p values <0.02). A significant interaction effect was found between group, tract and time for the right hemisphere [F(4, 72)=4.15, p =0.004], but not left hemisphere [F(4, 72) = 2.13, p = 0.085]. Post-hoc contrasts of the right UF, ILF, and SLF did not significantly differ
from controls at baseline or follow-up in PPA patient groups (all p values >0.2). This may be attributed to obtaining a mean FA value for the entire length of these tracts, reducing statistical power. LvPPA patients, however, showed significantly reduced FA across time in the right UF (p = 0.01), and a trend toward significance in the right ILF (p = 0.07) and SLF (p = 0.06) (fable 2). SvPPA patients showed significantly reduced FA across time only in the right UF (p=0.01). In the left hemisphere, significant differences were observed in the left UF and ILF for svPPA patients at baseline and follow-up (all p values <0.01), compared to controls. In contrast, lvPPA

121
S. Tu et al. I Divergent White Matter Degradation inPPA 859
Table2 Fractional anisotropy of the uncinate fasciculus, inferior longitudinal fasciculus, and superior longitudinal fasciculus tracts in healthy controls and PPA groups, at baseline and 1 year. Mean and standard
deviation reported
Baseline FA
UF ILF SLF
Control Left Hemisphere 0.39 (0.03) 0.42 (0.05) 0.44 (0.03) Rig ht Hemisphere 0.42 (0.04) 0.5 (0.04) 0.45 (0.03)
svPPA Left Hemisphere 0.31 (0.05)' 0.37 (O.Q4)' 0.44 (0.03) Right Hemisphere 0.43 (0.05) 0.51 (0.03) 0.46 (0.04)
lvPPA Left Hemisphere 0.39 (0.04) 0.4 (0.03) 0.42 (0.03) Right Hemisphere 0.43 (0.05) 0.49 (0 .04) 0.46 (0.03)
I Year UF ILF SLF
Control Left Hemisphere 0.39 (0.03) 0.43 (0.04) 0.44 (0.03) Right Hemisphere 0.42 (0.03) 0.5 (0.04) 0.46 (0.03)
svPPA Left Hemisphere 0.28 (O.OS)~b 0.36 (0.04)'-b 0.43 (0.04)b Right Hemisphere 0.4 (0.06)b 0.51 (0.05) 0.46 (0.04)
lvPPA Left Hemisphere 0.38 (0.04)b 0.38 (0.03)'·b 0.42 (0.03) Right Hemisphere 0.41 (0.04)b 0.48 (0.04) 0.45 (0.03)
UF, uncinate fasciculus; ILF, inferior longitudinal fasciculus; SLF,
superior longitudinal fasciculus. Significant atp < 0.05; 3 significant change compared to control; bsignificant change from baseline.
patients only showed significant differences in the left ILF at follow-up (p = 0.03), compared to controls. Across time, svPPA showed significantly reduced FA in the left UF, ILF, and SLF (allp-values <0.01), while lvPPA showed significant reduction in the left UF and ILF (all p-values <0.05).
DISCUSSION
This study examined longitudinal white matter integrity in lvPPA and svPPA with a focus on the pattern of change over a 1-year period. We uncovered different patterns in the degree of disease propagation in these patient groups, with lvPPA showing relatively diffuse white matter changes, compared to the focal changes observed in svPPA. Both PPA patient groups showed deterioration predominantly in the left hemispheric UF, ILF, and SLF, with additional involvement of their homologous tracts in the contralateral hemisphere. Clinically, the lvPPA and svPPA cohorts were well defined showing clear progressive decline on cognitive tasks and dissociation in degree of impaired speech production and comprehension, respectively. The divergent course of white matter degeneration in lvPPA and svPPA cohorts highlight the possibility of
utilizing advanced longitudinal neuroimaging to better clarify underlying pathology in vivo.
Degradation of the left UF and ILF has consistently been implicated as the main white matter tracts affected across the PPA spectrum [11, 24], with some evidence of SLF involvement [25, 26]. These tracts have also been identified as being critical components of the language network [12, 25, 27], in particular speech production and semantic retrieval [27, 28]. A recent cross-sectional study dissociated patterns of white matter change in lvPPA and svPPA [11]. Specifically, lvPPA demonstrate widespread left lateralized involvement of the UF and ILF while in svPPA white matter change is predominantly found in ventral tracts, involving bilateral UF and left ILF. Findings from the current study are consistent with the signature of white matter change in svPPA [11, 26]. Longitudinal imaging, however, revealed a complex pattern of progressive white matter degradation in lvPPA, in particular, dissociating from svPPA. Cross-sectional analyses at baseline demonstrated that early white matter pathology in lvPPA tend to be circumscribed to the posterior tail of the left ILF. Over a 12-month period, however, our analyses showed that changes became widespread, affecting the entire length of the left ILF and involving the UF and SLF in this group. These analyses further demonstrated a similar pattern of white matter changes in the right hemisphere, albeit to a lesser extent. These distinct differences in localization and laterality of white matter abnormality allude to divergent underlying pathology and their propagation in the logopenic and semantic variants of PPA.
The focal propagation of white matter dysfunction in svPPA followed a stereotypical pattern along anatomical tracts, suggesting that pathology spreads throughout neighboring axonal tracts [29]. In contrast, lvPPA showed a diffuse white matter involvement displaying a less stereotyped pattern of progression that not necessarily follows the trajectory of axonal bundles. This divergent pattern of white matter dysfunction mirrors extensive structural imaging and pathological evidence that shows widespread brain atrophy in lvPPA and circumscribed atrophy in svPPA [3, 30], and suggests fundamental mechanistic differences in how pathology spreads. Whereas the primary event driving white matter involvement in svPPA is axonal death (i.e., Wallerian degeneration) derived from focal neuronal death [29], the pattern of white matter involvement in lvPPA seems to be different. Our findings suggest a combined spreading characterized by eccentric white matter changes over time associated with distant multi-centric involvement. This

122
860 S. Tu et al. I Divergent White Matter Degradation in PPA
pattern suggests that pathology spreads in two different fashions: directly from neuron to neuron throughout axonal bundles, as well as multiple distant pathological seeds that do not necessarily follow anatomical connections. This pattern highlights the central role of extra-neuronal factors in lvPPA, in particular, the neurotoxic effect of amyloid-13. Interestingly, white matter dysfunction was also detected in the genu of the corpus callosum, perhaps presenting an avenue for transcallosal propagation. Further research is needed to investigate the differential regional effect of amyloid and neuronal vulnerability to clarify this issue.
White matter change is increasingly being used to address one of the prevalent issues in the field that remains unresolved from decades of structural imaging, which is the accurate prediction of pathology in vivo. Although important advances in the development of biomarkers have taken place in recent years, clinicopathological correlations at an individual level remain elusive. This issue is particularly acute in PPA cases due to AD, a pervasive pathology that can adopt varied neurocognitive patterns and is often present in unclassified PPA cases due to mixed deficits [31]. In this sense, white matter changes over time can provide a complementary account that can assist in the identification of AD cases. Current findings suggest the posterior tail of the left ILF is a sensitive marker of white matter change in the early stages of the disease.
In the current study, efforts were made in the careful selection of well-defined PPA cases fulfilling diagnostic criteria [1] with the aid of experienced neurologists. A limitation is, however, the lack of genetic testing and pathological confirmation of underlying disease pathology. PiB PET imaging was included as a proxy measure of AD pathology and was available for the majority of lvPPA patients, but only half of svPPA patients. For PPA patients with available PiB imaging, however, alllvPPA were found to be PiE-positive while svPPA were PiE-negative, consistent with expected pathology [8] and interpretation of current imaging findings. All patient and control follow-up assessments occurred within a 2-month margin of the 1-year timepoint, representing genuine longitudinal cognitive and neural change. Robust longitudinal imaging analyses were carried out using tensor-based registration for accurate registration of tract orientation [16] and localization of white matter changes. Current findings should, however, be replicated in a larger sample of PPA patients for reliability.
In conclusion, logopenic and semantic variants of PPA demonstrate marked behavioral and neural profiles resulting in progressive deterioration of distinct
neural networks. Modeling the progression of white matter degradation along tracts provides new insight into the molecular mechanisms underlying disease pathology.
ACKNOWLEDGMENTS
This work was supported by funding to Forefront, a collaborative research group dedicated to the study of frontotemporal dementia and motor neuron disease, from the National Health and Medical research Council (NHMRC) of Australia program grant (#1037746), the Australian Research Council (ARC) Centre of Excellence in Cognition and its Disorders Memory Node (#CE110001021) and an ARC Discovery Project grant (DP1 093279). ST is supported by Alzheimer's Australia Dementia Research Foundation and NHMRC of Australia awards. CEL is supported by DVC post-doctoral fellowship of the University of Sydney (S0716/U2644). OP is supported by a NHMRC Career Development Fellowship (APP1022684). MH is supported by Alzheimer Research UK and the Isaac Newton Trust. These funding sources had no involvement in the study design, collection, analysis and interpretation of data, writing the manuscript, and in the decision to submit the manuscript for publication. We are grateful to the research participants and carers involved with ForeFront research studies.
Authors' disclosures available online (http://j-alz. com/manuscript-disclosures/15-0626r1).
SUPPLEMENTARY MATERIAL
The supplementary material is available in the electronic version of this article: http:l/dx.doi.org/10.3233/ JAD-150626.
REFERENCES
(I] Gorno-Tempini ML, Hillis AE, Weintraub S, Kertesz A, Mendez M, Cappa SF, Ogar JM, Rohrer JD, BlackS, Boeve BF, Manes F, Dronkers NF, Vandenberghe R, Rascovsky K, Patterson K, Miller BL, Koopman DS, Hodges JR, Mesularn MM, Grossman M (2011) Classification of primary progressive aphasia and its variants. Neurology 76, 1006-1014.
[2] Gorno-Tempini ML, Dronkers NF, Rankin KP, Ogar JM, Phengrasamy L, Rosen HJ, Johnson JK, Weiner MW, Miller BL (2004) Cognition and anatomy in three variants of primary progressive aphasia. Ann Neurol55, 335-346.
[3] Rohrer JD, Ridgway GR, Crutch SJ, Hailstone J, Goll JC, Clarkson MJ, Mead S, Beck J, Mummery C, Ourselin S, Warrington EK, Rossor M:N, Warren JD (20 10) Progressive

123

124
Chapter 6 – Conclusion
The findings from this thesis show that the unique differential changes present in the neural
circuitry underlying spatial and episodic memory processes in neurodegenerative conditions can
be assessed cognitively. The longitudinal in-vivo imaging findings confirm there is dissociation
in anterior and posterior components of the Papez circuit in AD and FTD, consistent with recent
pathological studies (Hornberger et al., 2012; Tan et al., 2014). This dissociation in memory
circuit integrity highlights the potential for targeted cognitive measures, specific to the function
of discrete neural structures that show divergent patterns of atrophy in the early stages of
dementia pathologies. Notably, the virtual supermarket task of heading orientation and the long-
term contextual memory retrieval task are two novel and clinically feasible tasks that can be
easily applied with the potential to improve differential diagnostic accuracy of young onset
dementias. Findings from the virtual supermarket task are the first to concurrently examine
spatial orientation in AD and bvFTD, and demonstrate its merits for inclusion in dementia
screening. Furthermore, the finding that neural correlates beyond the hippocampus underlie
performance on spatial and long-term episodic memory provides continued insight for the
current direction of memory research aimed at exploring new neurological targets for
understanding clinical memory disorders (Aggleton, 2014). Findings from the experimental
studies in this thesis indicate that a greater emphasis should be placed on trying to understand the
changes affecting the posterior cingulate and anterior thalamus in neurodegenerative diseases.

125
The posterior cingulate and anterior thalamus are vulnerable regions underlying memory
impairments in AD and FTD: The posterior cingulate and anterior thalamus are both neural
hubs with strong connectivity to deep grey matter structures and higher-order cortical regions of
the brain. The findings demonstrate, in-vivo, that both structures are susceptible to degeneration
in the early stages of AD and FTD, and play a critical role in spatial and episodic memory
processing. Specifically, the retrosplenial cortex, located at the tail of the posterior cingulate, is a
focal region uniquely affected in AD, but preserved in FTD. In Chapter 2 it was demonstrated,
for the first time, a positive association between retrosplenial cortex volume and spatial
orientation performance in AD. Whilst not particularly surprising, in itself, this finding is
important given existing converging evidence from the human and animal literature, supporting
the role of retrosplenial cortex in integrating multi-modal sensory information (visual, motor)
and internal spatial map to determine heading direction during spatial navigation (Irish et al.,
2015; Vann, Aggleton, & Maguire, 2009). Using this information, an ecologically valid novel
cognitive measure of spatial orientation was implemented, the virtual supermarket task, which
demonstrated high diagnostic accuracy in differential diagnosis of AD and FTD. Development of
this task was aimed at providing a targeted assessment of a specific cognitive deficit in AD,
spatial disorientation, rather than more generalized spatial navigation or visuospatial episodic
memory recall tasks that draw more heavily upon the hippocampus (Epstein & Vass, 2013; Kim,
2016).
The anterior thalamus was another key structure found to be affected in the early stages
of both AD and FTD. In relation to memory, amnesia resulting from focal damage to the
thalamus is well documented in thalamic stroke, in particular when the anterior thalamic and
medio-dorsal nuclei are affected (Carlesimo, Lombardi, & Caltagirone, 2011; Tu, Miller, Piguet,

126
& Hornberger, 2014). Nevertheless, thalamic amnesia is viewed as a disruption in relay of
hippocampal information rather than driving amnesic symptoms, and overlooked in memory
associated disorders, such as AD and FTD. This is in part due to long-standing cognitive models
of long-term memory placing significant emphasis on hippocampal-cortical interaction, but
neglecting the role and impact of disruption to intermediary neural pathways necessary to
facilitate this communication (Preston & Eichenbaum, 2013; Squire, Clark, & Knowlton, 2001;
Winocur, Moscovitch, & Bontempi, 2010). Using a novel long-term contextual memory recall
task, Chapter 3 demonstrated that focal lesions affecting the medio-dorsal thalamic nuclei have a
significant impact on recall after a delay of 24 hours, despite intact acquisition and recall after a
1 hour delay. Two aspects strengthen the validity of the findings in this study: i) analyses were
carried out at the group level, which is rare in the literature due to significant variability in
laterality, size and location of lesions in thalamic stroke patients, ii) advanced tractography
demonstrated the principal pathway connecting the hippocampus to the anterior thalamus, the
mammillothalamic tract, which is often damaged as a consequence of thalamic stroke was intact
in our patient cohort (Carlesimo et al., 2011). Consequently, the structural abnormalities
underlying observed memory impairment could only be attributed to lesions involving the
medio-dorsal nuclei of the thalamus. Furthermore, an observation with significant clinical
implications was that the stroke patients showed relatively intact memory profile on standardised
neuropsychological measures. Recently, this finding has since been replicated in a study by
Danet and colleagues (Danet et al., 2015) who, in a similar cohort of thalamic stroke patients,
demonstrated mild memory impairment on standardised neuropsychological measures when
damage was confined to the medio-dorsal nucleus, but severe when the mammillothalamic tract
was also involved. The inclusion of memory recall measures with an extended delay (i.e., greater

127
than 24 hours) may hold increased sensitivity in characterising memory impairment in patients
with similar brain injury. When considering the absence in cognitive decline on standard visual
and verbal measures of memory recall in AD and FTD patients over 12 months (Chapter 4),
assessment of long-term memory retention may be a better indicator of monitoring functional
decline associated with disease progression. A key goal to further current understanding of the
role of the thalamus in memory processing will be to provide more accurate segregation of
individual thalamic nuclei and their relation to temporal, parietal, and frontal lobe areas,
longitudinally.
Longitudinal imaging provides insight into underlying disease pathology: The integration of
neuroimaging with novel experimental tasks is present in all studies conducted in this thesis and
provides insights into the underlying neural mechanisms associated with our cognitive findings.
Accurate characterisation of longitudinal in-vivo change in neurodegenerative diseases, such as
AD and FTD, hold significant implications for development of targeted cognitive and therapeutic
intervention. In regard to longitudinal analyses, accurate registration and group specific brain
templates for identifying regions of significant morphological change is an issue addressed
relatively poorly by well-established analysis packages, such as FSL. The longitudinal imaging
carried out in Chapter 4 incorporated additional stages of processing including: i) group
templates generated using a half-way transformation between two-time points, and ii) additional
tensor based orientation for diffusion imaging analyses. Both approaches provide greater
statistical power in detecting longitudinal change than the default processing pipeline provided in
FSL toolboxes (Douaud et al., 2009; Keihaninejad et al., 2013). In a clinical application of the
benefits of longitudinal imaging, Chapter 4 demonstrated that pathological differences in disease
progression can be detected, in-vivo, in clinically well-defined patient cohorts. Specifically,

128
patients with suspected AD and FTD pathology exhibited divergent multi-centric and focal white
matter degradation, respectively (Tu, Leyton, Hodges, Piguet, & Hornberger, 2015). While
underlying disease pathology can only be confirmed at post-mortem, proxy markers of pathology
in the two patient cohorts supported our findings.
The findings provided in this thesis provide important new insights into spatial and
episodic memory impairment in dementia and stroke, and their underlying neural substrates.
From a clinical perspective, divergent pathological change in neural structures associated with
memory processing can be used to drive development of more sensitive cognitive measures for
initial diagnosis and monitoring disease progression. While it is clear memory function cannot be
attributed to a single brain structure, the severity of impairment to specific processes can be
modulated by damage to relay structures or pathways in the brain’s memory circuit. In the
context of AD and FTD diagnosis, it has become increasingly evident that standard clinical
measures of memory performance can be improved both for diagnosis and measuring rate of
annual decline. Given the extensive atrophy to virtually all relay nodes comprising the Papez
circuit, novel measures with neural bases beyond the hippocampus, such as the virtual
supermarket task and long-term contextual recall task, are likely to be effective markers of
disease. It is hoped that continued exploration of the conditional relationship between neural
substrates and behavioural symptoms will drive development of effective disease intervention
and therapies in neurodegenerative conditions.

129
Reference List for Introduction and Conclusion Chapters
Acosta-Cabronero, J., & Nestor, P. J. (2014). Diffusion tensor imaging in alzheimer's disease:
Insights into the limbic-diencephalic network and methodological considerations. Frontiers
in Aging Neuroscience, 6, 266. doi:10.3389/fnagi.2014.00266 [doi]
Adlam, A. L., Patterson, K., & Hodges, J. R. (2009). "I remember it as if it were yesterday":
Memory for recent events in patients with semantic dementia. Neuropsychologia, 47(5),
1344-1351. doi:10.1016/j.neuropsychologia.2009.01.029
Aggleton, J. P. (2014). Looking beyond the hippocampus: Old and new neurological targets for
understanding memory disorders. Proceedings.Biological Sciences / the Royal Society,
281(1786), 10.1098/rspb.2014.0565. doi:10.1098/rspb.2014.0565 [doi]
Aggleton, J. P., Dumont, J. R., & Warburton, E. C. (2011). Unraveling the contributions of the
diencephalon to recognition memory: A review. Learning & Memory (Cold Spring Harbor,
N.Y.), 18(6), 384-400. doi:10.1101/lm.1884611; 10.1101/lm.1884611
Albert, M. S., DeKosky, S. T., Dickson, D., Dubois, B., Feldman, H. H., Fox, N. C., . . . Phelps,
C. H. (2011). The diagnosis of mild cognitive impairment due to alzheimer's disease:
Recommendations from the national institute on aging-alzheimer's association workgroups
on diagnostic guidelines for alzheimer's disease. Alzheimer's & Dementia : The Journal of
the Alzheimer's Association, 7(3), 270-279. doi:10.1016/j.jalz.2011.03.008 [doi]

130
Ashburner, J., & Friston, K. J. (2000). Voxel-based morphometry--the methods. NeuroImage,
11(6 Pt 1), 805-821. doi:10.1006/nimg.2000.0582
Bach, M., Laun, F. B., Leemans, A., Tax, C. M., Biessels, G. J., Stieltjes, B., & Maier-Hein, K.
H. (2014). Methodological considerations on tract-based spatial statistics (TBSS).
NeuroImage, 100, 358-369. doi:10.1016/j.neuroimage.2014.06.021 [doi]
Backman, L., Small, B. J., & Fratiglioni, L. (2001). Stability of the preclinical episodic memory
deficit in alzheimer's disease. Brain : A Journal of Neurology, 124(Pt 1), 96-102.
Batchelor, P., Atkinson, D., Hill, D., Calamante, F., & Connelly, A. (2003). Anisotropic noise
propagation in diffusion tensor MRI sampling schemes. Magnetic Resonance in Medicine,
49(6), 1143-1151.
Baumann, O., & Mattingley, J. B. (2010). Medial parietal cortex encodes perceived heading
direction in humans. The Journal of Neuroscience : The Official Journal of the Society for
Neuroscience, 30(39), 12897-12901. doi:10.1523/JNEUROSCI.3077-10.2010 [doi]
Bellassen, V., Igloi, K., de Souza, L. C., Dubois, B., & Rondi-Reig, L. (2012). Temporal order
memory assessed during spatiotemporal navigation as a behavioral cognitive marker for
differential alzheimer's disease diagnosis. The Journal of Neuroscience : The Official
Journal of the Society for Neuroscience, 32(6), 1942-1952. doi:10.1523/JNEUROSCI.4556-
11.2012 [doi]

131
Bertoux, M., de Souza, L. C., Corlier, F., Lamari, F., Bottlaender, M., Dubois, B., & Sarazin, M.
(2014). Two distinct amnesic profiles in behavioral variant frontotemporal dementia.
Biological Psychiatry, 75(7), 582-588. doi:10.1016/j.biopsych.2013.08.017 [doi]
Binetti, G., Locascio, J. J., Corkin, S., Vonsattel, J. P., & Growdon, J. H. (2000). Differences
between pick disease and alzheimer disease in clinical appearance and rate of cognitive
decline. Archives of Neurology, 57(2), 225-232.
Binetti, G., Magni, E., Padovani, A., Cappa, S. F., Bianchetti, A., & Trabucchi, M. (1996).
Executive dysfunction in early alzheimer's disease. Journal of Neurology, Neurosurgery,
and Psychiatry, 60(1), 91-93.
Bitar, R., Leung, G., Perng, R., Tadros, S., Moody, A. R., Sarrazin, J., . . . Roberts, T. P. (2006).
MR pulse sequences: What every radiologist wants to know but is afraid to ask.
Radiographics : A Review Publication of the Radiological Society of North America, Inc,
26(2), 513-537. doi:26/2/513 [pii]
Blackwell, A. D., Sahakian, B. J., Vesey, R., Semple, J. M., Robbins, T. W., & Hodges, J. R.
(2004). Detecting dementia: Novel neuropsychological markers of preclinical alzheimer's
disease. Dementia and Geriatric Cognitive Disorders, 17(1-2), 42-48.
doi:10.1159/000074081 [doi]
Boccia, M., Nemmi, F., & Guariglia, C. (2014). Neuropsychology of environmental navigation
in humans: Review and meta-analysis of FMRI studies in healthy participants.
Neuropsychology Review, 24(2), 236-251. doi:10.1007/s11065-014-9247-8 [doi]

132
Bookstein, F. L. (2001). "Voxel-based morphometry" should not be used with imperfectly
registered images. NeuroImage, 14(6), 1454-1462. doi:10.1006/nimg.2001.0770 [doi]
Braak, H., & Braak, E. (1991). Neuropathological stageing of alzheimer-related changes. Acta
Neuropathologica, 82(4), 239-259.
Braak, H., & Braak, E. (1998). Evolution of neuronal changes in the course of alzheimer's
disease. Journal of Neural Transmission.Supplementum, 53, 127-140.
Budson, A. E., Daffner, K. R., Desikan, R., & Schacter, D. L. (2000). When false recognition is
unopposed by true recognition: Gist-based memory distortion in alzheimer's disease.
Neuropsychology, 14(2), 277-287.
Burgess, N., Trinkler, I., King, J., Kennedy, A., & Cipolotti, L. (2006). Impaired allocentric
spatial memory underlying topographical disorientation. Reviews in the Neurosciences,
17(1-2), 239-251.
Caine, D., Patterson, K., Hodges, J. R., Heard, R., & Halliday, G. (2001). Severe anterograde
amnesia with extensive hippocampal degeneration in a case of rapidly progressive
frontotemporal dementia. Neurocase, 7(1), 57-64. doi:10.1093/neucas/7.1.57 [doi]
Carlesimo, G. A., Lombardi, M. G., & Caltagirone, C. (2011). Vascular thalamic amnesia: A
reappraisal. Neuropsychologia, 49(5), 777-789.
doi:10.1016/j.neuropsychologia.2011.01.026; 10.1016/j.neuropsychologia.2011.01.026
Carlesimo, G. A., Serra, L., Fadda, L., Cherubini, A., Bozzali, M., & Caltagirone, C. (2007).
Bilateral damage to the mammillo-thalamic tract impairs recollection but not familiarity in

133
the recognition process: A single case investigation. Neuropsychologia, 45(11), 2467-2479.
doi:10.1016/j.neuropsychologia.2007.03.025
Chare, L., Hodges, J. R., Leyton, C. E., McGinley, C., Tan, R. H., Kril, J. J., & Halliday, G. M.
(2014). New criteria for frontotemporal dementia syndromes: Clinical and pathological
diagnostic implications. Journal of Neurology, Neurosurgery, and Psychiatry, 85(8), 865-
870. doi:10.1136/jnnp-2013-306948 [doi]
Cipolotti, L., Husain, M., Crinion, J., Bird, C. M., Khan, S. S., Losseff, N., . . . Leff, A. P.
(2008). The role of the thalamus in amnesia: A tractography, high-resolution MRI and
neuropsychological study. Neuropsychologia, 46(11), 2745-2758.
doi:10.1016/j.neuropsychologia.2008.05.009; 10.1016/j.neuropsychologia.2008.05.009
Corkin, S., Amaral, D. G., Gonzalez, R. G., Johnson, K. A., & Hyman, B. T. (1997). H. M.'s
medial temporal lobe lesion: Findings from magnetic resonance imaging. The Journal of
Neuroscience : The Official Journal of the Society for Neuroscience, 17(10), 3964-3979.
Dalla Barba, G. (1997). Recognition memory and recollective experience in alzheimer's disease.
Memory (Hove, England), 5(6), 657-672. doi:10.1080/741941546 [doi]
Danet, L., Barbeau, E. J., Eustache, P., Planton, M., Raposo, N., Sibon, I., . . . Pariente, J. (2015).
Thalamic amnesia after infarct: The role of the mammillothalamic tract and mediodorsal
nucleus. Neurology, 85(24), 2107-2115. doi:10.1212/WNL.0000000000002226 [doi]

134
Davatzikos, C. (2004). Why voxel-based morphometric analysis should be used with great
caution when characterizing group differences. NeuroImage, 23(1), 17-20.
doi:10.1016/j.neuroimage.2004.05.010 [doi]
de Souza, L. C., Chupin, M., Bertoux, M., Lehericy, S., Dubois, B., Lamari, F., . . . Sarazin, M.
(2013). Is hippocampal volume a good marker to differentiate alzheimer's disease from
frontotemporal dementia? Journal of Alzheimer's Disease : JAD, 36(1), 57-66.
doi:10.3233/JAD-122293 [doi]
Douaud, G., Mackay, C., Andersson, J., James, S., Quested, D., Ray, M. K., . . . James, A.
(2009). Schizophrenia delays and alters maturation of the brain in adolescence. Brain : A
Journal of Neurology, 132(Pt 9), 2437-2448. doi:10.1093/brain/awp126 [doi]
Du, A. T., Schuff, N., Kramer, J. H., Rosen, H. J., Gorno-Tempini, M. L., Rankin, K., . . .
Weiner, M. W. (2007). Different regional patterns of cortical thinning in alzheimer's disease
and frontotemporal dementia. Brain : A Journal of Neurology, 130(Pt 4), 1159-1166.
doi:awm016 [pii]
Edelstyn, N. M., Hunter, B., & Ellis, S. J. (2006). Bilateral dorsolateral thalamic lesions disrupts
conscious recollection. Neuropsychologia, 44(6), 931-938.
doi:10.1016/j.neuropsychologia.2005.08.012
Einstein, A. (1905). Über die von der molekularkinetischen theorie der wärme geforderte
bewegung von in ruhenden flüssigkeiten suspendierten teilchen. Annalen Der Physik,
322(8), 549-560.

135
Epstein, R. A., Parker, W. E., & Feiler, A. M. (2007). Where am I now? distinct roles for
parahippocampal and retrosplenial cortices in place recognition. The Journal of
Neuroscience : The Official Journal of the Society for Neuroscience, 27(23), 6141-6149.
doi:27/23/6141 [pii]
Epstein, R. A., & Vass, L. K. (2013). Neural systems for landmark-based wayfinding in humans.
Philosophical Transactions of the Royal Society of London.Series B, Biological Sciences,
369(1635), 20120533. doi:10.1098/rstb.2012.0533 [doi]
Gaffan, D., & Gaffan, E. A. (1991). Amnesia in man following transection of the fornix. A
review. Brain : A Journal of Neurology, 114 ( Pt 6)(Pt 6), 2611-2618.
Gore, J. C. (2003). Principles and practice of functional MRI of the human brain. The Journal of
Clinical Investigation, 112(1), 4-9. doi:10.1172/JCI19010 [doi]
Gorno-Tempini, M. L., Hillis, A. E., Weintraub, S., Kertesz, A., Mendez, M., Cappa, S. F., . . .
Grossman, M. (2011). Classification of primary progressive aphasia and its variants.
Neurology, 76(11), 1006-1014. doi:10.1212/WNL.0b013e31821103e6 [doi]
Graham, A., Davies, R., Xuereb, J., Halliday, G., Kril, J., Creasey, H., . . . Hodges, J. (2005).
Pathologically proven frontotemporal dementia presenting with severe amnesia. Brain : A
Journal of Neurology, 128(Pt 3), 597-605. doi:awh348 [pii]
Graham, K. S., Becker, J. T., & Hodges, J. R. (1997). On the relationship between knowledge
and memory for pictures: Evidence from the study of patients with semantic dementia and

136
alzheimer's disease. Journal of the International Neuropsychological Society : JINS, 3(6),
534-544.
Graham, K. S., Simons, J. S., Pratt, K. H., Patterson, K., & Hodges, J. R. (2000). Insights from
semantic dementia on the relationship between episodic and semantic memory.
Neuropsychologia, 38(3), 313-324.
Granziera, C., Hadjikhani, N., Arzy, S., Seeck, M., Meuli, R., & Krueger, G. (2011). In-vivo
magnetic resonance imaging of the structural core of the papez circuit in humans.
Neuroreport, 22(5), 227-231. doi:10.1097/WNR.0b013e328344f75f;
10.1097/WNR.0b013e328344f75f
Hagmann, P., Jonasson, L., Maeder, P., Thiran, J. P., Wedeen, V. J., & Meuli, R. (2006).
Understanding diffusion MR imaging techniques: From scalar diffusion-weighted imaging
to diffusion tensor imaging and beyond. Radiographics : A Review Publication of the
Radiological Society of North America, Inc, 26 Suppl 1, S205-23. doi:26/suppl_1/S205 [pii]
Hampstead, B. M., & Koffler, S. P. (2009). Thalamic contributions to anterograde, retrograde,
and implicit memory: A case study. The Clinical Neuropsychologist, 23(7), 1232-1249.
doi:10.1080/13854040902936679; 10.1080/13854040902936679
Hodges, J. R., Davies, R. R., Xuereb, J. H., Casey, B., Broe, M., Bak, T. H., . . . Halliday, G. M.
(2004). Clinicopathological correlates in frontotemporal dementia. Annals of Neurology,
56(3), 399-406. doi:10.1002/ana.20203 [doi]

137
Hodges, J. R., & Graham, K. S. (2001). Episodic memory: Insights from semantic dementia.
Philosophical Transactions of the Royal Society of London.Series B, Biological Sciences,
356(1413), 1423-1434. doi:10.1098/rstb.2001.0943 [doi]
Hornberger, M., & Piguet, O. (2012). Episodic memory in frontotemporal dementia: A critical
review. Brain : A Journal of Neurology, 135(Pt 3), 678-692. doi:10.1093/brain/aws011 [doi]
Hornberger, M., Piguet, O., Graham, A. J., Nestor, P. J., & Hodges, J. R. (2010). How preserved
is episodic memory in behavioral variant frontotemporal dementia? Neurology, 74(6), 472-
479. doi:10.1212/WNL.0b013e3181cef85d [doi]
Hornberger, M., Wong, S., Tan, R., Irish, M., Piguet, O., Kril, J., . . . Halliday, G. (2012). In vivo
and post-mortem memory circuit integrity in frontotemporal dementia and alzheimer's
disease. Brain : A Journal of Neurology, 135(Pt 10), 3015-3025. doi:10.1093/brain/aws239;
10.1093/brain/aws239
Hort, J., Laczo, J., Vyhnalek, M., Bojar, M., Bures, J., & Vlcek, K. (2007). Spatial navigation
deficit in amnestic mild cognitive impairment. Proceedings of the National Academy of
Sciences of the United States of America, 104(10), 4042-4047. doi:0611314104 [pii]
Hsieh, S., Hodges, J. R., Leyton, C. E., & Mioshi, E. (2012). Longitudinal changes in primary
progressive aphasias: Differences in cognitive and dementia staging measures. Dementia
and Geriatric Cognitive Disorders, 34(2), 135-141. doi:000342347 [pii]
Hui Zhang, Avants, B. B., Yushkevich, P. A., Woo, J. H., Sumei Wang, McCluskey, L. F., . . .
Gee, J. C. (2007). High-dimensional spatial normalization of diffusion tensor images

138
improves the detection of white matter differences: An example study using amyotrophic
lateral sclerosis. Medical Imaging, IEEE Transactions On, 26(11), 1585-1597.
doi:10.1109/TMI.2007.906784
Hutchinson, A. D., & Mathias, J. L. (2007). Neuropsychological deficits in frontotemporal
dementia and alzheimer's disease: A meta-analytic review. Journal of Neurology,
Neurosurgery, and Psychiatry, 78(9), 917-928. doi:jnnp.2006.100669 [pii]
Iaria, G., Chen, J., Guariglia, C., Ptito, A., & Petrides, M. (2007). Retrosplenial and hippocampal
brain regions in human navigation: Complementary functional contributions to the
formation and use of cognitive maps. European Journal of Neuroscience, 25(3), 890-899.
doi:10.1111/j.1460-9568.2007.05371.x
Ino, T., Doi, T., Hirose, S., Kimura, T., Ito, J., & Fukuyama, H. (2007). Directional
disorientation following left retrosplenial hemorrhage: A case report with fMRI studies.
Cortex; a Journal Devoted to the Study of the Nervous System and Behavior, 43(2), 248-
254.
Irish, M., Halena, S., Kamminga, J., Tu, S., Hornberger, M., & Hodges, J. R. (2015). Scene
construction impairments in alzheimer's disease - A unique role for the posterior cingulate
cortex. Cortex; a Journal Devoted to the Study of the Nervous System and Behavior, 73, 10-
23. doi:S0010-9452(15)00292-0 [pii]
Irish, M., Piguet, O., Hodges, J. R., & Hornberger, M. (2014). Common and unique gray matter
correlates of episodic memory dysfunction in frontotemporal dementia and alzheimer's
disease. Human Brain Mapping, 35(4), 1422-1435. doi:10.1002/hbm.22263 [doi]

139
Jones, D. K. (2004). The effect of gradient sampling schemes on measures derived from
diffusion tensor MRI: A monte carlo study†. Magnetic Resonance in Medicine, 51(4), 807-
815.
Jung, Y. C., Chanraud, S., & Sullivan, E. V. (2012). Neuroimaging of wernicke's encephalopathy
and korsakoff's syndrome. Neuropsychology Review, 22(2), 170-180. doi:10.1007/s11065-
012-9203-4; 10.1007/s11065-012-9203-4
Kalova, E., Vlcek, K., Jarolimova, E., & Bures, J. (2005). Allothetic orientation and sequential
ordering of places is impaired in early stages of alzheimer's disease: Corresponding results
in real space tests and computer tests. Behavioural Brain Research, 159(2), 175-186.
doi:S0166-4328(04)00400-0 [pii]
Keihaninejad, S., Zhang, H., Ryan, N. S., Malone, I. B., Modat, M., Cardoso, M. J., . . . Ourselin,
S. (2013). An unbiased longitudinal analysis framework for tracking white matter changes
using diffusion tensor imaging with application to alzheimer's disease. NeuroImage, 72,
153-163. doi:10.1016/j.neuroimage.2013.01.044 [doi]
Kim, H. (2016). Default network activation during episodic and semantic memory retrieval: A
selective meta-analytic comparison. Neuropsychologia, 80, 35-46.
doi:10.1016/j.neuropsychologia.2015.11.006 [doi]
Kishiyama, M. M., Yonelinas, A. P., Kroll, N. E., Lazzara, M. M., Nolan, E. C., Jones, E. G., &
Jagust, W. J. (2005). Bilateral thalamic lesions affect recollection- and familiarity-based
recognition memory judgments. Cortex; a Journal Devoted to the Study of the Nervous
System and Behavior, 41(6), 778-788.

140
Kwok, T. C., Yuen, K. S., Ho, F. K., & Chan, W. M. (2010). Getting lost in the community: A
phone survey on the community-dwelling demented people in hong kong. International
Journal of Geriatric Psychiatry, 25(4), 427-432. doi:10.1002/gps.2361 [doi]
Laczo, J., Andel, R., Vyhnalek, M., Vlcek, K., Magerova, H., Varjassyova, A., . . . Hort, J.
(2012). From morris water maze to computer tests in the prediction of alzheimer's disease.
Neuro-Degenerative Diseases, 10(1-4), 153-157. doi:10.1159/000333121 [doi]
Laczo, J., Vlcek, K., Vyhnalek, M., Vajnerova, O., Ort, M., Holmerova, I., . . . Hort, J. (2009).
Spatial navigation testing discriminates two types of amnestic mild cognitive impairment.
Behavioural Brain Research, 202(2), 252-259. doi:10.1016/j.bbr.2009.03.041 [doi]
Land, M. F. (2014). Do we have an internal model of the outside world? Philosophical
Transactions of the Royal Society B: Biological Sciences, 369(1636)
doi:10.1098/rstb.2013.0045
Lee, A. C., Rahman, S., Hodges, J. R., Sahakian, B. J., & Graham, K. S. (2003). Associative and
recognition memory for novel objects in dementia: Implications for diagnosis. The
European Journal of Neuroscience, 18(6), 1660-1670. doi:2883 [pii]
Linn, R. T., Wolf, P. A., Bachman, D. L., Knoefel, J. E., Cobb, J. L., Belanger, A. J., . . .
D'Agostino, R. B. (1995). The 'preclinical phase' of probable alzheimer's disease. A 13-year
prospective study of the framingham cohort. Archives of Neurology, 52(5), 485-490.
Luzzi, S., Cafazzo, V., Damora, A., Fabi, K., Fringuelli, F. M., Ascoli, G., . . . Reverberi, C.
(2015). The neural correlates of road sign knowledge and route learning in semantic

141
dementia and alzheimer's disease. Journal of Neurology, Neurosurgery, and Psychiatry,
86(6), 595-602. doi:10.1136/jnnp-2014-309477 [doi]
Mackenzie, I. R., Neumann, M., Bigio, E. H., Cairns, N. J., Alafuzoff, I., Kril, J., . . . Mann, D.
M. (2010). Nomenclature and nosology for neuropathologic subtypes of frontotemporal
lobar degeneration: An update. Acta Neuropathologica, 119(1), 1-4. doi:10.1007/s00401-
009-0612-2 [doi]
Maddock, R. J., Garrett, A. S., & Buonocore, M. H. (2001). Remembering familiar people: The
posterior cingulate cortex and autobiographical memory retrieval. Neuroscience, 104(3),
667-76.
Maguire, E. A., Gadian, D. G., Johnsrude, I. S., Good, C. D., Ashburner, J., Frackowiak, R. S., &
Frith, C. D. (2000). Navigation-related structural change in the hippocampi of taxi drivers.
Proc Natl Acad Sci U S A, 97(8), 4398-403. doi:10.1073/pnas.070039597
Maguire, E. A., Nannery, R., & Spiers, H. J. (2006). Navigation around london by a taxi driver
with bilateral hippocampal lesions. Brain : A Journal of Neurology, 129(Pt 11), 2894-2907.
doi:10.1093/brain/awl286
Mahoney, C. J., Simpson, I. J., Nicholas, J. M., Fletcher, P. D., Downey, L. E., Golden, H. L., . .
. Fox, N. C. (2015). Longitudinal diffusion tensor imaging in frontotemporal dementia.
Annals of Neurology, 77(1), 33-46. doi:10.1002/ana.24296 [doi]

142
Marchette, S. A., Vass, L. K., Ryan, J., & Epstein, R. A. (2014). Anchoring the neural compass:
Coding of local spatial reference frames in human medial parietal lobe. Nature
Neuroscience, 17(11), 1598-1606. doi:10.1038/nn.3834 [doi]
McKhann, G. M., Knopman, D. S., Chertkow, H., Hyman, B. T., Jack, C. R.,Jr, Kawas, C. H., . .
. Phelps, C. H. (2011). The diagnosis of dementia due to alzheimer's disease:
Recommendations from the national institute on aging-alzheimer's association workgroups
on diagnostic guidelines for alzheimer's disease. Alzheimer's & Dementia : The Journal of
the Alzheimer's Association, 7(3), 263-269. doi:10.1016/j.jalz.2011.03.005 [doi]
Miller, J. P., Sweet, J. A., Bailey, C. M., Munyon, C. N., Luders, H. O., & Fastenau, P. S.
(2015). Visual-spatial memory may be enhanced with theta burst deep brain stimulation of
the fornix: A preliminary investigation with four cases. Brain : A Journal of Neurology,
138(Pt 7), 1833-1842. doi:10.1093/brain/awv095 [doi]
Moffat, S. D. (2009). Aging and spatial navigation: What do we know and where do we go?
Neuropsychology Review, 19(4), 478-489. doi:10.1007/s11065-009-9120-3 [doi]
Morganti, F., Stefanini, S., & Riva, G. (2013). From allo- to egocentric spatial ability in early
alzheimer's disease: A study with virtual reality spatial tasks. Cognitive Neuroscience, 4(3-
4), 171-180. doi:10.1080/17588928.2013.854762 [doi]
Morris, R. G., Garrud, P., Rawlins, J. N., & O'Keefe, J. (1982). Place navigation impaired in rats
with hippocampal lesions. Nature, 297(5868), 681-683.

143
Neary, D., Snowden, J. S., Gustafson, L., Passant, U., Stuss, D., Black, S., . . . Benson, D. F.
(1998). Frontotemporal lobar degeneration: A consensus on clinical diagnostic criteria.
Neurology, 51(6), 1546-1554.
Nedelska, Z., Andel, R., Laczó, J., Vlcek, K., Horinek, D., Lisy, J., . . . Hort, J. (2012). Spatial
navigation impairment is proportional to right hippocampal volume. Proceedings of the
National Academy of Sciences, 109(7), 2590-2594. doi:10.1073/pnas.1121588109
Nestor, P. J., Fryer, T. D., Ikeda, M., & Hodges, J. R. (2003). Retrosplenial cortex (BA 29/30)
hypometabolism in mild cognitive impairment (prodromal alzheimer's disease). The
European Journal of Neuroscience, 18(9), 2663-2667. doi:2999 [pii]
O'Keefe, J., & Nadel, L. (1978). The hippocampus as a cognitive map Oxford University Press.
Pai, M. C., & Jacobs, W. J. (2004). Topographical disorientation in community-residing patients
with alzheimer's disease. International Journal of Geriatric Psychiatry, 19(3), 250-255.
doi:10.1002/gps.1081 [doi]
Pai, M. C., & Yang, Y. C. (2013). Impaired translation of spatial representation in young onset
alzheimer's disease patients. Current Alzheimer Research, 10(1), 95-103. doi:CAR-EPUB-
20121002-1 [pii]
Papadakis, N. G., Murrills, C. D., Hall, L. D., Huang, C. L., & Carpenter, T. A. (2000). Minimal
gradient encoding for robust estimation of diffusion anisotropy☆. Magnetic Resonance
Imaging, 18(6), 671-679.

144
Papageorgiou, S. G., Beratis, I. N., Horvath, J., Herrmann, F. R., Bouras, C., & Kovari, E.
(2016). Amnesia in frontotemporal dementia: Shedding light on the geneva historical data.
Journal of Neurology, 263(4), 657-664. doi:10.1007/s00415-015-8019-6 [doi]
Papez, J. W. (1995). A proposed mechanism of emotion. 1937. The Journal of Neuropsychiatry
and Clinical Neurosciences, 7(1), 103-112.
Pengas, G., Hodges, J. R., Watson, P., & Nestor, P. J. (2010). Focal posterior cingulate atrophy
in incipient alzheimer's disease. Neurobiology of Aging, 31(1), 25-33.
doi:10.1016/j.neurobiolaging.2008.03.014 [doi]
Pengas, G., Patterson, K., Arnold, R. J., Bird, C. M., Burgess, N., & Nestor, P. J. (2010). Lost
and found: Bespoke memory testing for alzheimer's disease and semantic dementia. Journal
of Alzheimer's Disease : JAD, 21(4), 1347-1365.
Pennington, C., Hodges, J. R., & Hornberger, M. (2011). Neural correlates of episodic memory
in behavioral variant frontotemporal dementia. Journal of Alzheimer's Disease : JAD, 24(2),
261-268. doi:10.3233/JAD-2011-101668 [doi]
Pereira, J. M., Xiong, L., Acosta-Cabronero, J., Pengas, G., Williams, G. B., & Nestor, P. J.
(2010). Registration accuracy for VBM studies varies according to region and degenerative
disease grouping. NeuroImage, 49(3), 2205-2215. doi:10.1016/j.neuroimage.2009.10.068
[doi]
Perry, R. J., & Hodges, J. R. (2000). Differentiating frontal and temporal variant frontotemporal
dementia from alzheimer's disease. Neurology, 54(12), 2277-2284.

145
Piguet, O., Leyton, C. E., Gleeson, L. D., Hoon, C., & Hodges, J. R. (2015). Memory and
emotion processing performance contributes to the diagnosis of non-semantic primary
progressive aphasia syndromes. Journal of Alzheimer's Disease : JAD, 44(2), 541-547.
doi:10.3233/JAD-141854 [doi]
Pooley, R. A. (2005). AAPM/RSNA physics tutorial for residents: Fundamental physics of MR
imaging. Radiographics : A Review Publication of the Radiological Society of North
America, Inc, 25(4), 1087-1099. doi:25/4/1087 [pii]
Preston, A. R., & Eichenbaum, H. (2013). Interplay of hippocampus and prefrontal cortex in
memory. Current Biology : CB, 23(17), R764-73. doi:10.1016/j.cub.2013.05.041;
10.1016/j.cub.2013.05.041
Ranjith, N., Mathuranath, P. S., Sharma, G., & Alexander, A. (2010). Qualitative aspects of
learning, recall, and recognition in dementia. Annals of Indian Academy of Neurology,
13(2), 117-122. doi:10.4103/0972-2327.64639 [doi]
Rascovsky, K., Hodges, J. R., Knopman, D., Mendez, M. F., Kramer, J. H., Neuhaus, J., . . .
Miller, B. L. (2011). Sensitivity of revised diagnostic criteria for the behavioural variant of
frontotemporal dementia. Brain : A Journal of Neurology, 134(Pt 9), 2456-2477.
doi:10.1093/brain/awr179 [doi]
Rauchs, G., Piolino, P., Mezenge, F., Landeau, B., Lalevee, C., Pelerin, A., . . . Desgranges, B.
(2007). Autonoetic consciousness in alzheimer's disease: Neuropsychological and PET
findings using an episodic learning and recognition task. Neurobiology of Aging, 28(9),
1410-1420. doi:S0197-4580(06)00216-8 [pii]

146
Ridgway, G. R., Omar, R., Ourselin, S., Hill, D. L., Warren, J. D., & Fox, N. C. (2009). Issues
with threshold masking in voxel-based morphometry of atrophied brains. NeuroImage,
44(1), 99-111. doi:10.1016/j.neuroimage.2008.08.045 [doi]
Rohrer, J. D., Rossor, M. N., & Warren, J. D. (2012). Alzheimer's pathology in primary
progressive aphasia. Neurobiology of Aging, 33(4), 744-752.
doi:10.1016/j.neurobiolaging.2010.05.020 [doi]
Rudebeck, S. R., Scholz, J., Millington, R., Rohenkohl, G., Johansen-Berg, H., & Lee, A. C.
(2009). Fornix microstructure correlates with recollection but not familiarity memory. The
Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 29(47),
14987-14992. doi:10.1523/JNEUROSCI.4707-09.2009; 10.1523/JNEUROSCI.4707-
09.2009
Salmon, D. P. (2000). Disorders of memory in alzheimer’s disease. In L. S. Cermak (Ed.),
Handbook of neuropsychology, vol. 2: Memory and its disorders (2nd ed., pp. 155-195).
Amsterdam: Elsevier.
Salmon, D. P., & Bondi, M. W. (2009). Neuropsychological assessment of dementia. Annual
Review of Psychology, 60, 257-282. doi:10.1146/annurev.psych.57.102904.190024
Schoemaker, D., Gauthier, S., & Pruessner, J. C. (2014). Recollection and familiarity in aging
individuals with mild cognitive impairment and alzheimer's disease: A literature review.
Neuropsychology Review, 24(3), 313-331. doi:10.1007/s11065-014-9265-6 [doi]

147
Scoville, W., & Milner, B. (1957). Loss of recent memory after bilateral hippocampal lesions.
Journal of Neurology, Neurosurgery, and Psychiatry, 20, 11-21.
Seelaar, H., Rohrer, J. D., Pijnenburg, Y. A., Fox, N. C., & van Swieten, J. C. (2011). Clinical,
genetic and pathological heterogeneity of frontotemporal dementia: A review. Journal of
Neurology, Neurosurgery, and Psychiatry, 82(5), 476-486. doi:10.1136/jnnp.2010.212225;
10.1136/jnnp.2010.212225
Seeley, W. W., Crawford, R., Rascovsky, K., Kramer, J. H., Weiner, M., Miller, B. L., & Gorno-
Tempini, M. L. (2008). Frontal paralimbic network atrophy in very mild behavioral variant
frontotemporal dementia. Archives of Neurology, 65(2), 249-255.
doi:10.1001/archneurol.2007.38 [doi]
Serino, S., Cipresso, P., Morganti, F., & Riva, G. (2014). The role of egocentric and allocentric
abilities in alzheimer's disease: A systematic review. Ageing Research Reviews, 16, 32-44.
doi:10.1016/j.arr.2014.04.004 [doi]
Shah, A., Jhawar, S. S., & Goel, A. (2012). Analysis of the anatomy of the papez circuit and
adjoining limbic system by fiber dissection techniques. Journal of Clinical Neuroscience :
Official Journal of the Neurosurgical Society of Australasia, 19(2), 289-298.
doi:10.1016/j.jocn.2011.04.039; 10.1016/j.jocn.2011.04.039
Small, B. J., Fratiglioni, L., Viitanen, M., Winblad, B., & Backman, L. (2000). The course of
cognitive impairment in preclinical alzheimer disease: Three- and 6-year follow-up of a
population-based sample. Archives of Neurology, 57(6), 839-844. doi:noc90093 [pii]

148
Smith, S. M. (2002). Fast robust automated brain extraction. Human Brain Mapping, 17(3), 143-
155. doi:10.1002/hbm.10062 [doi]
Smith, S. M., Jenkinson, M., Johansen-Berg, H., Rueckert, D., Nichols, T. E., Mackay, C. E., . . .
Behrens, T. E. (2006). Tract-based spatial statistics: Voxelwise analysis of multi-subject
diffusion data. NeuroImage, 31(4), 1487-1505. doi:10.1016/j.neuroimage.2006.02.024
Smith, S. M., & Nichols, T. E. (2009). Threshold-free cluster enhancement: Addressing
problems of smoothing, threshold dependence and localisation in cluster inference.
NeuroImage, 44(1), 83-98. doi:10.1016/j.neuroimage.2008.03.061
Squire, L. R., Clark, R. E., & Knowlton, B. J. (2001). Retrograde amnesia. Hippocampus, 11(1),
50-55. doi:2-G
Squire, L. R., & Alvarez, P. (1995). Retrograde amnesia and memory consolidation: A
neurobiological perspective. Current Opinion in Neurobiology, 5(2), 169-177.
Takahashi, N., Kawamura, M., Shiota, J., Kasahata, N., & Hirayama, K. (1997). Pure
topographic disorientation due to right retrosplenial lesion. Neurology, 49(2), 464-469.
Tan, R. H., Wong, S., Kril, J. J., Piguet, O., Hornberger, M., Hodges, J. R., & Halliday, G. M.
(2014). Beyond the temporal pole: Limbic memory circuit in the semantic variant of
primary progressive aphasia. Brain : A Journal of Neurology, 137(Pt 7), 2065-2076.
doi:10.1093/brain/awu118 [doi]
Tsivilis, D., Vann, S. D., Denby, C., Roberts, N., Mayes, A. R., Montaldi, D., & Aggleton, J. P.
(2008). A disproportionate role for the fornix and mammillary bodies in recall versus

149
recognition memory. Nature Neuroscience, 11(7), 834-842. doi:10.1038/nn.2149;
10.1038/nn.2149
Tu, S., Leyton, C. E., Hodges, J. R., Piguet, O., & Hornberger, M. (2015). Divergent longitudinal
propagation of white matter degradation in logopenic and semantic variants of primary
progressive aphasia. Journal of Alzheimer's Disease : JAD, 49(3), 853-861.
doi:10.3233/JAD-150626 [doi]
Tu, S., Miller, L., Piguet, O., & Hornberger, M. (2014). Accelerated forgetting of contextual
details due to focal medio-dorsal thalamic lesion. Frontiers in Behavioral Neuroscience, 8
Valenstein, E., Bowers, D., Verfaellie, M., Heilman, K. M., Day, A., & Watson, R. T. (1987).
Retrosplenial amnesia. Brain : A Journal of Neurology, 110 ( Pt 6)(Pt 6), 1631-1646.
Van der Werf, Y. D., Scheltens, P., Lindeboom, J., Witter, M. P., Uylings, H. B., & Jolles, J.
(2003). Deficits of memory, executive functioning and attention following infarction in the
thalamus; a study of 22 cases with localised lesions. Neuropsychologia, 41(10), 1330-1344.
doi:S0028393203000599 [pii]
Vann, S. D., Aggleton, J. P., & Maguire, E. A. (2009). What does the retrosplenial cortex do?
Nature Reviews.Neuroscience, 10(11), 792-802. doi:10.1038/nrn2733 [doi]
Weintraub, S., Wicklund, A. H., & Salmon, D. P. (2012). The neuropsychological profile of
alzheimer disease. Cold Spring Harbor Perspectives in Medicine, 2(4), a006171.
doi:10.1101/cshperspect.a006171 [doi]

150
Whitwell, J. L., Jack, C. R.,Jr, Przybelski, S. A., Parisi, J. E., Senjem, M. L., Boeve, B. F., . . .
Josephs, K. A. (2011). Temporoparietal atrophy: A marker of AD pathology independent of
clinical diagnosis. Neurobiology of Aging, 32(9), 1531-1541.
doi:10.1016/j.neurobiolaging.2009.10.012 [doi]
Wicklund, A. H., Johnson, N., Rademaker, A., Weitner, B. B., & Weintraub, S. (2006). Word list
versus story memory in alzheimer disease and frontotemporal dementia. Alzheimer Disease
and Associated Disorders, 20(2), 86-92. doi:10.1097/01.wad.0000213811.97305.49 [doi]
Winocur, G., & Moscovitch, M. (2011). Memory transformation and systems consolidation.
Journal of the International Neuropsychological Society : JINS, 17(5), 766-780.
doi:10.1017/S1355617711000683
Winocur, G., Moscovitch, M., & Bontempi, B. (2010). Memory formation and long-term
retention in humans and animals: Convergence towards a transformation account of
hippocampal-neocortical interactions. Neuropsychologia, 48(8), 2339-2356.
doi:10.1016/j.neuropsychologia.2010.04.016
Yew, B., Alladi, S., Shailaja, M., Hodges, J. R., & Hornberger, M. (2013). Lost and forgotten?
orientation versus memory in alzheimer's disease and frontotemporal dementia. Journal of
Alzheimer's Disease : JAD, 33(2), 473-481. doi:10.3233/JAD-2012-120769 [doi]
Zaehle, T., Jordan, K., Wustenberg, T., Baudewig, J., Dechent, P., & Mast, F. W. (2007). The
neural basis of the egocentric and allocentric spatial frame of reference. Brain Research,
1137(1), 92-103. doi:S0006-8993(06)03577-3 [pii]

151
Zhang, Y., Brady, M., & Smith, S. (2001). Segmentation of brain MR images through a hidden
markov random field model and the expectation-maximization algorithm. IEEE
Transactions on Medical Imaging, 20(1), 45-57. doi:10.1109/42.906424

152
Appendices
Supplemental Material for Publications in Chapters
Page No.
Chapter 2 - Publication I 153
Chapter 2 – Publication II 157
Chapter 3 – Publication III 160
Chapter 4 – Publication IV 163
Chapter 5 – Publication V 164
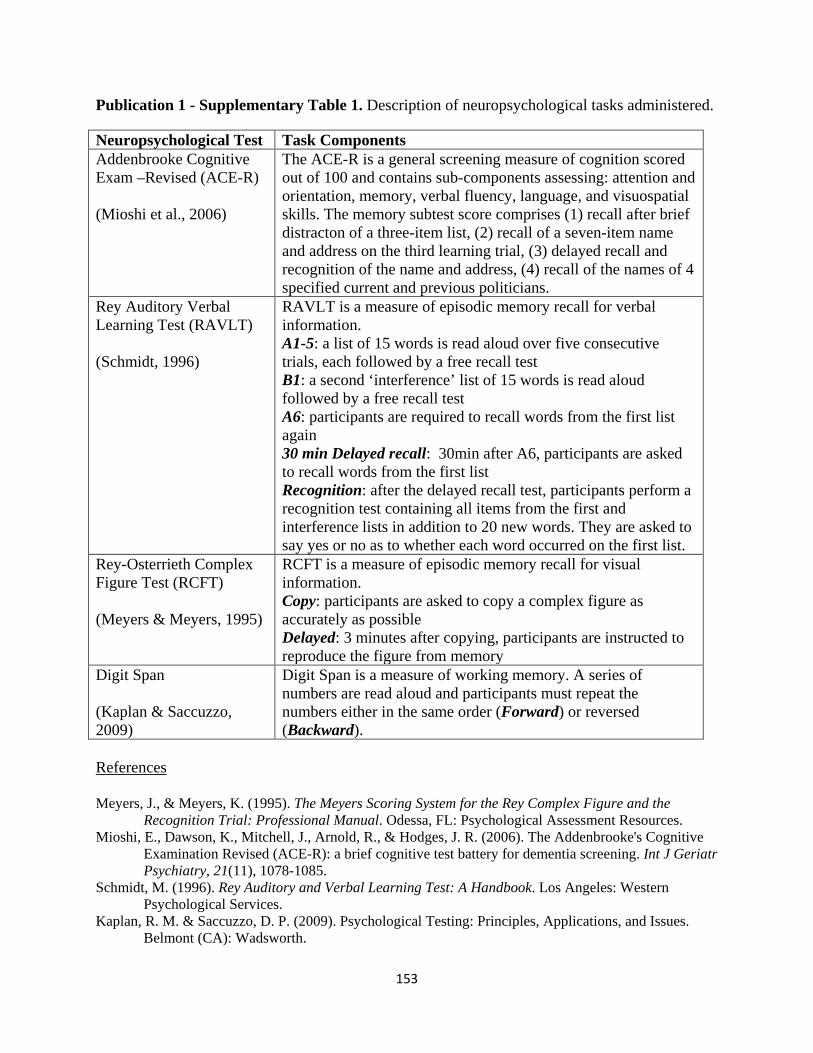
153
Publication 1 - Supplementary Table 1. Description of neuropsychological tasks administered.
Neuropsychological Test Task Components Addenbrooke Cognitive Exam –Revised (ACE-R) (Mioshi et al., 2006)
The ACE-R is a general screening measure of cognition scored out of 100 and contains sub-components assessing: attention and orientation, memory, verbal fluency, language, and visuospatial skills. The memory subtest score comprises (1) recall after brief distracton of a three-item list, (2) recall of a seven-item name and address on the third learning trial, (3) delayed recall and recognition of the name and address, (4) recall of the names of 4 specified current and previous politicians.
Rey Auditory Verbal Learning Test (RAVLT) (Schmidt, 1996)
RAVLT is a measure of episodic memory recall for verbal information. A1-5: a list of 15 words is read aloud over five consecutive trials, each followed by a free recall test B1: a second ‘interference’ list of 15 words is read aloud followed by a free recall test A6: participants are required to recall words from the first list again 30 min Delayed recall: 30min after A6, participants are asked to recall words from the first list Recognition: after the delayed recall test, participants perform a recognition test containing all items from the first and interference lists in addition to 20 new words. They are asked to say yes or no as to whether each word occurred on the first list.
Rey-Osterrieth Complex Figure Test (RCFT) (Meyers & Meyers, 1995)
RCFT is a measure of episodic memory recall for visual information. Copy: participants are asked to copy a complex figure as accurately as possible Delayed: 3 minutes after copying, participants are instructed to reproduce the figure from memory
Digit Span (Kaplan & Saccuzzo, 2009)
Digit Span is a measure of working memory. A series of numbers are read aloud and participants must repeat the numbers either in the same order (Forward) or reversed (Backward).
References
Meyers, J., & Meyers, K. (1995). The Meyers Scoring System for the Rey Complex Figure and the
Recognition Trial: Professional Manual. Odessa, FL: Psychological Assessment Resources. Mioshi, E., Dawson, K., Mitchell, J., Arnold, R., & Hodges, J. R. (2006). The Addenbrooke's Cognitive
Examination Revised (ACE-R): a brief cognitive test battery for dementia screening. Int J Geriatr Psychiatry, 21(11), 1078-1085.
Schmidt, M. (1996). Rey Auditory and Verbal Learning Test: A Handbook. Los Angeles: Western Psychological Services.
Kaplan, R. M. & Saccuzzo, D. P. (2009). Psychological Testing: Principles, Applications, and Issues. Belmont (CA): Wadsworth.
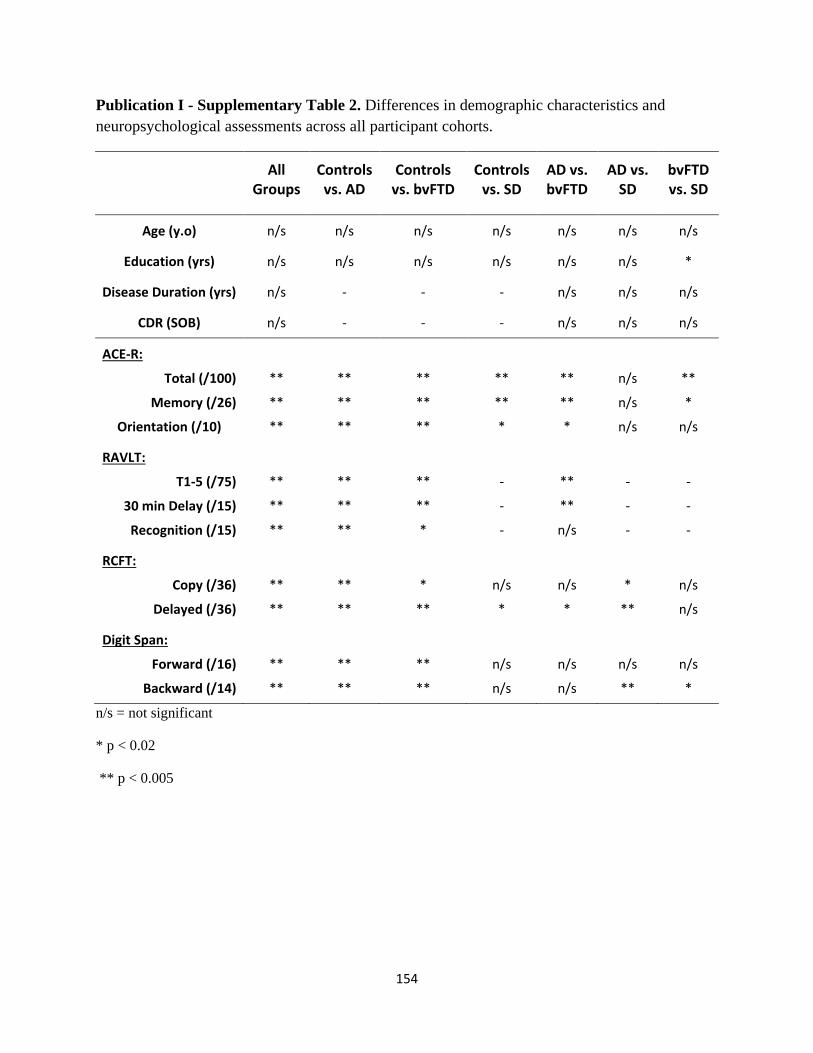
154
Publication I - Supplementary Table 2. Differences in demographic characteristics and neuropsychological assessments across all participant cohorts.
All Groups
Controls vs. AD
Controls vs. bvFTD
Controls vs. SD
AD vs. bvFTD
AD vs. SD
bvFTD vs. SD
Age (y.o) n/s n/s n/s n/s n/s n/s n/s
Education (yrs) n/s n/s n/s n/s n/s n/s *
Disease Duration (yrs) n/s - - - n/s n/s n/s
CDR (SOB) n/s - - - n/s n/s n/s
ACE-R:
Total (/100)
Memory (/26)
Orientation (/10)
**
**
**
**
**
**
**
**
**
**
**
*
**
**
*
n/s
n/s
n/s
**
*
n/s
RAVLT:
T1-5 (/75)
30 min Delay (/15)
Recognition (/15)
**
**
**
**
**
**
**
**
*
-
-
-
**
**
n/s
-
-
-
-
-
-
RCFT:
Copy (/36)
Delayed (/36)
**
**
**
**
*
**
n/s
*
n/s
*
*
**
n/s
n/s
Digit Span:
Forward (/16)
Backward (/14)
**
**
**
**
**
**
n/s
n/s
n/s
n/s
n/s
**
n/s
*
n/s = not significant
* p < 0.02
** p < 0.005
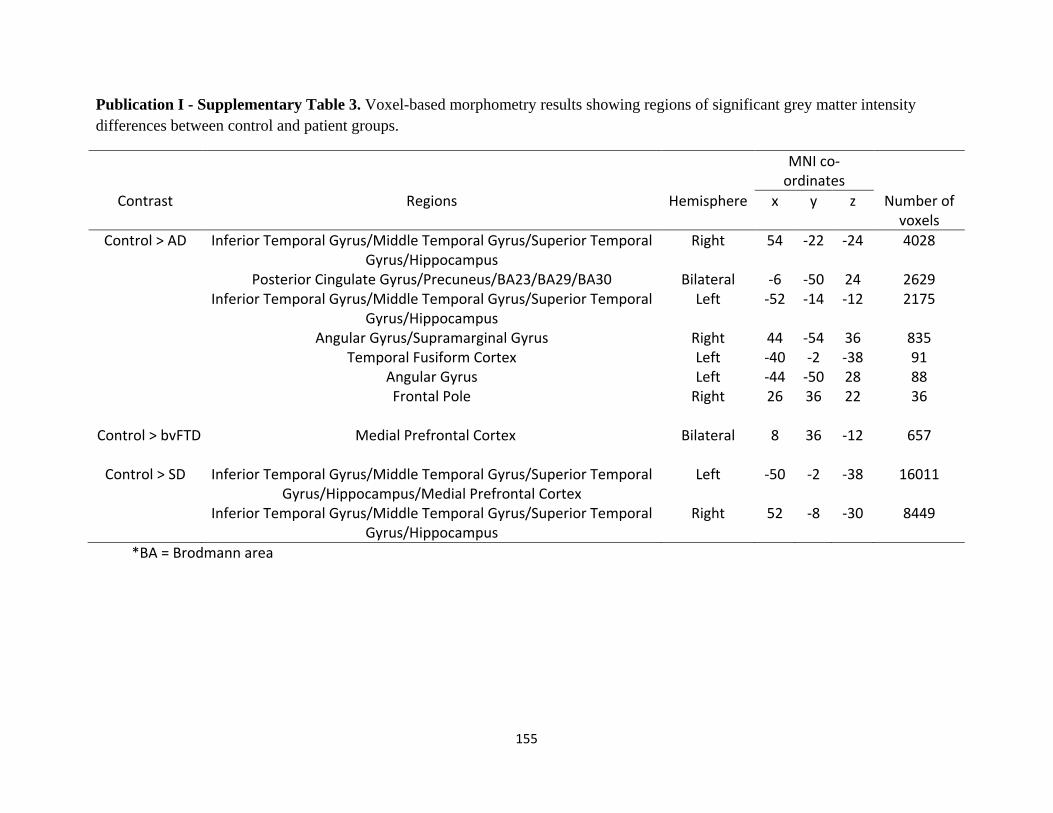
155
Publication I - Supplementary Table 3. Voxel-based morphometry results showing regions of significant grey matter intensity differences between control and patient groups.
MNI co-ordinates
Contrast Regions Hemisphere x y z Number of voxels
Control > AD Inferior Temporal Gyrus/Middle Temporal Gyrus/Superior Temporal Gyrus/Hippocampus
Right 54 -22 -24 4028
Posterior Cingulate Gyrus/Precuneus/BA23/BA29/BA30 Bilateral -6 -50 24 2629 Inferior Temporal Gyrus/Middle Temporal Gyrus/Superior Temporal
Gyrus/Hippocampus Left -52 -14 -12 2175
Angular Gyrus/Supramarginal Gyrus Right 44 -54 36 835 Temporal Fusiform Cortex Left -40 -2 -38 91 Angular Gyrus Left -44 -50 28 88 Frontal Pole Right 26 36 22 36
Control > bvFTD Medial Prefrontal Cortex Bilateral 8 36 -12 657
Control > SD Inferior Temporal Gyrus/Middle Temporal Gyrus/Superior Temporal Gyrus/Hippocampus/Medial Prefrontal Cortex
Left -50 -2 -38 16011
Inferior Temporal Gyrus/Middle Temporal Gyrus/Superior Temporal Gyrus/Hippocampus
Right 52 -8 -30 8449
*BA = Brodmann area
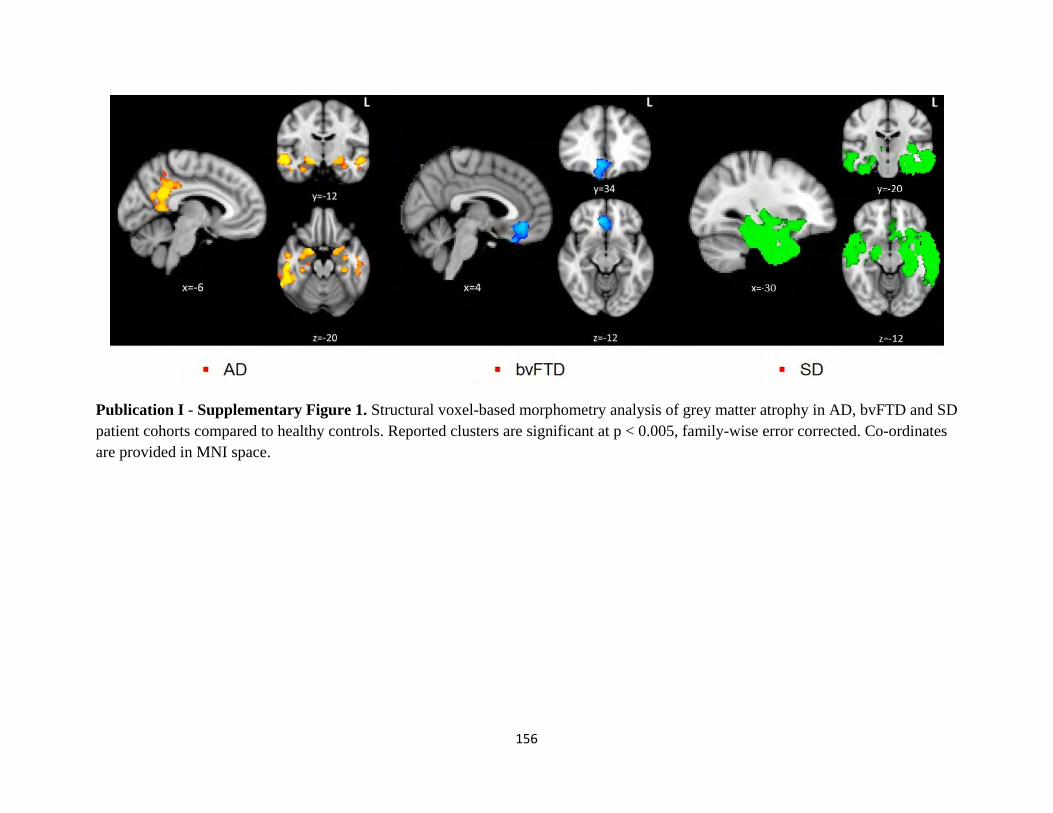
156
Publication I - Supplementary Figure 1. Structural voxel-based morphometry analysis of grey matter atrophy in AD, bvFTD and SD patient cohorts compared to healthy controls. Reported clusters are significant at p < 0.005, family-wise error corrected. Co-ordinates are provided in MNI space.
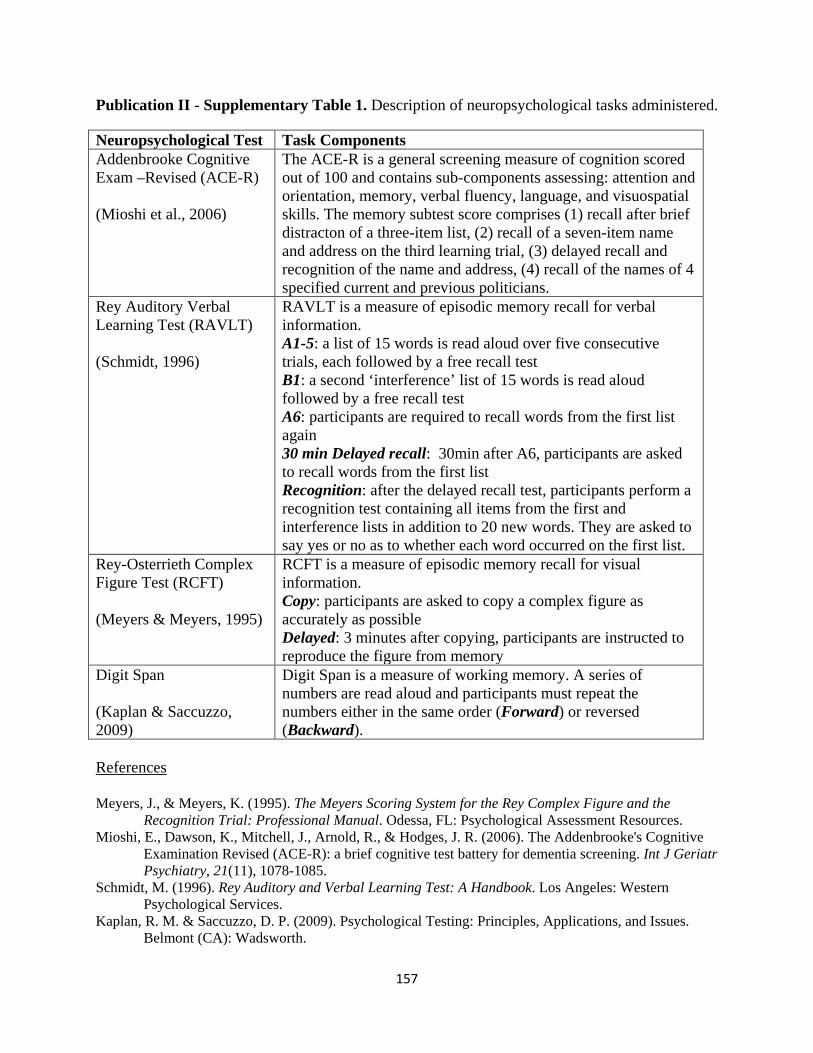
157
Publication II - Supplementary Table 1. Description of neuropsychological tasks administered.
Neuropsychological Test Task Components Addenbrooke Cognitive Exam –Revised (ACE-R) (Mioshi et al., 2006)
The ACE-R is a general screening measure of cognition scored out of 100 and contains sub-components assessing: attention and orientation, memory, verbal fluency, language, and visuospatial skills. The memory subtest score comprises (1) recall after brief distracton of a three-item list, (2) recall of a seven-item name and address on the third learning trial, (3) delayed recall and recognition of the name and address, (4) recall of the names of 4 specified current and previous politicians.
Rey Auditory Verbal Learning Test (RAVLT) (Schmidt, 1996)
RAVLT is a measure of episodic memory recall for verbal information. A1-5: a list of 15 words is read aloud over five consecutive trials, each followed by a free recall test B1: a second ‘interference’ list of 15 words is read aloud followed by a free recall test A6: participants are required to recall words from the first list again 30 min Delayed recall: 30min after A6, participants are asked to recall words from the first list Recognition: after the delayed recall test, participants perform a recognition test containing all items from the first and interference lists in addition to 20 new words. They are asked to say yes or no as to whether each word occurred on the first list.
Rey-Osterrieth Complex Figure Test (RCFT) (Meyers & Meyers, 1995)
RCFT is a measure of episodic memory recall for visual information. Copy: participants are asked to copy a complex figure as accurately as possible Delayed: 3 minutes after copying, participants are instructed to reproduce the figure from memory
Digit Span (Kaplan & Saccuzzo, 2009)
Digit Span is a measure of working memory. A series of numbers are read aloud and participants must repeat the numbers either in the same order (Forward) or reversed (Backward).
References
Meyers, J., & Meyers, K. (1995). The Meyers Scoring System for the Rey Complex Figure and the
Recognition Trial: Professional Manual. Odessa, FL: Psychological Assessment Resources. Mioshi, E., Dawson, K., Mitchell, J., Arnold, R., & Hodges, J. R. (2006). The Addenbrooke's Cognitive
Examination Revised (ACE-R): a brief cognitive test battery for dementia screening. Int J Geriatr Psychiatry, 21(11), 1078-1085.
Schmidt, M. (1996). Rey Auditory and Verbal Learning Test: A Handbook. Los Angeles: Western Psychological Services.
Kaplan, R. M. & Saccuzzo, D. P. (2009). Psychological Testing: Principles, Applications, and Issues. Belmont (CA): Wadsworth.
158
Publication II - Supplementary Figure 1. Representative responses on the spatial map component of the virtual supermarket task from participant groups. An example map of correct trial locations is shown at the top.
159
Publication II - Supplementary Figure 2. Mean retrosplenial cortex volume in participant groups. No significant differences were found between groups.
160
Publication III - Supplementary Table 1. Description of neuropsychological tasks administered.
Neuropsychological Test Task Components Addenbrooke Cognitive Exam –Revised (ACE-R) (Mioshi et al., 2006)
The ACE-R is a general screening measure of cognition scored out of 100 and contains sub-components assessing: attention and orientation, memory, verbal fluency, language, and visuospatial skills. The memory subtest score comprises (1) recall after brief distracton of a three-item list, (2) recall of a seven-item name and address on the third learning trial, (3) delayed recall and recognition of the name and address, (4) recall of the names of 4 specified current and previous politicians.
Doors subtest of the Doors & People Test (D&PT) (Baddeley, Emslie, & Nimmo-Smith, 1994)
This subtest examines visual recognition memory. It is comprised of two sections (A/B). In each section participants are first shown pictures of 12 different doors, then required to pick them out one at a time from arrays of 4 pictures (target and 3 distractors). In section B, the targets and distracters are more closely matched than in section A, and thus, more difficult.
Rey Auditory Verbal Learning Test (RAVLT) (Schmidt, 1996)
RAVLT is a measure of episodic memory recall for verbal information. A1-5: a list of 15 words is read aloud over five consecutive trials, each followed by a free recall test B1: a second ‘interference’ list of 15 words is read aloud followed by a free recall test A6: participants are required to recall words from the first list again 30 min Delayed recall: 30min after A6, participants are asked to recall words from the first list Recognition: after the delayed recall test, participants perform a recognition test containing all items from the first and interference lists in addition to 20 new words. They are asked to say yes or no as to whether each word occurred on the first list.
Rey-Osterrieth Complex Figure Test (RCFT) (Meyers & Meyers, 1995)
RCFT is a measure of episodic memory recall for visual information. Copy: participants are asked to copy a complex figure as accurately as possible Delayed: 3 minutes after copying, participants are instructed to reproduce the figure from memory
References
Baddeley, A. D., Emslie, H., & Nimmo-Smith, I. (1994). The Doors and People Test: a test of visual
and verbal recall and recognition. Bury St. Edmonds: Thames Valley Test Company. Meyers, J., & Meyers, K. (1995). The Meyers Scoring System for the Rey Complex Figure and the
Recognition Trial: Professional Manual. Odessa, FL: Psychological Assessment Resources. Mioshi, E., Dawson, K., Mitchell, J., Arnold, R., & Hodges, J. R. (2006). The Addenbrooke's
Cognitive Examination Revised (ACE-R): a brief cognitive test battery for dementia screening. Int J Geriatr Psychiatry, 21(11), 1078-1085.
Schmidt, M. (1996). Rey Auditory and Verbal Learning Test: A Handbook. Los Angeles: Western Psychological Services.
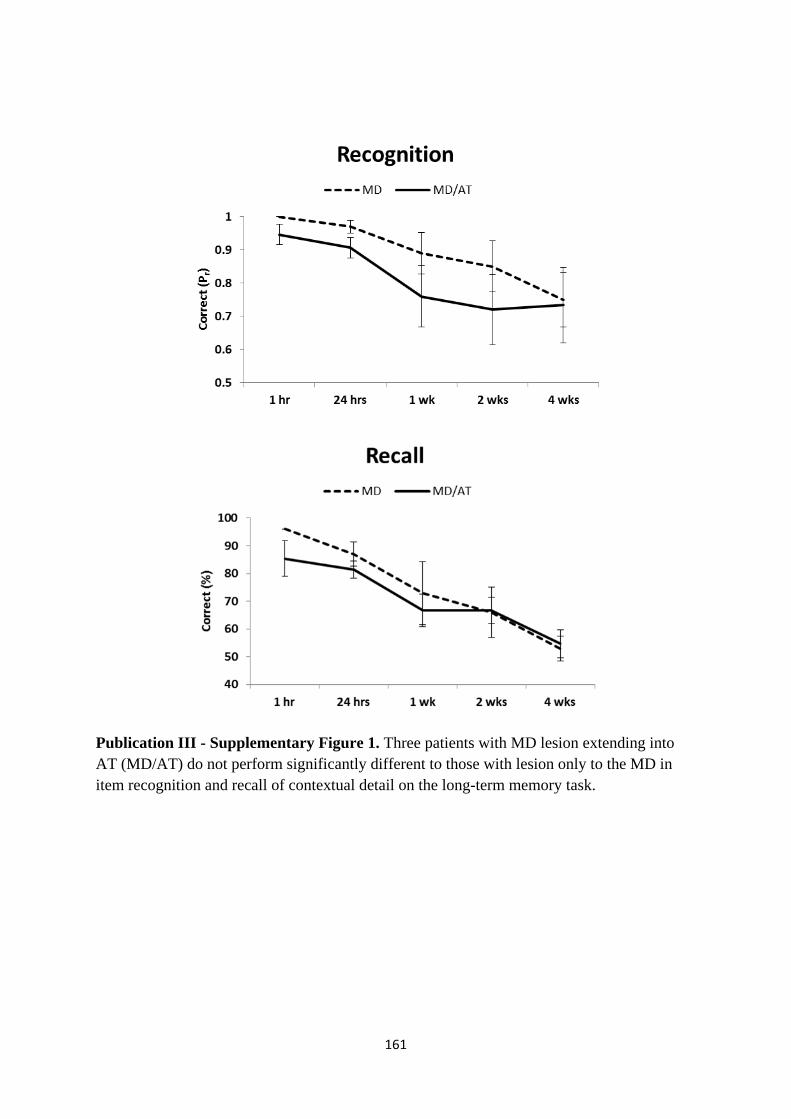
161
Publication III - Supplementary Figure 1. Three patients with MD lesion extending into AT (MD/AT) do not perform significantly different to those with lesion only to the MD in item recognition and recall of contextual detail on the long-term memory task.
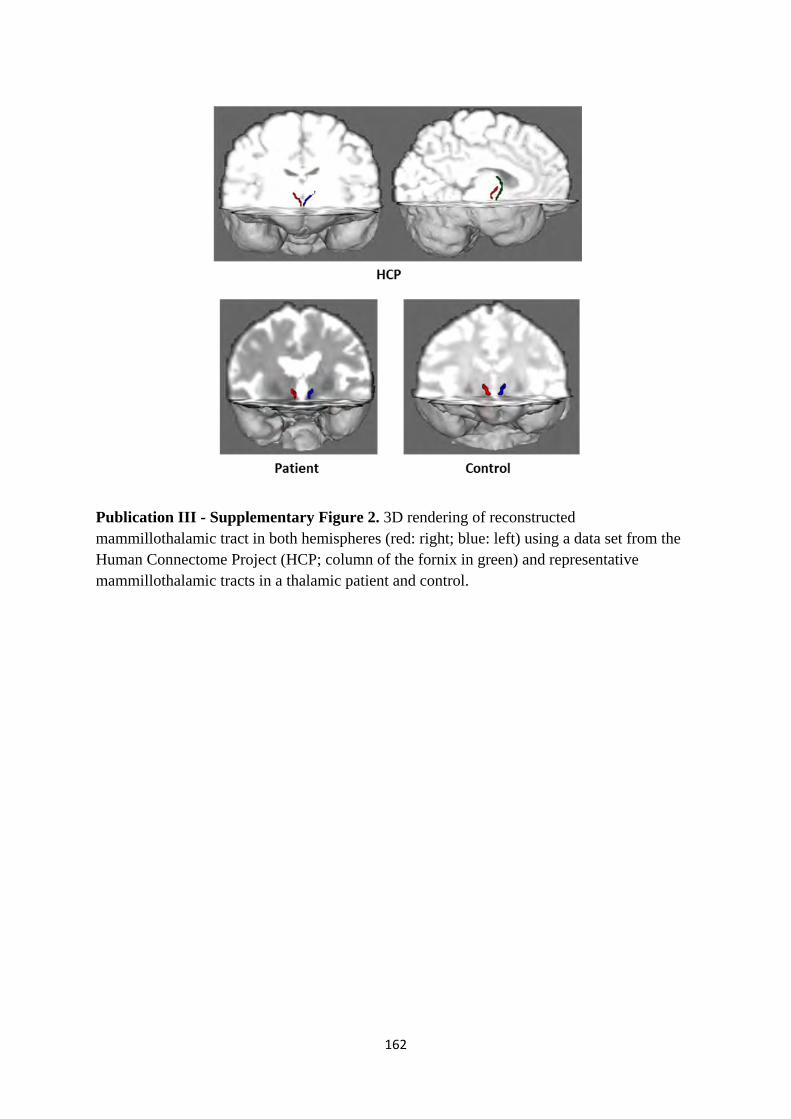
162
Publication III - Supplementary Figure 2. 3D rendering of reconstructed mammillothalamic tract in both hemispheres (red: right; blue: left) using a data set from the Human Connectome Project (HCP; column of the fornix in green) and representative mammillothalamic tracts in a thalamic patient and control.
163
Publication IV - Supplementary Table 1. Description of neuropsychological tasks administered.
Neuropsychological Test Task Components Addenbrooke Cognitive Exam –Revised (ACE-R) (Mioshi et al., 2006)
The ACE-R is a general screening measure of cognition scored out of 100 and contains sub-components assessing: attention and orientation, memory, verbal fluency, language, and visuospatial skills. The memory subtest score comprises (1) recall after brief distracton of a three-item list, (2) recall of a seven-item name and address on the third learning trial, (3) delayed recall and recognition of the name and address, (4) recall of the names of 4 specified current and previous politicians.
Rey Auditory Verbal Learning Test (RAVLT) (Schmidt, 1996)
RAVLT is a measure of episodic memory recall for verbal information. A1-5: a list of 15 words is read aloud over five consecutive trials, each followed by a free recall test B1: a second ‘interference’ list of 15 words is read aloud followed by a free recall test A6: participants are required to recall words from the first list again 30 min Delayed recall: 30min after A6, participants are asked to recall words from the first list Recognition: after the delayed recall test, participants perform a recognition test containing all items from the first and interference lists in addition to 20 new words. They are asked to say yes or no as to whether each word occurred on the first list.
Rey-Osterrieth Complex Figure Test (RCFT) (Meyers & Meyers, 1995)
RCFT is a measure of episodic memory recall for visual information. Copy: participants are asked to copy a complex figure as accurately as possible Delayed: 3 minutes after copying, participants are instructed to reproduce the figure from memory
References
Meyers, J., & Meyers, K. (1995). The Meyers Scoring System for the Rey Complex Figure and the
Recognition Trial: Professional Manual. Odessa, FL: Psychological Assessment Resources. Mioshi, E., Dawson, K., Mitchell, J., Arnold, R., & Hodges, J. R. (2006). The Addenbrooke's
Cognitive Examination Revised (ACE-R): a brief cognitive test battery for dementia screening. Int J Geriatr Psychiatry, 21(11), 1078-1085.
Schmidt, M. (1996). Rey Auditory and Verbal Learning Test: A Handbook. Los Angeles: Western Psychological Services.
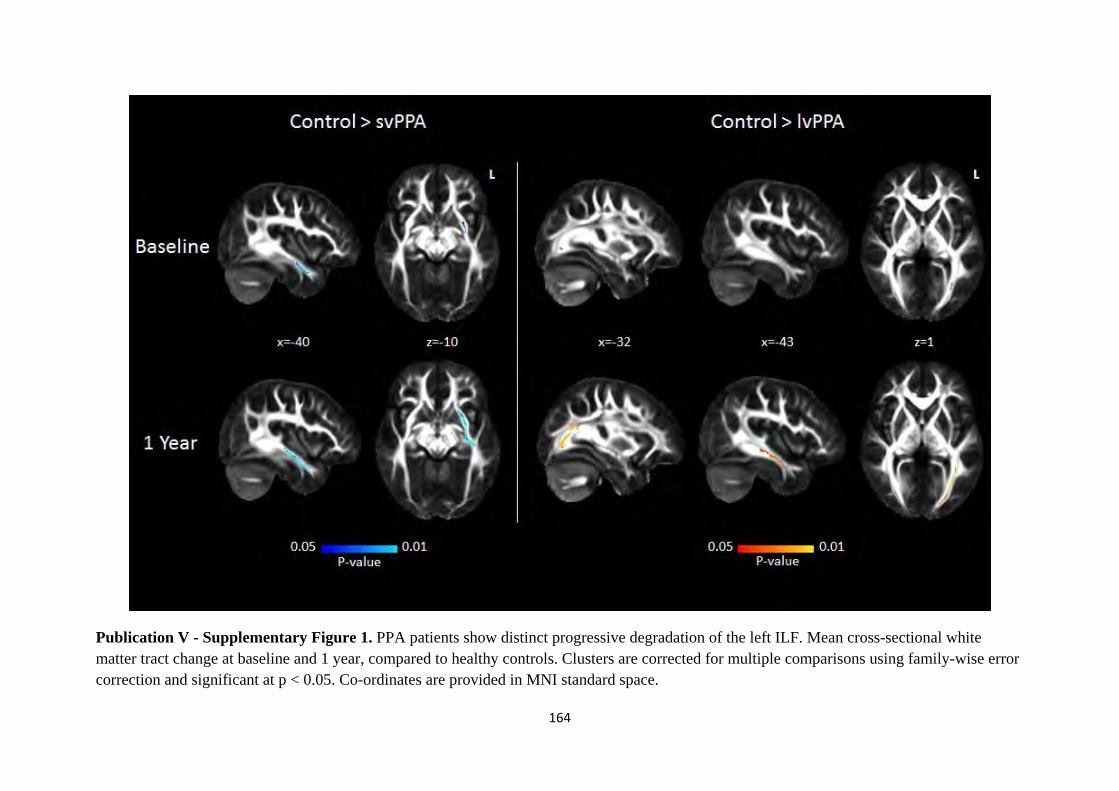
164
Publication V - Supplementary Figure 1. PPA patients show distinct progressive degradation of the left ILF. Mean cross-sectional white matter tract change at baseline and 1 year, compared to healthy controls. Clusters are corrected for multiple comparisons using family-wise error correction and significant at p < 0.05. Co-ordinates are provided in MNI standard space.
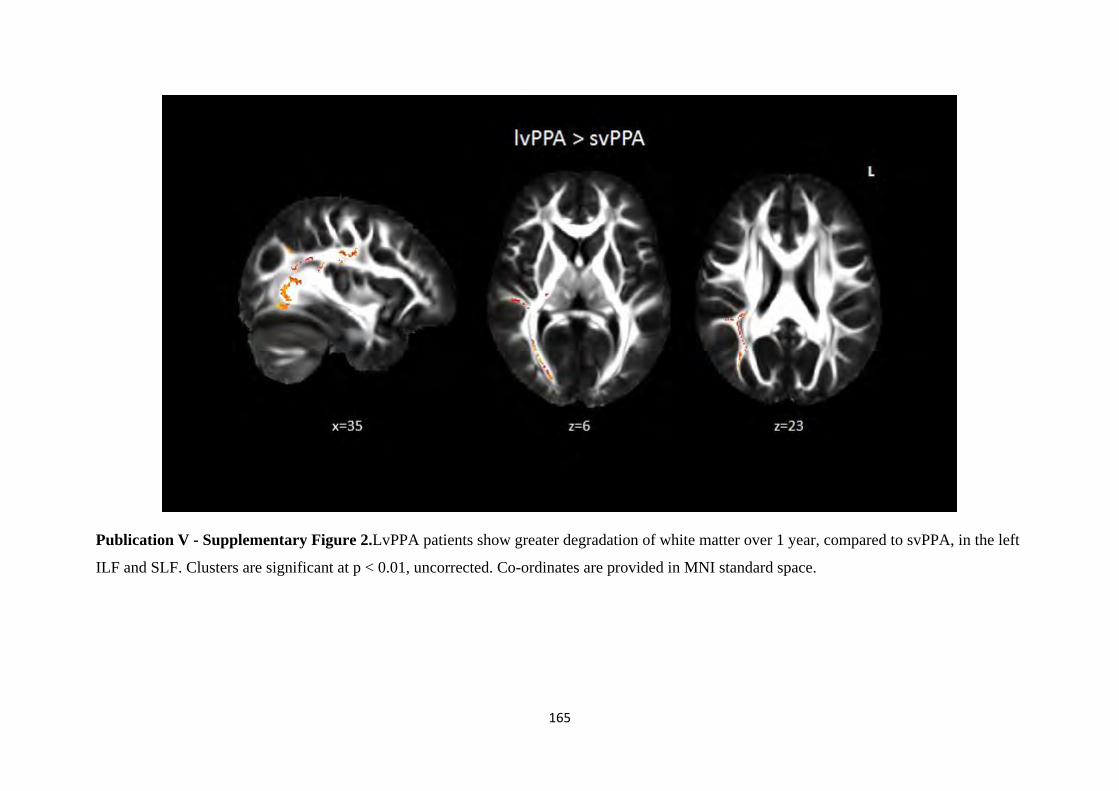
165
Publication V - Supplementary Figure 2.LvPPA patients show greater degradation of white matter over 1 year, compared to svPPA, in the left
ILF and SLF. Clusters are significant at p < 0.01, uncorrected. Co-ordinates are provided in MNI standard space.

166
Declaration Supporting Inclusion of Publications in the Thesis
Publication I – Tu, S., Wong, S., Hodges, J. R., Irish, M., Piguet, O., & Hornberger, M. (2015).
Lost in spatial translation - A novel tool to objectively assess spatial disorientation in alzheimer's
disease and frontotemporal dementia. Cortex; a Journal Devoted to the Study of the Nervous
System and Behavior, 67, 83-94. doi:10.1016/j.cortex.2015.03.016 [doi]
Publication II - Tu, S., Spiers, H. J., Hodges, J. R., Piguet, O., Hornberger, M. Egocentric vs.
allocentric spatial memory in behavioural variant frontotemporal dementia and Alzheimer’s
disease. (in submission)
Publication III - Tu, S., Miller, L., Piguet, O., & Hornberger, M. (2014). Accelerated forgetting
of contextual details due to focal medio-dorsal thalamic lesion. Frontiers in Behavioral
Neuroscience, 8
Publication IV - Tu, S., Piguet, O., Hodges, J. R., Hornberger, M. Longitudinal Papez circuit
integrity in Alzheimer’s disease and frontotemporal dementia. (in submission)
Publication V - Tu, S., Leyton, C. E., Hodges, J. R., Piguet, O., & Hornberger, M. (2015).
Divergent longitudinal propagation of white matter degradation in logopenic and semantic
variants of primary progressive aphasia. Journal of Alzheimer's Disease : JAD, 49(3), 853-861.
doi:10.3233/JAD-150626 [doi]

167
Declaration:
For each publication listed above, I certify that this publication was a direct result of my research
towards this PhD, and that reproduction in this thesis does not breach copyright regulations.
Signed:
Sicong Tu
Date: 30 May 2016





























