Embed Size (px)
Citation preview

INVITED REVIEWboj_1014 329..356
Murderous plants: Victorian Gothic, Darwin andmodern insights into vegetable carnivory
MARK W. CHASE FLS1*, MAARTEN J. M. CHRISTENHUSZ FLS2, DAWN SANDERS FLS3
and MICHAEL F. FAY FLS1
1Jodrell Laboratory, Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3DS, UK2Department of Botany, Natural History Museum, Cromwell Road, London SW7 5BD, UK3Scientific Associate, Department of Botany, Natural History Museum, Cromwell Road, London SW75BD, UK
Received 20 October 2009; accepted for publication 3 November 2009
Darwin’s interest in carnivorous plants was in keeping with the Victorian fascination with Gothic horrors, and hisexperiments on them were many and varied, ranging from what appears to be idle curiosity (e.g. what will happenif I place a human hair on a Drosera leaf?) to detailed investigations of mechanisms. Mechanisms for capture anddigestion of prey vary greatly among the six (or more) lineages of flowering plants that have well-developedcarnivory, and some are much more active than others. Passive carnivory is common in some groups, and one,Roridula (Roridulaceae) from southern Africa, is so passively carnivorous that it requires the presence of an insectintermediate to derive any benefit from prey trapped on its leaves. Other groups not generally considered to becarnivores, such as Stylidium (Stylidiaceae), some species of Potentilla (Rosaceae), Proboscidea (Martyniaceae) andGeranium (Geraniaceae), that have been demonstrated to both produce digestive enzymes on their epidermalsurfaces and be capable of absorbing the products, are putatively just as ‘carnivorous’ as Roridula. There is no clearway to discriminate between cases of passive and active carnivory and between non-carnivorous and carnivorousplants – all intermediates exist. Here, we document the various angiosperm clades in which carnivory has evolvedand the degree to which these plants have become ‘complete carnivores’. We also discuss the problems withdefinition of the terms used to describe carnivorous plants. © 2009 The Linnean Society of London, BotanicalJournal of the Linnean Society, 2009, 161, 329–356.
ADDITIONAL KEYWORDS: carnivorous plants – insectivorous plants – phylogenetics – proto-carnivory.
INTRODUCTION
Carnivorous or insectivorous plants have longinduced fascination in men, and they are among themost popular plants in cultivation; they are oftenoffered for sale in garden centres and over the Inter-net. There are many amateur botanical societies thatfocus upon them. The first living specimen of Dionaeamuscipula Ellis ex L. came to the attention of thepopulace of London in 1768, an event that ‘caused asensation throughout Europe’ (Magee, 2007: 49).Indeed, Linnaeus is reported to have declared
‘miraculum naturae’ (Magee, 2007) upon seeing D.muscipula. Prior to this event, John Bartram hadsent Patrick Collinson, a London botanical collector,several plant parts, after the specimen sent by Gov-ernor Dobbs of North Carolina had failed to arrive(Magee, 2007). Bartram used a popular name for D.muscipula, tipitiwitchet, a somewhat ribald Elizabe-than term for vulva (McKinley postscript to Nelson,1990). This connection between female sexualityand carnivorous plants continued into 19th centuryEngland and may have had something to do withtheir popularity and continued public fascination.
Insectivorous plants epitomize Victorian England’scultural interest in the Gothic form in literature,architecture and art and, in terms of natural history,*Corresponding author. E-mail: [email protected]
Botanical Journal of the Linnean Society, 2009, 161, 329–356. With 12 figures
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356 329
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

bizarre spectacles. These ‘queer flowers’, as GrantAllen described insectivorous plants in 1884, reacheda zenith of popular and artistic attention during themid to late 19th century. Allen’s essay demonstratedthe lure of the insectivorous plant as a floral femmefatale and in richly descriptive language describedits ‘murderous propensities’ (Allen in Smith, 2003).Smith (2003) considered Swinburne’s poem ‘TheSundew’ in relation to Allen’s essay and Darwin’s(1875) study on insectivorous plants. He noted that‘both Swinburne’s poem and Darwin’s book wereprominent elements in a cultural fascination with thesundew that extended from the 1860s well into the1880s’, and ‘in the aftermath of Insectivorous Plantsthe potentially subversive moral and cultural impli-cations of “The Sundew” become more difficult toignore’ (Smith, 2003: 130–131).
In one of the most fanciful of Victorian stories, theGerman explorer Carl Liche and members of thecave-dwelling Mkodo tribe were described as makinga trip through the Madagascan jungle. At one point,they come upon an amazing sight: a large plant witha bulbous trunk resembling a 2.5-m pineapple witheight elongate leaves, 3–4 m long, studded with hook-like thorns surrounding a depression filled withhoney-sweet liquid. At the top of the tree are a set oflong, hairy green tendrils and tentacles, ‘constantlyand vigorously in motion, with . . . a subtle, sinuous,silent throbbing against the air.’ The story goes on tosay that one of their women is forced at javelin pointto climb the trunk. Then ‘the atrocious cannibal tree,that had been so inert and dead, came to suddensavage life. The slender delicate palpi, with the furyof starved serpents, quivered a moment over herhead, then as if instinct with demoniac intelligencefastened upon her in sudden coils round and roundher neck and arms; then while her awful screams andyet more awful laughter rose wildly to be instantlystrangled down again into a gurgling moan, the ten-drils one after another, like great green serpents, withbrutal energy and infernal rapidity, rose, retractedthemselves, and wrapped her about in fold after fold,ever tightening with cruel swiftness and savagetenacity of anacondas fastening upon their prey.’ ‘Thegreat leaves slowly rose and stiffly, like the arms of aderrick, erected themselves in the air, approached oneanother and closed about the dead and hamperedvictim with the silent force of a hydraulic press andthe ruthless purpose of a thumbscrew.’ [‘Liche, 1881’(almost certainly a fictitious author; see below) citedby Osborn (1924)].
Some readers took this account seriously. Travellershad been returning from the jungles of the worldwith astonishing stories: ferocious man-like apes,vine-shrouded lost cities. Gorillas and the Mayanruins turned out to be real. Why not the man-eating
tree in far away Madagascar, when at home inEngland there was a vegetable carnivore, the sundew,known to everyone? In Victorian times, Gothic storiescombining horror and romance prevailed, and thestory of a man-eating tree or any carnivorous plantwas utterly exciting. Vegetable man-eaters matchedthe characters used in the supernatural tales aboutghosts, haunted mansions, werewolves etc.
Combined with the fantastic findings in naturalhistory by travellers, these man-eating trees tickledthe fancy of writers. Buel (1887), for example, in hisbook Sea and Land included a section on carnivorousplants, in which, following a description of the actionof Dionaea Ellis and Drosera L., he then wrote abouta plant from Central Africa (possibly Liche’s Mada-gascan plant) and tropical America, where it is knownas Ya-te-veo [I-see-you.] (Fig. 1), ‘that is not contentedwith the myriad of large insects which it catches andconsumes, but its voracity extends to making evenhumans its prey’. What fate awaited the ‘unfortunatetraveller’? ‘The body is crushed until every drop ofblood is squeezed out of it and becomes absorbed bythe gore-loving plant, when the dry carcass is thrownout and the horrid trap set again.’ Due to the spines
Figure 1. The man-eating tree Ya-te-veo reported to occurin Central America by Buel (1887).
330 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

reported to pierce the body of the victim, Buel madean analogy to the maiden, a torture instrument ‘ofthe dark ages’ (with inward pointing spikes) which‘was made, somewhat crudely, to represent a woman,hence the name applied to it’. Following more gorydetails about ulcers resulting from puncture woundsinflicted by the plant and mention of the ‘hundreds ofresponsible travelers [who] declare they have fre-quently seen it’, he concluded this section as follows:‘All of which, however, I am inclined to doubt; not thatthere is no foundation for such statements as travel-ers sometimes make about this astonishing growth,but that the facts are greatly exaggerated’. Thus evenwriters aiming to provide a vivid, but true, account ofthe wonders of nature managed to confuse real andfictional carnivorous plants.
Liche’s story was adopted by Chase Osborn (1924)in his book Madagascar, Land of the Man-Eating Tree.Osborn said missionaries had vouched for the exist-ence of the tree. No one has ever laid eyes again onthis carnivorous horror, or on the Mkodo tribe for that
matter, and Ley (1955) wrote that the Madagascanman-eating tree, the Mkodo tribe and even Carl Lichehimself were all fabrications. It is nevertheless agruesomely good story that may have its origins inolder works, such as the True History of LucianusSamosatensis, written in the 2nd century AD, in whichfemale grapevines consumed sailors who tried to matewith them (note again the references to carnivorousplants being alluring and female).
Darwin himself was fascinated by carnivorousplants. He came to believe, after much experimenta-tion, that the movement-sensing organ in sundews(Drosera) is far more sensitive than any nerve in thehuman body (Darwin, 1875). American naturalistMary Treat, who studied many of these plants in hergarden and understood their functioning in detail,discovered the function of bladderwort (UtriculariaL.) traps (Fig. 2). She communicated regularly withDarwin about carnivorous plants and helped himunderstand their habits better (Sanders & Gian-quitto, 2009). Another female correspondent and
Figure 2. Utricularia minor L., cultivated at RBG Kew. Photograph taken by Maarten J. M. Christenhusz.
MURDEROUS PLANTS 331
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

friend, Lady Ellen Lubbock (the wife of Darwin’sgreat supporter Sir John Lubbock, later LordAvebury) wrote a poem on reading Darwin’s Insectivo-rous Plants and sent it to him (see Milner, 2009). Itincluded the verse:
I never trusted DroseraSince I went there with a friend,And saw its horrid tentaclesBeginning all to bend.
We are accustomed to think of plants as beingimmobile and harmless, and there is something deeplyunnerving about the thought of carnivorous plants.The ongoing fascination with these stories, exaggerat-ing the traits of real-life carnivorous plants, indicatesthe deep horror we feel towards the idea of beingdevoured by a plant. No wonder the myths of theman-eating tree have stayed with us for centuries.
In 20th century Anglo-American culture, we saw ashift from the Gothic form to kitsch with the showLittle Shop of Horrors and the character Audrey II –an extraterrestrial carnivorous plant that constantlycries ‘feed me’ (Fig. 3). The Life of Pi (Martel, 2001)featured a floating mat of carnivorous algae. Veg-etable carnivores have also been adopted in sciencefiction stories such as Parasite Planet (Weinbaum,1935), in which a plant on Venus eats humans. In TheFellowship of the Ring (Tolkien, 1954), hobbits fallasleep and find themselves eaten by a plant. Hugecarnivorous trees are also dangers of the forests inBeyond the Deepwoods (Steward & Riddell, 1998).
In Comet in Moominland (Jansson, 1973) Moom-introll saves the Snork Maiden from the twining armsof a poisonous bush of the ‘dangerous Angosturafamily’ with his penknife.
A carnivorous plant, Tentacula, is also featured inthe Harry Potter series (Rowling, 1998), building uponthe romantic but horrific idea of plants devouring
people. Even some Pokémon characters (e.g. Bell-sprout, Weepinbell and Victreebell; Pokémon, 2009) arebased on carnivorous plants, bringing them intopopular modern culture. More recently, the discoveryof Nepenthes attenboroughii A.S.Robinson, S.McPher-son & V.B.Henrich (Robinson et al., 2009) has stimu-lated a flurry of internet-dispersed rumours of‘rat-eating plants’ (e.g. Instructables, 2009) andso a fascination with the ‘murderous propensities’of carnivorous plants continues to capture the publicimagination.
DEFINITION OF VEGETABLE CARNIVORY
Turning to a more serious side of the subject, severaldefinitions of botanical carnivory have been proposed,but most researchers still consider at least somespecies lacking some aspects to nonetheless be fullycarnivorous. The basic definition includes at least theability to absorb the products of decomposition, eitherdirectly on the leaves or through roots in the soil,thereby increasing their fitness, ultimately leading toincreased seed production. Including absorption byroots of nutrients released through one method ofdecay or another means that nearly all plants arecapable of a degree of carnivory, and indeed thisminimal definition is the one we follow in this paper (atleast in terms of the taxa we treat). However, research-ers who follow the ‘six lineages of carnivore’ argument(see below; Ellison & Gotelli, 2009) would add as wellthe provisos that these plants should also exhibit somemeans of (1) attracting prey to their traps (glisteningglandular hairs being a minimal common attractant),(2) capture of prey and (3) their digestion. Digestion inmany species generally held to be carnivores is notnecessarily through enzymes secreted by the plantitself, but this can be achieved via enzymes secreted bybacteria, fungi or even the stomachs of other animalsthat eat trapped prey with the subsequent absorptionof nutrients released by deposition of their faeces onnearby soil. The minimal glandular apparatus thatsecretes mucilage or other compounds needed to trapanimals is widespread in angiosperms, almost to thepoint of being universal, at least in eudicots. Croizat(1961) thought that carnivory was an ancestralangiosperm attribute, a concept not far removed fromthe sentiment of the previous sentence; extant fullydeveloped carnivores are merely more refined cases ofunderlying capacities present in all flowering plants.Formation of tanks into which prey and vegetabledetritus fall and decay is also frequent, particularly inthe commelinid monocots (e.g. bromeliads and others;see below). Proving that there is a net energetic benefitto carnivory when its occurrence is frequent and subtlemakes it more difficult (what would one use as thebackground case for angiosperms?). Studies have
Figure 3. The man-eating plant Audrey II from the filmLittle Shop of Horrors (Film Group Inc.; Griffith, 1960).
332 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

shown that construction costs and scaling relation-ships for leaf traits in ‘true’ carnivores are substan-tially greater when traps are active (Ellison & Gotelli,2009), but relatively few taxa have developed suchcomplex traps; active traps occur in only two families,Droseraceae and Lentibulariaceae. In the great major-ity of the taxa we discuss below, carnivory is simpler,and in terms of leaf construction the costs are less andscaling relationships much like those of ‘normal’plants. Overall, angiosperms of many types may beinvolved in a degree of carnivory and be ‘proto-carnivorous’; perhaps we should be more curious aboutwhy more plant species have not developed a ‘taste’ foranimal-derived nutrients.
DEVELOPMENT OF A PHYLOGENETIC FRAMEWORK
FOR CARNIVOROUS PLANTS
Phylogenetic studies of DNA sequences have greatlyaided efforts to assemble a complete tree of life for allangiosperms; based on this understanding of floweringplant phylogeny, full-blown carnivory has evolved inthe angiosperms at least six times (Albert, Williams &Chase, 1992; Chase et al., 1993). These phylogeneticstudies have also provided insights into morphologicalevolution of the varied mechanisms employed byplants to attract, retain, kill and digest animals andfinally absorb the released nutrients. Darwin (1875)himself was certain that there had been several inde-pendent origins of carnivory, and this assumption hasbeen broadly accepted by plant taxonomists since thetime of Darwin. However, acceptance of polyphyly ofcarnivorous plants has been against a backdrop ofmajor disputes over which groups of carnivores wereclosely vs. distantly related and to which other groupsof angiosperms they are related (e.g. Hutchinson,1973; Cronquist, 1981; Thorne, 1992; Takhtajan,1997). Suggestions of relationships between groups ofplants that we now know to be spurious were putforward by some of the most prominent botanists oftheir time: Hooker (1874; based on an idea fromLinnaeus), for example, believed that species of Sar-racenia L. were not only related to but had also evolvedfrom waterlilies as a result of the move onto dry land.Takhtajan (1969) placed Sarraceniales near theranalean complex, which included Nymphaeales,Papaverales and Ranunculales. It is equally clear thatthe origins of carnivory have been clouded by bothconvergent and divergent evolution and, before theadvent of molecular phylogenetics, this subject was atan impasse and could no longer progress.
The first phylogenetic assessment of carnivore rela-tionships was that of Albert et al. (1992), and thispaper set the stage for more complete subsequentinvestigations by other authors. Albert et al. (1992)were particularly interested in the degree to which the
carnivorous syndrome could provide evidence aboutgeneral macroevolutionary patterns and process inflowering plants. Indeed, this must have also been thereason why Darwin studied these plants and wroteInsectivorous Plants (Darwin, 1875). Extreme special-izations were thought to provide important insightsinto more general phenomena, such as the functions ofsticky glands. Sticky, mucilage secreting, glandularhairs occur commonly throughout the angiosperms,and their exact functions are not always easily deter-mined. Many angiosperms are undoubtedly passivecarnivores; once an insect has been trapped by stickyglands, even if prevention of herbivory is the actualrole of these hairs, its subsequent decay can providenutrients, taken up by roots of the trapping plants.This sort of passive carnivory is clearly taking place inthe southern African Roridula Burm. ex L. (Roridu-laceae, Ericales), aided by the action of a Nemo-like†bug that is unharmed by its host and lives its entire lifeon these plants, depositing on the soil around theplants its faeces, to the benefit of its host.
The ability to produce water-storage structures,whether through epiascidiation (inrolling of theabaxial leaf lamina with subsequent fusion of themargins as in Nepenthes L. and Sarracenia) or tankformation (as in the bromeliads Brocchinia Schult.f.and Catopsis Griseb.), provides the possibility of sub-sequent absorption of nutrients released by decay ofany matter, whether plant or animal, that ends upfalling into these structures. Other plants appear tobe mining these same sort of veins, but are notconsidered to be carnivores, although they certainlycould benefit from the decay of animals that happento be among the items they collect in either ‘trashbasket’ systems of roots (as in some orchids, includingspecies of Ansellia Lindl., Catasetum Rich. ex Kunthand Grammatophyllum Blume) or sterile leaves (as inthe ferns Platycerium Desv. and Drynaria (Bory)J.Sm.; Polypodiaceae).
The carnivorous syndrome in angiosperms haslargely been focused on the plants that are clearlytrapping, digesting and absorbing the resulting prod-ucts, such as Drosera, Utricularia and Genlisea A.St.-Hil., but many authors have noted that other speciesmay be going about the same business in a much lessobvious way. Darwin (1875) noted that many plantshave developed sticky glands that trap and killinsects, such as Erica tetralix L., Mirabilis longifloraL., Pelargonium zonale L’Hér., Primula sinensis Lour.and Saxifraga umbrosa L., but no one has furtherinvestigated these species to determine if somethingmore sinister is in fact taking place. Some of these,such Erica tetralix, grow in nutrient-poor soils and
†As in the movie, Finding Nemo, about a clownfish that livesin the tentacles of sea anemones without suffering from theirstings.
MURDEROUS PLANTS 333
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

are members of families related to those of carnivores(in this case, Sarraceniaceae), so they are likely can-didates for investigation. Earlier in the 20th century,several Italian researchers (Mameli, 1916; Mameli &Aschieri, 1920; Zambelli, 1929; see discussion of thesepapers by Simons, 1981) suggested that yet otherplants (Martynia L., Lychnis L. and Petunia Juss.,respectively) with sticky leaves were in fact digestingprey, although the experiments to support thesecontentions were not particularly sophisticated andtoday would be considered inconclusive.
In this part of our review, we focus upon (1) thephylogenetic arrangement of the families thatmost authors have thought to be carnivorous, (2)the degree of modifications made to enable carnivory(by describing the mechanisms by which prey aretrapped, killed and digested) and (3) the other generaof plants that have been hypothesized by at leastsome authors to be carnivorous and discuss theevidence for this.
PHYLOGENETIC RELATIONSHIPS AND
MODERN INSIGHTS
As stated above, Darwin assumed that some parallel-isms were likely among trapping types. Tank-formingplants such as the bromeliads Brocchinia and Catop-sis aside (because they inherited the trapping struc-ture from their ancestors and merely developedgreater absorptive capacities in order to becomecarnivorous), pitcher plants evolved three times inthe angiosperms, in Nepenthaceae (Nepenthes) inCaryophyllales, Cephalotaceae (Cephalotus Labill.) inOxalidales (a rosid order) and Sarraceniaceae (Dar-lingtonia DC., Heliamphora Benth. and Sarracenia)in Ericales (an asterid order), whereas flypaper leavesmay have evolved at least five times: Droseraceae(Drosera), Drosophyllaceae (Drosophyllum Link),Dioncophyllaceae (Triphyophyllum Airy Shaw), all inCaryophyllales, and Byblidaceae (Byblis Salisb.) andLentibulariaceae (Pinguicula L.), both in Lamiales(an asterid order). Most authors would add Roridu-laceae (Roridula) to this list, but, although the twospecies of this genus clearly trap insects, they neitherdigest nor absorb the released nutrients with special-ized cells on their leaves (see below). There is anequally parsimonious alternative to this becausethree of these families are closely related; flypapertraps may have evolved once in the common ancestorof Droseraceae, Drosophyllaceae and Dioncophyl-laceae, and then been lost in Ancistrocladus Wall. (thesole genus of Ancistrocladaceae), Habropetalum AiryShaw and Dioncophyllum Baill. (the other two generaof Dioncophyllaceae). Many other species exhibitsimilar capacities to trap insects and other arthro-pods, so classifying Roridula as a carnivore is arbi-
trary. Snap traps probably evolved only once, inDroseraceae in the common ancestor of Aldrovanda L.and Dionaea Ellis (Cameron, Wurdack & Jobson,2002). In Utricularia and Genlisea (Lentibulariaceae),we find two novel types of traps, bladders underpressure (formed on stolons, which are modifiedstems) and spiralled tubular traps (formed by subter-ranean leaves), respectively; Pinguicula in the samefamily has active flypaper traps.
Among angiosperms as a whole, flypaper traps areby far the most numerous, and it is easiest to considera flypaper trap as the logical evolutionary antecedentfor the development of more specialized traps. Oncetrapping/digesting of prey occurs, then further struc-tural modifications/physiological developments toimprove its function or specialize in particular types ofprey are easier to envisage. In nearly all cases of otherspecializations, flypaper traps occur among the relatedgenera, except in the case of Cephalotus (Cephalota-ceae, Oxalidales), which is the most distantly relatedspecies relative to the others known. Other families inOxalidales, for example, Cunoniaceae and Elaeocar-paceae, have mucilage secreting cells in their epider-mis, and a few genera (e.g. Eucryphia Cav.,Cunoniaceae) are glandular. In several other cases,stalked secretory glands are common among otherfamilies in orders with carnivorous taxa, for example,Caryophyllales (Lledó et al., 1998) and Lamiales.
OVERVIEW OF MURDEROUS PLANTS
Below we provide an overview of known carnivorousplants and their phylogenetic relationships, presentedin the order from APG III (2009). Considering thatthere are varying degrees of carnivory exhibited bymany angiosperms, this list will not be exhaustive,and good evidence for carnivory will most likely befound in other plant groups in the future. For eachcarnivorous genus, we provide habitat preferences,mechanisms of trapping insects, glandular modifica-tions to aid digestion and absorption of digestionproducts and an assessment of their degree of activecarnivory.
MONOCOTSPOALES
BromeliaceaeRosette-forming leaves of many bromeliads formtanks, in which they catch rainwater, allowing themto survive dry periods in the canopy of rainforests.Roots of most bromeliads are non-absorptive andfunction only to attach these plants to their substrateor host plant (in the case of the epiphytic species).Absorption by roots is replaced by absorption of min-
334 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

erals and potentially nutrients from their tanks viaspecialized scale glands, and these have been docu-mented to absorb labelled amino acids (reviewed inGivnish et al., 1984). In these ‘pitchers’, whole ecosys-tems can develop, and they are an important breedingground for tree-dwelling frogs in tropical America, aswell as a source of nutrients for the epiphytic speciesin particular.
Brocchinia reducta Baker, a terrestrial speciesfrom southern Venezuela and Guyana, grows insunny, wet, nutrient-poor habitats alongside othercarnivores such as Heliamphora. This species has aspecialized ‘pitcher’ formed by the leaf rosette inwhich water accumulates. The insides of the leavesare covered in waxy scales that reflect ultravioletlight, and many insects are attracted by this com-bination and lured into the trap. Brocchinia reductawas originally thought not to produce enzymes, anddigestion was then considered to be performed by acommunity of inquilines (Givnish et al., 1984). ThepH of the liquid in the pitcher of B. reducta mea-sured 2.8–3.0, so this too would aid in breakingdown material that fell into the liquid. It is knownto contain large numbers of drowned insects andhas recently been demonstrated to produce phos-phatases, albeit weakly (Płachno et al., 2006); thus,it can be considered a carnivorous plant in the strictsense. The trapped insects are broken down, andthe nutrients released are absorbed by the glandu-lar leaf scales. A closely related species, B. hechtio-ides Mez, has also been considered to be similarlycarnivorous, but this species has not been studied sofar in detail (Givnish et al., 1997).
Similarly, Catopsis berteroniana (Schult. &Schult.f.) Mez, an epiphytic species that can be foundfrom Florida to Brazil, appears to trap more insectsthan other bromeliads of a similar size (Frank &O’Meara, 1984). However, Givnish et al. (1984) per-formed a cost–benefit analysis, which showed thatonly plants growing in wet, bright conditions wouldbe likely to make full-blown carnivory a successfulstrategy. Epiphytic species should not be carnivorous,but it is clear that some species growing in dry areascan be carnivores (e.g. Drosophyllum, Drosophyl-laceae, and some Drosera spp., Droseraceae; Caryo-phyllales, see below). Some Nepenthes spp. growing inhumid environments can effectively be epiphytes, andmany grow in a high degree of shade.
Most species of Puya Molina have extremely sharp,outwardly pointing spines, probably to deter herbi-vores. In contrast, the spines of P. raimondii Harmspoint inward towards the rosette. This tree-likeAndean bromeliad is bird pollinated, and several birdspecies also use the rosette as a nest site, with theinward pointing spines providing stability for thenest. Some birds are also trapped by the spines and
killed. Rees & Roe (1978) counted a total of 44 deadbirds in only 17 plants. Because the leaves are curvedinto a trough shape, it may allow P. raimondii tobenefit from the added nutrients provided by run-offof accumulated nest debris and faeces, not to mentiondead birds.
EriocaulaceaePaepalanthus bromelioides Silv. shares the habitat ofBrocchinia reducta, and its leaves also form a tankthat may function in a similar fashion as that ofBrocchinia (Jolivet, 1998). This is, however, highlyspeculative, and no study has been carried out onpossible trapping, digesting or absorbing mechanismsof this species.
EUDICOTSSAXIFRAGALES
SaxifragaceaeDarwin (1875) reported that peduncles and petioles ofSaxifraga umbrosa L. are clothed in pink glands thatsecrete a yellowish viscid fluid, which can rarelyentrap minute Diptera. He proceeded to experimenton flower stems in a manner similar to, although lessextensive than, his studies on Drosera. Dipping theminto infusions of raw meat, he noted movements inthe protoplasm ‘exactly like those described in thecase of Drosera.’ With exposure to solutions of ammo-nium carbonate, he observed that ‘there is the closestresemblance to what takes place when a tentacle ofDrosera is immersed in a weak solution of the samesalt. The glands, however, absorb very much moreslowly than those of Drosera.’ He concluded that moreevidence was necessary ‘before we can fully admitthat the glands of this saxifrage can absorb . . . ani-mal matter from the minute insects which they occa-sionally and accidentally capture.’ For Saxifragarotundifolia L., he observed that the glands were ableto absorb ammonium carbonate much more quicklythan those of S. umbrosa. Darwin stated that ‘Themost interesting case for us [among the plants withglandular hairs that he studied] is that of the twospecies of Saxifraga, as this genus is distantly alliedto Drosera’. Although, molecular phylogenetic studieshave refuted this last statement, it appears that atleast some species of Saxifraga have glands that arecapable of trapping insects and that these sameglands can absorb a range of substances. The exu-dates from the glands have not been demonstrated tohave digestive properties, however, and it is unlikelythat these plants are true carnivores. Their habitatpreferences also do not appear to be those associatedwith the classic cases of carnivory.
MURDEROUS PLANTS 335
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

OXALIDALES
CephalotaceaeCephalotaceae are monotypic, the sole species beingCephalotus follicularis Labill. (Fig. 4A), which is onlyfound in southwestern Western Australia. Generallyknown as the Albany pitcher plant, it is also some-times referred to as the moccasin plant or WesternAustralian or (historically) New Holland pitcherplant. Described by Labillardière on the basis ofmaterial that has apparently been lost (Willis, 1965),further details of the taxonomic history of this speciesare given by McPherson (2009). Although described in1806, it appears to have been unknown to Darwinand is not mentioned in his book, although he didvisit Western Australia, which left him famouslyunimpressed:
FEBRUARY 17TH. – The Beagle sailed from Tasmania, and,on the 6th of the ensuing month, reached King George’sSound, situated near the S.W. corner of Australia. We staidthere eight days; and I do not remember, since leavingEngland, having passed a more dull, uninteresting time.(Darwin, 1839)
Although the pitchers are superficially similar tothose of Nepenthes, Brown (1832) stated that the twogenera ‘differentiate in so many other importantcharacters that they cannot be considered as nearlyrelated.’ Among other characters, Cephalotus Labill.differs from Nepenthes in its hermaphrodite flowers.Like Nepenthes, however, Cephalotus produces itsown digestive enzymes and is thus an active carni-vore (see McPherson, 2009 for further details). It alsogrows in wet, sunny sites where other carnivoroustaxa are common, particularly Drosera spp., includingthe enormous D. gigantea Lindl. Phylogenetic studiesindicate that Cephalotaceae belong to Oxalidales,within which they are related to Brunellia Ruiz &Pav. (Brunelliaceae) and then to Eleaocarpaceae andCunoniaceae; Cephalotus folicullaris is thus only dis-tantly related to any other carnivorous taxa (APG III,2009) and unusual in belonging to a clade in whichglands are not always present. Mucilage-producingepidermal cells or hairs (some genera of Elaeocar-paceae and Eucryphia, Cunoniaceae) do occur in thefamilies of Oxalidales and evidently can serve asantecedents for development of the digestive andabsorptive glands in Cephalotus.
MALPIGHIALES
PassifloraceaePassiflora L. is known for its glands that mimic insecteggs to fool especially butterflies that do not lay eggsif there are already some present on leaves. Similarly,the peculiar, pantropical, weedy passion flower, Pas-siflora foetida L., has long glandular hairs on thebracts subtending its white–green flowers (Fig. 4B).These multicellular, stalked glands that are remark-ably similar to those of taxa in Caryophyllales havebeen proven to produce digestive enzymes (Radha-mani, Sudarshana & Krishnan, 1995), suggestinginsects get trapped in the fine netting of the bractsand are digested. However, there is no mention ofsterilizing the bract surface or otherwise demonstrat-ing that production was endogenous, nor was absorp-tion definitively demonstrated. This may simply beanother case of a sticky floral defence against her-bivory. It does not fit the syndrome of habitat featuresobserved in other carnivorous taxa (Givnish et al.,1984); P. foetida inhabits wet tropical areas in whichcarnivores are otherwise absent. It is also an invasivein tropical ruderal zones worldwide.
ROSALES
RosaceaeMany members of Rosaceae are covered with glandu-lar, often sticky, processes. Insects often becomeentrapped, and it has widely been assumed thatthese glands have a defensive capability. However,members of the Potentilla arguta Pursh complex(sometimes placed in Drymocallis Fourr. ex Rydb.,including also P. glandulosa Lindl. and P. rupestris L.)appear capable of not only trapping insects, but alsodigesting them and absorbing the released nutrients(Spomer, 1999). These species occur in habitats thatcan support other carnivores, especially Drosera, andfit the syndrome of conditions associated with car-nivory, but Spomer (1999) admitted that he could notrule out surface microbes as the source of theenzymes he observed to be digesting insects trappedby the hairs on these species of Potentilla.
GERANIALES
GeraniaceaeAs in the case for Potentilla (Rosaceae; see above),Spomer demonstrated that Geranium viscosissimum
Figure 4. A, Cephalotus follicularis Labill., RBG Kew. B, Passiflora foetida L., showing flower with glandular bracts,Punaauia, Tahiti. C, Dionaea muscipula Ellis ex L., active traps, Carolina Beach State Park, North Carolina. D, Droserapulchella Lehm., with winged petioles, cultivated at RBG Kew. E, Drosera regia Stephens, regular circination of unfoldingleaf, cultivated at RBG Kew. F, Drosophyllum lusitanicum (L.) Link, reverse circination of unfolding leaf, RBG Kew. Allphotographs taken by Maarten J. M. Christenhusz.
�
336 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

MURDEROUS PLANTS 337
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

Fisch. ex C.A.May has the potential to be carnivorous.It appears to secrete digestive enzymes and be capableof absorbing amino acids. As for P. arguta, it cannot beruled out that surface microbes are responsible fordigestion, but, in the same series of experiments,leaves of Petunia did not exhibit this activity, so eitherthe microbes responsible do not occur on Petunia or theenzymes are released only by G. viscossisimum. Thehabitat preferences for this species (moist roadsidesand creek banks, sometimes in shade) do not seem tobe clearly those associated with the carnivorous syn-drome, and no other carnivorous taxa grow with it.Darwin (1875) also mentioned possible carnivory asso-ciated with the glandular hairs on Pelargonium zonaleL’Hér., but no one has ever investigated this further. Asfor G. viscossisimum, this seems an unlikely carnivore,given its habitat preferences.
BRASSICALES
BrassicaceaeCarnivorous seeds are perhaps a strange concept, buttests have been carried out on the seeds of shepherd’spurse, Capsella bursa-pastoris Medik. (Barber, 1978).It was shown that the mucilage layer of seeds uponimbibition contains chemicals that attract soil nema-todes, protozoa and soil bacteria and a toxin that killsthese organisms. The mucilage also produces protein-ases and labelled amino acids were found to be takenup by the seeds. It is easy to imagine that seedlingswould benefit nutritionally from the breakdown ofthese organisms.
CARYOPHYLLALES
CaryophyllaceaeVarious members of the genera Lychnis and Silene L.have sticky hairs and are known by the commonname of ‘catchfly’. Despite this, most modern authorshave given little credence to them being carnivorous.Barthlott et al. (2007), for example, reported thatLychnis viscaria L. is one of the non-carnivorousplants that ‘have developed sticky leaves as a defenseagainst feeding insects’ (along with Nicotianatabacum L. and Passiflora foetida), but continued thatthe insects thus trapped ‘are not digested becausethese plants have no digestive enzymes’. However,Mameli & Aschieri (1920) demonstrated that L. vis-caria ‘without doubt contains a proteolytic enzyme,that renders the proteinous substances slowly soluble’and that ‘the viscous zones of the stalks of the plantsare permeable’. Spomer (1999) demonstrated that thesticky exudates of some other members of Caryophyl-laceae (Cerastium arvense L. and Stellaria americana(Porter) Stanl. and S. jamesiana Torr.) showed pro-tease activity, but he did not investigate uptake of the
products of digestion. Further research would be nec-essary to demonstrate full carnivory in this family.
DioncophyllaceaeDioncophyllaceae consist of three monotypic genera,Dioncophyllum Baillon, Habropetalum Airy Shawand Triphyophyllum Airy Shaw, all lianas or shrubsendemic to tropical West Africa. Molecular studies(e.g. Fay et al., 1997; Cameron et al., 2002; Cuénoudet al., 2002; Heubl, Bringmann & Meimberg, 2006)have shown them to be most closely related to Ancis-trocladaceae, with a single genus AncistrocladusWallich consisting of 12 species of lianas in tropicalAfrica and Indomalaysia. This pair of families is mostclosely related to Drosophyllaceae and then moredistantly to Droseraceae, Nepenthaceae and othercaryophyllalean families.
Green, Green & Heslop-Harrison (1979) demon-strated that Triphyophyllum peltatum (Hutch. &Dalziel) Airy Shaw is carnivorous. The other twogenera of Dioncophyllaceae and Ancistrocladus lackthe carnivorous habit. Dioncophyllum appears to beeglandular, and the glands in Habropetalum aresimple and present only on young stems. This lack ofcarnivory probably represents losses in these threegenera, given the relationship to Drosophyllaceae andother carnivorous plant families (e.g. Heubl et al.,2006).
Triphyophyllum is heterophyllous, producing threetypes of leaves, two generally found on non-climbing,juvenile shoots and the third on adult climbing shoots.Short shoots firstly produce mostly eglandular oblan-ceolate leaves (Fig. 5A), but, just before the height ofthe rainy season, glandular filiform leaves (Fig. 5B)are produced, which are relatively short-lived, surviv-ing for only a few weeks. As in Drosophyllum, theseshow reverse circinnation. The glandular leaves bearstalked and sessile glands (Fig. 5C) and are efficient intrapping prey (Fig. 5B; mostly insects, with Coleopterabeing the most common). Large numbers of prey aretrapped: Green et al. (1979) found c. 21 identifiablecarcases per leaf plus other chitinous remains. Occa-sional leaves intermediate in form between these twotypes are found on the juvenile shoots (see for exampleMcPherson, 2008). The leaves of the mature axis areoblanceolate and have apical hooks (Fig. 5D), enablingthe liana to climb into the canopy. The account ofTriphyophyllum by McPherson (2008) included manymore photographs illustrating the different leaf typesand other features, including the prominently wingedpink seeds.
The glandular leaves possess stalked and sessileglands. The stalked glands fall into two size classes(2–3 mm and c. 1 mm) and bear secretion dropletsthat trap prey. The sessile glands are dry untilstimulated. The gland stalks are vascularized, as in
338 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

Figure 5. Triphyophyllum peltatum (Hutch. & Dalziel) Airy Shaw. A, eglandular leaves on juvenile shoots. B, glandularleaves on a juvenile shoot. C, close-up of glandular leaf showing two types of glands. D, mature axis with grappling hooks.Photographs taken by Stewart McPherson (2008).
MURDEROUS PLANTS 339
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

Drosera and Drosophyllum, and also in Passifloraceae(including Turneraceae), but this condition is other-wise unknown in the angiosperms. Green et al. (1979)stated that the stalked glands are ‘the most anatomi-cally elaborate known in the plant kingdom’. Theyalso demonstrated that the secretions of these glandscontain proteases and other enzymes.
Juniper, Robins & Joel (1989) referred to Triphyo-phyllum as the ‘part-time carnivorous plant’. It isunusual among carnivores in being well rooted andhas only a transient carnivorous phase. Green et al.(1979) reported that there is evidence that Triphyo-phyllum leaves are relatively rich in potassium,which is deficient in the surrounding soil, and theywrote ‘It seems possible that the transition from thejuvenile phase requires that some threshold levelshould be reached in nutrient reserves, and in theabsence of adequate soil resources carnivory could bethe means for gaining this threshold more rapidly.’They continued ‘The limited growth of the short shootis accompanied by an alternation of groups of long-lasting photosynthetic leaves and shorter-lived insect-capturing leaves. This growth pattern might beexpected to permit the hoarding of nutrients until themoment was reached when a switch could be made tothe development of the rapidly growing, efficientlyclimbing liane.’ More recent studies in the field byMcPherson (2008), however, indicated that plants areable to switch between primary and secondary foliagetypes more than once in their life cycle.
Carnivory appears to be well established in the caseof Triphyophyllum, although it grows in rather atypi-cal shady environments, not generally associated withcarnivorous plants (Juniper et al., 1989). Green et al.(1979) suggested that a parallel with Triphyophyllummay be seen in epiphytic and climbing Nepenthes,although the ability of Triphyophyllum to grow inshade and the lack of carnivory shown by the leavesof the mature liana clearly distinguish it frommembers of that genus.
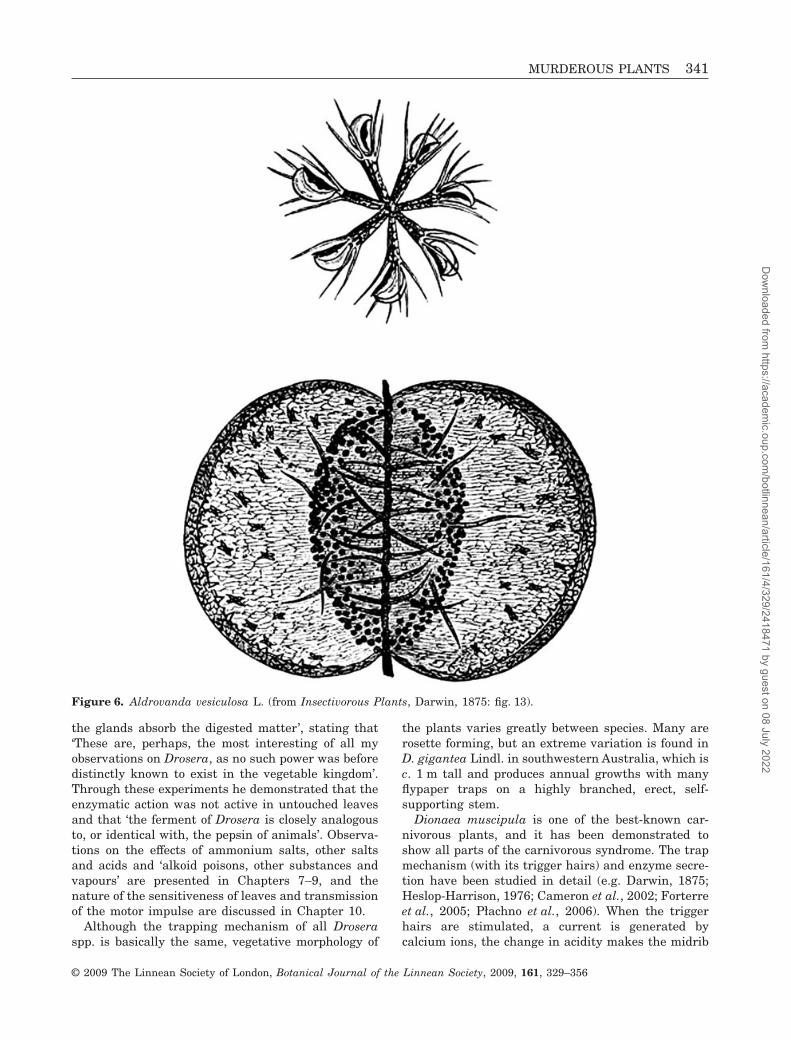
DroseraceaeDroseraceae comprise three genera, Aldrovanda,Dionaea and Drosera. Aldrovanda (one species) is awidespread aquatic in the Old World, and Dionaea(also a single species) is endemic to a small swampyarea in North and South Carolina. Drosera (morethan 180 species), in contrast, is extremely wide-spread, occurring on all continents except Antarctica.Notable radiations occur in Australia and SouthAfrica. Although Darwin (1875) considered Droser-aceae to include six genera (Aldrovanda, Byblis,Dionaea, Drosera, Drosophyllum and Roridula), phy-logenetic studies have demonstrated that Byblis,Drosophyllum and Roridula should be excluded, withDrosophyllum being more closely related to the other
caryophyllalean families (Ancistrocladaceae, Dionco-phyllaceae and Nepenthaceae) and Byblis andRoridula being members of Lamiales and Ericales,respectively. In spite of this narrowing of their cir-cumscription, Droseraceae nevertheless demonstrateremarkable diversity in morphology, specifically intheir mode of carnivory, with two major forms oftraps: flypaper traps in Drosera and snap traps inAldrovanda (Fig. 6) and Dionaea (Fig. 4C). Cameronet al. (2002) showed that Aldrovanda is sister toDionaea and this pair is sister to Drosera (Fig. 4D, E),but with relatively weak support for this pattern ofrelationships.
In analyses using rbcL sequence data alone, Riva-davia et al. (2003) with greater species samplingplaced Aldrovanda as sister to D. regia (Fig. 4E), butwith no support for these relationships. In theiranalyses of rbcL in combination with sequences of18S nuclear ribosomal DNA, however, they recovereda weakly supported monophyletic Drosera, and Aldro-vanda was moderately supported as sister to Dionaea.Heslop-Harrison (1976) referred to the traps ofDrosera as being passive and those of the other twogenera as active, although the power of movementdemonstrated by Darwin (1875) in Drosera leavesindicates that these are also active, if somewhatslower in their response.
Darwin (1875) devoted the great part of his book oncarnivorous plants to a series of experiments onDrosera, the first ten chapters (out of 18) dealingalmost exclusively with Drosera rotundifolia L.(Fig. 7), the common sundew, the eleventh summariz-ing the previous ten chapters and the twelfth detail-ing observations on other species of Drosera. Chapters13 and 14 cover his observations on Dionaea mus-cipula and Aldrovanda vesiculosa L., respectively.
Through his work on D. rotundifolia, Darwin dem-onstrated that the ‘tentacles’ are capable of rapidmovement in response to stimuli (beginning after aslittle as 10 seconds), the glands are absorptive andthat stimulating one tentacle caused movement inthe surrounding tentacles (Chapter 1). In Chapter 2,he showed that animal substances caused a muchquicker and stronger response than inorganic sub-stances or mechanical irritation and that movementonly occurred if a tentacle was touched briefly threeor more times. In Chapter 3, he described the processof aggregation of cellular contents in the glands afterexcitation. Chapter 4 deals with the effects of immer-sion in water at different temperatures and Chapter 5with the effect of nitrogenous and non-nitrogenousorganic fluids (comparing, for example, the effects ofdecoctions of green peas or cabbage with infusionsof raw meat). In Chapter 6, Darwin described theexperiments by which he demonstrated that ‘theleaves are capable of true digestion, and that
340 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022
the glands absorb the digested matter’, stating that‘These are, perhaps, the most interesting of all myobservations on Drosera, as no such power was beforedistinctly known to exist in the vegetable kingdom’.Through these experiments he demonstrated that theenzymatic action was not active in untouched leavesand that ‘the ferment of Drosera is closely analogousto, or identical with, the pepsin of animals’. Observa-tions on the effects of ammonium salts, other saltsand acids and ‘alkoid poisons, other substances andvapours’ are presented in Chapters 7–9, and thenature of the sensitiveness of leaves and transmissionof the motor impulse are discussed in Chapter 10.
Although the trapping mechanism of all Droseraspp. is basically the same, vegetative morphology of
the plants varies greatly between species. Many arerosette forming, but an extreme variation is found inD. gigantea Lindl. in southwestern Australia, which isc. 1 m tall and produces annual growths with manyflypaper traps on a highly branched, erect, self-supporting stem.
Dionaea muscipula is one of the best-known car-nivorous plants, and it has been demonstrated toshow all parts of the carnivorous syndrome. The trapmechanism (with its trigger hairs) and enzyme secre-tion have been studied in detail (e.g. Darwin, 1875;Heslop-Harrison, 1976; Cameron et al., 2002; Forterreet al., 2005; Płachno et al., 2006). When the triggerhairs are stimulated, a current is generated bycalcium ions, the change in acidity makes the midrib
Figure 6. Aldrovanda vesiculosa L. (from Insectivorous Plants, Darwin, 1875: fig. 13).
MURDEROUS PLANTS 341
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

cells change shape as a result of osmosis and the trapcloses. After several days the prey is reduced to itschitin skeleton, and the trap reopens for reuse.
The popularity of Venus’ fly traps in cultivation andthe limited area of distribution in the Carolina swampshas resulted in a great threat to the native popula-tions. Currently wild plants are protected, and wildcollections are illegal. Nevertheless, the sites areunder threat from urban development, and the speciesis therefore classified as ‘vulnerable’ (IUCN, 2009).Dionaea muscipula has been introduced to swamps inFlorida, New Jersey, California and Jamaica, wherepopulations have become well established.
Despite the difference in their habitat, the traps ofAldrovanda bear clear similarity to those of Dionaea
(e.g. Juniper et al., 1989), albeit somewhat miniatur-ized, and function in a similar way. Darwin (1875)described Aldrovanda as ‘a miniature, aquaticDionaea’, and Cameron et al. (2002) stated that ‘it isnot difficult to envision a Dionaea-like, terrestrialancestor of Aldrovanda vesiculosa becoming adaptedto a permanently aquatic lifestyle’.
Darwin’s fascination with carnivorous plants carriedforward into his studies of the power of movement inplants (Darwin, 1880), with observations on Dionaea,Drosera and Sarracenia. With reference to alteredbehaviour because of carnivory, he stated that:
Heliotropism prevails so extensively among the higher plants,that there are extremely few, of which some part, either the
Figure 7. Drosera rotundifolia L. (from Insectivorous Plants, Darwin, 1875: fig. 1).
342 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

stem, flower-peduncle, petiole, or leaf, does not bend towardsa lateral light. Drosera rotundifolia is one of the few plantsthe leaves of which exhibit no trace of heliotropism. Nor couldwe see any in Dionaea, though the plants were not so carefullyobserved. Sir J. Hooker exposed the pitchers of Sarracenia forsome time to a lateral light, but they did not bend towards it.We can understand the reason why these insectivorous plantsshould not be heliotropic, as they do not live chiefly by decom-posing carbonic acid; and it is much more important to themthat their leaves should occupy the best position for capturinginsects, than that they should be fully exposed to the light.
DrosophyllaceaeDrosophyllaceae include one species, Drosophyllumlusitanicum (L.) Link (Fig. 4F), native to the IberianPeninsula (Atlantic coast of Portugal, the southern-most tip of Spain and Gibraltar) and northern Africa(northernmost Morocco). Its common names are Por-tuguese sundew and dewy pine. It is unusual amongcarnivorous taxa in that it grows in dry basic ratherthan wet acidic soils, but the sites it inhabits arenonetheless nutrient deficient. No other carnivoroustaxa grow in these habitats. It was originallydescribed as a species of Drosera by Linnaeus inSpecies Plantarum (Linnaeus, 1753), and it doesresemble the species of that genus, except for itsreversely circinnate leaves (Fig. 4F; compare withFig. 4E) that do not move when capturing prey. Itforms an often stalked rosette of leaves covered withsticky, mucilage-secreting, long glands that trap prey(flypaper traps); plants reach approximately 40 cmin height. Once trapped, insects are dissolved byenzymes released by short glands on the leaves, andthe released nutrients are absorbed by the plant. It isthus an active carnivore because it has all the cellularmechanisms to trap prey, digest them and absorbnutrients. Molecular phylogenetic studies have dem-onstrated that Drosophyllum is more closely relatedto Dioncophyllaceae (see above), Ancistrocladaceae(not carnivorous) and probably Nepenthaceae than itis to Droseraceae (all Caryophyllales; Fay et al., 1997;Cuénoud et al., 2002). It was known to Darwin, whodrew sketches of the two types of glands on its leaves.
NepenthaceaeNepenthaceae include only one genus, Nepenthes L.,and the most recent major treatment lists 120 speciesand five ‘incompletely diagnosed taxa’ (McPherson,2009). The author of this work stated that
I have prepared this two volume work to provide a visuallyrich overview of the biology, ecology, diversity, distribution andconservation status of Nepenthes and Cephalotus. This work isnot intended as a botanical or taxonomic monograph
and referred readers to a number of ‘appropriateprinted sources’ for more detailed taxonomic treat-
ments. Despite this disclaimer, this work (reviewed byFay, 2009) represents the most complete work pub-lished in recent years, and the other works cited areregional treatments. The short account below islargely based on that of McPherson (2009).
Nepenthes is native to a large part of the Old WorldPalaeotropics (but not Africa), with the vast majorityof the species occurring on the Sunda Shelf in south-eastern Asia. Borneo, Sumatra, Sulawesi and thePhilippines are home to notable radiations of species.Madagascar, the Seychelles, Sri Lanka, northernIndia, Papua New Guinea, northeastern Australia,New Caledonia and some isolated Pacific islands arehome to small numbers of species. Nepenthes speciesgenerally grow in thin mats of leached organicmatter, which can overlay alkaline, neutral or acidsubstrates.
Although known from Madagascar since the 17th
century, Linnaeus (1753) based his description ofNepenthes on material from Sri Lanka, with a singlespecies, N. distillatoria L. Common names are tropi-cal or Asian pitcher plants and, for some species,monkey pots. McPherson (2009) listed a wide range ofcommon names in other languages.
Seedlings of Nepenthes form rosettes and, in themajority of species, plants then form a scrambling orclimbing stem. The traps of Nepenthes are pitchers(Fig. 8) formed from tendrils [similar in developmentto the grappling hooks in Triphyophyllum (Dionco-phyllaceae; see above)]. All leaves can bear tendrils orpitchers and, when growing conditions are not ideal,the tendrils fail to develop into pitchers. The pitchersare borne on the end of tendrils of variable length(Fig. 9A) and are hollow vessels with a mouth intowhich prey can fall. The size of the pitchers variesbetween species from only a few centimetres in lengthto > 30 cm, and they can hold three or more litres (formore details about the morphology of Nepenthes, seeMcPherson, 2009).
The pitchers are (at least in part) lined with secre-tory glands that produce the digestive liquid, and theupper part of the interior of the pitcher is generallysmooth and waxy. The digestive process is thought tobe partly because of the enzymes secreted in theliquid in the pitchers and partly because of micro-organisms in the liquid. In newly opened pitchers, theliquid can be highly acidic (pH 2.5; Heslop-Harrison,1976). Nepenthes species are thus active carnivoresbecause they trap prey, digest them and absorb nutri-ents. This active carnivory was already known in the19th century. In his sole reference to Nepenthes,Darwin (1875) reported that:
Dr Hooker likewise found that, although the fluid within thepitchers of Nepenthes possesses extraordinary power of diges-tion, yet when removed from the pitchers before they have
MURDEROUS PLANTS 343
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

Figure 8. Nepenthes eymae Shigeo Kurata, cultivated at RBG Kew. Photograph taken by Maarten J. M. Christenhusz.
344 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

been excited and placed in a vessel, it has no such power,although it is already acid; and we can account for this factonly on the supposition that the proper ferment [enzyme] isnot secreted until some exciting matter is absorbed.
Prey items are predominantly invertebrates,although occasional cases of the remains of amphib-ians and even small mammals being found in pitchershave been reported. Specialisation on particular typesof prey has been reported (Moran et al., 2001), andsome species appear to have forgone carnivory alto-gether. In addition to prey items, pitchers of Nepen-thes can provide a home for various micro-organisms,invertebrates and even vertebrates, which may takeadvantage of the trapping of other animals by theplants (see McPherson, 2009 for extensive review ofthe fauna of Nepenthes pitchers).
NyctaginaceaeDarwin (1875) reported that the stems and leavesof Mirabilis longiflora bear viscid hairs, and youngplants ‘caught so many minute Diptera, Coleoptera,and larvae, that they were quite dusted with them’.On the basis of experiments with ammonium carbon-ate and infusions of meat he concluded, however, thatthe glands on these hairs had no power of absorption.His final words on the subject were:
We may further infer that the innumerable insects caught bythis plant are of no more service to it than are those whichadhere to the deciduous and sticky scales of the leaf-buds ofthe horse-chestnut [Aesculus hippocastanum L.].
Claims that the seeds of Pisonia grandis R.Br. trapand kill seabirds for the added nutritional benefit ofgerminating near a decomposing corpse were investi-gated by Burger (2005). His study was based onnumerous reports of dead birds that apparently diedafter being entangled in the sticky infructescences ofthis coastal tree species. Burger found that anypossible benefit to the germinating plants was out-weighed by damage caused by scavenging crabs thatwere attracted to the decomposing birds. The killingof birds by P. grandis seeds, however morbid, seemsmore an adaptation to distribute seeds effectively tovarious oceanic islands than an actual adaptation forcarnivory.
PlumbaginaceaeLike Passiflora foetida, species of leadwort, PlumbagoL., have sticky, multi-celled, vascularized, glandularhairs in the inflorescence that capture many insects.Plumbago capensis Thunb. has been suggested to becarnivorous, but no evidence for protease and uptakewas shown (Rachmilevitz & Joel, 1976). The glandsare only produced in the inflorescence and are prob-ably more of a defence mechanism against herbivory
or a means to attach seeds to animals, aiding in theirdispersal. If at all carnivorous, Plumbago shows apassive form of proto-carnivory.
ERICALES
EricaceaeAlthough Darwin (1875) mentioned the possibilitythat Erica tetralix could be carnivorous because of ithaving glandular hairs that secrete ‘viscid matter’and occasionally catch minute insects, he showed thatthe glands have little or no power of absorption. Weare not aware of any further publications providingevidence of carnivory in this species.
PrimulaceaeIn Primula sinensis Lour., the flower stems, leavesand petioles are clothed in longer and shorter hairs,and the longer hairs have an enlarged terminal cell,forming a gland which secretes a variable amountof thick, slightly viscid, not acid, brownish–yellowmatter. Exposure to solutions and vapours of ammo-nium carbonate led Darwin (1875) to state that ‘inboth cases there could hardly be a doubt that the salthad been absorbed chiefly or exclusively by theglands’. Weak infusions of meat, however, had noeffect. Thus, although the glands appear to be capableof absorption of some material, there is no evidencefor uptake of nutrients or digestion.
RoridulaceaeThe single genus in this family Roridula consists oftwo species of South African subshrubs, R. dentata L.and R. gorgonias Planch. The leaves resemble those ofDrosera in being covered in long glandular hairs andexcrete a sticky sap that traps insects (Fig. 9B).Despite its morphological resemblance to sundews,they lack the specialized absorptive glands on theleaves that other carnivorous plants have, permittingthe uptake of dissolved prey, suggesting that this is nota fully carnivorous species. However, two species ofhemipteran, Pameridea roridulae and P. marthothii,are closely associated with Roridula, living their entirelives on these plants (Dolling & Palmer, 1991) withoutgetting trapped. It was initially thought that thesebugs intercepted the nutrients within trapped preybefore it could be absorbed by the plant. Ellis &Midgley (1996) studied this relationship further andfound that the faeces of Pameridea played an impor-tant role. This was the first account of a mutualistrelationship between a carnivorous plant and aninsect, in which Pameridia benefited from the trappedprey and Roridula benefited from the indirect diges-tion of nutrients. Anderson (2005) investigated themethod of absorption in Roridula and found absorptionthrough unusual cuticular gaps, which is most likely
MURDEROUS PLANTS 345
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

346 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

the way the nutrients from the Pameridia faeces areabsorbed by Roridula.
The population density of Pameridia on plants ofRoridula is negatively correlated with the density ofthe spider Synaema marlothi, a species that feedson both trapped prey and Pameridia (Anderson &Midgley, 2002). Pameridia also sucks the sap fromRoridula, and this herbivory reduces the benefit ofthe nutrients when the population of Pameridiabecomes too large. Synaema marlothii is important inthis mutualistic triangle to stabilize the populationsof Pameridia and maintain the balance (Anderson &Midgley, 2007).
SarraceniaceaeSarraceniaceae include three genera distributedmostly in North America (Sarracenia L., with c. 11species in eastern and northern North America and themonotypic Darlingtonia Torr. in the far west) with adisjunct distribution in northern South America (Heli-amphora Benth., Fig. 10, with more than 15 species,and more being described as additional tabletop moun-tains, tepuis, are investigated). The plants of all threegenera grow in nutrient-poor, acid bogs, and only thespecies of Sarracenia are full-fledged carnivores,exhibiting wax scales on the upper surfaces thatincrease the chances that an insect will lose its footingand fall into the pitcher, which produces digestiveenzymes and absorbs the released nutrients. Heliam-phora tatei Gleason, which is also unusual in theotherwise herbaceous family in being a shrub (up to4 m tall), has also been found to have wax scales andproduce digestive enzymes (Jaffe et al., 1992), so com-plete carnivores do exist in Sarrraceniaceae outsideSarracenia. Some species of Sarracenia also producenectar around the rim of the opening (reputed tocontain coniine, a narcotic, in S. flava L.; Fig. 9C, andnectar production appears to be more important inattracting prey than does the colour of pitchers(Bennett & Ellison, 2009; Schaefer & Ruxton, 2008)).Darlingtonia, like most species of Heliamphora, is apassive carnivore and depends on bacteria in thepitcher to digest prey, which it then absorbs.
In terms of phylogenetic relationships, Heliam-phora and Sarracenia are sister taxa, with Darling-tonia sister to this pair (Bayer, Hufford & Soltis,1996). This makes the biogeographical relationshipsmore difficult to explain, most authors having hypoth-esized a South American origin for the family
(Maguire, 1970; Juniper et al., 1989), and suggeststhat Darlingtonia has been an isolated taxon for arelatively long time. Some authors had also thoughtthat Heliamphora was the most primitive morpho-logically, but this does not immediately mean that itshould be sister to the other two genera. In any case,most features of Sarraceniaceae do not occur in any ofthe outgroup taxa, including Roridula, which mostanalyses have placed as sister to Sarraceniaceae(Albert et al., 1992; Chase et al., 1993) making polar-ization of morphological characters problematic.
[See Droseraceae, above, for discussion of lack ofheliotropism in Sarracenia (Darwin, 1880).]
GENTIANALES
GentianaceaeSaccifolium bandeirae Maguire & J.M.Pires, oftentreated as the sole member of Saccifoliaceae, has beenshown to be a member of Gentianaceae by Struweet al. (1998) and Thiv et al. (1999). This species hasstrange pouch-like leaves, leading to suggestions thatit might be carnivorous, but with no convincingevidence having been presented. Struwe et al. (1998)discussed the possibility of it being a ‘carnivorousgentian’, concluding that its position in the asteridsmade this a possibility, but they then indicated thatfurther research to demonstrate digestion and absorp-tion would be necessary. The opening to the leavesis, however, downward facing, so it seems highlyunlikely that the pouches could be effective trappingstructures.
LAMIALES
ByblidaceaeLong thought to comprise two species (Fig. 9D, E)endemic to Western Australia (although the firstknown collection was made in Queensland by JosephBanks et al. during Captain Cook’s first visit to Aus-tralia in 1770; McPherson, 2008), Byblidaceae nowconsist of seven species in the genus Byblis. Of these,two are perennial, occurring in Western Australianear Perth, and five are annuals distributed acrosstropical northern parts of Australia, with one speciesalso occurring on New Guinea (McPherson, 2008).
The superficial resemblance of Byblis to Drosophyl-lum led many to treat them as related, particularlyas the petals appear to be free. Close inspection,
Figure 9. A, Nepenthes sp., unfolding pitcher at the tip of a climbing tendril, cultivated at RBG Kew. B, Roridulagorgonias Planch. with a trapped bluebottle (Calliphora vomitoria L.), cultivated at RBG Kew. C, Sarracenia flava L.,Carolina Beach State Park, North Carolina. D, Byblis liniflora Salisb., showing the glands, cultivated at RBG Kew. E,Byblis gigantea Lindl., habit, near Perth, Western Australia. F, Proboscidea louisianica Thell., Turku Botanical Garden,Finland. Photographs A–D and F taken by Maarten J. M. Christenhusz. Photograph E taken by Michael F. Fay.�
MURDEROUS PLANTS 347
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

Figure 10. Heliamphora nutans Benth., cultivated at RBG Kew. Photograph taken by Maarten J. M. Christenhusz.
348 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

however, reveals that the petals of Byblis are basallyfused (e.g. Lloyd, 1942). Phylogenetic studies haveindicated placement in Lamiales, although with nostrong support for interfamilial relationships withinthat order (e.g. Müller et al., 2004, 2006).
Darwin (1875), on the basis of a dried specimen ofByblis gigantea Lindl. (Fig. 9E) sent from Kew, notedtwo forms of glands on the leaves, ‘sessile onesarranged in rows, and others supported on moder-ately long pedicels’. The latter type were
far more simple in structure than the so-called tentacles of thepreceding genera [of Droseraceae in the broad sense, i.e.Aldrovanda, Dionaea, Drosera, Drosophyllum], and they donot differ essentially from those borne by innumerable otherplants. . . . As no instance is known of unicellular structureshaving any power of movement, Byblis, no doubt, catchesinsects solely by the aid of its viscid secretion. These probablysink down besmeared with the secretion and rest on the smallsessile glands, which, if we may judge by the analogy ofDrosophyllum, then pour fourth [sic] their secretion and after-wards absorb the digested matter.
Darwin appears to have been more or less con-vinced that B. gigantea was carnivorous, although hewas unable to carry out his normal range of experi-ments to verify digestion and absorption because ofthe lack of living material at his disposal. Elsewherein the book, he stated that:
There can hardly be a doubt that all the plants belonging tothese six genera have the power of dissolving animal matterby the aid of their secretion, which contains an acid, togetherwith a ferment almost identical in nature with pepsin; andthat they afterwards absorb the matter thus digested. This iscertainly the case with Drosera, Drosophyllum, and Dionaea;almost certainly with Aldrovanda and, from analogy, veryprobable with Roridula and Byblis.
Bruce (1905) demonstrated that the sessile glands(but not the stalked glands) had the power to digestegg albumen, although Lloyd (1942) was unable todemonstrate digestion of fibrin. Hartmeyer (1997,1998) was not able to demonstrate enzyme productionby sessile glands of B. liniflora, but more recentstudies have shown that they produce enzymesin B. filifolia (Hartmeyer & Hartmeyer, 2005) andB. liniflora (Płachno et al., 2006; Fig. 9D).
Like Roridula, Byblis has been reported to havepotentially mutualistic relationships with variousarthropods and that at least some insects appear tobe able to move over the surface of the plant withoutbecoming entangled in the mucilage (Lloyd, 1942;McPherson, 2008). However, the dynamics of theserelationships are not well understood.
LentibulariaceaeLentibulariaceae consist of three genera: Utricularia(bladderworts; Fig. 2) that is sister to Pinguicula
(butterworts; Fig. 11), and Genlisea, which is sister tothis pair of genera (Müller et al., 2006). Utriculariaand Pinguicula have a more or less cosmopolitandistribution; Genlisea is only found in tropicalAmerica, Africa and Madagascar.
The morphologically variable genus Utriculariaconsists of approximately 200 species and occursnearly worldwide. It is especially diverse in thetropics, where species grow as aquatics, terrestrials,epiphytes and even vines. In many species, thereare no clear distinctions between the roots, stemsand leaves. Epiphytes (such as the Antillean U.alpina Jacq.) bear succulent tubers that containwater. In the North Temperate aquatic bladder-worts, roots are absent, even in seedlings. Thestems bear appendages that form bladder-like struc-tures with trapdoors (Fig. 2). When a trapdoortrigger is touched by a small animal, the trap opensabruptly, and the animal is sucked into the bladder;the trapdoor then re-closes, and the bladder secretesenzymes and absorbs the products of digestion.Aquatic species form bladders on photosyntheticbranches, and terrestrial species have their bladderson achlorophyllous runners. The traps can range insize from 0.2 mm to 1.2 cm (Taylor, 1989). Aquaticspecies possess bladders that are usually at thelarger limit and can feed on more substantial prey,such as water fleas (Daphnia), nematodes, fish fry,mosquito larvae and even young tadpoles. Manyspecies have chasmogamous and cleistogamousflowers in certain seasons and form buds to survivedry periods or winters. Adamec (2007) demonstratedlow (approaching zero) partial pressures of oxygenin traps of Utricularia and Genlisea and hypoth-esised that the prey may, at least in part, die ofanoxia. Sirová et al. (2009) showed that substantialproportions of newly fixed carbon are exuded intothe trap fluid, possibly as a source of nutrients forthe microbial community living there.
Utricularia humboldtii Schomb. is peculiar in thatit is only found in the water tanks of Brocchiniaspp. (Bromeliaceae) at high elevations. The Austra-lian U. multifida R.Br. and U. tenella R.Br. werepreviously placed in the genus PolypompholixLehm., based on a somewhat different shape of thetrapping bladders. Molecular studies have, however,shown that this and another segregate genus, Bio-vularia Kamienski, are embedded in Utricularia(Jobson et al., 2003).
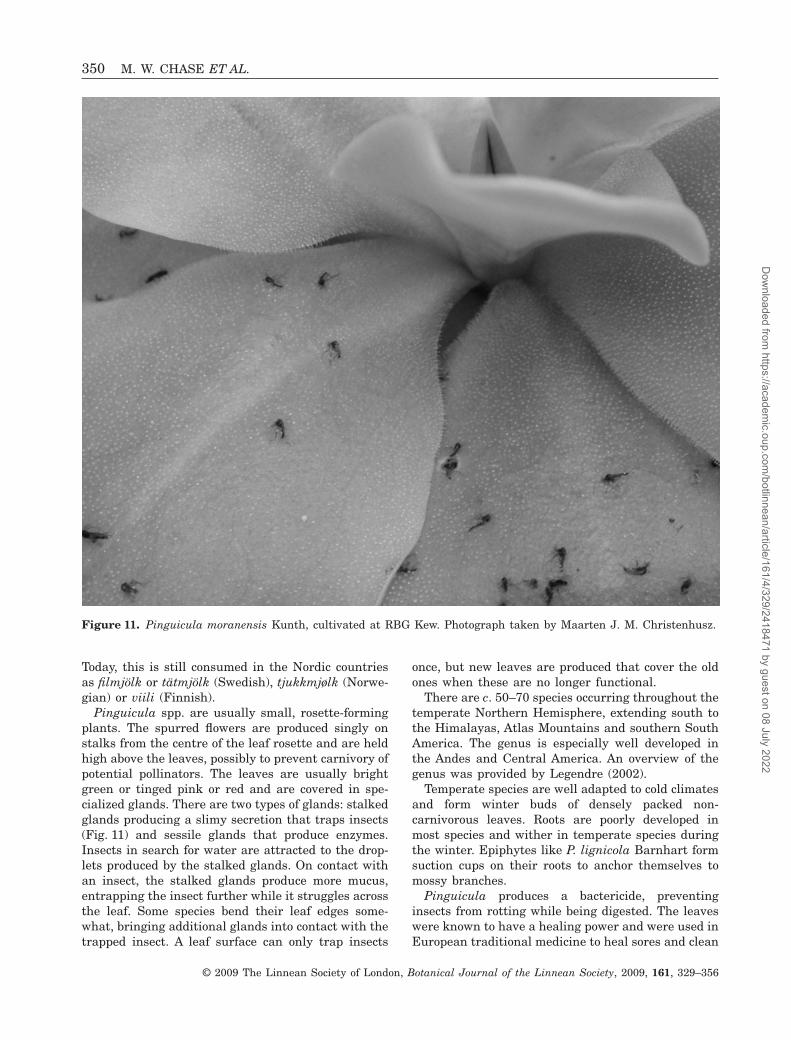
Pinguicula is named for its leaves that are greasyto the touch; pinguis meaning fat in Latin. TheEnglish common name butterwort results either fromthe juice being used on the udders of cows (eitheragainst magic or to treat chapping, etc.; Grigson,1955) or the leaves being used to curdle milk andform a buttermilk-like fermented dairy product.
MURDEROUS PLANTS 349
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022
Today, this is still consumed in the Nordic countriesas filmjölk or tätmjölk (Swedish), tjukkmjølk (Norwe-gian) or viili (Finnish).
Pinguicula spp. are usually small, rosette-formingplants. The spurred flowers are produced singly onstalks from the centre of the leaf rosette and are heldhigh above the leaves, possibly to prevent carnivory ofpotential pollinators. The leaves are usually brightgreen or tinged pink or red and are covered in spe-cialized glands. There are two types of glands: stalkedglands producing a slimy secretion that traps insects(Fig. 11) and sessile glands that produce enzymes.Insects in search for water are attracted to the drop-lets produced by the stalked glands. On contact withan insect, the stalked glands produce more mucus,entrapping the insect further while it struggles acrossthe leaf. Some species bend their leaf edges some-what, bringing additional glands into contact with thetrapped insect. A leaf surface can only trap insects
once, but new leaves are produced that cover the oldones when these are no longer functional.
There are c. 50–70 species occurring throughout thetemperate Northern Hemisphere, extending south tothe Himalayas, Atlas Mountains and southern SouthAmerica. The genus is especially well developed inthe Andes and Central America. An overview of thegenus was provided by Legendre (2002).
Temperate species are well adapted to cold climatesand form winter buds of densely packed non-carnivorous leaves. Roots are poorly developed inmost species and wither in temperate species duringthe winter. Epiphytes like P. lignicola Barnhart formsuction cups on their roots to anchor themselves tomossy branches.
Pinguicula produces a bactericide, preventinginsects from rotting while being digested. The leaveswere known to have a healing power and were used inEuropean traditional medicine to heal sores and clean
Figure 11. Pinguicula moranensis Kunth, cultivated at RBG Kew. Photograph taken by Maarten J. M. Christenhusz.
350 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

wounds. It took until the 19th century for this genus tobe recognized as carnivorous, by Darwin (1875), whowrote:
We thus see that numerous insects and other objects arecaught by the viscid leaves; but we have no right to infer fromthis fact that the habit is beneficial to the plant, any morethan in the before given case of the Mirabilis, or of thehorse-chestnut [Aesculus hippocastanum]. But it will pres-ently be seen that dead insects and other nitrogenous bodiesexcite the glands to increased secretion; and that the secretionthen becomes acid and has the power of digesting animalsubstances, such as albumen, fibrin, &c. Moreover, the dis-solved nitrogenous matter is absorbed by the glands, as shownby their limpid contents being aggregated into slowly movinggranular masses of protoplasm. The same results follow wheninsects are naturally captured, and as the plant lives in poorsoil and has small roots, there can be no doubt that it profitsby its power of digesting and absorbing matter from the preywhich it habitually captures in such large numbers.
Conversely, the role of bacteria in the digestionprocess has been discussed extensively and Lloyd(1942) concluded:
Pinguicula is a carnivorous plant inasmuch as it catches smallinsects and digests them, at least in part, by means of its ownferments. The possible part played by bacteria is not excluded.
The small genus Genlisea counts c. 20 species inthe tropics of the Americas, sub-Saharan Africa andMadagascar. They are small terrestrial or aquaticplants forming a rosette of leaves, and a few singleflowers are held above the rosette that is similar inshape and colour to those of Pinguicula or Utricu-laria. The most peculiar feature of this plant is foundunderground. There, leaves form a pair of capillarytubes joined at the tip in a V-shape; there are spiralgrooves down their lengths allowing soil-borne inver-tebrates and protozoa to enter. Inwardly pointinghairs prevent the prey from escaping; the only waythey can swim is towards the apex of the tube, wherethey are digested (Płachno, Kozieradzka-Kiszkurno &Swiatek, 2007).
It has been suggested that there is a water flow inthese capillary tubes, but this does not seem to bethe case, so an analogy with the digestive tract ofan animal seems misplaced. For example, Heslop-Harrison (1976) wrote:
Genlisea, a genus of the same family [as Utricularia] has adigestive system almost simulating the intestine of ananimal . . . A flow of soil fluid is maintained through the tubeby the secretion of water through the walls of the utricle andtube. The diet seems mainly to be of protozoa, but smallcrustacea and larval forms of other groups are also induced toenter the tube. Undigested remains accumulate in the utricle.
To simulate an animal digestive system it would be necessaryonly to replace the water flow by peristalsis and to provide theutricle with an anus!
Among angiosperms, Genlisea species have thesmallest known genomes (Greilhuber et al., 2006).Those of Pinguicula and Utricularia are larger, butstill also fall among the smallest angiosperm genomes.
MartyniaceaeMartyniaceae are closely related to Pedaliaceae(sesame, Sesamum indicum L., and relatives) andhave been suggested to be carnivorous (Mameli,1916). Martyniaceae occur in semi-dry to desert habi-tats from subtropical North America to tropical SouthAmerica, where the peculiarly shaped fruits are dis-tributed by large mammals, the hooves of which getcaught on the hooks of the fruits. Martynia andProboscidea are herbs with succulent stems, and theplants are covered in glandular hairs that produce afoul-smelling, sticky exudate. Even although the glan-dular hairs, especially of Proboscidea lutea (Lindl.)Stapf. (= Martynia lutea Lindl. = Ibicella lutea (Lindl.)Van Eselt.), catch many insects (Fig. 9F), no uptake ofamino acids has been demonstrated (Rice, 1999;Wallace, McGhee & Biology Class, 1999). Mameli(1916) wrote:
Martynia lutea Lindl. is an insectivorous plant. It catches theinsects by means of an abundant viscous substance, having anacid reaction, secreted by a large number of glandular hairswith which its above-ground parts are covered and by whichit dissolves albuminous substances by a proteolytic enzyme,which has the character of trypsin.
OrobanchaceaeVarious authors in the 19th century (see Groom, 1897)suggested that the glands and capitate hairs found inthe lacunae in the scale leaves of some membersof Orobanchaceae, including Lathraea, might beinvolved in nutrient absorption and entrapment anddigestion of small organisms. Groom (1897) dis-counted this, demonstrating that these structureswere instead involved in water excretion. Despitethis, the suggestion that these plants are potentiallycarnivorous has persisted in the literature (e.g.Heslop-Harrison, 1976; Fitter, 1987; Jolivet, 1998).However, carnivory in Lathraea and its relativesappears to be mere speculation (see Fay, 2009, forfurther details).
SOLANALES
SolanaceaeVarious members of the family have hairs that catchsmall insects. Darwin (1875) reported that Nicotianatabacum is covered with innumerable hairs of
MURDEROUS PLANTS 351
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

unequal lengths, which catch many minute insects,but, in his experiments with infusions of meat andammonium carbonate, he produced no convincingevidence of absorption. Zambelli (1929) statedthat ‘Petunia violacea and Petunia nyctaginiflora areinsectivorous plants’ on the basis of experiments dem-onstrating the presence of proteolytic enzymes, butwas also unable to produce convincing evidence ofabsorption. Spomer (1999) demonstrated that leavesof Solanum tuberosum L. secrete proteinases, butPetunia hybrida Vilm. does not. Simons (1981) sug-gested that crop plants and their relatives with stickyhairs (notably wild tomatoes and potatoes; species ofSolanum L.) are suitable study plants for investiga-tion of ‘degrees of carnivory’.
ASTERALES
StylidiaceaeThe Australian trigger plants of the genus Styl-idium Lour. are known to trap small insects such asgnats and midges using mucilage-secreting glandu-lar hairs on their inflorescences and stems. Becausetrigger plants are remarkable for their active polli-nation mechanism, they had not been examined forcarnivory. However, Stylidium uses the same mecha-nism to trap their pray as Drosera or Byblis, bothgenera that occur in similar habitats and often inthe same places where Stylidium is found. LikeDrosera, the glands produce proteases and can thusdigest the insects caught (Darnowski et al., 2006). Ithas been shown that amino acids are absorbed bythe surface of Stylidium, so some species of triggerplants appear to be fully carnivorous (Darnowski,Moberly & Płachno, 2007). They co-occur withspecies of Drosera in many places, so their habitatpreferences fit the syndrome associated with car-nivorous plants.
DIPSACALES
CaprifoliaceaeThe basal leaves of the common teasel, Dipsacusfullonum L., are united around the stem, forming acup-shaped structure that fills with water after rain.All sorts of debris and insects get trapped in it, butno evidence of digestive enzymes or foliar nutrientabsorption has been revealed. Christy (1923) did notethat the fluid collected in the basin formed by theleaves has a lower surface tension which could be anadaptation to kill prey.
LOSS OF CARNIVORY INCARNIVOROUS GENERA
In some predominantly carnivorous genera such asNepenthes and Utricularia a few detritivorous species
are known. Some Malaysian species, such as Nepen-thes ampullaria Jack and N. lowii Hook.f., and N.pervillei Blume from the Seychelles, catch few insects.Nepenthes ampullaria derives nutrition from fallenleaf litter, whereas the last two appear to obtainnutrients from the droppings of birds and tree shrewsthat feed on the nectaries in the pitchers and snaileggs laid on the rim of the pitchers (Corner, 1978;Clark et al., 2009).
Utricularia purpurea Walter may have partiallylost its appetite for meat. Trapping rates of preywere significantly lower than in other species ofUtricularia. The eastern purple bladderwort canstill trap and digest prey in its traps, but does sosparingly. Instead algae, zooplankton and debris arepresent in the bladders, suggesting that U. pur-purea favours mutualism instead of carnivory (Rich-ards, 2001).
CONCLUDING REMARKS
We think that it is evident from the descriptionsabove that many plant species have the capacityto trap and kill insects and other animals andthat some have refined the carnivorous syndrome toa high degree. Intermediates (the so-called ‘proto-carnivorous’ species) clearly do exist, and it is tempt-ing to consider many of these to be good carnivores.Hartmeyer (1998), referring to species that do notproduce their own digestive enzymes, stated:
The production of enzymes should not be a prerequisite for aplant to be considered carnivorous – a symbiosis with anotherdigesting agent should be sufficient. In the past, symbioseswere mistakenly considered strange exceptions, but now it isapparently a widespread syndrome with carnivorous plants.Indeed, with some plants it is an integrated part of thedigestive system!
Other species exhibit even less specialized cases, butthey too should not be discounted. The cost–benefitmodel for the evolution of carnivory indicates thatthere is a trade-off between the costs of carnivoryand benefits to photosynthetic output (and ulti-mately to enhanced seed production), made possibleby the additional nutrients acquired (Givnish et al.,1984; Benzing, 1987). The model also implicitlyimplies that, in some situations, carnivorous plantsshould have advantages over non-carnivorous plantsin the same habitats. In his review of publisheddata, Ellison (2006) showed that nitrogen, phospho-rus and potassium often limit the growth of carnivo-rous plants, and their use of these elements is20–50% that of non-carnivorous plants. All dataindicated as well that under no circumstances docarnivores have a clear-cut advantage over non-carnivorous plants, and thus it appears that car-
352 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

nivory is in fact a choice of the lesser of two evils indifficult circumstances. Carnivores have poor com-petitive abilities because their lineages becameadapted to living under conditions that were farfrom optimal in terms of nutrient availability, andcarnivory represents a slight improvement overwhat would otherwise be the case. The habitats inwhich carnivores typically occur are those in whichharsh abiotic conditions limit competition betweenspecies, and it is under such abiotic stress that car-nivory represents an improved although certainlynot optimal strategy. If the cost–benefit modelcannot be used to explain why vegetable carnivoryhas been adopted by species with the full-blown syn-drome, then it is difficult to see how it can beapplied in situations where the species are theseintermediate types described above. If mucilage-secreting hairs evolved in response to herbivoryor to attach seeds to animals to aid in their dis-persal, their co-option into providing increasednutrition necessitates minimal additional expense;it involves absorption of nutrients through roots,which happens as a matter of course. This minimallevel of carnivory must surely be beneficial andconfer some slight advantage. It is easy to imaginethat this limited response is further constrained bythe phylogenetic legacy inherited from the condi-tions under which their ancestors evolved and thelimitations imposed by environmental stress in theircurrent habitats. When confronted with two miser-able options, the less miserable wins the day. Full-blown vegetable carnivory obviously threads atwisting trajectory through the maze of cost andbenefit ‘peaks and valleys’ under highly stressfulenvironmental conditions, and these are notoriouslydifficult to model. The ‘intermediate’ cases describedabove are no less difficult to understand, particu-larly when the basic requirements for being carnivo-rous, mucilage-secreting hairs, are ubiquitous, atleast in the eudicots to which the majority of car-nivores and so-called ‘proto-carnivorous’ (i.e. inter-mediate) taxa belong. If carnivory is far morecommon than previously held because of manyspecies being subtly carnivorous, then the back-ground comparisons of ‘carnivore vs. non-carnivore’are also inappropriate because the latter categoryincludes perhaps many species that are subtly car-nivorous through symbioses with other organisms.The evolution of vegetable carnivores requires agreat deal more study, both in terms of better docu-mentation of the putatively intermediate cases aswell as understanding of its evolutionary context,which may be constrained as much by phylogenetichistory as by physiological responses. We may besurrounded by many more murderous plants thanwe think.
REFERENCES
Adamec L. 2007. Oxygen concentrations inside the traps ofthe carnivorous plants Utricularia and Genlisea (Lentibu-lariaceae). Annals of Botany 100: 849–856.
Albert VA, Williams SE, Chase MW. 1992. Carnivorousplants: phylogeny and structural evolution. Science 257:1491–1495.
Anderson B. 2005. Adaptations to foliar absorption of faeces:a pathway in plant carnivory. Annals of Botany 95: 757–761.
Anderson B, Midgley JJ. 2002. It takes two to tango butthree is a tangle: mutualists and cheaters on the carnivo-rous plant Roridula. Oecologia 132: 369–373.
Anderson B, Midgley JJ. 2007. Density-dependent out-comes in a digestive mutualism between carnivorous Ror-idula plants and their associated hemipterans. Oecologia152: 115–120.
APG III. 2009. An update of the Angiosperm PhylogenyGroup classification for the orders and families of floweringplants: APG III. Botanical Journal of the Linnean Society161: 105–121.
Barber JT. 1978. Capsella bursa-pastoris seeds: are they‘carnivorous’. Carnivorous Plant Newsletter 7: 39–42.
Barthlott W, Porembski S, Seine R, Theisen I. 2007. Thecurious world of carnivorous plants. A comprehensive guideto their biology and cultivation. Portland: Timber Press.
Bayer RJ, Hufford L, Soltis DE. 1996. Phylogenetic rela-tionships in Sarraceniaceae based on rbcL and ITSsequences. Systematic Botany 21: 121–134.
Bennett KF, Ellison AM. 2009. Nectar, not colour, may lureinsects to their death. Biology Letters 5: 469–472.
Benzing DH. 1987. The origin and rarity of botanical car-nivory. Trends in Ecology and Evolution 2: 364–369.
Brown R. 1832. On the structure and affinities of Cephalotusfollicularis. Edinburgh Philosophical Magazine 1: 314–317.
Bruce AN. 1905. On the activities of the glands of Byblisgigantea. Notes from the Royal Botanic Garden Edinburgh4: 9–14.
Buel JW. 1887. Sea and land. Philadelphia: H. W. Kelly.Burger AE. 2005. Dispersal and germination of seeds of
Pisonia grandis, an Indo-Pacific tropical tree associatedwith insular seabird colonies. Journal of Tropical Ecology21: 263–271.
Cameron KM, Wurdack KJ, Jobson RW. 2002. Molecularevidence for the common origin of snap-traps among car-nivorous plants. American Journal of Botany 89: 1503–1509.
Chase MW, Soltis DE, Olmstead RG, Morgan D, Les DH,Mishler BD, Duvall MR, Price RA, Hills HG, Qiu Y-L,Kron KA, Rettig JH, Conti E, Palmer JD, Manhart JR,Sytsma KJ, Michael HJ, Kress WJ, Karol KG, ClarkWD, Hedrén M, Gaut BS, Jansen RK, Kim KJ, WimpeeCF, Smith JF, Furnier GR, Strauss SH, Xiang QY,Plunkett GM, Soltis PS, Swensen SM, Williams SE,Gadek PA, Quinn CJ, Eguiarte LE, Golenberg E,Learn JrGH, Graham SW, Barrett SCH, DayanandanS, Albert VA. 1993. Phylogenetics of seed plants: an analy-
MURDEROUS PLANTS 353
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

sis of nucleotide sequences from the plastid gene rbcL.Annals of the Missouri Botanical Garden 80: 528–580.
Christy M. 1923. The common teasel as a carnivorous plant.Journal of Botany 61: 33–45.
Clark MC, Bauer U, Lee CC, Tuen AA, Rembold K,Moran JA. 2009. Tree shrew lavatories: a novel sequestra-tion strategy in a tropical pitcher plant. Biology Letters 5:632–635.
Corner EJH. 1978. The plant life. In: Luping M, Wen C,Dingley ER, eds. Kinabalu: summit of Borneo. Kota Kina-balu: The Sabah Society, 112–178.
Croizat L. 1961. Principia botanica. Caracas: published bythe author.
Cronquist A. 1981. An integrated system of classification offlowering plants. New York: Columbia University Press.
Cuénoud P, Savolainen V, Chatrou LW, Powell M,Grayer RJ, Chase MW. 2002. Molecular phylogeny ofCaryophyllales based on 18S rDNA, rbcL, atpB, and matK.American Journal of Botany 89: 132–144.
Darnowski DW, Carroll DM, Płachno B, Kabanoff E,Cinnamon E. 2006. Evidence of protocarnivory in trigger-plants (Stylidium spp.; Stylidiaceae). Plant Biology 8: 805–812.
Darnowski D, Moberly S, Płachno B. 2007. Triggerplants(Stylidium spp.; Stylidiaceae): a previously unrecognizedgenus of carnivorous plants. Proceedings of the 2007 Botanyand Plant Biology Joint Congress, Botanical Society ofAmerica. Available at http://www.2007.botanyconference.org/engine/search/index.php?func=detail & aid=18 (Abstractaccessed 22 October 2009).
Darwin C. 1839. Narrative of the surveying voyages of HisMajesty’s ships Adventure and Beagle, between the years 1826and 1836, describing their examination of the southern shoresof South America, and the Beagle’s circumnavigation of theglobe. In three volumes, Vol. III. London: Henry Colburn.
Darwin C. 1875. Insectivorous plants. London: John Murray.Darwin C. 1880. The power of movement in plants. London:
John Murray.Dolling WR, Palmer JM. 1991. Pameridea (Hemiptera:
Miridae): predaceous bugs specific to a highly viscid plantgenus Roridula. Systematic Entomology 16: 319–328.
Ellis AG, Midgley JJ. 1996. A new plant–animal mutualisminvolving a plant with sticky leaves and a resident hemi-pteran. Oecologia 106: 478–481.
Ellison AM. 2006. Nutrient limitation and stoichiometry ofcarnivorous plants. Plant Biology 8: 740–747.
Ellison AM, Gotelli NJ. 2009. Energetics and the evolutionof carnivorous plants – Darwin’s ‘most wonderful plants inthe world’. Journal of Experimental Botany 60: 19–42.
Fay MF. 2009. Book review. Botanical Journal of the LinneanSociety 161: 449–450.
Fay MF. 2009. 663. Lathraea clandestina. Orobanchaceae.Curtis’s Botanical Magazine 26: doi: 10.1111/j.1467-8748.2009.01674.x.
Fay MF, Cameron KM, Prance GT, Lledó MD, ChaseMW. 1997. Familial relationships of Rhabdodendron (Rhab-dodendraceae): plastid rbcL sequences indicate a caryophyl-lid placement. Kew Bulletin 52: 923–932.
Fitter A. 1987. Wild flowers of Britain and Northern Europe.London: Collins.
Forterre Y, Skotheim JM, Dumais J, Mahadevan L.2005. How the Venus flytrap snaps. Nature 433: 421–425.
Frank JH, O’Meara GF. 1984. The bromeliad Catopsis bert-eroniana traps terrestrial arthropods but harbors Wyeomyialarvae (Diptera: Culicidae). Florida Entomologist 67: 418–424.
Givnish TJ, Burkhardt EL, Happel RE, Weintraub JD.1984. Carnivory in the bromeliad Brocchinia reducta, witha cost/benefit model for the general restriction of carnivo-rous plants to sunny, moist, nutrient-poor habitats. TheAmerican Naturalist 124: 479–497.
Givnish TJ, Sytsma KJ, Smith JE, Hahn WJ, BenzingDH, Burkhardt EL. 1997. Molecular evolution and adap-tive radiation in Brocchinia (Bromeliaceae: Pitcairnioideae)atop tepuis of the Guayana Shield. In: Givnish TJ, SytsmaKJ, eds. Molecular evolution and adaptive radiation. Cam-bridge: Cambridge University Press, 259–311.
Green S, Green TL, Heslop-Harrison Y. 1979. Seasonalheterophylly and leaf gland features in Triphyophyllum(Dioncophyllaceae), a new carnivorous plant genus. Botani-cal Journal of the Linnean Society 78: 99–116.
Greilhuber J, Borsch T, Müller K, Worberg A, Poremb-ski S, Barthlott W. 2006. Smallest angiosperm genomesfound in Lentibulariaceae, with chromosomes of bacterialsize. Plant Biology 8: 770–777.
Griffith CB. 1960. The little shop of horrors. Film directed byR. Corman. The Filmgroup Inc.
Grigson G. 1955. The Englishman’s flora. London: PhoenixHouse.
Groom P. 1897. On the leaves of Lathraea squamaria and ofsome allied Scrophulariaceae. Annals of Botany 11: 385–398.
Hartmeyer S. 1997. Carnivory of Byblis revisited: a simplemethod for enzyme testing on carnivorous plants. Carnivo-rous Plant Newsletter 26: 39–45.
Hartmeyer S. 1998. Carnivory in Byblis revisited II: thephenomenon of symbiosis on insect trapping plants. Car-nivorous Plant Newsletter 27: 110–113.
Hartmeyer I, Hartmeyer S. 2005. Byblis filifolia als echteKarnivore rehabilitiert. Das Taublatt 3: 4–5.
Heslop-Harrison Y. 1976. Carnivorous plants, a centuryafter Darwin. Endeavour 126: 114–122.
Heubl G, Bringmann G, Meimberg H. 2006. Molecularphylogeny and character evolution of carnivorous plant fami-lies in Caryophyllales – revisited. Plant Biology 8: 821–830.
Hooker JD. 1874. Address to the Department of Zoology andBotany of the British Association, Belfast, August 21, 1874.
Hutchinson J. 1973. The families of flowering plantsarranged according to a new system based on their probablephylogeny, 3rd edn. Oxford: Clarendon Press.
Instructables. 2009. Giant rat-eating plant discovered. Avail-able at http://www.instructables.com/community/Giant-rat-eating-plant-discovered/ (accessed 1 November 2009).
IUCN. 2009. IUCN Red List of threatened species. Version2009.2. Available at http://www.iucnredlist.org/ (accessed 15November 2009).
354 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

Jaffe K, Michelangeli F, Gonzaléz JM, Miras B, RuizMC. 1992. Carnivory in pitcher plants of the genus Heli-amphora (Sarraceniaceae). New Phytologist 122: 733–744.
Jansson T. 1973. Comet in Moominland. Harmondsworth:Puffin Books.
Jobson RW, Playford J, Cameron KM, Albert VA. 2003.Molecular phylogenetics of Lentibulariaceae inferred fromplastid rps16 intron and trnL-F DNA sequences: implica-tions for character evolution and biogeography. SystematicBotany 28: 157–171.
Jolivet P. 1998. Interrelationship between insects and plants.Boca Raton: CRC Press.
Juniper BE, Robins RJ, Joel DM. 1989. The carnivorousplants. London: Academic Press.
Legendre L. 2002. The genus Pinguicula L. (Lentibulari-aceae): an overview. Acta Botanica Gallica 141: 77–95.
Ley W. 1955. Salamanders and other wonders. New York:Viking Press.
Linnaeus C. 1753. Species plantarum. Stockholm: Lauren-tius Salvius.
Lledó MD, Crespo MB, Cameron KM, Fay MF, ChaseMW. 1998. Systematics of Plumbaginaceae based upon cla-distic analysis of rbcL sequence data. Systematic Botany 23:21–29.
Lloyd FE. 1942. The carnivorous plants. Waltham: ChronicaBotanica.
McPherson S. 2008. Glistening carnivores. The sticky-leavedinsect-eating plants. Poole: Redfern Natural HistoryProductions.
McPherson S. 2009. Pitcher plants of the Old World. Poole:Redfern Natural History Publications, 2 vols.
Magee J. 2007. The art and science of William Bartram.Philadelphia: The Pennsylvania State University Press.
Maguire B. 1970. On the flora of the Guayana Highland.Biotropica 2: 85–100.
Mameli E. 1916. Ricerche anatomiche, fisiologiche e biolog-iche sulla Martynia lutea Lindl. Atti dell’Istituto Botanicodella Università di Pavia, Series 2 16: 137–188.
Mameli E, Aschieri E. 1920. Ricerche anatomiche e bio-chimiche sul Lychnis viscaria Linn. Atti dell’Istituto Bota-nico della Università di Pavia, Series 2 17: 119–129.
Martel Y. 2001. Life of Pi. New York: Knopf Canada.Milner R. 2009. Darwin’s universe: evolution from A to Z.
Berkeley: University of California Press.Moran JA, Merbach MA, Livingston NJ, Clarke CM,
Booth WE. 2001. Termite prey specialization in the pitcherplant Nepenthes albomarginata–evidence from stableisotope analysis. Annals of Botany 88: 307–311.
Müller K, Borsch T, Legendre L, Porembski S, BarthlottW. 2006. Recent progress in understanding the evolution ofcarnivorous Lentibulariaceae (Lamiales). Plant Biology 8:748–757.
Müller K, Borsch T, Legendre L, Porembski S, TheisenI, Barthlott W. 2004. Evolution of carnivory in Lentibu-lariaceae and the Lamiales. Plant Biology 6: 477–490.
Nelson EC. 1990. Aphrodite’s mousetrap: a biography ofVenus’s flytrap with facsimiles of an original pamphlet and
the manuscripts of John Ellis, F. R. S. Aberystwyth: Boet-hius Press.
Osborn CS. 1924. Madagascar, land of the man-eating tree.New York: Republic Publishing Company.
Płachno BJ, Adamec L, Lichtscheidl IK, Peroutka M,Adlassnig W, Vrba J. 2006. Fluorescence labelling of phos-phatase activity in digestive glands of carnivorous plants.Plant Biology 8: 813–820.
Płachno BJ, Kozieradzka-Kiszkurno M, Swiatek P. 2007.Functional ultrastructure of Genlisea (Lentibulariaceae)digestive hairs. Annals of Botany 100: 195–203.
Pokémon. 2009. The official Pokémon website. Available athttp://www.pokemon.com/ (accessed 1 November 2009).
Rachmilevitz T, Joel DM. 1976. Ultrastructure of the calyxglands of Plumbago capensis Thunb. in relation to theprocess of secretion. Israel Journal of Botany 25: 159–168.
Radhamani TR, Sudarshana L, Krishnan R. 1995.Defense and carnivory: dual role of bracts in Passiflorafoetida. Journal of Biosciences 20: 657–664.
Rees WA, Roe NA. 1978. Puya raimondii (Pitcairnioideae,Bromeliaceae) and birds: an hypothesis on nutrientrelationships. Canadian Journal of Botany 58: 1262–1268.
Rice B. 1999. Testing the appetites of Ibicella luteaand Drosophyllum. Carnivorous Plant Newsletter 28: 40–43.
Richards JH. 2001. Bladder function in Utricularia pur-purea (Lentibulariaceae): is carnivory important? AmericanJournal of Botany 88: 170–176.
Rivadavia F, Kondo K, Kato M, Hasebe M. 2003. Phylog-eny of the sundews, Drosera (Droseraceae), based on chlo-roplast rbcL and nuclear 18S ribosomal DNA sequences.American Journal of Botany 90: 123–130.
Robinson AS, Fleischmann AS, McPherson S, Volker H,Gironella EP, Pena CQ. 2009. A spectacular new speciesof Nepenthes L. (Nepenthaceae) pitcher plant from centralPalawan, Philipines. Botanical Journal of the LinneanSociety 159: 195–202.
Rowling JK. 1998. Harry Potter and the chamber of secrets.London: Bloomsbury.
Sanders D, Gianquitto T. 2009. The ‘irritable power’ ofcarnivorous plants: Mary Treat, Charles Darwin and floralmetaphors in the evolutionary narrative. Lecture at TheLinnean Society of London, Burlington House, London, 17September 2009.
Schaefer HM, Ruxton GD. 2008. Fatal attraction: carnivo-rous plants roll out the red carpet to lure insects. BiologyLetters 4: 153–155.
Simons P. 1981. How exclusive are carnivorous plants? Car-nivorous Plant Newsletter 10: 65–68, 79–80.
Sirová D, Borovec J, Šantrucková H, Šantrucek J, VrbaJ, Adamec L. 2009. Utricularia carnivory revisited: plantssupply photosynthetic carbon to traps. Journal of Experi-mental Botany doi:10.1093/jxb/erp286.
Smith J. 2003. Une fleur du mal? Swinburne’s ‘The Sundew’and Darwin’s Insectivorous Plants. Victorian Poetry 41: 131–150.
Spomer GG. 1999. Evidence of protocarnivorous capabilitiesin Geranium viscosissimum and Potentilla arguta and other
MURDEROUS PLANTS 355
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022

sticky plants. International Journal of Plant Sciences 160:98–101.
Steward P, Riddell C. 1998. Beyond the deepwoods. London:Doubleday.
Struwe L, Thiv M, Kadereit JW, Pepper ASR, Motley TJ,White PJ, Rova JHE, Potgieter K, Albert VA. 1998.Saccifolium (Saccifoliaceae), an endemic of Sierra de laNeblina on the Brazilian–Venezuelan border, is related to atemperate-alpine lineage of Gentianaceae. Harvard Papersin Botany 3: 199–214.
Takhtajan A. 1969. Flowering plants, origin and dispersal.Edinburgh: Oliver & Boyd.
Taylor P. 1989. The genus Utricularia. Kew Bulletin Addi-tional Series XIV. London: HMSO.
Takhtajan A. 1997. Diversity and classification of floweringplants. New York: Columbia University Press.
Thiv M, Struwe L, Albert VA, Kadereit JW. 1999.The phylogenetic relationships of Saccifolium bandeirae
Maguire & Pires (Gentianaceae) reconsidered. HarvardPapers in Botany 4: 519–526.
Thorne RF. 1992. An updated phylogenetic classification ofthe flowering plants. Aliso 13: 365–389.
Tolkien JRR. 1954. The fellowship of the ring. London:George Allen and Unwin.
Wallace J, McGhee R, Biology Class. 1999. Testing forcarnivory in Ibicella lutea. Carnivorous Plant Newsletter 28:49–50.
Weinbaum SG. 1935. Parasite planet. Astounding Stories,February 1935.
Willis JH. 1965. Historical notes on the W.A. pitcher plant,Cephalotus follicularis including the provenance of the typematerial. The Western Australian Naturalist 10: 1–7.
Zambelli E. 1929. Ricerche anatomofisiologiche sulla Petuniaviolacea e sulla Petunia nyctaginiflora come piante insetti-vore. Atti dell’Istituto Botanico della Università di Pavia 1:75–87.
356 M. W. CHASE ET AL.
© 2009 The Linnean Society of London, Botanical Journal of the Linnean Society, 2009, 161, 329–356
Dow
nloaded from https://academ
ic.oup.com/botlinnean/article/161/4/329/2418471 by guest on 08 July 2022





























