Embed Size (px)
Citation preview

ARTICLEdoi:10.1038/nature13116
Molecular basis of nitrate uptake by theplant nitrate transporter NRT1.1Joanne L. Parker1 & Simon Newstead1,2
The NRT1/PTR family of proton-coupled transporters are responsible for nitrogen assimilation in eukaryotes and bacteriathrough the uptake of peptides. However, in most plant species members of this family have evolved to transport nitrateas well as additional secondary metabolites and hormones. In response to falling nitrate levels, NRT1.1 is phosphorylatedon an intracellular threonine that switches the transporter from a low-affinity to high-affinity state. Here we present boththe apo and nitrate-bound crystal structures of Arabidopsis thaliana NRT1.1, which together with in vitro binding andtransport data identify a key role for His 356 in nitrate binding. Our data support a model whereby phosphorylationincreases structural flexibility and in turn the rate of transport. Comparison with peptide transporters further reveals howthe NRT1/PTR family has evolved to recognize diverse nitrogenous ligands, while maintaining elements of a conservedcoupling mechanism within this superfamily of nutrient transporters.
Nitrogen is a key element in biology, required for the synthesis of aminoand nucleic acids and is a fundamental nutrient for cellular metabolism.The PTR or peptide transporter family, also known as the POT, orproton dependent oligopeptide transporter family, has an importantrole in nitrogen assimilation in bacteria, fungi and mammals throughthe uptake of short peptides from the environment1,2. In contrast, in themajority of plant species nitrogen is largely obtained through the uptakeof nitrate (NO3
2) from the soil3. This is achieved through the actionsof specific nitrate transporters in the plasma membrane of root cells4
and is controlled through the NRT1 and NRT2 gene families thatencode for low (KM < mM) and high (KM <mM) affinity transporters,respectively5,6. Intriguingly, the NRT1 family of nitrate transportersphylogenetically belongs to the PTR family, and may have evolved froman ancestral peptide transport protein7.
In plants the NRT1/PTR family, recently renamed the NPF (NRT1/PTR family)8, has functionally diverged with individual members recog-nizing peptides, glucosinolate defence compounds, plant hormones ornitrate9–12. The NRT1/PTR family belongs to the major facilitator super-family (MFS) of secondary active transporters13,14 that use the protonelectrochemical gradient to drive substrate uptake into the cell15–17. MFStransporters operate through an alternating access mechanism, whereina centrally located binding site is re-orientated to either side of the mem-brane to uptake and release substrate18–20. Recent crystal structures ofbacterial members of the NRT1/PTR family have identified pairs of saltbridges that orchestrate the orientation of the transporter and revealedproton binding residues that are conserved across the family fromprokaryotes to eukaryotes21,22. An important question therefore is howthe binding site in this transporter family has evolved to recognize andtransport diverse molecules while retaining elements of the same coup-ling mechanism.
The Arabidopsis thaliana nitrate transporter, NRT1.1 (NPF6.3), isthe founding member7 of the NRT1/PTR family of transporters andshares significant sequence identity to mammalian and bacterial PTRpeptide transporters (Extended Data Fig. 1). Interestingly, NRT1.1 hastwo KM values for nitrate23; in conditions of high nitrate availability(.1 mM) NRT1.1 behaves as a low-affinity transporter (KM < 4 mM).However, when nitrate levels fall below 1 mM, NRT1.1 is switchedinto a high-affinity mode (KM < 40mM). This switch occurs following
phosphorylation of an intracellular threonine, Thr 101, by the kinaseCIPK2324. This regulatory mechanism allows for the rapid adaption tochanging nitrate levels before the dedicated high affinity NRT2 trans-porter family is expressed25. These observations suggest a complex inter-play between transporter activity and post-translational modificationat the molecular level that currently has no structural or biochemicalbasis for explanation.
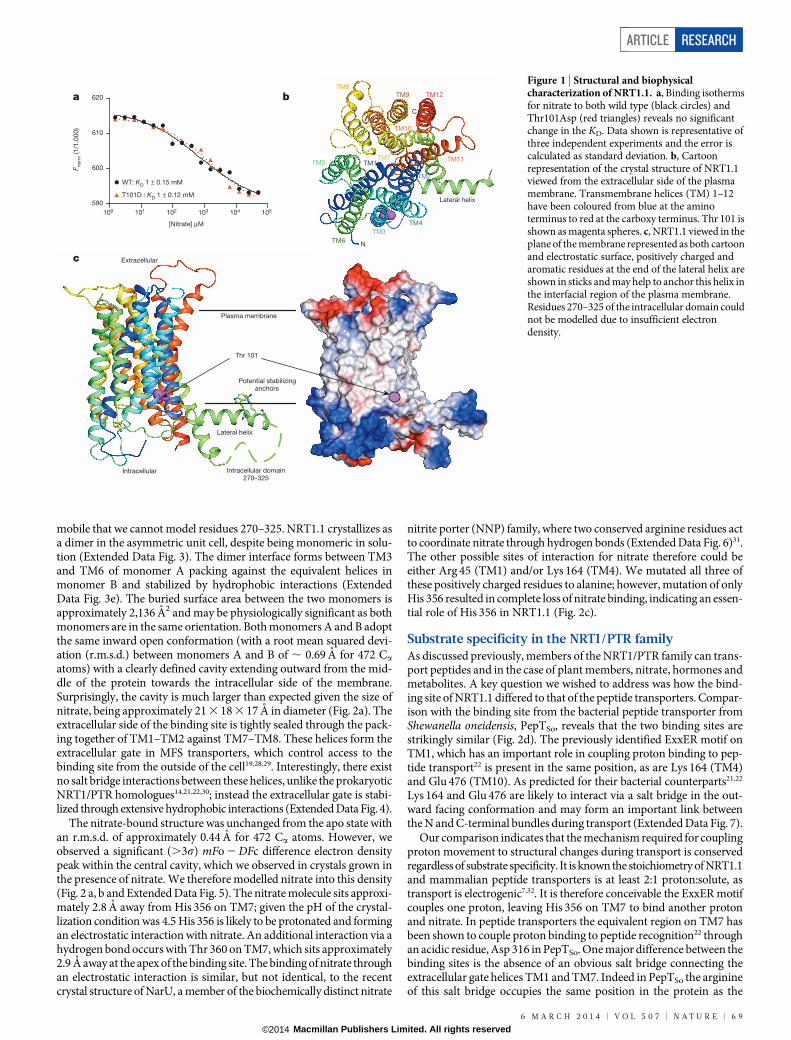
Structural basis of nitrate recognition in NRT1.1To study the binding of nitrate to NRT1.1 we developed a microscalethermophoresis assay26,27 (Extended Data Fig. 2a) and determined theKD for nitrate to be 1 6 0.15 mM (Fig. 1a and Extended Data Fig. 2b).A wide range of nitrate concentrations were tested (1mM–50 mM) andwe did not observe a high affinity state. We further tested a range ofdifferent ligands and show that NRT1.1 specifically recognizes bothnitrate and chlorate, a herbicide for which the NRT1.1 protein was ori-ginally named (chlorate resistance 1)7 but does not recognize nitrite,alanine, sulphate, phosphate or the di-peptide Ala-Ala (Extended DataFig. 2c). A key question surrounding the function of NRT1.1 is theswitch between high and low affinity states. Using the phosphomimeticvariant Thr101Asp, we investigated the effect on nitrate binding. TheKD for nitrate binding to the Thr101Asp variant was the same as forwild type (KD 5 1 6 0.12 mM) (Fig. 1a), indicating that phosphoryla-tion is unlikely to alter the nitrate-binding site directly. To follow upthis finding and identify the nitrate-binding site we crystallized NRT1.1in the presence and absence of nitrate. The crystal structures weredetermined to 3.7 A (Extended Data Table 1).
The apo structure of NRT1.1 contains 12 transmembrane (TM)-spanning alpha helices, consisting of amino-(TM1–TM6) and carboxy-terminal (TM7–TM12) bundles forming the canonical MFS fold (Fig. 1b).Separating these two bundles is a previously uncharacterized intracel-lular domain consisting of 84 amino acids. This domain is predominantlyalpha helical and extends outwards from the transporter domain at anapproximately 90u angle, which we have termed the lateral helix (Fig. 1c).At the distal end of this helix are three conserved positively chargedresidues that may help to stabilize this domain on the intracellular sideof the membrane. It is unlikely that this domain is a crystallographicartefact as it does not make any crystal contacts and is sufficiently
1Department of Biochemistry, University of Oxford, Oxford OX1 3QU, UK. 2Research Complex at Harwell, Rutherford Appleton Laboratory, Didcot OX11 0FA, UK.
6 8 | N A T U R E | V O L 5 0 7 | 6 M A R C H 2 0 1 4
Macmillan Publishers Limited. All rights reserved©2014
mobile that we cannot model residues 270–325. NRT1.1 crystallizes asa dimer in the asymmetric unit cell, despite being monomeric in solu-tion (Extended Data Fig. 3). The dimer interface forms between TM3and TM6 of monomer A packing against the equivalent helices inmonomer B and stabilized by hydrophobic interactions (ExtendedData Fig. 3e). The buried surface area between the two monomers isapproximately 2,136 A2 and may be physiologically significant as bothmonomers are in the same orientation. Both monomers A and B adoptthe same inward open conformation (with a root mean squared devi-ation (r.m.s.d.) between monomers A and B of , 0.69 A for 472 Ca
atoms) with a clearly defined cavity extending outward from the mid-dle of the protein towards the intracellular side of the membrane.Surprisingly, the cavity is much larger than expected given the size ofnitrate, being approximately 21 3 18 3 17 A in diameter (Fig. 2a). Theextracellular side of the binding site is tightly sealed through the pack-ing together of TM1–TM2 against TM7–TM8. These helices form theextracellular gate in MFS transporters, which control access to thebinding site from the outside of the cell19,28,29. Interestingly, there existno salt bridge interactions between these helices, unlike the prokaryoticNRT1/PTR homologues14,21,22,30; instead the extracellular gate is stabi-lized through extensive hydrophobic interactions (Extended Data Fig. 4).
The nitrate-bound structure was unchanged from the apo state withan r.m.s.d. of approximately 0.44 A for 472 Ca atoms. However, weobserved a significant (.3s) mFo 2 DFc difference electron densitypeak within the central cavity, which we observed in crystals grown inthe presence of nitrate. We therefore modelled nitrate into this density(Fig. 2 a, b and Extended Data Fig. 5). The nitrate molecule sits approxi-mately 2.8 A away from His 356 on TM7; given the pH of the crystal-lization condition was 4.5 His 356 is likely to be protonated and formingan electrostatic interaction with nitrate. An additional interaction via ahydrogen bond occurs with Thr 360 on TM7, which sits approximately2.9 A away at the apex of the binding site. The binding of nitrate throughan electrostatic interaction is similar, but not identical, to the recentcrystal structure of NarU, a member of the biochemically distinct nitrate
nitrite porter (NNP) family, where two conserved arginine residues actto coordinate nitrate through hydrogen bonds (Extended Data Fig. 6)31.The other possible sites of interaction for nitrate therefore could beeither Arg 45 (TM1) and/or Lys 164 (TM4). We mutated all three ofthese positively charged residues to alanine; however, mutation of onlyHis 356 resulted in complete loss of nitrate binding, indicating an essen-tial role of His 356 in NRT1.1 (Fig. 2c).
Substrate specificity in the NRT1/PTR familyAs discussed previously, members of the NRT1/PTR family can trans-port peptides and in the case of plant members, nitrate, hormones andmetabolites. A key question we wished to address was how the bind-ing site of NRT1.1 differed to that of the peptide transporters. Compar-ison with the binding site from the bacterial peptide transporter fromShewanella oneidensis, PepTSo, reveals that the two binding sites arestrikingly similar (Fig. 2d). The previously identified ExxER motif onTM1, which has an important role in coupling proton binding to pep-tide transport22 is present in the same position, as are Lys 164 (TM4)and Glu 476 (TM10). As predicted for their bacterial counterparts21,22
Lys 164 and Glu 476 are likely to interact via a salt bridge in the out-ward facing conformation and may form an important link betweenthe N and C-terminal bundles during transport (Extended Data Fig. 7).
Our comparison indicates that the mechanism required for couplingproton movement to structural changes during transport is conservedregardless of substrate specificity. It is known the stoichiometry of NRT1.1and mammalian peptide transporters is at least 2:1 proton:solute, astransport is electrogenic7,32. It is therefore conceivable the ExxER motifcouples one proton, leaving His 356 on TM7 to bind another protonand nitrate. In peptide transporters the equivalent region on TM7 hasbeen shown to couple proton binding to peptide recognition22 throughan acidic residue, Asp 316 in PepTSo. One major difference between thebinding sites is the absence of an obvious salt bridge connecting theextracellular gate helices TM1 and TM7. Indeed in PepTSo the arginineof this salt bridge occupies the same position in the protein as the
100 101 102 103 104 105
590
600
610
620
[Nitrate] μM
F no
rm (1
/1,0
00
)
WT: KD 1 ± 0.15 mM
T101D : KD 1 ± 0.12 mM
a b
TM1
TM2
TM7TM5
TM3
TM4
TM6
TM8TM9
TM10
TM11
TM12
Lateral helix
N
C
Lateral helix
Potential stabilizing anchors
Extracellular
Intracellular
Plasma membrane
Thr 101
Intracellular domain270–325
c
Figure 1 | Structural and biophysicalcharacterization of NRT1.1. a, Binding isothermsfor nitrate to both wild type (black circles) andThr101Asp (red triangles) reveals no significantchange in the KD. Data shown is representative ofthree independent experiments and the error iscalculated as standard deviation. b, Cartoonrepresentation of the crystal structure of NRT1.1viewed from the extracellular side of the plasmamembrane. Transmembrane helices (TM) 1–12have been coloured from blue at the aminoterminus to red at the carboxy terminus. Thr 101 isshown as magenta spheres. c, NRT1.1 viewed in theplane of the membrane represented as both cartoonand electrostatic surface, positively charged andaromatic residues at the end of the lateral helix areshown in sticks and may help to anchor this helix inthe interfacial region of the plasma membrane.Residues 270–325 of the intracellular domain couldnot be modelled due to insufficient electrondensity.
ARTICLE RESEARCH
6 M A R C H 2 0 1 4 | V O L 5 0 7 | N A T U R E | 6 9
Macmillan Publishers Limited. All rights reserved©2014

nitrate (Fig. 2b, d). Another significant difference is the replacement ofthe two conserved aromatic residues on TM1 (Phe 28, Tyr 29), whichare responsible for peptide specificity21 with residues containing smallerside chains helping to create space for nitrate to bind. Given that di-peptides are coordinated horizontally between the N- and C-terminalbundles22,30, it is conceivable that the increased width of the cavity inNRT1.1 can no longer coordinate the amino and carboxy termini simul-taneously. Coupled with the absence of the salt bridge and tyrosines,which help position the peptide and coordinate movement of TM1 andTM7, these alterations could explain why NRT1.1 cannot recognize ortransport peptides.
Functional role of phosphorylation at Thr 101Our structures show that Thr 101 is situated at the bottom of TM3 andpoints towards a hydrophobic pocket constructed from residues inTM2 and TM4 (Fig. 3a). Given the position of Thr 101 it is highly likelythat phosphorylation would cause localized structural distortion in thepacking of TM3 with TM1 and TM4 on the intracellular side of theprotein. Consistent with this hypothesis we discovered that the Thr101Aspvariant of NRT1.1 was significantly less stable compared to wild-typeprotein (9 uC lower melting temperature) (Fig. 3b). To investigate theeffect of the Thr101Asp substitution on nitrate uptake we reconstitutedNRT1.1 into liposomes and monitored the relative uptake of nitrateas a function of proton movement using a pH-sensitive fluorophore(Extended Data Fig. 8). As NRT1.1 is a proton-coupled nitrate trans-porter this assay provides a reliable readout for nitrate transport. Wediscovered that compared to the wild-type protein, the Thr101Aspvariant showed increased uptake of approximately 2.8-fold, whereasthe His356Ala variant showed no detectable transport, consistent withan essential role in nitrate recognition (Fig. 3c).
Given the high sequence conservation of the N-terminal bundle withinthe NRT1/PTR family (Extended Data Figs 1 and 9a), we mutated theequivalent threonine in PepTSo (Thr 87) to investigate any general effectsdisrupting the packing between these helices may have on this family.Interestingly the Thr87Asp variant also resulted in an increase in the
rate of peptide transport, whereas other variants Thr87Ala/Ser/Lys showedwild-type-like rates (Extended Data Fig. 9b–d), showing that this regionis sensitive to the addition of a negative charge. Comparable to what weobserved for NRT1.1, the Thr87Asp variant was also less stable thanwild-type protein by 4 uC (Extended Data Fig. 9e). However, a Thr87Argvariant was inactive, indicating that gross structural distortion in thisregion cannot be tolerated. This shows that disrupting the helix packing
Extracellular
Nitrate
Exit tunnel
21 Å
N-terminalbundle
C-terminalbundle
a
c
NRT1.1 PepTSo
100 101 102 103 104 105580
590
600
610
[Nitrate] μM
F no
rm (1/1
,000)
590
600
610
620
600
610
620
Lys164AlaArg45AlaHis356Ala
KD 1.2 ± 0.1 mM KD 0.9 ± 0.1 mM
b d
H356H356
Y388Y388
E476E476
K164K164
E41E41
E44E44R45R45
T48T48L49L49
NONO3–
T360T360
TM1TM1 TM7TM7
TM4TM4
TM10TM10
TM8TM8
D316D316
E419E419
K127K127
E21E21
E24E24R25R25
F28F28 Y29Y29
R32R32
F315F315H356
Y388
E476
K164
E41
E44R45
T48L49
NO3–
T360
TM1 TM7
TM4
TM10
TM8
D316
E419
K127
E21
E24R25
F28 Y29
R32
F315
F no
rm (1/1
,000)
100 101 102 103 104 105
[Nitrate] μM
100 101 102 103 104 105
[Nitrate] μM
F no
rm (1/1
,000)
Figure 2 | Crystal structure of nitrate bound NRT1.1. a, Section throughthe protein volume showing the binding site in the plane of the membrane, withthe mFo 2 DFc difference electron map contoured at 3s (green) identifying theposition of nitrate. b, Zoomed in view of the binding site. Shown in sticks are theconserved NRT1/PTR residues and His 356 that can be seen coordinatingnitrate within the binding site, supported by Thr 360. Dashed lines indicate apotential interaction network between His 356, Tyr 388 and Glu 476 in the
C-terminal bundle. c, Binding isotherms showing the effect of mutatingpositively charged residues in the binding site on the affinity of NRT1.1 fornitrate. Data shown is representative of three independent experiments and theerror is calculated as standard deviation. d, Similarity between the binding sitesof the peptide transporter PepTSo and NRT1.1. The residues important forpeptide transport are shown in sticks. The relative position of nitrate from theNRT1.1 structure is superimposed (green mesh).
TM1
TM2
TM3
TM4
TM5
TM6
Thr 101
NRT1.1
a b
c
No
prote
in
Wild
type
His35
6Ala
Thr1
01Asp
0
100
200
300
400
Rela
tive n
itra
te t
ran
sp
ort
(%
wild
typ
e)
0 20 40 60 80 100–15
–10
–5
0
Temperature (°C)
CD
mill
ideg
rees
WT: Tm 42 °C
T101D: Tm 33 °C
Hydrophobicpocket
Figure 3 | Proposed mechanism for the effect of phosphorylation at Thr 101on nitrate transport. a, Thr 101 (represented as magenta spheres) in NRT1.1 issituated in a hydrophobic cavity formed at the intracellular ends of TM2 andTM4. b, Thermal stability as assessed by the loss of alpha helical secondarystructure using circular dichroism shows a reduction in the Thr101Asp variantwhen compared to wild-type protein. c, The Thr101Asp variant of NRT1.1shows a significant increase in transport when compared to wild-type proteinin a reconstituted assay system, whereas the His356Ala mutant shows noactivity. Data represent the mean value from eight experiments and error barsare standard deviation.
RESEARCH ARTICLE
7 0 | N A T U R E | V O L 5 0 7 | 6 M A R C H 2 0 1 4
Macmillan Publishers Limited. All rights reserved©2014

between TMs 1, 2, 3 and 4 in the N-terminal bundle of the NRT1/PTRfamily can markedly affect transport rates. Similar studies have shownthat altering the structural flexibility within the MFS fold can affect therate-limiting step of transport33,34. We propose that a similar mech-anism may occur in NRT1.1 to switch the protein between a high andlow KM state. Phosphorylation causes a localized disruption of theN-terminal bundle helices that leads to an increased flexibility of theprotein. This in turn leads to an increase in the transport rate, thatwould result in the lower KM observed in the previous study24.
Model for proton-coupled nitrate transport via NRT1.1Taken together, a working model for proton-coupled nitrate transportemerges from our study (Fig. 4). The protonation of His 356 is clearly akey component of the transport mechanism. Our model predicts thatprotonation must occur before nitrate binding and may be conditionalupon the presence of nitrate to stabilize the charged state of His 356.Following closure of the extracellular gate and adoption of the occludedstate, the release of both proton and nitrate from His 356 must be coupledto the opening of the intracellular gate. A possible mechanism in achiev-ing this is that adoption of the occluded state brings His 356 into closeproximity to Glu 476, causing the disruption of the intracellular gatesalt bridge (Extended Data Fig. 7). The interaction between His 356and Glu 476 may be facilitated by Tyr 388 (TM8), which can be seenmaking a hydrogen bond to Glu 476 in the crystal structures. The bind-ing of additional protons, potentially to the ExxER motif, might alsoaffect the strength of the salt bridge between Lys 164 and Glu 476, facil-itating opening of the intracellular gate and release of bound nitrateand protons into the interior of the cell. Our data suggest that in NRT1.1the effect of phosphorylation is to cause a localized disruption in theN-terminal bundle that in turn may affect the position of the ExxERmotif with respect to Lys 164. The increase in transport rate that occursis possibly the result of altering the efficiency in the formation andbreakage of the Lys 164–Glu 476 salt bridge and release of nitrate from
the binding site. Perhaps the most surprising finding from our study,however, is that only minor changes to the previously characterizedpeptide transporter binding site are required to accommodate nitrateand that these are predominantly located in the C-terminal bundle.The ability of the PTR family to recognize multiple ligands may there-fore stem from the separation of a fundamentally conserved protoncoupling mechanism that resides largely in the N-terminal bundle andorchestrates gross conformational changes between TM4–TM5 andTM10–TM11, while leaving the C-terminal bundle to evolve to recog-nize different nitrogenous ligands.
METHODS SUMMARYNRT1.1 was cloned into a modified, cleavable green fluorescent protein–histidine(GFPHis) fusion vector and expressed in Saccharomyces cerevisiae35. The fusionprotein was solubilized in 1% n-dodecyl-b-D-maltopyranoside (DDM) and puri-fied to homogeneity. For MST binding measurements the GFPHis protein was leftattached to the C terminus. For crystallization the GFPHis was removed throughtobacco etch virus protease cleavage. Crystals were grown at pH 4.5 by the vapourdiffusion method at 4 uC. Data were collected on beamlines IO2, IO3, IO4 and IO4-1at the Diamond light source, UK. The crystals were derivatized with K2Au(CN)2
overnight and the phases calculated using single anomalous dispersion. For thenitrate co-complex, purification and crystallization was carried out in the presenceof 10 mM sodium nitrate. The final apo and nitrate-bound structures were refinedto 3.7 A resolution. MST binding studies were carried out as described in methodsfollowing the standard protocols for the instrument. Binding isotherm data werefitted to a sigmoidal dose–response equation. Nitrate uptake was assayed in pro-teoliposomes containing wild-type, His356Ala, Thr101Asp variants of NRT1.1 orno protein controls. Nitrate uptake was measured as a function of coupled protonmovement driven by an inwardly directed proton gradient through the quenchingof the fluorescent pH indicator dye pyranine.
Online Content Any additional Methods, Extended Data display items and SourceData are available in the online version of the paper; references unique to thesesections appear only in the online paper.
Received 2 October 2013; accepted 31 January 2014.
Published online 26 February 2014.
1. Steiner, H.-Y., Naider, F. & Becker, J. M. The PTR family: a new group of peptidetransporters. Mol. Microbiol. 16, 825–834 (1995).
2. Daniel, H., Spanier, B., Kottra, G. & Weitz, D. From bacteria to man: archaic proton-dependent peptide transporters at work. Physiology (Bethesda) 21, 93–102(2006).
3. Crawford, N. M. Nitrate: nutrient and signal for plant growth. Plant Cell 7, 859–868(1995).
4. Tsay, Y. F. & Hsu, P. K. The role of plasma membrane nitrogen transporters innitrogen acquisition and utilization. Plant Cell Monogr. 19, 223–236 (2011).
5. Orsel, M. et al. Characterization of a two-component high-affinity nitrate uptakesystem in arabidopsis. physiology and protein-protein interaction. Plant Physiol.142, 1304–1317 (2006).
6. Dechorgnat, J. et al. From the soil to the seeds: the long journey of nitrate in plants.J. Exp. Bot. 62, 1349–1359 (2011).
7. Tsay, Y. F., Schroeder, J. I., Feldmann, K. A. & Crawford, N. M. The herbicidesensitivity gene CHL1 of Arabidopsis encodes a nitrate-inducible nitratetransporter. Cell 72, 705–713 (1993).
8. Leran, S. et al. A unified nomenclature of NITRATE TRANSPORTER 1/PEPTIDETRANSPORTER family members in plants. Trends Plant Sci. 19, 5–9 (2013).
9. Tsay, Y.-F., Chiu, C.-C., Tsai, C.-B., Ho, C.-H. & Hsu, P.-K. Nitrate transporters andpeptide transporters. FEBS Lett. 581, 2290–2300 (2007).
10. Boursiac, Y. et al. ABA transport and transporters. Trends Plant Sci. 18, 325–333(2013).
11. Nour-Eldin, H. H., Andersen, T. G., Burow, M.& Madsen, S. R. NRT/PTR transportersare essential for translocation of glucosinolate defence compounds to seeds.Nature 488, 531–534 (2012).
12. Krouk, G. et al. Nitrate-regulated auxin transport by NRT1.1 defines a mechanismfor nutrient sensing in plants. Dev. Cell 18, 927–937 (2010).
13. Reddy, V. S., Shlykov, M. A., Castillo, R., Sun, E. I. & Saier, M. H. The major facilitatorsuperfamily (MFS) revisited. FEBS J. 279, 2022–2035 (2012).
14. Newstead, S. et al. Crystal structure of a prokaryotic homologue of the mammalianoligopeptide-protonsymporters, PepT1and PepT2. EMBOJ. 30, 417–426 (2011).
15. Fei, Y. J. et al. Expression cloning of a mammalian proton-coupled oligopeptidetransporter. Nature 368, 563–566 (1994).
16. Nakajima, H. et al. Cloning and functional expression in Escherichia coli of the geneencoding the di- and tripeptide transport protein of Lactobacillus helveticus. Appl.Environ. Microbiol. 63, 2213–2217 (1997).
17. Chiang, C.-S., Stacey, G. & Tsay, Y.-F. Mechanisms and functional properties of twopeptide transporters, AtPTR2 and fPTR2. J. Biol. Chem. 279, 30150–30157(2004).
P
+NONO3–
+
+
NONO3– +
+
+
ExtracellularExtracellular
IntracellularIntracellular
a b
cd
POPO3–
E476E476K164K164
H356H356
N C
e
1 1
114
4 4
4
7 7
77
1010 1010
1010 1010
OutwardOutwardopenopen
InwardInwardopenopen
Nitrate boundNitrate boundoccludedoccluded
Nitrate-boundNitrate-boundoutward openoutward open
T101T101
NONO3–
P
+NO3–
+
+
NO3– +
REXXE
REXXE
RE
XX
E
+
+RE
XX
E
Extracellular
Intracellular
a b
cd
PO3–
E476K164
H356
N C
e
1 1
114
4 4
4
7 7
77
10 10
10 10
Outwardopen
Inwardopen
Nitrate-boundoccluded
Nitrate-boundoutward open
T101
NO3–
Figure 4 | Alternating access model for nitrate-proton symport. a, In theoutward facing state the extracellular gate, constructed from TM1–TM2 (darkblue) and TM7–TM8 (yellow) is open and the intracellular gate, constructedfrom TM4–TM5 (cyan) packing against TM10–TM11 (orange) is closed andstabilized by a salt bridge between Lys 164 (TM4) and Glu 476 (TM10),similarly to the bacterial homologues. b, Following protonation of His 356,nitrate is able to bind triggering closure of the extracellular gate. Additional sitesof protonation exist possibly within the ExxER motif (TM1). c, Upontransition to the occluded state the intracellular gate salt bridge will break as aresult of Glu 476 moving to form a new salt bridge with His 356. d, This willresult in the release of nitrate and protons into the interior of the cell.e, Phosphorylation of Thr 101 (purple sphere) by CIPK23 results in increasedflexibility within the N-terminal bundle, increasing the overall transport rate.
ARTICLE RESEARCH
6 M A R C H 2 0 1 4 | V O L 5 0 7 | N A T U R E | 7 1
Macmillan Publishers Limited. All rights reserved©2014

18. Radestock, S. & Forrest, L. R. The alternating-access mechanism of MFStransporters arises from inverted-topology repeats. J. Mol. Biol. 407, 698–715(2011).
19. Yan, N. Structural advances for the major facilitator superfamily (MFS)transporters. Trends Biochem. Sci. 38, 151–159 (2013).
20. Madej, M. G. & Kaback, H. R. Evolutionary mix-and-match with MFS transporters.Proc. Natl Acad. Sci. USA 110, E4831–E4838 (2013).
21. Solcan, N. et al. Alternating access mechanism in the POT family of oligopeptidetransporters. EMBO J. 31, 3411–3421 (2012).
22. Doki, S. et al. Structural basis for dynamic mechanism of proton-coupled symportby the peptide transporter POT. Proc. Natl Acad. Sci. USA 110, 11343–11348(2013).
23. Liu, K. H., Huang, C. Y. & Tsay, Y. F. CHL1 is a dual-affinity nitrate transporter ofArabidopsis involved in multiple phases of nitrate uptake. Plant Cell 11, 865–874(1999).
24. Liu, K.-H. & Tsay, Y.-F. Switching between the two action modes of the dual-affinitynitrate transporter CHL1 by phosphorylation. EMBO J. 22, 1005–1013 (2003).
25. Wang, Y.-Y., Hsu, P.-K. & Tsay, Y.-F. Uptake, allocation and signaling of nitrate.Trends Plant Sci. 17, 458–467 (2012).
26. Baaske, P., Wienken, C., Willemsen, M. J. & Braun, D. Protein-binding assays inbiological liquids using microscale thermophoresis. J. Biomol. Tech. 22, S55(2011).
27. Wienken, C. J., Baaske, P., Rothbauer, U. & Braun, D. Protein-binding assays inbiological liquids using microscale thermophoresis. Nature Commun. 1, 100(2010).
28. Sun, L. et al. Crystal structure of a bacterial homologue of glucose transportersGLUT1–4. Nature 490, 361–366 (2012).
29. Dang, S. et al. Structure of a fucose transporter in an outward-open conformation.Nature 467, 734–738 (2010).
30. Guettou,F.et al.Structural insights intosubstrate recognition inproton-dependentoligopeptide transporters. EMBO Rep. 14, 804–810 (2013).
31. Yan, H. et al. Structure and mechanism of a nitrate transporter. Cell Rep. 3,716–723 (2013).
32. Chen, X.-Z., Zhu, T., Smith, D. E. & Hediger, M. A. Stoichiometry and kinetics of thehigh-affinity H1-coupled peptide transporter PepT2. J. Biol. Chem. 274,2773–2779 (1999).
33. Chandrasekaran, A., Ojeda, A. M., Kolmakova, N. G. & Parsons, S. M. Mutational andbioinformatics analysis of proline- and glycine-rich motifs in vesicularacetylcholine transporter. J. Neurochem. 98, 1551–1559 (2006).
34. Ugolev, Y., Segal, T., Yaffe, D., Gros, Y. & Schuldiner, S. Identification ofconformationally sensitive residues essential for inhibition of vesicularmonoamine transport by the noncompetitive inhibitor-tetrabenazine. J. Biol.Chem. 288, 32160–32171 (2013).
35. Newstead, S. et al. High-throughput fluorescent-based optimization of eukaryoticmembrane protein overexpression and purification in Saccharomyces cerevisiae.Proc. Natl Acad. Sci. USA 104, 13936–13941 (2007).
Acknowledgements We thank R. Flaig for help with additional access to beamline IO4and we thank Diamond Light Source for access to beam lines I02, I03, I04 and I04-1(MX7345) that contributed to the results presented here. This research was fundedthrough the Medical Research Council (MRC) Career Development Award grantG0900399 and Royal Society grants (RG110211 and IE111401) to S.N.
Author Contributions J.L.P. and S.N. designed, performed and analysed allexperiments and wrote the manuscript.
Author Information The atomic coordinates and structure factors of the apo andnitrate bound NRT1.1 are deposited in the Protein Data Bank with accession codes4cl4 and 4cl5, respectively. Reprints and permissions information is available atwww.nature.com/reprints. The authors declare no competing financial interests.Readers are welcome to comment on the online version of the paper.Correspondence and requests for materials should be addressed to J.L.P.([email protected]) or S.N. ([email protected]).
RESEARCH ARTICLE
7 2 | N A T U R E | V O L 5 0 7 | 6 M A R C H 2 0 1 4
Macmillan Publishers Limited. All rights reserved©2014

METHODSCloning, expression and purification of NRT1.1. The gene encoding NRT1.1was amplified from an Arabidopsis image clone and cloned into a modified formof a C-terminal GFPHis fusion yeast expression vector35. The yeast vector had anadditional selection marker the LEU2 gene under the control of a truncatedpromoter to increase copy number within Saccharomyces cerevisiae36. Wild-typeand mutant NRT1.1 were expressed by growing overnight in media with 2% glucosebut lacking leucine, this culture was diluted tenfold in media with 2% lactate butlacking leucine. After 8–10 h expression was induced thorough the addition of 2%galactose from a 25% stock. The yeast was collected after 20 h and membranes wereprepared. NRT1.1 and mutants were purified to homogeneity using standard IMACprotocols in n-dodecyl-b-D-maltopyranoside (DDM) detergent as described prev-iously37. For MST binding experiments the C-terminal GFPHis was not removed.The protein was concentrated to 10 mg ml21 for crystallization or 0.5 mg ml21 forbinding assays and stored at 280 uC.Crystallization and structure determination. Crystals of NRT1.1 were initiallyobtained in the MemGold crystallization screen38 and optimized to 22% PEG 400,0.05 M sodium citrate pH 4.5, 0.07 M sodium chloride and 1.5% PEG 600 usingthe hanging drop vapour diffusion technique at 4 uC. For cryoprotection the crys-tals were transferred to a solution containing 33% PEG 400, before being flash-cooled in liquid nitrogen. The crystals always showed anisotropic diffraction, withthe best crystals diffracting X-rays to Bragg spacing’s of 3.50 A in the strongestdirection. Initial phases were calculated using single wavelength anomalous dis-persion (SAD) using a single gold-cyanide-derivatized crystal. Data were collectedon beamlines I02, I03, I04 and I04-1 at the Diamond Light Source, UK. Data wereprocessed by the XIA239,40 pipeline to XDS41 and scaled using AIMLESS42. Diffractiondata were collected to 3.5 A (nitrate bound) and 3.6 A (apo) resolutions and thesedata were used for calculating maps during model building; however, the finaldeposited model was refined against the 3.7 A data shown in Extended Data Table 1,as the I/sI (measured intensity of the reflections divided by the standard deviationof the intensity for the data set) and completeness of the data dropped significantlyafter this resolution cut off. The space group was determined to be P212121 withtwo molecules in the asymmetric unit forming a head-to-head dimer (ExtendedData Fig. 3e). Two gold sites were initially located using SHELXC/D43 with theirpositions further refined and initial phases calculated using SHARP with solventflattening in SOLOMON44. The resulting experimental maps were of sufficientquality to see all 12 trans membrane helices from each of the two molecules. Furtherimprovement in map quality was obtained following cross crystal averaging inDMMulti45,46 with the native data set. The lateral helix appeared in the maps fol-lowing model building and refinement.Model building and refinement. The model was built into experimental mapscalculated from SHARP and DMMulti, using O47. The partial models were furthercycled back into SOLOMON to improve the initial solvent envelope used for thesolvent flipping procedure. The amino acid side chains were then built using anhomology model of NRT1.1 built from the template of PepTSt (4APS) and GkPOT(4IKV) using modeller v9.1248. Refinement of the model was carried out inBUSTER49 with inclusion of experimental phase information (Extended Data Fig. 5).Model validation was carried out using the Molprobity server50. Images were pre-pared using PyMol51.Microscale thermophoresis binding assay. Binding was calculated for NRT1.1 andthe control POT family transporter, PepTSo14, using microscale thermophoresis26.A range of concentrations of the required ligand (range from 0.1mM to 50 mM)was incubated with 0.8mM of purified GFP-tagged protein 5 min in assay buffer(20 mM Bis-Tris pH 6.5, 150 mM NaCl, 0.03% DDM). The sample was loaded intothe NanoTemper glass capillaries and microthermophoresis carried out using 10%LED power and 80% MST. KD were calculated using the mass action equation viathe NanoTemper software from duplicate reads of triplicate experiments. Theinstrument used was a NanoTemper monolith NT.115.PepTSo transport assays using a proton driven proteoliposome system. PepTSo
was purified as previously described14 and reconstituted by the dilution method.The efficiency of reconstitution was analysed by densitometry of a fixed amountof proteoliposomes using a serial dilution of purified protein as a standard. Forproton driven uptake assays, artificially imposed potassium ion diffusion poten-tials were generated as previously described52. Proteoliposomes were thawed andcentrifuged at 90,000g for 30 min at 4 uC and re-suspended in 20 mM potassiumphosphate, pH 6.50, 100 mM potassium acetate, 2 mM magnesium sulphate, fol-lowed by 11 cycles of extrusion through a 400-nm polycarbonate filter to obtainsmall unilamellar vesicles of relatively homogenous size53. Proteoliposomes weresubsequently diluted 1:50 (v/v) to final protein concentration of 0.25mM intoexternal buffer containing 20 mM sodium phosphate, pH 6.5, 2 mM magnesiumsulphate with 10mM valinomycin and 3H-labelled di-Ala peptide (40mM). Uptakeof 3H substrate was assayed at 25 uC. Diluting aliquots into 1.5 ml of ice-cold 0.1 Mlithium chloride stopped the reaction. Proteoliposomes were collected on 0.22-mm
nitrocellulose filters and washed under vacuum with 0.1 M lithium chloride beforescintillation counting. The 3H signal was converted to molar concentrations ofpeptide using standard curves for each substrate.Reconstitution of NRT1.1 into liposomes. NRT1.1 was reconstituted into lipo-somes using the biobead method. Polar soy lipids (Avanti polar lipids) were washedtwice in pentane and resuspended at 5 mg ml21 in lipid buffer (50 mM potassiumphosphate pH 7.5). The lipids were extruded through a 0.4-mm filter and titratedwith Triton X-100 until the absorbance at 540 nm was approximately 90% ofmaximal. Purified NRT1.1 (0.2 mg) was added to the lipids at a final lipid:proteinratio of 60:1 and incubated for 1 h on ice or for the no protein liposome control thesame volume of gel filtration buffer used to purify NRT1.1 was added. After thistime the required amount of biobeads (,0.1 mg) were added and incubated at 4 uCwith rotary mixing for 3 h. The biobead addition was repeated a further three timeswith the final incubation being overnight. The biobeads were removed and pro-teoliposomes were collected by centrifugation at 120,000g for 30 min. Proteoliposomeswere resuspended in lipid buffer at a final lipid concentration of 30mgml21 anddialysed overnight in lipid buffer to remove any traces of residual detergent. Afterdialysis the proteoliposomes were collected and resuspended as before and sub-jected to three rounds of freeze-thawing in liquid nitrogen before storage at280 uC. Protein amount was quantified by SDS–PAGE and densitometry.Nitrate transport assays using pyranine dye. NRT1.1 proteoliposomes werethawed and collected at by centrifugation at 120,000g for 25 min at 4 uC . The lipo-somes were resuspended in inside buffer (5 mM HEPES pH 6.8, 120 mM KCl,2 mM MgSO4) and 1 mM pyranine and loaded by two rounds of freeze-thawingin liquid nitrogen followed by extrusion through first a 0.4-mm filter then a 0.2-mmfilter. Excess pyranine was removed by washing once in inside buffer and then byapplication to a G25 gel filtration (GE Healthcare) column equilibrated in insidebuffer. After this step the liposomes were split into 8mg aliquots (enough for oneassay per tube) and collected by centrifugation at 120,000g for 25 min at 15 uC.
The prepared liposomes were resuspended in 99 ml external buffer (5 mM MESpH 6.0, 1 mM KCl, 2 mM MgSO4, 109 mM NaCl) and the fluorescence was readimmediately at 460/510 nm and 415/510 nm in a SpectraMax M3 plate readerusing 96-well black clear-bottom plates (Molecular Devices). Each assay wasmeasured individually. Either 10 mM sodium nitrate or 10 mM sodium chloride(no nitrate control) from 1 M stocks were added and again fluorescence readimmediately after the addition and mixing. Data were collected for 30 s in 10-sincrements. The difference between nitrate and no nitrate after 20 s was calculated(averaged from 8 experiments) and used to compare the variants of NRT1.1 (datashown in Fig. 3b).
36. Kazemi Seresht, A., Nørgaard, P., Palmqvist, E. A., Andersen, A. S. & Olsson, L.Modulating heterologous protein production in yeast: the applicability oftruncated auxotrophic markers. Appl. Microbiol. Biotechnol. 97, 3939–3948(2013).
37. Drew, D. et al. GFP-based optimization scheme for the overexpression andpurification of eukaryotic membrane proteins in Saccharomyces cerevisiae. NatureProtocols 3, 784–798 (2008).
38. Newstead, S., Ferrandon, S. & Iwata, S. Rationalizing a-helical membrane proteincrystallization. Protein Sci. 17, 466–472 (2008).
39. Winter,G., Lobley,C.M.C.&Prince,S.M.Decisionmaking inxia2. ActaCrystallogr. D69, 1260–1273 (2013).
40. Winter, G. xia2: an expert system for macromolecular crystallography datareduction. J. Appl. Cryst. 43, 186–190 (2009).
41. Kabsch, W. XDS. Acta Crystallogr. D 66, 125–132 (2010).42. Evans, P. R. An introduction to data reduction: space-group determination, scaling
and intensity statistics. Acta Crystallogr. D 67, 282–292 (2011).43. Sheldrick, G. M. Experimental phasing with SHELXC/D/E: combining
chain tracing with density modification. Acta Crystallogr. D 66, 479–485(2010).
44. Abrahams, J. P. & Leslie, A. G. W. Methods used in the structuredetermination of bovine mitochondrial F1 ATPase. Acta Crystallogr. D52, 30–42 (1996).
45. Cowtan, K. DM: an automated procedure for phase improvement by densitymodification. Joint CCP4 and ESF-EACBM Newsletter on Protein Crystallography 31,34–38 (1994).
46. Pedersen, B. P., Morth, J. P. & Nissen, P. Structure determination using poorlydiffracting membrane-protein crystals: the H1-ATPase and Na1,K1-ATPase casehistory. Acta Crystallogr. D 66, 309–313 (2010).
47. Jones, T. A., Zou, J. Y., Cowan, S. W.& Kjeldgaard,M. Improvedmethods for buildingproteinmodels inelectron density mapsand the location oferrors in thesemodels.Acta Crystallogr. A 47, 110–119 (1991).
48. Yang, Z. et al. UCSF Chimera, MODELLER, and IMP: An integrated modelingsystem. J. Struct. Biol. 179, 269–278 (2012).
49. Blanc, E. et al. Refinement of severely incomplete structures withmaximum likelihood in BUSTER-TNT. Acta Crystallogr. D 60, 2210–2221(2004).
50. Davis, I. W., Murray, L. W., Richardson, J. S. & Richardson, D. C. MOLPROBITY:structure validation and all-atom contact analysis for nucleic acids and theircomplexes. Nucleic Acids Res. 32, W615–W619 (2004).
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2014

51. Schrodinger, LLC. The PyMOL Molecular Graphics System, Version 1.3r1.52. Foucaud, C. & Poolman, B. Lactose transport system of Streptococcus
thermophilus. Functional reconstitution of the protein and characterizationof the kinetic mechanism of transport. J. Biol. Chem. 267, 22087–22094(1992).
53. Knol, J.et al. Unidirectional reconstitution intodetergent-destabilized liposomes ofthe purified lactose transport system of Streptococcus thermophilus. J. Biol. Chem.271, 15358–15366 (1996).
54. Smirnova, I. et al. Sugar binding induces an outward facing conformation of LacY.Proc. Natl Acad. Sci. USA 104, 16504–16509 (2007).
55. Zheng, H., Wisedchaisri, G. & Gonen, T. Crystal structure of a nitrate/nitriteexchanger. Nature 497, 647–651 (2013).
56. Katoh, K. & Toh, H. Recent developments in the MAFFT multiple sequencealignment program. Brief. Bioinform. 9, 286–298 (2008).
57. Clamp, M., Cuff, J., Searle, S. M. & Barton, G. J. The Jalview Java alignment editor.Bioinformatics 20, 426–427 (2004).
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Figure 1 | Alignment of NRT1.1 with PTR family members.Amino acid alignment of A. thaliana NRT1.1 (Uniprot: Q05085) withPepTSo (Q8EKT7), PepTSt (Q5M4H8), human PepT1 (B2CQT6) and PepTGk
(Q5KYD1) using MAFFT56 with manual adjustment in JalView57. AtNRT1.1shares 23% identity to human PepT1 and 22% identity to PepTSo at the primarystructure level. Identical residues shared across the NRT1/PTR family arehighlighted in red with key functionally conserved residues highlighted by bluetriangles. Nitrate binding site residues His 356, Tyr 388 are highlighted asmagenta stars. Thr 101 is conserved in all mammalian PTR family membersand a small subset of bacterial homologues including PepTSo. NRT1.1 topology:the central cavity is shown as an open triangle, representing the inwardopen state observed in the crystal structure. The 12 TM helices identifiedfrom the crystal structures are coloured from blue to red and the observedintracellular domain between TM6–TM7 is indicated in grey.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Figure 2 | Microscale thermophoresis binding assay as atechnique to calculate KD for ligand binding to NRT1/PTR transporters.a, The accuracy of the microscale thermophoresis binding assay was examinedthrough comparison with data obtained from transport assays. PepTSo wasfunctionally reconstituted into liposomes as described and an IC50 valuecalculated for Ala-Ala was determined and found to be approximately 50mM(a, left). Using the NanoTemper monolith NT.115 instrument the KD forAla-Ala was calculated to be approximately 35mM (a, right) similar to the IC50
calculated from proton-driven transport assays. In this assay the orientation ofthe PepTSo molecules are highly likely to adopt multiple conformations, asobserved for other members of the MFS54, and so the KD we have determined is
a global parameter for this protein and should be the same for eitherconformation of the transporter. b, The binding of nitrate to NRT1.1 wasdetermined at pH 7.0, 6.5 and 6.0; the KD calculated at each of these pH valuesdid not markedly differ from one another. However, as the pH was reducedthe fluorescent signal produced by GFP was partially quenched, therefore allsubsequent analysis was performed at a pH of 6.5. Nitrite, alanine, di-alanine,phosphate and sulphate show no observable interaction with AtNRT1.1 usingthe MST binding assay under the same conditions as for nitrate. c, Chlorate wasshown to bind NRT1.1 with a KD of approximately 5 mM. Data shown arerepresentative of three independent experiments and mean and standarddeviations are shown.
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Figure 3 | Purification, secondary structure analysis andcrystallization of NRT1.1. a, Gel filtration trace of the purified NRT1.1 aftertobacco etch virus cleavage of the GFPHis. The protein elutes as a single peakcorresponding to a size of approximately 65 kDa following subtraction of theDDM micelle (,50 kDa), which would correspond to a monomer in solution.b, The protein was purified to homogeneity as judged by Coomassie-stainedSDS–PAGE. c, Analysis of secondary structure content of NRT1.1 usingcircular dichroism spectroscopy shows the protein is predominately alphahelical with 22% disordered regions and no significant beta strands. d, Typicalcrystal obtained with scale bar at 100mm. e, AtNRT1.1 crystallizes as a dimer inthe asymmetric unit. The buried surface area between the two monomers is
2,135 A2 as calculated using PDBe PISA (http://www.ebi.ac.uk/msd-srv/prot_int/) and may be physiologically significant as both monomers are in thesame orientation. Structural comparisons between bacterial members of thePTR family in different conformations of the transport cycle suggest theN-terminal bundle is less flexible compared to the C-terminal helices21, whichmay explain why we observe NRT1.1 packing through these helices in thecrystal. The oligomeric state of the PTR family in the membrane is currentlyunclear, with a recent bacterial member conjectured to form a tetramer30.In this case however, the dimer interface was found to be via the C- rather thanthe N-terminal bundle.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Figure 4 | Extracellular gate of NRT1.1 is stabilized throughhydrophobic interactions as opposed to the salt bridges observed in thebacterial and mammalian members of this family. NRT1.1 is shown in theplane of the membrane with a slice through the protein volume highlighting the
central nitrate binding site and the exit tunnel. The extracellular gate formedfrom TM1 and TM2 packing against TM7 and TM8 is highlighted and theinterface between these helices is illustrated using electrostatic surfacerepresentations.
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Figure 5 | Representative electron density maps used duringmodel building and refinement. Stereo view of final 2mFo 2 DFc electrondensity from refinement, contoured at 1s around chain A (PDB: 4CL5). Viewshown is equivalent to Fig. 2b in main text (PDB: 4CL4). Nitrate binding site:the position of modelled nitrate was resolved much better in the electrondensity maps in chain A compared to chain B, possibly due to higher occupancyin this molecule and the analysis in the paper was therefore conducted onchain A. In chain B the nitrate was seen to occupy a slightly different position
within the binding site, displaced down approximately 1.7 A compared to theposition in molecule A but still sitting close to and interacting with His 356.Given the resolution is 3.7 A; it is difficult to ascertain the significance of thisobservation with respect to the coordination of nitrate. The low KD valuedetermined from the binding experiments could be consistent with nitratebeing able to adopt slightly different positions within the binding site withrespect to His 356.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Figure 6 | Structural comparison between nitrate bindingsites in NRT1.1 and the bacterial NNP family members NarK and NarU.Although NRT1.1 shares no sequence identity to the nitrate-nitrite porter(NNP) family, they do both belong to the major facilitator superfamily, andshare the same canonical architecture of 12 transmembrane helices4.a, Superposition of NarU and NarK (both from E. coli) with NRT1.1; NarU andNarK crystal structures were recently published in complex with nitrate andnitrite, respectively31,55. Consistent with previously determined crystalstructures for ligand-bound MFS family members the ligand-binding site islocated in the centre of the molecule, constructed from side chains contributedby the N- and C-terminal bundles. In NRT1.1 the nitrate in chain A is locatedmuch further into the cavity, approximately 9 A, than observed in either NarUor NarK and makes far fewer interactions with side chain functional groups.However, as discussed in the main text the binding sites do exhibit similarphysiochemical features. b, c, As illustrated in the close-up of the binding
sites (b) and depicted in the cartoon schematic below (c), the nitrate in NarU isheld tightly by two electrostatic interactions to Arg 87 and Arg 303 and bytwo further hydrogen bonds to Asn 173 and Tyr 261 (yellow sticks). Theseinteractions are further sandwiched between two layers of hydrophobicresidues, which will probably increase the relative strength of the electrostaticinteractions by lowering the dielectric constant in the immediate vicinity of thenitrate. By comparison and as discussed in the main text, in NRT1.1 the nitrateis also held by an electrostatic interaction with a protonated His 356 withcontributions in chain A from Thr 360 (purple sticks). The immediate vicinityof the nitrate in the present conformation of NRT1.1 is also hydrophobic andhighlighted (purple sticks and black oval). However, as shown in the cartoonschematic, the number and strength of these interactions are substantiallylower compared to the NNP family of transporters, potentially explaining themuch lower KD we observe for NRT1.1 (1 mM) compared to NarU (33mM).
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Figure 7 | Superposition of NRT1.1 onto the outward facingcrystal structure of the fucose permease, FucP. The coordinates for theNRT1.1 structure was divided into the N-and C-terminal six-helix bundles,coloured blue and orange, respectively, before being superimposed onto thestructure of FucP (PDB: 3O7Q) using the CCP4 Superpose program. Ther.m.s.d. values are indicated. Helices 2, 7 and 11 were removed for clarity.Superposition of both N and C bundles of NRT1.1 onto the proton-coupledfucose transporter FucP places Lys 164 in the same position as a conservedGlu 135 in FucP, which upon protonation is predicted to trigger the switch tothe inward-facing state29. Similarly, in superimposing the respective C-terminal
domains, Glu 476 of NRT1.1, which this study suggests is involved in structuralaspects of nitrate binding through an electrostatic interaction with His 356 (seeFig. 2b and Extended Data Fig. 5), is close to Tyr 365 in FucP, which is theproposed interaction partner of Glu 135. A speculative function of Lys 164 maytherefore be to form a salt bridge with Glu 476 in the outward openconformation (Fig. 4a), facilitating the closer packing of helices H4 and H10and sealing the binding site to the interior of the cell in the outward-facing state.Supporting this interaction in the POT family are mutational data from bothPepTSt and GkPOT, which show these residues are important for proton-driven peptide transport in the POT family21,22.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2014

RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Figure 8 | Proton transport is increased in the Thr101Aspvariant of NRT1.1 when compared to wild-type protein. A reconstitutedproteoliposome system was used to study the effect of the Thr101Asp varianton nitrate uptake. a, In this assay the membrane impermeant pH indicator,pyranine (trisodium 8-hydroxypyrene-1,3,6-trisulfonate) was trapped withinthe NRT1.1 reconstituted liposomes. In the presence of a pH gradient, NRT1.1couples the import of nitrate with protons into the lumen of the liposome.b, The trapped pyranine becomes protonated, whereupon the excitationmaximum shifts from 460 to 415 nm, the emission maximum remainingconstant at 510 nm. Therefore proton-coupled nitrate transport can bemeasured as a function of the proton-dependent quenching of pyranine.c, Protein was reconstituted into liposomes as detailed in the Methods andassays were performed on the same amount of protein for the wild-type,Thr101Asp and His356Ala variants. d, Representative data obtained from thenitrate transport assays showing three independent experiments for both
protein free liposomes and those containing the variants were studied. Nitrateuptake was driven using a pH gradient (pH 6.8 inside, 6.0 outside) as theaddition of valinomycin caused an anomalous proton leak in NRT1.1-containing liposomes. The difference between the addition of nitrate (blue)verses a no nitrate control (black) is shown, data were normalized to the valuesobtained before the addition of nitrate. Due to the use of a plate reader tomeasure the experiments, after the addition of nitrate there is a slight delay(,10 s) before the first measurement is taken (time zero) which is why at timezero a drop in fluorescence is observed. The presence of wild-type NRT1.1caused a larger change in fluorescence (indicative of acidification) in thepresence of nitrate, which was not seen with the inactive His356Ala variant. TheThr101Asp variant, however, caused a substantial increase in acidificationindicative of more protons and therefore nitrate being transported by thisvariant.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2014

RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Figure 9 | Effects of mutating Thr 87 in PepTSo on proteinstability, peptide binding and proton-coupled peptide uptake in areconstituted proteoliposome system. a, The N-terminal domains of NRT1.1(also shown in Fig. 3a) and the bacterial peptide transporters are very similar.The location of Thr 101 and the equivalent threonine in PepTSo, Thr 87, arehighlighted as magenta spheres, both threonines are located in a similarhydrophobic cavity formed at the intracellular ends of TM2 and TM4.b, Thr87Asp variant of PepTSo showed a fourfold increase transport rate,whereas the Thr87Arg variant was inactive. c, Mutation to alanine, serine orlysine had no significant impact on the rate of transport. Error bars represents.d. from three independent experiments. d, Comparison of substrate specificity
between wild type and The87Asp variant showed no significant change,indicating that the binding site has not been adversely affected. Error barsrepresent s.d. from three independent experiments. e, The stability as assessedusing a high-performance circular dichroism spectrometer (Jasco J-1500instrument) of the Thr87Asp (Tm 5 52 uC) and Arg (Tm 5 46 uC) mutants isreduced when compared to wild-type protein (56 uC). Loss of alpha helicalsecondary structure was monitored continuously at 220 nm while thetemperature was increased from 20 uC to 80 uC over 1-degree increments. 10mgof protein were monitored in a 100-ml cuvette. f, Binding isotherms showingthere is no change in affinity for peptide upon mutating Thr 87 to Asp.
ARTICLE RESEARCH
Macmillan Publishers Limited. All rights reserved©2014

Extended Data Table 1 | Data collection and refinement statistics
RESEARCH ARTICLE
Macmillan Publishers Limited. All rights reserved©2014





























