Embed Size (px)
Citation preview

18 Leonardi et al.Archives of Insect Biochemistry and Physiology 39:18–35 (1998)
© 1998 Wiley-Liss, Inc.
Modification of the Nutritional Parameters and of MidgutBiochemical and Absorptive Functions Induced by the
IGR Fenoxycarb in Bombyx mori LarvaeM. Giovanna Leonardi1* Silvia Cappellozza,2 Magda de Eguileor,3 Annalisa Grimaldi,3 Luciano Cappellozza,2
Morena Casartelli,1 and Barbara Giordana1
1Department of Biology, University of Milan, Milan, Italy2Istituto Sperimentale per la Zoologia Agraria, Sezione Specializzata per la Bachicoltura, Padova, Italy
3Department of Structural and Functional Biology, University of Insubria, Varese, Italy
Fifth instar larvae of B. mori were topically or orally treatedwith increasing amounts of the Insect Growth Regulator (IGR)fenoxycarb in a single application, in order to determine itseffects on the nutritional parameters, the midgut functionalactivities and the growth of the silk glands. The IGR affectedin a dose-dependent manner the progress of the life cycle ofthe insect, causing a delay or inhibition of spinning, alterationof the feeding behaviour, decrease of the nutritional parameters,impairment of the growth of the silk glands, and an increasedmortality during larval-pupal transformation. Measurement ofleucine uptake into midgut brush border membrane vesicles andmidgut histochemistry revealed a reduced absorption of leucineby the midgut and a large alteration of a number of midgut en-zyme activities as a result of treatments with a high dose offenoxycarb (2.5 mg). Treatments with a dose of 2.5 femto g/larvacaused an increase in leucine uptake by the midgut, an in-creased weight of the cocoon shell, and a modification of somemidgut enzyme activities. The lepidopteran midgut appears tobe a larval organ that responds promptly to the exposure tofenoxycarb. The epithelial columnar cells modify their absorp-tive functions, at least with regard to amino acid uptake, aswell as their metabolic activity, with a modification of the oxi-dative status of the cells that is detectable with a single doseof the chemical as low as few fg/larva. Arch. Insect Biochem.Physiol. 39:18–35, 1998. © 1998 Wiley-Liss, Inc.
*Abbreviations used: AcP = acid phosphatase; AD = approxi-mate digestibility; AlP = alkaline phosphatase; AM = ante-rior-middle; BBMV = brush border membrane vesicles; BrdU= 5-Bromodeoxyuridine; CI = Consumption index; CW =weight of the diet consumed; ECD = efficiency of conversionof digested food to body substance; ECI = efficiency of con-version of ingested food to body substance; FCCP = carbo-nyl cyanide p-trifluoromethoxyphenyl-hydrazone; Feno-xycarb = Ethyl[2-(p-phenoxy-phenoxy) ethyl] carbamate; GR= growth rate; IGR = Insect Growth Regulator; JH = juve-nile hormone; LD50 = Dose wich is lethal to 50% of the or-ganisms tested; Methoprene = 11-Methoxy-3,7,11-trimethyl-
2,4-dodecadienoic acid 1-methylethyl ester; NADH-TR =NADH tetrazolium reductase; O.R.O = oil red O; P = poste-rior; PAS = periodic acid Schiff stain; ppm = part per mil-lion; SDH = succinic dehydrogenase.
Contract grant sponsor: Ministero dell’Università e dellaRicerca Scientifica e Tecnologica, Italy (MURST ex 60%).
*Correspondence to: Dr. M. Giovanna Leonardi, Departmentof Biology, University of Milan, via Celoria 26, 20133 Milan,Italy. E-mail: [email protected]
Received 14 April 1998; accepted 22 July 1998

Fenoxycarb and Midgut Functions 19
Key words: fenoxycarb; Bombyx mori; nutritional parameters; larval midgut;histochemistry; absorption
INTRODUCTION
Fenoxycarb* is an insect growth regulator(IGR) with a potent toxicity towards target pests.It is a large spectrum insecticide (Grenier andGrenier, 1993), able to produce toxic effects to dif-ferent classes of invertebrates (Gordon et al.,1996; Stoka, 1996).
Unfortunately, the IGR has proved harmfulto a number of beneficial insects (Grenier andGrenier, 1993). For instance, it produces a highmortality in the parasitoid, Phanerotoma ocularis,used for pest control when applied to the hostlarvae Ephestia kuehniella (Moreno et al., 1993).In the silkworm Bombyx mori, fenoxycarb inhib-its spinning (Cappellozza et al.,1990; Dedos andFugo, 1992) and induces the formation of dauerlarvae (Dedos et al., 1993; Monconduit andMauchamp, 1997) by altering the complex hor-monal interations (Plantevin et al., 1991; Cap-pellozza et al., 1995; Dedos and Fugo, 1996;Monconduit and Mauchamp, 1997). Silkwormsare sensitive to extremely low doses of the IGR(Cappellozza et al., 1990), consequently spinninginhibition was first observed in the sericulture farmseven in the absence of a contamination of the mul-berry leaves detectable by standard analytical meth-ods. Only careful laboratory trials (Cappellozza etal., 1995; Kamimura, 1995) could demonstrate thatfenoxycarb was indeed responsible for the spinninginhibition observed in the field.
The IGR enters the silkworm essentially byingestion so that the midgut epithelium is the lar-val tissue that is first exposed to the chemical.We have observed that its presence in the larvalhemolymph, after a topical application of con-trolled amounts of the IGR to larvae (Insegar,commercial formulation), affected the ability ofthe midgut to translocate leucine via the K+/aminoacid symporters located on the brush border mem-brane of midgut columnar cells. Fenoxycarbstimulated leucine transport at a low dose (fg/larva) and inhibited it at a high dose (µg/larva)(Leonardi et al., 1996).
We have now investigated how the oral or topi-cal administration of fenoxycarb to B. mori larvaeaffected midgut functional properties in vivo by de-termining the nutritional parameters, and in vitroby measuring leucine uptake into brush bordermembrane vesicles. We have also investigated thepossible metabolic alterations induced by the IGR
in the midgut cells, by histochemical determinationof a number of enzyme activities and substrates,as part of a screening for precocious indicators ofexposure to the insecticide.
MATERIALS AND METHODSExperimental Animals
Fifth instar larvae of a tetramoulter polyhy-brid (BG20 × BG 8)×(BC21 × BC24) B. mori wereused. The larvae were kept in laboratory condi-tions, under a 16L:8D photoperiod, 24 ± 1°C and6570% RH. From the beginning of the fifth in-star, the larvae were kept in transparent plasticcups covered with muslin cloth and were rearedindividually on artificial diet (Yakult Co., Japan),unless otherwise indicated.
Treatments With Fenoxycarb
Fenoxycarb (95% pure) was a gift from Prof.M. Dolci, Dipartimento di Valorizzazione e Prote-zione delle Risorse Agroforestali, University ofTorino, Italy. For topical treatments the chosenamount of IGR dissolved in 1 µl ethanol wasplaced on the first thoracic tergite. For oral treat-ments, the IGR dissolved in 1 µl of water wasdelivered in the oral cavity with a 10 µl Hamiltonsyringe equipped with a blunt needle. The treat-ment was carried out once, at the beginning ofthe last instar just after the fourth ecdysis un-less otherwise specified.
Determination of the Nutritional Parameters
Larvae were divided into four groups, 40 foreach group, and each insect in each of the groupswas topically treated with: (1) 1 µl ethanol; (2)2.5 fg of fenoxycarb; (3) 2.5 pg of fenoxycarb; or(4) 2.5 µg of fenoxycarb. The different groups werereared in separate rooms.
Following the same scheme, four groups wereorally treated with: (1) 1 µl of water; (2) 2.5 fg offenoxycarb; (3) 2.5 pg of fenoxycarb; or (4) 2.5 µg offenoxycarb.
To determine the effect of the treatments per-formed on the nutritional parameters, the larvaewere provided a known weight of artificial dietthat was weighed daily. To evaluate water loss,weighed samples of diet were kept in similar cups.The faeces and uneaten diet were removed andweighed daily until the end of the larval instar.

20 Leonardi et al.
The corrected weight of the diet consumed(CW), the efficiency of conversion of ingested foodto body substance (ECI), and the efficiency of con-version of digested food to body substance (ECD)were calculated according to Waldbauer (1968).Consumption index (CI), growth rate (GR), andapproximate digestibility (AD) were defined ac-cording to the equations given by Soo Hoo andFraenkel (1966). These parameters were calcu-lated both daily and for the entire period of thelast larval instar. To make the measurements uni-form both for untreated and treated animals, thedata were recorded from the beginning of the lastinstar (corresponding to the first feeding afterecdysis) until the insects started to show a de-crease in their body weight. The duration of thefifth instar, the percentage of larval mortality, theweight of cocoon (cocoon shell + pupa), of cocoonshell and of pupa were also recorded.
ANOVA analysis and Duncan’s test were car-ried out on the data reported in Tables 1, 2, and3. Corrected mortality was calculated accordingto the method of Abbott modified by Schneider-Orelli, as reported by Tremblay (1982):
Dose mortality -Control mortality
Corrected mortality = × 100100 – Control mortality
Continuous Application of fg-Dosesof Fenoxycarb
Fifth instar larvae, reared on mulberryleaves, were treated with consecutive topical ororal applications of 2.5 fg of fenoxycarb. Larvaewere divided into three groups of 50 animals eachand they were reared in cups containing five ani-mals each. The first group was of untreated lar-vae (controls), the second group was force-fed with2.5 fg of fenoxycarb for 4 consecutive days (im-mediately prior to the first feeding after ecdysisand then after 24, 48, and 72 h), and the thirdgroup was topically treated with the same amountof the IGR and the same treatment protocol. Eachtreated larva, therefore, received a total dose of10 fg of fenoxycarb. Previous experiments hadshown that sham-injected or sham force-fed con-
trols did not differ from untreated larvae withregard to the parameters under examination (un-published results).
The following parameters were recorded: theduration of the fifth instar, the percentage of lar-val mortality, and the weight of cocoon, cocoonshell, and pupa.
Analysis of Silk Gland Growth
Five male and five female 2.5 pg-treated lar-vae (one-dose at the beginning of the fifth instar)were weighed and sacrificed on each day of the lastinstar to assess the silk gland growth in compari-son to the control. The larvae were anaesthetisedwith chloroform and dissected to remove the intactsilkglands. These were immediately rinsed in physi-ological solution, blotted on blotting-paper and thenweighed. The experiment was stopped on the dayof spinning of the control larvae
Isolation of Midgut Tissue, Preparation ofBrush Border Membrane Vesicles andTransport Experiments
Controls and silkworms topically or orallytreated with a single application of 2.5 fg/larvaor 2.5 µg/larva, corresponding to the lowest andthe maximal dose, were sacrificed on the sixthday after treatment. Larvae were kept on crushedice for 15–20 min and then cut immediately be-hind the last pair of thoracic legs and the fourthpair of abdominal appendages, to exclude the fo-regut and the hindgut. The integument was cutaway and the exposed midgut, deprived of theperitrophic membrane and the Malpighian tu-bules, was dissected longitudinally and rinsedthoroughly with ice-cold 210 mM sucrose, 45 mMKCl, 10 mM Tris-HCl at pH 7.2. The anterior-middle (AM) and posterior (P) regions (Giordanaet al., 1994) were separated, slightly blotted,weighed, placed in cryotubes, and frozen by im-mersing the vial in liquid nitrogen. The cryotubeswere then stored in liquid nitrogen. For prepara-tion of BBMV from the two midgut regions, fro-zen samples were rapidly thawed by immersingthe cryotube in a 37°C water bath. BBMV wereobtained by differential calcium precipitation asdescribed by Hanozet et al. (1980). The final
TABLE 1. Effects of the Topical Application of Fenoxycarb
IGR dose Length of fifth instar (day) Mortality (%) Cocoon weight (g ± S.D.) Shell weight (g ± S.D.)
None 8 0 1.88 ± 0.24 0.41 ± 0.072.5 pg 11 52 2.33 ± 0.33* 0.43 ± 0.032.5 µg 16 100 — —
*P< 0.01.

Fenoxycarb and Midgut Functions 21
pellet was resuspended in 90 mM Hepes, 34 mMTris at pH 7.2 by 10 passes through a 22-gaugeneedle. The protein concentration was deter-mined with the Coomassie Brilliant Blue G-250(Pierce, Rockford, IL) protein assay, using bo-vine serum albumin as standard, and adjustedto a final concentration of 5 mg/ml.
Amino acid transport experiments were per-formed in quadruplicate at 22°C by rapid filtrationof the vesicle suspension through a pre-wetted cel-lulose nitrate filter. For tritiated-leucine uptake ex-periments, 10 µl of the vesicle suspension was mixedwith 40 µl of the radiolabelled incubation medium,with the following final concentration (mM): 18Hepes, 55 Tris-OH at pH 8.8, 0.11 L-leucine, 2.22and 1.48 MBq/ml L-[4,5-3H]leucine (Radiochemi-cal Centre Amersham International, Amersham,UK) for AM-BBMV and P-BBMV, respectively, 50K2SO4, 100 µM FCCP. These experimental set-tings simulated at best the “physiological” condi-tions, i.e., a nearly neutral pH inside the vesiclesas in cytoplasm, a high alkalinity outside theBBMV as in midgut lumen, and an electrical po-tential difference of about 90 mV, interior nega-tive, across the vesicle membrane (obtained bythe proton diffusion along its electrochemical gra-dient induced by the protonophore) to mimic thehigh transapical voltage of midgut columnar cells.The incubations lasted 7 s and each measurementwas performed in quadruplicate. Uptake wasstopped by diluting the incubation mixture with100 vol of ice-cold 150 mM NaCl, 10 mM Hepes-Tris at pH 7.2. The suspension was filtered througha prewetted cellulose nitrate filter (0.45 µm poresize, Micro Filtration Systems, Dublin, CA), thencounted for radioactivity in a scintillation spectrom-eter (Tri-Carb model 300 C, Packard, Camberra, IL).
The results reported in Figure 3 represent atypical experiment. Experiments from three dif-ferent BBMV preparations gave similar results.
Histochemistry
Frozen sections of four midguts from eachgroup of controls and larvae topically treated withsingle dose of 2.5 fg/larva or 2.5 µg/larva, sacri-
ficed on the sixth day after treatment, were cutin a cryostat at –20°C, mounted on glass slides,and processed according to Dubowitz and Brooke(1973), utilizing a histoenzymatic kit (Bio-Optica)for: NADH-TR (NADH tetrazolium reductase),SDH (succinic dehydrogenase), both reflecting theutilisation of various metabolic intermediates ofthe Krebs cycle and related pathways, namelymitochondrial activity; O.R.O. (oil red O), indi-cating neutral lipids reserves; acid (AcP) and al-kaline phosphatase (AlP) activities, markers,respectively, of lysosomal activity in relation tocatabolic processes and brush border membranefunctional integrity; PAS to reveal the presenceof glycogen or other polysaccharides. For all re-actions, controls involving the omission of the sub-strate were carried out except for the PAS stain,the specificity of which was checked by using dia-stase digestion. In all cases, negative results wereobtained, thus proving the specific nature of thereactions. Cell proliferation was monitored usingDNA incorporation of the substituted nucleotide,5-BrdU. A monoclonal antibody against BrdU(Amersham, Buckinghamshire, UK) was used toreveal the incorporation. The immunocychem-ical technique was used to detect the BrdU in-corporated by S-phase cells. Four larvae wereinjected with a solution at a final concentra-tion of 0.05% and were sacrificed 6 h later.Specimens were prepared using the usual pro-cedure for electron microscopy.
Light Microscopy and TransmissionElectron Microscopy
For routine transmission electron micros-copy, four midguts from each group were cut intosmall pieces fixed in 2% glutaraldeyde in 0.1 Mcacodylate buffer at pH 7.2 for 2 h and thesamples were postfixed for 2 h with 1% osmic acidin cacodylate buffer (pH 7.2). After standard de-hydration in an ethanol series, specimens wereembedded in an Epon812-Araldite mixture.
Sections were obtained with a ReichertUltracut E ultratome (Leica, Nassloch, Germany).Semi-thin sections were stained by conventional
TABLE 2. Effect of the Application of 2.5 fg/Larva of Fenoxycarb for Four Consecutive Days
IGR dose (cumulative) Length of fifth instar (day) Mortality (%) Cocoon weight (g ± S.D.) Shell weight (g ± S.D.)
None 8 0 1.98 ± 0.216 0.43 ± 0.0342.5 fg (oral) 9 34 2.05 ± 0.230 0.46 ± 0.049*2.5 fg (topical) 10 34 2.21 ± 0.237** 0.55 ± 0.063**
*P < 0.05.**P < 0.01.
22Leonardi et al.
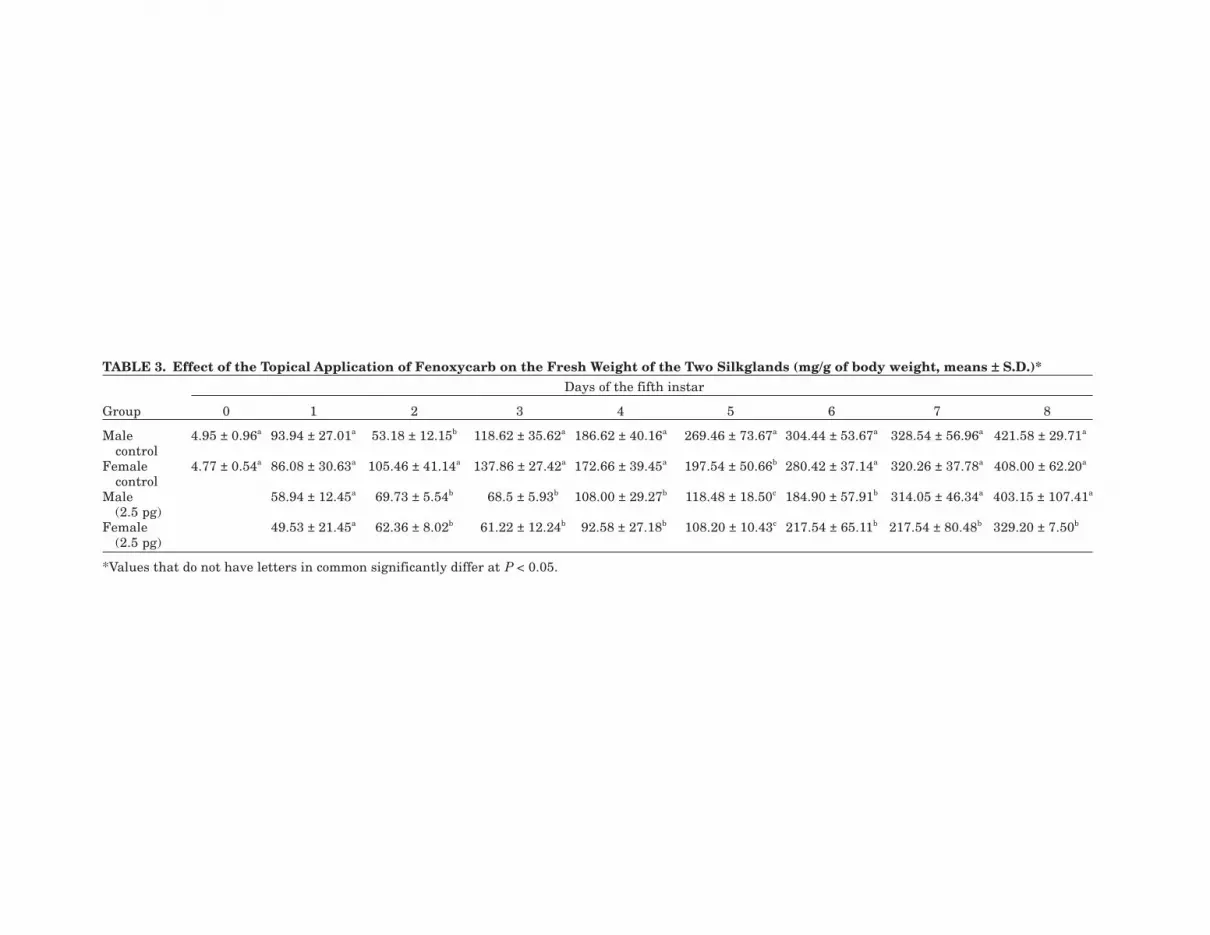
TABLE 3. Effect of the Topical Application of Fenoxycarb on the Fresh Weight of the Two Silkglands (mg/g of body weight, means ± S.D.)*Days of the fifth instar
Group 0 1 2 3 4 5 6 7 8
Male 4.95 ± 0.96a 93.94 ± 27.01a 53.18 ± 12.15b 118.62 ± 35.62a 186.62 ± 40.16a 269.46 ± 73.67a 304.44 ± 53.67a 328.54 ± 56.96a 421.58 ± 29.71a
controlFemale 4.77 ± 0.54a 86.08 ± 30.63a 105.46 ± 41.14a 137.86 ± 27.42a 172.66 ± 39.45a 197.54 ± 50.66b 280.42 ± 37.14a 320.26 ± 37.78a 408.00 ± 62.20a
controlMale 58.94 ± 12.45a 69.73 ± 5.54b 68.5 ± 5.93b 108.00 ± 29.27b 118.48 ± 18.50c 184.90 ± 57.91b 314.05 ± 46.34a 403.15 ± 107.41a
(2.5 pg)Female 49.53 ± 21.45a 62.36 ± 8.02b 61.22 ± 12.24b 92.58 ± 27.18b 108.20 ± 10.43c 217.54 ± 65.11b 217.54 ± 80.48b 329.20 ± 7.50b
(2.5 pg)
*Values that do not have letters in common significantly differ at P < 0.05.
Fenoxycarb and Midgut Functions 23
methods (crystal violet, basic fuchsin) and thenobserved in an Olympus light microscope.
Thin sections were stained with uranyl ac-etate and lead citrate and were observed with aJeol (Peabody, MA) 1010 electron-microscope.
RESULTS
Effects of a Single Application on FeedingBehaviour and Nutritional Parameters
As expected, in the groups of larvae exposedto fenoxycab, the spinning was retarded or sup-
pressed according to the dose applied either topi-cally or orally. Controls started to spin on theeighth day, 2.5 pg-treated larvae on the eleventhday, while larvae treated with the highest dosedied as dauer larvae. The single application of2.5 fg fenoxycarb, either topically or orally, didnot delay spinning.
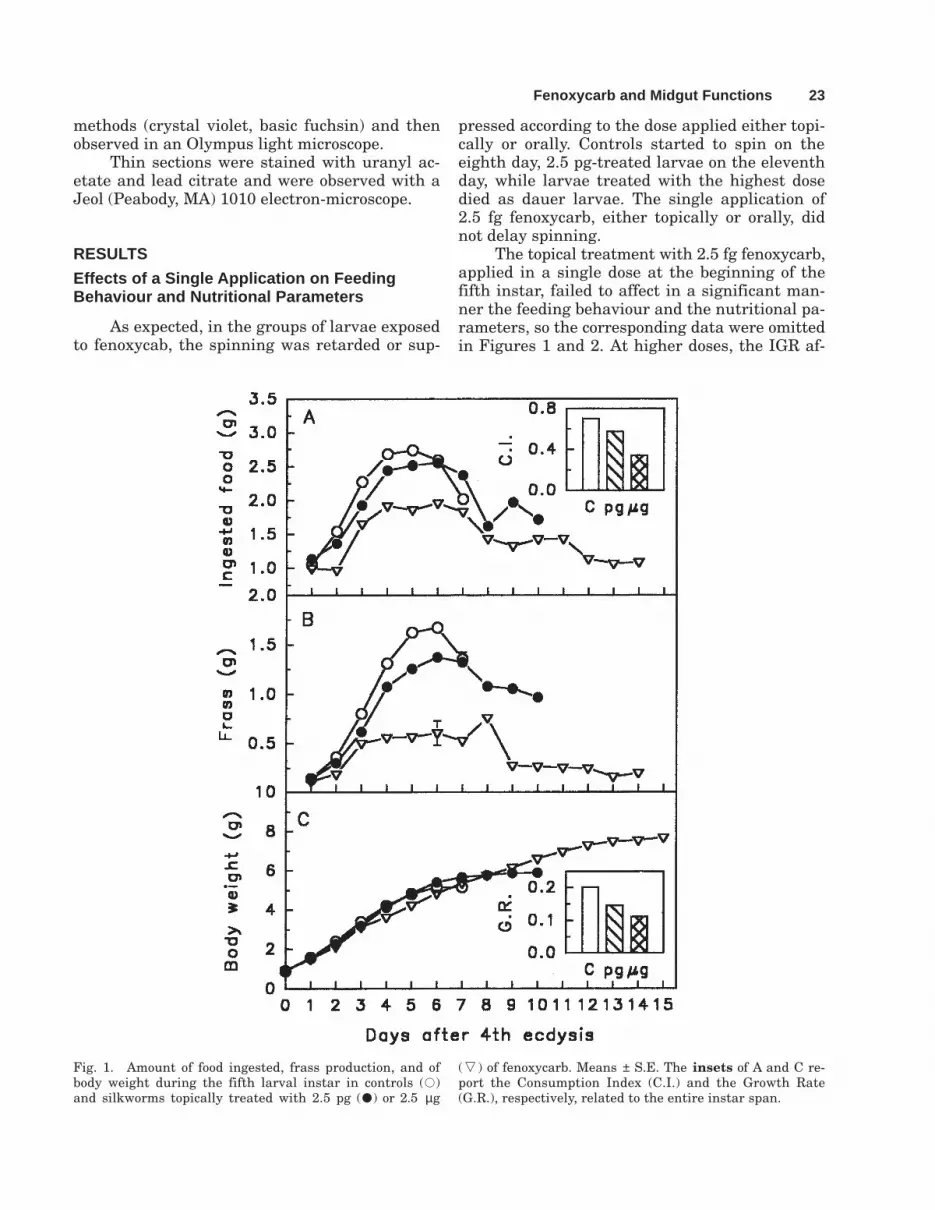
The topical treatment with 2.5 fg fenoxycarb,applied in a single dose at the beginning of thefifth instar, failed to affect in a significant man-ner the feeding behaviour and the nutritional pa-rameters, so the corresponding data were omittedin Figures 1 and 2. At higher doses, the IGR af-
Fig. 1. Amount of food ingested, frass production, and ofbody weight during the fifth larval instar in controls (7)and silkworms topically treated with 2.5 pg (2) or 2.5 µg
(S) of fenoxycarb. Means ± S.E. The insets of A and C re-port the Consumption Index (C.I.) and the Growth Rate(G.R.), respectively, related to the entire instar span.

24 Leonardi et al.
fected voluntary food ingestion of silkworm lar-vae during the entire length of the instar (Fig.1A). The effect varied according to the dose andwas significant (analysed by Student’s t-test)from the second day of the last instar. Identi-cal results were recorded with the oral inges-tion of the IGR as regards this and all theparameters considered in this section, so onlythe data obtained with the topical applicationare reported in Figures 1 and 2.
Voluntary food ingestion in silkworms intheir last instar can be divided into three phases:(1) the poor appetite stage, immediately after
moulting until the second day, which is followedby (2) the good appetite stage until the fifth day,and (3) the pre-spinning stage, which comes justbefore spinning and is characterised again by lowfood consumpion. Considering the increase inbody weight during the feeding period in controland topically treated larvae (Fig. 1C), it was ob-served that growth curves of similar shape wererecorded, but the increase in weight was delayedin treated larvae. The weight in control andtreated larvae was the same during the firststage, controls were much heavier than the 2.5µg-treated larvae during the second stage, while
Fig. 2. Nutritional parameters during the fifth larval in-star in controls (7) and silkworms topically treated with2.5 pg (2) or 2.5 µg (S) of fenoxycarb. AD: ApproximateDigestibility, E.C.I.: Efficiency of Conversion of Ingested food
to body substance, E.C.D.: Efficiency of Conversion of Di-gested food to body substance. Means ± S.E. The inset of Creports E.C.D referred to the entire instar span.

Fenoxycarb and Midgut Functions 25
the weight of all treated larvae was higher in thethird period.
One of the most remarkable and early ef-fects of fenoxycarb was the drastic reduction offrass production not only in orally treated larvae,where it could be expected as a result of the feed-ing inhibition behaviour associated with the in-gestion of potentially toxic compounds (Mulderand Gljswijt, 1973; Abid et al., 1978; Retnakaranet al., 1983) but also in topically treated silkworms(Fig. 1B). It should be observed that larval weightdid not decrease in the first 3 days in 2.5 µg-treatedlarvae even though they had reduced feeding (Fig.1A), because the reduction of defecation was alsoalmost immediate and urination did not occur atall. This low frass excretion, due to the insecticideaction, affected all the physiological parameters con-sidered in this study, i.e., CI (inset in Fig. 1A), GR(inset in Fig. 1C), AD (Fig. 2A), ECI (Fig. 2B), ECD(Fig. 2C). For example, AD was much higher intreated than in control insects, since treated silk-worms ate less but excreted even smaller amountsof faeces than controls. In this case, the parameterwas not an index of midgut functionality; on thecontrary, it indicated that the digestive apparatuswas not working properly.
If the growth rate is calculated for the wholefifth instar (inset in Fig. 1C; 7 and 10 days be-fore spinning for controls and 2.5 pg-treated lar-vae, 15 days before the end of weight gain for 2.5µg-treated silkworms), it is evident that treatedinsects were not able to grow as fast as the con-trols. Furthermore, ECD related to the whole lastinstar (inset in Fig. 2C) suggests that treated lar-vae were not able to convert food to body weightas efficaciously as controls.
The µg-dose caused 100% dauer larvae while2.5 pg-dose resulted in 55.3% mortality, which oc-curred mostly during pupation, inside the cocoon.
Effect of a Continuous Application of fg-Dosesto Silkworms
Since no significant differences, either positiveor negative, could be observed on the nutritionalparameters by treating the silkworms with a singleapplication of a dose of 2.5 fg, despite the fact thatin a previous study such a low dose of the IGR, inits commercial formulation Insegar, did effectivelyexert some influence on midgut functions (Leonardiet al., 1996; see next paragraph), a set of experi-ments was performed in which the 2.5 fg-dose offenoxycarb, applied either topically or orally, wasadministrated on 4 consecutive days early in thelast instar. The results are reported in Table 2. Spin-
ning in control larvae started on the eighth day,while orally treated larvae started one day laterand topically treated silkworms two days later. Be-ginning of spinning was less synchronous withintreated groups. Corrected mortality was signifi-cantly higher (P < 0.01) in treated larvae, thoughnot different between the two modes of administra-tion. Mortality occurred mainly at the time of spin-ning, the larvae remaining dauer and forming loosecocoons, or dead within the cocoon not metamor-phosed into pupae. No larval-pupal intermediateswere observed.
Effect on the Silk Glands
The effect of fenoxycarb on the silk glandsis shown in Table 3. The chemical caused the in-hibition of silk gland growth from the beginningof the fifth instar (second-third day), both in themale and in the female silkworm, while there wasa certain recovery at the time of control spinning.The weight of the two silk glands is expressed asmg/g of larval body weight. In fact, as previouslystated, fenoxycarb affected the growth of thewhole larval body, so a poor growth of the silkglands may have been due to the general impair-ment of the larvae. Nevertheless, it is clear thatfenoxycarb effect on the silk gland growth wasmore remarkable than that on the whole bodyweight. These findings are in agreement with thedata reported in Table 1, where it is shown thatthe cocoon shells spun by 2.5 pg-treated larvaewere not thicker although the cocoons wereheavier and spinning was delayed by 3 days.
BBMVs and Leucine Uptake
In the silkworm, midgut absorption of theamino acids occurs along its entire length but pre-sents functional differences between the anterior-middle (AM) and the posterior (P) regions (Gioraet al., 1994; Shinbo et al., 1996). These regionsare also characterized by marked morphologicaldifferences, especially as regards the goblet cells(Cioffi, 1979) and the extension of the brush bor-der of columnar cells (de Eguileor, personal com-munication). We have investigated the effects ofthe IGR on the ability of the gut epithelium toperform the absorption of amino acids, by pre-paring BBMV from the two midgut regions andmeasuring the uptake of labelled leucine into thevesicles. Figure 3 reports initial uptake rate ofthe amino acid in AM- (Fig. 3A and B) and P-BBMV (Fig. 3C and D) in controls and in silk-worms exposed to either topical (Fig. 3A,C) or oral(Fig. 3B,D) applications of the IGR.

26 Leonardi et al.
Fig. 3. Leucine uptakes into BBMV from the anterior-middle (A,B) and posterior regions (C,D) of the midgut ofcontrols and silkworms topically or orally treated with 2.5
fg, 2.5 pg and 2.5 µg of fenoxycarb. Bars represent means ±S.E. of a typical experiment performed in triplicate. **P <0.01, ***P < 0.001.
The data obtained as a result of the topicaltreatments substantially confirm our previous re-sults with the commercial formulation, Insegar(Leonardi et al., 1996): a consistent inhibition ofleucine uptake was observed at a high dose ofthe IGR, while a significant increase was causedby the application of a single dose of 2.5 fg. Boththe negative and positive effects were moremarked in AM-BBMV than in P-BBMV.
Conversely, the oral application of the IGRdid not have marked effects on leucine uptakerates: uptake was significantly reduced with thehigh dose in AM-BBMV but not in P-BBMV, anda barely detectable increase of the uptake wasseen with 2.5 fg in P-BBMV.
Histochemistry
A morphological and histochemical charac-terization of the midgut epithelium in larvaetreated with the maximal or lowest dose by topi-cal application was performed. The silkworm mid-guts showed a peculiar histochemical picture inresponse to different fenoxycarb dosages.
The control epithelium was strongly stained
for both NADH tetrazolium reductase and suc-cinic dehydrogenase reactions and the stain pro-portionally decreased according to the dose in thespecimens from treated larvae (Figs. 4–9). Thecontrol epithelium was lightly stained while theoil red O stain increased in intensity in thetreated specimens, with a strong positive stain-ing in the midguts from larvae treated with 2.5µg of fenoxycarb (Figs. 10–12). Alkaline phos-phatase reaction, localized at the brush bordermembrane of the epithelial cells, was stronglypositive in control epithelia and in specimens fromlarvae treated with the low dose while the staindramatically decreased in the epithelia from lar-vae treated with the high dose of the IGR (Figs.13–15). Acid phosphatase staining was completelynegative in controls and in larvae treated with2.5 µg (Figs. 16, 18), while it was detectable inmidguts from larvae treated with 2.5 fg (Fig. 17).The epithelia of controls and low dose-treated lar-vae were not stained with PAS reagent (Figs. 19,20) while a positive reaction was evident in themidgut cells of larvae exposed to 2.5 µg of theIGR, where many granules could be observed (Fig.

Fenoxycarb and Midgut Functions 27
Figs. 4–6. Frozen sections. NADH-tetrazolium reductasethe activity of which reflects the capacity of intracellularoxidation. The black intense staining (arrowheads) de-creased clearly in relation to the dose of fenoxycarb. 4,control × 33; 5, dose of 2.5 fg/larva x 65; 6, dose of 2.5 µg/larva × 65.Figs. 7–9. Frozen sections. SDH (succinic dehydrogenase)reflecting the mithocondrial activity. The black intense stain-
ing (arrowheads) practically disappeared with the higherdose of the IGR (Fig. 9). × 65.Figs. 10–12. Frozen sections. O.R.O. The positivity of thereaction, evidenced by a conspicuous amount of magenta redgranules, increased in relation to the dose of fenoxycarb.With 2.5 µg/larva (Fig. 12), lipid metabolism was drasticallyaltered. 10, control × 65; 11, dose of 2.5 fg/larva × 65; 12,dose of 2.5 µg/larva × 33.

28 Leonardi et al.
Figs. 13–15. Frozen sections. The alkaline phosphatase ac-tivity, marker of brush border membrane functional integ-rity, was strongly positive (arrowheads) at the level of thebrush border in control and low-dose treated larvae (Figs.13, 14). The positivity dramatically decreased in the epithe-lium from µg-treated insects. × 65.Figs. 16–18. Frozen sections. Acid phosphatase, markerof lysosomal activity. The reaction was negative in con-
trols and in larvae treated with 2.5 µg. An evident posi-tivity (arrowheads) was present in specimens from 2.5 fg-treated larvae. × 65.Figs. 19–21. Frozen sections. PAS. The epithelium of con-trols and low dose-treated larvae were unstained while alarge amount of granules (arrowheads) were evident in lar-vae heavily exposed to fenoxycarb. × 65.

Fenoxycarb and Midgut Functions 29
21). The qualitative changes caused by IGR treat-ment are summarized in Table 4.
Optical and Electron Microscopy
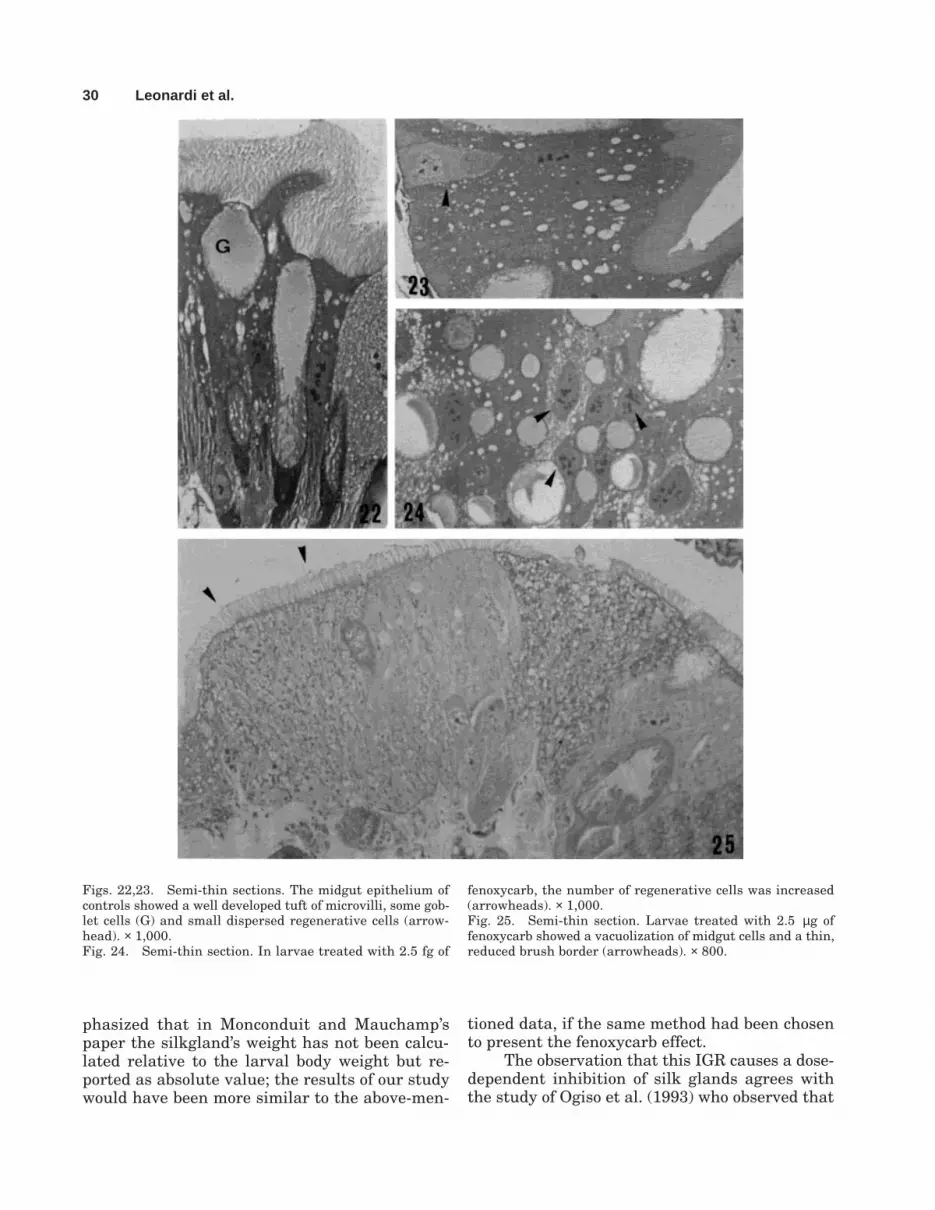
The larval midgut epithelium of B. mori iscomposed, as in all lepidopterans, of three typesof cells: columnar, goblet, and regenerative cells(Figs. 22–29).
Each columnar cell has a thick brush borderat the apical border, which appeared reduced intreated larvae (Figs. 25, 30, 31). In larvae exposedto the high dose of fenoxycarb, midgut microvilliwere few and shorter than those of control or lowdose-treated ones, and the cytoplasm showed a sig-nificant increase of vesicles (Figs. 30–32).
Morphology of the goblet cells was unaltered(Fig. 28). The regenerative cells, responsible forthe growth and the maintenance of the epithe-lium, as well as for the substitution of damagedcells, appeared more abundant with fenoxycarbapplication of 2.5 fg (Fig. 24). These cells were infact in S-phase as demonstrated by incorporationof BrdU (Fig. 33).
DISCUSSION
It was already demonstrated (Cappellozzaand Cappellozza, 1994) that the continuous in-gestion of fenoxycarb by silkworm larvae duringthe fifth instar results in a more harmful effectin comparison to the topical one-dose treatmentapplied at the beginning of the instar, even if ina higher amount. In addition, it was also shownthat if the same amount used in topical treat-ment was divided into several doses and appliedon 5 consecutive days, a further delay in spin-ning and an increase in the mortality rate werecaused (Cappellozza and Cappellozza, 1994). Amortality rate around 34% was recorded whenthe larvae were given 10 fg of the chemical onconsecutive days, while Kamimura (1995) ob-served a mortality rate of 22% in fifth instar
silkworm larvae when the animals were topi-cally-treated with 1 pg of fenoxycarb at the be-ginning of the last instar. The results of theexperiments carried out in the current studyagree with these previous findings.
On the other hand, this study gives evidencethat there are some differences between the oraland topical treatments even if they are adminis-tered on the same time schedule. These differ-ences concern the progress of the life cycle fromthe juvenile to the adult stage and silk produc-tion (Table 2; see later in the Discussion for thedifferent effects on leucine transport).
The action of the IGR on the feeding be-haviour and the nutritional parameters has beenwidely described in Results. The relationship be-tween these parameters and the silk glands isworthy of consideration. In fact, the growth ofthese glands is inhibited at a dose of 2.5 pg, whichaffects also the physiology of the midgut and theabove-mentioned parameters. As stated in Re-sults, the cocoon shell spun by 2.5 pg-treated lar-vae was not thicker than that of the controlsdespite a 3-day delay in spinning. Worthy of fur-ther investigations is to which extent the absorp-tion processes by the midgut are affected byfenoxycarb (even if leucine transport does notseem to be influenced at this concentration) andto which extent these mechanisms are involvedin the response of the silkgland to the IGR. Alarge number of data indicate that fenoxycarb ata dosage near to the LD50 does not act as a JH-mimic, its effects being more similar to those of anon specific toxic drug than to that of a JH-ana-logue such as methoprene. However, despite an al-teration of some enzyme activities (Table 4), aconsistent activation of leucine transport was ob-served (Fig. 3) when the larva was topically treatedwith 2.5 fg of IGR in which it is conceivable thatit may act as a JH-mimic. Amino acid absorptionthrough the gut is increased, and possibly this isresponsible for the increased weight of the cocoonshell, when silkworms were subjected to repeated(four times) topical application of a low dose offenoxycarb (Table 2).
Monconduit and Mauchamp (1997) observeda similar dose-dependent stimulation-inhibitionof silk gland growth in a polyvoltine silkwormstrain, in which the treatment with 100 pg/larvaproduced larger silk glands (after the period whencontrol larvae completed spinning; while no dif-ferences were recorded in the first part of the in-star), while 1 ng of fenoxycarb/larva stronglyinhibited silk gland development. It should be em-
TABLE 4. Summary of the Histochemical ResultsObserved in the Midgut Epithelium of Controls andSilkworms Topically Treated with Fenoxycarb
Reaction Controls 2.5 fg 2.5 µg
NADH-TR +++ ++ —SDH +++ + —O.R.O — ++ +++AlP +++ +++ +AcP — ++ —PAS — + +++
30 Leonardi et al.
Figs. 22,23. Semi-thin sections. The midgut epithelium ofcontrols showed a well developed tuft of microvilli, some gob-let cells (G) and small dispersed regenerative cells (arrow-head). × 1,000.Fig. 24. Semi-thin section. In larvae treated with 2.5 fg of
fenoxycarb, the number of regenerative cells was increased(arrowheads). × 1,000.Fig. 25. Semi-thin section. Larvae treated with 2.5 µg offenoxycarb showed a vacuolization of midgut cells and a thin,reduced brush border (arrowheads). × 800.
phasized that in Monconduit and Mauchamp’spaper the silkgland’s weight has not been calcu-lated relative to the larval body weight but re-ported as absolute value; the results of our studywould have been more similar to the above-men-
tioned data, if the same method had been chosento present the fenoxycarb effect.
The observation that this IGR causes a dose-dependent inhibition of silk glands agrees withthe study of Ogiso et al. (1993) who observed that

Fenoxycarb and Midgut Functions 31
Figs. 26–29. Thin sections. In control midguts were visible:the epithelial columnar cells (arrowheads), some goblet cells(G) with their typical closing system (arrow) and some cells(E) characterised by a developed reticulum. The columnar
cells showed at the apical border a thick brush border (ar-rowhead) and some dispersed vesicles of cytoplasm (VC). 26,×2,000; 27, × 3,100; 28, × 4,500; 29, × 2,000.

32 Leonardi et al.
Figs. 30,31. Thin sections showing the reduced brush bor-der (arrowheads) in relation to the dose of fenoxycarb ap-plied (Fig. 30: 2.5 fg × 12,500; Fig. 31: 2.5 µg × 5,000).Fig. 32. Thin section. The cytoplasm of treated larvaeshowed a vacuolization (arrowheads). × 3,700.
Fig. 33. Frozen section. On the basis of BrdU incorpora-tion, actively proliferating elements could be identified inspecimens from treated larvae.

Fenoxycarb and Midgut Functions 33
fenoxycarb treatment of last instar silkworm lar-vae affected the weight of the cocoon shell, evenif heavier cocoons were produced. This phenom-enon is not peculiar to fenoxycarb but it has beenalready recorded as an effect of juvenile hormone(JH) and JH-analogues on the silkworms. Chow-dary et al. (1986, 1990) explained that the in-crease in silk weight due to the prolongation ofthe larval period induced by some JH-analogueslasted for only 1–3 days; afterwards, the silk ra-tio was negative because there was a greater in-crease in the pupal weight than in the silk layers.The same behaviour was found in methoprene-treated larvae at concentrations higher than 10ppm. In addition, low-dosage treatments withmethoprene inhibited silk gland growth in com-parison to that of the controls. Silk gland weightsat silkworm maturity were greater than those ofthe control group only because of the one-day pro-longation of the last instar (Anonymous, 1973).
The silkworm midgut performs a complexseries of activities to accomplish a large spectrumof tasks, which include the ability to reduce tox-icity of allelochemicals and bacteria (Appel, 1994)and the ability to extract nutrients from leavesin order to maintain the ionic and chemical ho-meostasis. A number of functional differentiationshave been observed between the morphologicallydifferent midgut regions (Cioffi, 1979), e.g., pro-duction of digestive enzymes (Terra, 1990) andthe ability to absorb amino acids (Giordana et al.,1994; Shinbo et al., 1996). We have preparedBBMV from the anterior-middle and posterior re-gions to detect the influence of the IGR on aminoacid absorption.
Leucine absorption appears to be more ex-tensively affected by the IGR in the anterior-middle region (Figs. 3A and C). The percentactivation of leucine uptake at a low dose as wellas the percent inhibition of transport at a highdose were larger in AM- than in P-BBMV. More-over, the IGR administered in a single dose ismore effective by topical application than by in-gestion. Probably, the insecticide is readily per-meable through the larval cuticle and epidermisand can be rapidly conveyed to the insect internalenvironment when dissolved in ethanol. Conversely,ingested fenoxycarb may encounter physico-chemi-cal alterations in the midgut lumen, filled with anextremely alkaline fluid rich in active enzymes, andhas to cross the epithelial barrier to reach the silk-worm hemolymph. These events may justify thesmaller effects on amino acid uptake. However, theoverall effects of the ingested IGR on gut functions
and larval behaviour are more extensive and com-plex than those exerted on amino acid absorp-tion, since the dose-response relationship on thenutritional parameters was similar with the twokinds of administration.
The analysis of midgut cell morphology andfine structure as well as the histochemical stainingsperformed to investigate enzyme activities and lipidand carbohydrate metabolism clearly show thatfenoxycarb introduced directly into the insecthemolymph causes a definite alteration of midgutcolumnar cell morphology and biochemical activi-ties. The midgut epithelium of the larvae exposedto a high dose showed cells with a vacuolated cy-toplasm and shorter and irregular microvilli. Atthe same time, an activation of the regenerativecapability of the epithelium was apparent inspecimens from low dose-treated larvae. A simi-lar regenerative process was observed in the mid-gut epithelium of Manduca sexta larvae after theoral administration of sublethal amounts of acrystal endotoxin of Bacillus thuringiensis (Spiesand Spence, 1985).
The histochemical investigation suggested thepresence of altered metabolic processes in midgutcells following treatement of larvae with the highor the low dose of the IGR. An increased activity ofacid phosphatase was detectable after applicationof 2.5 fg (Fig. 17, Table 4). In agreement with thealteration of the morphology of the microvilli ob-served with the high dose of fenoxycarb, the brushborder membrane marker enzyme alkaline phos-phatase, typical of absorptive epithelia, was stronglyinhibited. This finding agrees with the relevant re-duction of leucine uptake into AM- and P-BBMVfrom µg-treated larvae (Fig. 3A and C).
Fenoxycarb was responsible for a severe dis-ruption of lipid and carbohydrate metabolism inthe fat body of Choristoneura fumiferana topicallytreated with a dose of 0.26 µg/insect (Mulye andGordon, 1993a). The specific histochemical reac-tions show that this is true also for B. mori mid-gut epithelial cells. A large number of stainedgranules, showing the presence of abnormalamounts of polysaccharides, were detected in mid-gut specimens from µg-treated larvae stained withPAS reaction (Fig. 21). Similarly, O.R.O stainingindicated an alteration of lipid metabolism, withan accumulation of neutral lipids with increas-ing doses of the IGR (Figs. 11, 12). Instead, inthe fat body of C. fumiferana fenoxycarb causeda reduction of lipid biosynthesis, with a suppres-sion of the synthesis of complex lipids (Mulye andGordon, 1993b).

34 Leonardi et al.
The accumulation of fuel molecules (carbo-hydrates and lipids) observed in the midgut cellsof treated larvae is associated with a strong de-crease of cellular reducing power as indicated bythe large reduction of NADH (Figs. 5, 6). In addi-tion, the activity of the mitochondrial enzymeSDH was clearly lowered in specimens from lowdose-treated larvae (Fig. 8). Even if the aminoacids are the major source for energy metabolism,which is mainly aerobic in B. mori midgut (Par-enti et al., 1985), the histochemical resultsstrongly suggest a considerable impairment ofenergy generation.
The administration of fenoxycarb at a dose(fg) which may mimic hormonal effects, as sug-gested by the increased rate of uptake of leucinein AM-BBMV (Fig. 3), seems to cause an impair-ment of some metabolic processes. However, theparallel activation, as a response to the noxiouseffect, of the regenerative cells, normally presentas a scanty number of single cells at the base ofthe epithelium during the feeding period of fifthinstar larvae (Turbeck, 1974), with their subse-quent entry into S-phase (Fig. 33), may providean adequate recovery of midgut metabolic andbarrier functions, which can explain the lack ofeffect on the nutritional parameters.
In conclusion, the lepidopteran midgut is alarval organ that responds promptly to the expo-sure of the insect to fenoxycarb, when the chemi-cal reaches the epithelium from either the luminalor the basal side. The columnar cells modify theirabsorptive functions, at least as regards aminoacid uptake, and their metabolic activity, with amodification of the oxidative status of the cellsalready detectable at a dose of the IGR as low asfew femtograms/larva. Whether this representsa JH-like effect or not remains to be determined.
ACKNOWLEDGMENTS
This research was supported by a grant fromthe Ministero dell’Università e della RicercaScientifica e Tecnologica, Italy (MURST ex 60%).
LITERATURE CITED
Abid MK, Chobrial A, El Haideri HS, Abbas SA (1978):Dimilin (TH-6040): Effects on the spiny bollworm,Earias insulana Boisd. (Lepidoptera: Phalaenidae). ZAm Gew Ent 85:321–324.
Anonymous (1973): Cocoon and silk enhancing agent MantaR
for silkworm culture: handbook of the use of MantaR.Zoecon Corporation, Palo Alto, CA: Zoecon Corporation,pp. 24
Appel HM (1994): The chewing herbivore gut lumen: physi-cochemical conditions and their impact. In BernaysEA (ed): Insect-Plant Interactions. Boca Raton: CRCPress, vol. V, pp. 209–223.
Cappellozza L, Miotto F, Moretto E (1990): Effetti delfenoxycarb a basse concentrazioni sulle larve diBombyx mori (Lepidoptera Bombycidae). REDIA 73:517–529.
Cappellozza L, Cappellozza S (1994): Relation between thespinning of the cocoon and oral and topical treatmentswith various dosages of fenoxycarb in a Bombyx moristrain. REDIA 77:369–374.
Cappellozza L, Cappellozza S, Sbrenna G (1995): Changesin the developmental programme of Bombyx moricaused by oral treatment with fenoxycarb during thelast larval instar. Sericologia 35:427–436.
Chowdary SK, Raju PS, Ogra RK (1990): Effect of JH-ana-logues on silkworm, Bombyx mori L., growth and de-velopment of silk gland. Sericologia 30:155165.
Chowdary SK, Senhal F, Ray SK, Raju PS, Mathu Sarita(1986): Giant cocoon formation in Bombyx mori L.,topically treated with juvenile hormone SJ-42-F.Sericologia 26:455–459.
Cioffi M (1979): The morphology and fine structure of thelarval midgut of a moth (Manduca sexta) in relationto active ion transport. Tissue Cell 11:467–479.
Dedos GS, Fugo H (1992): The usage of fenoxycarb and itsinfluence on sericulture. Seric Sci Technol 31:30–33.
Dedos GS, Asahina M, Fugo H (1993): Effect of fenoxycarbapplication on the pupal-adult development of the silk-worm . J Seric Sci Jpn 62:276–285.
Dedos GS, Fugo H (1996): Effect of fenoxycarb on the secre-tory activity of the prothoracic glands in the fifth instarof the silkworm Bombyx mori. Gen Comp Endocrinol 104:213–224.
Dubowitz V, Brooke MH (1973): Muscle Biopsy: A ModernApproach. London: Saunders.
Giordana B, Leonardi MG, Tasca M, Villa M, Parenti P.(1994): The amino acid/K+ symporters for neutralamino acids along the midgut of lepidopteran larvae:functional differentiations. J Insect Physiol 40:1059–1068.
Gordon R, Chippett J, Tilley J (1996): Effects of two car-bamates on infective juveniles of Steinernema carpo-capsae All strain and Steinernema feltiae Umea strain.J Nematol 28:310–317.
Grenier S, Grenier AM (1993): Fenoxycarb, a fairly new In-sect Growth Regulator: A review of its effects on in-sects. Ann Appl Biol 122:369–403.
Hanozet GM, Giordana B, Sacchi VF (1980): K+-dependent

Fenoxycarb and Midgut Functions 35
phenylalanine uptake in membrane vesicles isolatedfrom the midgut of Philosamia cynthia larvae. BiochimBiophys Acta 596:481–486.
Kamimura M (1995): Effects of a juvenile hormone analogue,fenoxycarb, on larval growth of the silkworm Bombyxmori (Lepidoptera: Bombycidae). Appl Entomol Zool30:487–489.
Leonardi MG, Cappellozza S, Ianne P, Cappellozza L, ParentiP, Giordana B (1996): Effects of the topical applica-tion of an Insect Growth Regulator (Fenoxycarb) onsome physiological parameters in the fifth instar lar-vae of the silkworm Bombyx mori. Comp BiochemPhysiol 113B:361–365.
Monconduit H, Mauchamp B (1997): Effects of the non-ter-penoid carbamate, fenoxycarb, on JH-regulated param-eters in the silkworm, Bombyx mori. XVIIth Congressof International Sericultural Commission, 22-26thApril, Londrina, Brazil.
Moreno J, Hawlitzky N, Jimenez R (1993): Effect of the ju-venile hormone analogue fenoxycarb applied via thehost on the parasitoid Phanerotoma (Phanerotoma)ocularis Kohl (Hym., Braconidae). J Insect Physiol 39:183–186.
Mulder R, Gljswijt MJ (1973): The laboratory evaluation oftwo promising new insecticides which interfere withcuticle deposition. Pestic Sci 4:149–158.
Mulye H, Gordon R (1993a): Effects of two hormone juve-nile analogs on hemolymph and fat-body metabolitesof the eastern spruce budworm, Choristoneura fumi-ferana (Clemens) (Lepidoptera: Tortricidae). Can J Zool71:1169–1174.
Mulye H, Gordon R (1993b): Effects of Fenoxycarb, a juve-nile hormone analog, on lipid metabolism of the east-ern spruce budworm, Choristoneura fumiferana. JInsect Physiol 39:721–727.
Ogiso M, Okhawa K, Asayama T (1993): Influence of fen-oxycarb on the development of Bombyx mori. Proceed-ings of the 63rd Annual Meeting of the JapaneseSericultural Society, April 67th, Ustunomiya, IbarakiPrefec., Japan, 13.
Parenti P, Giordana B, Sacchi VF, Hanozet GM, GuerritoreA (1985): Metabolic activity related to the potassiumpump in the midgut of Bombyx mori larvae. J ExpBiol 116:69–78.
Plantevin G, Grenier S, Chavancy G (1991): Effets d’unRégulateur de Croissance d’Insectes, le fénoxycarbe,sur le développement postembryonnaire de Bombyxmori (Lepidoptera, Bombycidae). C R Acad Sci Paris313:513–519.
Retnakaran A, Lauzon H, Fast P (1983): Bacillus thurin-giensis induced anorexia in the spruce budworm,Choristoneura fumiferana. Ent Exp Appl 34:233–239.
Shinbo H, Konno K, Hirayama C, Watanabe K (1996): Di-gestive sites of dietary proteins and absorptive sitesof amino acids along the midgut of the silkworm,Bombyx mori. J Insect Physiol 42:1129–1138.
Spies AG, Spence KD (1985): Effect of sublethal Bacillusthuringiensis crystal endotoxin treatment on the lar-val midgut of a moth, Manduca: SEM study. TissueCell 17:379–394.
Soo Hoo CF, Fraenkel G (1966): The consumption, diges-tion and utilization of food plants by a poliphagousinsect, Prodenia eridania (Cramer). J Insect Physiol12:711–730.
Stoka AM (1996): Activity of the juvenile hormone and juve-nile hormone analogues on the growth of Tripanosomacruzi. J Steroid Biochem Mol Biol 59:495–500.
Terra W (1990): Evolution of digestive systems of insects.Ann Rev Entomol 35:181–200.
Tremblay E (1982): Generalità e mezzi di controllo. InEntomologia applicata. Napoli: Liguori Editore, Vol.1, pp.1–155.
Turbeck BO (1974): A study of the concentrically laminatedconcretions, ‘spherites,’ in the regenerative cells of themidgut of lepidopterous larvae. Tissue Cell 6:627–640.
Walbauer GP (1968): The consumption and utilization of foodby insects. Adv Insect Physiol 5:229–282.
![The optical/NIR counterpart of the INTEGRAL obscured source IGR J16318-4848 : a sgB[e] in a HMXB ?](https://img.dokumen.tips/doc/110x75/635077338624f8c9710e835b/the-opticalnir-counterpart-of-the-integral-obscured-source-igr-j16318-4848-a.jpg)




























