Embed Size (px)
Citation preview

Medical Hypotheses 7: 285-310, 1981
MOBIUS DNA,THE PAPER TIGER
B. Daunter, P. Silburn and R. Hill. Department of Obstetrics and
Gynaecology, Clinical Sciences Building, Royal Brisbane Hospital,
Brisbane, Queensland, Austral ia.
ABSTRACT
Deoxyribonucleic acid (DNA), the biological molecule that deter-
mines the evolutionary architecture of all known life forms has been
largely ignored as regards its own evolution from the prokaryotic ccl 1
(ccl 1 s without nuclear membranes), to the eukaryotic ccl 1 (ccl 1s with
nuclear membranes). In addition, the properties of prokaryotic DNA,
have in some cases, been imposed on eukaryotic DNA resulting in the loss of evolutionary continuity of DNA. Regardless of the source of DNA,
only small sections are usually examined as a salt extract. It is
devoid of its associated proteins, which in the eukaryotic cell are
responsible for the packing of DNA into chromosomes, and from its intimate connections with the nuclear membrane. From such studies
various structures for DNA have been proposed which have not found
general acceptance. Thus, any hypothesis on the structure of DNA must be able to incorporate most, if not al 1, other reported structures. Similarly, any structure proposed for DNA must be considered in terms of its own evolution as distinct from its phenotypic evolutionary
expression.
A single strand of DNA has opposite terminal ends, a 3’OH group and
a 5’ phosphate group. These ends may join by a 5’ - 3’ phosphate 1 ink-
age forming circular DNA. Both these single stranded linear and cir-
cular forms of DNA are considered to belong to “primitive” prokaryotic ccl Is. A higher order of circular prokaryotic DNA may be formed if at one end, a single strand of DNA is turned through 1800, resulting in the
3’OH and 5’ phosphate groups being diagonally opposite each other (3’/
5’). This circular DNA has a continuous phosphate backbone and a discontinuous base sequence, Mobius circular DNA. The discontinuous base sequence may provide a start/stop signal for DNA and RNA polymerase. This Mobius DNA (M. 1.) may give rise to single circular, double linear
or double circular DNA.
285

During the rep1 ication of M. 1, DNA it may be possible for the two complementary strands to form one continuous Mobius DNA circle (M.2.
DNA) by two 5’/3’ phosphate 1 inkages. Thus, there are two start/stop
signals for DNA and RNA polymerase, therefore DNA rep1 ication would occur as two separate strands and requi.re splicing together. In
addition, M.2. DNA may be the first example of gene duplication and
represent the first “primitive” eukaryotic DNA. Another form of Mobius DNA may be derived from M. I., which represents the higher order in eukaryotic cells. This may be formed by two complementary strands of M.1. forming a Mobius DNA circle (M.3. DNA) by 5’ - 3’ and 5’/3’ phos-
phate 1 inkages. This will result in a double right handed a helical configuration which can be formed into a linear side-by-side configur-
ation with a double loop at one end, which is a left handed helix. The side-by-side configuration allows base pairing, but in the double loop,
the bases are facing in the same direction and are displaced, therefore
they cannot base pair. Thus, single stranded DNA is existing with based pa i red DNA, It is proposed that the single stranded DNA represents unique copy and the base paired DNA repetitive copy, and that a number
of these M.3. DNA’s constitute a gene.
The DNA of eukaryotic cells is wound around a number of histone
cores. Between each core there is a linking region of DNA upon which the histone Hl is attached. The core plus I inker, represents a nucleo- some, and under the electronmicroscope resembles beads on a string. A
number of configurations have been proposed for these core histones
which enable DNA to be packed to form chromosomes. The various con- figurations proposed for the histones can be achieved by considering
each histone as a Mobius circle. The association of these Mobius histones results in the formation of cores in the form of wedge-shaped
cyl inders, which associate to form an helical spring-like solenoid with a central hole, when M.3. DNA undergoes compaction.
INTRODUCTION
Double a He1 ical Structure
Deoxyribonucleic acid (DNA) is a biological molecule that dete;;anes
the architecture of all known life forms directly or indirectly.
possible general .structure and sequence of genetic information carried
by this molecule was first proposed by Watson and Crick in 1853 (1).
This was achieved by comparing stereochemical models of DNA with X-ray
diffraction data of DNA extracts from various sources such as calf
thymus, spermatozoa, leucocytes and bacteria. The results suggested that DNA consists of a pair of very long molecules, polynucleotides, twisted spirally around each other to form a double right hand a hel ix. Each
polynucleotide is made up of mononucleotides which consist of the carbo-
hydrate deoxyri bose, a phosphate group and a nitrogenous base, of which
there are four different types; two pyrimidines, thymine and cytosine
and two purines, adenine and guanine. In each polynucleotide chain the
deoxyribose moieties are linked by phosphate groups via carbon 5’ of one
deoxyribose molecule to carbon 3’ of another (5’ - 3’) and the poly- nucleotide chains run paralIe1 to each other, but in the opposite direction. The two polynucleotide chains are brought in to close
286

FIG. 1.
Double a helix Side by side
association by hydrogen bonding between the nitrogenous bases at carbon
1, on each deoxyribose molecule. Thus the phosphate groups and the bases are stacked above each other to form a double cx helix with regular inter-
nal base pairing which varies to give the genetic code. The base pairing is specific, in that adenine always forms hydrogen bonds with thymine and guanine with cytosine in the opposite polynucleotide chain.
Various reports and proposals suggest that DNA may have more than one conformational state. This has recently been reviewed (2). Fibre diffraction analysis of DNA suggests the existence of three distinct forms
based on the flexibility of the phosphate backbone. However in solution
the structure has always been assumed to be an c1 helix (B conformation).
Similarly, suggestions have been made that some DNA with unusual base
sequences could form somewhat different conformations to that of the
B DNA. This has been verified, to some extent, by the use of synthetic
DNA polymers of simple alternating sequences of adenine and thymine re-
sulting in alternating conformations of the backbone in solution, but
still in the B form. The hexanucleotide guanine and cytosine however have been reported to form a left-handed double he1 ix. This has been referred
to as Z DNA because of the staggered zig-zag course of the backbone. It has been proposed that the B and Z forms of DNA are interchangeable, however , the transition from B to Z occurs at an unphysiological salt con- centration. The transition therefore of DNA from the B form to the Z form in vi~,o is unlikely unless polyamines or specific proteins are involved in the transition.
287

Side by Side He1 ical Structure
Models for DNA not based on the double o helix have also been proposed.
This has been reviewed briefly by Arnott (3). Basically, these models
suggest that the two antiparal lel complementary base-paired polynucleotide
chains lie side by side. One such structure which alternates the helical
conformation every half-repeating distance results in intermeshing rather
than intertwining of the polynucleotide chains (4). Thus the structure
is to some degree, both a right and left-handed helix (Figure 1). A
simi lar structure has been proposed by Sasisekharan and co-workers based
on an alternating right and left-handed helix (5).
Supercoiling of DNA
Regardless of structural preferences for DNA it must be coiled and packed to form chromosomes in the eukaryotic cells (cells with nuclear
membranes). This is partially achieved by winding DNA around a core of histone proteins consisting of two molecules each of histones H2A, H2B,
H3 and H4. The core complex is rigidly held in place by the inter- action of the basic amino termini of the histones and the phosphate
groups of the DNA. Between each core there is a linking DNA strand upon which histone Hl is attached. The core plus linker represents a nucleo- some and under the electronmicroscope resembles beads on a string. There
are 2.5 turns of DNA per nucleosome representing on average 200 base pairs of DNA of which 140 are associated with the core octamer, which is
288

equivalent to 1.75 turns. Thus, on average, 60 base pairs are contained
in the Hl bound linker region imparting another 0.75 turn in DNA. Over-
all this is thought to result in a left-handed solenoidal supercoil of
DNA as it winds from nucleosome to nucleosome (6, 7). This solenoidal
supercoil is then compacted, by an undertermined mechanism, to form the
metaphase or mitotically visible chromosome. Hence the vehicles of in-
heritance are viewed as one long strand of DNA complexed with protein,
RNA and 1 ipids to form chromatin which is wound upon itself to form a
chromosome.
Prokaryot ic DNA
Prokaryotic cells (cells without nuclear membranes), such as
bacteria, have been extensively investigated in terms of DNA structure
and function relative to eukaryotic DNA. The DNA of prokaryot ic ccl 1 s
may be single or double linear or circular. Some prokaryotic DNA may
be considered to form nucleosomes but the majority of such DNA is found
loosely associated with only a few non-histone proteins. Therefore
nucleosome formation is predominantly found in eukaryotic cells (8).
Double stranded DNA Replication
Briefly the replication of double a helical DNA is considered to
involve the unwinding of the a helix by unwinding proteins. This forms
what is called a replication fork which consists of some 200 molecules
of unwinding protein each complexed to 8 - 10 nucleotides resulting in
approximately 2000 unpaired bases. The enzymes DNA polymerase then
replicates short segments of the DNA in the direction of the phosphate
5’ - 3’ 1 inkage (Figure 2) which are then joined by DNA 1 igase (8).
Single Stranded DNA Rep1 ication
The DNA of a number of bacterial viruses exists as a single strand
which serves as a template in the host for the formation of a comple- mentary strand. The resulting double c1 helix then serves as a template for the formation of new single stranded viral DNA. This form of replication is necessary in this case, because if the viral DNA was able
to undergo replication per se the resulting daughter cell would contain
completely different genetic formation (8).
Double Stranded Circular DNA Rep1 ication ---
The DNA of some viruses and bacteria is a duplex ring complexed by
proteins. The removal of these proteins results in the twisting of the
DNA molecule upon itself, to form supercoils, due to a change in the
number of base pairs per turn of the doub)e he1 ix. The chains become
linked together by covalent bonds, any subsequent change in the number
of base pairs per helical turn must then be compensated by the formation
of a supercoil in the opposite direction. Thus any purified circular
DNA containing negative supercoils, coiling opposite to the a helix, is
assumed to be coated with protein during the DNA replication process. However, some purified circular DNA molecules lack supercoiling. In
addition, some DNA vi ruses have 1 i near DNA and in order to undergo replication it is converted into a circle in the host, for example,
bacteriophage X DNA (8).
289

Real ication of Circular DNA
The replication of circular DNA does not necessarily involve the
temporary formation of linear DNA, for example, in E coli. Obviously the untwisting of the circular helix must take place without causing
supercoi 1 ing. It appears that both paternal strands remain intact
throughout most of the replication cycle, the unrepl icated portion be- coming periodically supercoiled. For this to occur it is proposed that transient cuts are made in one of the polynucleotide chains to allow
them to untwist. This results in the DNA taking on the appearance of
a B sign. The rep1 ication of the DNA may start within a section of
covalently closed DNA and replication may cease due to the steric
hinderance of supercoiling of the unreplicated portion. The DNA re-
plication may resume only when the enzyme swivelase creates a revers-
ible single strand cut to relax the supercoiled molecule into an un-
twisted circular form (8).
An alternative way to rep1 icate circular DNA is by the Roll ing Circle technique. This involves a specific cut in the paternal duplex
circle resulting in a polynucleotide strand with a free 3’ OH group on
its carbohydrate (deoxyribose) moiety at one end and a 5’ phosphate
group at the other. DNA polymerase adds deoxynucleotides to the free
3’ OH end and the 5’ phosphate group end is rolled out as replication
proceeds by rotation of the double helix about its axis. Complement- ary fragments are synthesized on the elongating tail to form a double CL he1 ix. The termination of tail growth may result from a specific
cut with an endonuclease and hence the formation of the daughter circular DNA. The formation of single stranded DNA by the Rolling
Circle technique can also be achieved. This involves the DNA tail
being immediately covered with structural proteins as it is synthe- sized (8).
Transcription
The genetic code of DNA is transcribed by ribonucleic acid poly- merase (RNA polymerase) into a messenger ribonucleic acid (mRNA) which
has a complementary sequence to the DNA. mRNA is generally single
stranded and does not usually have complementary base ratios and only
one of the DNA chains is transcribed by RNA polymerase in the 5’ - 3’
phosphate group direction (8). The nitrogenous basis in mRNA are the
same as those in DNA with the exception that thymidine has been
replaced by uracil. The mRNA then leaves the nucleus to associate with
ribosomes in the cytoplasm. Specific transfer (t) RNAs with a comple-
mentary base sequence to sections of mRNA, assemble amino acids into
peptides in the order determined by the mRNA within the ribosomes (8).
PROBLEMS
Relative to the voluminous 1 iterature on the structure and genet-
ics of prokaryotic DNA, that of the eukaryotic cell is scanty, which
reflects the complexity of eukaryotic DNA. Thus as the inbred mouse is
to the immunologist so is bacterial DNA, especially that of E coli, to the molecular biologist. Both attempting to make that quantum 1 i bet
evolutionary leap by applying what is known to that which is not known,

in that Primate of primates, man. Thus it is inconceivable that DNA
the architect of evolution, has largely been ignored as regards its
own evolution from the prokaryot ic ccl 1 to the eukaryot ic ccl 1 .
The polymer of DNA, regardless of its source, is never studied in
its entirety, only small sections are examined as a salt extract. It is devoid of its associated proteins, which in the eukaryotic cell are responsible for the packing of DNA into chromosomes, and from its
intimate connections with the nuclear membranes. Such DNA is examined by X-ray diffraction whose diffraction patterns may be more imaginary
than they are real. This may account for the various reported struct-
ures or configurations of DNA (2, 3, 4, 5) and its associated proteins
(6, 7, 9). Alternatively the various structures proposed for DNA may
be real in that they represent the source of the original starting
material and the methods used in the extraction of this biological mole- cule. Thus any hypothesis on the structure of DNA must be able to in-
corporate most, if not all, other reported structures. In addition the following must also be taken into consideration.
I . What happens to both ends of the DNA in vivo?
II. Is the structure of DNA, the right handed c1 he1 ix, the result of removing its associated proteins?
III. Does DNA in ZJ~VO contain both right handed and left handed a
he1 ixes, or is this an in vitro effect resulting from the removal of its associated proteins?
IV. Does 1 I and I I I apply to the various forms of DNA, A, B, C, D, and 21
V. Can both DNA chains be transcribed at any time?
VI. The unwinding of DNA in replication and RNA in transcription.
Other problems which have not been referred to will become apparent
as the alternate structure for DNA unfolds as opposed to unwinding.
HYPOTHESIS
It is proposed that the evolution of the structural configuration
of DNA, as opposed to its phenotypic evolutionary expression, involved
two terminal pathways. One of these pathways involves the formation
of single stranded circular DNA from its linear configuration. Similar-
ly, double stranded DNA may have been derived from two complementary
1 inear or circular forms of DNA. The above DNA configurations are con-
sidered to have been retained by the prokaryotic cells. Eukaryot ic DNA
is considered to have evolved from a complex form of DNA, referred to
as Mobius circular DNA. However, before we continue further, let us consider what a Mobius circle is.
291

Mobius Circle
If one takes a suitable strip of paper and joins the ends to form a
circle, it will be found that it is only possible to travel on one side
of the strip (outer or inner surface) in the same direction. To change
from the inner to the outer surface, or vice versa, one must move
through 90° to the original path. Thus the inner and outer surfaces are
not continuous. However, if we now join the strip by first turning one
end in towards oneself (180~) both the inner and outer surfaces become
con t i nuous , a Mobius strip. If this Mobius strip or circle is cut along
its midline in an attempt to form two circles, only one “circle” will
result, twice as large as the first, in a double right handed c1 helical
configuration (Figure 3). However the natural symmetry of this strip is
to form a double loop at one end. This double loop forms a single left
handed helix with the rest of the strip lying side-by-side (Figure 4).
If the initial turn in the strip is made in the opposite direction, a
double left handed helix will be observed and the double loop will become
a right hand he1 ix (Figure 5).
EVOLUTION OF MOB IUS DNA
A single strand of DNA, with its associated proteins, has opposite
terminal ends, a 3’OH group and a 5’ phosphate group. Obviously these
ends may join by a 5’ - 3’ phosphate linkage forming circular DNA. How-
ever, a Mobius circle will be formed if one end of the DNA molecule is
turned out from the plane of this page towards the reader (180’). This
will result in the 3’OH and 5’ phosphate groups being diagonally opposite
each other (5’/3’), but they may form a conventional phosphate linkage
(5’ - 3’) since the phosphate backbone is known to undergo alternate con-
format ional changes (2). Thus, Mobius DNA has a continuous phosphate
backbone and a discontinuous base sequence. That is, at the point of
twist in the DNA molecule the bases will be facing in opposite direct-
ions. This twisting may provide an identification point for DNA and RNA
polymerase, with start and stop defined by the bases being in the opposite
direction (Figure 6). In the case of DNA polymerase, complementary
bases will be laid down until the terminal 5’OH end is reached where
the original bases terminate and the phosphate backbone starts (Figure
7) * If the complementary strand of DNA separates, this too will be in
a Mobius configuration and obviously coding for different proteins then the original template. This may have occurred in our evolutionary
past giving rise occasionally to DNA with a viable phenotypic ex-
pression.
To produce a copy of the original base sequence, the template and
complementary strand of DNA, at the points of twist, may form normal
5’ - 3’, as opposed to 5’/3’ phosphate 1 inkages. This will result in one large circle of non-Mobius DNA. This circular form of DNA may then replicate as a Rolling Circle, the complementary strands being spliced to form a single Mobius DNA. Alternatively, the circle may base pair to form a double strand of linear DNA, which may replicate as previously
discussed, and continue its existence in the double linear form. Similarly, the double linear strand of DNA may form a circle with a nick in one strand to prevent supercoil ing, and rep1 icate as such. Alter- natively in the absence of a nick, a supercoiled structure may result (Figure 8).
292
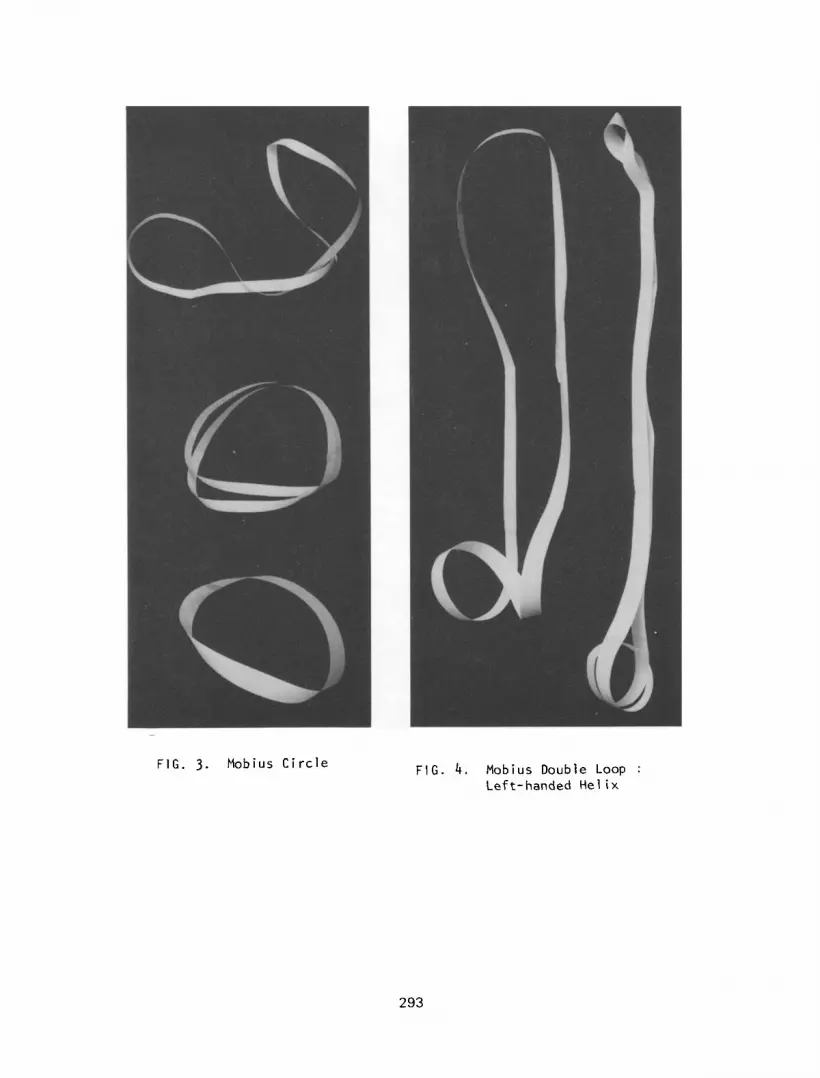
FIG. 3. Mobius Circle FIG. 4. Mobius Double Loop :
Left-handed Helix
293
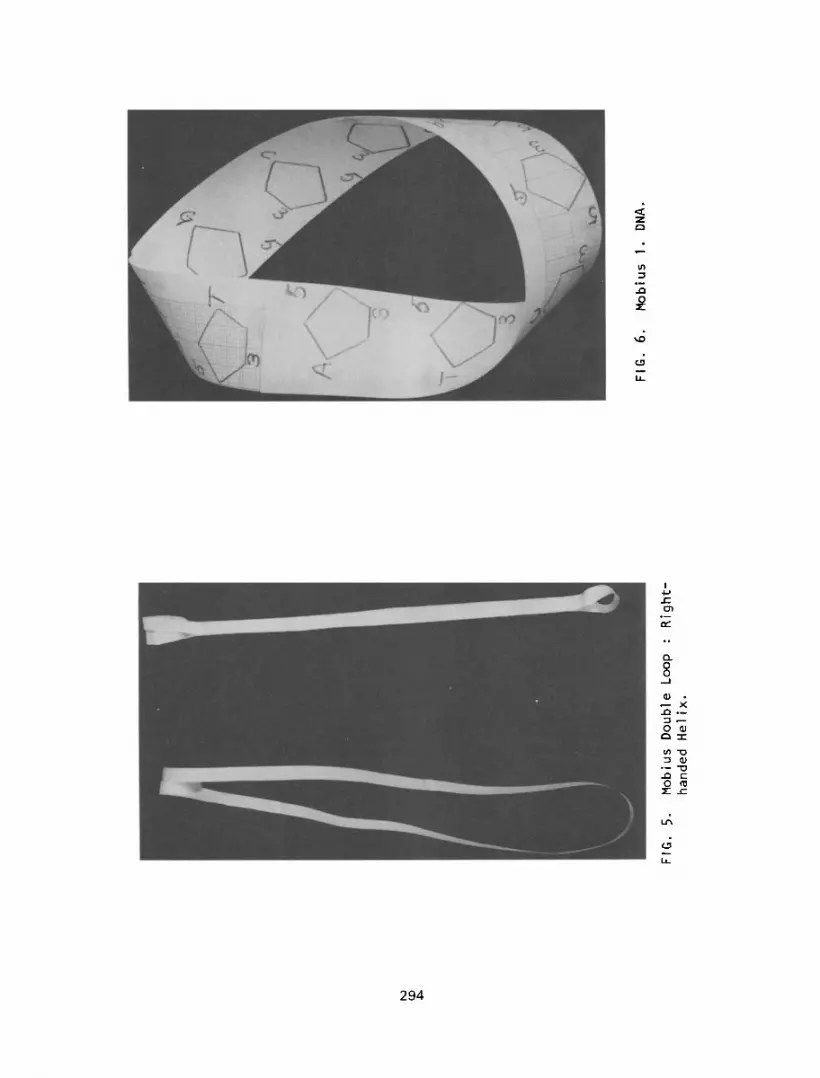
FIG
. 5.
M
obiu
s D
oubl
e Lo
op
: R
ight
-
hand
ed
Helix
.
FIG
. 6.
M
obiu
s 1.
DN
A.

Eukaryot ic DNA
During the replication of Mobius DNA it may be possible for the two
complementary strands to form one continuous Mobius by two 5’/3’ phos-
phate linkages. The formation of double linear of circular DNA from this Mobius may occur as previously described. However , the properties of this Mobius DNA (Mobius 2) are different from that of the original
Mobius (Mobius l), in that the inner and outer surfaces are not contin- uous. In addition, there are two points of change from bases on one
edge to a phosphate backbone, that is, two start and stop points for DNA and/or RNA polymerase. Thus, for example, DNA replication may start
in one of two positions or simultaneously, This would result in two sections of DNA being synthesized and would require joining (splicing)
together. This may explain the general observation that some DNA’s
replicate in short sections and then become spliced together (8). It is not difficult to visualize a number of these Mobius 2 loops joined
together via 2 x 5’/3’ phosphate linkages to form possibly one of the
first primitive eukaryotic chromosomes (-Figure 9) that may have under-
gone further rep1 ication. This may explain the duplication of genes and chromosomes in the evolution of phenotypic expression (10).
A model of the DNA conformat ional state described above may be
visualized if one takes a strip of paper and turns one end out (180’)
and the other in (180~) relative to the plane of this page. If this strip is then cut longitudinally along its midline, two connected loops
will be obtained which must then be joined to each other by two 5’!3’
phosphate 1 inkages.
Another form of Mobius DNA may also be found, Mobius 3 (M-3 DNA). Consider the conformation of Mobius 1 undergoing replication and a 5’/3’
and a 5’ - 3’ phosphate linkage joins the two DNA strands. This will
result in a double d he1 ical configuration which can be formed into a
linear side by side configuration with a double loop at one end (Figure
10). The bases in the linear side by side configuration are paired.
However in the double loop, which is a left-handed helix, the nucleotides
are facing the same direction and therefore cannot pair with each other.
In addition, base pairing is further prevented by the fact that the base
sequence has been displaced by one base (Figure 11). This situation may
explain the isolation of single stranded DNA from eukaryotic cells (11 -
14). We propose that the single stranded DNA double loop represents
unique DNA copy and that the pai red bases represents repetitive copy.
The splicing together of M-3 DNA’s may constitute the chromosomal DNA of
higher eukaryotic ccl 1s.
The 1 inkage of M-3 DNA may be visualized by cutting a Mobius 1 model longitudinally along the midline to form an M-3 model and repeat-
ing the process with the newly formed M-3 model. The resultant M-3 is
linked via the double loop of the original M-3 (Figure 12). The actual
linkage of M-3 DNA’s to each other will be discussed in detail later.
The overall evolutionary pathway for DNA from the prokaryot ic to
the eukaryotic cell is represented in Figure 13.
295

FIG. 7. Replication of Mobius 2. DNA.
.wBIUS Am mw-mBI”* LwA
FIG. 8. Evolution of Mobius DNA (Prokaryotic).
296
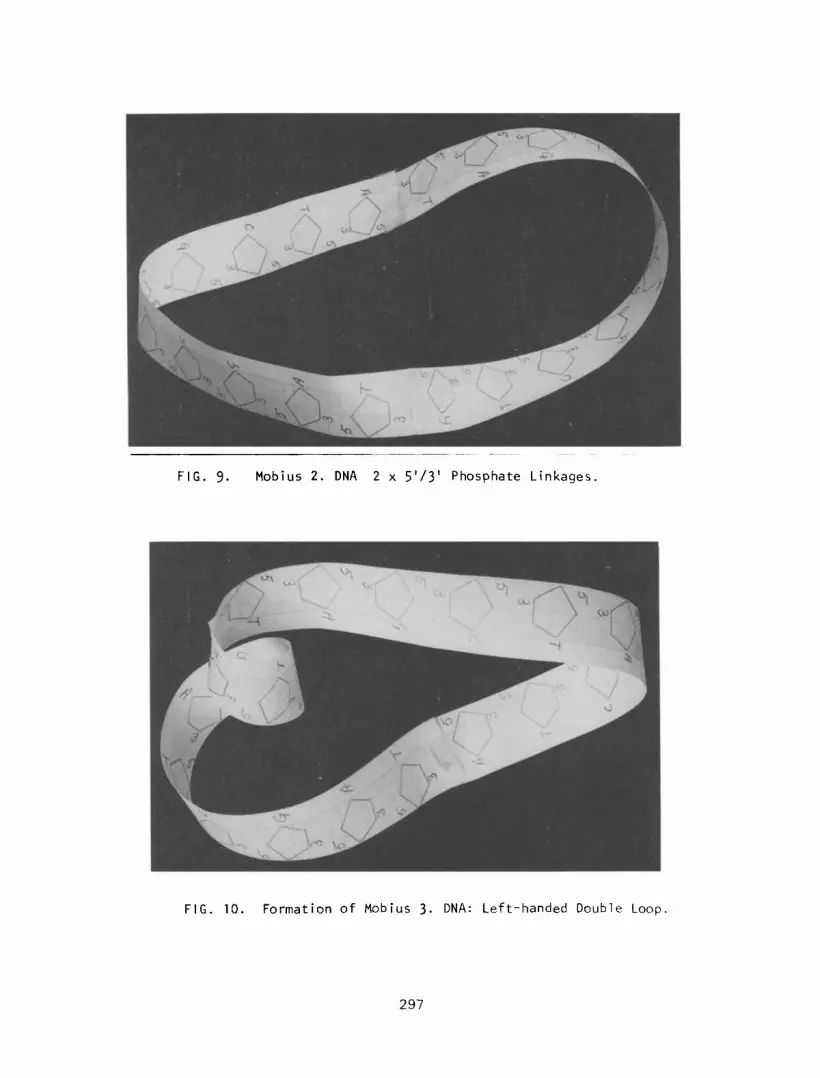
FIG. 9. Mobius 2. DNA 2 x 5’/3’ Phosphate Linkages.
FIG. 10. Formation of Mobius 3. DNA: Left-handed Double Loop.
297

FIG. 11. Mobius 3. DNA :
Side by Side Base
Pairing and Left-
handed Double Loop.
FIG. 12. Model for
Linkage of
Mobius 3. DNA
298

Single linear DNA
I Single circular DNA (5’ - 3’ linkage)
(5’ - 3’ linkage) Ll Mabius.1. (5’/3’ linkage)
DDuble linear DNA (5' - 3' linkage)
I Bkks.3. (5'/3', 51-3')
FIG. 13. Evolution of DNA from the Prokaryotic to the Eukaryotic state
Nucleosomes
The complex structure of nucleosomes are found predominantly in
eukaryotic ccl 1s. As previously discussed eight histones (2 x H2A, H2B,
H3 and H4) form the core proteins around which the DNA is wound. These
histones may be associated in equivalent groups of 4 with each strand
of duplex DNA and lock together to form the histone core per se (15). The configuration of the histone core has been described as a helical
(16), globular (17), as a solenoid with a hole down the central axis
(18)) he1 ical circular “spring” (19) and now as a wedge shaped cyl inder
(9). To confuse the situation even more, two alternative configurations
for the histone core are proposed. The first takes into consideration
more the globular and locking nature of the histones. The second con-
siders the locking, helical solenoid and wedge shaped cylindrical
arrangement of the histones.
Mobius 1 Core Histones
It is proposed that two Mobius shaped histones may associate and
a similar associated pair may space fill the first pair. A complement- ary structure may also be arranged and the two will associate by a
twist locking arrangement (Figure 14 - 16). The side by side DNA may
then wind around this core in the longitudinal direction.
299

300

FIG. 15. Space Fitting of Histone Pairs Forming Complementary Structures (tetramers).
FIG. 16. Association of Two Tetramer Histone Structures to Form an Octomer.
301

. h
302

FIG. 18. Association of Histone Wedge-shaped Cylinders.
FIG. 19. Association of Wedge-shaped Histone Cylinders.
303

FIG. 20. Association of Histone Wedge-shaped Cylinders Forming
He1 ical Spring-l ike Solenoid
304

FIG. 21. Helical Spring-like Solenoid Showing a
Hole along its Central Axis.
FIG. 22. Association of Mobius 3. DNA’ s Forming
Active and Inactive Gene Regions.
305

Mobius Shell Histones (M-Shell)
If Mobius histones are placed side by side as before (Figure 14)
but this time instead of arranging in pairs, arrange in octomers, the
result is an approximation to a wedge shaped cylinder (Figure 17).
Consider the DNA travel 1 ing from the first set of core histones on the
left to the second set on the right (Figure 17). In order to pack the
DNA into a compact form, to form a chromosome, the core histones
must come into contact with each other. In this situation a helical
spring-like, solenoid core with a hole down the central axis is built
up from wedge shaped cylinders (Figure 18, 19, 20, 21). This structure
may then coil on itself to form the metaphase chromosome with its
associated non-histone proteins.
Mobius Chromosomes
We have proposed that Mobius chromosomes of higher eukarotic cells
consist of a number of Mobius 3 DNA’s (M-3 DNA) in a linear arrangement.
The linkage of these Mobius 3 DNA’s to each other may be via the single
tail loops through two 5’/3’ phosphate linkages of each M-3 DNA thus
retaining the Mobius configuration. This would result in the necessity
of linking two double loops, single stranded DNA, of two M-3 DNA’s.
This may be achieved by partly base pairing of the double loops to form
sections of double stranded DNA, possibly complexed in a structural
framework of non-histone proteins. Similarly the linking of a double
loop and a single tail loop of two M-3 DNA may be achieved by complexes
of histone HT in a structural framework of non-histone proteins (Figure 22). Thus there may be sequences of these various linkages in the DNA
of eukaryotic ccl Is. The importance of these 1 inkages is as fol lows.
The double loop of M-3 DNA, single stranded DNA, is considered as
unique copy and has the potential to be transcribed when linked to a
second M-3 DNA by the tail loop. If the double loops of two M-3 DNA’s
are partly base paired, the loops will not be transcribed. Such regions
on the chromosome may be described as inactive genes. Thus the double-
loop / single loop linkage may be described as active genes and may be
transcribed when the histone HT complexes are removed. The side by side
pairing of repetitive M-3 DNA is considered to be associated with the
histone cores. Thus one M-3 DNA may constitute a nucleosome. Alter-
natively, the repetitive M-3 DNA may be associated with more than one
hi stone core. The proposed structure would explain the replication of
DNA in short segments since each M-3 has one stop/start point as in
Mobius 1 DNA. The short DNA segments may then be spliced together to
form chromosomal DNA per se. In addition the proposed structure also
al lows for the variation .in the number of histone cores and nucleosomes
per gene (21).
DISCUSSION AND CONCLUSIONS
Simple prokaryotic DNA no doubt exists in an c1 he1 ical configur- at ion, be it single, or double in a I inear or circular form. However
this appears to represent the evolution of DNA in prokaryotes and its
phenotvoic expression may lead to prokarvotic speciation. Thus the irst involve the evolution evolution and’ speciation’of eukarybtes must f
of DNA which may retain some of its prokaryot
The structure of purified DNA, from proka is obviously different to that in V~UO since
ic characteristics.
ryotic or eukaryotic cells it is devoid of its
306

associated proteins, which are required for the compacting of DNA,
especially in the eukaryotic cell. It is possible that the removal of
such proteins from eukaryotic DNA may result in the collapse of the
higher conformational state of the DNA to form a double right-handed a
helix by random base pairing. They may be visual ized by the formation
of a duplex in the first conformational state of M-3 DNA. The second
conformational state of M-3 DNA is the formation of the double loop,
which is not base paired, and is in a single left handed helix. This
results in the major portion of M-3 DNA being base paired side by side.
It is important to remember that M-3 DNA has been derived from the
association of 2 M-l DNA’s by a 5’/3’. 5’ - 3’ phosphate linkages,
(5’/3’, 5’/3’ phosphate linkages give rise to possibly a primitive form
of eukaryotic DNA, and 5’ - 3’, 5’ - 3’ phosphate linkages give rise to
circular DNA that may rep1 icate to give rise to M-l DNA). Thus M-3 DNA
incorporates a conformational change in the phosphate backbone (2), a
double a helix (1), a single left hand helix as opposed to a double
(20), single stranded DNA (11 - 14) and a side by side base pairing
(4, 5). Similarly, histone cores around which DNA is wound, the M-shell
incorporates a number of structures proposed by other investigators,
helical (16), solenoidal (18), circular spring (18) and a wedge shaped
cyl inder (9). The globular Mobius histone core may be a primitive
eukaryotic form and possibly be associated with M2 DNA. Thus the change
from the globular Mobius histone core to the M-shell form may represent
the evolution of histones.
The structure of histones and their relationship to DNA has recent-
ly been reviewed (22). It appears that the histones H3 and H4 show only
slight evolutionary variations in structure. Whereas H2A and H2B show
substantial structural evolutionary changes which may have occurred to
allow an optimal degree of interaction between these histones and the
evolving DNA. Similarly structural variations also occur in histone HT
and there appears to be an H5 histone that is structurally similar to HT.
In addition histones appear to be species and tissue specific and cells
undergoing differentiation contain a number of variants of H2A, H2B and
H, histones. These histone variants therefore cannot be completely
explained by the evolution of Mobius core histones or M-shell histones
to provide a framework for DNA. It is possible that this histone
variation, in part, is involved in regulating gene activity.
The association of histones with M-3 DNA has been discussed. We
have considered that one M-3 DNA may constitute a nucleosome. A convent-
ional nucleosome consists of 170 - 250 bases (21) and a gene consist.s of
approximately 1,200 bases (8) which suggests approximately six nucleo-
somes (200 bases) per gene. However it has been proposed that the double
loop of single stranded DNA in the M-3 nucleosome constitutes the unique
copy and this may represent a maximum of 110 bases. The tai 1 of M-3 DNA
with its single loop, repetitive DNA, must represent approximately 140
base pairs associated with the histone core. Thus the structure is
within the limits of the conventional nucleosome, 250 bases (21). This means that approximately 10 - 11 M-3 nucleosomes make up a gene of l,llO-
1,220 bases. It is possible that the number of bases in the double loop of single stranded M-3 DNA may vary. Thus a minimum of 30 bases would result in 40 M-3 nucleosomes per gene and a nucleosome length of 170 bases. The structure proposed above may find objections in that RNA
307

polymerase may be required to “jump” the gap from one double loop of
single strand DNA to the next. It has been suggested however, that mRNA
may be synthesized in short pieces and spliced together (21). Alter-
natively, the single stranded DNA may also code for a linker reqion in
the mRNA. This may allow RNA polymerase to move to the next double
loop of single stranded DNA with mRNA in tow. The mRNA per se would
then be formed by removing the linker sections and spl icing together the
sect ions of mRNA. This may explain why there is usually more mRNA
produced in the nucleus than is actually used in the cytoplasm (21).
Similarly, the double loop of single stranded DNA may represent one gene,
containing approximately 1,200 bases with approximately six histone cores
and associated H, histones. However the histone cores will be in halves,
each half associated with a single strand section of DNA. In this
situation one complete mRNA will be produced.
In both gene models it would be theoretically possible to increase
or decrease the size of the repetitive or unique DNA copy by “slipping”
of the M-3 DNA. Thus an increase in repetitive DNA may be achieved by
increased base pairing and the association of the half histone cores to
form histone cores per se. This would reduce the size of the double
loops of single stranded unique copy DNA. The reverse situation would
result in an increase of unique copy and the dissociation of the histone
cores into halves. This situation may be described as a pulsatile gene
which may have been of evolutionary significance. Such slipping of DNA
molecules has been proposed to explain the random appearance of nucleo-
somes found in DNA preparations (21). The slip situation for M-3 DNA
may be visualized by examination of Figure 11. Whichever gene model is
convenient both offer single strand DNA for the attachment of RNA poly-
merase and a termination point for the synthesis of mRNA, as opposed to
the non-committal phosphate backbone and base pairs in the a he1 ix. In
addition the proposed structures for DNA nucleosomes, including inactive
gene regions, can explain the variable number of bases that constitute
nucleosomes from various tissues (24). Finally neither model may require
kinked DNA to wind around the histone cores as in the a helical model
(23).
The replication of DNA and the way in which the various enzymes are
used appears to be mOre complex than was originally thought. Three DNA polymerases have been identified which obviously must bind single
stranded DNA, as previously discussed, if rep1 ication of DNA is to pro-
teed. The DNA polymerases each have exonuclease activity (removal of
internal bases) in the 3’ - 5’ direction as well as polymerase activity in the 5’ - 3’ direction. The exonuclease activity enables the proof
reading of the polymerase activity. The role of DNA polymerase I and I I I appear to have been defined but that of DNA polymerase I I is more obscure.
The point we wish to make here is that these enzymes may be in the shape of a circle. The active site of the exonuclease would be sterically in
the opposite direction and on the opposite side of the molecule to the
active site of the polymerase. This would enable the enzyme to rotate about its central axis as it inserts bases and proof reads them at the
same time it moves along the DNA strand.
In conclusion the proposed structure for eukaryotic DNA follows an
evolutionary pathway and embraces most of the proposed structures for DNA and its associated proteins and enzymes.
308

References
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
Watson JD, Crick FHC. Molecular structure of nucleic acids. A
structure for deoxyribose nucleic acid. Nature 248: 765, 1974.
Davies DR, Zimmerman S. A new twist for DNA? Nature 283: 11,
1980.
Arnott S. Is DNA really a double helix? Nature 278: 780, 1979.
Rodley GA, Scobie RS, Bates RHT, Lewitt RM. A possible conform-
ation for double-stranded polynucleotides. Proceedings of the
National Academy of Sciences of the United States of America 73:
Y: 2959, 1976.
Sasisekharan V, Pattabiraman N, Gupta G. Some implications of an
alternative structure for DNA. Proceedings of the National Academy
of Sciences of the United States of America 75: 9: 4092, 1978.
Sonnenbichler J. Advances in chromatin research. Naturwissen-
schaften 66: 244, 1979.
Robertson M. Beads of 1 ife. New Scientist, July 1979.
Watson JD. Molecular Biology of the Gene. WH Benjamin Inc. 1978.
Dubochet J, No1 1 M. Nucleosome arcs and he1 ices. Science 202:
280, 1978.
Ohno S. Evolution by Gene Duplication. Springer-Verlag, 1970.
Tapiero H, Leibovitch SA, Shaool D, Monier MN, Hare1 J. Isolation
of single-stranded DNA related to the transcriptional activity of
animal ccl 1s. Nucleic Acids Research 3: 953, 1976.
Hanania N, Shaool D, Poncy C, Tapiero H, Hare1 J. Isolation of
single-stranded transcription sites from human nuclear DNA. Cell
Biology International Reports 1: 309, 1977.
Shaool D, Hanania N, Hare1 J, May E. Relationship between single- stranded DNA isolated from mouse cells transformed by Simian Virus
40 and transcription of cellular and virus genes. Journal of General Virology 43: 571, 1979.
Bjursell G, Gussander E, Lindahl T. Long regions of single- stranded DNA in human ccl Is. Nature 280: 420, 1979.
Staynov DZ, Spencer M, Allan J, Gould HJ. A reversible conform-
ational transition in chromatin. Nature 279: 263, 1979.
Bradbury EM, Crane-Robinson C. Physical and conformational studies of histones and nucleohistones. p. 85 in Histones. (DMP Phil1 ips, ed) Plenum Press, 1971.
Kornberg RD. Structure of chromatin. Annual Review of Biochemis- try 46: 931, 1977.
Felsenfeld G. Chromatin. Nature 271: 115, 1978.
Trifonov E. The he1 ical model of the nucleosome core. Nucleic Acids Research 5: 4: 1371, 1978.
309

20.
21.
22.
23.
Wang AH-J, Quigley GJ, Kolpak FJ, Crawford JL, van Boom JH,
van der Mare1 G, Rich A. Molecular structure of a left-handed double he1 ical DNA fragment at atomic resolution. Nature 282:
680, 1979.
Chambon P. Summary: The molecular biology of the eukaryotic
genome is coming of age. p. 1209 in Proceedings from the Cold
Spring Harbour Symposium, U.S.A., 1978.
Von Holt C, Strickland WN, Brandt WF, Strickland MS. More
histone structures. FEBS Letters 100: 2: 201, 1979.
Crick FHC, Klug A. Kinky helix. Nature 255: 530, 1975.
Acknowl egement
The authors are grateful to the Wenkart Foundation for its
support.
310





























