Embed Size (px)
Citation preview

Macromolecular transport in synapseto nucleus communicationNicolas Panayotis1, Anna Karpova2, Michael R. Kreutz2, and Mike Fainzilber1
1 Department of Biological Chemistry, Weizmann Institute of Science, Rehovot 76100, Israel2 Neuroplasticity Group, Leibniz Institute for Neurobiology, Brenneckestraße 6, Magdeburg 39118, Germany
Review
Local signaling events at synapses or axon terminalsmust be communicated to the nucleus to elicit transcrip-tional responses. The lengths of neuronal processespose a significant challenge for such intracellular com-munication. This challenge is met by mechanisms rang-ing from rapid signals encoded in calcium waves toslower macromolecular signaling complexes carried bymolecular motors. Here we summarize recent findingson macromolecular signaling from the synapse to thenucleus, in comparison to those employed in injurysignaling along axons. A number of common themesemerge, including combinatorial signal encoding bypost-translational mechanisms such as differential phos-phorylation and proteolysis, and conserved roles forimportins in coordinating signaling complexes. Neuronsmay integrate ionic flux with motor-transported signalsas a temporal code for synaptic plasticity signaling.
Communication between synapse and somaNeurons communicate at synaptic contacts by signalingevents, yet their long-term responses to such stimuli re-quire changes in gene expression at the nucleus. In con-trast to many other cell types, the intracellular distancesthat separate synapse from soma in neurons are wellbeyond the effective range of diffusion-dependent mecha-nisms [1–4]. How then are distant stimuli sensed anddecoded by the nucleus? Conversely, how are changes intranscription or translation in the soma conveyed to spe-cific synapses or terminals? A series of events, includingmembrane depolarization and calcium influx, was shownto contribute to information transfer in neurons viaelectrophysiological encoding, while the physical transportof macromolecules was thought to serve secondary roles[5–7]. In contrast, long-distance transport of protein mes-sengers in axons is well established as being essential forneuronal maintenance, survival, and regeneration [8–10]. Can localized activation of macromolecules combinedwith intracellular trafficking also enable communicationbetween the synapse and the soma [11]? Here we reviewrecent advances in the understanding of macromolecularcommunication between the synapse and the nucleus, and
0166-2236/
� 2014 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.tins.2014.12.001
Corresponding author: Fainzilber, M. ([email protected]).Keywords: synaptic plasticity; dendrite; axonal transport; nucleocytoplasmic trans-port; dynein; kinesin; myosin; importin; karyopherin.
108 Trends in Neurosciences, February 2015, Vol. 38, No. 2
discuss how these advances can be integrated with themore established fast coupling mechanisms underlyingsynaptic plasticity.
Nuclear integration of synaptic events by fast couplingmechanismsThe prevailing viewpoint in the literature is that calciumsignals are the major route for communication of synapticactivity to the nucleus [7,12]. According to this model, theactivation of receptor-gated calcium channels in the distaldendrites is propagated by action potentials or calciumwaves to the soma, thereby enabling the local activation ofcalcium-dependent pathways connected to gene transcrip-tion. The fast genomic response to sustained rises innuclear calcium is generally believed to be important forthe induction of immediate early gene expression (IEG)within a few minutes of the stimulus [5,7,13,14]. It hasbeen suggested that IEG transcription can be subdividedinto rapid versus delayed transcription [15]. A distinctivefeature is the near-instantaneous transcription by thestalling of RNA polymerase II directly downstream ofthe transcription start site and it is likely that very rapidtranscription events require fast calcium signaling [15,16].
In the electrochemical coupling model, the activation ofcalcium-permeable ion channels in distal dendrites gen-erates back-propagating action potentials or regenerativecalcium waves that are conveyed along the endoplasmicreticulum (ER) to the soma [12]. Their arrival at thenucleus enables the local activation of calcium-dependentpathways connected to gene transcription. This mode ofsynapse to nucleus communication has been intensivelystudied for excitatory synapses and calcium influx gener-ated by activation of synaptic NMDA receptors, L-typecalcium channels, or intracellular calcium release [17–20]. The resulting action potentials generated at the initialaxon segment may propagate retrogradely in the soma,and spread into dendrites [21]. Propagation of such signalsis influenced by the geometry of the dendritic arbor as wellas spatiotemporal integration of excitatory and inhibitoryinputs in the system and back-propagating action poten-tials were suggested to complement and enhance synapticcalcium influx [22,23].
Many forms of long-term potentiation (LTP) requireNMDA receptor (NMDAR) activation and postsynapticcalcium entry [24]. Interestingly, the calcium transientsgenerated upon synaptic activation of NMDARs are spa-tially restricted in spines, with low diffusion into the

Review Trends in Neurosciences February 2015, Vol. 38, No. 2
dendritic shaft [25,26]. However, although calcium signalsare dampened by uptake and clearance systems, regener-ative wave mechanisms may allow them to cover distancesof hundreds of micrometers in a number of milliseconds,bridging the gap between the synapse and the nucleus.Thus, synaptic activation can mobilize calcium from inter-nal stores like the ER [27–29], which extends from thenucleus into the distal processes in both axons and den-drites [30,31]. Regenerative calcium waves may be initiat-ed by calcium signals arising from voltage-gated ionchannels, NMDARs, or the activation of metabotropicglutamate receptors (mGluRs) [32], and propagated bycalcium-induced calcium release mediated by the activa-tion of inositol-1,4,5-triphosphate (IP3) and ryanodinereceptors (RyRs) located on the ER [33].
Although waves represent a plausible mechanism toconvey calcium signals to the nucleus, the source of nuclearcalcium and the mechanism of its elevation by synapticactivity are still a matter of debate [7,34]. Of note, thenucleus can generate autonomous calcium transients and avery recent study suggests that BK-type calcium-sensitivepotassium channels reside on the nuclear envelope andregulate the coupling between synaptic activity and geneexpression [35]. BK channels are typically plasma mem-brane proteins that contain a large conserved C-terminalextension, which is involved in channel gating by intracel-lular ions or second-messenger systems [36]. Li et al. [35]demonstrated the presence of BK channels on the nuclearenvelope of rodent hippocampal neurons. Pharmacologicalblockade of nuclear BK channels in both intact neuronsand isolated nuclei induced calcium release from the nu-clear envelope and cAMP response element-binding pro-tein (CREB)-dependent transcription [35]. These resultssuggest that local signaling at the nuclear envelope couldlink synaptic activity to nuclear calcium transients.
The rapid mechanisms described above have been aprimary focus for most of the field in recent years. Aconsiderably slower process that depends on the nuclearimport of proteins released from synapses may be of par-ticular importance in coupling local synaptic activity tospecific gene expression programs and potentially moresustained and delayed gene transcription [11,37]. What isthe evidence for such macromolecular signaling in synapseto nucleus communication?
ERK signaling from synapse to nucleus requiresmacromolecular traffickingAlthough diverse signaling molecules have been proposedto physically translocate from synapse to soma [11], thefunctional significance of such transport events has beencontentious. NMDARs play a pivotal role in regulation ofactivity-dependent gene expression and the multi-proteinNMDAR complex is a potentially rich source for long-distance protein messengers [38–40]. The mitogen-activat-ed protein kinase (MAPK) cascade plays an important rolein transducing synaptic signals to the nucleus, and theMAPK extracellular signal-regulated kinase (ERK) regu-lates gene transcription in learning and memory [41]. ERKactivation occurs both at the vicinity of the stimulatedsynapse [4,42] and at the nucleus [6]. Strikingly, two veryrecent studies have now combined advanced imaging with
biochemical approaches to provide convincing evidence ofthe need for transport of ERK signaling complexes inspecific paradigms of synapse to nucleus communication[43,44].
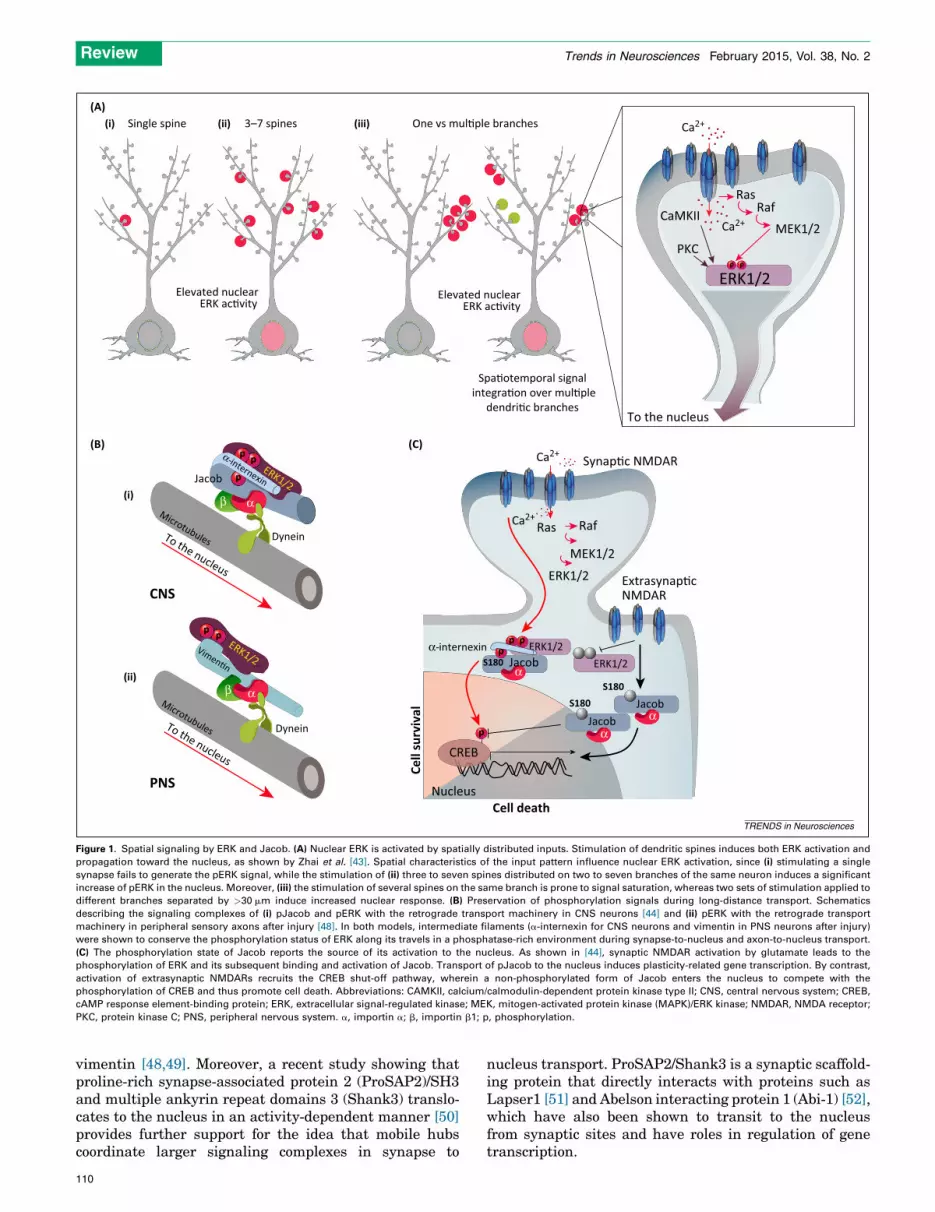
Zhai et al. [43] used a fluorescent ERK activity reporterto examine spatial parameters of signaling under differentparameters of LTP. They evaluated the effect of inducingLTP in single versus multiple dendritic spines of rat CA1hippocampal pyramidal neurons on the propagation ofERK signaling to the nucleus. Stimulation of three or morespines generated a signal that propagated to the nucleus, ifthe stimulated spines were located in at least two distinctdendritic branches (Figure 1A). Moreover, Zhai et al. fur-ther showed that this mechanism allows both spatialintegration of signals from multiple branches and temporalsummation of spatially distinct signals separated by inter-vals of 30 min. The onset of the nuclear response wasdelayed by tens of minutes, in correlation with the distanceof the stimulation, suggesting that propagation was via arelatively slow mechanism [43].
The effective diffusion range of ERK has been estimatedto be approximately 30 mm in hippocampal dendrites [2]and, moreover, the signaling range might be additionallyconstrained by phosphatase activity in the cytoplasm[3]. How then might a phosphorylated signaling moleculesuch as ERK traverse distances of a few hundred micronsin a phosphatase-rich environment from synapse to nucle-us while maintaining its signaling capacity over periods oftens of minutes or more? Karpova et al. [44] characterizedsuch a mechanism in the course of a study aimed atunderstanding how the nucleus can distinguish betweensignaling of synaptic versus extrasynaptic NMDARs. Syn-aptic and extrasynaptic NMDARs have distinct roles insynaptic plasticity, transcription, and cell death[45,46]. The transcriptional response to synaptic NMDARactivation is biased toward cell survival and plasticitygenes, whereas extrasynaptic NMDAR signaling primarilyinduces expression of transcripts involved in cell deathpathways [44,47]. ERK activity was found to be requiredfor nuclear translocation of the protein Jacob after specificactivation of synaptic NMDARs. Although activation ofextrasynaptic NMDARs also induced accumulation of Ja-cob in the nucleus, ERK activity was not required in thelatter case. A series of biochemical, proteomic, and cellbiological analyses then showed that Jacob is an ERKsubstrate, and that synaptic, but not extrasynaptic,NMDAR activation leads to the phosphorylation of Ser-ine180 in Jacob by ERK. Phosphorylated Jacob was pro-tected from dephosphorylation en route to the nucleus byassociation with proteolytic cleavage fragments of theintermediate filament a-internexin, together with ERK(Figure 1B). The phosphorylation state of Jacob uponarrival in the nucleus then determines the transcriptionalresponse and physiological outcome (Figure 1C), and it istempting to speculate that Jacob operates as a mobile hubthat docks NMDAR-derived signalosomes to CREB andpotentially other nuclear target sites [44]. This transportmechanism is strikingly analogous to that previously de-scribed for protected transport of phosphorylated ERKfrom injured peripheral axons to the soma in associationwith proteolytic fragments of the intermediate filament
109
p pVimen�n
ERK1/2
p p
p
Dynein
Microtubules
Jacob
α-internexin
To the nucleus
MicrotubulesTo the nucleus
CNS
PNS
β α
Ca2+
To the nucleus
RasRaf
MEK1/2CaMKII
PKC
Elevated nuclearERK ac�vity
β α
Spa�otemporal signal integra�on over mul�ple
dendri�c branches
(A)
(B) (C)
ERK1/2p p
Extrasynap�cNMDAR
Nucleus
CREBp
Cell death
Cell
surv
ival
Synap�c NMDAR
ERK1/2JacobS180
p pp
α
α-internexinERK1/2
JacobS180
αJacobS180
α
Ras Raf
MEK1/2
ERK1/2
Single spine(i)
(i)
(ii)
(ii) (iii) 3–7 spines One vs mul�ple branches
Elevated nuclearERK ac�vity
ERK1/2
Dynein
Ca2+
Ca2+
Ca2+
TRENDS in Neurosciences
Figure 1. Spatial signaling by ERK and Jacob. (A) Nuclear ERK is activated by spatially distributed inputs. Stimulation of dendritic spines induces both ERK activation and
propagation toward the nucleus, as shown by Zhai et al. [43]. Spatial characteristics of the input pattern influence nuclear ERK activation, since (i) stimulating a single
synapse fails to generate the pERK signal, while the stimulation of (ii) three to seven spines distributed on two to seven branches of the same neuron induces a significant
increase of pERK in the nucleus. Moreover, (iii) the stimulation of several spines on the same branch is prone to signal saturation, whereas two sets of stimulation applied to
different branches separated by >30 mm induce increased nuclear response. (B) Preservation of phosphorylation signals during long-distance transport. Schematics
describing the signaling complexes of (i) pJacob and pERK with the retrograde transport machinery in CNS neurons [44] and (ii) pERK with the retrograde transport
machinery in peripheral sensory axons after injury [48]. In both models, intermediate filaments (a-internexin for CNS neurons and vimentin in PNS neurons after injury)
were shown to conserve the phosphorylation status of ERK along its travels in a phosphatase-rich environment during synapse-to-nucleus and axon-to-nucleus transport.
(C) The phosphorylation state of Jacob reports the source of its activation to the nucleus. As shown in [44], synaptic NMDAR activation by glutamate leads to the
phosphorylation of ERK and its subsequent binding and activation of Jacob. Transport of pJacob to the nucleus induces plasticity-related gene transcription. By contrast,
activation of extrasynaptic NMDARs recruits the CREB shut-off pathway, wherein a non-phosphorylated form of Jacob enters the nucleus to compete with the
phosphorylation of CREB and thus promote cell death. Abbreviations: CAMKII, calcium/calmodulin-dependent protein kinase type II; CNS, central nervous system; CREB,
cAMP response element-binding protein; ERK, extracellular signal-regulated kinase; MEK, mitogen-activated protein kinase (MAPK)/ERK kinase; NMDAR, NMDA receptor;
PKC, protein kinase C; PNS, peripheral nervous system. a, importin a; b, importin b1; p, phosphorylation.
Review Trends in Neurosciences February 2015, Vol. 38, No. 2
vimentin [48,49]. Moreover, a recent study showing thatproline-rich synapse-associated protein 2 (ProSAP2)/SH3and multiple ankyrin repeat domains 3 (Shank3) translo-cates to the nucleus in an activity-dependent manner [50]provides further support for the idea that mobile hubscoordinate larger signaling complexes in synapse to
110
nucleus transport. ProSAP2/Shank3 is a synaptic scaffold-ing protein that directly interacts with proteins such asLapser1 [51] and Abelson interacting protein 1 (Abi-1) [52],which have also been shown to transit to the nucleusfrom synaptic sites and have roles in regulation of genetranscription.

Review Trends in Neurosciences February 2015, Vol. 38, No. 2
The findings outlined above reveal how transport ofERK-containing signaling complexes allows the neuronto distinguish between localized versus distributed synap-tic stimulation [43], or between synaptic versus extrasy-naptic activation of the same receptor [44]. In addition toERK and Jacob, a burgeoning list of signaling moleculeshas been implicated in synapse to nucleus communication[11,37,53]. Transcriptional targets have been identified forsome of these macromolecular messengers, for exampleArc/Arg3.1 and Bdnf for Jacob [44,54], c-fos, Arc, andZif268 for CREB-regulated transcription coactivator 1(CRTC1) [55], and Nap1, JunD, and HLA-DOB for nuclearfactor-kB (NF-kB) [56]. Many of these candidate proteinmessengers, including axin interactor, dorsalization asso-ciated-1D (AIDA1D), CREB2, NF-kB, and Jacob itself,contain nuclear localization signals (NLS) that can berecognized by classical nuclear import factors from theimportin a family [57]. Moreover, ERK itself can beimported to the nucleus via phosphorylation of a shortthree-amino acid motif that interacts with importin 7[58]. These findings have motivated closer examinationof possible roles of nuclear import factors in the cyto-plasmic transport of protein messengers.
Nuclear targeting of synaptic signalsEarly suggestions that nuclear import mechanisms mightbe implicated in long-distance retrograde signaling werebased on the hypothesis of shared regulatory mechanismsfor both neuronal plasticity and neuronal injury responses[59]. Direct evidence for long-distance transport of impor-tin a with the dynein retrograde motor was first obtainedin injured sensory axons [60] and then also shown inmodels of long-term synaptic plasticity [61]. It should benoted that at least six importin a genes are expressed inany given mammal, and multiple importin a gene productsand isoforms are expressed in different cells[57,62,63]. Hence, for the purpose of clarity in this review,we will use the generic term importin a throughout, with-out specifying which individual isoforms are implicated ineach case.
Different modes of regulation of importin complex for-mation have been described in axonal injury versus syn-aptic signaling. In the axon injury model, a series of studieshave demonstrated that calcium-dependent local transla-tion of importin b1 is a critical trigger for retrogradecomplex formation (Figure 2A) [60,64,65]. In contrast,studies of the postsynaptic compartment demonstratedan association of importin a with the cytoplasmic NLSof the NR1-1a subunit of the NMDAR [66]. Activation ofNMDARs in cultured hippocampal neurons or triggering oflate-phase CA3–CA1 LTP in hippocampal slices releasesimportin a from NMDARs. Interestingly, this release is notobserved for stimuli that trigger early-phase LTP[66]. Thus, activity regulation and phosphorylation ofNMDARs can free importin a for cargo binding and trans-port in specific paradigms of synaptic plasticity(Figure 2B).
In addition to the importins, NLS bearing cargo mole-cules are also activated and regulated by translational andpost-translational mechanisms. Transcription factorsmust access the nucleus to fulfill their primary roles in
cells, and therefore have importin-binding capacity. Forexample, the RelA/p65 subunit of NF-kB is recruited toimportins upon phosphorylation-mediated release of itsinhibitor IkBa, following synaptic activation of NMDARs(Figure 2B) [67]. Intracellular transport of NF-kB canoccur by either dynein-dependent transport or facilitateddiffusion [68,69], and studies in different cell types haverevealed that dynein-dependent transport is critical main-ly for the longer distances of transport required in neurons[70,71]. Moreover, the NLS of RelA/p65 was shown to beessential for NF-kB association with the dynein motorcomplex after synaptic stimulation of hippocampal neu-rons [71].
Other transcription factors have been reported to trans-locate over distance in neuronal processes under diversephysiological paradigms. Early observations on the phos-phorylation dynamics of signal transducer and activator oftranscription 3 (STAT3) in axon versus soma led to thesuggestion that it might act as a retrograde signaling factor[72]. A recent proteomic array approach revealed a batteryof transcription factors associated with dynein in injuredaxons, one of which was STAT3 [73]. Activated STAT3 wasthen shown to arise from local translation and phosphor-ylation in the injured axon, and its transport with dyneinand importins provides a retrograde injury signal to thesoma in both sensory and hippocampal neurons [73–75]. In-terestingly, STAT3 signaling was recently implicated inlong-term depression (LTD), although nuclear transloca-tion was not required for this function [76]. Indeed, STAT3may exert local cytoplasmic effects on the cytoskeleton thatare distinct from its transcriptional activity [77], thus itmay have multiple roles in synapse to nucleus signaling atdifferent subcellular junctions.
Finally, the transcription factor CREB is one of the moststudied components in neuronal plasticity pathways, and awealth of studies have described how LTP-inducing stimulitrigger CREB phosphorylation and transcriptional activa-tion [6,78]. Electrochemical mechanisms are thought to bethe primary route for conveying synaptic events to CREB-mediated transcription [7], and although CREB itself hasbeen reported in both embryonic dendrites [79] and matureperipheral axons [80], evidence of a function for dendriticallytransported CREB in neuronal plasticity is unavailable todate. However, recent studies have revealed functionallyimportant trafficking of CREB modulators and effectorsfrom the synapse. The transcription repressor CREB2was reported to translocate from synapse to nucleus underexperimental paradigms of LTD, but not LTP [81]. Animportin a is required for CREB2 transport from distaldendrites to the soma in both rodent hippocampal andAplysia sensorimotor synapses [81]. Interestingly, testingof Jacob under similar NMDAR stimulation paradigms inhippocampal neurons revealed importin-dependent nuclearaccumulation under LTP, but not LTD [82]. CRTC1 (alsoknown as TORC) is another CREB pathway modulatorrecently reported to translocate from synapse to nucleus[55]. CRTC1 binds CREB to promote transcription, andsynaptic activity regulates its transfer from synapse tonucleus in hippocampal neurons. Activity-dependentCRTC1 nuclear translocation is required for the completetranscriptional response of specific CRE-containing genes,
111

Axon to nucleus(A)
(B)
Nucleus
Microtubule
Imp-αDynein
+
++
+ –
AAAAAARanBP1
AAAAAA
(i) (ii) (iii)Lesion
CASα
Signalingcargo
NLS β
Ran
Ca2+
Ca2+Ca2+
AAAAAA
RanBP1AAAAAA
Imp-β1
Imp-β1
αα
Synapse to nucleusPKC
CAMKIIp
p
+
α
Jacob
ERK
p50p65
p
IκBα
H�HAP1
Dynein
Impor�ns
F-ac�n
CREB2α
α
α
RanBP1
Microtubule
TRENDS in Neurosciences
Figure 2. Importin-dependent transport in neuronal processes. (A) Importin-dependent transport of signals from axon to nucleus after peripheral nerve injury. Under
normal conditions (i) an importin a is found in axons in a complex with CAS and the retrograde molecular motor dynein, while importin b1 and RanBP1 are present
predominantly as mRNA. Following lesion (ii), both RanBP1 and importin b1 are translated locally in a calcium-dependent manner. RanBP1 and other cofactors (not shown)
free importin a for association with importin b1, thus generating high affinity binding sites for NLS-bearing signaling proteins. (iii) The complex is then retrogradely
transported toward the nucleus by virtue of the interaction of importin a with dynein [10]. (B) Importins are implicated in activity-dependent synapse to nucleus transport in
CNS neurons. Synaptic activity and NMDAR activation trigger calcium entry and the subsequent activation of protein kinases such as PKC and CAMKII. Jeffrey et al. [66]
demonstrated that an importin a can be anchored at the synapse under basal activity via an interaction with certain splice isoforms of the NR1 subunit of the NMDAR.
Phosphorylation of the latter by PKC elicits the release of importin a, which is then available to bind Jacob (itself released from an interaction with caldendrin upon synaptic
calcium elevation) for retrograde transport to the soma [44]. Synaptic activation of NMDARs also leads to phosphorylation of the NF-kB inhibitor IkBa by CAMKII, thereby
unmasking the NF-kB p50/RelA(p65) NLS for importin a binding and subsequent transport [67]. Abbreviations: CAMKII, calcium/calmodulin-dependent protein kinase type
II; CAS, Crk-associated substrate; CREB2, cAMP response element-binding protein 2; ERK, extracellular signal-regulated kinase; HAP1, huntingtin-associated protein 1; Htt,
huntingtin; NF-kB, nuclear factor-kB; NLS, nuclear localization signal; NMDAR, NMDA receptor; PKC, protein kinase C; RanBP1, Ran-specific binding protein 1. a, importin a;
b, importin b1; IkBa, NF-kB inhibitor alpha.
Review Trends in Neurosciences February 2015, Vol. 38, No. 2
and localized glutamate stimulation reveals that CRTC1exit is specific to the stimulated dendrite [55]. These findingssuggest that physical translocation of CREB repressors ormodulators may provide spatial or quantitative informationto the nucleus on the stimulated synapses.
Multiple regulatory and transport pathways fromsynapse to nucleusThe NMDAR complex, as outlined above, fulfills centralroles in synaptic signaling and neuronal plasticity. A di-verse array of molecules with dual synaptic and nuclear
112
functions have been shown to associate with the complex(Figure 3), however, the mechanisms of activation or re-lease of these long-distance modulators are, as yet, poorlyunderstood. It will be critical to determine the completerange of mechanisms that enable local NMDAR signalingto induce divergent nuclear responses. One interestingoption of localized encoding for long-distance signaling isthe utilization of local protein synthesis, as has beendescribed for axonal injury responses [10]. Indeed, trans-lational control of dendritic mRNAs is required for variousforms of synaptic plasticity [83,84], and synaptic protein

+
Nucleus F-ac�n
+
+
+
Abi-1
Dynein
α
-
NMDAR ac�va�on LTP LTD
Microtubules
α
Ca2+ Ca2+Ca2+
AIDA1
Jacob
ERK Calcineurin
CRTC1 CREB2
Abi-1
β
H�
RelA/p65
1
2
3 4
5
6
7Synap�c
cargo
Synap�ccargo
Myosin
α
α
TRENDS in Neurosciences
Figure 3. Long-distance transport from synapse to nucleus. (1) Dynamic
microtubules can transiently invade a subset of spine synapses following
NMDAR activation in a strictly Ca2+-dependent manner. (2) Neuronal importins
can associate with several synapto-nuclear messengers at synaptic sites but it is
currently unclear how this interaction might be regulated. (3,4) Signaling
downstream of NMDAR that eventually elicits nuclear transport might also
encode that type of NMDAR activation. (3) Ca2+-dependent (Jacob and CRTC1)
and independent (AIDA1, blue) dissociation of protein messengers from NMDAR
has been reported and local phosphorylation via ERK in the case of Jacob or
dephosphorylation via calcineurin in the case of CRTC1 is required to exit the
synapse. (4) Interestingly, NMDAR-dependent LTP but not LTD induces nuclear
translocation of Jacob, while the reverse occurs for CREB2. (5) For many proteins
binding to a dynein motor, likely via association with an importin a, is a
prerequisite for long-distance transport. Whether this transport requires
microtubule invasion of dendritic spines is presently unclear. (6) Alternatively,
proteins like Abi-1 might leave the synapse utilizing a myosin motor. (7) At present
it is also unknown whether the macromolecular transport complex is assembled in
spines or in the dendritic shaft below the synapse. Another interesting question is
whether Htt might substitute for importin b1 in some cases. It has been reported
that an association with Htt is essential for the dissociation of RelA/p65 from
synaptic sites. Abbreviations: Abi-1, Abelson interacting protein 1; AIDA1, axin
interactor, dorsalization associated 1; CREB2, cAMP response element-binding
protein 2; CRTC1, CREB-regulated transcription coactivator 1; ERK, extracellular
signal-regulated kinase; Htt, huntingtin; LTD, long-term depression; LTP, long-term
potentiation; NMDAR, NMDA receptor.
Review Trends in Neurosciences February 2015, Vol. 38, No. 2
transcripts traffic from nucleus to synapse in translation-ally silenced ribonucleoprotein granules [85,86]. This mo-dality of nucleus-to-synapse communication sets the stagefor local translation of mRNAs that are critical for synapticplasticity, but it is not yet clear if such local translationevents also generate synapse-to-nucleus messengers suchas Jacob [87]. Regardless, local translation can induce long-lasting changes in synapse activity [88], and ribosomalcomplexes are enriched at postsynaptic sites [89]. Localprotein synthesis can be stimulated downstream ofNMDARs, mGluR, and the neurotrophin receptor tyrosinekinase TrkB, supporting the involvement of dendritictranslation of trafficked mRNAs in synaptic plasticity[90–92]. It will be interesting to examine how such mecha-nisms might be integrated at the NMDAR with Ca2+
signals and kinase and phosphatase pathways that mightalso be activated by other membrane receptors (Figure 3).
It is also still unknown whether the transport complexfor nuclear trafficking is assembled at synaptic sites or indendrites (Figure 3). As already outlined above, importin a
can directly associate with certain NMDARs [66]. A directtransit from receptor association to transport complexformation might be possible when dynamic microtubulestransiently invade a subset of spine synapses [93]. Incontrast, certain protein messengers such as Abi-1 needactin-based motors for nuclear transport and their traffick-ing from synapse to nucleus is not necessarily microtubule-based [52,94]. Interestingly, microtubule entry to spinesrequires sustained NMDAR-dependent Ca2+-transients[95], and a connection between microtubule transportand NMDAR complexes might be facilitated by their inde-pendent interactions with neural cell adhesion molecule(NCAM) [96,97]. It will be interesting to test whetherimportin a already associates with a dynein motor foractive retrograde transport along microtubules in dendrit-ic spines, as is the case in axons (Figure 2).
Finally, it will be interesting to find out whether signal-ing endosomes might also play a role in synapse to nucleussignaling, analogous to their well-established role in sur-vival and maintenance signaling in the axonal compart-ment [98,99]. A recent study used microfluidics chambersto investigate dendrite to nucleus signaling in embryoniccortical neurons, demonstrating that dendritic applicationof BDNF elicited a transcriptional response in the soma[100]. Intriguingly the response was dependent on activityof the BDNF receptor TrkB in the soma, consistent withinvolvement of a signaling endosome [100]. BDNF-inducedretrograde transport of TrkB in the dendrites of striatalneurons was reported to require Huntingtin (Htt), theprotein mutated in Huntington’s disease [101]. Interesting-ly, Htt can also function as a molecular adaptor for non-vesicular cargoes [102], and a role for Htt in synapse tonucleus transport has also been proposed (Figure 3). It isnot clear at this stage whether this might reflect a specificbinding adaptor role of Htt in signaling complexes[101,102], or a more general role in energy supply totransport complexes [103]. From all of the above evidence,it is increasingly apparent that multiple mechanisms areimplicated in synapse to nucleus trafficking events.
Concluding remarks: How do neurons integratedifferent modalities of synapse to soma signaling?The recent progress described above raises the pressingquestion of how and to what end would a neuron integrateelectrophysiological and motor-driven modalities of syn-apse to soma signaling? One intriguing possibility is thatthe motor-dependent signals provide a temporal compo-nent for information transfer from synapse to soma. Theaverage velocities of motor-driven transport [104] areorders of magnitude lower than those of ionic waves orelectrophysiological signals [105,106], thus a synapticevent transduced by both mechanisms will generate bi-phasic fast and slow signals that arrive in the soma atdifferent times. There is precedent for such temporalencoding in transcription factor regulation, where fast[calcium/calmodulin-dependent protein kinase type IV
113

Box 1. Outstanding questions
� How is the synaptic origin of protein messengers encoded? Is it by
translational or post-translational mechanisms at the synapse
itself, or by temporal dynamics of signal trafficking?
� How are protein messengers released from the synapse?
� What are the main transporters of protein messengers from the
synapse to the nucleus? Are these importin complexes, kinase
scaffolding molecules, or others?
� Does dendritic trafficking of macromolecular complexes influence
transport of axonal signals and vice versa? How do neurons
integrate pre- and post-synapse communication with the nucleus?
� How are ‘fast’ calcium signals integrated with ‘slow’ motor-
dependent signals in synapse to nucleus communication and
neuronal plasticity?
� What are the functionally important transcriptional targets of
macromolecular synapse-to-nucleus messengers? Are these the
same transcriptional targets as those activated by calcium-based
mechanisms?
� Which aspects of neuronal plasticity definitively require macro-
molecular communication between the synapse and the nucleus?
Does it serve an auxiliary role for other aspects of plasticity?
Review Trends in Neurosciences February 2015, Vol. 38, No. 2
(CaMKIV)-dependent] and slow (Ras- or MAPK-depen-dent) pathways were shown to converge on CREB, withdifferent downstream consequences [107,108].
Even if the ultimate molecular messenger in the soma isshared by both signaling mechanisms, as may be the case forERK, the pulsatile nature of temporal encoding may relayinformation to the nucleus. The specificity of such signalingsystems is dependent on noise levels in the intracellularmilieu, and this might be a significant challenge for decodingpulses of ubiquitous signaling molecules such as ERK[109]. Indeed, computational modeling of the fidelity ofinformation transfer in a system comprising a fast ionicwave signal and slower molecular motor trafficked signalssuggested that multiple slow signals would be required toensurerobustnessof informationtransfer[110].Alternative-ly, frequency encoding is likely to be more robust and toconvey more information than amplitude encoding for in-tracellular signaling, since information can be encoded bothin the amplitude of the oscillations and in their frequency, aswell as the potential for encoding multiple frequencies in acomplex oscillating signal [111]. Both theoretical [112] andexperimental analyses in non-neuronal cells [113–116] haveshown that an oscillating signal can lead to a more consis-tent output than a non-oscillating signal, and frequencyencoded signaling was recently proposed to function in axongrowth and length control in neurons [117,118]. Regardlessof whether the macromolecular signals in synapse to nucle-us communication are amplitude or frequency encoded, itwill be intriguing to determine which aspects of plasticityrequire long-distance protein messengers. New geneticmodels and methods to interrogate systems at the subcellu-lar level will be key to answering these questions (Box 1).
AcknowledgmentsOur research on these topics was supported by the European ResearchCouncil (Grant Agreement # 339495) and the Chaya Professorial Chair inMolecular Neuroscience (M.F.); the French Ministry of Foreign AffairsInternational Volunteers Program (N.P.); the Center for Behavioral BrainSciences Magdeburg (A.K.); and the German Research Foundation, theLeibniz Foundation, the European Union Marie Curie InternationalTraining Network NPlast, and a Deutsch-Israelische Projektforderunggrant (M.R.K.).
114
References1 Neves, S.R. et al. (2008) Cell shape and negative links in regulatory
motifs together control spatial information flow in signaling networks.Cell 133, 666–680
2 Wiegert, J.S. et al. (2007) Diffusion and not active transport underliesand limits ERK1/2 synapse-to-nucleus signaling in hippocampalneurons. J. Biol. Chem. 282, 29621–29633
3 Kholodenko, B.N. et al. (2010) Signalling ballet in space and time. Nat.Rev. Mol. Cell Biol. 11, 414–426
4 Harvey, C.D. et al. (2008) The spread of Ras activity triggered byactivation of a single dendritic spine. Science 321, 136–140
5 Greer, P.L. and Greenberg, M.E. (2008) From synapse to nucleus:calcium-dependent gene transcription in the control of synapsedevelopment and function. Neuron 59, 846–860
6 West, A.E. and Greenberg, M.E. (2011) Neuronal activity-regulatedgene transcription in synapse development and cognitive function.Cold Spring Harb. Perspect. Biol. 3, a005744
7 Bading, H. (2013) Nuclear calcium signalling in the regulation of brainfunction. Nat. Rev. Neurosci. 14, 593–608
8 Howe, C.L. and Mobley, W.C. (2005) Long-distance retrogradeneurotrophic signaling. Curr. Opin. Neurobiol. 15, 40–48
9 Ibanez, C.F. (2007) Message in a bottle: long-range retrogradesignaling in the nervous system. Trends Cell Biol. 17, 519–528
10 Rishal, I. and Fainzilber, M. (2014) Axon-soma communication inneuronal injury. Nat. Rev. Neurosci. 15, 32–42
11 Karpova, A. et al. (2012) Long-distance signaling from synapse tonucleus via protein messengers. Adv. Exp. Med. Biol. 970, 355–376
12 Adams, J.P. and Dudek, S.M. (2005) Late-phase long-termpotentiation: getting to the nucleus. Nat. Rev. Neurosci. 6, 737–743
13 Bading, H. et al. (1993) Regulation of gene expression in hippocampalneurons by distinct calcium signaling pathways. Science 260, 181–186
14 Cohen, S. and Greenberg, M.E. (2008) Communication between thesynapse and the nucleus in neuronal development, plasticity, anddisease. Annu. Rev. Cell Dev. Biol. 24, 183–209
15 Saha, R.N. and Dudek, S.M. (2013) Splitting hares and tortoises: aclassification of neuronal immediate early gene transcription based onpoised RNA polymerase II. Neuroscience 247, 175–181
16 Saha, R.N. et al. (2011) Rapid activity-induced transcription of Arcand other IEGs relies on poised RNA polymerase II. Nat. Neurosci. 14,848–856
17 Adams, J.P. et al. (2009) NMDA receptor-independent control oftranscription factors and gene expression. Neuroreport 20, 1429–1433
18 Zhao, M. et al. (2005) Pattern-dependent role of NMDA receptors inaction potential generation: consequences on extracellular signal-regulated kinase activation. J. Neurosci. 25, 7032–7039
19 Dolmetsch, R.E. et al. (2001) Signaling to the nucleus by an L-typecalcium channel-calmodulin complex through the MAP kinasepathway. Science 294, 333–339
20 Thiagarajan, T.C. et al. (2005) Adaptation to synaptic inactivity inhippocampal neurons. Neuron 47, 725–737
21 Waters, J. et al. (2005) Backpropagating action potentials inneurones: measurement, mechanisms and potential functions.Prog. Biophys. Mol. Biol. 87, 145–170
22 Koester, H.J. and Sakmann, B. (1998) Calcium dynamics in singlespines during coincident pre- and postsynaptic activity depend onrelative timing of back-propagating action potentials andsubthreshold excitatory postsynaptic potentials. Proc. Natl. Acad.Sci. U.S.A. 95, 9596–9601
23 Nakamura, T. et al. (1999) Synergistic release of Ca2+ from IP3-sensitive stores evoked by synaptic activation of mGluRs pairedwith backpropagating action potentials. Neuron 24, 727–737
24 Bliss, T.V. and Collingridge, G.L. (1993) A synaptic model of memory:long-term potentiation in the hippocampus. Nature 361, 31–39
25 Yuste, R. et al. (2000) From form to function: calciumcompartmentalization in dendritic spines. Nat. Neurosci. 3, 653–659
26 Hagenston, A.M. and Bading, H. (2011) Calcium signaling in synapse-to-nucleus communication. Cold Spring Harb. Perspect. Biol. 3,a004564
27 Finch, E.A. and Augustine, G.J. (1998) Local calcium signalling byinositol-1,4,5-trisphosphate in Purkinje cell dendrites. Nature 396,753–756
28 Takechi, H. et al. (1998) A new class of synaptic response involvingcalcium release in dendritic spines. Nature 396, 757–760

Review Trends in Neurosciences February 2015, Vol. 38, No. 2
29 Emptage, N. et al. (1999) Single synaptic events evoke NMDAreceptor-mediated release of calcium from internal stores inhippocampal dendritic spines. Neuron 22, 115–124
30 Merianda, T.T. et al. (2009) A functional equivalent of endoplasmicreticulum and Golgi in axons for secretion of locally synthesizedproteins. Mol. Cell Neurosci. 40, 128–142
31 Berridge, M.J. (1998) Neuronal calcium signaling. Neuron 21, 13–2632 Berridge, M.J. (2009) Inositol trisphosphate and calcium signalling
mechanisms. Biochim. Biophys. Acta 1793, 933–94033 Jaffe, D.B. and Brown, T.H. (1994) Metabotropic glutamate receptor
activation induces calcium waves within hippocampal dendrites. J.Neurophysiol. 72, 471–474
34 Bootman, M.D. et al. (2009) An update on nuclear calcium signalling.J. Cell Sci. 122, 2337–2350
35 Li, B. et al. (2014) Nuclear BK channels regulate gene expression viathe control of nuclear calcium signaling. Nat. Neurosci. 17, 1055–1063
36 Salkoff, L. et al. (2006) High-conductance potassium channels of theSLO family. Nat. Rev. Neurosci. 7, 921–931
37 Jordan, B.A. and Kreutz, M.R. (2009) Nucleocytoplasmic proteinshuttling: the direct route in synapse-to-nucleus signaling. TrendsNeurosci. 32, 392–401
38 Emes, R.D. et al. (2008) Evolutionary expansion and anatomicalspecialization of synapse proteome complexity. Nat. Neurosci. 11,799–806
39 Husi, H. et al. (2000) Proteomic analysis of NMDA receptor-adhesionprotein signaling complexes. Nat. Neurosci. 3, 661–669
40 Trinidad, J.C. et al. (2008) Quantitative analysis of synapticphosphorylation and protein expression. Mol. Cell. Proteomics 7,684–696
41 Carasatorre, M. and Ramirez-Amaya, V. (2013) Network, cellular,and molecular mechanisms underlying long-term memory formation.Curr. Top. Behav. Neurosci. 15, 73–115
42 Hardingham, G.E. et al. (2001) A calcium microdomain near NMDAreceptors: on switch for ERK-dependent synapse-to-nucleuscommunication. Nat. Neurosci. 4, 565–566
43 Zhai, S. et al. (2013) Long-distance integration of nuclear ERKsignaling triggered by activation of a few dendritic spines. Science342, 1107–1111
44 Karpova, A. et al. (2013) Encoding and transducing the synaptic orextrasynaptic origin of NMDA receptor signals to the nucleus. Cell152, 1119–1133
45 Hardingham, G.E. et al. (2002) Extrasynaptic NMDARs opposesynaptic NMDARs by triggering CREB shut-off and cell deathpathways. Nat. Neurosci. 5, 405–414
46 Ivanov, A. et al. (2006) Opposing role of synaptic and extrasynapticNMDA receptors in regulation of the extracellular signal-regulatedkinases (ERK) activity in cultured rat hippocampal neurons. J.Physiol. 572, 789–798
47 Hardingham, G.E. and Bading, H. (2010) Synaptic versusextrasynaptic NMDA receptor signalling: implications forneurodegenerative disorders. Nat. Rev. Neurosci. 11, 682–696
48 Perlson, E. et al. (2005) Vimentin-dependent spatial translocation ofan activated MAP kinase in injured nerve. Neuron 45, 715–726
49 Perlson, E. et al. (2006) Vimentin binding to phosphorylated Erksterically hinders enzymatic dephosphorylation of the kinase. J.Mol. Biol. 364, 938–944
50 Grabrucker, S. et al. (2014) The PSD protein ProSAP2/Shank3displays synapto-nuclear shuttling which is deregulated in aschizophrenia-associated mutation. Exp. Neurol. 253, 126–137
51 Schmeisser, M.J. et al. (2009) Synaptic cross-talk between N-methyl-D-aspartate receptors and LAPSER1-beta-catenin at excitatorysynapses. J. Biol. Chem. 284, 29146–29157
52 Proepper, C. et al. (2007) Abelson interacting protein 1 (Abi-1) isessential for dendrite morphogenesis and synapse formation. EMBOJ. 26, 1397–1409
53 Mosca, T.J. and Schwarz, T.L. (2010) The nuclear import of Frizzled2-C by Importins-beta11 and alpha2 promotes postsynapticdevelopment. Nat. Neurosci. 13, 935–943
54 Dieterich, D.C. et al. (2008) Caldendrin-Jacob: a protein liaison thatcouples NMDA receptor signalling to the nucleus. PLoS Biol. 6, e34
55 Ch’ng, T.H. et al. (2012) Activity-dependent transport of thetranscriptional coactivator CRTC1 from synapse to nucleus. Cell150, 207–221
56 Kaltschmidt, B. et al. (2006) NF-kappaB regulates spatial memoryformation and synaptic plasticity through protein kinase A/CREBsignaling. Mol. Cell. Biol. 26, 2936–2946
57 Perry, R.B. and Fainzilber, M. (2009) Nuclear transport factors inneuronal function. Semin. Cell Dev. Biol. 20, 600–606
58 Chuderland, D. et al. (2008) Identification and characterization of ageneral nuclear translocation signal in signaling proteins. Mol. Cell31, 850–861
59 Ambron, R.T. and Walters, E.T. (1996) Priming events and retrogradeinjury signals. A new perspective on the cellular and molecularbiology of nerve regeneration. Mol. Neurobiol. 13, 61–79
60 Hanz, S. et al. (2003) Axoplasmic importins enable retrograde injurysignaling in lesioned nerve. Neuron 40, 1095–1104
61 Thompson, K.R. et al. (2004) Synapse to nucleus signaling duringlong-term synaptic plasticity: a role for the classical active nuclearimport pathway. Neuron 44, 997–1009
62 Yasuhara, N. et al. (2009) The role of the nuclear transport system incell differentiation. Semin. Cell Dev. Biol. 20, 590–599
63 Marfori, M. et al. (2011) Molecular basis for specificity of nuclearimport and prediction of nuclear localization. Biochim. Biophys. Acta1813, 1562–1577
64 Yudin, D. et al. (2008) Localized regulation of axonal RanGTPasecontrols retrograde injury signaling in peripheral nerve. Neuron 59,241–252
65 Perry, R.B. et al. (2012) Subcellular knockout of importin beta1perturbs axonal retrograde signaling. Neuron 75, 294–305
66 Jeffrey, R.A. et al. (2009) Activity-dependent anchoring of importinalpha at the synapse involves regulated binding to the cytoplasmictail of the NR1-1a subunit of the NMDA receptor. J. Neurosci. 29,15613–15620
67 Meffert, M.K. et al. (2003) NF-kappa B functions in synaptic signalingand behavior. Nat. Neurosci. 6, 1072–1078
68 Shrum, C.K. et al. (2009) Stimulated nuclear translocation of NF-kappaB and shuttling differentially depend on dynein and thedynactin complex. Proc. Natl. Acad. Sci. U.S.A. 106, 2647–2652
69 Wellmann, H. et al. (2001) Retrograde transport of transcriptionfactor NF-kappa B in living neurons. J. Biol. Chem. 276, 11821–11829
70 Mikenberg, I. et al. (2006) TNF-alpha mediated transport of NF-kappaB to the nucleus is independent of the cytoskeleton-basedtransport system in non-neuronal cells. Eur. J. Cell Biol. 85, 529–536
71 Mikenberg, I. et al. (2007) Transcription factor NF-kappaB istransported to the nucleus via cytoplasmic dynein/dynactin motorcomplex in hippocampal neurons. PLoS ONE 2, e589
72 Lee, N. et al. (2004) STAT3 phosphorylation in injured axons beforesensory and motor neuron nuclei: potential role for STAT3 as aretrograde signaling transcription factor. J. Comp. Neurol. 474, 535–545
73 Ben-Yaakov, K. et al. (2012) Axonal transcription factors signalretrogradely in lesioned peripheral nerve. EMBO J. 31, 1350–1363
74 Ohara, R. et al. (2011) Axotomy induces axonogenesis in hippocampalneurons through STAT3. Cell Death Dis. 2, e175
75 Shin, J.E. et al. (2012) Dual leucine zipper kinase is required forretrograde injury signaling and axonal regeneration. Neuron 74,1015–1022
76 Nicolas, C.S. et al. (2012) The Jak/STAT pathway is involved insynaptic plasticity. Neuron 73, 374–390
77 Selvaraj, B.T. et al. (2012) Local axonal function of STAT3 rescuesaxon degeneration in the pmn model of motoneuron disease. J. CellBiol. 199, 437–451
78 Lonze, B.E. and Ginty, D.D. (2002) Function and regulation of CREBfamily transcription factors in the nervous system. Neuron 35, 605–623
79 Crino, P. et al. (1998) Presence and phosphorylation of transcriptionfactors in developing dendrites. Proc. Natl. Acad. Sci. U.S.A. 95, 2313–2318
80 Cox, L.J. et al. (2008) Intra-axonal translation and retrogradetrafficking of CREB promotes neuronal survival. Nat. Cell Biol. 10,149–159
81 Lai, K.O. et al. (2008) Importin-mediated retrograde transport ofCREB2 from distal processes to the nucleus in neurons. Proc. Natl.Acad. Sci. U.S.A. 105, 17175–17180
82 Behnisch, T. et al. (2011) Nuclear translocation of jacob inhippocampal neurons after stimuli inducing long-term potentiationbut not long-term depression. PLoS ONE 6, e17276
115

Review Trends in Neurosciences February 2015, Vol. 38, No. 2
83 Holt, C.E. and Schuman, E.M. (2013) The central dogmadecentralized: new perspectives on RNA function and localtranslation in neurons. Neuron 80, 648–657
84 Swanger, S.A. and Bassell, G.J. (2011) Making and breaking synapsesthrough local mRNA regulation. Curr. Opin. Genet. Dev. 21, 414–421
85 Speese, S.D. et al. (2012) Nuclear envelope budding enables largeribonucleoprotein particle export during synaptic Wnt signaling. Cell149, 832–846
86 Fritzsche, R. et al. (2013) Interactome of two diverse RNA granuleslinks mRNA localization to translational repression in neurons. CellRep. 5, 1749–1762
87 Kindler, S. et al. (2009) Dendritic mRNA targeting of Jacob and N-methyl-D-aspartate-induced nuclear translocation after calpain-mediated proteolysis. J. Biol. Chem. 284, 25431–25440
88 Kindler, S. and Kreienkamp, H.J. (2012) Dendritic mRNA targetingand translation. Adv. Exp. Med. Biol. 970, 285–305
89 Zalfa, F. et al. (2006) mRNPs, polysomes or granules: FMRP inneuronal protein synthesis. Curr. Opin. Neurobiol. 16, 265–269
90 Graber, T.E. et al. (2013) Reactivation of stalled polyribosomesin synaptic plasticity. Proc. Natl. Acad. Sci. U.S.A. 110, 16205–16210
91 Klann, E. and Dever, T.E. (2004) Biochemical mechanisms fortranslational regulation in synaptic plasticity. Nat. Rev. Neurosci.5, 931–942
92 Buffington, S.A. et al. (2014) Translational control in synapticplasticity and cognitive dysfunction. Annu. Rev. Neurosci. 37, 17–38
93 Jaworski, J. et al. (2009) Dynamic microtubules regulate dendriticspine morphology and synaptic plasticity. Neuron 61, 85–100
94 Jordan, B.A. et al. (2007) Activity-dependent AIDA-1 nuclearsignaling regulates nucleolar numbers and protein synthesis inneurons. Nat. Neurosci. 10, 427–435
95 Merriam, E.B. et al. (2013) Synaptic regulation of microtubuledynamics in dendritic spines by calcium, F-actin, and drebrin. J.Neurosci. 33, 16471–16482
96 Sytnyk, V. et al. (2006) NCAM promotes assembly and activity-dependent remodeling of the postsynaptic signaling complex. J.Cell Biol. 174, 1071–1085
97 Perlson, E. et al. (2013) Dynein interacts with the neural celladhesion molecule (NCAM180) to tether dynamic microtubulesand maintain synaptic density in cortical neurons. J. Biol. Chem.288, 27812–27824
98 Cosker, K.E. and Segal, R.A. (2014) Neuronal signaling throughendocytosis. Cold Spring Harb. Perspect. Biol. 6, a020669
99 Bronfman, F.C. et al. (2014) Spatiotemporal intracellular dynamics ofneurotrophin and its receptors. Implications for neurotrophinsignaling and neuronal function. Handb. Exp. Pharmacol. 220, 33–65
116
100 Cohen, M.S. et al. (2011) Neurotrophin-mediated dendrite-to-nucleussignaling revealed by microfluidic compartmentalization of dendrites.Proc. Natl. Acad. Sci. U.S.A. 108, 11246–11251
101 Liot, G. et al. (2013) Mutant Huntingtin alters retrograde transport ofTrkB receptors in striatal dendrites. J. Neurosci. 33, 6298–6309
102 Marcora, E. and Kennedy, M.B. (2010) The Huntington’s diseasemutation impairs Huntingtin’s role in the transport of NF-kappaBfrom the synapse to the nucleus. Hum. Mol. Genet. 19, 4373–4384
103 Zala, D. et al. (2013) Vesicular glycolysis provides on-board energy forfast axonal transport. Cell 152, 479–491
104 Schiavo, G. et al. (2013) Cytoplasmic dynein heavy chain: the servantof many masters. Trends Neurosci. 36, 641–651
105 Munoz-Garcia, J. and Kholodenko, B.N. (2010) Signalling over adistance: gradient patterns and phosphorylation waves withinsingle cells. Biochem. Soc. Trans. 38, 1235–1241
106 Cho, Y. et al. (2013) Injury-induced HDAC5 nuclear export is essentialfor axon regeneration. Cell 155, 894–908
107 Finkbeiner, S. et al. (1997) CREB: a major mediator of neuronalneurotrophin responses. Neuron 19, 1031–1047
108 Wu, G.Y. et al. (2001) Activity-dependent CREB phosphorylation:convergence of a fast, sensitive calmodulin kinase pathway and aslow, less sensitive mitogen-activated protein kinase pathway. Proc.Natl. Acad. Sci. U.S.A. 98, 2808–2813
109 Voliotis, M. et al. (2014) Information transfer by leaky, heterogeneous,protein kinase signaling systems. Proc. Natl. Acad. Sci. U.S.A. 111,E326–E333
110 Kam, N. et al. (2009) Can molecular motors drive distancemeasurements in injured neurons? PLoS Comput. Biol. 5, e1000477
111 Cheong, R. and Levchenko, A. (2010) Oscillatory signaling processes:the how, the why and the where. Curr. Opin. Genet. Dev. 20, 665–669
112 Tostevin, F. et al. (2012) Reliability of frequency and amplitudedecoding in gene regulation. Phys. Rev. Lett. 108, 108104
113 Yissachar, N. et al. (2013) Dynamic response diversity of NFATisoforms in individual living cells. Mol. Cell 49, 322–330
114 Cai, L. et al. (2008) Frequency-modulated nuclear localization burstscoordinate gene regulation. Nature 455, 485–490
115 Albeck, J.G. et al. (2013) Frequency-modulated pulses of ERK activitytransmit quantitative proliferation signals. Mol. Cell 49, 249–261
116 Aoki, K. et al. (2013) Stochastic ERK activation induced by noise andcell-to-cell propagation regulates cell density-dependent proliferation.Mol. Cell 52, 529–540
117 Albus, C.A. et al. (2013) Cell length sensing for neuronal growthcontrol. Trends Cell Biol. 23, 305–310
118 Rishal, I. et al. (2012) A motor driven mechanism for cell lengthsensing. Cell Rep. 1, 608–616





























