Embed Size (px)
Citation preview

B American Society for Mass Spectrometry, 2013DOI: 10.1007/s13361-013-0687-9
J. Am. Soc. Mass Spectrom. (2013)
RESEARCH ARTICLE
LptA Assembles into Rod-Like Oligomers InvolvingDisorder-to-Order Transitions
Carlo Santambrogio,1 Paola Sperandeo,1 Riccardo Villa,2 Frank Sobott,3
Alessandra Polissi,1 Rita Grandori1
1Department of Biotechnology and Biosciences, University of Milano-Bicocca, 20126 Milan, Italy2Department of Biosciences, University of Milan, 20133 Milan, Italy3Department of Chemistry and CFP-CeProMa, University of Antwerp, 2020 Antwerp, Belgium
Abstract. LptA is a periplasmic protein involved in the transport of lipopolysac-charide (LPS) from the inner membrane (IM) to the outer membrane (OM) ofGram-negative bacteria. Growing evidence supports a model in which LptAassembles into oligomers, forming a physical bridge connecting IM and OM. Thiswork investigates assembly and architecture of LptA oligomers. Circular dichroismand “native” electrospray-ionization ion-mobility mass spectrometry (ESI-IM-MS)are employed to test concentration dependence of LptA structural features and toanalyze the morphology of higher-order aggregates. The results show that LptAprogressively assembles into rod-like oligomers without fixed stoichiometry, andgrows by an n + 1 mechanism up to at least the pentamer. The oligomerization
process induces disorder-to-order transitions in the polypeptide chain. Comparison with crystallographic andcomputational data suggests that these conformational changes likely involve short disordered regions at theN- and C-termini of monomeric LptA. The protein response to thermal denaturation displays strongconcentration dependence, indicating that oligomerization increases protein stability. LptA conformationalstability can also be enhanced by in vitro LPS binding. The genesis of these fibrillar structures could berelevant for the correct transport of LPS across the bacterial periplasm.Key words: Gram-negative bacteria, Lipopolysaccharide, Lpt machinery, Induced folding, Electrosprayionization-ion mobility-mass spectrometry, Circular dichroism
Received: 5 April 2013/Revised: 20 May 2013/Accepted: 20 May 2013
Introduction
The cell envelope of Gram-negative bacteria is a complex,multilayered structure, composed of a symmetric cyto-
plasmic (inner) membrane (IM) and an asymmetric outermembrane (OM). These are separated by an aqueouscompartment, the periplasm, which contains a thin layer ofpeptidoglycan [1]. In the OM, lipids are asymmetricallyarranged, so that the inner leaflet is composed of phospholipidsand the outer leaflet contains lipopolysaccharide (LPS) [2].LPS is essential in most Gram-negative bacteria [3] and largely
contributes to the selective permeability properties exhibited bythe OM [4]. Biosynthesis of LPS begins in the cytoplasm and iscompleted at the periplasmic side of the IM [5, 6]. Then, LPS istransported across the periplasm to its final location, the cellsurface, by a transenvelope device composed of seven proteins,LptA-G [7, 8] (Figure 1). At the IM, the LptB2FG complexconstitutes an ABC transporter that provides the energy forLPS transport [9]. LptC is a small bitopic protein [10] thatresides in the IM and interacts with the LptB2FG complex [9]and with the periplasmic protein LptA [11, 12]. LptA is thoughtto transfer LPS to the LptDE complex residing at the OM.Thus, LptA is the periplasmic protein that connects the IM Lptcomponents to the OMLPS translocon (LptDE), which ensuresthe assembly of LPS at the cell surface [13–15].
The crystal structures of LptA and LptC have beensolved. Both proteins present a similar fold, consisting of a“twisted boat” structure, with two β-sheets coming into
Electronic supplementary material The online version of this article(doi:10.1007/s13361-013-0687-9) contains supplementary material, whichis available to authorized users.
Correspondence to: Frank Sobott; e-mail: [email protected],Alessandra Polissi; e-mail: [email protected], Rita Grandori;e-mail: [email protected]

apposition [16, 17] and capable of binding LPS in vitro [17,18]. Interestingly, a similar structure is predicted in the N-terminal periplasmic domain of LptD [13] and in theperiplasmic region of LptF and LptG [8, 19], suggestingthat the assembly of the Lpt machinery is based oninteractions among structurally homologous domains.
The process by which hydrophobic LPS molecules aretransported across two membranes and the interveningperiplasmic space is not fully understood. The current modelpostulates that the Lpt proteins create a transenvelope bridgethat connects IM and OM [13, 19], thus forming acontinuous channel through which LPS is moved towardsthe exterior environment [20]. According to this model, theC-terminus of LptC interacts with the N-terminus of LptAand the C-terminus of LptA interacts with the N-terminalperiplasmic domain of LptD [13]. Several lines of evidencesuggest that the transenvelope complex is not a staticstructure and that its assembly is finely regulated byphysiological and environmental conditions [13, 19]. Inparticular, the length of the oligomeric structure must beadapted, depending on location, cellular conditions, andorganism [13]. As the protein that connects IM and OM,LptA plays a key role in the assembly of the Lpt machinery.The crystal structure of LptA obtained in the presence ofLPS shows a fibrillar arrangement with a head-to-tailorganization of subunits [16], consistent with its in vivorole. A rod-like morphology of the LptA oligomers has alsobeen suggested on the basis of size-exclusion chromatogra-phy-laser light scattering (SEC-LS) [21]. The stoichiometry,
however, and the assembly mechanism of LptA oligomershave not been elucidated yet [13].
Complementary biophysical techniques are employedhere to analyze LptA structural properties as a function ofprotein concentration and LPS presence, gaining newinsights on the mechanism and the intermediates of thisimportant protein oligomerization process.
MethodsProtein Expression and Purification
Cultures of BL21(DE3) carrying pET/LptA-H plasmid andexpressing LptA fused to a C-terminal His6 tag (LptA-SGRVEHHHHHH) [16] were grown overnight in 0.2%glucose M9 minimal medium, supplemented with ampicillin(100 μg/mL). Aliquots were diluted 1:100 in fresh medium andgrown up to mid-logarithmic phase (OD600, 0.6) at 30 °C.Expressionwas induced by the addition of 0.5 mM isopropyl β-D-1-thiogalactopyranoside (IPTG; Sigma-Aldrich, St. Louis,MO, USA) and further incubation for 16–18 h at 18 °C. Cellswere harvested by centrifugation at 4 °C (5000 g, 20 min). Thecell pellet was resuspended in buffer A (50 mM sodiumphosphate, pH 8.0, 300 mM NaCl, 10 mM imidazole, 10%glycerol), followed by 30min incubation at 4 °C, shaking in thepresence of lysozyme (1 mg/mL), DNase (100 μg/mL), 10 mMMgCl2, and 1 mM phenylmethanesulfonylfluoride (PMSF;Sigma-Aldrich, St. Louis, MO, USA). Cells were disrupted bya single cycle of French press (One ShotModel Cell Disrupter;
Figure 1. LPS transport machinery. Schematic representation of the transenvelope complex responsible for LPS translocationfrom IM to OM. The seven Lpt components are labeled by the corresponding letters. The LPS molecule is extracted from the IMby the ABC transporter LptB2CFG and transferred to the LptDE complex at OM by LptA. The number (n) of LptA moleculesrequired to form the transenvelope complex is not yet known
C. Santambrogio et al.: LptA Rod-Like Oligomers

Constant Systems Ltd., Low March, Daventry, UK) at 25,000psi. Unbroken cells and cell debris were removed bycentrifugation at 4 °C (39,000 g, 30 min). The soluble proteinwas purified from the supernatant by nickel-nitrilotriacetic acid(Ni-NTA) affinity columns (Qiagen, Hilden, Germany). Thecolumn was washed with 10 column volumes (CV) of 4%buffer B (50 mM sodium phosphate, pH 8.0, 300 mM NaCl,500 mM imidazole, 10% glycerol) in buffer A. Proteins wereeluted by a stepwise gradient at 10%, 20%, 50%, 70%, and100% buffer B. At each step, 1 CV was passed through thecolumn. Fractions were analyzed by SDS-PAGE on 12.5%polyacrylamide gels [22]. Pooled fractions containing thepurified protein (990 %) were dialyzed against 50 mM sodiumphosphate, pH 8.0, 150 mM NaCl (cellulose membranes,12,000-Da cut-off; Sigma-Aldrich, St. Louis, MO, USA).Protein concentration was determined by a Coomassie (Brad-ford) assay kit (Thermo-Pierce, Rockford, IL, USA), usingbovine serum albumin as a standard. LptA was buffer-exchanged before MS analyses by two cycles on Micro-Bio-Spin P6 chromatography columns (Bio-Rad, Hercules, CA,USA). The molecular mass of the purified protein determinedby ESI-MS corresponds to the processed product, in which the27 N-terminal residues of the signal sequence (M1–A27) havebeen removed (calculated mass 18646.7 Da, experimentalmass under denaturing conditions and harsh parameters18646.3 ± 0.5 Da).
Sequence Analysis
The LptA amino acid sequence, truncated by the first 27residues and including the His6 tag, was subjected to disorderprediction. Algorithms of the PONDR package (Predictors ofNatural Disordered Regions, www.pondr.com) [23, 24] wereemployed. The predictors are feed-forward neural networkstrained on distinct sets of ordered and disordered sequences.The fractional output score is compared with the generallyapplied, empiric threshold of 0.5.
Mass Spectrometry
Nondenaturing ESI-MS and ESI-IM-MS analyses wereperformed on a Synapt G2 instrument (Waters, Milford,MA, USA) equipped with a TriVersa Nanomate ion source(Advion, Ithaca, NY, USA). The samples were injected, afterdesalting, at different protein concentrations (1–8 μM nom-inal monomer concentration) and pH values (3 and 8) in 200mM aqueous ammonium acetate at room temperature.Instrument parameters were carefully optimized to minimizedisruption of noncovalent interactions. The following instru-mental settings were applied: ion spray voltage 1.7 kV,sampling cone voltage 20 V, trap collision energy 3 V (4.5 Vunder harsh conditions), transfer collision energy 0 V, bias40 V (60 V under harsh conditions), backing pressure 4.5mbar, trap pressure 2 × 10–2 mbar, IMS pressure 2.5 mbar.Spectra were averaged over 2 min and acquired by thesoftware MassLynx 4.1 (Waters, Milford, MA, USA).
Circular Dichroism
CD spectra were acquired on a J-815 spectropolarimeter(JASCO Corporation, Tokyo, Japan) equipped with a Peltiersystem for temperature control. Samples were measured in50 mM sodium phosphate, 50 mM NaCl at different proteinconcentrations (1, 3, 4.5, 6, 9, 12, and 25 μM) and pH values(3 and 8). Thermal ramps were performed in the temperaturerange 20–65 °C, with increments of 5 °C at each step. Thesamples were allowed to equilibrate for 2 min at eachtemperature before acquisition. Quartz cuvettes with differ-ent path lengths (1–5 mm) were employed, depending onprotein concentration. The following instrumental settingswere applied: acquisition range 200–260 nm, data pitch 0.1nm, scan speed 20 nm/min, bandwidth 1 nm. Final spectraare the average of two measurements subtracted by theblank.
LPS-Binding Assay
The in vitro LPS binding assay was based on the protocoldescribed previously [18], with a few minor modifications.Briefly, assays (500 μL) were carried out in buffer C (50mM sodium phosphate, pH 8, 50 mM NaCl) containing 25μM of purified LptA and a 10-fold molar excess of purifiedsmooth LPS from E. coli serotype O55:B5 (Sigma-Aldrich,St. Louis, MO, USA). To calculate the molar concentrationof the ligand, its molecular weight was assumed to be10,000 Da.
The reactions were incubated at room temperature for1 h on a rotary shaker to allow for the formation ofLPS-protein complexes. Ni-NTA resin (200 μL, His-Select nickel affinity gel; Sigma-Aldrich, St. Louis, MO,USA), washed in 1 mL of buffer C, was added to thereaction mixtures and incubated for another hour toallow binding of LPS–protein complexes via the His6tag. The reaction mixture was centrifuged at 13,000 g for1 min, and the supernatant was collected. The resin wasthen washed four times with buffer C and the protein–LPS complexes were eluted in two steps, with 500 μL ofbuffer C containing 300 mM imidazole (E1) or 500 mMimidazole (E2), respectively.
SDS-PAGE
To monitor LPS-LptA complex formation, equal volumes (20μL) of the collected chromatografic fractions were analyzed bydenaturing gel electrophoresis. For LPS visualization, sampleswere run on N-tris(hydroxymethyl)methylglycine (tricine)-SDS-PAGE (18% acrylamide) [25] and developed by silverstaining as described previously [26]. LptA, instead, wasanalyzed by SDS-PAGE (12.5% acrylamide) [22] andWesternblot. After SDS-PAGE, proteins were transferred ontoHybond-ECL nitrocellulose membrane (GE-Healthcare, LittleChalfont, Buckinghamshire, UK) and probed by mouse anti-polyhistidine monoclonal antibody (1:3,000) (Sigma-Aldrich,
C. Santambrogio et al.: LptA Rod-Like Oligomers

St. Louis, MO, USA) according to the manufacturer’s pro-cedures. Donkey anti-mouse antibodies conjugated to IRDye800 (LI-COR GmbH, Bad Homburg, Germany) were used at adilution of 1:15,000 for filter development and bands werevisualized by an Odyssey Fc Imaging System (LI-COR GmbH,Bad Homburg, Germany).
Results and DiscussionLptA Contains Unstructured Regions Closeto the N- and C-Termini
The published crystallographic data for monomeric LptAsuggest that the β-rich structure of this protein also containssome degree of conformational disorder (Figure 2a) [16].Several residues in the N- and C-terminal regions are missingin the crystal structures because of diffuse scattering in X-raydiffraction (Figure S1 in Online Resource). In order to predictthe extent of structural disorder in solution, the LptA sequencewas analyzed by the PONDR algorithms for the evaluation ofdisorder propensity [23, 24]. Figure 2c shows the profiles of thefractional disorder score calculated by four predictors. Theresidues with a score above the conventional threshold of 0.5by at least two predictors are classified as unstructured andindicated by thick black bars on the threshold line of the graph.
These residues come to lie in three distinct regions of theLptA sequence: the first 24 amino acids at the N-terminus ofthe mature protein (region 1), an internal group of 11 amino
acids (aa 72–83, region 2), and the last 34 amino acids(region 3). The positions of these predicted disordered tractscorrespond quite closely to the missing residues of thecrystallographic structure (Figure 2a, Figure S1 in OnlineResource), especially in the case of regions 2 and 3. Region1 is not completely unstructured in the crystals, showingfolded elements, such as a short helix and two small β-strands. Altogether, these results strongly hint at thepresence of intrinsically disordered regions in monomericLptA in solution [27].
Conformational Transitions of LptA at VariablepH and Temperature
Protein secondary and tertiary structures were analyzed byfar-UV CD and ESI-MS, respectively, using 1 μM LptAsolutions at variable pH and temperature (Figure 3). Theextent of protein ionization by electrospray is known todepend strongly on the overall structural compactness,leading to lower charge states for folded, globular confor-mations [28, 29]. Under nondenaturing conditions (pH 8,room temperature), the LptA mass spectrum shows a narrowcharge-state distribution, consisting of a few peaks atrelatively low charge states (7+ to 9+) (Figure 3a). Thiskind of peak envelope is typically associated with a highdegree of structural compactness maintained by the proteinduring the ionization process, reflecting a globular fold inthe solution phase [28].
Figure 2. Disordered regions in LptA. (a) Crystal structure of monomeric LptA (PDB code: 2R19, unit a). The three regionspredicted to be disordered are depicted in black. The gap corresponding to missing residues is represented by a dashed line.(b) Crystal structure of oligomeric LptA (PDB code: 2R1A), obtained in the presence of LPS. The regions in black in the centralunit are the same as in (a). (c) Graphical output of disorder prediction. Different PONDR predictors are used: CAN_XT (thin solidline), VLXT (thick solid line), XL1_XT (dashed line), and VL3 (dotted line). Residues with a disorder score higher than the empiricthreshold of 0.5 by at least two predictors are marked by a black bar on the threshold line
C. Santambrogio et al.: LptA Rod-Like Oligomers

At higher m/z values, the spectrum shows peaks corre-sponding to a minor population of dimeric LptA (~10%).This partial association of LptA into dimeric structures, atrelatively low protein concentration, indicates a propensityto oligomerize, in line with previous reports by othermethods [13, 21]. The monomer prevails under the hereemployed experimental conditions, while the protein appearspredominantly dimeric by SEC-LS measurements at similarpH, ionic strength and protein concentration [21]. It is likelythat the dimer is lost to some extent under electrosprayconditions, due to in-source dissociation [30]. Notably theprotein concentration is well below the generally observedthreshold (~100 μM) for unspecific complex formationduring electrospray [31], suggesting that the detected dimersreflect a specific oligomeric state of the protein in thesolution phase.
Upon acidification to pH 3, a new peak envelope appearsin LptA mass spectra (Figure 3b). The lower m/z values ofthis component around the 11+ ion (monomer) indicates lossof compact conformation, implying that the protein un-dergoes denaturation under acidic conditions [28]. Therelative amounts of the two components in the spectrumindicate that a native-like, globular conformation remainspredominant at pH 3.
The influence of pH on protein conformation was alsostudied by far-UV CD, in order to monitor changes insecondary structure [32]. Under non-denaturing conditions(pH 8, 20 °C), the LptA spectrum is characteristic of apredominantly β-sheet protein, with a broad minimum in therange 210–220 nm [32] (Figure 3c). This result is inagreement with the features highlighted by X-ray crystal-lography [16] (Figure 2a). At pH 3, the band at ~215 nm isless intense and the spectrum presents a new globalminimum at lower wavelengths (~203 nm), indicating partialloss of ordered secondary structure [32] (Figure 3c). Theisodichroic point at ~207 nm is typical of β-sheet-to-coiltransitions [32].
Circular dichroism was also employed to monitor LptAdenaturation by heat, as illustrated in Figure 3d. The spectrarecorded during a thermal ramp in the range 20–65 °C showan unfolding transition that reaches a plateau at ~50 °C.Maintenance of the isodichroic point at ~207 nm indicatesan apparent two-state transition involving β-sheet-to-coilconversion. Comparison between panels c and d of Figure 3shows that acid-induced unfolding is not complete at pH 3,in agreement with the results obtained by ESI-MS.
In conclusion, LptA presents a compact, β-rich confor-mation in solution that is prone to protein-protein interac-
Figure 3. LptA structural characterization. (a) and (b) Nano-ESI-MS spectra of 1 μM LptA; (c) and (d) Far-UV CD spectra of 1μM LptA. (a) 200 mM ammonium acetate, pH 8, 20 °C; (b) 200 mM ammonium acetate, pH 3, 20 °C; (c) 50 mM sodiumphosphate (pH 8, solid line and pH 3, dashed line), 50 mM NaCl, 20 °C; (d) 50 mM sodium phosphate, pH 8, 50 mM NaCl, 20–65 °C. The most intense ion of each component in panels (a) and (b) is labeled according to quaternary structure (M, monomer;D, dimer) and charge state. Arrows in panel (d) indicate the direction of spectral changes as the temperature is increased. Thevertical line across panels (c) and (d) indicates the position of the isodichroic point
C. Santambrogio et al.: LptA Rod-Like Oligomers

tions, and undergoes apparent two-state unfolding transitionsinduced by acids and heat.
Morphology and Stoichiometry of LptA Oligomers
The propensity of LptA to acquire quaternary structure wasfurther investigated by native ESI-MS [33]. Samples atdifferent protein concentrations (1–8 μM) were infused intothe mass spectrometer to test whether LptA displays concen-tration-dependent oligomerization under electrospray condi-tions. Figure 4a shows the mass spectrum obtained with aprotein concentration of 8 μM. Several peaks can be identifiedin the high-m/z region of the spectrum, representing distinctLptA oligomeric species. The protein assembles sequentiallyinto progressively higher-order oligomers from the monomerto the pentamer, the highest stoichiometry identified under thehere employed conditions. A comparison between Figure 3aand Figure 4a shows that an increase of LptA concentrationfrom 1 to 8 μM promotes the transition from a nearly mono-disperse population of monomers to a heterogeneous ensembleof oligomeric species. Intermediate concentrations gave inter-mediate results (data not shown).
While the average charge state of each peak envelope increaseswith the molecular weight of the complex, the average charge persubunit remains relatively low, suggesting that all the detectedspecies maintain a native-like fold (Figure 4a). Furthermore, theself-association of LptA molecules does not seem to follow anyobligate stoichiometry. As the total protein concentration in-creases, higher-order structures accumulate by progressive addi-tion of one subunit. This feature suggests that the arrangement ofLptA subunits inside the complexes is not constrained by a closedarchitecture but is, on the contrary, prone to grow progressively,consistent with the head-to-tail interaction model [12, 13, 21].
ESI-IM-MS was employed in order to analyze the mor-phology of LptA oligomers. This method introduces anadditional observable, the collision cross section, based onion drift through a buffer gas, besides conventional time-of-flight (TOF) m/z determination [34–41]. These measurementsyield 2D plots of drift time versus m/z, with a “heat-map” styleintensity (Figure 4c). Since the mobility of the ions within thedrift cell depends on their overall shape and subunit topology, itis possible to separate different conformers and oligomers evenwithin the same m/z value. When performing this kind ofexperiments, it is important to limit collisional heating of thespecies in the gas phase, in order to avoid protein dissociation
Figure 4. LptA oligomerization. (a) Nano-ESI-MS spectrum of LptA 8 μM in 200 mM ammonium acetate, pH 8, 20 °C. Peaksare labeled according to quaternary structure (M, monomer; D, dimer; T, trimer; Q, tetramer; P, pentamer), and charge state. (b)Enlargement of panel (a) in the m/z range 4500–6000. (c) Nano-ESI-IM-MS spectrum of LptA 8 μM in 200 mM ammoniumacetate, pH 8, room temperature. Each spot is labeled according to quaternary structure and charge state. The spotscorresponding to the same oligomeric species are grouped into dotted ellipses. The spots labeled by asterisks refer to speciesthat accumulate as harsher parameters are applied (see also Figure S2). (d) Plot of normalized and averaged drift time as afunction of molecular weight. Each point is labeled according to quaternary structure. The line is a linear interpolation of theexperimental data (R2 = 0.99)
C. Santambrogio et al.: LptA Rod-Like Oligomers
or unfolding. The instrumental parameters were thereforecarefully tuned and the settings were chosen to optimizepreservation of non-covalent structures.
Figure 4c shows a typical drift plot. These data confirm theexistence of monomer-to-pentamer species distributions, al-ready observed in the normal ESI-MS spectrum. The signals ofeach distinct oligomeric state are enclosed in dashed ovals.Since drift time is also affected by charge, the signals of a givenstructure at different charge states lie on oblique lines. Most ofthe detected species give rise to one, well-defined spot in theplot, indicating the presence of a specific structure for eachparticular combination of subunit number and charge state.However, especially for large oligomers and high charge states,some ions present two or three distinct spots, corresponding todifferent drift times, indicating coexisting conformers. Thisbehavior most likely reflects structural perturbation of thecomplexes in the gas phase rather than a real structuralheterogeneity in solution. Indeed, the effect involves only thehigher charge states and become more pronounced as harsherinstrumental settings are applied (Figure S2 in OnlineResource). The spots whose intensity increases with harsherparameters are labeled by asterisks in Figure 4c. These resultsindicate that LptA assembles into oligomeric species of definedshape and, therefore, unique architecture.
ESI-IM-MS data can also provide some information on themorphology of the detected complexes by evaluating therelationship between ion drift times and oligomer size.Figure 4d shows a plot of the average drift time of each oligomerversus mass. The drift time of each ion type was corrected for thecharge state and the time delay between mobility measurementand entry into the TOF [36]. Upon such normalizations, the datapoints come to lie on a straight line. This behavior is typical ofrod-like supramolecular assemblies [36, 42, 43]. Indeed, it hasbeen shown that globular and extended structures can bediscriminated by their different profiles of average collisionalcross section as a function of mass, with globular speciessignificantly deviating from the linear dependence of extendedstructures [42, 43]. This result is consistent with the currentmodel for LptA oligomerization [13, 16, 21]. Thus, combiningESI-MS and ESI-IM-MS results, it is possible to conclude thatLptA oligomerizes with a rod-like architecture by single-subunitaddition, up to at least the pentamer. This observation providesfurther support and detail to the transenvelope bridge model[13]. The mean width estimated for the periplasmic spacesuggests that at least two molecules of LptA are necessary tocover the distance between IM and OM [21], although the inter-membrane distance can vary significantly [13]. The openarchitecture of LptA oligomers should allow membrane connec-tion irrespective of the required number of subunits, and iscompatible with the dynamic nature of the assembly.
Oligomerization Affects LptA Structureand Stability
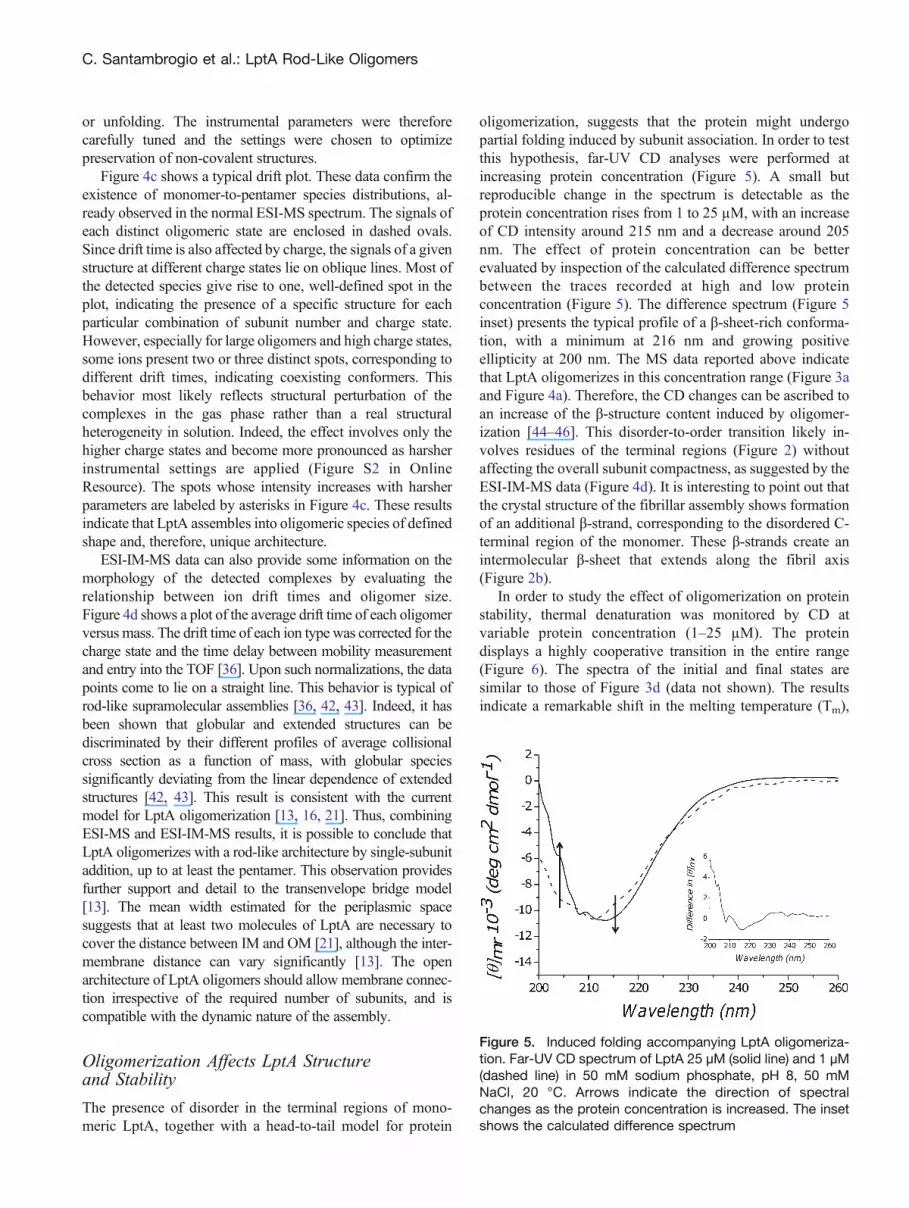
The presence of disorder in the terminal regions of mono-meric LptA, together with a head-to-tail model for protein
oligomerization, suggests that the protein might undergopartial folding induced by subunit association. In order to testthis hypothesis, far-UV CD analyses were performed atincreasing protein concentration (Figure 5). A small butreproducible change in the spectrum is detectable as theprotein concentration rises from 1 to 25 μM, with an increaseof CD intensity around 215 nm and a decrease around 205nm. The effect of protein concentration can be betterevaluated by inspection of the calculated difference spectrumbetween the traces recorded at high and low proteinconcentration (Figure 5). The difference spectrum (Figure 5inset) presents the typical profile of a β-sheet-rich conforma-tion, with a minimum at 216 nm and growing positiveellipticity at 200 nm. The MS data reported above indicatethat LptA oligomerizes in this concentration range (Figure 3aand Figure 4a). Therefore, the CD changes can be ascribed toan increase of the β-structure content induced by oligomer-ization [44–46]. This disorder-to-order transition likely in-volves residues of the terminal regions (Figure 2) withoutaffecting the overall subunit compactness, as suggested by theESI-IM-MS data (Figure 4d). It is interesting to point out thatthe crystal structure of the fibrillar assembly shows formationof an additional β-strand, corresponding to the disordered C-terminal region of the monomer. These β-strands create anintermolecular β-sheet that extends along the fibril axis(Figure 2b).
In order to study the effect of oligomerization on proteinstability, thermal denaturation was monitored by CD atvariable protein concentration (1–25 μM). The proteindisplays a highly cooperative transition in the entire range(Figure 6). The spectra of the initial and final states aresimilar to those of Figure 3d (data not shown). The resultsindicate a remarkable shift in the melting temperature (Tm),
Figure 5. Induced folding accompanying LptA oligomeriza-tion. Far-UV CD spectrum of LptA 25 μM (solid line) and 1 μM(dashed line) in 50 mM sodium phosphate, pH 8, 50 mMNaCl, 20 °C. Arrows indicate the direction of spectralchanges as the protein concentration is increased. The insetshows the calculated difference spectrum
C. Santambrogio et al.: LptA Rod-Like Oligomers

which increases from ~43 °C to ~51 °C in the exploredconcentration range. The inset shows the Tm profile as afunction of protein concentration. These results indicate thatLptA oligomerization drastically increases protein thermalstability. The loss of disordered regions likely adds to theincrease in particle size determining the observed increase inTm [47].
LPS Binding Increases LptA Stability
We also tested the effect of LPS on LptA stability. LptA-LPScomplex formation was assessed by co-purification on Ni-NTA-affinity columns and the results were analyzed bydenaturing electrophoresis (Figure S3 in Online Resource).LPS detected by silver staining displays the expectedheterogeneous profile. In the absence of LptA, LPS isdetected in the flow-through fraction, indicating that it doesnot bind to the resin (Figure S3a in Online Resource). Whenthe LptA-LPS mixtures are loaded, instead, LPS is retained onthe column and is eluted by imidazole, together with theprotein (Figure S3b in Online Resource). This result indicatesformation of a non-covalent complex between LptA and LPSunder the employed conditions, in agreement with previousreports [18].
The same conditions were then employed for CDmeasurements, in order to compare LptA response totemperature ramps in the absence or presence of LPS. In
the absence of the ligand (Figure S3c in OnlineResource), the spectra show the typical β-sheet-to-coiltransition. The midpoint is shifted towards highertemperature compared to lower protein concentrations(Figure 3d and Figure 6), consistent with the stabilizingeffect of protein oligomerization. The response of thesample containing the LptA-LPS complex, instead, isquite different (Figure S3d in Online Resource), indicat-ing substantial conservation of native-like secondarystructure, even at the highest tested temperature (65°C). This result indicates that LPS binding significantlystabilizes LptA against thermal denaturation. In the lightof the results presented in this work, LPS-mediatedstabilization of oligomeric structures likely contributesto the observed effect. Such a mechanism could berelevant from the biological point of view, and supportsthe hypothesis that oligomeric LptA is the form active inLPS transport.
ConclusionsThe results presented here indicate that LptA oligomerizes,even in the absence of LPS, into rod-like structures thatelongate by an n + 1 mechanism up to at least thepentamer. The oligomers present a well-defined architec-ture, though no fixed stoichiometry. The elongationmechanism is consistent with a head-to-tail interactionbetween subunits. The process is accompanied by adisorder-to-order transition, likely involving the terminalregions, and by an increase in thermostability. Thedisordered tracts near the chain ends could provideconformational plasticity for the instauration of highlyintertwined interaction surfaces. Altogether these resultsstrongly suggest that oligomeric LptA is present in activetransenvelope bridges. LPS binding to LptA also increasesprotein thermal stability. Together with previous crystal-lographic studies, this observation suggests that thepresence of LPS is an additional factor that favors LptAoligomerization.
Several residues implicated in LptA dimerization (I36,H37, S44, F54, F153, and V163), or interactions betweenLptA and LptC (H37), LptA and LptD (V163), or LptAand LPS (T32 and I36) are located in the disorderedtracts of the protein [13, 20, 21]. Interestingly, some ofthese residues (I36, H37, V163) are implicated inmultiple interactions. It thus appears that the disorderedregions of LptA may be reorganized by interactions withother LptA subunits, with different partners (LptC orLptD), or with the ligand (LPS) during transport.
Growing evidence suggests that the Lpt machinery is adynamic system, whose components assemble in response tothe environmental conditions [13, 19]. Our data arecompatible with a model where the LPS ligand promotes
Figure 6. Induced stability accompanying LptA oligomeriza-tion. Thermal unfolding transition by the ellipticity at 215 nm,for LptA 1 μM (black circles, dashed line) and 25 μM (blacksquares, solid line) in 50 mM sodium phosphate, pH 8, 50mM NaCl. The lines are sigmoidal interpolations of theexperimental data (R2 = 0.99). The arrow indicates thedirection of profile changes as the protein concentration isincreased. The inset shows the changes in the meltingtemperature (inflection point of the fitting curves) as afunction of protein concentration
C. Santambrogio et al.: LptA Rod-Like Oligomers

LptA oligomerization and thus the assembly of thetransenvelope bridge.
AcknowledgmentsThe authors acknowledge suppport for this work byFondazione Cariplo (grant 2010.0653), Regione Lombar-dia Cooperazione scientifica e tecnologica internazionale(grant 16876 SAL-18), MIUR-Regione Lombardia (ID30190679), and Fondazione per la Ricerca sulla FibrosiCistica (grant FFC#13/2010) with the contribution ofDonatori SMS solidale 2010. Funding from the HerculesFoundation (Belgium; for the Synapt instrument) and theFrancqui Foundation (personal grant to F.S.) is gratefullyacknowledged.
References1. Silhavy, T.J., Kahne, D., Walker, S.: The bacterial cell envelope. Cold
Spring Harb. Perspect. Biol. 2, 1–16 (2010)2. Nikaido, H.: Transport across the bacterial oter membrane. J. Bioenerg.
Biomembr 25, 581–589 (1993)3. Bos, M.P., Robert, V., Tommassen, J.: Biogenesis of the Gram-negative
bacterial outer membrane. Annu. Rev. Microbiol 61, 191–214 (2007)4. Nikaido, H.: Molecular basis of bacterial outer membrane permeability
revisited. Microbiol. Mol. Biol. Rev 67, 593–656 (2003)5. Raetz, C.R., Whitfield, C.: Lipopolysaccharide endotoxins. Ann. Rev.
Biochem 71, 635–700 (2002)6. Zhou, Z., White, K.A., Polissi, A., Georgopoulos, C., Raetz, C.R.:
Function of Escherichia coli MsbA, an essential ABC familytransporter, in lipid A and phospholipid biosynthesis. J. Biol. Chem273, 12466–12475 (1998)
7. Sperandeo, P., Dehò, G., Polissi, A.: The lipopolysaccharide transportsystem of Gram-negative bacteria. Biochim. Biophys. Acta 1791, 594–602 (2009)
8. Sperandeo, P., Villa, R., Dehò, G., Polissi, A.: The outer membrane ofGram-negative bacteria: lipopolysaccharide biogenesis and transport. In:Bacterial Membranes: Structural and Molecular Biology. Remaut, H.,Fronzes, R., Eds; Horizon Scientific Press: in press (2013).
9. Narita, S., Tokuda, H.: Biochemical characterization of an ABCtransporter LptBFGC complex required for the outer membrane sortingof lipopolysaccharides. FEBS Lett 583, 2160–2164 (2009)
10. Sperandeo, P., Lau, F.K., Carpentieri, A., De Castro, C., Molinaro, A.,Dehò, G., Silhavy, T.J., Polissi, A.: Functional analysis of the proteinmachinery required for transport of lipopolysaccharide to the outermembrane of Escherichia coli. J. Bacteriol 190, 4460–4469 (2008)
11. Sperandeo, P., Cescutti, R., Villa, R., Di Benedetto, C., Candia, D.,Dehò, G., Polissi, A.: Characterization of lptA and lptB, two essentialgenes implicated in lipopolysaccharide transport to the outer membraneof Escherichia coli. J. Bacteriol 189, 244–253 (2007)
12. Sperandeo, P., Villa, R., Martorana, A.M., Samalikova, M., Grandori,R., Dehò, G., Polissi, A.: New insights into the Lpt machinery forlipopolysaccharide transport to the cell surface: LptA-LptC interactionand LptA stability as sensors of a properly assembled transenvelopecomplex. J. Bacteriol 193, 1042–1053 (2011)
13. Freinkman, E., Okuda, S., Ruiz, N., Kahne, D.: Regulated assembly ofthe transenvelope protein complex required for lipopolysaccharideexport. Biochemistry 51, 4800–4806 (2012)
14. Chng, S.S., Gronenberg, L.S., Kahne, D.: Proteins required forlipopolysaccharide assembly in Escherichia coli form a transenvelopecomplex. Biochemistry 49, 4565–4567 (2010)
15. Freinkman, E., Chng, S.S., Kahne, D.: The complex that insertslipopolysaccharide into the bacterial outer membrane forms a two-protein plug-and-barrel. Proc. Natl. Acad. Sci. USA 108, 2486–2491(2011)
16. Suits, M.D., Sperandeo, P., Dehò, G., Polissi, A., Jia, Z.: Novelstructure of the conserved gram-negative lipopolysaccharide trans-port protein A and mutagenesis analysis. J. Mol. Biol 380, 476–488(2008)
17. Tran, A.X., Dong, C., Whitfield, C.: Structure and functional analysis ofLptC, a conserved membrane protein involved in the lipopolysaccharideexport pathway in Escherichia coli. J. Biol. Chem 285, 33529–33539(2010)
18. Tran, A.X., Trent, M.S., Whitfield, C.: The LptA protein of Escherichiacoli is a periplasmic lipid A-binding protein involved in the lipopoly-saccharide export pathway. J. Biol. Chem 283, 20342–20349 (2008)
19. Villa, R., Martorana, A.M., Okuda, S., Gourlay, L.J., Nardini, M.,Sperandeo, P., Dehò, G., Bolognesi, M., Kahne, D., Polissi, A.: TheEscherichia coli Lpt transenvelope protein complex for lipopolysaccha-ride export is assembled via conserved structurally homologousdomains. J. Bacteriol 195, 1100–1108 (2013)
20. Okuda, S., Freinkman, E., Kahne, D.: Cytoplasmic ATP hydrolysispowers transport of lipopolysaccharide across the periplasm in E. coli.Science 338, 1214–1217 (2012)
21. Merten, J.A., Schultz, K.M., Klug, C.S.: Concentration-dependentoligomerization and oligomeric arrangement of LptA. Protein Sci 21,211–218 (2012)
22. Laemmli, U.K.: Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227, 680–685 (1970)
23. Romero, P., Obradovic, Z., Li, X., Garner, E.C., Brown, C.J., Dunker, A.K.:Sequence complexity of disordered protein. Proteins 42, 38–48 (2001)
24. Li, X., Romero, P., Rani, M., Dunker, A.K., Obradovic, Z.: Predictingprotein disorder for N-, C-, and internal regions. Genome Informatics10, 30–40 (1999)
25. Lesse, A.J., Campagnari, A.A., Bittner, W.E., Apicella, M.A.: Increasedresolution of lipopolysaccharides and lipooligosaccharides utilizingtricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis.J. Immunol. Methods 126, 109–117 (1990)
26. Hitchcock, P.J., Brown, T.M.: Morphological heterogeneity amongSalmonella lipopolysaccharide chemotypes in silver-stained polyacryl-amide gels. J. Bacteriol 154, 269–277 (1983)
27. Longhi, S., Lieutaud, P., Canard, B.: Conformational disorder. MethodsMol. Biol 609, 307–325 (2010)
28. Kaltashov, I.A., Abzalimov, R.R.: Do ionic charges in ESI MS provideuseful information on macromolecular structure? J. Am. Soc. MassSpectrom 19, 1239–1246 (2008)
29. Grandori, R., Santambrogio, C., Brocca, S., Invernizzi, G., Lotti, M.:Electrospray-ionization mass spectrometry as a tool for fast screening ofprotein structural properties. Biotechnol. J 4, 73–87 (2009)
30. Robinson, C.V., Chung, E.W., Kragelund, B.B., Knudsen, J., Aplin,R.T., Poulsen, F.M., Dobson, C.M.: Probing the nature of noncovalentinteractions by mass spectrometry. A study of protein-CoA ligandbinding and assembly. J. Am. Chem. Soc 118, 8646–8653 (1996)
31. Invernizzi, G., Grandori, R.: Detection of the equilibrium foldingintermediate of beta-lactoglobulin in the presence of trifluoroethanol bymass spectrometry. Rapid Commun. Mass Spectrom 21, 1049–1052(2007)
32. Kelly, S.M., Jess, T.J., Prince, N.C.: How to study proteins by circulardichroism. Biochim. Biophys. Acta 1751, 119–139 (2005)
33. Ngounou Wetie, A., Sokolowska, I., Woods, A.G., Roy, U., Loo, J.A.,Darie, C.C.: Investigation of stable and transient protein–proteininteractions: Past, present, and future. Proteomics 13, 538–557 (2013)
34. Uetrecht, C., Rose, R.J., van Duijn, E., Lorenzen, K., Heck, A.J.: Ionmobility mass spectrometry of proteins and protein assemblies. Chem.Soc. Rev 39, 1633–1655 (2010)
35. Kaddis, C.S., Loo, J.A.: Native protein MS and ion mobility largeflying proteins with ESI. Anal. Chem 79, 1778–1784 (2007)
36. Ruotolo, B.T., Benesch, J.L., Sandercock, A.M., Hyung, S.J., Robinson,C.V.: Ion mobility-mass spectrometry analysis of large protein complexes.Nat. Protoc 3, 1139–1152 (2008)
37. Woods, L.A., Radford, S.E., Ashcroft, A.E.: Advances in ionmobility spectrometry-mass spectrometry reveal key insights intoamyloid assembly. Biochim. Biophys. Acta 1834(6), 1257–1268(2013)
38. Brocca, S., Testa, L., Sobott, F., Samalikova, M., Natalello, A., Papaleo,E., Lotti, M., De Gioia, L., Doglia, S.M., Alberghina, L., Grandori, R.:
C. Santambrogio et al.: LptA Rod-Like Oligomers

Compaction properties of an intrinsically disordered protein: Sic1 andits kinase-inhibitor domain. Biophys. J 100, 2243–2252 (2011)
39. Konijnenberg, A., Butterer, A., Sobott, F.: Native ion mobility-massspectrometry and related methods in structural biology. Biochim.Biophys. Acta 1834(6), 1239–1256 (2013)
40. Sobott, F.: Characterization of proteins and protein complexes by ionmobility-mass spectrometry. Chemistry Today 29, 20–24 (2011)
41. Jurneczko, E., Barran, P.E.: How useful is ion mobility massspectrometry for structural biology? The relationship between proteincrystal structures and their collision cross sections in the gas phase.Analyst 136, 20–28 (2011)
42. Smith, D.P., Radford, S.E., Ashcroft, A.E.: Elongated oligomers inbeta2-microglobulin amyloid assembly revealed by ion mobilityspectrometry-mass spectrometry. Proc. Natl. Acad. Sci. USA 107,6794–6798 (2010)
43. Borysik, A.J., Robinson, C.V.: The ‘sticky business’ of cleaning gas-phase membrane proteins: a detergent oriented perspective. Phys.Chem., Chem. Phys 14, 14439–14449 (2012)
44. Receveur-Brechot, V., Bourhis, J.M., Uversky, V.N., Canard, B.,Longhi, S.: Assessing protein disorder and induced folding. Proteins62, 24–45 (2006)
45. Tompa, P.: Intrinsically disordered proteins: a 10-year recap. TrendsBiochem. Sci 37, 509–516 (2012)
46. Dunker, A.K., Brown, C.J., Lawson, J.D., Iakoucheva, L.M.,Obradovic, Z.: Intrinsic disorder and protein function. Biochemistry41, 6573–6582 (2002)
47. Melnik, B.S., Povarnitsyna, T.V., Glukhov, A.S., Melnik, T.N., Sarma,R.H.: SS-stabilizing proteins rationally: intrinsic disorder-based designof stabilizing disulphide bridges in GFP. J. Biomol. Struct. Dyn 29,815–824 (2012)
C. Santambrogio et al.: LptA Rod-Like Oligomers





























