Embed Size (px)
Citation preview

1886
Ecology, 83(7), 2002, pp. 1886–1898q 2002 by the Ecological Society of America
LIGHT, NUTRIENTS, AND P:C RATIOS IN ALGAE: GRAZERPERFORMANCE RELATED TO FOOD QUALITY AND QUANTITY
DAG O. HESSEN, PER J. FÆRØVIG, AND TOM ANDERSEN
University of Oslo, Department of Biology, P.O. Box 1027 Blindern, N-0316 Oslo, Norway
Abstract. Continuous cultures of the green algae Selenastrum capricornutum weregrown at different concentrations of total phosphorus (1–50 mmol P/L) and different lightlevels (10–200 mmol quanta·m22·s21). Growth yields in terms of C, N, and P were positivelycorrelated to total P level. Total biomasses in terms of C and N, but not P, were alsopositively correlated to light level. The cell quotas of C were positively correlated withlight, whereas cell quotas of P were negatively correlated with light. The resulting elementalratios gave a quite consistent pattern, where high light caused significant reductions in bothN:C and P:C ratios, as well as strong reductions in chlorophyll to carbon ratios. The increasein P:C ratio with increasing total P and decreasing light can be interpreted as an adaptiveresponse to self-shading. In that case, our results indicate that light adaptation not onlyinvolves a cost in terms of increased chlorophyll synthesis, but also in terms of increasedP demands. This provides new insight not only into the physiological regulation of C andP uptake in algae, but it could also explain deviations from the Redfield ratio.
The growth of juvenile Daphnia magna fed S. capricornutum from the different lightand phosphorus treatments was studied in a series of short-term assays (7 d) covering agradient of food concentrations. The response of Daphnia growth rate along this quantity(0.5–5.0 mg C/L) and quality (0.5–12 mg atomic P·[mg atomic C]21) gradient gave a closefit to a double hyperbola model. Changes in elemental ratios of the algae were reflected inthe growth rate of Daphnia, such that up to 40% reduction in its growth rate could beattributed to increased C:P ratios. This study demonstrates that the physiological responsesof phototrophs in terms of chlorophyll content and elemental composition depend stronglyon ambient light and nutrient regimes. It also confirms that these patterns can yield con-trasting responses on herbivore growth responses along the food quantity and quality axes.
Key words: Daphnia; food quality; grazer; green algae (Selenastrum capricornutum); light;nutrients; photosynthesis.
INTRODUCTION
Aquatic ecosystem productivity is governed primar-ily by the inputs of light and nutrients, both in absoluteand relative terms. While the quantitative yield of pri-mary production is related to the quantity of photo-synthetic active radiation (PAR) and key nutrients, re-cent studies demonstrate that the qualitative output interms of elemental ratios may be determined by thebalance of light and nutrients (Urabe and Sterner 1996,Sterner et al. 1997). At high light levels, photosyntheticcarbon fixation is kept high, and unless the total supplyof inorganic phosphorus is also high, the uptake of Cand P may be desynchronized, causing decreasing P:Cratios. This may be manifested as an accumulation ofC-rich storage compounds such as lipids or starch inalgal cells (Berman-Frank and Dubinsky 1999). Con-versely, P:C ratios tend to be high under low light andabundant P (Urabe and Sterner 1996).
Algae have a large capacity for acclimating theirphotosynthetic machinery to different light conditions.Increasing the investment into light-harvesting pig-
Manuscript received 19 December 2000; revised 28 May2001; accepted 19 June 2001; final version received 10 October2001.
ments, as reflected by the chlorophyll to carbon ratio,is a well-documented response to light limitation acrossa wide variety of algal taxa (Geider 1987). Moreover,there is a growing body of evidence indicating thatphotoadaptation to low light conditions also carries acost in terms of increased N demands, mainly due tothe high protein content of photosynthetic machinery(Raven 1984). While there is evidence that the syn-thesis of chlorophyll, and thus the capacity for lightadaptation, may be constrained by N limitation (Pre-zelin and Matlick 1983), there appear to be no dataconsidering any corresponding effect of P limitation.
Primary producers may exhibit a highly flexible cellstoichiometry with P:C ratios spanning more than anorder of magnitude (Andersen 1997, DeMott et al.1998). Metazoan herbivores, in comparison, have amuch tighter regulation of their stoichiometry, withzooplankton species like Daphnia spp. having far high-er P:C ratios than that normally encountered in theirfood (Andersen and Hessen 1991, Sterner and Hessen1994). When the P:C ratio of the algal food decreasesbeyond the threshold for balanced net intake of C rel-ative to P, growth efficiency in terms of C will declinesince an increasing share of gross ingested C cannot

July 2002 1887ALGAL PHOTOSYNTHESIS AND GRAZER FITNESS
be utilized for balanced somatic growth (Hessen 1992,Sterner and Hessen 1994, DeMott et al. 1998).
A number of experiments have verified that thegrowth of zooplankton herbivores, notably P-demand-ing species like Daphnia spp., indeed may be con-strained at sufficiently low food P:C ratios (Urabe andWatanabe 1992, Sterner 1993, Urabe et al. 1997,DeMott et al. 1998). The existence of actual thresholdsfor P limitation is disputable, however. Based on the-oretical and empirical evidence, P:C ratios over therange from 3–7 mg atomic P·[mg atomic C]21 (atomicratio C:P 5 150–300) have been proposed as thresholdsfor the onset of P limitation in Daphnia (Hessen 1992,Urabe and Watanabe 1992, Sterner 1993). More recentexperiments by DeMott et al. (1998) point to a thresh-old for reduced growth that exceeds 10 mg atomicP·[mg atomic C]21 for juvenile Daphnia (C:P atomicratio ,100). This will not be a fixed limit, however,but will depend on the grazers’ assimilation of elementsfrom the food particles and on the grazers’ own somaticstoichiometry, which is not entirely fixed. The foodquantity also may in itself be a determinant of thresh-olds for P limitation, since when the grazer encountersfood concentrations where net C intake approachesmaintenance metabolism, the C:P threshold for poten-tial P limitation will rise to infinity (Sterner and Rob-inson 1994).
The bottom line of the P limitation argument is thathigh growth rates increase the demand for RNA (Hes-sen 1990, Elser et al. 2000b), and particularly ribo-somal RNA. P deficiency thus translates into lowergrowth rates (DeMott et al. 1998). The evolutionaryexplanation for such a mismatch may, from the auto-trophs’ perspective, be seen as an adaptation to causelower grazer fitness (at the expense of its own growth,however; cf. White 1993), while the grazers’ dilemmawould be the trade-off between high protein synthesisand the risk of direct mineral limitation.
Under conditions with light limitation and P satu-ration, algal biomass will be low, but the algal cellswill contain high P:C ratio (i.e., high food quality forherbivores). Following a transition towards light sat-uration while maintaining P deficiency, algal biomasswill increase and finally reach the carrying capacity,but the P:C ratio of the cells will decrease. This shiftin elemental stoichiometry could cause a trade-off sce-nario for grazers along the same gradient of increasedlight and decreased P (Urabe and Sterner 1996). Theherbivores’ growth rate would be predicted to increaseinitially due to increased biomass of high quality algae,and to reach a peak growth rate at the optimum light:nutrient balance, before finally declining due to de-creasing food P:C (i.e., decreased quality).
The relative importance of food quality vs. foodquantity limitation will be of vital importance for as-sessments of ecosystem productivity. While food quan-tity limitation is assumed to be more important thanfood quality limitation in most localities, the latter may
pose additional constraints not only for zooplanktonsecondary production, but also for a wide diversity ofterrestrial grazers ranging from insects to mammals(McNaughton 1990, White 1993, Elser et al. 2000a).The P:C or N:C ratios of phototrophs are commonlyfar below those of their grazers in both terrestrial andaquatic ecosystems (Elser et al. 2000a), and this stoi-chiometric mismatch may have profound effects on en-ergy flow and community structure. For example, Hes-sen and Faafeng (2000) found a mean sestonic P:C ratioof 4.5 mg atomic P·[mg atomic C]21 in 110 Norwegianlakes, and estimated that P limitation could potentiallyconstrain growth rates of P-demanding taxa like Daph-nia spp. in a majority of the surveyed lakes.
If food quality can be governed by large-scale pat-terns of light and nutrients, this raises a set of intriguingissues related to both the within-lake variability of foodquality that occurs over season and depth, and the be-tween-lake variability that exist with respect to pro-ductivity and light penetration. It also poses far-reach-ing questions regarding the control of carbon seques-tration, storage, and export in ecosystems. In the pres-ent account we focus on the steady state responses ofalgae grown under light and nutrient limitation in con-tinuous culture, and their subsequent short- and inter-mediate-term effects on herbivore grazing and growthrates.
MATERIAL AND METHODS
Experimental setup
The experiments were performed in a continuous,two-step, culture system, consisting of a primary pho-totroph chamber and a secondary herbivore chamber.The primary step was a 2-L glass vessel receiving anominal light intensity (I ) of 70 mmol quanta·m22·s21
from 25-W blue-white fluorescent tubes. Light gradi-ents were implemented by varying the distances fromthe light sources, and by wrapping culture vessels inneutral density filters. For the first test, experimentswere run in duplicates over a four-step gradient of light(I ) from 23 to 200 mmol quanta·m22·s21. Since the high-est light level (200 mmol quanta·m22·s21) tended to givechlorotic and unstable cultures, the remaining experi-ments were run in triplicates under only two light re-gimes: 70 mmol quanta·m22·s21 (light saturation) and10 mmol quanta·m22·s21 (light limitation). The primaryproducer was the green algae Selenastrum capricor-nutum grown in COMBO medium (Kilham et al. 1998).A total P gradient was achieved by reducing P in themedium, yielding nominal P concentrations of 1, 1.8,2.5, 5.0, 10, and 50 mmol/L. For all levels, actual con-centrations were monitored by routine analysis of totalP in the reservoir tanks. The full COMBO medium has1000 mmol N/L and 50 mmol P/L and this N:P ratio of20 (atomic : atomic) is close to the optimal N:P ratiofor Selenastrum capricornutuum of 22 (Rhee and Goth-am 1980). At 50 mmol P/L, we assume no nutrient

1888 DAG O. HESSEN ET AL. Ecology, Vol. 83, No. 7
limitation, while for all treatments with 10 mmol P/Land less (i.e., N:P $ 100), there will be an increasingdegree of P limitation.
Peristaltic pumps supplied medium at a flow corre-sponding to a dilution rate of 0.42 d21 in the phototrophstep. Timer-controlled magnetic stirrers mixed the pri-mary chambers. Stirring was found preferable to airbubbling because air bubbles tended to cause super-saturation problems in the downstream Daphnia cul-tures.
Selenastrum cultures were allowed to run for 2 wkbefore the start of the Daphnia growth experiments, inorder to obtain a stable level of cell numbers, cell vol-umes, and particulate C, N, and P. Algae from all cul-tures, i.e., from all light and nutrient treatments, werediluted to standardized particulate carbon concentra-tions of 0.5, 1.0, and 1.5 mg/L before being fed to theDaphnia cultures. For the 1.0, 1.8, and 10 mmol P/Ltreatments, an additional high food concentration treat-ment level (5 mg C/L) was also run. Individual growthrates of Daphnia were followed in COMBO medium(without N and P to avoid changes in algal stoichi-ometry) for the first 7 d after hatching under all ex-perimental protocols. Neonates, ,24 h old, obtainedfrom the same well-fed stock culture, were placed in-dividually in 20-mL vials and fed standardized foodconcentrations. Somatic growth rates were measuredon five parallel vials for each food concentration andeach combination of light and P treatments. Growthrates were based on weight estimates obtained by anondestructive image analysis technique, which will bedescribed in full detail elsewhere (Færøvig et al. 2002).Comparison of measured dry masses and volumes ob-tained by image analysis gave a close fit over the entiregradient from neonates to senescent individuals (r2 50.96, n 5 60).
Previous studies have suggested that the increasedcell wall thickness in nutrient-limited Selenastrumcould induce a grazing resistance (Van Donk and Hes-sen 1993, Van Donk et al. 1997). The reduced assim-ilation efficiency of thick-walled cells will be perceivedas reduced clearance rates in short-term grazing ex-periments, since ingested algae would be defecated aswhole cells, and interpreted as noningested cells. Sincea considerable share of nutrient-stressed algae may passintact thorough the gut, this would also translate toreduced growth rates of the grazers. Two grazing ex-periments with different sized Daphnia on differentfood qualities (C:P ratios) were performed to check forthis possibility. Both grazing experiments used algaegrown in 1.8 mmol P/L medium at low and high lightintensity, yielding P:C ratios of 2.5 and 1.0 mg atomicP·[mg atomic C]21, diluted to a fixed food concentration(5 mg C/L). The first experiment used four differentage groups (2, 4, 8, and 25 d old) of Daphnia raisedin high quality food (high P:C). The second experimentused a single cohort of animals (starting from ,24-h-old neonates), raised in the same food qualities as used
in the grazing assays. Food and media were replenishedevery day, while individual growth rates were followedover this period. Grazing rates were measured after 2,4, and 8 d. Each grazing experiment was run in trip-licate with 30 Daphnia per 200 mL beaker, againstbeakers with zero grazers as controls. Ingestion andclearance rates were estimate by flow cytometric(FCM) analysis of cell numbers and autofluorescenceproperties. Samples for FCM were collected after 1 and10 min, and then every 30 min over a 6-h period.
Analytical protocols
The phototroph growth chambers were sampled atleast twice a week. Subsamples of 100 mL were filteredon pre-ignited (5008C, 2 h) GF/F filters and analyzedfor particulate C and N on a Carlo-Erba CHN 1106elemental analyzer (CE Elantech, Lakewood, New Jer-sey, USA). Particulate P was measured on correspond-ing filters placed in 15 mL distilled water that wasacidified (150 mL of 4.0 mol/L H2SO4), added perox-odisulfate (0.15 g K2S2O8), and autoclaved (1218C, 1h). The particulate fraction of P was then analyzedspectrophotometrically by the standard ammonium-molybdate method. Total P and dissolved P were an-alyzed correspondingly on unfiltered and GF/F filteredwater samples. The freshwater food web literature of-fers a confusing variety of units for reporting elementalratios, e.g., as both C:P or P:C ratios, on both weightor molar basis. There are arguments in favor of allalternatives, but for compositional ratios it appears log-ical to put the major element in the denominator. Thus,throughout this paper, ratios are expressed as P:C, mgatomic P·[mg atomic C]21. Due to convention, concen-trations of P are expressed in molar units, while bio-masses and yields are expressed in mass units.
Algal cell numbers and size distributions were an-alyzed by flow cytometry. Samples of 1.0 mL werecollected in Eppendorf vials, preserved with 20 mL50% glutaraldehyde (1% final concentration), andstored in a refrigerator until analysis. FCM analyseswere performed on a Skatron Argus 100 flow cytometer(Skatron A/S, Oslo, Norway) equipped with high-pow-er mercury arc lamp (103 W) and a 100-mm nozzle.The instrument was modified with gas-pressure-con-trolled sheath flow delivery and a syringe-based sampleinjection system, which minimized sample cross-con-tamination. Monodisperse latex beads (PolysciencesNo. 23517, Europa GmbH, Eppelheim, Germany; meandiameter 1.0 mm, CV 5 1.27%) were used for instru-mental alignment and calibration. By using autofluores-cence as trigger channel, egested cells would be ex-cluded from FCM counts (cell autofluorescence wasseverely reduced by passage through the animal gut).Triggering on forward scatter and gating on fluores-cence enabled differentiation between bacteria and Se-lenastrum in the FCM counts. Even under low lightand high P, the share of bacteria gave a minor contri-bution (,5%) to total biomass. For determinations of

July 2002 1889ALGAL PHOTOSYNTHESIS AND GRAZER FITNESS
FIG. 1. Culture yields in terms of particulate organic car-bon (POC), particulate organic nitrogen (PON), particulateorganic phosphorus (POP), and chlorophyll a in the contin-uous cultures over a gradient of total P from 1 to 50 mmol/L. Open bars represent high light treatments (70 mmolquanta·m22·s21), and shaded bars are the low light treatments(10 mmol quanta·m22·s21). Each bar represents the mean 1 1SD for a minimum of six separate measurements over a 2-wkperiod.
chlorophyll a, 25-mL subsamples were filtered in thedark on GF/F filters and stored at 2258C in the darkbefore analysis. After testing with both acetone (90%)and methanol (100%) extraction, the latter method waschosen due to its higher extraction efficiency.
RESULTS
Effects on autotrophs
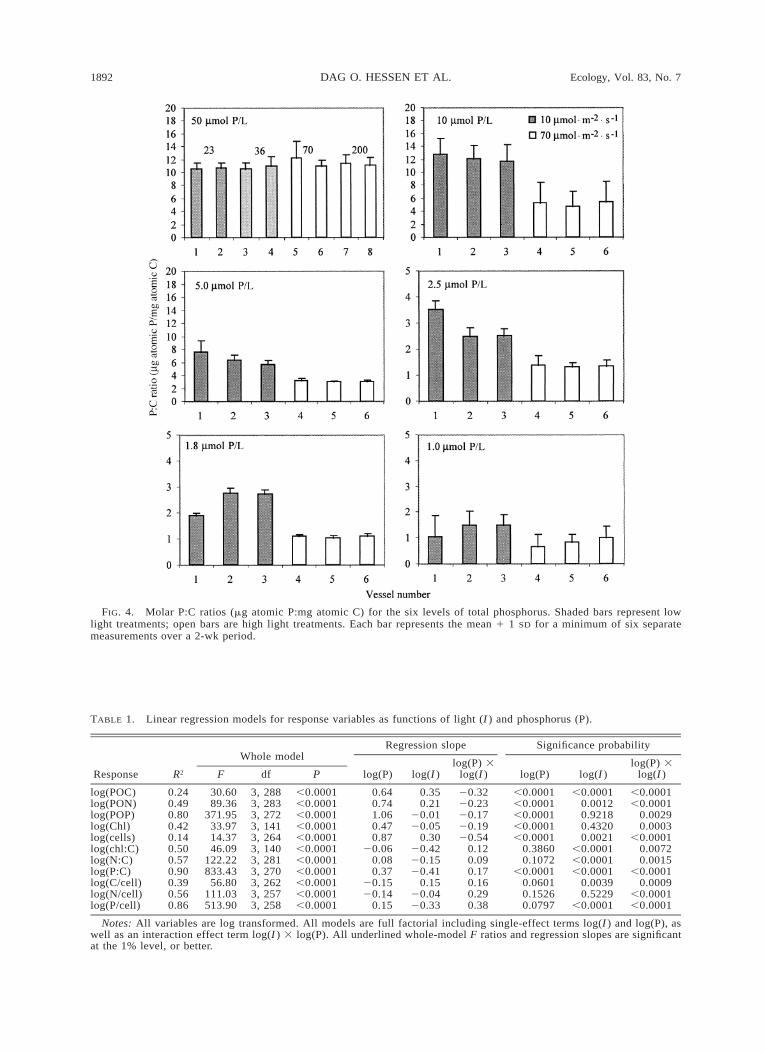
The effects of different light levels on phytoplanktonyield in terms of particulate C, N, and P as well aschlorophyll were quite pronounced (Fig. 1). In lowlight, algal biomass increased threefold across the totalP gradient (from ;4 to 12 mg C/L), while the highlight treatment at the same total P level typically dou-bled the carbon biomass yield. The low yield in the 10mmol P/L treatment could have been caused by sub-stantial growth on the growth chamber walls under highlight. Particulate N yield resembled the pattern for par-ticulate C with a less pronounced effect of light, whilethe yield of particulate P increased more than tenfoldover the total P gradient for both light regimes. Cellquotas of C showed low variability and ranged from20 to 60 pg/cell in all #10 mmol P/L treatments, butexceeded 100 pg/cell at the highest total P and lightlevels (Fig. 2). This contrasts with the cell quotas ofP, which increased from near 0.1 pg/cell at low ambientP to more than 3 pg/cell at 50 mmol P/L (Fig. 3). Thedifferent responses of cell quotas of C and P along theP gradient at the two light regimes were manifested asa consistent pattern in cell P:C ratios. In full-strengthCOMBO medium (50 mmol P/L), no changes in Se-lenastrum P:C were detected over a gradient from 23to 200 mmol quanta·m22·s21 (Fig. 4). P:C was close to12 mg atomic P·[mg atomic C]21, with no significantdifferences between treatments. When ambient P wasreduced, there was a successive decrease in the P:Cratio to a level of 2.5–1.5 mg atomic P·[mg atomic C]21
at low light, and ,0.8 mg atomic P·[mg atomic C]21 athigh light.
Responses of phototroph biomass yield and com-position to the experimental treatment variables wereanalyzed statistically by regression analysis. Depen-dent variables included biomass yields (as particulateC, N, P, chlorophyll a, and cell abundance), cell quotas(as C, N, P per cell), and cell composition ratios (N:C,P:C, chlorophyll:C). As both independent variablesspanned more than an order of magnitude, we choseto log10 transform all variables (both dependent andindependent). All regression models used the same fullfactorial set of dependent variables, including two sin-gle effect terms and a multiplicative interaction term.All yield-type response variables (particulate C, N, P,chlorophyll a, and cell abundance) showed significantpositive relationships with total P level (Table 1).Yields of particulate C, N, and cell number—but notyields of particulate P and chlorophyll a—also in-creased significantly with light level. It is noticeable
that the regression slopes of the total P effect are gen-erally close to 1, while the light effect slopes are sub-stantially ,1. This implies close to linear proportion-ality to total P and sublinear proportionality to light in

1890 DAG O. HESSEN ET AL. Ecology, Vol. 83, No. 7
FIG. 2. Cell quotas of carbon for six levels of total phosphorus. Shaded bars represent low light treatments; open barsare high light treatments. Each bar represents the mean 1 1 SD for a minimum of six separate measurements over a 2-wkperiod. The first (50 mmol P/L) treatment was designed differently from the others, with duplicate runs for the four lightlevels indicated above (as mmol quanta·m22·s21), instead of triplicate runs at two light levels.
the equivalent, antilog transformed model. The subline-arity of the light effect is most likely a reflection ofthe nonlinearities typically associated with light satu-ration phenomena. Finally, all yield-type variables hadsignificant negative slopes with respect to the light 3nutrient interaction term.
The consistent pattern of signs and magnitudes ofthe regression slopes (Table 1) strongly suggests thatthe interaction term may be interpreted as the modu-lating effect of total P on the response to light con-ditions. Thus, a given increment in light level appearsto give a larger biomass yield increase at low than athigh total P. It is reasonable to attribute this to theincreased self-shading effects with increasing carryingcapacity, and the corresponding cellular adaptive re-sponses to compensate for this. This interpretation issupported by the pattern of regression slopes for thecell-composition-type variables in Table 1 (chloro-
phyll:C, N:C, P:C), which all have negative light ef-fects and positive interaction terms. In other words, thecarbon specific cell content of chlorophyll, N, or P canbe increased by either decreasing the light at constanttotal P, or by increasing total P at constant light. Theoverall similarity between the adaptive response to selfshading and light limitation in terms of P:C is roughlythe same as for chlorophyll:C and N:C. The regressionmodels for the cell quota-type variables (C/cell, N/cell,P/cell) all have nonsignificant total P effects and sig-nificant positive interaction effects, while the light ef-fects show different patterns (positive for C/cell, non-significant for N/cell, and negative for P/cell).
The strong interaction between the total P and lightlevel on the size distributions of Selenastrum cells isalso clearly by the flow cytometric analyses. The for-ward scatter signal in flow cytometry is generally foundto be closely related to cell size, although it is also

July 2002 1891ALGAL PHOTOSYNTHESIS AND GRAZER FITNESS
FIG. 3. Cell quotas of phosphorus for six levels of total phosphorus. Shaded bars represent low light treatments; openbars are high light treatments. Each bar represents the mean 1 1 SD for a minimum of six separate measurements over a 2-wk period.
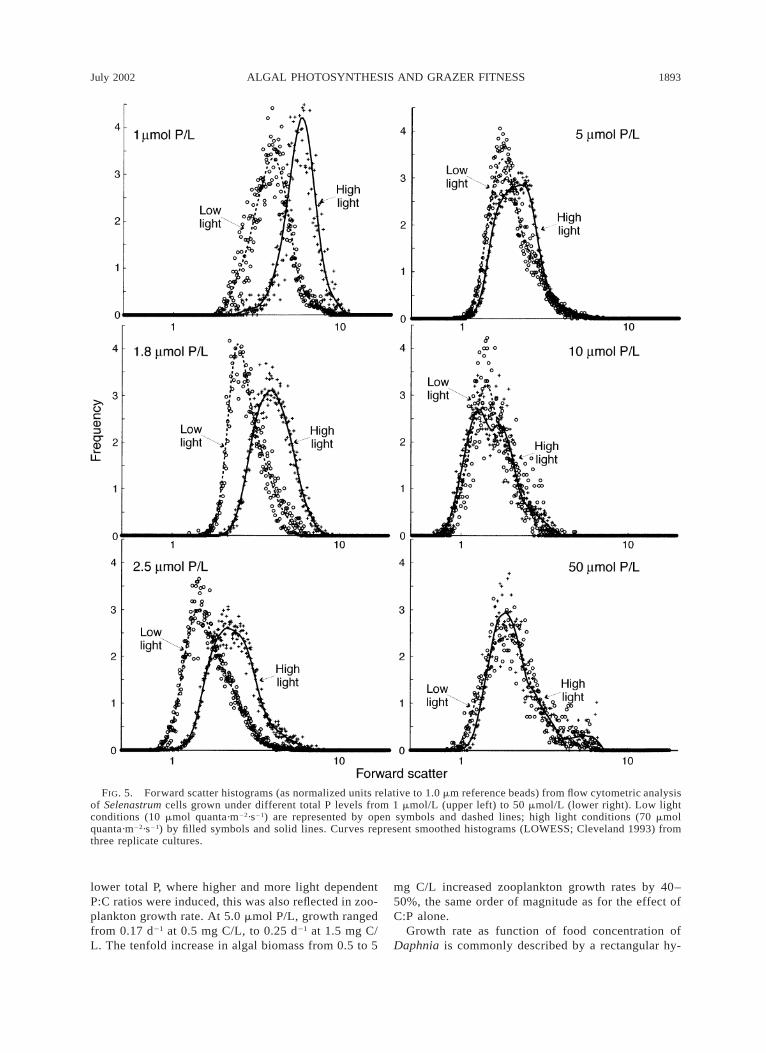
known to be influenced by other factors like cell shape,texture, and refractive index (Shapiro 1995). The for-ward-scatter cytograms (Fig. 5) show no effect of lightlevel on cell size or shape at the highest total P level(50 mmol/L). At total P concentrations of 5 mmol/Land below, there was both a gradual increase in meanforward scatter signal with decreasing total P, and anincreasing deviation between the high and low lighttreatments. Cell size distributions were almost identicalamong the three replicated cultures for all total P andlight treatments.
Effect on grazers
The additive effect of high light and low P yieldeda substantial increase in cell size, but the cells wouldstill be within a grazable range for all size categoriesof Daphnia. There could, however, be more qualitativechanges associated with the increased volume, such aschanged cell wall properties and biochemical compo-
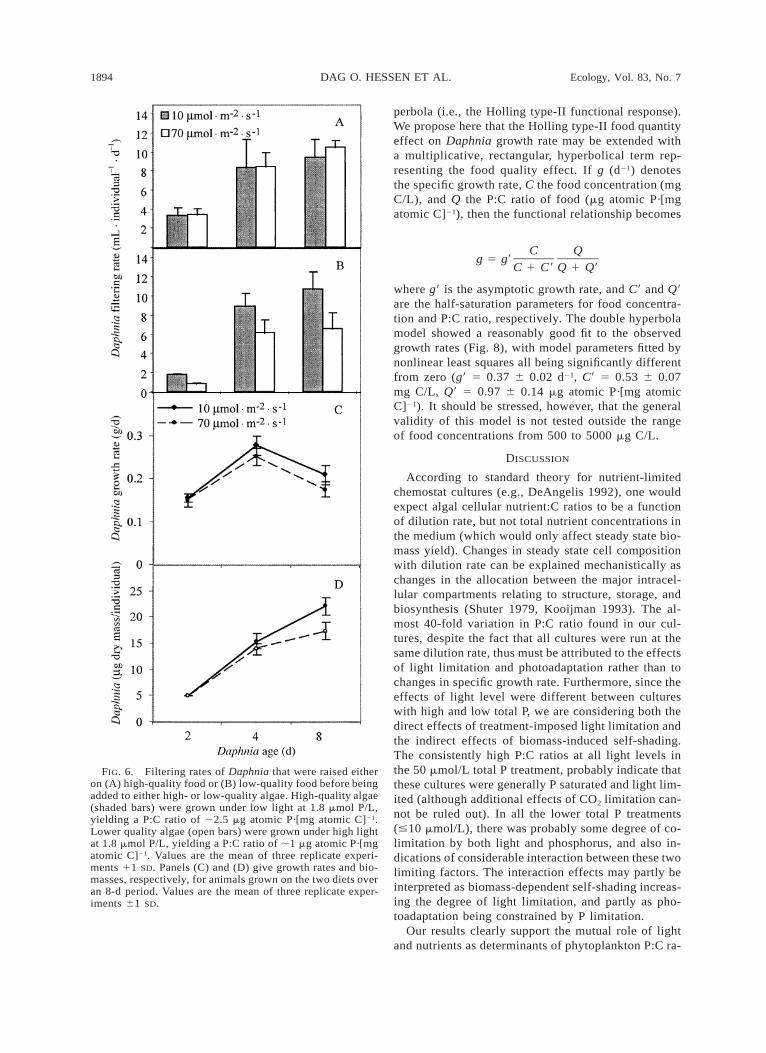
sition that would affect the next trophic level. The twoseparate grazing assays demonstrated two different as-pects (Fig. 6). If the animals were raised on an optimal(low C:P) diet, and then offered algae with differentC:P ratios, there were no difference in clearance rateseither in 2, 4, or 8-d-old animals. A separate grazingassay was also performed with large (25-d-old) animalson the same food, and also this yielded almost identicalclearance rates (52–55 mL·individual21·d21). If, how-ever, the animals were raised on low and high C:P diets,respectively, the individuals fed high C:P (low quality)food had reduced clearance rates. This is most obvi-ously a consequence of lower growth rates (Fig. 6) andthus reduced per capita clearance rates due to reducedage-specific size.
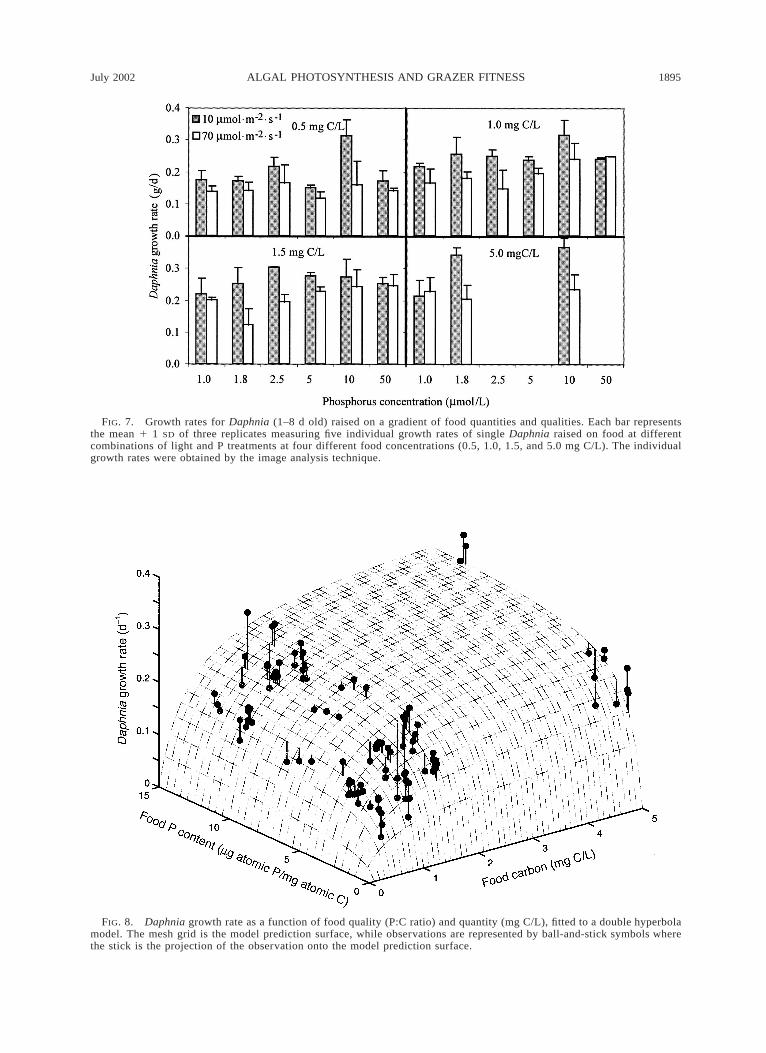
For algae grown in full COMBO medium (50 mmolP/L), there was no difference between light treatmentson Daphnia growth rates (Fig. 7), which ranged from0.22 d21 at 0.5 mg C/L to 0.28 d21 at 1.5 mg C/L. At
1892 DAG O. HESSEN ET AL. Ecology, Vol. 83, No. 7
FIG. 4. Molar P:C ratios (mg atomic P:mg atomic C) for the six levels of total phosphorus. Shaded bars represent lowlight treatments; open bars are high light treatments. Each bar represents the mean 1 1 SD for a minimum of six separatemeasurements over a 2-wk period.
TABLE 1. Linear regression models for response variables as functions of light (I ) and phosphorus (P).
Response R2
Whole model
F df P
Regression slope
log(P) log(I )log(P) 3
log(I )
Significance probability
log(P) log(I )log(P) 3
log(I )
log(POC)log(PON)log(POP)log(Chl)log(cells)log(chl:C)log(N:C)log(P:C)log(C/cell)log(N/cell)log(P/cell)
0.240.490.800.420.140.500.570.900.390.560.86
30.6089.36
371.9533.9714.3746.09
122.22833.43
56.80111.03513.90
3, 2883, 2833, 2723, 1413, 2643, 1403, 2813, 2703, 2623, 2573, 258
,0.0001,0.0001,0.0001,0.0001,0.0001,0.0001,0.0001,0.0001,0.0001,0.0001,0.0001
0.640.741.060.470.87
20.060.080.37
20.1520.14
0.15
0.350.21
20.0120.05
0.3020.4220.1520.41
0.1520.0420.33
20.3220.2320.1720.1920.54
0.120.090.170.160.290.38
,0.0001,0.0001,0.0001,0.0001,0.0001
0.38600.1072
,0.00010.06010.15260.0797
,0.00010.00120.92180.43200.0021
,0.0001,0.0001,0.0001
0.00390.5229
,0.0001
,0.0001,0.0001
0.00290.0003
,0.00010.00720.0015
,0.00010.0009
,0.0001,0.0001
Notes: All variables are log transformed. All models are full factorial including single-effect terms log(I ) and log(P), aswell as an interaction effect term log(I ) 3 log(P). All underlined whole-model F ratios and regression slopes are significantat the 1% level, or better.
July 2002 1893ALGAL PHOTOSYNTHESIS AND GRAZER FITNESS
FIG. 5. Forward scatter histograms (as normalized units relative to 1.0 mm reference beads) from flow cytometric analysisof Selenastrum cells grown under different total P levels from 1 mmol/L (upper left) to 50 mmol/L (lower right). Low lightconditions (10 mmol quanta·m22·s21) are represented by open symbols and dashed lines; high light conditions (70 mmolquanta·m22·s21) by filled symbols and solid lines. Curves represent smoothed histograms (LOWESS; Cleveland 1993) fromthree replicate cultures.
lower total P, where higher and more light dependentP:C ratios were induced, this was also reflected in zoo-plankton growth rate. At 5.0 mmol P/L, growth rangedfrom 0.17 d21 at 0.5 mg C/L, to 0.25 d21 at 1.5 mg C/L. The tenfold increase in algal biomass from 0.5 to 5
mg C/L increased zooplankton growth rates by 40–50%, the same order of magnitude as for the effect ofC:P alone.
Growth rate as function of food concentration ofDaphnia is commonly described by a rectangular hy-
1894 DAG O. HESSEN ET AL. Ecology, Vol. 83, No. 7
FIG. 6. Filtering rates of Daphnia that were raised eitheron (A) high-quality food or (B) low-quality food before beingadded to either high- or low-quality algae. High-quality algae(shaded bars) were grown under low light at 1.8 mmol P/L,yielding a P:C ratio of ;2.5 mg atomic P·[mg atomic C]21.Lower quality algae (open bars) were grown under high lightat 1.8 mmol P/L, yielding a P:C ratio of ;1 mg atomic P·[mgatomic C]21. Values are the mean of three replicate experi-ments 11 SD. Panels (C) and (D) give growth rates and bio-masses, respectively, for animals grown on the two diets overan 8-d period. Values are the mean of three replicate exper-iments 61 SD.
perbola (i.e., the Holling type-II functional response).We propose here that the Holling type-II food quantityeffect on Daphnia growth rate may be extended witha multiplicative, rectangular, hyperbolical term rep-resenting the food quality effect. If g (d21) denotesthe specific growth rate, C the food concentration (mgC/L), and Q the P:C ratio of food (mg atomic P·[mgatomic C]21), then the functional relationship becomes
C Qg 5 g9
C 1 C9 Q 1 Q9
where g9 is the asymptotic growth rate, and C9 and Q9are the half-saturation parameters for food concentra-tion and P:C ratio, respectively. The double hyperbolamodel showed a reasonably good fit to the observedgrowth rates (Fig. 8), with model parameters fitted bynonlinear least squares all being significantly differentfrom zero (g9 5 0.37 6 0.02 d21, C9 5 0.53 6 0.07mg C/L, Q9 5 0.97 6 0.14 mg atomic P·[mg atomicC]21). It should be stressed, however, that the generalvalidity of this model is not tested outside the rangeof food concentrations from 500 to 5000 mg C/L.
DISCUSSION
According to standard theory for nutrient-limitedchemostat cultures (e.g., DeAngelis 1992), one wouldexpect algal cellular nutrient:C ratios to be a functionof dilution rate, but not total nutrient concentrations inthe medium (which would only affect steady state bio-mass yield). Changes in steady state cell compositionwith dilution rate can be explained mechanistically aschanges in the allocation between the major intracel-lular compartments relating to structure, storage, andbiosynthesis (Shuter 1979, Kooijman 1993). The al-most 40-fold variation in P:C ratio found in our cul-tures, despite the fact that all cultures were run at thesame dilution rate, thus must be attributed to the effectsof light limitation and photoadaptation rather than tochanges in specific growth rate. Furthermore, since theeffects of light level were different between cultureswith high and low total P, we are considering both thedirect effects of treatment-imposed light limitation andthe indirect effects of biomass-induced self-shading.The consistently high P:C ratios at all light levels inthe 50 mmol/L total P treatment, probably indicate thatthese cultures were generally P saturated and light lim-ited (although additional effects of CO2 limitation can-not be ruled out). In all the lower total P treatments(#10 mmol/L), there was probably some degree of co-limitation by both light and phosphorus, and also in-dications of considerable interaction between these twolimiting factors. The interaction effects may partly beinterpreted as biomass-dependent self-shading increas-ing the degree of light limitation, and partly as pho-toadaptation being constrained by P limitation.
Our results clearly support the mutual role of lightand nutrients as determinants of phytoplankton P:C ra-
July 2002 1895ALGAL PHOTOSYNTHESIS AND GRAZER FITNESS
FIG. 7. Growth rates for Daphnia (1–8 d old) raised on a gradient of food quantities and qualities. Each bar representsthe mean 1 1 SD of three replicates measuring five individual growth rates of single Daphnia raised on food at differentcombinations of light and P treatments at four different food concentrations (0.5, 1.0, 1.5, and 5.0 mg C/L). The individualgrowth rates were obtained by the image analysis technique.
FIG. 8. Daphnia growth rate as a function of food quality (P:C ratio) and quantity (mg C/L), fitted to a double hyperbolamodel. The mesh grid is the model prediction surface, while observations are represented by ball-and-stick symbols wherethe stick is the projection of the observation onto the model prediction surface.

1896 DAG O. HESSEN ET AL. Ecology, Vol. 83, No. 7
tios. Under the whole range of light and nutrients ap-plied in these experiments, an almost 40-fold decreasein algal P:C was induced by reducing the total P from50 to ;1 mmol P/L, and by increasing the light from10 to 70 mmol quanta. Urabe and Sterner (1996) sim-ilarly recorded a range in algal P:C from ;1 mg atomicP·[mg atomic C]21 at low P (,1 mmol/L) and high lightto .10 at high P and low light. A corresponding rangeof elemental ratios has been reported in previous ex-periments with P-starved and P-saturated freshwaterchlorophytes in batch cultures (Van Donk and Hessen1993, Van Donk et al. 1997) and continuous cultures(DeMott et al. 1998). The latter authors obtained ‘‘P-sufficient’’ Scenedesmus at 100 mmol P/L, yielding aP:C ratio of 12.5 mg atomic P·[mg atomic C]21. Byreducing the dilution rate from 1.0 d21 to 0.15 d21, andreducing medium P to 6 mmol P/L, P:C ratio over arange of 1.2–1 mg atomic P·[mg atomic C]21 was pro-duced. Lake seston is of heterogeneous origin, of whichdetritus with high P:C frequently make up a consid-erable share (cf. Sterner and Hessen 1994). Thus lakeseston stoichiometry may not necessarily mimic theelemental composition of its algal component. Nev-ertheless, the span of C:P ratios in lake seston corre-sponds well with that achieved in algal cultures (cf.Heckey et al. 1993, Elser and Hassett 1994, Sterner etal. 1997, Hessen and Faafeng 2000).
The range of light and nutrients applied in theseexperiments is well within the limits of natural varia-tion within (light) and between (nutrients) lakes. Whilethe low-light treatment was 14% of full light in ourexperiments, the compensation depths where photo-synthesis balances respiration is commonly assumed tobe 1% of surface radiation. Phytoplankton within a lakemay experience the full range of light exposure bothon a diurnal time scale and on other time scales dueto mixing regimes. The concentration of total phos-phorus (TP) will also be one of the main determinantsof the underwater light climate in lakes, since highlevels of chlorophyll a will effectively attenuate pho-tosynthetic active radiation (PAR). This does not implya covariation between high TP and low light, however,since cell quotas of P may be depleted under situationswith low supply and high phytoplankton biomass.
Algal cells may respond to increased light levels invarious ways, and the decreased P:C could be inter-preted as a decoupling of C and P uptake that wouldbe accentuated by low P (cf. Urabe and Sterner 1996).While the final outcome of an increased light : nutrientratio in terms of increased C:P seems consistent, theresponses in terms of C and P uptake and metabolismmay be more subtle. Apparently the algal cell quota ofC increases under high light, consistent with the theoryof storage of C-rich compounds resulting from ‘‘ex-cess’’ C generated by C fixation at a high rate. The cellquota of P did not, however, show any consistent pat-tern, although for all runs above 1 mmol P/L, the cellP quota tended to decrease under high light. This was
reversed at 50 mmol P/L, where cell quotas of both Cand P increased, yielded uniform (and high) cellularP:C ratios.
An increased cell size in P-limited green algae haspreviously been demonstrated, and seems to be linkedto the storage of excess C as starch granules (van Donket al. 1997). For Selenastrum (NIVA CHL 1, the sameclone as used in these experiments) a doubling in cellvolume occurred upon transition from P saturation toP limitation (van Donk and Hessen 1993, van Donk etal. 1997). This morphological change was supportedby the forward scatter cytograms (representing a com-bined signal of optical density and particle size).
One very intriguing outcome of these experimentswas the apparent dependency of cell-specific chloro-phyll not only on light, but also on ambient P. Judgingfrom the decreasing chlorophyll a : carbon ratio overthe gradient of ambient P, chlorophyll a synthesis mayin fact be directly constrained by the availability of P.Under low light, there is a need for more light-har-vesting machinery, and apparently the synthesis of themembrane-rich chloroplasts could be constrained notonly by N (Prezelin and Matlick 1983), but also by theavailability of P for the phospholipid synthesis. Thismay induce several feedback effects both on total lightharvesting and C fixation that may in turn affect cellbiochemistry and stoichiometry.
One basic question is whether or not such inducedeffects on algal cell nutrient quotas should be regardedas transient or persistent. This may hardly be sortedout from batch culture experiments. The fact that cellquotas and P:C ratios remained stable over a .3-wkrun under all treatments in these experiments suggestthat these cellular responses may be quite persistentunder given scenarios. Data from a wide range of lakesdo also suggest some inherent properties of seston stoi-chiometry accredited to nutrients, light, and mixingregimes across lakes (Sterner et al. 1997).
The relevance of these elemental changes for foodweb production hinges upon the potential for P limi-tation in zooplankton. The cladoceran family Daphniais most relevant in this regard, not only because it hashigh P:C ratios (and thus high specific requirementsfor P), but also because of their prominent position inthe food webs of many lakes. There are, however, nofixed thresholds of P:C for the onset of P limitation inDaphnia. Hessen (1992) estimated a threshold near 8mg atomic P·[mg atomic C]21 while Urabe and Watan-abe (1992) applied a threshold for onset of P limitationat 3 mg atomic P·[mg atomic C]21. Based a set of verythorough experiments, DeMott et al. (1998) found on-set on P limitation at P:C-ratios as high as 11 mg atomicP·[mg atomic C]21. Urabe and Sterner (1996) found astrong negative effect on biomass accumulation in ju-venile Daphnia when P:C decreased from ;4 to 2 mgatomic P·[mg atomic C]21, in accordance with theirproposed threshold of 3 mg atomic P·[mg atomic C]21.These differences in proposed thresholds for onset of

July 2002 1897ALGAL PHOTOSYNTHESIS AND GRAZER FITNESS
P limitation are by no means trivial. A threshold of 11mg atomic P·[mg atomic C]21 would mean that phy-toplankton in a majority of lakes would offer subop-timal quality with respect to P requirements for Daph-nia growth. Hessen and Faafeng (2000) estimated thatDaphnia in 88% out of 110 Norwegian lakes wouldtheoretically be P limited judged from sestonic P:Cratios when applying the high threshold of 11 mg atom-ic P·[mg atomic C]21, but even if the threshold wasreduced to 3 mg atomic P·[mg atomic C]21, some 40%of the Daphnia populations would theoretically face Plimitation. This does not imply that food-quality con-straints are more important than quantity constraints,however. Rather, P limitation may be superimposed onC limitation, providing an additional tax on the netuptake of C by the animals.
The Holling type-II response to food quality wouldindicate no pronounced threshold, but an increasingeffect with decreased P:C ratios in food. Since the graz-ers to some extent may adjust their own somatic de-mands for P relative to C, i.e., they are not strictlyhomeostatic (DeMott et al. 1998), there will be no fixedthreshold that will apply to all situations. The responseof food C:P ratios will depend also on the assimilationof C relative to P. A high assimilation of P relative toC will raise the threshold and vice versa. Finally, thefood quantity itself could influence the potential onsetof P limitation (Sterner and Robinson 1994, Sterner1997). As food quantity in terms of C approaches thethreshold for positive net growth, an increasing shareof the assimilated C will be used for maintenance me-tabolism, and at zero net growth the somatic need forP will approach zero. The model of food quantity vs.food quality limitations (Fig. 8) may thus not be validfor food concentrations below the observed range offood concentrations (500–5000 mg C/L), and particu-larly not when approaching the threshold carbon con-centration for net growth in Daphnia (;50 mg C/L).
The effects of lowered P:C in the diet on Daphniagrowth has been repeatedly demonstrated in laboratoryexperiments (Urabe and Watanabe 1992, Sterner 1993,DeMott et al. 1998) as well as in field studies (Sterneret al. 1997), and the potential for direct P limitationhas been verified experimentally (Urabe et al. 1997).Although alternative explanations for reduced growthof Daphnia on P-deficient food have been advocated,such as deficiencies in the supply of polyunsaturatedfatty acids (Muller-Navarra 1995, Brett and Muller-Navarra 1997), the potential role of these may be over-ruled by strict P deficiency under low P:C ratios in thefood (Sundbom and Vrede 1997, Boersma 2000). Theeffect of increased algal cell-wall thickness and grazingresistance under strong P deficiency has also been dem-onstrated for batch cultures of green algae like Selen-astrum and Chlamydomonas (Van Donk and Hessen1994, Van Donk et al. 1997). The removal of algal cellsfrom cultures by Daphnia grazing was strongly reducedwhen the algae was strongly P limited, simply due to
passage of intact cells through the gut. Correspondingeffects have not been reported in continuous cultures,however (Sterner 1993, DeMott et al. 1998). Our graz-ing experiments over a range of Daphnia ages at dif-ferent P and light treatments indicated neither reducedgrazing rates or gut passage of intact algae. The lowerclearance rates found for animals raised on low P dietreflected simply lower growth rates of these animalscompared with control. This may illustrate the potentialfor overall lower community grazing in P-limited sys-tems due to food quality depression among herbivores.
The study demonstrates a quite subtle regulation ofcellular levels of nutrients and chlorophyll in plank-tonic algae that is related to ambient light and nutrientlevels. While high light causes elevated cell quotas ofC, there is an apparent increased demand for P thatmay be seen as an adaptation to self shading and in-creased cellular levels of chlorophyll. This adaptationwas reflected as increased cell quotas of P with reducedlight. These responses may have profound effects oncellular stoichiometry, nutrient allocation, and carbonsequestration of autotrophs, and these effects in turncan propagate up the food chain. Clearly, a short-termresult of these responses on high-P-demanding grazerscould be reduced herbivore growth rates that in turncould translate into reduced total grazing rates whensestonic P:C ratios are low. This sequence of eventswould typically occur during in the late phase of algalblooms containing high biomasses of nutrient deficientcells. These food quality effects will be superimposedon the food quantity effects, but the relative importanceof these factors over a seasonal cycle is not settled,however. These findings could also have strong impli-cations for our understanding of carbon sequestrationin aquatic ecosystems. They also provide new insights(and questions), not only of the physiological regula-tion of C and P uptake and demands in algae, but alsocould explain systematic deviations from the Redfieldratio, a cornerstone in plankton ecology and aquatic Cmodels.
While these aspects of elemental regulation in au-totrophs and the subsequent effects on grazers havebeen explored primarily in freshwater habitats, we be-lieve that these findings could be of general validity.In particular the elemental mismatch between auto-trophs and herbivores also seems to appear also in ma-rine and terrestrial ecosystems (White 1993, Elser etal. 2000a). Since autotroph production and biomass iscommonly considered to be N limited in these systems,the vegetation and consumer N:C ratios would presum-ably be more important than the P:C ratios studied infreshwater systems. Yet the role of P limitation bothfor autotroph regulation of photosynthesis and successof grazers calls for further scrutiny also in these sys-tems.
LITERATURE CITED
Andersen, T. 1997. Grazers as sources and sinks for nutrients.Ecological Series 129. Springer-Verlag, New York, NewYork, USA.

1898 DAG O. HESSEN ET AL. Ecology, Vol. 83, No. 7
Andersen, T., and D. O. Hessen. 1991. Carbon, nitrogen andphosphorus content of freshwater zooplankton. Limnologyand Oceanography 36:807–814.
Berman-Frank, I., and Z. Dubinsky. 1999. Balanced growthin aquatic plants: myth or reality? BioScience 49:29–37.
Boersma, M. 2000. The nutritional quality of P-limited algaefor Daphnia. Limnology and Oceanography 45:1157–1161.
Brett, M. T., and D. C. Muller-Navarra. 1997. The role ofhighly unsaturated fatty acids in aquatic food-web pro-cesses. Freshwater Biology 38:483–500.
Cleveland, W. S. 1993. Visualizing Data. Hobart, Summit,New Jersey, USA.
DeAngelis, D. L. 1992. Dynamics of nutrient cycling andfood webs. Chapman and Hall, New York, New York, USA.
DeMott, W. R., R. D. Gulati, and K. Siewertsen. 1998. Effectsof phosphorus- deficient diets on the carbon and phospho-rus balance of Daphnia magna. Limnology and Oceanog-raphy 43:1147–1161.
Elser, J. J., W. F. Fagan, R. F. Denno, D. R. Dobberfuhl, A.Folarin, A. Huberty, S. Interlandi, S. S. Kilham, E. Mc-Cauley, K. L. Schulz, E. H. Siemann, and R. W. Sterner.2000a. Nutritional constraints in terrestrial and freshwaterfood webs. Nature 408:578–580.
Elser, J. J., and R. P. Hassett. 1994. A stoichiometric analysisof the zooplankton–phytoplankton interactions in marineand freshwater ecosystems. Nature 370:211–213.
Elser, J. J., R. W. Sterner, E. Gorokhova, W. F. Fagan, T. A.Markow, J. B. Cotner, J. F. Harrison, S. E. Hobbie, G. M.Odell, and L. J. Weider. 2000b. Biological stoichiometryfrom genes to ecosystems. Ecology Letters 3:540–550.
Færøvig, P., T. Andersen, and D. O. Hessen. 2002. Imageanalysis of Daphnia populations: nondestructive determin-ing of demography and biomass in cultures. FreshwaterBiology, in press.
Geider, R. J. 1987. Light and temperature dependence of thecarbon to chlorophyll ratio in microalgae and cyanobac-teria: implications for physiology and growth of phyto-plankton. New Phytologist 106:1–34.
Hecky, R. E., P. Campell, and L. L. Hendzel. 1993. Thestoichiometry of carbon, nitrogen and phosphorus in par-ticulate matter of lakes and oceans. Limnology and Ocean-ography 38:709–724.
Hessen, D. O. 1990. Carbon, nitrogen and phosphorus statusin Daphnia magna at varying food conditions. Journal ofPlankton Research 12:1239–1249.
Hessen, D. O. 1992. Nutrient element limitation of zooplank-ton production. American Naturalist 140:799–814.
Hessen, D. O., and B. A. Faafeng. 2000. Elemental ratios infreshwater seston; implications for community structureand energy transfer in food webs. Archiv fur HydrobiologieSpecial Issues Advances in Limnology 55:349–363.
Kilham, S. S., D. A. Kreeger, S. G. Lynn, C. E. Goulden, andL. Herrera. 1998. COMBO: a defined freshwater culturemedium for algae and zooplankton. Hydrobiology 377:147–159.
Kooijman, S. A. L. M. 1993. Dynamic energy budgets in
biological systems. Cambridge University Press, Cam-bridge, UK.
McNaughton, S. J. 1990. Mineral nutrition and seasonalmovement of African migratory ungulates. Nature 345:613–615.
Muller-Navarra, D. 1995. Evidence that highly unsaturatedfatty acids limits Daphnia growth in nature. Archiv furHydrobiologie 132:297–307.
Prezelin, B. B., and H. A. Matlick. 1983. Nutrient-dependentlow-light adaptation in the dinoflagellate Gonyaulax poly-hedra. Marine Biology 74:141–150.
Raven, J. A. 1984. A cost–benefit analysis of photon ab-sorption by photosynthetic unicells. New Phytologist 98:593–625.
Rhee, G.-Y., and I. J. Gotham. 1980. Optimum N:P ratiosand coexistence of planktonic algae. Journal of Phycology16:468–489.
Shapiro, H. M. 1995. Practical flow cytometry. Third edition.Wiley, New York, New York, USA.
Shuter, B. 1979. A model of physiological adaptation in uni-cellular algae. Journal of Theoretical Biology 78:519–552.
Sterner, R. W. 1993. Daphnia growth on varying quality ofScenedesmus: mineral limitation of zooplankton. Ecology74:2351–2360.
Sterner, R. W. 1997. Modeling interactions of food qualityand quantity in homeostatic consumers. Freshwater Biol-ogy 38:473–481.
Sterner, R. W., J. J. Elser, E. J. Fee, S. J. Guilford, and T. H.Chrzanowski. 1997. The light : nutrient ratio in lakes: thebalance of energy, and materials affect ecosystems struc-ture, and process. American Naturalist 150:663–684.
Sterner, R. W., and D. O. Hessen. 1994. Algal nutrient lim-itation and the nutrition of aquatic herbivores. Annual Re-view of Ecology and Systematics 25:1–29.
Sterner, R. W., and J. Robinson. 1994. Thresholds for growthin Daphnia magna with high and low phosphorus diets.Limnology and Oceanography 39:1228–1232.
Sundbom, M., and T. Vrede. 1997. Effects of fatty acid andphosphorus content of food on the growth, survival andreproduction of Daphnia. Freshwater Biology 38:665–674.
Urabe, J., J. Clasen, and R. W. Sterner. 1997. Phosphorus-limitation of Daphnia growth: is it real? Limnology andOceanography 42:1436–1443.
Urabe, J., and R. W. Sterner. 1996. Regulation of herbivoregrowth by the balance of light and nutrients. Proceedingsof the National Academy of Science (USA) 93:8465–8469.
Urabe, J., and Y. Watanabe. 1992. Possibility of N and Plimitation for planktonic cladocerans: an experimental test.Limnology and Oceanography 37:244–251.
Van Donk, E., and D. O. Hessen. 1993. Grazing resistancein nutrient stressed phytoplankton. Oecologia 93:508–511.
Van Donk, E., M. Lurling, D. O. Hessen, and C. Lokhorst.1997. Changed cell wall morphology and reduced grazervulnerability in nutrient deficient phytoplankton. Limnol-ogy and Oceanography 42:357–364.
White, T. C. R. 1993. The inadequate environment: nitrogenand the abundance of animals. Springer-Verlag, New York,New York, USA.





























