Embed Size (px)
Citation preview

Developmental Brain Research, 11 (1983) 93-106 93 Elsevier
Light Experience During Development Affects Asymmetry of Forebrain Function in Chickens
J. V. ZAPPIA and L. J. ROGERS
Department of Pharmacology, Monash University, Wellington Road, Clayton 3168, Victoria (Australia)
(Accepted June 30th, 1983)
Key words: functional lateralization - - chicken forebrain - - glutamate - - attack and copulation - - light exposure - - embryonic development
The chicken brain has asymmetry of forebrain function for attack and copulation behaviors. This study reports that this asymmetry, revealed by intracranial injections of glutamate, is affected by light exposure. Eggs were incubated in 3 forced-draught incubators matched for noise, humidity and temperature control, so that light and laboratory noise were the only systematic variables. All eggs were incubated in darkness for the first 17 days of incubation after which they were allocated to one of 3 incubating conditions, L÷S +, L-S +, L÷S -, where + or - - indicates the presence or absence of light (L) or laboratory noise (S). Light exposure was necessary to es- tablish lateralization of attack and copulation, sound was not since only the L÷S + and L÷S - groups were lateralized. The lateralization of auditory habituation was not affected by these incubating conditions.
Further experiments established that on day 19 of incubation exposure for 4 h to a light of constant intensity produced lateraliza- tion. 1 h exposure to a light of constant intensity or to a flickering light (1.7 Hz) failed to produce lateralization.
Examination of the data for individuals suggests that light exposure synchronizes the direction of lateralization rather than gener- ates its presence. Environmental and genetic factors interact to produce the lateralization of attack and copulation.
INTRODUCTION
It is becoming increasingly evident that lateraliza-
tion of brain function, once thought to be unique to
humans, is present in a number of non-human spe-
cies, both mammal ian 15 and avian31, 33. Rogers and
colleagues 21.35 have demons t ra ted asymmetry of
forebrain function in chickens by injecting a single
dose of cycioheximide, a r ibosomal prote in synthesis
inhibitor, or monosod ium glutamate (MSG) , a puta-
tive neurot ransmit ter , into ei ther the right or left
forebrain hemisphere on day 2 post-hatch. Behavio-
ral testing during the second week post-hatch re-
vealed that chicks t rea ted in the left hemisphere have
re tarded visual and audi tory learning and elevated
attack and copula tory responses while t rea tment of
the right hemisphere did not affect these behaviors.
Adminis t ra t ion of cycloheximide or MSG after the
first week of life no longer effects these behaviors.
The mechanisms underlying the effects of these drugs
are discussed elsewhere 37. While unilateral adminis-
trat ion of MSG or cycloheximide can be used as a
useful tool to reveal la teral izat ion, lateral izat ion can
"also be demons t ra ted in unt rea ted chicks tested mo-
nocularly (Rogers , Bullock and Zappia , in prepara-
tion).
The ontogeny of lateral izat ion in the chicken, or in
any other species including humans, is not clear.
Many researchers speculate that it is p redominant ly
inheri ted, ei ther genetically 25 or cytoplasmically12, 27.
Others argue that la teral izat ion results from the in-
fluence of both genetic and environmental fac-
tors4,10, the lat ter being brought into play since the
organism lives in a biased ( lateral ized) world. Rogers
and Anson35 proposed an ontogenet ic mechanism
which may account for the lateral izat ion observed in
chickens. They suggested that unilateral sensory in-
put during incubation may st imulate deve lopment of
the forebrain hemispheres differentially, and that
this subsequently results in the lateral izat ion of func-
tion observed post-hatch. This hypothesis is based on
a knowledge of the anatomical deve lopment of the
chicken embryo and its or ienta t ion in the egg which
enables unilateral sensory st imulat ion to occur.
Throughout embryonic deve lopment the left eye and
ear of the chick are shielded ei ther by the yolk sac or
0165-3806/83/$03.00 © 1983 Elsevier Science Publishers B.V.

94
the chicken's body Is. During the first 14 days of incu- bation the chick lies on its left side on the yolk sac with its right eye facing outwards to the shell. While
the embryo is migrating in the egg (day 15 to 16) the yolk comes to cover the entire head. During the last 3
days of incubation, when the yolk sac has retreated from the head, the head is turned against the left side of the body 32 such that the left eye and ear are oc-
cluded leaving the right eye and ear better exposed to receive perceptual inputs through the shell and air
sac. The position of the head during the last 3 days of incubation is likely to be critical in terms of environ-
mental influences on the brain since the peripheral visual system becomes functionally connected to the forebrain at this time 18~4~. Rogers and Anson 35 pre- dict that light and sound reaching the left eye and ear
would be attenuated relative to the right eye and ear. In fact, Lauber and Shutze 24 have reported data
which indicates that light exposure during incubation increases the size of the right eye relative to the left. Avian visual pathways decussate completely 13 and
most information entering one eye is processed by the contralateral forebrain hemisphere. The fore- brain hemispheres of the embryonic chick should therefore receive differential stimulation. This asym- metry of stimulation would be expected to increase
blood flow in the left, stimulated hemisphere, as BondyS,7 has demonstrated for the contralateral hemisphere in monocutarly-trained chickens, and may therefore increase protein synthesis in the left hemisphere 3.38. Such events could lead to the devel- opment of the left hemisphere in advance of the
right. If this differential stimulation is responsible for the
lateralization of function observed in chickens, chicks incubated in darkness to eliminate asymmet- rical visual stimulation would not be expected to show lateralization. In fact, Rogers 34 has observed lateralization for attack and copulation in chickens hatched from eggs that received light during incuba- tion but not for those incubated in complete dark- ness. As little as 2-3 h of light input just prior to pip- ping (i.e. on day 19 of incubation) was found to be sufficient to cause elevated attack and copulation in groups of chicks treated with MSG in the left hemi- sphere but not in those treated in the right.
Rogers 34 noticed that the non-lateralized groups exhibited greater variation in attack and copulation
scores than did chicks in the lateralized groups. This
suggested that the groups which did not display lat- eralization may still have contained individuals which
had hemispheric asymmetry, but half of these indi- viduals were asymmetrical in one direction and half in the other direction leaving no overall group bias, or lateralization. Light exposure therefore appears to synchronize the direction of laieralization such
that most individuals in the population are lateralized in the same direction. This highlights the importance
of distinguishing between population and individual lateralization ~5.
There is a critical period when light exposure re-
sults in population lateralization. This coincides with the emergence of a functional visual system, around day 18 or 19 of incubation 4°. Light exposure prior to
this time fails to synchronize lateralization. However, while Rogers' study 34 supports the origi-
nal hypothesis it is important to realize that tactile, vestibular, proprioceptive and auditory systems as well as the visual system begin to function during in- cubation 18,41. It is therefore possible that all these
systems may interact with the outside world and that several of these systems may synergize to produce the effect described by Rogers 34. In the experiments to be described here particular attention is paid to the auditory and visual systems in order to delineate how inputs in these modalities influence lateralization of attack and copulation and, in addition, lateralization of auditory habituation. Moreover, the nature of the visual input is manipulated and its effect on the lat- eralization of attack and copulation assessed.
METHODS
Animals and housing Eggs, from australorp-leghorn cross hens, were ob-
tained from a local poultry farm and incubated under conditions described below. After hatching the chicks were housed in groups of 4-6 animals for the first 4 days and kept thereafter in individual cages 36. Starter crumbs (Baristoc, Victoria) and water were available ad libitum. Constant warmth (25 °C) and light were provided by overhead bulbs.
Drug administration The drugs were administered to conscious animals
on day 2 post-hatch. Injections were performed free-

hand using sterile 25-gauge syringes which were
fitted with stops to ensure that the needles pen-
etrated the cranium at a depth o f n o more than 2.5
mm. The injection sites were midway between the
anterior and posterior extremities of each forebrain
hemisphere and ranged between 1 and 5 mm on ei-
ther side of the midline.
Chicks received either glutamate to the left hemi-
sphere and saline to the right (this group will be re-
ferred to as left-treated group), or glutamate to the
right hemisphere and saline to the left (referred to as
the right-treated group). The monosodium salt of L-
glutamic acid (British Drug House) was dissolved in
sterile pyrogen-free water, and a dose of 5/A of 100
mM glutamate per hemisphere was used. Saline in-
jections consisted of 5 fll of 0.9% saline in sterile, py-
rogen-free water.
Behavioral testing
Attack and copulation Standard hand thrust tests developed by Andrew1.2
and modified by Young and Rogers 46 were used to
measure juvenile attack and copulatory behaviors.
Attack behavior was measured by the response ob- served to a hand held facing the chicken with the fin-
gers arched over towards the beak moved rapidly
back and forth 10 times. The responses were assessed
95
using a rank scoring scale ~vhich ranged from an avert
gaze (zero score) through to pecking and attack leap-
ing at the hand (maximum score of 10) (see Fig. 1).
Copulation was elicited by a hand held horizontal-
ly with the palm facing downward and thrust gently
and rapidly 10 times at the chick's chest, and then
held still at a level which allowed the chick to step
onto it with ease. The copulation response (Fig. 2) was rated according to a ranked scoring scale. The
responses ranged from walking over the hand (no
score) to a full crouch with pecking treading and pel-
vic thrusting, which attained the maximum score of
10.
Each test was repeated 3 times daily and a mean
calculated for each day. Inter-rater reliability co-effi-
cients were calculated between two trained observ-
ers. The two observers were found to be in
agreement for 91% of 144 incidences of attack and
94% of 144 incidences of copulation.
Auditory habituation Auditory habituation was tested according to pro-
cedures used by Rogers et al. 36 and Howard et al. 21.
Chicks were deprived of food for 4 h and then allow-
ed to feed for 1 min before presenting an auditory
stimulus, produced by striking once a metal disc 15
cm in diameter with a metal spoon. The response in-
volved ceasing to feed and orienting to the novel
Fig. 1. Sample of responses in the hand-thrust tests for attack. In A the chicken has turned away from the thrusting hand and would score 0. In B the chicken is actively sparring and showing an intention to attack and leap; it would score 7. This test is repeated 3 times consecutively and a mean daily score is obtained.
96
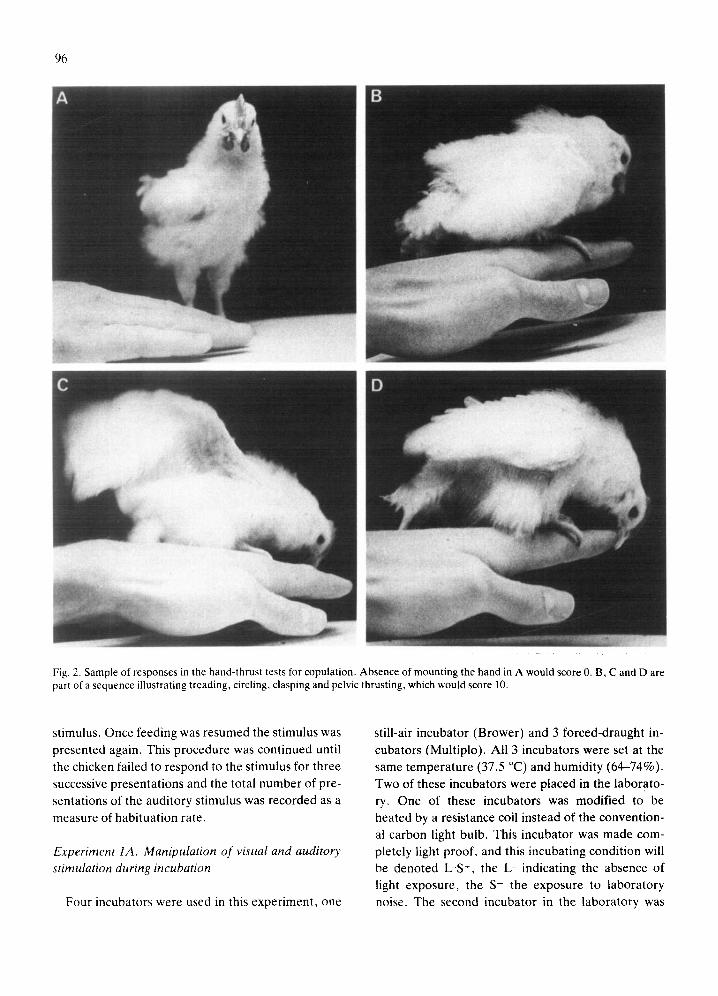
Fig. 2. Sample of responses in the hand-thrust tests for copulation. Absence of mounting the hand in A would score 0. B, C and D are part of a sequence illustrating treading, circling, clasping and pelvic thrusting, which would score 10.
stimulus. Once feeding was resumed the stimulus was
presented again. This procedure was cont inued until
the chicken failed to respond to the stimulus for three
successive presentat ions and the total number of pre-
sentations of the audi tory stimulus was recorded as a
measure of habi tuat ion rate.
Experiment 1A. Manipulation of visual and auditory stimulation during incubation
Four incubators were used in this exper iment , one
still-air incubator (Brower) and 3 forced-draught in-
cubators (Multiplo). Al l 3 incubators were set at the
same tempera ture (37.5 °C) and humidi ty (64-74%).
Two of these incubators were placed in the laborato-
ry. One of these incubators was modif ied to be
heated by a resistance coil instead of the convention-
al carbon light bulb. This incubator was made com-
pletely light proof , and this incubating condit ion will
be denoted L-S +, the L- indicating the absence of
light exposure, the S ÷ the exposure to labora tory
noise. The second incubator in the labora tory was

heated by a carbon light bulb and received external
light input through the incubator's observation win-
dows; eggs in it therefore received both light and sound exposure, this condition is denoted L+S ÷. The third forced-draught incubator was also heated by a
light bulb and received external light input through observation windows. It was located in a sound-proof room and not exposed to laboratory noises, this con- dition is denoted L+S -, the S- indicating lack of expo-
sure to laboratory noise, although constant noise from the incubator fan was still present.
The overall intensity of illumination, at the level of
the eggs, in conditions L+S + and L+S - was free to vary but remained within the range of 100-500 lux (Weston photometer).
The intensity of the noise generated by the motor- ized fan was measured by a IE 10A Audio Spectrum
Analyzer (IVIE Electronics) and was found to be the same for all forced-draught incubators. The greatest intensity was 80 dB at the lower frequencies (32-63 Hz) and this steadily waned to 40 dB at the higher fre- quencies (8-16 kHz). The ambient intensity of labo- ratory noise ranged for 40-100 dB, and consisted of voices, vocalizations from other chickens, door bang- ing, etc. These external noises increased the sound intensity within the incubator despite the background noise of the incubator fans. Chicks calls could in- crease the peak sound pressure level within the incu- bator by 10 dB at 4 kHz but more importantly loud talking increased the level by an average of 20 dB over the frequency range of 250 Hz-4 kHz.
For the first 17 days of incubation all eggs were in- cubated at 37.5 (+ 1) °C in a light-tight, still-air incu- bator located in a darkroom. All eggs were briefly ex- posed to light during handling on day 8. On day 17 the eggs were randomly allocated to one of the 3 in- cubating conditions L+S ÷, L+S - or L-S +. The eggs al- located to L-S + were transported in a light-tight con- tainer and were placed inside the incubator without being exposed to light. These incubators were main- tained at 37.5 (+ 0.3) °C and at a relative humidity of 64--74%. None of the eggs were turned after day 17. The dark incubator (L-S ÷) was not opened until the chicks had hatched.
Chicks were removed from these incubators 4-6 h after hatching and housed in conditions described above. The groups of chicks were of mixed sex and no attempt was made to sex them since attack and co-
97
pulatory behaviors are lateralized in both males and
females 34. There were 8-11 chicks in each treatment group.
Testing of attack and copulation commenced on day 6 post-hatch and continued until day 13. Audito- ry habituation was tested on day 14.
Experiment lB. Auditory habituation after incubation in silence
This experiment was designed to assess whether external auditory input of any kind during incubation was necessary to establish the lateralization of audi-
tory habituation since the eggs in Experiment 1A were exposed to the sound of the incubator fans from day 17 to hatching. Throughout hatching eggs in Ex- periment 1B were incubated in complete darkness in two silent, still-air incubators (Brower) located in a
sound-proof room. After hatching the chicks were housed and treated with MSG in the manner de- scribed above. Auditory habituation was tested on day 12. Attack and copulation were not tested. There were 12 chicks per treatment group.
Experiment2. Manipulation of the nature of the visual input
(A) Exposure to 4 h of constant illumination The light exposure in the L + groups in Experiment
1A was of varying intensity since some of the light was supplied intermittently by the heating bulbs. Moreover the intensity of the extraneous light which could enter the incubators via observation windows was also free to vary. Therefore an experiment was designed to see whether exposure to a light of con- stant intensity results in population lateralization.
Eggs were exposed to light of constant intensity for either 1 or 4 h on day 19 of incubation. The eggs were kept in still-air incubators throughout incubation ex- cept on day 19 of incubation, at which time they were randomly allocated to one of the two treatment con- ditions of light exposure which entailed exposure to a light source of the same constant intensity. For this purpose the modified forced-draught incubator in the conditions L-S + of Experiment 1A was further mod- ified; a 6 W fluorescent lamp was attached to the in- side roof such that the eggs were 12 cm below the light source. The eggs were placed in the centre of the tray with their air sacs upwards, so that the right

98
eye of the embryo was directly exposed to light enter-
ing the egg. The luminosity at the level of the eggs was measured with a Weston photometer . Each egg
received light exposure of a constant intensity; the
amount falling on the flat recording disc placed at the
level of the egg ranged from 800 lux directly under
the light source to 500 lux for the eggs furthest away
from the light source. Eggs in one condition were ex-
posed to constant light intensity for 1 h (CL-1), and
those in the second condition were exposed for 4 h
(CL-4). MSG treatment and testing for attack and
copulation proceeded in the same manner as de-
scribed above. There were 10-12 chicks in each treat-
ment group.
(B) Exposure to 1 h flickering light It has been shown previously that flashing (on-off)
or flickering lights are more effective imprinting
stimuli than are continuously on lights of constant in- tensity 17. Flashing lights also have different biochem-
ical effects. Rose et al. 38 demonstrated that a flashing
light leads to a more rapid increase in R N A synthesis
in the forebrain roof and midbrain of neonate chicks than does a light of constant intensity.
Therefore there is a distinct possibility that a peri- od of light exposure previously shown to be ineffec-
tive to cause lateralization, say, 1 h, might be made
more effective by using an on-off flashing visual stim-
ulus. In the following experiment eggs incubated in
complete darkness for the first 17 days of incubation
were then exposed for 1 h to either a light of constant
intensity as in Experiment 2A (CL-1) or a light flick-
ering at 1.7 Hz (FL-1). Embryonic visual experience at frequencies between 1.16 Hz and 3.50 Hz have been shown to be maximally effective in eliciting ap- proach behavior 17. After hatching these chicks were
treated with MSG and tested for attack and copula-
tion as described above. There were between 19 and 22 chicks in each treatment group.
Statistical analysis If attack and copulation are lateralized at the pop-
ulation level, a significant difference should be ob-
served between the group means of left- and right- treated animals. This comparison was assessed by an analysis of variance taking into account the repeated measurement nature of the experimental design (i.e. across all testing days). The Geisser-Greenhouse
correction 22 was used to evaluate comparisons which involved the repeated measure.
In addition, the data from day 13 was analyzed by
the Levene test for variance effects 22 to test for ho-
mogeneity of variance. A square-root transfor-
mation was performed on attack and copulation
scores to minimize heterogeneity of variance. The
transformed data was analyzed by analysis of vari-
ance with unequal sample sizes. Simple main effects
and Newman-Keuls procedures were employed for multiple comparisons 22.
RESULTS
Experiment 1. Manipulation of auditory and visual exposure
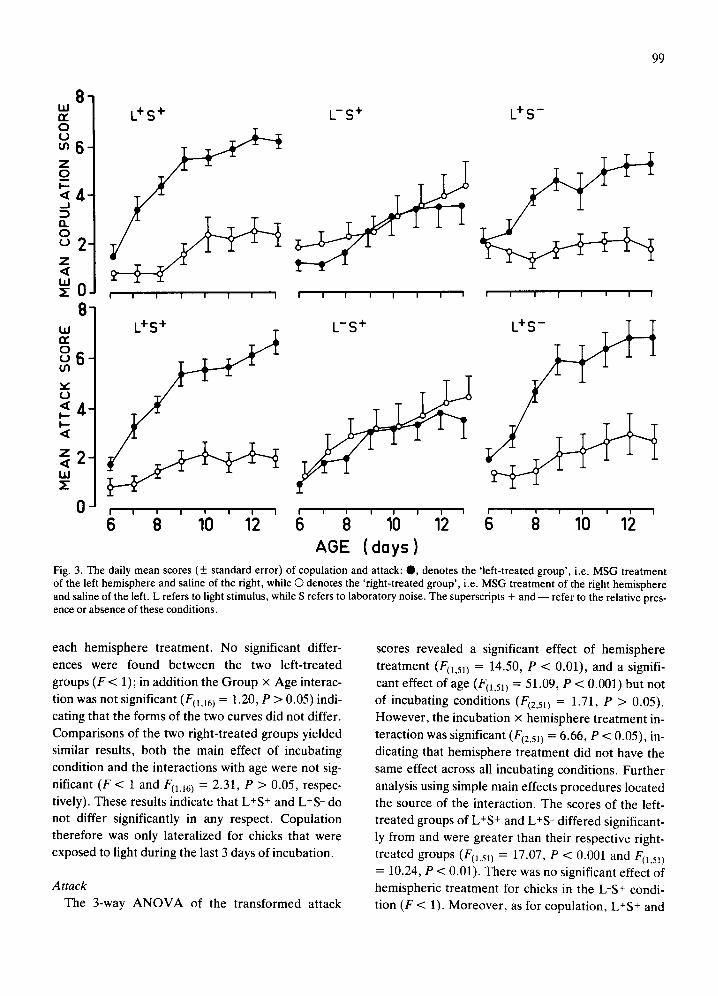
The daily mean values of copulation and attack scores over the 8 testing days are presented in Fig. 3.
To assess whether individuals responded consistently on repetitions of the same test, p roduct -moment cor-
relations were calculated between individual scores
on day 12 and day 13 for each of the treatment condi- tions. There was a significant correlation for attack
scores (the correlation coefficients ranged from 0.72 to 0.98; P < 0.05), and also for copulation (coeffi-
cients ranged from 0.63 to 0.94; P < 0.05). These re- sults indicate that individuals responded consistently
on repetitions of the same test.
Copulation The 3-way A N O V A of the transformed copulation
scores revealed a significant effect of hemisphere
treatment (F(1.51) = 39.40, P < 0.001), but not for the incubating conditions (F < 1). However, their interac-
tion was significant (F(2,511 = 13.44, P < 0.001). There was also a significant effect of age, or day of
testing (Fll,511 = 60.10, P < 0.001). Further analysis using simple main effects procedures 22 revealed that
the left-treated groups differed significantly from the right-treated groups for conditions L-S + and L+S-, scores for the left-treated groups being greater than those of right-treated groups. Thus copulation was
lateralized for groups L -S + and L+S - (F~t,sll = 32.08, P < 0.001 and FIl,51) = 33.86, P < 0.001) but not for L-S + ( F < 1).
Analyses of variance with repeated measures across age were used to compare L+S + and L+S - for
99
e LIJ n, 0 tJ ~6 z o ..J
0. o c) 2 - Z
W ~ 0
8- W
0 ~ 6 'v"
< 4 I,,- I--
Z 2 - ,< w I [
0
L + S +
I I I I I I I I
L- S + L+S -
I I I I I I I I I I I I I I I !
L-S+ L + ~
' 8 ' l b ' 6 ' 8 ' lb ' 1 2 ' 6 ' 8 ' lb ' AGE (days)
Fig. 3. The daily mean scores (+ standard error) of copulation and attack: O, denotes the 'left-treated group', i.e. MSG treatment of the left hemisphere and saline of the right, while O denotes the 'right-treated group', i.e. MSG treatment of the right hemisphere and saline of the left. L refers to light stimulus, while S refers to laboratory noise. The superscripts + and - - refer to the relative pres- ence or absence of these conditions.
each hemisphere treatment. No significant differ-
ences were found between the two left-treated
groups (F < 1); in addition the Group x Age interac-
tion was not significant (F(1,16) = 1.20, P > 0.05) indi- cating that the forms of the two curves did not differ.
Comparisons of the two right-treated groups yielded similar results, both the main effect of incubating
condition and the interactions with age were not sig-
nificant (F < 1 and F(1.16) = 2.31, P > 0.05, respec- tively). These results indicate that L+S ÷ and L+S - do not differ significantly in any respect. Copulation therefore was only lateralized for chicks that were exposed to light during the last 3 days of incubation.
Attack The 3-way A N O V A of the transformed attack
scores revealed a significant effect of hemisphere
treatment (F(1,5]) = 14.50, P < 0.01), and a signifi-
cant effect of age (F(1,51) = 51.09, P < 0.001) but not
of incubating conditions (/7(2,51) = 1.71, P > 0.05). However, the incubation x hemisphere treatment in-
teraction was significant (F(2,51) = 6.66, P < 0.05), in- dicating that hemisphere treatment did not have the
same effect across all incubating conditions. Further
analysis using simple main effects procedures located
the source of the interaction. The scores of the left- treated groups of L+S + and L+S - differed significant- ly from and were greater than their respective right-
treated groups (F(1,5]) = 17.07, P < 0.001 and F(1,5~) = 10.24, P < 0.01). There was no significant effect of hemispheric treatment for chicks in the L-S + condi-
tion (F < 1). Moreover, as for copulation, L+S + and
1 U U
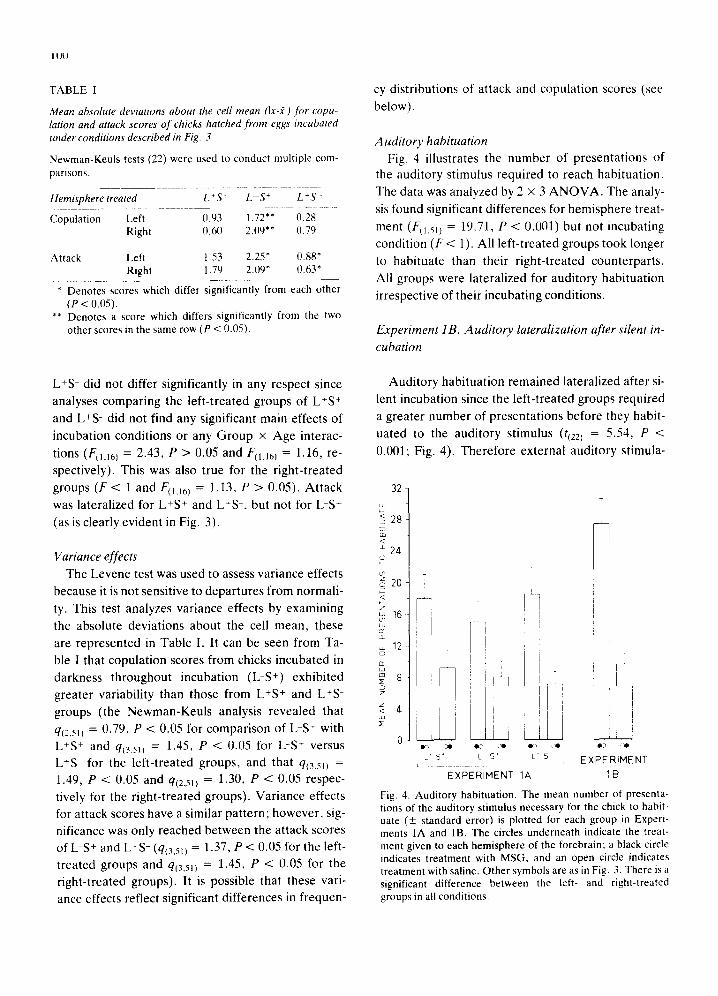
TABLE I
Mean absolute deviations about the cell mean (Ix-,fl) for copu- lation and attack scores of chicks hatched from eggs incubated under conditions described in Fig. 3
Newman-Keuls tests (22) were used to conduct multiple com- parisons.
Hemisphere treated L * S + L-S + L *,7-
Copulation Left (I.93 1.72"* 0,28 Right (t,60 2.09** 0.79
Attack Left t.53 2.25* (1.88" Right 1,79 2.[)9* 0.63*
* Denotes scores which differ significantly from each other ( P < 0.05).
** Denotes a score which differs significantly from the two other scores in the same row (P < 0.05).
cy d is t r ibut ions of at tack and copula t ion scores (see
below).
Audi tory habituation
Fig. 4 i l lustrates the n u m b e r of p resen ta t ions of
the audi tory s t imulus requi red to reach habi tua t ion .
The data was analyzed by 2 × 3 A N O V A . The analy-
sis found significant differences for hemisphere treat-
men t (FII,5 n = 19.71, P < 0.001) bu t not incuba t ing
condi t ion ( F < 1). All lef t - t reated groups took longer
to habi tua te than their r ight - t rea ted counterpar t s .
All groups were la teral ized for audi tory hab i tua t ion
irrespective of their i ncuba t ing condi t ions .
Experiment lB . Audi tory lateralization after silent in-
cubation
L + S - did not differ significantly in any respect since
analyses compar ing the lef t - t reated groups of L+S +
and L+S - did no t find any signif icant ma in effects of
incuba t ion condi t ions or any G r o u p x Age interac-
t ions (F(1,161 = 2.43, P > 0.05 and F( l ,16) = 1.16, re-
spectively). This was also true for the r ight - t rea ted
groups (F < 1 and F(H6) = 1.13, P > 0.05). At tack
was lateral ized for L+S + and L+S -, bu t not for L-S +
(as is clearly ev ident in Fig. 3).
Variance effects
The Levene test was used to assess var iance effects
because it is no t sensi t ive to depar tu res f rom normal i -
ty. This test analyzes var iance effects by examin ing
the absolute devia t ions abou t the cell me an , these
are represen ted in Tab le I. It can be seen from Ta-
ble I that copula t ion scores f rom chicks incuba ted in
darkness th roughou t incuba t ion (L-S +) exhibi ted
greater variabil i ty than those f rom L+S ÷ and L+S -
groups (the N e w m a n - K e u l s analysis revealed that
q(2.51/ = 0.79, P < 0.05 for compar i son of L-S + with
L+S + and qi3,sn = 1.45, P < 0.05 for L-S + versus
L+S - for the lef t - t reated groups, and that q(3511 =
1.49, P < 0.05 and qI2,sn = 1.30, P < 0.05 respec-
t ively for the r ight- t reated groups) . Var iance effects
for at tack scores have a similar pa t t e rn ; however , sig-
nificance was only reached be tween the at tack scores
of L-S + and L+S - (q(35~) = 1.37, P < 0.05 for the left-
t rea ted groups and q(3.5n = 1.45, P < 0.05 for the
r ight- t reated groups) . It is possible that these vari-
ance effects reflect significant differences in f r equen-
Aud i to ry hab i tua t ion r e m a i n e d la teral ized after si-
lent incuba t ion since the lef t - t reated groups requi red
a greater n u m b e r of p resen ta t ions before they habit-
uated to the audi tory s t imulus (t(221 = 5.54, P <
0.001; Fig. 4). There fo re externa l audi tory s t imula-
32
28
24.
20
I
12 ~ F I
8
0
I i
, i !I i
L'S + L S ~ L'5
ExPERiMENT IA
T
[
L EXPERIMENT
1B
Fig. 4. Auditory habituation. The mean number of presenta- tions of the auditory stimulus necessary for the chick to habit- uate (+ standard error) is plotted for each group in Experi- ments 1A and lB. The circles underneath indicate the treat- ment given to each hemisphere of the forebrain; a black circle indicates treatment with MSG, and an open circle indicates treatment with saline. Other symbols are as in Fig. 3. There is a significant difference between the left- and right-treated groups in all conditions.
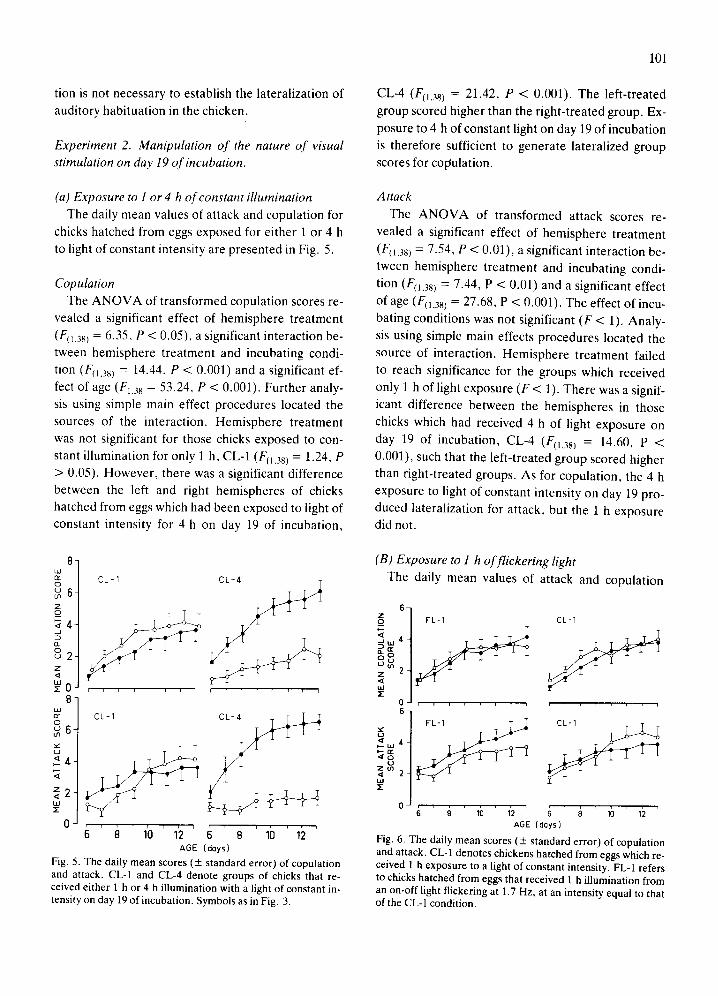
tion is not necessary to establish the lateralization of
auditory habituation in the chicken.
Experiment 2. Manipulation of the nature of visual stimulation on day 19 of incubation.
(a) Exposure to 1 or 4 h of constant illumination The daily mean values of attack and copulation for
chicks hatched from eggs exposed for either 1 or 4 h
to light of constant intensity are presented in Fig. 5.
Copulation
The A N O V A of transformed copulation scores re- vealed a significant effect of hemisphere treatment
(F(~.38) = 6.35, P < 0.05), a significant interaction be- tween hemisphere treatment and incubating condi-
tion (F(L38) = 14.44, P < 0.001) and a significant ef-
fect of age (FL38 = 53.24, P < 0.001). Further analy- sis using simple main effect procedures located the
sources of the interaction. Hemisphere treatment
was not significant for those chicks exposed to con- stant illumination for only 1 h, CL-1 (F(1.38) = 1.24, P
> 0.05). However, there was a significant difference
between the left and right hemispheres of chicks hatched from eggs which had been exposed to light of constant intensity for 4 h on day 19 of incubation,
8 - w
o
~6- z o_
£L 0 U 2 "
z
~o 8
o ~6 ~2
~4- <
UA IE
O-
CL-1 CL-4
i i r i i p i p i T r , i , ~ i
CL-1
p i r r i i ~ ~ i , J i , r , ~
6 8 10 12 6 8 10 12 AGE (days)
Fig. 5. The daily mean scores (+ standard error) of copulation and attack. CL-1 and CL-4 denote groups of chicks that re- ceived either 1 h or 4 h illumination with a light of constant in- tensity on day 19 of incubation. Symbols as in Fig. 3.
101
CL-4 (F(t.38) = 21.42, P < 0.001). The left-treated group scored higher than the right-treated group. Ex-
posure to 4 h of constant light on day 19 of incubation is therefore sufficient to generate lateralized group scores for copulation.
Attack
The A N O V A of transformed attack scores re-
vealed a significant effect of hemisphere treatment
(F/L3~) = 7.54, P < 0.01), a significant interaction be- tween hemisphere treatment and incubating condi-
tion (F{1,38) = 7.44, P < 0.01) and a significant effect
of age (F(1,38) = 27.68, P < 0.001). The effect of incu- bating conditions was not significant (F < 1). Analy-
sis using simple main effects procedures located the
source of interaction. Hemisphere treatment failed
to reach significance for the groups which received
only 1 h of light exposure (F < 1). There was a signif-
icant difference between the hemispheres in those
chicks which had received 4 h of light exposure on
day 19 of incubation, CL-4 (F(1,38) = 14.60, P <
0.001), such that the left-treated group scored higher than right-treated groups. As for copulation, the 4 h
exposure to light of constant intensity on day 19 pro-
duced lateralization for attack, but the 1 h exposure did not.
(B) Exposure to 1 h o f flickering light
The daily mean values of attack and copulation
'] z FL-I CL-1
~ 2
- CL -1
8 . . . . . . , ,
AGE (days)
Fig. 6. The daily mean scores (+ standard error) of copulation and attack. CL-1 denotes chickens hatched from eggs which re- ceived 1 h exposure to a light of constant intensity. FL-1 refers to chicks hatched from eggs that received 1 h illumination from an on-off light flickering at 1.7 Hz, at an intensity equal to that of the CL-1 condition.
102
2O
li I LATER~LIZE~ (n=51) 12
20 LAIERALIZED 16 CO {n=52)
1
0
m 12 Go (n=56)
l i ]~[ ~ 12 co NON-LATERALIZED (n: 57)
~1 ATTACK SCORE ON DAY 1t POSTHATCH
20 li1 2 L'TE''L'ZED'n:'I'
_ = _ L - - ]
24
, z , r - - n = = = _ _ = = _ _ _ _
O
c ~ ~ c n c n c n ¢ ~ o ~ o ~ o ~ o ~
I , _ i i i J i
COPULATION SCORE ON DAY 11 POSTHATCH
scores for chicks from eggs which received either lh of constant light exposure, CL-1, or 1 h of flickering
light on day 19 of incubation, are presented in Fig. 6.
The analysis of transformed copulation scores did not
reveal any significant effect of hemisphere treat-
ment, incubating condition or any interaction be-
tween the two (F < 1 in all cases), although a signifi-
cant effect of age was present (F(l.761 = 84.38, P <
0.001). The analysis of transformed attack scores did not reveal any significant effects of hemisphere treat-
ment (F < 1), incubating condition or their interac-
tion (F < 1 in all cases), although a significant effect
of age was present (F/l,761 = 72.29, P < 0.001). Thus
both exposure to a flickering light of 1.7 Hz for 1 h on day 19 of incubation proved insufficient to estab-
lish the lateralization of attack and copulation.
Distribution of scores for lateralized and non-later- alized populations
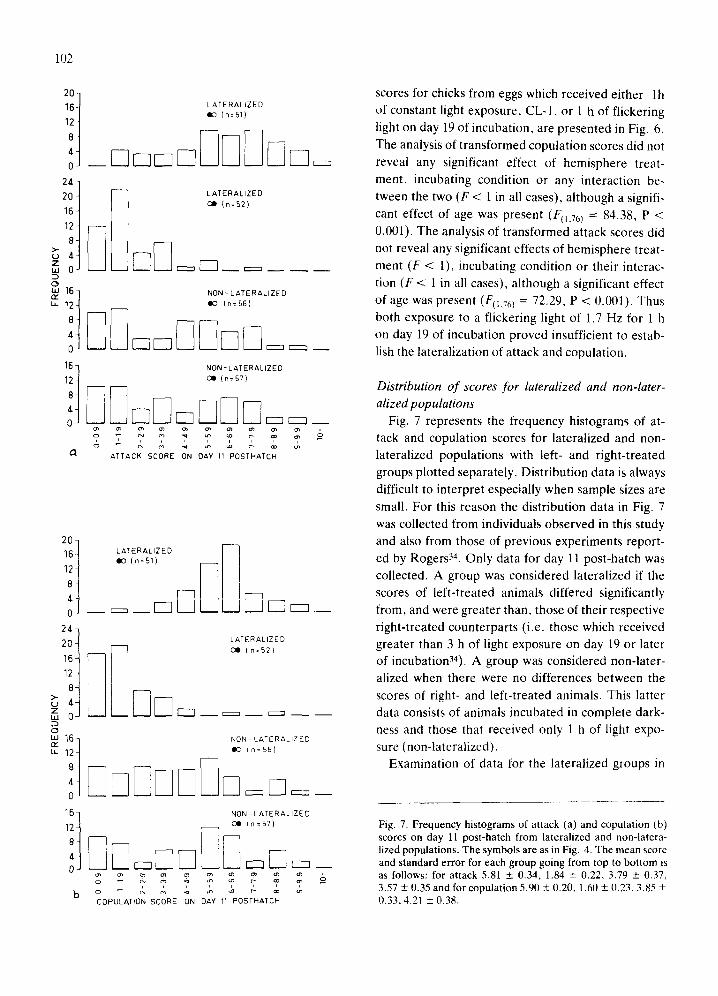
Fig. 7 represents the frequency histograms of at- tack and copulation scores for lateralized and non-
lateralized populations with left- and right-treated
groups plotted separately. Distribution data is always difficult to interpret especially when sample sizes are
small. For this reason the distribution data in Fig. 7
was collected from individuals observed in this study
and also from those of previous experiments report- ed by Rogers 34. Only data for day 11 post-hatch was
collected. A group was considered lateralized if the
scores of left-treated animals differed significantly
from, and were greater than, those of their respective
right-treated counterparts (i.e. those which received greater than 3 h of light exposure on day 19 or later of incubation34). A group was considered non-later-
alized when there were no differences between the
scores of right- and left-treated animals. This latter data consists of animals incubated in complete dark- ness and those that received only 1 h of light expo-
sure (non-lateralized). Examination of data for the lateralized groups in
Fig. 7. Frequency histograms of attack (a) and copulation (b) scores on day ll post-hatch from lateralized and non-latera- lized populations. The symbols are as in Fig. 4. The mean score and standard error for each group going from top to bottom is as follows: for attack 5.81 + 0.34. 1.84 + 0.22, 3.79 + 0.37, 3.57 _+ 0.35 and for copulation 5.90 _+ 0.20, 1.60 + 0.23, 3.85 + 0.33, 4.21 _+ 0.38.

Fig. 7 clearly demonstrates that the distributions are skewed and the direction depends on the hemisphere treated. The left-treated groups are positively skewed towards the high attack and copulation scores, while the modes for the right-treated groups tend to be closer to zero. These distributions confirm the validity of observations made by using group
means as measures of laterality (viz. left-right hemi- sphere asymmetry).
In the non-lateralized groups a different picture emerges. A general statement that would apply to the data collected thus far is that the distribution of these non-lateralized populations are not skewed and
probably not unimodal. There is some indication that these populations are bimodal, this appears to be the case for attack and copulation scores of all the non- lateralized groups (Fig. 7), except perhaps the copu- lation scores of the left-treated, non-lateralized ani- mals. However, the distribution of the latter scores are still different from their lateralized counterparts as they are definitely not skewed in the same manner. As more data is collected the shapes of these distribu- tions will be delineated with greater clarity. The dif- ferences between these distributions also reflect the finding above that non-lateralized groups have great- er variances than lateralized groups.
DISCUSSION
The data presented demonstrates that the chicken forebrain functions asymmetrically with respect to the control of attack and copulatory behavior and
that this asymmetry can be manipulated by light ex- posure of the embryo during the last few days of incu- bation.
The high correlation between the scores which in- dividuals obtained on days 12 and 13 attests that indi- vidual chicks respond consistently on repetitions of the same test across days. Hence we need hold no reservations about the reliability of the measures of lateralityll,44.
Experiment 1A revealed the factors responsible for, or contributing to, lateralization; viz. that light exposure during the last 3 days of incubation is a suf- ficient condition for determining lateralization at the group or population level. Humidity, temperature control and fan noise were eliminated, not as unnec- essary but as insufficient to cause lateralization of at-
103
tack and copulation in the tested population, since
these factors were present in all incubating condi- tions to the same degree. Lateralization was present only in groups of chickens hatched after incubation in the conditions which received light. Presence or ab- sence of laboratory noise during the last 3 days of in-
cubation had no influence on either the presence or the degree of lateralization of attack or copulation, since lateralization was observed in the absence of
laboratory noise, L+S -, and exposure to laboratory noise per se, L-S +, was not sufficient to cause popula- tion lateralization of attack and copulation. Given that Woolf et al. 45 have demonstrated that a similar
level of auditory stimulation over incubator fan noise
can accelerate general development, it is unlikely that the failure of laboratory noise to influence lat- eralization can simply be attributed to its being masked by fan noise.
The results of the Levene tests on data for day 13 in Experiment 1A are difficult to interpret, since the variances of non-lateralized groups (L-S +) were not always significantly larger than those of the later-
alized groups (L+S ÷, L+S-). Nevertheless these tests revealed that non-lateralized groups tend to exhibit greater variance than lateralized groups, a finding consistent with Rogers' report 34. Moreover this dif- ference was associated with fundamental differences in frequency distributions; see below for further dis- cussion (Fig. 7).
In contrast to attack and copulation, the incubat- ing conditions in Experiment 1A did not alter the lat- eralization of auditory habituation. In chicks from all
3 incubating conditions, the left-treated groups re- quired more presentations of the auditory stimulus to habituate than did their right-treated counterparts. This is consistent with the findings of Howard et ai. 21 . This selective effect of light exposure on the laterali- zation of the visually guided responses of attack and copulation but not on auditory habituation argues
against global hypotheses which state that lateraliza- tion of all functions is determined in the same way. It could be that the auditory system has different re- quirements in terms of the nature of the perceptual input, the duration of exposure, or a different sensi- tive period.
In fact auditory habituation remained lateralized, even after the eggs were incubated in silent, still-air incubators (i.e. no fan noise) located in a sound-

104
proof room. Thus it seems unlikely that asymmetric- ally perceived auditory stimulation from an external
source during incubation is necessary to establish the lateralization of auditory habituation. However, it
must be noted that we did not control for vocaliza- tions or other sounds made by the embryos them- selves. Given that vocalizations or click sounds made by avian embryos can influence rate of devel- opment 18,~5 it may be possible that embryo-produced sounds provide sufficient auditory input to influence
lateralization in groups of eggs incubated together. Experiment 1A demonstrated that exposure to
light of randomly varying intensity (100-500 lux) 3 days prior to hatching will result in population latera-
lization of attack and copulation. Experiment 2A ex- tends this finding and demonstrates that exposure to a light of constant intensity for 4 h on day 19 can also cause population lateralization. One hour of con- stant-intensity light exposure on day 19 proved insuf- ficient to cause population lateralization. This find- ing is consistent with Rogers 34 study, which demon- strated that between 2 and 3 h of light of varying inten- sity on day 19 of incubation is sufficient to establish
lateralization at the population level. The ability of 4 h constant illumination to so radically affect brain organization may provide a useful system for studying biochemical and structural aspects of brain
development and plasticity. The intensity of the light in Experiment 2A at the
level of the eggs was measured to be between 500 and 800 lux depending on the location of the egg within the incubator, but as the eggs were not moved it was constant for any given egg. The intensity of the light
measured after passing through the egg shell with at- tendant membranes present was found to be between 30 and 60 lux. It is therefore quite conceivable that eggs incubated by hens would be exposed to light of
sufficient intensity to produce population lateraliza- tion given that the hen does not sit on the eggs contin-
uously. Experiment 2B examined whether 1 h exposure to
a flickering light (at 1.7 Hz) would be a more potent influence on lateralization of attack and copulation than 1 h exposure to a light of constant illumination. This expectation was based on the finding that flick- ering lights are more potent stimuli in inducing cer- tain behavioral and biochemical changes r-39. How- ever, the 1 h exposure used in our experiment did not
prove to be sufficient for causing lateralization in the population. Given that only one flicker frequency was used, we should remain cautious in our interpre- tations concerning the effects of flickering lights.
Our demonstration of the role of light in determin- ing asymmetry for attack and copulation supports the proposal that lateralization of forebrain function in the avian brain results from lateralized perceptual in-
put during development the right eye receiving more input than the left 35. This lateralized perceptual input
is the result of the accumulation of several factors; the orientation of the chicken in the egg 1~.32, the com-
plete decussation of the visual pathways at the optic chiasm 13, a functional visual system after the 18th day of incubation 40, and of course the fact that light can penetrate the egg shell 17,4°. This initial bias in devel- opment may be the basis for subsequent differentia- tion of asymmetry for attack and copulation between the hemispheres, and perhaps other pathways func- tionally associated with the visual system. Indeed, there is evidence to suggest that the side of the ner- vous system which leads in development exerts an in- hibitory influence on the trailing side, thus contribut- ing to the emergence of functional asymmetry j2.
Morphological and biochemical changes have
been demonstrated when dark-reared animals are exposed rather briefly to light. Cragg H found changes in the diameter of synaptic terminals in the visual cortex after dark-reared rats were exposed to 3 h of light. Similarly, it has been shown in chickens that brief exposure (3 h) to light shortly after hatch- ing markedly increases the responsiveness of cells in the hyperstriatum accessorium to diffuse retinal illu- mination9 and that this is accompanied by morpho- logical changes~. Other studies have shown that di- rect stimulation for as little as a few minutes produces morphological changes in the visual tract 19. Visual exposure is also associated with an array of biochem- ical and metabolic changes ~w.
The studies of cerebral blood flow in neonate chicks by Bondy and his associates are of particular interest with respect to our findings. They found that visual deprivation decreased blood flow through the contralateral hemisphere 7 and that visual attention affects brain blood flow~. Altered blood flow is inter- preted as an index of the energy requirements of ce- rebral tissue. If the effects of unilateral experiences prior to hatching parallel those observed in chickens

105
after hatching by Bondy, then there would be a
strong physiological basis for one hemisphere to de-
velop in advance of, or at least out of phase with, the
other 12. Unilateral visual experience may also increase
protein synthesis in the contralateral hemisphere 3,39.
In the rabbit the right eye has been found to open
several hours before the left eye and this asymmetric-
al visual experience is sufficient to accelerate mye- lination in the stimulated optic nerves 28.29. Moreover
unequal periods of monocular deprivation in cats
causes asymmetries in ocular dominance columns 42
and asymmetries in the size of visual fields 43 favour-
ing the stimulated visual pathways. It seems that pathways in the forebrain must be in-
volved in modulating attack and copulation in the
chicken, as the asymmetry becomes apparent after
unilateral MSG-treatment of the forebrain hemi-
spheres. However, as there is evidence that retinohy-
pothalamic projections may be mediating effects of
visual stimulation of endocrine processes associated
with the pineal and hypothalamic regulation of gona- dal function 23,26, asymmetry in lower pathways may
also be involved. In fact, asymmetry in hypothalamic
functioning has only recently been reported by Nor-
deen and Yahr 30 for the rat.
The influence of early experiences on brain lateral- ity has only recently been recognized 16,34. Denen-
berg16 has suggested that the rat brain is latently or
potentially lateralized, the latter's presence becom- ing manifest provided that the rats receive stimula- tion (handling) during infancy. He suggests that this
non-specific stimulation can only act on and exagger- ate a pre-existing genetically-determined brain asymmetry which is assumed to be in the same direc-
tion for all individuals in a population. In contrast
Rogers and Ansons '35 proposal does not need to as- sume an initial asymmetry in the brain either at the
population level or at the individual level.
The lateralization of attack and copulation in
chickens appears to be qualitatively different from
the type of lateralization described for rats by De-
nenberg 16. Examination of the frequency distribu-
tions of attack and copulation scores for groups of
non-lateralized animals revealed that both high and
low scoring animals occur in both the left-treated
group and in the right-treated group. Such a distribu-
tion would explain the variance effects found by the
Levene test in Experiment 1A. This is consistent
with the suggestion made by Rogers 34 that individu-
als in the non-lateralized groups still exhibit laterali-
zation for attack and copulation but half are later-
alized in one direction and half in the other so that con-
sistent lateralization at the population level fails to
emerge. Light therefore appears to synchronize the
direction of lateralization such that most individuals
in the population are lateralized in the same direc-
tion. In contrast to the effects of environmental expe-
riences described by Denenberg 16 for rats, light expe-
rience appears to affect the direction but not the de-
gree of brain asymmetry in chickens. However, both
examples clearly illustrate instances where environ- mental factors affect the presence of lateralization in
a population of individuals, and that lateralization of
brain function in these individuals results from the
complex interaction of genetic and environmental factors.
ACKNOWLEDGEMENTS
The authors express their thanks to Dr. Stella A.
Crossley for her discussion and suggestions. This re-
search was supported, in part, by the Australian Re-
search Grants Committee, Grant A77/153 to Profes- sor A. L. A. Boura and L. J. R.
REFERENCES
1 Andrew, R. J., Precocious adult behaviour in the young chick, Anita. Behav., 14 (1966) 485-500.
2 Andrew, R. J., Effects of testosterone on the behaviour of the domestic chick. II. Effects present in males, but not in females, Anita. Behav., 23 (1975) 156--168.
3 Bateson, P. P. G., Horn, G. and Rose, S. P. R., Effects of early experience on regional incorporation of precursors into RNA and protein in the chick brain, Brain Res., 39 (1973) 449--465.
4 Bianki, V. L., Lateralization of function in the animal brain, Int. J. Neurosci., 15 (1981) 37--47.
5 Bondy, S. C. and Harrington, M. E, Brain blood flow: al- teration by prior exposure to a learned task, Science, 199 (1978) 318-319.
6 Bondy, S. C., Lehman, R. A. W. and Purdy, J. L., Visual attention affects brain blood flow, Nature (Lond.), 248 (1974) 440--441.
7 Bondy, S. C. and Morelos, B. S., Stimulus deprivation and cerebral blood flow, Exp. Neurol., 31 (1971)200-206.
8 Bradley, P. and Horn, G., Effects of visual experience on

106
the chick brain: a light and electron microscopic study of the hyperstriatum accessorium, Brain Res., 162 (1979) 148--153.
9 Brown, M. W. and Horn, G., Neuronal plasticity in the chick brain electrophysiological effects of visual experience on hypostriatal neurones, Brain Res., 162 (1979) 142-147.
10 Collins, R. L., Toward an admissable genetic model for the inheritance of the degree and direction of asymmetry. In S. Harnard, R. W. Doty, L. Goldstein, J. Jaynes and G. Krauthamer (Eds.), Lateralization in the Nervous System, Academic Press, New York, 1977, pp. 137-150.
11 Collins, R. L., On asymmetries exhibiting a near-equi- probable distribution of direction, Behav. Brain Sci., 4 (1981)23--24.
12 Corballis, M. C. and Morgan, M. J., On the biological basis of human laterality, Behav. Brain Sci., 2 (1978) 261-336.
13 Cowan, W. M., Adamson, L. and Powell, T. P. S., An ex- perimental study of the avian visual system, J. Anat. (Lond.), 95 (1961) 545-563.
14 Cragg, B. G,, Changes in synaptic numbers and dimensions following first exposure of rats to light, Nature (Lond.), 215 (1967) 215-253.
15 Denenberg, V. H., Hemispheric laterality in animals and the effects of early experience, Behav. Brain Sci., 4 (1981) 1-21.
16 Denenberg, V. H., Hofmann, M., Garbanati, J. A., Sher- man, G. F., Rosen, G. D. and Yutzey, D. A., Handling in infancy, taste aversion, and brain laterality in rats, Brain Res., 200 (1980) 123-133.
17 Dimond, S. J. and Adam, J., Approach behaviour and em- bryonic visual experience in chicks: Studies of the effect of rate of visual flicker, Anirn. Behav., 20 (1972) 413-420.
18 Freeman, B. M. and Vince, M. A., Development o f the A vi- an Embryo, Chapman and Hall, London, 1974.
19 Friere, M., Effects of dark rearing on dendritic spines in layer IV of the mouse visual cortex. A quantitative electron microscopical study, J. Anat. (Lond.). 126 (1978) 193-201.
20 Gottlieb, G. and Simner, M. L., Auditory versus visual flicker in directing the approach response of domestic chicks, J. comp. physiol. Psychol., 67 (1969) 58-63.
21 Howard, K. J., Rogers, L. J. and Boura, A. L. A., Func- tional lateralisation of the chick forebrain revealed by use of intracraniat glutamate, Brain Res., 188 (1980) 369-382.
22 Keppel, G., Design and Analysis: A Researcher's Hand- book, Prentice-Hall, Englewood Cliffs, New Jersey, 1973.
23 Klein, D. C., The pineal gland: a model of neuroendocrine regulation. In S. Reichlin, R. J. Baldessarini and J. N. Mar- tin (Eds.), The Hypothalamus, Raven Press, New York, 1978, pp. 303-327.
24 Lauber, J. K. and Shutze, J. V., Accelerated growth of em- bryo chicks under the influence of light, Growth. 28 (1964) 179-190.
25 Levy, J., Psychobiological implications of bilateral asym- metry. In J. J. Dimond and J. G. Beaumont (Eds.), Hemi- sphere Function in the Human Brain. Elek. Science, Lon- don, 1974, pp. 121-183.
26 Meier, R. E., Autoradiographic evidence for a direct ret- ino-hypothalamic projection in the avian brain, Brain Res., 53 (1973) 417--42l.
27 Morgan, M. J., Embryology and inheritance of asymmetry. In S. R. Harnard, D. W. Doty, L. Goldstein, J. Jaynes and G. Krauthamer (Eds.), Lateralization in the Nervous Sys- tem, Academic Press, New York, 1977, pp. 173-194.
28 Narang, H. K., Right-left asymmetry of myelin devel- opment in epiretinal portion of rabbit optic nerve, Nature (Lond.), 266 (1977) 855-856.
29 Narang, N. K. and Wisniewski, H. M., The sequence of myelination in the epiretinal portion of the optic nerve in the rabbit, Neuropath. appl. Neurobiol., 3 (1977) 15-27.
30 Nordeen, E. J. and Yahr, P., Hemispheric asymmetries in the behavioural and hormonal effects of sexually differen- tiating mammalian brain, Science, 218 (1982) 391-394.
31 Nottebobm, F., Neural lateralization of vocal control in a Passerine bird. II. Sub-song, calls and a theory of vocal learning, J. exp. Zool., 179 (1972) 35-50.
32 Oppenheim, R. W., Prehatching and hatching behavior: a comparative and physiological consideration. In G. Gott- lieb (Ed.), Studies on the Development of Behavior and the Nervous System, Behavioural Embryology Vol. 1, Aca- demic Press, New York, 1978, pp. 163-244.
33 Rogers, L. J., Lateralisation in the avian brain, Bird Be- haw. 2 (1981)) 1-12.
34 Rogers, L. J., Light experience and asymmetry of brain function in chickens, Nature (Lond.), 297 (1982) 223-225.
35 Rogers, L. J. and Anson, J. M., Lateralisation of function in the chicken forebrain, Pharmacol. Biochem. Behav., 10 (1979) 679-686.
36 Rogers, L. J., Dr.ennen, H. D. and Mark, R. F., Inhibition of memory formation in the imprinting period: irreversible action of cycloheximide in young chickens, Brain Res., 79 (1974) 213-233.
37 Rogers, L. J. and Hambley, J. W., Specific and non-specific effects of neuro-excitatory amino acids on learning and oth- er behaviours in the chicken, Behav. Brain Res., 4 (1982) 1-18.
38 Rose, S. P. R., Bateson, P. P. G., Horn, A. L. D. and Horn, G., Effects of an imprinting procedure on regional incorporation of tritiated uracil into chick brain RNA, Na- ture (Lond.), 225 (1970) 650-651.
39 Rose, S. P. R. and Haywood, J., Experience, learning and brain metabolism. In A. N. Davison (Ed.), Biochemical Correlates of Brain Structure and Function, Academic Press, London, 1977, pp. 249-292.
40 Sedl~i~ek, J., Development of the optic afferent system in chick embryos. In G. Newton and A. Reisen (Eds.), Ad- vances in Psychobiology, Vol. 1, Wiley-Interscience, New York, 1972, pp. 129-170.
41 Shapiro, L. J., Pre-hatching influences that can potentially mediate post-hatch attachments in birds, Bird Behav., 3 (1981) 1-18.
42 Shatz, C. J. and Stryker, M. P., Occular dominance in layer IV of the cat's visual cortex and the effects of monocular deprivation, J. Physiol. (Lond.), 281 (1978) 267-284.
43 Tumosa, N., Tieman, S. B. and Hirsch, H. U. B.. Unequal alternating monocular deprivation causes asymmetric visu- al fields in cats, Science, 25 (1980) 421-423.
44 Warren, J. W., Laterality and natural selection, Behav. Brain Sci., 4 (1981) 36-37.
45 Woolf, N. K., Bixby, J. L. and Capranica, R. R., Prenatal experience and avian development; brief auditory stimula- tion accelerates the hatching of Japanese quail, Science. 194 (1976) 959-960.
46 Young, C. E. and Rogers, L. J., Effects of steroidal hor- mones on sexual behavior, attack and search behavior in the isolated male chick, Horm. Behav., 10 (1978) 107-t 17.





























