Embed Size (px)
Citation preview

Mar Biol
DOI 10.1007/s00227-008-0915-1RESEARCH ARTICLE
Latitudinal symbiont zonation in Stylophora pistillata from southeast Africa
Angus H. H. Macdonald · Eugenia M. Sampayo · Tyrone Ridgway · Michael H. Schleyer
Received: 6 August 2007 / Accepted: 14 January 2008© Springer-Verlag 2008
Abstract Studies on latitudinal gradients in Symbiodi-nium diversity on scleractinian corals are largely restrictedto warm-water low latitude locations, and it appears thatthere is a shift in symbiont distributions with increasing lat-itude. The Symbiodinium assemblages of high latitude coralcommunities have largely been undocumented despiteoccupying an important transitional zone between tropicaland temperate regions. Using a combination of the internaltranscribed spacer region 2 (ITS2) and denaturing gradientgel electrophoresis (DGGE), we assessed the cladal andsubcladal variability of Symbiodinium in the widely distrib-uted species Stylophora pistillata along a latitudinal tran-sect in southeast African waters which extended into highlatitude locations. All colonies examined belonged to cladeC. Six unique ITS2-DGGE banding proWles (designatedCspa to Cspf) were observed, which showed a latitudinal dis-tribution from north to south, most likely a result of a gradi-ent in water temperature and irradiance driven by riverineinput in the southern regions. Sequence analysis revealedthat all sequences except one did not match previouslyidentiWed clade C sub-types, probably due to the lack ofregional information in the Western Indian Ocean whencompared to the Caribbean and PaciWc. This study furthersupports the applicability of ITS2-DGGE in studies on
Symbiodinium diversity, and highlights that potentially eco-logically informative biogeographic patterns may be over-looked when only cladal designations are employed.
Introduction
Coral reefs are restricted to tropical waters, and the pole-ward distribution of these systems is largely limited bywater temperature, or one of its environmental or ecologicalcorrelates. Kleypas and coworkers (1999) reviewed thephysiochemical factors that limit coral reef developmentand reported that water temperature, light penetration, andaragonite saturation levels are the most signiWcant factorsdetermining reef development, with biological factors ofonly local signiWcance. Scleractinian coral species numbersgradually decline along a latitudinal gradient and thedecline tends to be heightened at around 24° to 25° latitude,largely due to temperature (data extracted from Veron1993). Given the attention that climate change andincreased sea temperatures have been aVorded over the pastdecade, the southward/northward migration of corals tohigher latitude locations has been proposed as a possiblesurvival strategy for coral reefs (Done 1999).
The occurrence of genetically diVerent clades (A–H) ofSymbiodinium (e.g. Rowan and Powers 1991; Baker 2003for review) within coral tissues has been used to explainzonation patterns as well as environmental tolerancebetween corals (e.g. Rowan and Knowlton 1995). In recentyears, more variable genetic markers have been employed,whereby the cladal system has seen a rapid expansion into alarge number of subcladal types (e.g. van Oppen et al.2001; LaJeunesse 2002). The expansion of the cladal sys-tem into subcladal types has uncovered a high degree ofhost–symbiont speciWcity as well as Wne-scale biogeograpical
Communicated by M.I. Taylor.
A. H. H. Macdonald (&) · M. H. SchleyerOceanographic Research Institute, PO Box 10712, Marine Parade, Durban 4056, South Africae-mail: [email protected]
E. M. Sampayo (&) · T. RidgwayCentre for Marine Studies, University of Queensland, St Lucia, QLD 4072, Australiae-mail: [email protected]
123

Mar Biol
and ecological patterns that would have otherwise remainedundetected (Iglesias-Prieto et al. 1992; LaJeunesse et al.2004; Warner et al. 2006; Sampayo et al. 2007).
Symbiodinium diversity has been assessed on broadlatitudinal (e.g. Loh et al. 2001; van Oppen et al. 2005) orlongitudinal gradients (e.g. Burnett 2002), and on local reefscales (e.g. LaJeunesse et al. 2003; Fabricius et al. 2004;Sampayo et al. 2007) but these studies are largely restrictedto low latitude locations. Nevertheless, limited evidencesuggests that widely distributed host species show a shift intheir symbiont composition at high latitude locations.LaJeunesse and Trench (2000) found that the sea anemoneAnthopleura elegantissima from the temperate northernsites of Washington and Oregon contained a diVerent sym-biont (clade B) complement in comparison to southernCalifornian sites. A latitudinal pattern also occurs in thecoral species Plesiastrea versipora (Rodriguez-Lanetty et al.2001) along the East Australian coast, whereby a cladaltransition occurs from clade C to B in the cooler, high-lati-tude communities. Rowan and Powers (1991) also reportthe presence of clade B in Astrangia danae from Massachu-setts, but no collections were made in tropical or sub-tropi-cal locations. More recently, Lien et al. (2007) investigatedSymbiodinium diversity in Oulastrea crispata from Thai-land (6°N) to Japan (35°N), and found that the temperateJapanese populations harbored only clade D, whereas thetropical locations harbored both clade C and D.
Based on the limited studies available it appears that ashift in symbiont distributions occurs with increasing lati-tude, a process which may be driven by changes in temper-ature, irradiance, turbidity or the possible isolation ofhigher latitude host populations. Comparative latitudinalstudies that extend into temperate waters generally requirespecies with widespread distributions from tropical to tem-perate locations, which is somewhat restrictive in sclerac-tinian corals. There is a deWnite need to increase latitudinaldistribution studies on Symbiodinium diversity into highlatitude environments, and the East African coastline pro-vides an ideal latitudinal gradient to test the hypothesis thatSymbiodinium distributions change with increasing latitude.
Southern Mozambique and South Africa’s coral commu-nities represent a small subset of reefs that lack most of thegeomorphological traits typical of tropical reefs. Speciesnumbers on these reefs decrease signiWcantly with latitudewith Nine-mile Reef (27°S) reported to have about 90species of scleractinian coral (Celliers and Schleyer 2002),Aliwal Shoal (30°S) only 15 species, and Pondoland (31°S)only three species (Stylophora pistillata, Anomastrea irreg-ularis and Coscinarea sp.) which represents only 0.2%of the benthic cover (Celliers et al. 2007). S. pistillata,although widely distributed throughout the Indo-PaciWc isregionally rare in certain locations. It is distributed through-out the range of coral habitat along the southeast African
coast and thus presents an ideal candidate to assess Symbi-odinium latitudinal zonation. The symbionts in S. pistillataare vertically transmitted to the oVspring (i.e. they areacquired directly from the mother colony) (Atoda 1947), astrategy that has been suggested to ensure the persistence ofa successful symbiotic relationship (Douglas 1998; Lohet al. 2001). While being released with a full complementof symbionts may impose limitations on the Xexibility, tol-erance and dispersal ranges of this species, a recent detailedassessment of Symbiodinium diversity at Heron Island onthe Great Barrier Reef (Sampayo et al. 2007) showed thatS. pistillata does have the capacity to harbor multiple sym-biont types.
Given the results of prior studies from diVerent symbi-otic hosts and geographic areas (LaJeunesse and Trench2000; Rodriguez-Lanetty et al. 2001; Lien et al. 2007), wehypothesized that Symbiodinium populations in S. pistillatawould vary along a latitudinal transect along the southeastAfrican coast in the Western Indian Ocean. We use a com-bination of the internal transcribed spacer region 2 (ITS2)and denaturing gradient gel electrophoresis (DGGE) toassess the cladal and subcladal variability of Symbiodiniumalong this geographic gradient.
Materials and methods
Sample collection
Samples of S. pistillata were collected from four siteswithin the high-latitude communities of South Africa, andan additional reef location from Mozambique (InhacaIsland) (Fig. 1, Table 1). In addition, three opportunistic
Fig. 1 Sampling locations of Stylophora pistillata (inset) along thesoutheast African coast. GPS co-ordinates of the sampling locationsappear in Table 1
123
Mar Biol
samples were obtained from a true reef location from anexpedition to Madagascar by one of the authors (MHS)(Fig. 1; Table 1). The latitudinal range of the sample loca-tions encompasses 2500 km, which includes a decreasingcline in temperature from north to south (Table 1; Reynoldsand Smith 1994). Due to the somewhat low abundance of S.pistillata at some sites, the level of access and permitrestrictions of working in regional Marine Protected Areas,sample sizes at Inhaca Island and Nine-mile Reef were lessthan originally planned. At each site, tissue samples werecollected randomly by removing 4 cm2 fragments from col-onies at least 3 m apart. Samples were placed in DMSObuVer (DMSO 20% v/v, EDTA 0.25 M saturated withNaCl) for storage, and DNA was extracted using a standardPhenol:Chloroform extraction (Sambrook and Russell2001).
ITS2 PCR and DGGE
The extracted DNA was ampliWed for the internal tran-scribed spacer region 2 (ITS2) of the ribosomal DNA usingthe primers ITSintfor (5�-GAA TTG CAG AAC TCC GTG-3�) and ITS2clamp (5�- CGC CCG CCG CGC CCC GCGCCC GTC CCG CCG CCC CCG CCC GGG ATC CATATG CTT AAG TTC AGC GGG T-3�) (LaJeunesse 2002).The PCR conditions were: 10 min at 94°C; 30 cycles at 45s94°C, 45s 60°C and 1°min 72°C; with a Wnal extension of20 min at 72°C. The ampliWed ITS2 products were visual-ized using denaturing gradient gel electrophoresis (DGGE)using a Biorad DCode system. 10 �l of the ampliWed ITS2DNA was loaded onto the gel with 10 �l of 2£ gel loadingdye (Biorad DCode Dye Solution). Samples were loaded
onto 1 mm thick 8% acrylamide gels (16 cm) with an inter-nal gradient of 30–60% denaturants (formamide and urea)(cf. Sampayo et al. 2007). Gels were run at 100 V for 12 h,stained for 20–30 min using SYBR Green (Amresco,10.000 £ diluted from stock) and visualized using an UVI-Doc digital camera gel imaging system. Gels were scoredbased on unique banding proWles that included the entirecomplexity of bands present in the proWle.
Sequence analysis
The dominant repeat of the rDNA was obtained by exci-sion of the dominant band(s) from three representativesamples of each unique DGGE proWle (LaJeunesse 2002;LaJeunesse et al. 2004). Bands were eluted overnight in500 �l dH2O and re-ampliWed the following day using theprimers for DGGE. AmpliWcations of excised bands werererun on the DGGE alongside the original sample fromwhich they were excised to ensure that the correct bandwas obtained without contamination. After this conWrma-tion process, the excised bands were ampliWed followingthe methods outlined in LaJeunesse (2002) and puriWedusing the UltraClean PCR puriWcation Kit (MolecularBiology Laboratories, USA). PuriWed DNA was sent forsequencing using the forward (ITSintfor2) or reverse(ITS2reverse) primers in separate runs at the AustralianGenome Research Facility (ABI BigDye Terminator(v3.1) chemistry run on AB3730xl sequencer). All chro-matograms were visually checked using the programSeqman (Lasergene software package), and the resultingconsensus sequence (representing the dominant band onthe DGGE for the proWle of interest) was blasted against
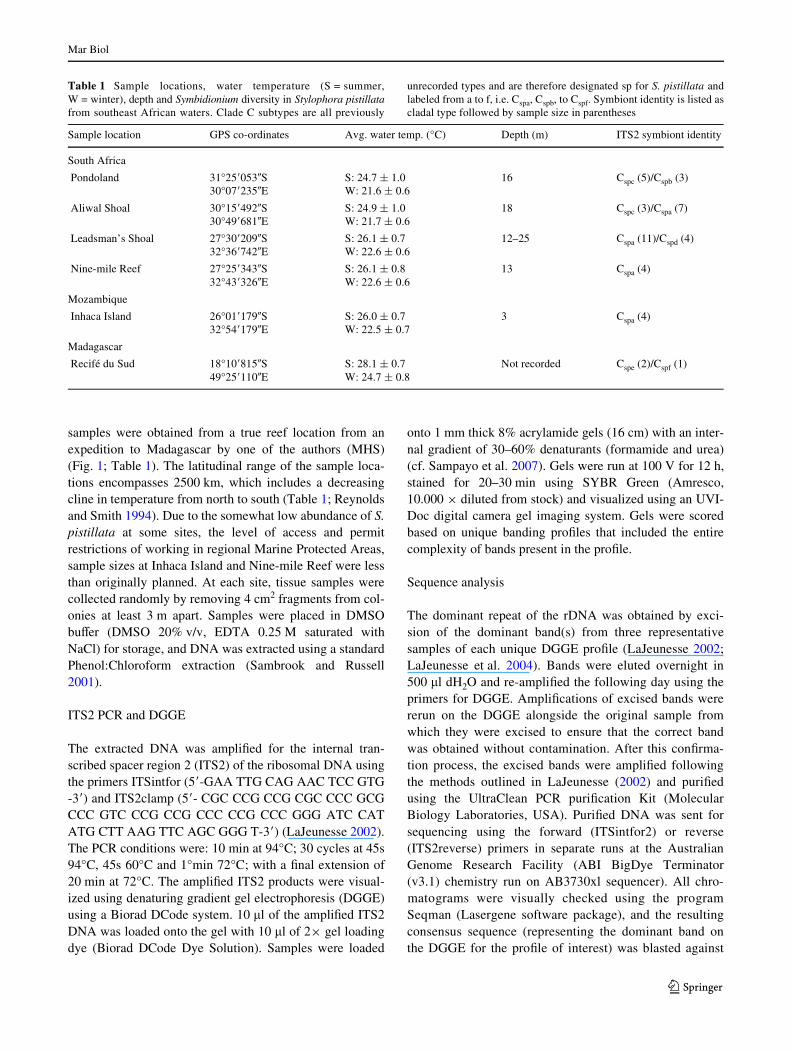
Table 1 Sample locations, water temperature (S = summer,W = winter), depth and Symbidionium diversity in Stylophora pistillatafrom southeast African waters. Clade C subtypes are all previously
unrecorded types and are therefore designated sp for S. pistillata andlabeled from a to f, i.e. Cspa, Cspb, to Cspf. Symbiont identity is listed ascladal type followed by sample size in parentheses
Sample location GPS co-ordinates Avg. water temp. (°C) Depth (m) ITS2 symbiont identity
South Africa
Pondoland 31°25�053�S30°07�235�E
S: 24.7 § 1.0W: 21.6 § 0.6
16 Cspc (5)/Cspb (3)
Aliwal Shoal 30°15�492�S30°49�681�E
S: 24.9 § 1.0W: 21.7 § 0.6
18 Cspc (3)/Cspa (7)
Leadsman’s Shoal 27°30�209�S32°36�742�E
S: 26.1 § 0.7W: 22.6 § 0.6
12–25 Cspa (11)/Cspd (4)
Nine-mile Reef 27°25�343�S32°43�326�E
S: 26.1 § 0.8W: 22.6 § 0.6
13 Cspa (4)
Mozambique
Inhaca Island 26°01�179�S32°54�179�E
S: 26.0 § 0.7W: 22.5 § 0.7
3 Cspa (4)
Madagascar
Recifé du Sud 18°10�815�S49°25�110�E
S: 28.1 § 0.7W: 24.7 § 0.8
Not recorded Cspe (2)/Cspf (1)
123

Mar Biol
previously recorded Symbiodinium types on GenBankhttp://www.ncbi.nih.gov. All sequences were alignedusing ClustalX (Thompson et al. 1997) with the inclusionof the ancestral generalist C-types, C1 and C3 (LaJeunesse2005). The sequence alignment was annotated to highlightimportant structural components of the ITS2 region, whichare characteristically speciWc of clade C symbionts(Hunter et al. 2007). All sequences were assessed, again incomparison to types C1 and C3, to determine whether theobserved base changes conferred negative disruptions tothe ITS2 secondary structure (Hunter et al. 2007; Thornhillet al. 2007). Finally, a phylogenetic analysis was per-formed using the structurally sound dominant repeatsequences of the ITS2. Analysis was run under the maxi-mum parsimony criterion treating gaps as a Wfth characterstate and indels as a single evolutionary event. Bootstrapvalues were calculated based on 1,000 replicates and allanalysis was done using the program PAUP (version4.0b10).
Results
ITS2 PCR and DGGE
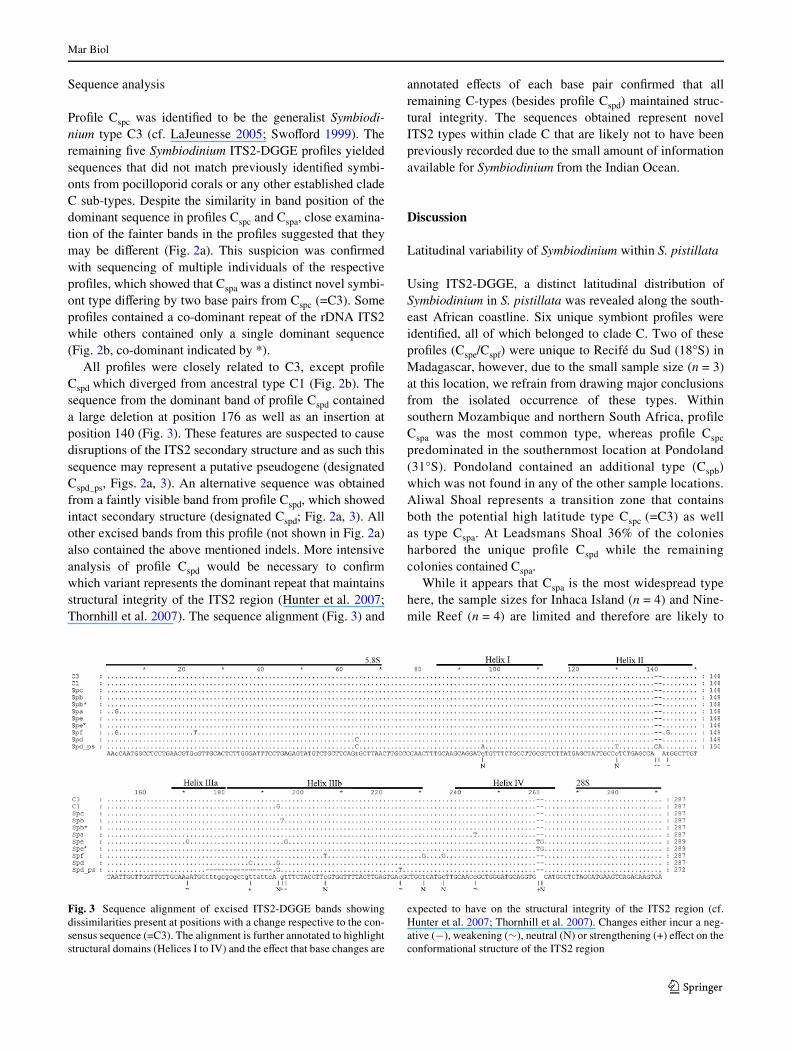
All colonies of southeast African S. pistillata containedclade C symbionts, and further separation into ITS2 subcla-dal types uncovered a latitudinal distribution. In total, sixITS2-DGGE banding proWles were identiWed (Cspa to Cspf,Fig. 2a). Two of these proWles (Cspe/Cspf) were unique toRecifé du Sud in Madagascar (Fig.2a; Table 1). Cspa
occurred at Inhaca Island, Nine-mile Reef, Leadsman’sShoal and Aliwal Shoal, whereas Cspc was only presentin the southern locations at Aliwal Shoal and Pondoland.Two other symbiont types Cspb and Cspd were present atPondoland and Leadsmans Shoal respectively (Table 1).Symbiont type Cspd was strongly correlated to a diVerentmorphotype of the host that was previously establishedusing these samples and a multivariate analysis of hostmorphometrics (Macdonald 2005).
Fig. 2 a Symbiont ITS2-types in Stylophora pistillata from southeastAfrican waters. Dominant and co-dominant (*) bands in the ITS2-DGGE proWles which were excised are indicated with arrowheads.Corresponding sequences were deposited on GenBank with accessionnumbers EU431995, EU431996 and EU436642- EU436648. Note thatthe dominant band of proWle Cspa and proWle Cspc runs to a similar po-sition in the gel but the diVerence between proWles is apparent from thefainter bands and is conWrmed by a 2-base pair sequence diVerence. bMost parsimonious un-rooted tree of the ITS2 rDNA dominant repeats
identiWed by ITS2-DGGE and direct sequencing of excised bands.Bootstrap values are based on 1,000 replicates and are only representedif larger than 50%. Ancestral types C1 and C3 are represented by largergrey circles. Circle size is not representative of abundance and branchdistance is represented by length of connections. Variant Cspd_ps likelyrepresents a pseudogene and contains a large deletion (see also Fig. 3)that was treated as single gap. Branch length for this variant is thereforenot representative of the total number of base changes and is shown asa dashed line
(a) (b)
123
Mar Biol
Sequence analysis
ProWle Cspc was identiWed to be the generalist Symbiodi-nium type C3 (cf. LaJeunesse 2005; SwoVord 1999). Theremaining Wve Symbiodinium ITS2-DGGE proWles yieldedsequences that did not match previously identiWed symbi-onts from pocilloporid corals or any other established cladeC sub-types. Despite the similarity in band position of thedominant sequence in proWles Cspc and Cspa, close examina-tion of the fainter bands in the proWles suggested that theymay be diVerent (Fig. 2a). This suspicion was conWrmedwith sequencing of multiple individuals of the respectiveproWles, which showed that Cspa was a distinct novel symbi-ont type diVering by two base pairs from Cspc (=C3). SomeproWles contained a co-dominant repeat of the rDNA ITS2while others contained only a single dominant sequence(Fig. 2b, co-dominant indicated by *).
All proWles were closely related to C3, except proWleCspd which diverged from ancestral type C1 (Fig. 2b). Thesequence from the dominant band of proWle Cspd containeda large deletion at position 176 as well as an insertion atposition 140 (Fig. 3). These features are suspected to causedisruptions of the ITS2 secondary structure and as such thissequence may represent a putative pseudogene (designatedCspd_ps, Figs. 2a, 3). An alternative sequence was obtainedfrom a faintly visible band from proWle Cspd, which showedintact secondary structure (designated Cspd; Fig. 2a, 3). Allother excised bands from this proWle (not shown in Fig. 2a)also contained the above mentioned indels. More intensiveanalysis of proWle Cspd would be necessary to conWrmwhich variant represents the dominant repeat that maintainsstructural integrity of the ITS2 region (Hunter et al. 2007;Thornhill et al. 2007). The sequence alignment (Fig. 3) and
annotated eVects of each base pair conWrmed that allremaining C-types (besides proWle Cspd) maintained struc-tural integrity. The sequences obtained represent novelITS2 types within clade C that are likely not to have beenpreviously recorded due to the small amount of informationavailable for Symbiodinium from the Indian Ocean.
Discussion
Latitudinal variability of Symbiodinium within S. pistillata
Using ITS2-DGGE, a distinct latitudinal distribution ofSymbiodinium in S. pistillata was revealed along the south-east African coastline. Six unique symbiont proWles wereidentiWed, all of which belonged to clade C. Two of theseproWles (Cspe/Cspf) were unique to Recifé du Sud (18°S) inMadagascar, however, due to the small sample size (n = 3)at this location, we refrain from drawing major conclusionsfrom the isolated occurrence of these types. Withinsouthern Mozambique and northern South Africa, proWleCspa was the most common type, whereas proWle Cspc
predominated in the southernmost location at Pondoland(31°S). Pondoland contained an additional type (Cspb)which was not found in any of the other sample locations.Aliwal Shoal represents a transition zone that containsboth the potential high latitude type Cspc (=C3) as wellas type Cspa. At Leadsmans Shoal 36% of the coloniesharbored the unique proWle Cspd while the remainingcolonies contained Cspa.
While it appears that Cspa is the most widespread typehere, the sample sizes for Inhaca Island (n = 4) and Nine-mile Reef (n = 4) are limited and therefore are likely to
Fig. 3 Sequence alignment of excised ITS2-DGGE bands showingdissimilarities present at positions with a change respective to the con-sensus sequence (=C3). The alignment is further annotated to highlightstructural domains (Helices I to IV) and the eVect that base changes are
expected to have on the structural integrity of the ITS2 region (cf.Hunter et al. 2007; Thornhill et al. 2007). Changes either incur a neg-ative (¡), weakening (»), neutral (N) or strengthening (+) eVect on theconformational structure of the ITS2 region
123

Mar Biol
represent an underestimation of the regional diversity.This may help explain the isolated occurrence of type Cspd
at Leadmans Shoal given the larger sample size (n = 15)at this location. Larger sample sizes would have been theideal, but diYculties associated with working in thesesoutheast African locations did restrict the numbersobtained to a mean n = 7 per location. However, similarstudies on P. versipora (Rodriguez-Lanetty et al. 2001)and Oulastrea crispata (Lien et al. 2007) had mean sam-ple sizes per location of n = 8 and n = 6 respectively.Most Symbiodinium diversity assessments on single siteson coral reefs have thus far been represented by relativelysmall sample sizes per host species (e.g. van Oppen et al2001, 2005; LaJeunesse et al. 2003, 2004). Althoughadditional symbiont types may be uncovered by increasedsampling of a single species (Sampayo et al. 2007), itshould be pointed out that the number of symbionts withwhich a single host can associate (particularly at a singlelocation) is likely to be limited. Host–symbiont associa-tions are highly speciWc even on global scales (LaJeu-nesse 2005) and the ability to host multiple symbionttypes between diVerent colonies of a single host species isgenerally limited (van Oppen et al. 2001; Baker 2003;LaJeunesse et al. 2003; Thornhill et al. 2006; Warneret al. 2006). S. pistillata on the Great Barrier Reef hasbeen found to associate with up to four subcladal symbi-ont types at a single location (Sampayo et al. 2007), andalthough that study showed that ecological patterns maybe confounded by the use of limited sampling eVort, italso showed that the full complexity of host–symbiontassociations could generally be established with lowersamples sizes (6–10). At Harry’s Bommie for example, 31colonies were sampled at a depth of 5 m, and three symbi-ont proWles were found at frequencies of 52, 32, and 16%(Sampayo et al. 2007). Similarly, sampling 40 colonies ata depth of 18 m, three symbiont proWles were found at fre-quencies of 60, 37, and 3% (Sampayo et al. 2007).Although in this study only two symbiont types were dis-covered in most locations, the predominant symbiont typeoccurred in frequencies of 60–70%, similar to the largersample sizes of Sampayo et al. (2007) for the same spe-cies. Also, no pronounced depth pattern was observed inthat Cspa occurred at depths ranging from 3 to 25 m(Table 1). Thus, the major eVect of the sample sizes is apotential underestimation of the subcladal diversitywithin sites, and a resultant subsequent inXation of theproportion of the potentially dominant symbiont type ateach location. Whilst we acknowledge the limitations thatthe low sample sizes may have on the present data, thereis a clear shift in symbiont types at the latitudinal extentof S. pistillata’s range in South Africa.
This pattern is best explained by regional temperaturediVerences as the sample locations span a decreasing
temperature continuum from north to south (Table 1). Asmentioned earlier, a similar latitudinal pattern occurs inthe coral species P. versipora (Rodriguez-Lanetty et al.2001) in Australia, whereby a cladal transition occursfrom clade C to B in the cooler, high latitude communi-ties. Similarly, LaJeunesse and Trench (2000) found thatthe temperature diVerence between northern and centralparts of the West coast of the USA. drives cladal and sub-cladal diVerences in symbiont populations of the seaanemone A. elegantissima. However, temperature is notthe only signiWcant environmental variable that changespredictably with latitude, and irradiance follows a similarpattern. Both Aliwal Shoal and Pondoland are stronglyinXuenced by riverine run-oV with subsequent increasesin sediment loads compared to the northern SouthAfrican locations. Thus, not only is there a reduction inscleractinian coral diversity from Leadmans Shoal toPondoland, but also a reduction in their abundance and achange in their morphology that may be attributable tothe lower sea temperatures and reduced light availability(Celliers et al. 2007). The occurrence of S. pistillata oVthe Pondoland coast not only represents the southernmostlimit of reef-building coral distribution along the south-ern African coastline (Celliers et al. 2007), but also asouthward transition in colony form from digitate/spatu-late to submassive/massive forms (Macdonald 2005).ITS sequencing of the host conWrmed that all coloniesrepresent a single species (Macdonald 2005). Thus themorphological changes observed mirror those of theSymbiodinium patterns uncovered and appear to accom-pany changes in water quality (turbidity increase–reduced irradiance) and temperature (temperature reduc-tion).
However, diVerentiation can also occur as a result ofgeographical isolation but information on connectivity ofcoral populations in the study area is limited. A recentreview highlighted that genetic knowledge in the WesternIndian Ocean is very limited in terms of marine ecosystemvariability and connectivity (Ridgway and Sampayo 2005)with only one genetic study available on a single coral spe-cies at the local scale (Ridgway et al. 2001). High levels ofgenetic exchange are present between Nine-mile Reef andLeadsman’s Shoal for Pocillopora verrucosa (a member ofthe same coral family; Ridgway et al. 2001), yet thereappears to be limited genetic exchange in P. verrucosabetween Bazaruto Island (21°S) in central Mozambique andthe South African reefs (Ridgway et al. 2007). No furtherknowledge on the connectivity of the South African andsouthern Mozambican communities is available but thesoutherly direction of the Agulhas Current as well as thesimilarity in composition of the reefs at Inhaca Island andSouth Africa suggests that interplay of temperature andirradiance rather than isolation plays a major role in shaping
123
Mar Biol
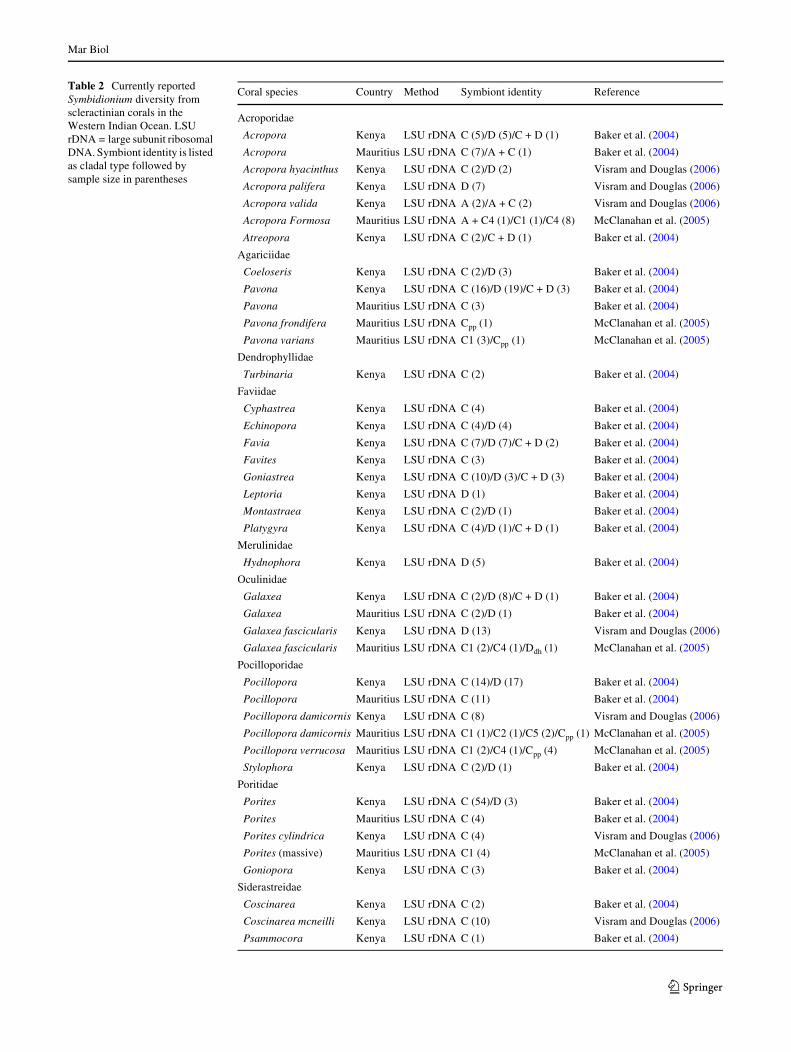
Table 2 Currently reported Symbidionium diversity from scleractinian corals in the Western Indian Ocean. LSU rDNA = large subunit ribosomal DNA. Symbiont identity is listed as cladal type followed by sample size in parentheses
Coral species Country Method Symbiont identity Reference
Acroporidae
Acropora Kenya LSU rDNA C (5)/D (5)/C + D (1) Baker et al. (2004)
Acropora Mauritius LSU rDNA C (7)/A + C (1) Baker et al. (2004)
Acropora hyacinthus Kenya LSU rDNA C (2)/D (2) Visram and Douglas (2006)
Acropora palifera Kenya LSU rDNA D (7) Visram and Douglas (2006)
Acropora valida Kenya LSU rDNA A (2)/A + C (2) Visram and Douglas (2006)
Acropora Formosa Mauritius LSU rDNA A + C4 (1)/C1 (1)/C4 (8) McClanahan et al. (2005)
Atreopora Kenya LSU rDNA C (2)/C + D (1) Baker et al. (2004)
Agariciidae
Coeloseris Kenya LSU rDNA C (2)/D (3) Baker et al. (2004)
Pavona Kenya LSU rDNA C (16)/D (19)/C + D (3) Baker et al. (2004)
Pavona Mauritius LSU rDNA C (3) Baker et al. (2004)
Pavona frondifera Mauritius LSU rDNA Cpp (1) McClanahan et al. (2005)
Pavona varians Mauritius LSU rDNA C1 (3)/Cpp (1) McClanahan et al. (2005)
Dendrophyllidae
Turbinaria Kenya LSU rDNA C (2) Baker et al. (2004)
Faviidae
Cyphastrea Kenya LSU rDNA C (4) Baker et al. (2004)
Echinopora Kenya LSU rDNA C (4)/D (4) Baker et al. (2004)
Favia Kenya LSU rDNA C (7)/D (7)/C + D (2) Baker et al. (2004)
Favites Kenya LSU rDNA C (3) Baker et al. (2004)
Goniastrea Kenya LSU rDNA C (10)/D (3)/C + D (3) Baker et al. (2004)
Leptoria Kenya LSU rDNA D (1) Baker et al. (2004)
Montastraea Kenya LSU rDNA C (2)/D (1) Baker et al. (2004)
Platygyra Kenya LSU rDNA C (4)/D (1)/C + D (1) Baker et al. (2004)
Merulinidae
Hydnophora Kenya LSU rDNA D (5) Baker et al. (2004)
Oculinidae
Galaxea Kenya LSU rDNA C (2)/D (8)/C + D (1) Baker et al. (2004)
Galaxea Mauritius LSU rDNA C (2)/D (1) Baker et al. (2004)
Galaxea fascicularis Kenya LSU rDNA D (13) Visram and Douglas (2006)
Galaxea fascicularis Mauritius LSU rDNA C1 (2)/C4 (1)/Ddh (1) McClanahan et al. (2005)
Pocilloporidae
Pocillopora Kenya LSU rDNA C (14)/D (17) Baker et al. (2004)
Pocillopora Mauritius LSU rDNA C (11) Baker et al. (2004)
Pocillopora damicornis Kenya LSU rDNA C (8) Visram and Douglas (2006)
Pocillopora damicornis Mauritius LSU rDNA C1 (1)/C2 (1)/C5 (2)/Cpp (1) McClanahan et al. (2005)
Pocillopora verrucosa Mauritius LSU rDNA C1 (2)/C4 (1)/Cpp (4) McClanahan et al. (2005)
Stylophora Kenya LSU rDNA C (2)/D (1) Baker et al. (2004)
Poritidae
Porites Kenya LSU rDNA C (54)/D (3) Baker et al. (2004)
Porites Mauritius LSU rDNA C (4) Baker et al. (2004)
Porites cylindrica Kenya LSU rDNA C (4) Visram and Douglas (2006)
Porites (massive) Mauritius LSU rDNA C1 (4) McClanahan et al. (2005)
Goniopora Kenya LSU rDNA C (3) Baker et al. (2004)
Siderastreidae
Coscinarea Kenya LSU rDNA C (2) Baker et al. (2004)
Coscinarea mcneilli Kenya LSU rDNA C (10) Visram and Douglas (2006)
Psammocora Kenya LSU rDNA C (1) Baker et al. (2004)
123

Mar Biol
the latitudinal zonation of the diverse symbiont communityobserved in S. pistillata.
Western Indian ocean symbiont diversity
The Western Indian Ocean encompasses Mozambique,Somalia, South Africa, Tanzania, Comores, Madagascar,Reunion, and the Seychelles, all of which have shownmajor signs of environmental degradation as well asdeclines in natural resources and biodiversity (Berg et al.2002). Whilst obviously degrading, the current level of sci-entiWc information in the Western Indian Ocean is limited,which coupled with the levels of reef degradation high-lights the immediate need to expand scientiWc knowledge inthe region. In terms of Symbiodinium diversity, only fourstudies are available for the Western Indian Ocean (Burnett2002; Baker et al. 2004; McClanahan et al. 2005; Visramand Douglas 2006), which is in direct contrast to the well-categorized Caribbean and greater PaciWc regions. Three ofthese studies focus on scleractinian corals but are restrictedto study sites around Kenya and Mauritius while the fourthis on a zoanthid (Palythoa caesia) covering sites from boththe Eastern and Western Indian Ocean. The current knowl-edge of Symbiodinium diversity in reef-building corals inthe Western Indian Ocean is based on a broad cladal leveland is limited to nine families of hosts from Kenya andMauritius (Table 2). In general, clades C and D predomi-nate the reefs of Kenya and Mauritius, with a few isolatedoccurrences of clade A in Acropora. The prevalence ofclade D in Kenyan corals has been suggested to be theresult of bleaching episodes because clade D may be morethermally tolerant compared to clade C (Baker et al. 2004),however clade D has also been found to occur in turbid/low-light (van Oppen et al. 2001; Toller et al. 2001) andhigh-latitude/colder environments (Lien et al. 2007).
Unlike LaJeunesse and Trench (2000) and Rodriguez-Lanetty et al. (2001), no cladal transitions were found inthis study. However symbiont diversity in S. pistillata onthe Great Barrier Reef showed that very closely related sub-cladal symbiont types not only represent distinct evolution-ary lineages, but also occupy a distinct niche within theirhost (Sampayo et al. 2007). Subcladal types have also beenshown to be physiologically distinct and can determinehost ranges and performance (Iglesias-Prieto et al. 1992,2004; Warner et al. 2006). While heterogeneity within theribosomal array may complicate the study of symbiontdiversity, particularly with more variable DNA markers(Thornhill et al. 2007), the patterns linking subcladal sym-biont types with ecological diversiWcation or to speciWchosts are not explained by this heterogeneity (LaJeunesse2005; Sampayo et al. 2007). Most of the symbiont typesdescribed in the present study have not previously beenfound in any other host species and a global analysis of
symbionts from pocilloporid species indicates that IndianOcean symbionts represent a distinct group from theirPaciWc counterparts (Sampayo et al., in preparation). Thisis congruent with other studies that identify the IndianOcean as a distinct biogeographical province (e.g. Randall1998; Rocha et al. 2007).
Conclusion
Symbiotic associations between invertebrate hosts andSymbiodinium are predominant in tropical seas, but symbi-otic associations are also found in environments wheretemperature and light may not be optimal (e.g. Hoegh-Guldberg and Hinde 1986; Rodriguez-Lanetty et al. 2001).These high latitude communities have been suggested to bemore vulnerable to climate change because they occur atthe end of range margins and often represent small, isolatedpopulations (Hughes et al. 2002). This study provides theWrst subcladal assessment of Symbiodinium diversity in theWestern Indian Ocean and provides the Wrst information onSymbiodinium diversity in South Africa. There is a clearshift in symbiont subcladal types at the latitudinal extent ofS. pistillata’s range in South Africa, which is likely drivenby the interplay of changing temperature and irradiance.Finally, this study highlights that potentially important bio-geographic and ecological patterns may be overlookedwhen employing cladal designations and reinforces theneed for follow up physiological studies to establish whycertain cladal or subcladal types are preferred at higherlatitudes.
Acknowledgments Thanks are extended to Prof. Ove Hoegh-Guld-berg for Wnancial support and laboratory usage, Dr. M. Meusel for useof laboratory facilities, Dr. L. Celliers for manuscript feedback, andSAAMBR and the NRF for an MSc scholarship to AM.
References
Atoda K (1947) The larva and postlarval development of some reef-building corals. II. Stylophora pistillata (Esper). Science ReportsTohoku University 4th Series (Biol) 18:48–65
Baker AC (2003) Flexibility and speciWcity in coral–algal symbiosis:diversity, ecology, and biogeography of Symbiodinium. AnnuRev Ecol Ev Syst 34:661–689
Baker AC, Starger C, McClanahan T, Glynn P (2004) Corals’ adaptiveresponse to climate change. Nature 430:741
Berg H, Francis J, Souter P (2002) Support to marine research for sus-tainable management of marine and coastal resources in the West-ern Indian ocean. Ambio 31:597–601
Burnett WJ (2002) Longitudinal variation in algal symbionts (zooxan-thellae) from the Indian ocean zoanthid Palythoa caesia. MarEcol Prog Ser 234:105–109
Celliers L, Schleyer MH (2002) Coral bleaching on high-latitude mar-ginal reefs at Sodwana Bay, South Africa. Mar Pollut Bull44:1380–1387
123

Mar Biol
Celliers L, Mann BQ, Macdonald AHH, Schleyer MH (2007) A ben-thic survey of the rocky reefs oV Pondoland, South Africa. Afr JMar Sci 29:65–77
Done TJ (1999) Coral community adaptability to environmentalchange at the scales of regions, reefs, and reef zones. Am Zool39:66–79
Douglas AE (1998) Host beneWt and the evolution of specialization insymbiosis. Heredity 81:599–603
Fabricius KE, Mieog JC, Colin PL, Idip D, van Oppen MJH (2004)Identity and diversity of coral endosymbionts (zooxanthellae)from three Palauan reefs with contrasting bleaching, temperatureand shading histories. Mol Ecol 13:2445–2458
Hoegh-Guldberg O, Hinde R (1986) Studies on a nudibranch that con-tains zooxanthellae I. Photosynthesis, respiration and the translo-cation of newly Wxed carbon by zooxanthellae in Pteraeolidiaianthina. Proc R Soc Lond B 228:493–509
Hughes TP, Bellwood DR, Connolly SR (2002) Biodiversity hotspots,centres of endemicity, and the conservation of coral reefs. EcolLett 5:775–784
Hunter RL, LaJeunesse TC, Santos SR (2007) Structure and evolutionof the rDNA internal transcribed spacer (ITS) region 2 in the sym-biotic dinoXagellates (Symbiodinium, Dinophyta). J Phycol43:120–128
Iglesias-Prieto R, Matta JL, Robins WA, Trench RK (1992) Photosyn-thetic response to elevated-temperature in the symbiotic dinoXa-gellate Symbiodinium microadriaticum in culture. Proc Natl AcadSci USA 89:10302–10305
Iglesias-Prieto R, Beltran VH, LaJeunesse TC, Reyes-Bonilla H,Thome PE (2004) DiVerent algal symbionts explain the verticaldistribution of dominant reef corals in the eastern PaciWc. Proc RSoc B 271:1757–1763
Kleypas JA, McManus JW, Menez LAB (1999) Environmental limitsto coral reef development: where do we draw the line? Am Zool39:146–159
LaJeunesse TC (2002) Diversity and community structure of symbioticdinoXagellates from Caribbean coral reefs. Mar Biol 141:387–400
LaJeunesse TC (2005) Species radiations of symbiotic dinoXagellatesin the Atlantic and Indo-PaciWc since the Miocene–Pliocene tran-sition. Mol Biol Evol 22:570–581
LaJeunesse TC, Trench RK (2000) Biogeography of two species ofSymbiodinium (Freudenthal) inhabiting the intertidal sea anem-one Anthopleura elegantissima (Brandt). Biol Bull 199:126–134
LaJeunesse TC, Loh WKW, van Woesik R, Hoegh-Guldberg O,Schmidt GW, Fitt WK (2003) Low symbiont diversity in southernGreat Barrier Reef corals, relative to those of the Caribbean. Lim-nol Oceanogr 48:2046–2054
LaJeunesse TC, Bhagooli R, Hidaka M, deVantier L, Done T, SchmidtGW, Fitt WK, Hoegh-Guldberg O (2004) Closely related Symbi-odinium spp. diVer in relative dominance in coral reef host com-munities across environmental, latitudinal and biogeographicgradients. Mar Ecol Prog Ser 284:147–161
Lien Y-T, Nakano Y, Plathong S, Fukami H, Wang J-T, Chen CA(2007) Occurrence of the putatively heat-tolerant Symbiodiniumphylotype D in high-latitudinal outlying coral communities. CoralReefs 26:35–44
Loh WKW, Loi T, Carter D, Hoegh-Guldberg O (2001) Geneticvariability of the symbiotic dinoXagellates from the wide rangingcoral species Seriatopora hystrix and Acropora longicyathus inthe Indo-West PaciWc. Mar Ecol Prog Ser 222:97–107
McClanahan TR, Maina J, Moothien-Pillay KR, Baker AC (2005)EVects of geography, taxa, water Xow, and temperature variationon coral bleaching intensity in Mauritius. Mar Ecol Prog Ser298:131–142
Macdonald AHH (2005) The tramp coral, Stylophora pistillata, in thesouth–west Indian Ocean: ecomorph or speciation. MSc thesis,University of KwaZulu-Natal, South Africa
Randall JE (1998) Zoogeography of shore Wshes of the Indo-PaciWcregion. Zool Stud 37:227–269
Reynolds RW, Smith TM (1994) Improved global sea surface temper-ature analyses. J Clim 7:929–948
Ridgway T, Sampayo EM (2005) Population genetic status of theWestern Indian Ocean: What do we know? WIO J Mar Sci 4:1–9
Ridgway T, Hoegh-Guldberg O, Ayre DJ (2001) Panmixia in Pocillo-pora verrucosa from South Africa. Mar Biol 139:175–181
Ridgway T, Riginos C, Davis J, Hoegh-Guldberg O (2007) Pocillo-pora verrucosa genetic connectivity patterns in Southern AfricanMarine Protected Areas. Mar Ecol Prog Ser In press
Rocha LA, Craig MT, Bowen BW (2007) Phylogeography and theconservation of coral reef Wshes. Coral Reefs 26:501–512
Rodriguez-Lanetty M, Loh WKW, Carter D, Hoegh-Guldberg O(2001) Latitudinal variability in symbiont speciWcity within thewidespread scleractinian coral Plesiastrea versipora. Mar Biol138:1175–1181
Rowan R, Powers DA (1991) Molecular genetic identiWcation of sym-biotic dinoXagellates (zooxanthellae). Mar Ecol Prog Ser 71:65–73
Rowan R, Knowlton N (1995) IntraspeciWc diversity and ecologicalzonation in coral algal symbiosis. Proc Natl Acad Sci USA92:2850–2853
Sambrook J, Russell DW (2001) Molecular cloning: a laboratory man-ual 3rd edn. Cold Spring Harbor Laboratory Press, New York
Sampayo EM, Franceschinis L, Hoegh-Guldberg O, Dove S (2007)Niche diversiWcation of closely related symbiotic dinoXagellates.Mol Ecol 16:3721–3733
SwoVord DL (1999) PAUP*, phylogenetic analysis using parsimony(*and other methods), version 4.0b10, Sinauer Associates, Sun-derland
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG(1997) The ClustalX windows interface:Xexible strategies formultiple sequence alignment aided by quality analysis tools.Nucleic Acids Res 24:4876–4882
Thornhill DJ, LaJeunesse TC, Kemp DW, Fitt WK, Schmidt GW(2006) Multi-year, seasonal genotypic surveys of coral–algalsymbioses reveal prevalent stability or post-bleaching reversion.Mar Biol 148:711–722
Thornhill DJ, LaJeunesse TC, Santos SR (2007) Measuring rDNAdiversity in eukaryotic microbial systems:how intragenomic var-iation, pseudogenes, and PCR artifacts confound biodiversityestimates. Mol Ecol 16:5326–5340
Toller WW, Rowan R, Knowlton N (2001) Zooxanthellae of the Mon-tastraea annularis species complex: patterns of distribution offour taxa of Symbiodinium on diVerent reefs and across depths.Biol Bull 201:348–359
van Oppen MJH, Palstra FP, Piquet AMT, Miller DJ (2001) Patterns ofcoral–dinoXagellate associations in Acropora:signiWcance oflocal availability and physiology of Symbiodinium strains andhost–symbiont selectivity. Proc R Soc Lond B 268:2617–2617
van Oppen MJH, Mahiny AJ, Done TJ (2005) Geographic distributionof zooxanthella types in three coral species on the Great BarrierReef sampled after the 2002 bleaching event. Coral Reefs24:482–487
Veron JEN (1993) A biogeographic database of hermatypic corals:Species of the Central Indo-PaciWc, Genera of the World.Australian Institute of Marine Science Monograph Series 10.433 pp
Visram S, Douglas AE (2006) Molecular diversity of symbiotic algae(zooxanthellae) in scleractinian corals of Kenya. Coral Reefs25:172–176
Warner ME, LaJeunesse TC, Robison JE, Thur RM (2006) The ecolog-ical distribution and comparative photobiology of symbiotic dino-Xagellates from reef corals in Belize: potential implications forcoral bleaching. Limn Oceanogr 51:1887–1897
123





























