Embed Size (px)
Citation preview

Page 1/19
Iso�avone consumption reduces in�ammationthrough modulation of phenylalanine and lipidmetabolismRachel L. Shrode
University of IowaNicole Cady
University of IowaSamantha N. Jensen
University of IowaNicholas Borcherding
Washington UniversityAshutosh K. Mangalam ( [email protected] )
University of Iowa
Research Article
Keywords: phytoestrogen, iso�avone, in�ammation, untargeted metabolomics, microbiome, multiplesclerosis
Posted Date: July 21st, 2022
DOI: https://doi.org/10.21203/rs.3.rs-1871117/v1
License: This work is licensed under a Creative Commons Attribution 4.0 International License. Read Full License

Page 2/19
AbstractIntroduction
Phytoestrogen found in soy, fruits, peanuts, and other legumes, has been identi�ed as a metabolitecapable of providing bene�cial effect in multiple pathological conditions due to its ability to mimicendogenous estrogen. Interestingly, the health promoting effect of phytoestrogens such as iso�avone isdependent on the presence of gut bacteria with ability to metabolize dietary phytoestrogen intometabolite such as equol which have higher a�nity for endogenous estrogen receptors. We havepreviously shown that patients with multiple sclerosis (MS), a neuroin�ammatory disease lack gutbacteria with the ability to metabolize phytoestrogen. We have validated the importance of phytoestrogenand gut bacteria in disease protection utilizing an animal model of MS. Speci�cally, we have shown thatan iso�avone rich diet can protect from neuroin�ammatory diseases and that protection was dependenton the ability of gut bacteria to metabolize iso�avone into equol. Additionally, mice on a diet withiso�avone showed an anti-in�ammatory response compared to the mice on a phyto-free diet. However, itis unknown how iso�avone and/or equol mediates its protective effect especially the effect of iso�avoneon host metabolomics.
Objectives
In this study we utilized untargeted metabolomics to identify the metabolites modulated by the presenceof dietary iso�avone.
Results
We found that the consumption of iso�avone increases anti-in�ammatory monounsaturated fatty acidsand bene�cial polyunsaturated fatty acids while reducing pro-in�ammatory glycerophospholipids,sphingolipids, phenylalanine metabolism, and arachidonic acid derivatives.
Conclusion
Iso�avone consumption alters the systemic metabolic landscape through resultant increase inmonounsaturated fatty acids and bene�cial polyunsaturated fatty acids while reducing proin�ammatorymetabolites and pathways. This highlights a potential mechanism by which an iso�avone diet modulateimmune mediated disease.
1. IntroductionRecently, diet has emerged as an important regulator of human health. Aside from nutrition it providesnumerous bioactive molecules such as polyphenols, �avonoids, phytoestrogens, carotenoids, andvitamins necessary for maintaining a healthy state (Sun et al., 2019). Trillions of bacteria (microbiota)living in our gut play an important role in mediating the bene�cial effects of dietary compounds as theyhelp in breaking down original plant compounds into more potent metabolites (Jensen et al., 2021). Thus,

Page 3/19
consuming a healthy diet full of vegetables and fruits will enrich the growth of healthy gut microbiota,which in turn will help produce bene�cial metabolites necessary for maintaining health. For example,plant-based foods containing phytoestrogen compounds are thought to be bene�cial for human healthdue to their structural similarity with estrogens which allow them to bind and activate endogenousestrogen receptors (ERs). Phytoestrogen compounds present in plant-based food have weak a�nity forERs, but numerous bacteria present in the human gut have enzymes that can metabolize dietaryphytoestrogens into secondary metabolites to enhance their ability to bind and activate endogenous ERs(Aboushanab et al., 2021). Certain gut bacteria such as Parabacteroides, Adlercreutzia, Slackia,Lactobacillus and Bi�dobacterium genera can metabolize the iso�avone daidzein into S-equol which hashigher activity than daidzein itself (Mayo et al., 2019). For simplicity S-equol will be referred to as equolthroughout the rest of this work. The loss of these bacteria and absence of equol may create a void inwhich non-commensal bacteria may establish themselves and predispose to disease by inducing theproduction of pro-in�ammatory mediators.
Therefore, it is not surprising that multiple human diseases are linked with the loss of bene�cialmetabolites and gut microbiota (dysbiosis) which is characterized by an increase in pro-in�ammatorymediators plus pathogenic gut bacteria and a decrease in anti-in�ammatory mediators (Mangalam et al.,2021). Interestingly, we have previously shown that people with multiple sclerosis (MS), a chronicin�ammatory disease of the central nervous system lack gut bacteria with the ability to metabolizedietary phytoestrogen (Chen et al., 2016). Besides MS, iso�avone has been associated with protectionagainst other diseases such as cardiovascular disease, osteoporosis, prostate cancer, breast cancer,atherosclerosis, and diabetic conditions (Pabich et. al, 2019; Sathyapalan et al., 2018). Iso�avones arealso being studied as a treatment strategy for atrophy, menopause, and postmenopausal symptoms(Pabich et al., 2019).
We validated the importance of phytoestrogen metabolizing bacteria in MS utilizing experimentalautoimmune encephalomyelitis (EAE), an animal model of MS (Jensen et al., 2021). In this study weshowed that mice on an iso�avone diet developed milder disease compared to those on a phytoestrogen-free (phyto-free) diet. Most importantly, the iso�avone diet-mediated disease protection was dependenton the presence of phytoestrogen metabolizing gut bacteria speci�cally those lacking in patients with MS(pwMS). We also observed that compared to mice on a phyto-free diet, mice on an iso�avone dietdemonstrated lower in�ammatory responses as measured by antigen-speci�c T cell proliferation andcytokine levels (Jensen et al., 2021). In summary, published studies and our data established animportant role of an iso�avone diet in suppressing in�ammatory diseases (Aboushanab et al., 2021; Chenet al., 2016; Jensen et al., 2021; Mayo et al., 2019; Pabich et al., 2019; Sathyapalan et al., 2018).
Due to structural similarity of equol to endogenous estrogen, it can interact with ERs which are expressedin majority of mammalian cells and tissue (Warner et al., 2017). Thus, equol could have systemic effectbeyond the gut. Further, recent studies have also highlighted thousands of metabolites present in thehuman blood play an important role in maintaining the overall health by regulating numerousphysiological processes (François et al., 2021; Poisson et al., 2015; Rangel-Huerta et al., 2017; Zhang et

Page 4/19
al., 2015). Based on these data, we hypothesized that iso�avone and its metabolite equol may mediateits disease-protective effect through modulation of the host metabolome speci�cally the balance betweenpro and anti-in�ammatory metabolites.
Therefore, in this study we investigated whether mice on a diet with iso�avone had a distinct metabolitepro�le compared to mice on diet lacking iso�avone (and all other phytoestrogens). We observedsigni�cant differences in the metabolome compositions between the two diets. In fact, we observedenrichment of metabolites linked with a healthy state in mice on a diet with iso�avone and a consequentincrease in metabolites linked with disease state in mice on a phyto-free diet. Ours is the �rst study topro�le the metabolome using untargeted metabolomics in mice on a diet with or without iso�avone.These �ndings give us an insight into how iso�avone might provide protection from diseases throughalteration of speci�c metabolites which can be harnessed for development of future therapy for MS aswell as other in�ammatory diseases.
2. Materials And Methods
2.1 Experimental mice and diet treatmentC57BL/6J male mice (4–6 weeks old) were purchased from the Jackson Laboratories (Bar Harbor, ME)and allowed to adjust to their new environment for 1–2 weeks. Mice (n = 10) were then switched to a dietwith iso�avone [genistein (0.24 g/kg of diet) and daidzein (0.22 g/kg of diet)]] or on a diet without anyphytoestrogen (Envigo, Indianapolis, IN) ad libitum for 6 weeks as described previously (Jensen et al.,2021).
2.2 Serum sample collectionAt the end of the six-week diet, blood was collected, and the serum was separated by standard bloodclotting method from all mice. These serum samples were stored at -80℃ and shipped to Metabolon(Morrisville, NC) for metabolite identi�cation.
2.3 Untargeted metabolomicsMetabolic pro�ling was performed by Metabolon Inc. (Morrisville, NC) using proprietary methods aspreviously described by our group (Mangalam et al., 2013). In brief Metabolon Inc. extracts proteins andsmall molecules from the sample utilizing methanol and intense shaking followed by centrifugation. Formetabolite identi�cation, Metabolon then utilizes reverse-phase ultra-high performance chromatographymass spectrometry with negative and positive ion mode electrospray ionization and hydrophilicinteraction ultra-performance liquid chromatography mass spectrometry. Metabolon also ran severalcontrols along with our samples to assure quality measurements. Compounds were then identi�ed basedon their retention time/index (RI), mass to charge ratio (m/z), and chromatographic data (MS/MSspectral data) by comparison to a library of compounds. The output of the work�ow was a data tablecontaining the abundance of each metabolite present in each sample.

Page 5/19
2.4 Statistical analysis of metabolitesBoth MetaboAnalyst 5.0 (Pang et al., 2021)d (R Core Team, 2022) (version 4.1.1) were utilized forstatistical analyses of the metabolites observed in the serum samples from both groups. Overall datawas �rst �ltered based on missing values. If a given metabolite had missing values for more than 50% ofthe samples in each group, it was removed. All other missing values were �lled with the minimum valuefor the given metabolite. Data were then normalized by sum scaling and log (base 10) transforming. Forunivariate testing we utilized a Wilcoxon Rank Sum test and the p-values were adjusted by utilizingBenjamini-Hochberg procedure. Results were considered signi�cant if the p-value < 0.05 and the adjustedp-value was < 0.1. Univariate tests were further visualized utilizing ggplot2 (Hadley Wickham, 2016).Pathway analysis utilized Holm-Bonferroni for a stricter p-value adjustment. As this correction is stricter,we consider a Holm’s p < 0.20 signi�cant.
3. Results
3.1 Iso�avone alters the metabolomeFirst, we generated a Principal Coordinate Analysis (PCA) plot to determine whether iso�avone affectedthe overall composition of the metabolome (Fig. 1). Based on the PCA, the �rst and second PCcomponents explained 36.2% and 15.8% of the variance, respectively, between the two diets. The mice ona diet with iso�avone clustered separately from mice on a phyto-free diet suggesting the metabolitecomposition was distinct between the two diets.
We next performed hierarchical clustering based on the Euclidean distance of the top 25 features (Fig. 2).The heatmap generated by this analysis showed distinct groupings of metabolites based on diet.Speci�cally, 8 of the 25 metabolites were increased and 17 were decreased in the iso�avone diet groupcompared to the phyto-free diet. The metabolites found to be increased in the iso�avone group wereequol sulfate, 4-ethylphenyl sulfate, linolenate (18:3n3), palmitoleate (16:1n7), linoleate (18:2n6), 10-heptadecenoate, oleate/vaccenate (18:1), and palmitate (16:0). The metabolites found to be decreased iniso�avone were p-cresol sulfate, 1-(1-enyl-palmitoyl)-2-palmitoyl-GPC (P-16:0/16:0), hydroxyproline, 2-amino-p-cresol sulfate, phenylalanine, 1-stearoyl-2-docosahexaenoyl-GPE, sphingosine 1-phosphate, 1-oleoyl-2-docosahexaenoyl-GPE, 1-oleoyl-2-arachidonoyl-GPE, sphingomyelin (d18:1/20:0, d16:1/22:0),sphingomyelin (d18:1/22:1, d18:2/22:0, d161/24:1), 1-2-dipalmitoyl-GPC, stearoyl sphingomyelin,adenosine, sphingomyelin (d18:2/24:1/18:0), 1-palmitoyl-2-docosahexaenoyl-GPC, 1-stearoyl-2-linoleoyl-GPCF. Thus, hierarchical clustering-based analysis suggested that presence of iso�avone in diet had aneffect on host metabolites as highlighted by enrichment or depletion of speci�c metabolites.
3.2 Individual metabolites are altered on an iso�avone dietTo identify metabolite-based biomarkers speci�c to the iso�avone diet, we used the nonparametricWilcoxon Rank Sum test. While only considering metabolites signi�cant if their p-value was < 0 .05 and

Page 6/19
their adjusted p-value (FDR) was < 0.10, we found 132 signi�cantly different metabolites. 109 of thesemetabolites were signi�cantly increased while 23 were decreased in the iso�avone diet compared to thephyto-free diet.
All 8 metabolites found to be increased in the heatmap were also identi�ed by univariate test assigni�cantly increased: equol sulfate, 4-ethylphenyl sulfate, linolenate (18:3n3), palmitoleate (16:1n7),linoleate (18:2n6), 10-heptadecenoate, oleate/vaccenate (18:1), and palmitate (16:0) (Fig. 3A-H).
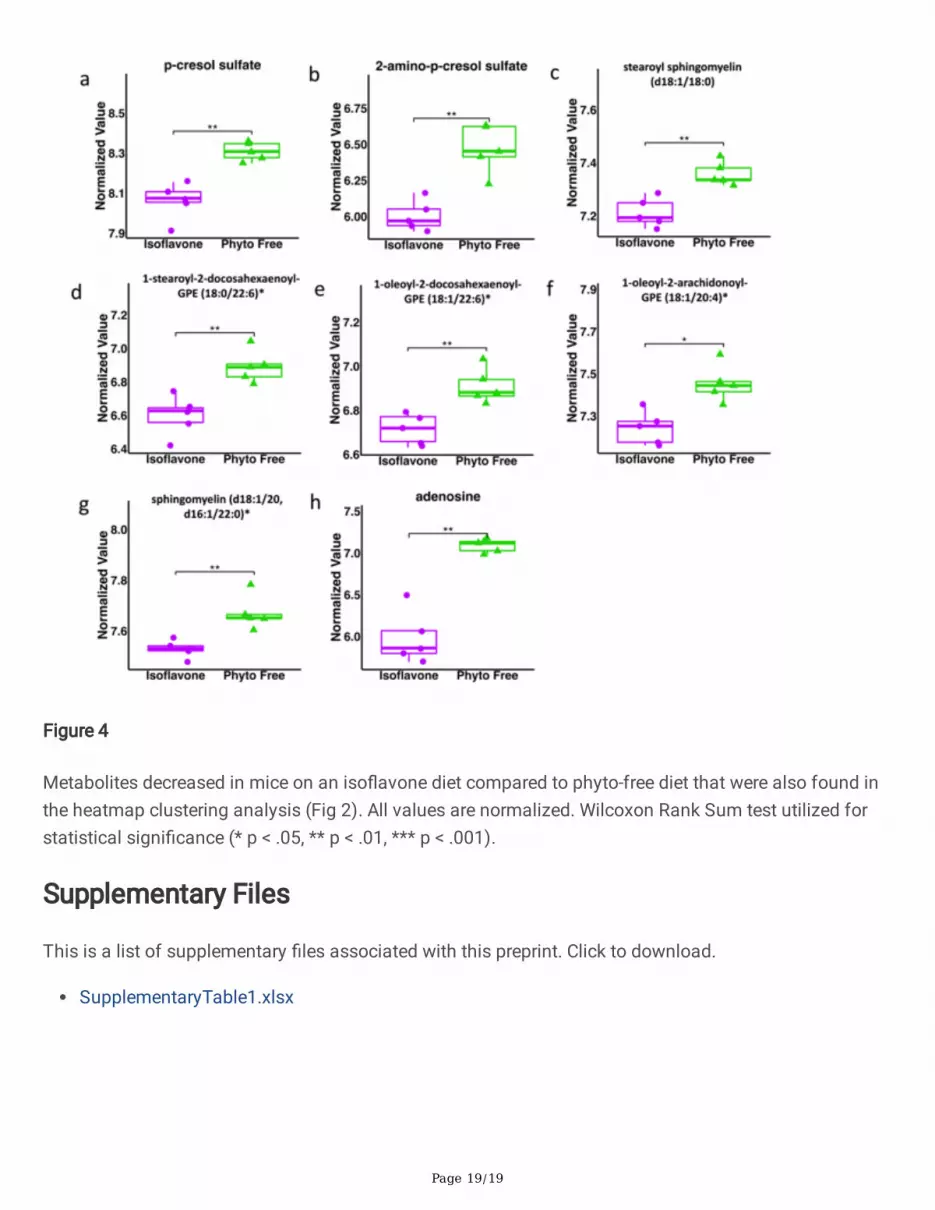
The univariate test identi�ed 8 of the 17 metabolites previously found in the heatmap analysis assigni�cantly decreased in the iso�avone-consuming mice. These metabolites include p-cresol sulfate, 2-amino-p-cresol sulfate, 1-stearoyl-2-docosahexaenoyl-GPE, sphingomyelin (d18:1/20:0, d16:1/22:0), 1-oleoyl-2-docosahexaenoyl-GPE, stearoyl sphingomyelin, adenosine, and 1-oleoyl-2-arachidonoyl-GPE(18:1/20:4) (Fig. 4A-H). Signi�cant values are in Table 1. Similar to hierarchical clustering-based analysis,univariate test also suggested that dietary iso�avone in�uence overall host metabolite composition withmodulation of speci�c metabolites.

Page 7/19
Table 1Individual metabolites altered in mice on an iso�avone diet. This table contains the metabolites
signi�cantly altered in both the heatmap clustering and univariate analysis. The adjusted p-value wascalculated utilizing the Benjamini-Hochberg procedure.
Metabolite p-value
BH adjusted p-value
Iso�avone diet vs Phyto-freediet
Equol sulfate .0079 .0804 Increased
4-ethylphenyl sulfate .0079 .0804 Increased
Linolenate (18:3n3) .0079 .0804 Increased
Linoleate (18:2n6) .0079 .0804 Increased
10-heptadecenoate .0079 .0804 Increased
oleate/vaccenate (18:1) .0079 .0804 Increased
Palmitate (16:0) .0079 .0804 Increased
Palmitoleate (16:1n7) .0079 .0804 Increased
P-cresol sulfate .0079 .0804 Decreased
2-amino-p-cresol sulfate .0079 .0804 Decreased
1-stearoyl-2-docosahexaenoyl-GPE .0079 .0804 Decreased
sphingomyelin (d18:1/20:0,d16:1/22:0)
.0079 .0804 Decreased
1-oleoyl-2-docosahexaenoyl-GPE, .0079 .0804 Decreased
stearoyl sphingomyelin .0079 .0804 Decreased
adenosine .0079 .0804 Decreased
1-oleoyl-2-arachidonoyl-GPE(18:1/20:4)
.0159 .0962 Decreased
3.3 Metabolic pathways are altered on an iso�avone dietAs multiple metabolites varied between plasma from mice on a diet with or without iso�avone, we nextinvestigated speci�c metabolic pathways affected by the diet. This analysis was performed withMetaboAnalyst, and the pathway analysis parameters included the Global Test for the enrichmentmethods and the Relative-betweenness Centrality for the topology analysis. The KEGG Mus musculuspathway library was chosen for identi�cation. The pathways were considered signi�cant if they had a p-value < 0.05 and a Holm’s p < 0.20. With these parameters we found 15 pathways signi�cantly altered:Purine Metabolism, Selenocompound Metabolism, Tryptophan Metabolism, Thiamine Metabolism,Aminoacyl-tRNA Biosynthesis, Galactose Metabolism, Arachidonic Acid Metabolism, Linoleic AcidMetabolism, Alpha-Linolenic Acid Metabolism, Starch and Sucrose Metabolism, Phenylalanine

Page 8/19
Metabolism, Vitamin B6 Metabolism, Phosphatidylinositol Signaling System, Glutathione Metabolism,and Sphingolipid Metabolism. These pathways and their signi�cant values can be seen in Table 2. Thus,the pathway analysis showed that presence of iso�avone in diet resulted in modulation of speci�cmetabolic pathways.
Table 2Metabolic pathways altered in mice on an iso�avone diet. This tablecontains the pathways signi�cantly altered between iso�avone andphyto-free diet based on the Global Test. The adjusted p-value was
calculated utilizing the Holm-Bonferroni method.Pathway p-value Holm’s p-value
Purine Metabolism . 00024 .0127
Selenocompound Metabolism .00042 .0127
Tryptophan Metabolism .0012 .024
Thiamine Metabolism .0028 .0410
Aminoacyl-tRNA Biosynthesis .0056 .0410
Galactose Metabolism .0062 .0410
Arachidonic Acid Metabolism .0068 .0410
Linoleic Acid Metabolism .0068 .0410
Alpha-linoleic Acid Metabolism .0068 .0410
Starch and Sucrose Metabolism .0068 .0410
Phenylalanine Metabolism .00836 .0456
Vitamin B6 Metabolism .012 .062
Phosphatidylinositol Signaling System .0324 .141
Glutathione Metabolism .0328 .141
Sphingolipid Metabolism .0453 .171
4. DiscussionThe signi�cance of diet and gut microbiota in the pathobiology of MS is increasingly appreciated,speci�cally their potential as novel therapeutic agents (Bjørnevik et al., 2017; Yu et al., 2022; Zahoor et al.,2021). We and others have previously shown that MS patients have gut dysbiosis with depletion ofbene�cial gut bacteria, notably those able to metabolize phytoestrogens (Chen et al., 2016; Storm-Larsenet al., 2019). Further, using the animal model of MS, we have shown that an iso�avone (phytoestrogen)rich diet can protect mice from EAE, but this protection was dependent on the presence of speci�c gutbacteria and their ability to metabolize dietary phytoestrogen into equol (Jensen et al., 2021). In this study

Page 9/19
we performed untargeted metabolomics in the sera of mice on a diet with or without iso�avone and haveshown enrichment or reduction of speci�c metabolites in mice on a diet with or without iso�avone.
We observed that iso�avone diet affected host metabolism as highlighted by a distinct metabolitecomposition in mice on a diet with or without iso�avone. Several studies have shown that the bene�cialeffects of a diet or a dietary supplement are associated with global change in host metabolome (Díaz-Rubio et al., 2015; Rangel-Huerta et al., 2017; Torres Santiago et al., 2019). Healthy adults consumingantioxidant rich juice (n = 28) (Díaz-Rubio et al., 2015) or consuming orange juice (n = 30) (Rangel-Huertaet al., 2017) showed a distinct metabolic pro�le compared to the baseline. Similarly, Santiago et al.reported a change in urinary and fecal metabolite in Wistar rats consuming oral resveratrol compared tothe water group (Torres Santiago et al., 2019). Thus, our data showing a distinct metabolic pro�le in miceon an iso�avone diet is in accordance with previous studies reporting that dietary supplement can have aglobal effect on host metabolites.
Heat map clustering analysis, which measures the samples’ Euclidean distances from one another,showed modulation of the top 25 metabolites; with 8 being increased and 17 being decreased in mice onthe iso�avone diet compared to the phyto-free diet. Using Wilcoxon Signed Rank Test for a univariateanalysis, we identi�ed 132 metabolites to be signi�cantly different between these two groups.Speci�cally, there were 110 metabolites signi�cantly increased and 22 metabolites signi�cantlydecreased on the iso�avone diet compared to the phyto-free diet. Of note, equol sulfate was the mostincreased of any metabolite, and this is expected as iso�avone is broken down into equol which is thenconverted into equol sulphate. This �nding serves as a positive control that our diets did in fact differ bypresence of iso�avone which was metabolized into equol by gut bacteria (Díaz-Rubio et al., 2015).Analysis of speci�c pathways showed that several pathways were different between the two groups,speci�cally phenylalanine metabolism, linoleic acid metabolism, alpha-linolenic acid metabolism,sphingomyelin metabolism, and arachidonic acid metabolism.
Numerous signi�cantly altered metabolites were linked to phenylalanine (Phe) metabolism. Phe is anaromatic amino acid, which is essential for the host and obtained from diet as the host does not naturallyproduce it. Dietary Phe is broken down into Tyrosine (Tyr) or 4-hydroxyphenylpyruvic acid (Gryp et al.,2017). Both of these metabolites can further be broken down into 4-ethylphenyl sulfate, p-cresol sulfate,and p-cresol glucuronide (Gryp et al., 2017). Phenylalanine was decreased in the iso�avone diet alongwith its secondary metabolites p-cresol sulfate (PCS) and p-cresol glucuronide (PCG). PCS and PCG arebroken down from Phe and Tyr through anaerobic intestinal bacteria (Wishart et al., 2022). Interestingly,pwMS have higher levels of PCS in plasma as well as cerebrospinal �uids and PCS can induce axonaldamage in cultured neurons (Ntranos et al., 2022). Additionally, pwMS treated with dimethyl fumarateshowed decreased levels of PCS compared to untreated group (Ntranos et al., 2022). PCS had also beenproposed as a biomarker for autism spectrum disorder (ASD) as its levels are elevated in the urine andfeces of children with ASD (el Aidy et al., 2021). The current understanding is that PCS passes the blood-brain-barrier and affects microglia and neurons to induce neuroin�ammation and axonal damages in

Page 10/19
neurological diseases such as MS and ASD. Thus, iso�avone may reduce neuroin�ammation bymodulation of phenylalanine metabolism speci�cally by reduction in the level of pro-in�ammatory PCS.
Further investigation of the signi�cant metabolites revealed iso�avone had altered fatty acid metabolism.Firstly, we found palmitoleate, oleate, linoleate, and linolenate were all signi�cantly increased iniso�avone diet. Palmitoleate is the salt form of the monounsaturated fatty acid, palmitoleic acid, and ismodulated in several in�ammatory diseases but depending on the disease, palmitoleate levels had beenshown to be either increased or decreased (Akazawa et al., 2021). Oleate is also the salt form of amonounsaturated fatty acid, oleic acid, which had shown to possess anti-in�ammatory activity. In fact,fatty tissue from pwMS lack oleic acid and it is suggested to be responsible for regulatory T cell (Treg)dysfunction (Pompura et al., 2021). The exposure of Tregs from pwMS and healthy controls (HC) witholeic acid in culture restored suppressive function of these Tregs (Pompura et al., 2021). Additionally,presence of both palmitoleate and oleate had been linked with reduced in�ammation in skeletal musclecells (Coll et al., 2008). Thus, iso�avone may also mediate its anti-in�ammatory effect through inductionof the monounsaturated fatty acids palmitoleate and oleate both of which had been linked with thehealthy state of the host.
Additionally, linoleate and linolenate, both polyunsaturated fatty acids (PUFAs) and the salt form oflinoleic acid and linolenic acid, respectively, were increased in the mice on an iso�avone diet. Metabolismof linoleic acid and alpha-linolenic acid (ALA) were also found to be pathways signi�cantly modulatedbased on our pathway analysis. PUFAs have been linked to protection from many in�ammatory diseasesincluding MS. A study evaluating effects of dietary intake of PUFA and MS risk showed that among allPUFA tested, only ALA showed inverse association with MS risk (Bjørnevik et al., 2017). In a follow-upstudy, the group reported that the higher serum levels of ALA were associated with lower disease activityin pwMS (Bjornevik et al., 2019). Additionally, a recent study had shown that pwMS with high ALA levelsshowed reduced risk of relapse and longer time to convert from clinically isolated syndrome to clinicallyde�nite multiple sclerosis (Munger et al., 2022). Utilizing an animal model of MS, we have also identi�edmetabolites linked with PUFA, such as ALA and linoleic acid as signi�cant biomarkers of disease(Mangalam et al., 2013; Poisson et al., 2015). Additionally, we showed that resolvin D1 (RvD1), adownstream metabolite of ALA, ameliorated disease in a chronic EAE model (Poisson et al., 2015; Zahooret al., 2021). Thus, increased levels of bene�cial PUFAs in mice on a diet with iso�avone suggest thatPUFAs may have a role in creating an anti-in�ammatory environment due to the consumption ofiso�avone.
As for fatty acids that were depleted on an iso�avone diet compared to phyto-free, glycerophospholipidsand sphingolipids from Phosphatidylcholine (PC) metabolism and ceramide metabolism weresigni�cantly decreased. Speci�cally, the glycerophospholipids involved in PC metabolism that weredepleted in the iso�avone group include 1-stearoyl-2-docosahexaenoyl-GPE, 1-palmitoyl-2-docosahexaenoyl-GPC, 1-2-dipalmitoyl-GPC, 1-stearoyl-2-linoleoyl-GPC, and 1-oleoyl-2-docosahexaenoyl-GPE (Wishart et al., 2022). A study on the cerebral spinal �uid of MS and non-MS patients found that 4 ofthe 6 glycerophospholipids analyzed were signi�cantly increased in MS patients (Nogueras et al., n.d.).

Page 11/19
Suggesting that glycerophospholipid composition alteration contributes to the disease state and thus thesuppression of these metabolites on a diet with iso�avone supports an anti-in�ammatory role ofiso�avone.
PC also contributes to the production of sphingolipids with the help of ceramides (Li et al., 2008).Ceramides are lipid molecules that are metabolized into sphingolipids, namely sphingomyelin andsphingosine, which can be further broken down into sphingosine-1-phosphate (Hannun et al., 2010).Many structures of sphingomyelin, stearoyl sphingomyelin, and sphingosine-1-phosphate were found tobe signi�cantly decreased on the iso�avone diet. Additionally, sphingolipid metabolism was found to be adistinguishing pathway between our two diets based on our pathway analysis. Sphingomyelins are foundin animal cell membranes and most abundantly in the myelin sheath protecting nerve cell axons (Jana &Pahan, 2010). Sphingosine-1-phosphate is a signaling molecule also found in animal cell membranes arepresent during in�ammation (Spiegel et al., 2011). In fact, sphingosine 1-phosphate is activated by pro-in�ammatory cytokines and is involved in T-cell tra�cking. One MS treatment involves the use ofsphingosine 1-phosphate receptor modulators to mediate its bene�cial effect in pwMS (Mcginley et al.,2021). Due to the reduction in PC metabolism and these ceramide derivatives, our results suggestiso�avone promotes an anti-in�ammatory environment by reducing the metabolism ofglycerophospholipids and sphingolipids.
Another lipid alteration found on the iso�avone diet was a reduction in the arachidonic acid derivative, 1-oleoyl-2-arachidonoyl-GPE. Arachidonic acid was also identi�ed as a signi�cant pathway in our pathwayanalysis. Arachidonic acid metabolism produces prostaglandins (Jewell et al., 2022), which are lipids thatare produced in damaged and infected tissues to control in�ammation, clotting, and overall blood �ow(Ricciotti et al., 2011). The reduction of arachidonic acid metabolism in mice on iso�avone diet suggeststhat the iso�avone diet can reduce in�ammation through suppression of pro-in�ammatory arachidonicmetabolism.
In conclusion, dietary iso�avone alters the host metabolome by promoting anti-in�ammatory metabolitesand suppressing pro-in�ammatory metabolites. Speci�cally, we found that the major alterations wereseen in phenylalanine metabolism and lipid metabolism. Several monounsaturated and bene�cial PUFAswere increased on an iso�avone diet compared to phyto-free. Whereas several pro-in�ammatorymetabolites, including glycerophospholipids, sphingolipids, and fatty acids derived from arachidonic acid,were all depleted in the iso�avone diet. The pathways that were important in distinguishing the two dietsdirectly relate to these �ndings as the metabolism of linoleic acid, ALA, phosphatidylcholine, ceramide,sphingomyelin, and arachidonic acid were all considered signi�cant based on our pathway analysis.These pathways, and many of their metabolites, have been associated with in�ammatory diseases asaforementioned. Overall, these results indicate that dietary iso�avone induces changes in the systemicmetabolome potentially by alteration of gut microbiome and/or host response. Our study provides arationale for future studies investigating the mechanism through which dietary iso�avone modulate hostmetabolic network speci�cally phenylalanine and lipid metabolism.

Page 12/19
DeclarationsAcknowledgements and Funding: We acknowledge funding from the National Institutes of health/NIAID1RO1AI137075 (A.K.M), Veteran Affairs Merit Award 1I01CX002212 (A.K.M), University of IowaEnvironmental Health Sciences Research Center, NIEHS/NIH P30 ES005605 (A.K.M), Gift from P.Heppelmann and M. Wacek to A.K.M and Carver Trust Pilot Grant (A.K.M). R.L.S. was supported by theInformatics Fellowship from the University of Iowa. S.N.J was supported on an institutional training grant(T32AI007485 to G. Bishop) and diversity supplement award to A.K.M on parent 1RO1AI137075.
Author Contributions: Conceptualizations, Nicole Cady, Samantha N. Jensen, and Ashutosh K. Mangalam;methodology, Rachel L. Shrode., Nicole Cady, Samantha N. Jensen, Nick Borcherding, and Ashutosh K.Mangalam; validation, Rachel L. Shrode , Nick Borcherding, and Ashutosh K. Mangalam; formal analysis,investigation, resources, and data curation, Rachel L. Shrode; writing – original draft preparations RachelL. Shrode; writing – review and editing, Rachel L. Shrode, Nicole Cady, Samantha N. Jensen, NickBorcherding, and Ashutosh K. Mangalam; supervision Nick Borcherding, and Ashutosh K. Mangalam,funding acquisitions Ashutosh K. Mangalam. All authors have read and agreed to the published versionof the manuscript.
Institutional Review Board Statement: All procedures were done according to the Institutional Animal Careand Use Committee (IACUC) guidelines at the University of Iowa.
Data Availability Statement: Metabolomic data utilized for analysis is located in the supplementary dataas Table S1.
Con�icts of Interest: A.K.M. is one of the inventors of a technology claiming the use of Prevotella histicolato treat autoimmune diseases. A.K.M. received royalties from Mayo Clinic (paid by Evelo Biosciences).However, no fund or product from the patent were used in the present study. All other authors declare nocommercial or �nancial relationships that could be a potential con�ict of interest.
References1. Aboushanab, S. A., Khedr, S. M., Gette, I. F., Danilova, I. G., Kolberg, N. A., Ravishankar, G. A., Ambati,
R. R., & Kovaleva, E. G. (2021). Iso�avones derived from plant raw materials: bioavailability, anti-cancer, anti-aging potentials, and microbiome modulation. Critical Reviews in Food Science andNutrition. Ltd: Bellwether Publishing. https://doi.org/10.1080/10408398.2021.1946006
2. Akazawa, Y., Morisaki, T., Fukuda, H., Norimatsu, K., Shiota, J., Hashiguchi, K., Tabuchi, M., Kitayama,M., Matsushima, K., Yamaguchi, N., Kondo, H., Fujita, F., Takeshita, H., Nakao, K., & Takeshima, F.(2021). Signi�cance of serum palmitoleic acid levels in in�ammatory bowel disease. Scienti�cReports, 11(1), https://doi.org/10.1038/s41598-021-95923-6
3. Bjørnevik, K., Chitnis, T., Ascherio, A., & Munger, K. L. (2017). Polyunsaturated fatty acids and the riskof multiple sclerosis. Multiple Sclerosis, 23(14), 1830–1838.

Page 13/19
https://doi.org/10.1177/1352458517691150
4. Bjornevik, K., Myhr, K. M., Beiske, A., Bjerve, K. S., Holmøy, T., Hovdal, H., Midgard, R., Riise, T.,Wergeland, S., & Torkildsen, Ø. (2019). α-Linolenic acid is associated with MRI activity in aprospective cohort of multiple sclerosis patients. Multiple Sclerosis Journal, 25(7), 987–993.https://doi.org/10.1177/1352458518779925
5. Chen, J., Chia, N., Kalari, K. R., Yao, J. Z., Novotna, M., Soldan, M. M. P., Luckey, D. H., Marietta, E.,Jeraldo, P. R., Chen, X., Weinshenker, B. G., Rodriguez, M., Kantarci, O. H., Nelson, H., Murray, J. A., &Mangalam, A. K. (2016). Multiple sclerosis patients have a distinct gut microbiota compared tohealthy controls. Scienti�c Reports, 6. https://doi.org/10.1038/srep28484
�. Coll, T., Eyre, E., Rodríguez-Calvo, R., Palomer, X., Sánchez, R. M., Merlos, M., Laguna, J. C., & Vázquez-Carrera, M. (2008). Oleate reverses palmitate-induced insulin resistance and in�ammation in skeletalmuscle cells. Journal of Biological Chemistry, 283(17), 11107–11116.https://doi.org/10.1074/jbc.M708700200
7. Díaz-Rubio, M. E., Pérez-Jiménez, J., Martínez-Bartolomé, M., Álvarez, I., & Saura-Calixto, F. (2015).Regular Consumption of an Antioxidant-rich Juice Improves Oxidative Status and CausesMetabolome Changes in Healthy Adults. Plant Foods for Human Nutrition, 70(1), 9–14.https://doi.org/10.1007/s11130-014-0455-4
�. el Aidy, S., Burokas, A., Cristiano, C., Marzal, P. L., Pardo, P. P., Zheng, Y., Bek, M. K., Prince, N. Z.,Marzal, P., Garssen, L. N., Perez Pardo, J., P., & Kraneveld, A. D. (2021). The Role of Bacterial-DerivedAromatic Amino Acids Metabolites Relevant in Autism Spectrum Disorders: A Comprehensive Review.https://doi.org/10.3389/fnins.2021.738220
9. François, M., Karpe, A., Liu, J. W., Beale, D., Hor, M., Hecker, J., Faunt, J., Maddison, J., Johns, S.,Doecke, J., Rose, S., & Leifert, W. R. (2021). Salivaomics as a Potential Tool for Predicting Alzheimer’sDisease During the Early Stages of Neurodegeneration. Journal of Alzheimer’s Disease, 82, 1301–1313. https://doi.org/10.3233/JAD-210283
10. Gryp, T., Vanholder, R., Vaneechoutte, M., & Glorieux, G. (2017). p-cresyl sulfate. Toxins, 9(2),https://doi.org/10.3390/TOXINS9020052
11. Hadley Wickham (2016). ggplot2: Elegant Graphics for Data Analysis. In ggplot2: Elegant Graphicsfor Data Analysis (978th-3rd-319th-24277th–4th ed.)
12. Hannun, Y. A., Gault, C. R., & Obeid, L. M. (2010). Sphingolipids as Signaling and RegulatoryMolecules An Overview of Sphingolipid Metabolism: From Synthesis to Breakdown
13. Jana, A., & Pahan, K. (2010). Sphingolipids in Multiple Sclerosis. Neuromolecular Medicine.https://doi.org/10.1007/s12017-010-8128-4
14. Jensen, S. N., Cady, N. M., Shahi, S. K., Peterson, S. R., Gupta, A., Gibson-Corley, K. N., & Mangalam, A.K. (2021). Iso�avone diet ameliorates experimental autoimmune encephalomyelitis throughmodulation of gut bacteria depleted in patients with multiple sclerosis. In Sci. Adv (Vol. 7, Issue 9).https://www.science.org

Page 14/19
15. Jewell, D. E., & Jackson, M. I. (2022). Dietary Betaine and Fatty Acids Change Circulating Single-Carbon Metabolites and Fatty Acids in the Dog. Animals, 12(6),https://doi.org/10.3390/ani12060768
1�. Li, Z., & Vance, D. E. (2008). Phosphatidylcholine and choline homeostasis. In Journal of LipidResearch (Vol. 49, Issue 6, pp. 1187–1194). https://doi.org/10.1194/jlr.R700019-JLR200
17. Mangalam, A., Poisson, L., Nemutlu, E., Datta, I., Denic, A., Dzeja, P., Rodriguez, M., Rattan, R., & Giri, S.(2013). Pro�le of Circulatory Metabolites in an Animal Model of Multiple Sclerosis using GlobalMetabolomics. Journal of Clinical & Cellular Immunology, 04(03), https://doi.org/10.4172/2155-9899.1000150
1�. Mangalam, A., Yadav, M., & Yadav, R. (2021). The emerging world of microbiome in autoimmunedisorders: Opportunities and challenges. Indian Journal of Rheumatology (Vol, 16(1), 57–72.https://doi.org/10.4103/injr.injr_210_20. Wolters Kluwer Medknow Publications
19. Mayo, B., Vázquez, L., & Flórez, A. B. (2019). Equol: A bacterial metabolite from the Daidzeiniso�avone and its presumed bene�cial health effects. Nutrients, 11(9),https://doi.org/10.3390/NU11092231
20. Mcginley, M. P., & Cohen, J. A. (2021). Therapeutics Sphingosine 1-phosphate receptor modulators inmultiple sclerosis and other conditions. The Lancet, 398, 1184–1194.https://doi.org/10.1016/S0140-6736(21)00244-0
21. Munger, K., Bjornevik, K., Cortese, M., Edan, G., Freedman, M., Hartung, H. P., Montalban, X., Sandbrink,R., Radue, E. W., Barkhof, F., Wicklein, E. M., Kappos, L., & Ascherio, A. (2022). A Prospective Study ofSerum Levels of Polyunsaturated Fatty Acids and Effects on Multiple Sclerosis Disease Activity andProgression (S40.006). In Supplement) THURSDAY (Vol. 98, Issue 7)
22. Nogueras, L., Gonzalo, H., Jové, M., Sol, J., Gil-Sanchez, A., Hervás, J., Valcheva, P., Solana, M. J.,peralta, S., pamplona, R., & Brieva, L. (n.d.). Lipid pro�le of cerebrospinal �uid in multiple sclerosispatients: a potential tool for diagnosis. https://doi.org/10.1038/s41598-019-47906-x
23. Ntranos, A., Park, H. J., Wentling, M., Tolstikov, V., Amatruda, M., Inbar, B., Kim-Schulze, S., Frazier, C.,Button, J., Kiebish, M. A., Lublin, F., Edwards, K., & Casaccia, P. (2022). Bacterial neurotoxicmetabolites in multiple sclerosis cerebrospinal �uid and plasma. Brain, 145(2), 569–583.https://doi.org/10.1093/brain/awab320
24. Pabich, M., & Materska, M. (2019). Biological effect of soy iso�avones in the prevention ofcivilization diseases. Nutrients, 11(7), https://doi.org/10.3390/nu11071660
25. Pang, Z., Chong, J., Zhou, G., de Lima Morais, D. A., Chang, L., Barrette, M., Gauthier, C., Jacques, P., Li,S., & Xia, J. (2021). MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functionalinsights. Nucleic Acids Research, 49(W1), W388–W396. https://doi.org/10.1093/nar/gkab382
2�. Poisson, L. M., Suhail, H., Singh, J., Datta, I., Deni, A., Labuzek, K., Hoda, N., Shankar, A., Kumar, A.,Cerghet, M., Elias, S., Mohney, R. P., Rodriguez, M., Rattan, R., Mangalam, A. K., & Giri, S. (2015).Untargeted plasma metabolomics identi�es endogenous metabolite with drug-like properties in

Page 15/19
chronic animal model of multiple sclerosis. Journal of Biological Chemistry, 290(52), 30697–30712.https://doi.org/10.1074/jbc.M115.679068
27. Pompura, S. L., Wagner, A., Kitz, A., LaPerche, J., Yosef, N., Dominguez-Villar, M., & Ha�er, D. A. (2021).Oleic acid restores suppressive defects in tissue-resident FOXP3 Tregs from patients with multiplesclerosis. Journal of Clinical Investigation, 131(2), https://doi.org/10.1172/JCI138519
2�. R Core Team (2022). R: A Language and Environment for Statistical Computing (4.1.1)
29. Rangel-Huerta, O. D., Aguilera, C. M., Perez-de-la-Cruz, A., Vallejo, F., Tomas-Barberan, F., Gil, A., &Mesa, M. D. (2017). A serum metabolomics-driven approach predicts orange juice consumption andits impact on oxidative stress and in�ammation in subjects from the BIONAOS study. MolecularNutrition and Food Research, 61(2), https://doi.org/10.1002/mnfr.201600120
30. Ricciotti, E., & Fitzgerald, G. A. (2011). Prostaglandins and In�ammation. Arteriosclerosis,Thrombosis, and Vascular Biology. https://doi.org/10.1161/ATVBAHA.110.207449
31. Sathyapalan, T., Aye, M., Rigby, A. S., Thatcher, N. J., Dargham, S. R., Kilpatrick, E. S., & Atkin, S. L.(2018). Soy iso�avones improve cardiovascular disease risk markers in women during the earlymenopause. Nutrition Metabolism and Cardiovascular Diseases, 28(7), 691–697.https://doi.org/10.1016/j.numecd.2018.03.007
32. Spiegel, S., & Milstien, S. (2011). The outs and the ins of sphingosine-1-phosphate in immunity.Nature Reviews Immunology (Vol, 11(6), 403–415. https://doi.org/10.1038/nri2974
33. Storm-Larsen, C., Myhr, K. M., Farbu, E., Midgard, R., Nyquist, K., Broch, L., Berg-Hansen, P., Buness, A.,Holm, K., Ueland, T., Fallang, L. E., Burum-Auensen, E., Hov, J. R., & Holmøy, T. (2019). Gut microbiotacomposition during a 12-week intervention with delayed-release dimethyl fumarate in multiplesclerosis-a pilot trial. https://doi.org/10.1177/2055217319888767
34. Sun, Y., Tao, W., Huang, H., Ye, X., & Sun, P. (2019). Flavonoids, phenolic acids, carotenoids andantioxidant activity of fresh eating citrus fruits, using the coupled in vitro digestion and humanintestinal HepG2 cells model. Food Chemistry, 279, 321–327.https://doi.org/10.1016/j.foodchem.2018.12.019
35. Torres Santiago, G., Serrano Contreras, J. I., Meléndez Camargo, M. E., & Zepeda Vallejo, L. G. (2019).NMR-based metabonomic approach reveals changes in the urinary and fecal metabolome caused byresveratrol. Journal of Pharmaceutical and Biomedical Analysis, 162, 234–241.https://doi.org/10.1016/j.jpba.2018.09.025
3�. Warner, M., Huang, B., & Gustafsson, J. A. (2017). Estrogen Receptor β as a Pharmaceutical Target.Trends in Pharmacological Sciences (38 vol., pp. 92–99). Elsevier Ltd.1https://doi.org/10.1016/j.tips.2016.10.006
37. Wishart, D. S., Guo, A., Oler, E., Wang, F., Anjum, A., Peters, H., Dizon, R., Sayeeda, Z., Tian, S., Lee, B. L.,Berjanskii, M., Mah, R., Yamamoto, M., Jovel, J., Torres-Calzada, C., Hiebert-Giesbrecht, M., Lui, V. W.,Varshavi, D., Varshavi, D., & Gautam, V. (2022). HMDB 5.0: the Human Metabolome Database for2022. Nucleic Acids Research, 50. https://doi.org/10.1093/nar/gkab1062

Page 16/19
3�. Yu, H., Bai, S., Hao, Y., & Guan, Y. (2022). Fatty acids role in multiple sclerosis as “metabokines.Journal of Neuroin�ammation, 19(1), 157. https://doi.org/10.1186/s12974-022-02502-1
39. Zahoor, I., Rui, B., Khan, J., Datta, I., & Giri, S. (2021). An emerging potential of metabolomics inmultiple sclerosis: a comprehensive overview. Cellular and Molecular Life Sciences (78 vol., pp.3181–3203). Springer Science and Business. https://doi.org/10.1007/s00018-020-03733-2. MediaDeutschland GmbH
40. Zhang, A., Sun, H., Yan, G., Wang, P., & Wang, X. (2015). Review Article Metabolomics for BiomarkerDiscovery: Moving to the Clinic. https://doi.org/10.1155/2015/354671
Figures
Figure 1

Page 17/19
Principal Component Analysis (PCA) with variance scores for PC1 of 36.2% and PC2 of 15.8%. Pinkrepresents the metabolites from mice on an iso�avone group and green represents those from phyto-freegroup.
Figure 2
Hierarchical cluster analysis based on the Euclidean distance and Ward’s linkage clustering algorithm.Clustering shown for top 25 metabolites as heatmap, cell color based on normalized abundance. The leftcolumn with a green bar represents metabolite data from mice on a phyto-free diet, while the right columnwith a red bar represents data from mice on an iso�avone diet.

Page 18/19
Figure 3
Metabolites increased in mice on an iso�avone diet compared to phyto-free diet that were also found inthe heatmap clustering analysis (Fig 2). All values are normalized. Wilcoxon Rank Sum test utilized forstatistical signi�cance (* p < .05, ** p < .01, *** p < .001).
Page 19/19
Figure 4
Metabolites decreased in mice on an iso�avone diet compared to phyto-free diet that were also found inthe heatmap clustering analysis (Fig 2). All values are normalized. Wilcoxon Rank Sum test utilized forstatistical signi�cance (* p < .05, ** p < .01, *** p < .001).
Supplementary Files
This is a list of supplementary �les associated with this preprint. Click to download.
SupplementaryTable1.xlsx





























