Embed Size (px)
Citation preview

Bioelectrochemistry 87 (2012) 9–14
Contents lists available at ScienceDirect
Bioelectrochemistry
j ourna l homepage: www.e lsev ie r.com/ locate /b ioe lechem
Investigation of the mediated electron transfer mechanism of cellobiosedehydrogenase at cytochrome c-modified gold electrodes
David Sarauli a, Roland Ludwig b, Dietmar Haltrich b, Lo Gorton c, Fred Lisdat a,⁎a Biosystems Technology, Wildau Technical University of Applied Sciences, Bahnhofstr. 1, Wildau, 15745 Wildau, Germanyb Division of Food Biotechnology, Department of Food Sciences and Technology, BOKU-University of Natural Resources and Life Sciences Vienna, Muthgasse 18, A-1190 Vienna, Austriac Department of Analytical Chemistry/Biochemistry and Structural Biology, Lund University, P.O. Box 124, SE-22100 Lund, Sweden
⁎ Corresponding author. Tel.: +49 3375 508 456; faxE-mail address: [email protected] (F. Lisdat).
1567-5394/$ – see front matter © 2011 Published by Edoi:10.1016/j.bioelechem.2011.07.003
a b s t r a c t
a r t i c l e i n f oArticle history:Received 6 June 2011Received in revised form 14 July 2011Accepted 19 July 2011Available online 24 July 2011
Keywords:Cellobiose dehydrogenaseDirect electron transferMediated electron transferCytochrome cImmobilised enzyme
The present study reports on the comparison of direct and mediated electron transfer pathways in theinteraction of the fungal enzyme cellobiose dehydrogenase (CDH)with the redox protein cytochrome c (cyt c)immobilised at a modified gold electrode surface. Two types of CDHs were chosen for this investigation: abasidiomycete (white rot) CDH from Trametes villosa and a recently discovered ascomycete from thethermophilic fungus Corynascus thermophilus. The choice was based on the pH-dependent interaction of theseenzymes with cyt c in solution containing the substrate cellobiose (CB). Both enzymes show rather similarcatalytic behaviour at lower pH, dominated by a direct electron exchange with the electrode. With increasingpH, however, also cyt c-mediated electron transfer becomes possible. The pH-dependent behaviour in thepresence and in the absence of cyt c is analysed and the potential reaction mechanism for the two enzymeswith a different pH-behaviour is discussed.
: +49 3375 508 971.
lsevier B.V.
© 2011 Published by Elsevier B.V.
1. Introduction
Electron transfer (ET) phenomena in proteins represent one ofthe most common types of biochemical reactions, which play acentral role in energy conversion pathways in living cells [1–4]. Thecoupling of redox enzymes/proteins to an electrode via a hetero-geneous ET reaction is interesting in terms of providing a powerfultool for examining ET kinetics and mechanisms, understanding ofthe dynamic operation of macromolecular interfaces, and forapplication in biosensors and biofuel cells [5–10]. Especially forbiosensors it is important to elucidate the ET pathways oninterfaces, such as direct electron transfer (DET) and mediatedelectron transfer (MET) [11,12]. DET avoids the use of shuttlingmolecules and reduces potential interferences and side reactions.However, DET on different kinds of electrodes, including gold andvarious carbon materials has been obtained only for a restrictednumber of redox proteins and enzymes, and the majority of themwere shown to contain metallocentres [7,10,13–15]. Cellobiosedehydrogenase (CDH) is an interesting example of a two domainprotein for which DET could be obtained on a variety of differentelectrode materials [16].
CDH is an extracellular enzyme produced by a number wooddegrading and phytopathogenic fungi [17]. This enzyme is a monomericprotein and consists of two domains connected by a linker region. The
larger, catalytically active flavodehydrogenase domain contains FAD ascofactor, the smaller cytochrome domain harbors a b-type heme [18].Among the large number of one electron acceptors and reaction partnersfor CDH, the redoxprotein cytochrome c canbe accentuated. In contrast tolowmolecularweight oneelectronacceptors cyt c communicateswith thehemedomain of CDH [16,17]. Cyt chas been intensively investigatedwithrespect to the heterogeneous electron transfer process on modified goldelectrodes [19–22], and a cyt cmonolayer was found to efficiently oxidisesuperoxide radicals, representing a third-generation biosensor [23–25].Moreover, the reaction of cyt cwith CDHwas analysed electrochemically,demonstrating a prominent catalytic activity of the enzyme with theredox protein [26]. The capability of the protein to undergo quasi-reversible redox conversion at an electrode and to exchange electronswith specific enzymes makes cyt c an interesting building block indesigning biomimetic signal chains. This has been shown not only withthe protein in solution [27,28], but also with the adsorbed protein [26,29]and more recently even with cyt c immobilised in multiple layers [8,30].
DET between CDH and thiol-modified Au has also beeninvestigated, involving CDHs from the white-rot basidiomycetefungi Phanerochaete chrysosporium[31,32], Phanerochaete sordidaand Trametes villosa[15], the plant pathogenic basidiomyceteSclerotium rolfsii[31], the soft-rot ascomycetes Humicola insolens[31] and Myriococcum thermophilum[15,33]. The first four enzymesbelong to the basidiomycete class I (white rot) CDHs, the latter twoto the ascomycete class II CDHs [17]. In particular, Stoica et al. [15]demonstrated the ability of CDHs from different sources to directlyexchange electrons with different thiol-modified Au-electrodes,leading to the occurrence of DET in the presence of cellobiose.

10 D. Sarauli et al. / Bioelectrochemistry 87 (2012) 9–14
The goal of this study is the electrochemical investigation of theelectron transfer between the enzyme CDH and the redox proteincyt c immobilised on a thiol-modified Au surface, so that a METbetween the enzyme and the electrode surface via the immobilisedcyt c might be achieved. This situation is compared with the DET ofCDH at different pHs. CDH from the white rot fungus T. villosa, classI CDH [34], as well as a novel one from the thermophilic fungusCorynascus thermophilus, class II CDH [35], have been chosen forthis study.
2. Materials and methods
2.1. Materials
2.1.1. ChemicalsHorse-heart cytochrome c (cyt c), D-(+)-cellobiose (98%), 11-
mercapto-1-undecanoic acid (MUA), and 11-mercapto-1-undecanol(MU) are purchased from Sigma-Aldrich; potassium hydrogenphosphate and potassium dihydrogen phosphate are taken fromMerck. All reagents are used without further purification. 18 MΩMillipore water (Eschborn, Germany) is used for all types ofmeasurements.
2.1.2. EnzymesSolutions of CDHs from the following two organisms are used:
T. villosa (5 mg/ml; 88 U/ml) and C. thermophilus (2.6 mg/ml;17.2 U/ml). T. villosa CDH has been recently characterised [34] andhas a molecular mass of 98 kDa (SDS-PAGE) and a pI of 4.4. C.thermophilus CDH has a molecular mass of 87 kDa and pI of 4.1[35].
2.2. Apparatus
2.2.1. ElectrochemistryAll electrochemical measurements are done in a 1 ml cell using
an Ag|AgCl|1 M KCl reference (Biometra, Germany) and a Pt-wirecounter electrode. The working electrodes are modified gold wires(diameter 0.5 mm) obtained from Goodfellow (Bad Nauheim,Germany). Cyclic voltammetric experiments are carried out withthe Autolab PGSTAT 30 device (Metrohm, Germany). The potentialrange has been chosen to be between −0.4 and +0.4 V. Themeasurements are performed in 5 mMphosphate buffer of differentpHs with a scan rate of 5 mV/s. In the course of experiments first theelectrodes are studied in the respective buffer solution, then theenzyme is added and the electrodes are scanned again. In order toevaluate a potential catalytic behaviour cellobiose is added from aconcentrated stock solution and the electrodes are again voltam-metrically studied. For the establishment of a uniform concentra-tion in the measuring solution magnetic stirring is used before andbetween the voltammetric scans. Besides modified gold electrodeswith adsorbed cyt c (Au-MUA/MU-cyt c) also modified electrodeswithout cyt c (Au-MUA/MU) have been tested.
2.2.2. Kinetic measurementsUV–Vis measurements are performed with Helios α, Thermo
Spectronic spectrophotometer. Kinetic data are determined withconstant concentrations of both cellobiose (5 mM) and cyt c (10 and5 μM) at pHs 5 and 7 and 25 °C in 5 mM potassium phosphate buffer.The concentrations of CDH are varied between 0.02 and 0.2 μM. Theextinction coefficient (ε=19.6 mM−1 cm−1) is taken for the calcu-lations [36].
2.3. Preparation of cytochrome c monolayers
Prior to modification gold wire electrodes are intensivelycleaned: firstly they are boiled in KOH for 4 h, rinsed with water,
and stored in concentrated H2SO4. After that the electrodes aretreated for 15 min with concentrated nitric acid, and rinsed againcarefully with water. Modification of the electrodes with alkanethiols is carried out by 48 h incubation in a 5 mM MUA/MU (1:3)(mercaptoundecanoic acid/mercaptoundecanol) ethanol solutionand subsequent rinsing with ethanol and phosphate buffer (pH 7).A cyt c monolayer is prepared on top of the MUA/MU layer by 2 hadsorption from a 20 μMcyt c solution in 5 mMpotassium phosphatebuffer solution (pH 7) [23]. These monolayer electrodes are rinsedwith 5 mM potassium phosphate buffer prior to experiments in theenzyme solution.
3. Results
In this study CDH from two different sources, T. villosa and C.thermophilus have been investigated with respect to a cyt c-mediatedelectron-transfer and a direct electron exchange at the electrode. Forthis purpose MUA/MU-modified gold electrodes are used, since thiscarboxylic acid/hydroxy-terminated surface layer provides a suitableinterface for cyt c and CDH conversion [15,23]. Furthermore, thisinterface allows adsorption of cyt c from solutions of low ionicstrength in the range of a monolayer. Different pH values of theelectrolyte-containing enzyme solutions have been evaluated for theinteraction with such a kind of electrode.
3.1. Cellobiose dehydrogenase from T. villosa
Fig. 1 shows cyclic voltammograms of T. villosa CDH in solution onan electrode with adsorbed cyt c monolayer (Au-MUA/MU-cyt c) inthe pH range between pH 4.5 and 7 in the absence and in the presenceof the substrate cellobiose. In the absence of cellobiose a stable andquasi-reversible electrochemical response of cyt c immobilised on theMUA/MU-modified gold was observed and is in agreement withalready published results [23]. A shift of the redox potential of theFeIII/II heme redox couple in cyt cwith different pH values can be seen,confirming the involvement of proton-exchange in the electrontransfer reaction. The averaged value of the surface coverage of cyt cdetermined by measuring 6 different electrodes at pH 7 is aboutΓ=10±2 pmol/cm2.
Upon addition of cellobiose a prominent catalytic current appears.However, the catalytic behaviour is strongly pH-dependent. At pHvalues 4.5 and 5 the catalytic wave starts at about−0.15 V vs. Ag|AgCl,indicating that the electron transfer fromCDH to the electrode surface isnot mediated by cyt c, since the catalytic behaviour occurs already atpotentials lower than oxidation of cyt c takes place and rather in thepotential range of the heme domain of the CDH.
At pH 6 the catalytic current starts at higher potentials (E=0mV vs.Ag|AgCl), near the potential of cyt c oxidation. The shape of this catalyticcurrent differs significantly from thatmeasured at lowerpH. Even thoughthe magnitude of this current is only in the range of the Faradaic cyt ccurrent, a clear evidence for the change in reaction mechanism throughthe presence of adsorbed cyt c can be elucidated. The adsorbed proteinactshere as amediator. Surprisingly, evenatpH7a small catalytic currentoccurs upon addition of the same amount of cellobiose, which seems tobemediated by cyt c. The catalytic process is unambiguously less efficientthan at pH 6.
In order to evaluate the contribution of DET in CDH electrontransfer, the same set of experiments has been performed, however,without adsorbed cyt c. The results for the mixed thiol-modified goldelectrodes measured in solutions of T. villosa CDH are summarised inFig. 2 in the presence and in the absence of cellobiose in the same pHrange.
It can be demonstrated that the catalytic currents at lower pHvalues (pHs 4.5 and 5) are pronounced, with higher current intensitiesin more acid solution. Almost no difference in the shapes of the CVscan be seen in comparison with the results obtained when also
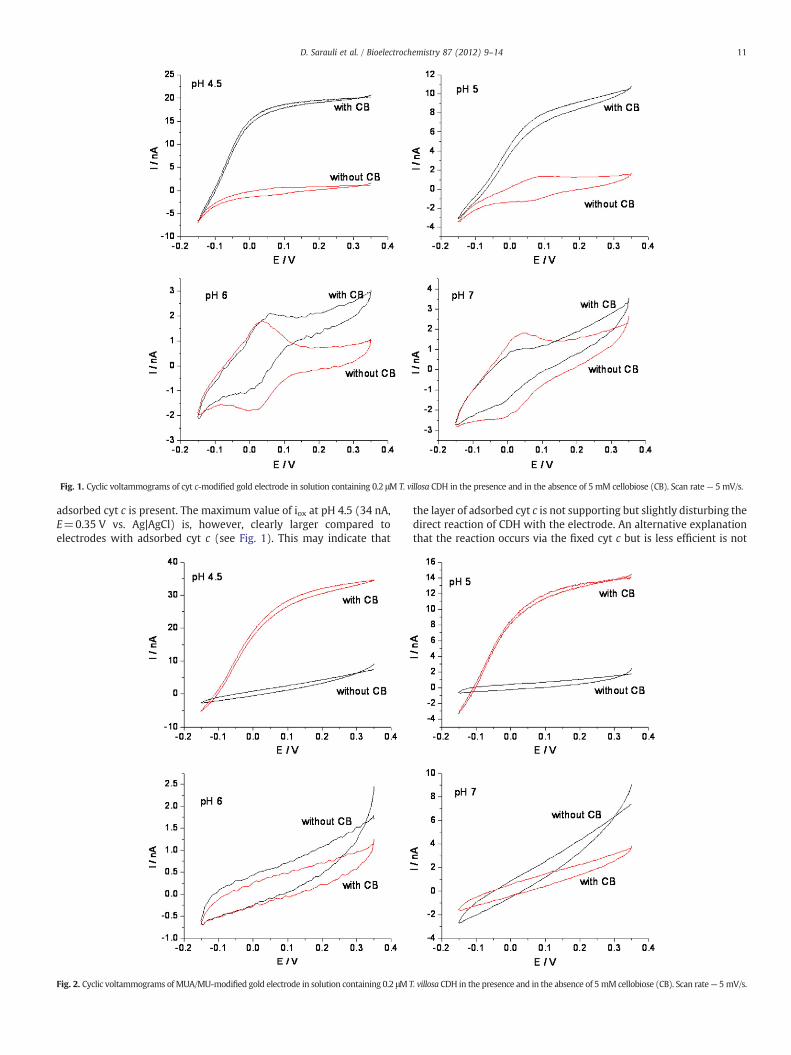
Fig. 1. Cyclic voltammograms of cyt c-modified gold electrode in solution containing 0.2 μM T. villosa CDH in the presence and in the absence of 5 mM cellobiose (CB). Scan rate— 5 mV/s.
11D. Sarauli et al. / Bioelectrochemistry 87 (2012) 9–14
adsorbed cyt c is present. The maximum value of iox at pH 4.5 (34 nA,E=0.35 V vs. Ag|AgCl) is, however, clearly larger compared toelectrodes with adsorbed cyt c (see Fig. 1). This may indicate that
Fig. 2. Cyclic voltammograms ofMUA/MU-modified gold electrode in solution containing 0.2 μM T
the layer of adsorbed cyt c is not supporting but slightly disturbing thedirect reaction of CDH with the electrode. An alternative explanationthat the reaction occurs via the fixed cyt c but is less efficient is not
. villosa CDH in the presence and in the absence of 5 mM cellobiose (CB). Scan rate— 5 mV/s.

12 D. Sarauli et al. / Bioelectrochemistry 87 (2012) 9–14
very likely since the catalytic reaction starts at more negativepotentials than the redox potential of cyt c.
At pHs 6 and 7 no catalytic current appears when only CDH ispresent. Obviously no direct enzyme conversion can take place,and thus electrons cannot be shuttled to the electrode. Thisrepresents a clear contrast to the case when cyt c is present and aresponse signal from the enzyme to the electrode via cyt c is foundupon addition of the substrate.
The electrochemical results are in good agreement with thereaction between cyt c and the substrate-reduced enzyme insolution evaluated photometrically. Results of measurements atpHs 5 and 7 are summarised in Table 1. No activity of this CDH atpH 7 can be detected, whereas the rate constant between CDH andcyt c (k) at pH 5 is 0.28×105 M−1 s−1.
3.2. Cellobiose dehydrogenase from C. thermophilus
Fig. 3 summarises the CVs measured with Au-MUA/MU-cyt celectrodes in solutions of C. thermophilus CDH in the presence and inthe absence of cellobiose. The same pH range as for T. villosa CDH waschosen for the measurements.
At pH 4.5 a pronounced catalytic current starts at a rather lowpotential at about −0.1 V vs. Ag|AgCl, but with less intensitycompared to T. villosa CDH. Again cyt c oxidation seems to be noprecondition for the catalytic oxidation current. The same is validfor pH 5 and also in this case the current intensity is smallercompared to T. villosa CDH. With increasing pH to 6 still a clearcatalytic current is visible, but it starts at higher potentials. At pH 7the intensity of this current is very weak.
In order to characterise the role of DET for C. thermophilus CDH,measurements in the absence of cyt c have been performed. Theresults are given in Fig. 4. At pHs 4.5 and 5 the behaviour is rathersimilar to the reaction of T. villosa CDH at MUA/MUmodified gold. Thecurrent densities are, however, clearly smaller in comparison to T.villosa CDH. By comparing the catalytic behaviour of C. thermophilusCDH at pH 4.5 in the presence and absence of cyt c it is evident thathere the presence of the redox protein has no positive effect on thecatalytic current and thus the current densities with and without cyt care rather similar.
At pHs 6 and 7 no reaction is found for C. thermophilus CDH atthe MUA/MU-modified electrode — analogous to the T. villosasystem. Thus, the catalytic activity found at pH 6 with the enzymeand adsorbed cyt c seems to be mediated by the redox protein.
Similar to T. villosa CDH, the reaction of C. thermophilus CDHwith cyt cin solution has been studied photometrically to evaluate the overall rateconstant, k, for substrate oxidation using cyt c as electron acceptor. Theresults for pH 5 and pH 7 are summarised in Table 1. This enzyme showssignificantly higher activity at both pHs 5 and 7, compared to T. villosaCDH. The values of k were determined to be 1.9×105 M−1 s−1 and1.3×105 M−1 s−1 at pH 5 and pH 7, respectively. Contrary to T. villosaCDH, a catalytic reaction at neutral pH is feasible.
4. Discussion
At pHs 4.5 and 5 the DET for both CDHs is dominating at themodified gold electrodes regardless of the presence of cyt c. The
Table 1Summary of second-order reaction rate constants for both CDHs, measured photomet-rically. Experimental conditions: [cyt c]=10 μM, [cellobiose]=5 mM, t=25 °C.
pH Stoichiometrya k, 105 M−1 s−1
T. villosa C. thermophilus
5 2 0.28 1.97 2 1.3
a Molecules of cyt c reduced per molecule of reduced CDH.
reaction cycle can be explained by the enzyme mechanism: thecatalytic oxidation of cellobiose takes place at the FAD, when electronsand protons are transferred from cellobiose. Next, the reduced FADcan directly transfer electrons one by one to the heme cofactor via aninternal electron transfer mechanism between the flavodehydrogen-ase and the heme b of the cytochrome domain. The reducedcytochrome domain can then be oxidised directly at the electrode. Itis also possible that the cytochrome subunit releases electrons to 1 e−
acceptors such as cyt c[31,32], however, this seems not to be veryefficient in the acidic pH range, when the redox protein isimmobilised. In addition, it should be pointed out that even thoughthe activity of C. thermophilus CDH in solution at pH 5 is determined tobe much higher, than that of T. villosa CDH, a higher catalytic currentfor the latter CDH at the modified gold surface appears. This isindicative of a much better interaction of T. villosa CDH with themodified electrode surface.
At pHs 6 and 7, however, no DET for T. villosa CDH at a MUA/MUmodified electrode can be detected. In previous studies on CDHelectrochemistry it has been shown that this behaviour can beexplained by a significantly decreased internal electron transfer forthis CDH at pHs higher than 5 [15,36,37]. Whereas this process isefficient at pH 4.5, at pHs 6 and 7 it is shown [17] that the interactionof the two domains decreases due to electrostatic repulsion ofdeprotonated amino acid residues on the surface of the two domains.The DET at these pH values is almost completely switched off bylack of internal electron transfer. Our photometrical evaluation isalso in agreement with this statement, since no activity of T. villosaCDH is detected in solution at pH 7, in comparison with the kineticsat pH 5.
When cyt c is present on the electrode surface a moderate, butclearly visible catalytic current occurs in the presence of T. villosa CDH.This means that at pH 6 (and to a much smaller extent at pH 7) thesignal chain from cellobiose via the enzyme to the electrode ismediated by cyt c. However, the question remains how electrons aretransferred to the electrode when the internal pathway between bothsubunits is diminished. This question cannot be fully answered at thepresent stage. The structure of this CDH is a more open one at neutralpH because of the electrostatic repulsion of surface groups on the twodomains resulting in a larger distance between the FAD and the hemedomains [34]. This may allow an electron transfer from the FADdomain directly to cyt c immobilised on the electrode surface oralternatively that the immobilised cyt c supports the interaction ofboth subunits of the enzyme. At least one can state that somesubstances such as low molecular weight mediators or Os-complexescan also interact directly with the FAD domain [38]. However cyt c israther different from these molecules and a direct interaction is notvery likely.
Another interesting result is the catalytic behaviour of C. thermo-philus CDH at pHs 6 and 7. No DET for this enzyme at a MUA/MUmodified electrode can be detected, although C. thermophilus CDHshows high activity in solution also at pH 7. Obviously here theinteraction with the electrode is weak, resulting in a non productiveelectron transfer at neutral pH. In contrast when adsorbed on graphiteit shows rather efficient DET at this pH [39,40].
In conclusion it has to be stated that both CDHs show a rathersimilar behaviour at the MUA/MU modified electrode, although adifferent mechanism is behind. In the presence of immobilised cyt cC. thermophilus CDH shows a similar catalytic behaviour as T. villosaCDH, since a moderate, but clearly visible catalytic current occurs inthe presence of the substrate. It means that the redox protein canestablish the communication towards the electrode, although CDHalone cannot. Surprisingly, at pH 7 no catalytic current occursalthough the enzyme can react with the protein in solution, andcyt c electrochemistry is efficient at this pH. This cannot be fullyunderstood, but it seems to be caused by the immobilised state ofthe redox protein.

Fig. 3. Cyclic voltammograms of cyt c-modified gold electrode in solution containing 0.2 μM C. thermophilus CDH in the presence and in the absence of 5 mM cellobiose(CB). Scan rate — 5 mV/s.
13D. Sarauli et al. / Bioelectrochemistry 87 (2012) 9–14
In order to explain the shuttling ability of immobilised cyt c forboth investigated CDHs found at pH 6, the rotational flexibility inthe adsorbed state has to be considered. Although fixed on the
Fig. 4. Cyclic voltammograms of MUA/MU-modified gold electrode in solution containing 0Scan rate — 5 mV/s.
electrode surface, the protein shows rotational movement, allowingelectron shuttling between a reaction partner and the electrode[29,30,41].
.2 μM C. thermophilus CDH in the presence and in the absence of 5 mM cellobiose (CB).

14 D. Sarauli et al. / Bioelectrochemistry 87 (2012) 9–14
5. Conclusion
The electrochemical behaviour of CDH from two different sources,T. villosa and C. thermophilus, has been investigated with respect to amediated electron transfer by an immobilised layer of cyt c and adirect electron transfer communication at a thiol-modified electrode.The behaviour of the two different enzymes shows a strongdependence on solution pH. At lower pH (≤5) a pronounced catalyticcurrent is found with both enzymes, whereas T. villosa CDH showshigher catalytic currents in the presence of cellobiose. Despite thepresence of a protein layer on the electrode, DET of CDH to theelectrode is here the dominating electron transfer pathway for bothenzymes. At pH 6, however, no DET occurs, but a mediation bysurface-bound cyt c can be verified for the CDHs. For T. villosa CDH thiscan be explained by a bridging of the lack of internal electron transferby immobilised cyt c, whereas for the C. thermophilus CDH cyt cmediation is necessary, since the protein cannot productively interactwith the modified gold electrode. For both CDH types, a signal chainstarting from cellobiose in solution via the enzyme and immobilisedredox protein can be obtained at well defined pH values.
Acknowledgements
The authors thank the following agencies for financial support: theSwedish Research Council (L. G.), MWFK Brandenburg (project: 3508-14/13) and the BMBF (project: 03IS2201I), Germany.
References
[1] P.S. Brookes, A.L. Levonen, S. Shiva, P. Sarti, V.M. Darley-Usmar, Mitochondria:regulators of signal transduction by reactive oxygen and nitrogen species, FreeRadic. Biol. Med. 33 (2002) 755–764.
[2] G.W. Brudvig, R.H. Crabtree, Mechanism for photosynthetic O2− evolution, Proc.
Natl Acad. Sci. USA 83 (1986) 4586–4588.[3] A.N. Glazer, A. Melis, Photochemical reaction centers — structure, organization,
and function, Annu. Rev. Plant Physiol. Plant Mol. Biol. 38 (1987) 11–45.[4] J.F. Turrens, Mitochondrial formation of reactive oxygen species, J. Physiol.London
552 (2003) 335–344.[5] V. Coman, Heterogeneous electron transfer studies with ligninolytic redox
enzymes and living bacteria. Applications in biosensors and biofuel cells,Department of Analytical Chemistry, Lund University, Lund, 2009, p. 76.
[6] W.B. Frommer, M.W. Davidson, R.E. Campbell, Genetically encoded biosensorsbased on engineered fluorescent proteins, Chem. Soc. Rev. 38 (2009) 2833–2841.
[7] L. Gorton, A. Lindgren, T. Larsson, F.D. Munteanu, T. Ruzgas, I. Gazaryan, Directelectron transfer between heme-containing enzymes and electrodes as basis forthird generation biosensors, Anal. Chim. Acta 400 (1999) 91–108.
[8] F. Lisdat, R. Dronov, H. Mohwald, F.W. Scheller, D.G. Kurth, Self-assembly ofelectro-active protein architectures on electrodes for the construction ofbiomimetic signal chains, Chem. Commun. (2009) 274–283.
[9] J.M. Pingarron, P. Yanez-Sedeno, A. Gonzalez-Cortes, Gold nanoparticle-basedelectrochemical biosensors, Electrochim. Acta 53 (2008) 5848–5866.
[10] W.J. Zhang, G.X. Li, Third-generation biosensors based on the direct electrontransfer of proteins, Anal. Sci. 20 (2004) 603–609.
[11] L.J.C. Jeuken, Conformational reorganisation in interfacial protein electrontransfer, Biochim. Biophys. Acta, Bioenerg. 1604 (2003) 67–76.
[12] C.C. Page, C.C. Moser, X.X. Chen, P.L. Dutton, Natural engineering principles ofelectron tunnelling in biological oxidation–reduction, Nature 402 (1999) 47–52.
[13] T. Ikeda, Direct redox communication between enzymes and electrodes, Frontiersin Biosensorics, Birkhäuser Verlag, Basel, 1997, pp. 243–266.
[14] W. Schuhmann, Amperometric enzyme biosensors based on optimised electro-n-transfer pathways and non-manual immobilisation procedures, J. Biotechnol.82 (2002) 425–441.
[15] L. Stoica, N. Dimcheva, D. Haltrich, T. Ruzgas, L. Gorton, Electrochemicalinvestigation of cellobiose dehydrogenase from new fungal sources on Auelectrodes, Biosens. Bioelectron. 20 (2005) 2010–2018.
[16] R. Ludwig,W. Harreither, F. Tasca, L. Gorton, Cellobiose dehydrogenase: a versatilecatalyst for electrochemical applications, ChemPhysChem 11 (2010) 2674–2697.
[17] M. Zamocky, R. Ludwig, C. Peterbauer, B.M. Hallberg, C. Divne, P. Nicholls, D.Haltrich, Cellobiose dehydrogenase — a flavocytochrome from wood-degrading,phytopathogenic and saprotropic fungi, Curr. Protein Pept. Sci. 7 (2006) 255–280.
[18] B.M. Hallberg, T. Bergfors, K. Backbro, G. Pettersson, G. Henriksson, C. Divne, A newscaffold for binding haem in the cytochrome domain of the extracellularflavocytochrome cellobiose dehydrogenase, Struct. Fold. Des. 8 (2000) 79–88.
[19] S. Arnold, Z.Q. Feng, T. Kakiuchi, W. Knoll, K. Niki, Investigation of the electrodereaction of cytochrome c through mixed self-assembled monolayers of alka-nethiols on gold(111) surfaces, J. Electroanal. Chem. 438 (1997) 91–97.
[20] J.E. Frew, H.A.O. Hill, Direct and indirect electron-transfer between electrodes andredox proteins, Eur. J. Biochem. 172 (1988) 261–269.
[21] I. Taniguchi, K. Toyosawa, H. Yamaguchi, K. Yasukouchi, Voltammetric response ofhorse heart cytochrome c at a gold electrode in the presence of sulfur bridgedbipyridines, J. Electroanal. Chem. 140 (1982) 187–193.
[22] B. Willner, E. Katz, I. Willner, Electrical contacting of redox proteins bynanotechnological means, Curr. Opin. Biotechnol. 17 (2006) 589–596.
[23] B. Ge, F. Lisdat, Superoxide sensor based on cytochrome c immobilized onmixed-thiol SAMwith anewcalibrationmethod,Anal. Chim.Acta454 (2002)53–64.
[24] K.V. Gobi, F. Mizutani, Efficient mediatorless superoxide sensors using cyto-chrome c-modified electrodes: surface nano-organization for selectivity andcontrolled peroxidase activity, J. Electroanal. Chem. 484 (2000) 172–181.
[25] F. Lisdat, B. Ge, E. Ehrentreich-Forster, R. Reszka, F.W. Scheller, Superoxidedismutase activity measurement using cytochrome c modified electrode, Anal.Chem. 71 (1999) 1359–1365.
[26] V. Fridman, U. Wollenberger, V. Bogdanovskaya, F. Lisdat, T. Ruzgas, A. Lindgren, L.Gorton, F.W. Scheller, Electrochemical investigation of cellobiose oxidation bycellobiose dehydrogenase in the presence of cytochrome c as mediator, Biochem.Soc. Trans. 28 (2000) 63–70.
[27] R. Dronov, D.G. Kurth, F.W. Scheller, F. Lisdat, Direct and cytochrome c mediatedelectrochemistry of bilirubin oxidase on gold, Electroanalysis 19 (2007)1642–1646.
[28] W. Jin, U. Wollenberger, F.F. Bier, A. Makower, F.W. Scheller, Electron transferbetween cytochrome c and copper enzymes, Bioelectrochem. Bioenerg. 39 (1996)221–225.
[29] W. Jin, U. Wollenberger, E. Kargel, W.H. Schunck, F.W. Scheller, Electrochemicalinvestigations of the intermolecular electron transfer between cytochrome c andNADPH cytochrome P450 reductase, J. Electroanal. Chem. 433 (1997) 135–139.
[30] R. Dronov, D.G. Kurth, H. Moehwald, F.W. Scheller, F. Lisdat, Communication in aprotein stack: electron transfer between cytochrome c and bilirubin oxidasewithin a polyelectrolyte multilayer, Angew. Chem. Int. Ed. 47 (2008) 3000–3003.
[31] A. Lindgren, L. Gorton, T. Ruzgas, U. Baminger, D. Haltrich, M. Schulein, Directelectron transfer of cellobiose dehydrogenase from various biological origins atgold and graphite electrodes, J. Electroanal. Chem. 496 (2001) 76–81.
[32] A. Lindgren, T. Larsson, T. Ruzgas, L. Gorton, Direct electron transfer between theheme of cellobiose dehydrogenase and thiol modified gold electrodes, J.Electroanal. Chem. 494 (2000) 105–113.
[33] V. Coman, W. Harreither, R. Ludwig, D. Haltrich, L. Gorton, Investigation ofelectron transfer between cellobiose dehydrogenase from Myriococcum thermo-philum and gold electrodes, Chem. Anal. 52 (2007) 945–960.
[34] R. Ludwig, A. Salamon, J. Varga, M. Zamocky, C.K. Peterbauer, K.D. Kulbe, D.Haltrich, Characterisation of cellobiose dehydrogenases from the white-rot fungiTrametes pubescens and Trametes villosa, Appl. Microbiol. Biotechnol. 64 (2004)213–222.
[35] W. Harreither, C. Sygmund, M. Augustin, M. Narciso, M.L. Rabinovich, L. Gorton, D.Haltrich, R. Ludwig, Catalytic properties and classification of cellobiose de-hydrogenases from ascomycetes, Appl. Environ. Microbiol. 77 (2011) 1804–1815.
[36] L. Stoica, R. Ludwig, D. Haltrich, L. Gorton, Third-generation biosensor for lactosebased on newly discovered cellobiose dehydrogenase, Anal. Chem. 78 (2006)393–398.
[37] L. Stoica, T. Ruzgas, R. Ludwig, D. Haltrich, L. Gorton, Direct electron transfer — afavorite electron route for cellobiose dehydrogenase (CDH) from Trametes villosa.Comparison with CDH from Phanerochaete chrysosporium, Langmuir 22 (2006)10801–10806.
[38] F. Tasca, L. Gorton, W. Harreither, D. Haltrich, R. Ludwig, G. Noll, Comparison ofdirect and mediated electron transfer for cellobiose dehydrogenase fromPhanerochaete soridida, Anal. Chem. 81 (2009) 2791–2798.
[39] V. Coman, R. Ludwig, W. Harreither, D. Haltrich, L. Gorton, T. Ruzgas, S. Shleev, Adirect electron transfer-based glucose/oxygen biofuel cell operating in humanserum, Fuel Cells 10 (2010) 9–16.
[40] F. Tasca, M.N. Zafar, W. Harreither, G. Noll, R. Ludwig, L. Gorton, A third generationglucose biosensor based on cellobiose dehydrogenase from Corynascus thermo-philus and single-walled carbon nanotubes, Analyst 136 (2010) 2033–2036.
[41] J.J. Wei, H.Y. Liu, D.E. Khoshtariya, H. Yamamoto, A. Dick, D.H. Waldeck,Electron-transfer dynamics of cytochrome c: a change in the reaction mechanismwith distance, Angew. Chem. Int. Ed. 41 (2002) 4700–4703.





























