Embed Size (px)
Citation preview

Intracellular Mediators of Insulin Action
W . Kirby Gottschalk and Leonard Jarett
University of Pennsylvania School of Medicine. Department of Pathology and Laboratory Medicine
I . Introduction ................................................................. I1 . The First Direct Evidence for Mediators of Insulin Action ................... 111 . Tissue Sources of Insulin Mediators .........................................
A . Plasma Membrane Sources .............................................. 8 . Whole Cells and Intact Tissue ...........................................
2 . Muscle ................................................................ 3 . Liver ..................................................................
IV . Enzymes Modulated by Insulin Mediators ................................... A . Glycogen Synthase ...................................................... B . Pyruvate Dehydrogenase ................................................ C . Glucose-6-Phosphatase .................................................. D . Phospholipid Methyitransferase ......................................... E . Acetyl-CoA Carboxylase ................................................. F . (Ca" + Mg2')-ATPase . ................................................ G . CAMP-Dependent Protein Kinase ........................................ H . Enzymes of cAMP Metabolism ..........................................
2 . Low-K, cAMP Phosphodiesterase .................................... V . Evidence for a Family of Mediators of Insulin Action ........................
VI . Physicochemical Properties of Insulin Mediators ............................ VII . Physiologic and Pathophysiologic Roles of Insulin Mediators ...............
VIII . Conclusion ..................................................................
1 . Adipocytes ...........................................................
1 . Adenylate Cyclase ....................................................
229 231 233 233 236 236 236 237 240 240 241 242 242 243 243 244 244 244 245 245 248 251 252
I . INTRODUCTION
Insulin. the major anabolic hormone of the body. has pleiotropic effects on cellular metabolism . Within seconds of interacting with its receptor. insulin promotes changes at the plasma membrane that result in increased transport of carbohydrates. amino acids. fatty acids. and ions; within minutes. it redirects metabolism by activating anabolic pathways and inhibiting catabolic events; and. over a period of hours. it exerts control over nuclear events resulting in
Diabetes/Metabolism Reviews. Vol . 1. No . 3. 229-259 (1985) 0 1985 by John Wiley & Sons. Inc . CCC 0742-4221/85/03229-31$04.00

230 INSULIN MEDIATORS
increased cell growth. Despite much intense effort spanning nearly the 60 years since insulin’s discovery, a detailed picture of the mechanisms of insulin action has yet.to emerge.
One early theory of insulin action proposed that insulin affected metabolism by interacting directly with intracellular enzymes.’ By the late 1940s, however, attention was directed at events occurring at the plasma membrane by the dis- coveries that (1) radioactively labeled insulin became attached to the surface of sensitive cells2z3 and (2) insulin promoted an increased rate of hexose transport into many sensitive cells.44 As a result of these discoveries, it was proposed that most of the actions of insulin on metabolism were consequences of the hormone’s effects on membrane transport systems. However, this unitarian model of insulin action became untenable after the discovery by Villar-Palasi and Larner that the effects of insulin on glycogen synthase could be dissociated from the hormone’s effects on glucose ~ p t a k e . ~
Conclusive proof that the binding of insulin to its receptor initiates the cellular response to insulin was adduced by Freychet et a1.,8,9 who developed techniques of labeling insulin to high specific activity and methodology to study the specific binding of ligands to their receptors. A variety of signals, including membrane hyperpolarization, cyclic nucleotides, ions, and hydrogen peroxide, have been proposed to mediate the effects of insulin. None of these, however, has been found to mimic insulin’s pleiotropic effects on both intact cells and in broken cell preparations, and, thus, none are likely to represent physiologicalIy relevant insulin second messengers.
Recently, attention has focused on a family of low-molecular-weight sub- stances whose synthesis and/or release is enhanced by the interaction of insulin with its receptors on the surfaces of sensitive cells. Larner first discussed the theoretical implications of intracellular mediators of insulin action, lo and, with his co-workers, was the first to identify material with the biochemical properties of an insulin mediator in muscle extracts. At about the same time, but inde- pendently of Larner, Turakulov et al. l1 partially purified a low-molecular-weight material from livers of insulin-treated rats that regulated mitochondria1 Ca’+ transport. Also working independently of Larner, Jarett’s laboratory was the first to directly demonstrate the production by insulin of a mediator from the plasma membrane: they showed, in a subcellular system, that the interaction of insulin with its receptor on isolated plasma membranes brought about the generation and/or release of a low-molecular-weight material that could activate pyruvate dehydrogenase in isolated mitochondria. l2
The remainder of this review will focus on six topics. First, the initial ex- periments in which the mediators of insulin action were first detected will be discussed. Next the variety of tissue sources from which mediators have been obtained will be reviewed, followed by a description of the insulin-sensitive enzymes whose activities are modulated by the mediators. Fourth, evidence will be presented which clearly demonstrates that the second messengers of insulin action are a family of closely related molecules, not just a single entity. Fifth, the physicochemical properties of the mediators, as they are presently known, are summarized. Finally, the physiologic and pathologic importance of these mediators is discussed.

GOTTSCHALK AND JARETT 231
11. THE FIRST DIRECT EVIDENCE FOR MEDIATORS OF INSULIN ACTION
Larner’s proposal for an intracellular mediator of insulin action was based on several findings. First, Villar-Palasi and Larner showed that glycogen syn- thase could be activated by insulin in the absence of glucose in the media, that is, without concurrently stimulating the transport system and without, there- fore, increasing intracellular glucose levels, which could ”push” the glycogen synthase r e a ~ t i o n . ~ , ~ ~ , ’ ~ These independent effects of insulin on two insulin- sensitive events could be explained by an insulin-dependent mediator.” In ad- dition, Larner took note of the facts that (1) insulin inactivated CAMP-dependent protein kinase and (2) basal cAMP levels were not detectably affected by insulin, while it blocked the increase of cAMP elicited by glycogenolytic hormones. Larner suggested that these observations could be explained by the insulin- promoted generation of an intracellular messenger that was distinct from cAMP but interconvertible with it. lo As initially envisioned, this intermediate would block the activation of CAMP-dependent protein kinase by preventing increases in cAMP levels promoted by glycogenolytic hormones, as well as by acting as a sink for cAMP formed under basal conditions. In fact, it was later found that insulin inactivated the CAMP-dependent protein kinase both by promoting the formation of the holoenzyme (inactive) form and by desensitizing the holoen- zyme to stimulation by CAMP.'^,'^
Larner’s laboratory provided the first direct experimental evidence for the putative intracellular messenger of insulin action. Larner et al. extracted from skeletal muscle of insulin-treated rabbits a low-molecular-weight, heat- and acid- stable material that inhibited CAMP-dependent protein kinase.17 This material was shown to be distinct from a cAMP antagonist18 by chromatography on Dowex-1, and, although it co-eluted from this column with cGMP, the protein kinase inhibitory material was much less stable than the cyclic nucleotide at pH 7, and was therefore distinct from it. In addition, unlike the protein kinase inhibitor that had previously been isolated in Kreb’s laboratory,” the inhibitor identified by Larner et al. was soluble in TCA and its amounts or activity were increased by insulin treatment of the rabbits before extraction. When the muscle extracts were subjected to paper chromatography to remove nucleotides and then sized on gel filtration columns, material in the same column fraction that possessed the protein kinase inhibitory material also activated glycogen syn- thase. 2o
Evidence was first provided by Jarett’s laboratory that insulin mediators are generated and/or released by the plasma membrane subsequent to the inter- action of insulin with its receptor. Jarett and Smith developed the first plasma membrane system that responded directly to insulin in vitro.21 They showed that insulin enhanced the magnesium-dependent hydrolysis of ATP by isolated adipocyte plasma membranes. Since protein phosphorylation, especially of in- sulin-sensitive enzymes, plays an important role in intracellular regulation,22 the effects of insulin on the phosphorylation of adipocyte plasma membranes were examined. The addition of insulin to assay mixtures containing plasma membranes and [y-32P]ATP inhibited phosphorylation of membranes, in a con-

232 INSULIN MEDIATORS
centration-dependent manner, compared with control mixtures.23 The effect was specific for insulin and was detected as early as 30 s after the addition of insulin.
These results were examined further by using SDS-PAGE to analyze the phosphoproteins of the labeled membrane^.^^,^^ Five phosphorylated bands, three minor bands, and two major bands of 120,000 and 42,000 daltons, re- spectively, were identified. Insulin promoted the decreased phosphorylation of the two major bands, and this decrease entirely accounted for the decreases observed with the intact membranes. The identity of the 120,000-dalton band has not been established to date, but the 42,000-dalton band was shown to be the a-subunit of pyruvate dehydrogenase (PDH), which was contained in mi- tochondria that contaminated the plasma membrane preparations to the extent of 3-10%. Insulin had no effect on the phosphorylation of the 42,000-dalton band when added to mitochondria alone, but the hormone’s effect was restored when plasma membranes were added back to the system. These findings sug- gested that the effects of insulin on the 42,000-dalton band were secondary to its interaction with the insulin receptor on the plasma membrane, resulting in the generation and/or release of a mediator that affected the a-subunit in mi- tochondria. In a recent report, Akers and Routtenberg26 showed that the ad- dition of insulin to homogenates of rat hippocampal tissue promoted the dephosphorylation of the a-subunit of pyruvate dehydrogenase.
The effect of insulin in these subcellular systems is similar to its effects in vivo. Insulin treatment of rat adipocytes decreased phosphorylation of the a- subunit of the PDH, and this dephosphorylation was correlated with increased PDH It had been shown previously that PDH is a phosphoprotein whose activity is inhibited by increased phosphorylation, and conversely is stimulated by depho~phorylat ion.~~,~~ These results, together with the in vitro results from Jarett’s laboratory, suggested that the material whose generation/ release from the membrane was promoted by insulin might mediate insulin’s effects on the PDH complex and stimulate PDH activity concomitantly with promoting its dephosphorylation.
Seals and Jarett” showed that the addition of insulin to mixtures of adi- pocyte plasma membrane and mitochondria activated the PDH, compared with control mixtures receiving BSA in place of insulin. The concentration-effect curve of insulin action was biphasic, increasing over the concentration range 50-100 p,U/ml, and decreasing from 100 to 250 p,U/ml. The range of insulin’s effectiveness was similar to the physiologic range of insulin action on fat cell metabolism. The effect of insulin was specific for the intact hormone, since neither boiled insulin nor the desoctapeptide of insulin had any detectable effect on the enzyme when they were tested at the concentration at which native insulin was maximally active. The insulin-mimetic agents Concanavalin-A and anti-insulin receptor antibody also stimulated the PDH in dose-dependent, bi- phasic manners. None of the active agents had any detectable effects on PDH activity when they were added directly to mitochondria in the absence of plasma membrane. In addition, Seals and Czech33 and Kiechle et a1.34 showed that material which stimulated PDH activity could be recovered in the supernatant fraction after treating adipocyte plasma membranes with insulin and subse- quently removing the membranes by centrifugation. This ruled out the possi-

GOTTSCHALK AND JARETT 233
bility that direct contact between mitochondria and plasma membranes in the presence of insulin was necessary for the insulin effect.
In collaboration with Larner’s group, Jarett and Seals35 found that the ma- terial extracted from rabbit skeletal muscle activated PDH in isolated adipocyte mitochondria without added plasma membranes, and that extracts from insulin- treated animals activated the enzyme to a greater extent than did extracts from control animals. These results suggested that the extracts of insulin-treated mus- cle contained an active principle that was similar to, if not identical with, the mediator generated by interaction of insulin with its receptors on the adipocyte plasma membrane.
Thus, by the early 1980s, nearly 60 years after the first reported isolation of insulin in homogenous form, evidence adduced in two laboratories, working independently of each other and following different experimental approaches, showed that insulin elicits at least some of its biological effects by promoting the generationhelease of unique intracellular mediator(s).
111. TISSUE SOURCES OF INSULIN MEDIATORS
Insulin mediators have been detected from a variety of sources, including plasma membranes from insulin-sensitive cells, whole cells, and intact tissue from insulin-treated animals. A number of enzymes and metabolic systems have been employed to detect mediators in these studies, including glycogen-syn- thase D phosphatase, CAMP-dependent protein kinase, pyruvate dehydrogen- ase phosphatase, adenylate cyclase, phospholipid methyltransferase, acetyl-CoA carboxylase, low-K, CAMP phosphodiesterase, glucose-6-phosphatase, anti- lipolysis, and lipogenesis. The manner in which insulin mediators affect these enzymes is discussed in greater detail in section IV.
A. Plasma Membrane Sources
Plasma membrane preparations have been employed as convenient sources of small quantities of insulin mediators in a number of laboratories. Table I.A. summarizes the membrane systems that have been used to generate mediators of insulin action and the enzymes whose activities these mediators have been found to modulate.
Preliminary characterization of both the generation of mediator activity and certain of the mediator’s physicochemical properties were conducted in two laboratories, both using rat adipocyte membranes. Kiechle et al.34 found that mediator activity was spontaneously released from adipocyte membranes that had been prepared in Tris buffer, and more activity, as judged by the stimulation of PDH, was released by membranes prepared in Tris buffers containing EDTA or EGTA than in Tris buffers alone. The mediator-activity content of the Tris- treated membranes could be depleted by repeated incubation and centrifugation of the membranes. The material release from Tris-treated membranes resulted in a linear dose-response curve on PDH activity, and was stable at 4°C for at least 16 h at pH 7.0 but not pH 7.4. It eluted as a single, low-molecular-weight peak during chromatography on Sephadex G-15 or G-25.

234 INSULIN MEDIATORS
Table I. Sources of Insulin Mediator Enzymes and Metabolic
Source System Affected Reference A. Membrane sources Rat adipocyte PDH
LOW-K, PDE' (Ca2 + + Mg2 + )-ATPase Lipolysis Lipogenesis Intracellular cAMP accumulation
Adenylate cyclase Acetyl-CoA carboxylase Lipogenesis
Glucose-6-phosphatase
Rat liver PDH
Human placenta PDH
B. Intact cell and tissue sources Rat adipocyte PDH
Rabbit skeletal CAMP-dependent protein kinase muscle Glycogen synthase phosphatase Rat skeletal muscle PDH
LOW-K, PDE*
Adenylate cyclase
Phospholipid methyltransferase Lipolysis
Adenylate cyclase
Adenylate cyclase Phospholipid methyltransferase Mitochondria1 Ca2+ transport
Adenylate cyclase
Adenylate cyclase
LOW-K, PDE'
Rat heart PDH
Rat liver PDH
Rat hepatocytes PDH
Hepatoma cells PDH
LOW-K, PDE* Human monocytes PDH
IM-9 lymphocytes PDH Glucose-6-phosphatase
Adenylate cyclase
*Low-& cAMP phosphodiesterase. tMacaulay, Kelly, and L. Jarett, unpublished observations. $Parker and L. Jarett, unpublished observations. $Both t and 1 after treatment with dexamethasone.68
T , 1 t T 1 t
.1 t1.l t
T l . 1 1
T, 1 T 1 t T, .1 1 t 1 1 t
t t 1
38 49 96 45 45 45 37 38 41 46 43,44 44
47,48 49 20 20 52, 58 58 58 t t 57 57 58 58 t 11 69 69 67 t 67 44 44 71 fi
Membranes prepared in phosphate buffers did not spontaneously release insulin-dependent mediator, but insulin treatment of such membranes did re- sult in the release of mediator a ~ t i v i t y . ~ ~ , ~ ' These results were confirmed by Kiechle et al. ,34 who also showed that the insulin-elicited material from phos- phate-treated membranes eluted in the same position as the material derived from Tris-treated membranes during chromatography on Sephadex G-25. Seals and Czech showed that the mediator generated by insulin from phosphate-

GOTTSCHALK AND JARETT 235
treated membranes was sensitive to treatment with pro tease^,^^ was stable for short times (12 min) at high temperatures (100°C) and under basic conditions (0.1 N NaOH), and bound reversibly to both anion-exchange and hydroxyapa- tite columns at pH 7.4.36 They also found that mediator generated by super high concentrations of insulin displayed biphasic dose-response effects on py- ruvate dehydrogenase activity, consistent with results of Seals and Jarett.I2 This result suggested either that high concentrations of mediator inhibited the PDH, or that insulin elicited the generation of both PDH-inhibiting and PDH-activat- ing activities. The multiplicity of mediator activities generated by the interaction of insulin with plasma membranes will be discussed in section V.
Rat liver membranes have also been used in a number of investigations as sources of insulin mediator activity. Jarett and K i e ~ h l e ~ ~ showed that liver mem- branes spontaneously released material that activated either liver or adipocyte mitochondria1 pyruvate dehydrogenase. Insulin treatment of a liver particulate fraction in phosphate buffer enhanced the generation of activities that modulate PDH,3840 adenylate c y ~ l a s e , ~ ' , ~ ~ acetyl-CoA c a r b o ~ y l a s e , ~ ~ , ~ ~ and glucose-6- p h ~ s p h a t a s e . ~ ~ Ethanol extraction of the lyophilized supernatant of insulin-treated membranes yielded an ethanol-insoluble fraction that stimulated PDH and ace- tyl-CoA carboxylase and an ethanol-soluble fraction that inhibited PDH and adenylate c y c l a ~ e ~ ~ and acetyl-CoA carboxyla~e.~~ The PDH-stimulating material was active when either adipocyte or liver mitochondria were used in the assay.41
Placenta is an insulin-responsive tissue, rich in insulin receptors, and thus a potentially useful source of insulin mediators. Two groups have shown that physiologically relevant concentrations of insulin promote the generation of PDH-modulating activity from human placental membranes. Sakamoto et al.43 reported that activity was detectable only after gel filtration of the supernatant fractions of insulin-treated membranes, and was not observed in the crude supernatants before fractionation. No stimulating activity was detected, either before or after chromatography, in supernatant fractions from the control mem- branes, and only PDH-inhibiting activity was detected after chromatography. This inhibitory material was not characterized further.
Suzuki et al.44 also found PDH-stimulating material from placental mem- branes. However, in contrast to the results of Sakamoto et al., activity was detected both in the crude supernatant fractions and after gel filtration; no significant amounts of inhibiting material was observed after gel filtration of either control or insulin-treated supernatants. The reason for this discrepancy was not clear. Suzuki et al. also showed that, similarly to PDH mediator from adipocyte membranes,36 the PDH-stimulating material from placental mem- branes bound reversibly to both anion-exchange and hydroxyapatite chromato- graphic supports.
Two groups have used assays that are based on intact cells, rather than cell homogenates or partially purified enzymes, to examine the effects of insulin mediators extracted from plasma membranes on metabolic systems, such as lipolysis, which are not completely amenable to in vitro analysis. Zhang et al.45 tested extracts of insulin-treated adipocyte plasma membranes for their ability to mimic insulin action on intact adipocytes. Before testing, insulin was removed from the extracts by gel filtration chromatography. The extracts inhibited both basal and hormone (glucagon + forsko1in)-stimulated CAMP accumulation. The

236 INSULIN MEDIATORS
basis of this action is not understood, however, since the extract had no effects on either adenylate cyclase or low-K, CAMP phosphodiesterase in broken cell preparations. The extracts also inhibited both basal and hormone-stimulated lipolysis and stimulated lipogenesis in a dose-dependent manner. Whether or not all three biological effects are due to the same substances awaits further purification.
An insulin-dependent lipogenic activity was also recovered from a liver particulate fraction by Car0 et a1.& These workers tested for lipogenesis using intact cultured hepatocytes, and they found that, in addition to lipogenesis, activity in the supernatant fraction also promoted insulin-receptor downregulation. These biological effects were not due to contaminating insulin in the supernatant fraction, since no insulin was detected in the supernatants by RIA, or when 1251-insulin was used to generate the mediator substance. The active principles have not been characterized further, and its (their) relationship to other mediator activities, especially the lipogenic activity reported by Zhang et al.,45 awaits purification and chemical characterization of the respective active materials.
Plasma membranes from both liver and adipocytes have been used exten- sively as sources of insulin mediators in studies examining the physiologic roles played by the mediators. These results are reviewed in section VIII. Most of the physicochemical properties of the insulin mediators have been determined us- ing material generated from plasma membranes, and these results are sum- marized in section VI.
B. Whole Cells and Intact Tissue
The presence of mediators in tissue extracts, in quantities that are de- pendent on the physiologic condition of the tissue, is one proof that these substances subserve biologically important roles. In addition, intact tissues are probably the only sources rich enough to provide the amounts of material that are needed for the full chemical characterization and identification of insulin mediators. Table I.B. summarizes the tissues from which insulin mediators have been extracted and the enzymes whose activities they modulate.
2. Adipocytes Kiechle et al.47 showed that PDH-stimulating mediator activity could be
extracted from intact adipocytes, and that more activity was extractable from insulin-treated cells than from control cells that had been treated with BSA. These results were confirmed by Seals and Czech.48 Kiechle and Jarett49 also demon- strated that an insulin-dependent mediator of low-Y, CAMP phosphodiesterase was extractable from intact adipocytes. This material had no measurable effects on a high-Km CAMP phosphodiesterase from adipocytes, consistent with the reported lack of effect of insulin treatment of intact cells on this enzyme.50
2. Muscle
Lamer et al.17 were the first to demonstrate the presence of insulin mediator activity in insulin-treated tissue. They observed that extracts of skeletal muscle from insulin-treated rabbits inhibited cAMP-dependent protein kinase to a greater extent than muscle extracts from control animals. The crude extract was frac-

GOTTSCHALK AND JARETT 237
tionated by heat denaturation in acid to remove the bulk of the protein, by paper chromatography to remove nucleotides, and then gel filtered on Sephadex G-25.“ Mediator activity eluted from the G-25 column as a single peak, in the molecular-weight range of 1500 daltons. It specifically inhibited CAMP-de- pendent protein kinase and had no detectable effects on CAMP-independent kinases. Material in this column fraction also stimulated glycogen synthase D phosphatase and pyruvate dehydrogenase.
Recently, by employing combinations of gel filtration and HPLC ion-ex- change chromatography, Thompson et al.52 and Cheng et a1.53J03 have reported the purification to near homogeneity of both the CAMP-dependent protein ki- nase and the PDH-stimulating mediator activities.
Jarett’s laboratory has identified four mediator activities in extracts of rat m ~ s c l e . ~ ” ~ Both skeletal and heart muscle extracts possess PDH-stimulating and adenylate cyclase-inhibiting mediator activities, and the levels of both were enhanced by insulin treatment. Insulin treatment also enhanced the levels in skeletal muscle extracts of both a phospholipid methyltransferase-inhibiting me- diator (K. L. Kelly, S. L. Macaulay, and L. Jarett, unpublished observations) and a Iow-K, CAMP phosphodiesterase-stimulating mediator,58 but the effect of heart extracts on these enzymes has not yet been studied. On the basis of dilution assays, heart appears to be the richest source of the PDH and the adenylate cyclase mediators yet examined. Thompson et al.52 and Cheng et al.’03 have also identified a PDH-stimulating mediator in rat skeletal muscle.
The effects of the skeletal muscle extract on adipocyte metabolism have been examined using a whole (intact) cell assay system. Partially purified me- diator stimulated PDH activity in a dose-dependent f a ~ h i o n ~ ~ , ~ ~ and, in addition, stimulated glycogen synthase but had no effect on either glucose oxidation or glucose t r a n ~ p o r t . ~ ~ Mediator from skeletal muscle also inhibited isoproterenol- stimulated phospholipid methyltransferase and lipolysis (K. L. Kelly, S. L. Macaulay, and L. Jarett, unpublished observations). Effects on either adenylate cyclase or lipid metabolism have not been examined to date.
3. Liver In a series of studies on the action of insulin on liver mitochondria, Gain-
utdinov et a1.11,6042 reported the presence in liver extracts of a low-molecular- weight regulator of mitochondrial Ca2+ transport. The activity, which stim- ulated mitochondrial Ca2+ uptake in vitro, was enhanced in liver extracts of insulin-treated rats. In addition to, or perhaps as a result of, its effects on mitochondrial Ca2+ transport, the insulin-dependent activity inhibited epineph- rine-stimulated oxidation of pyruvate and succinate in rat liver mitochondria in vitro. It also exhibited a number of in vivo biological effects that are relevant to insulin action, including antagonizing both gluconeogenesis and epinephrine- stimulated glucose release from rat liver slices, and lowering blood glucose levels in rats made hyperglycemic by alloxan treatment. It was partially purified by anion-exchange and gel filtration chromatography and shown to possess the properties of a heat-stable glycopeptide; 43 amino acids were identified after acid hydrolysis of the active material.60 However, no evidence was provided that these amino acids were part of the mediator molecule. Further studies are

238 INSULIN MEDIATORS
required to establish the relationship of this mediator to the other mediators discussed in this review.
A number of researchers have shown that insulin affects nuclear functions. Purrello et al.63 demonstrated that insulin activated a nucleoside triphosphatase located on the nuclear envelope, and further showed that treatment with insulin or insulin-mimetic agents decreased 32P-incorporation into nuclear envelope protein^.^,^^ This may be one way in which insulin regulates mRNA metabo- lism. In this context, it is of interest that Horvat66 reported that insulin promoted the generation of a low-molecular-weight moiety in liver that stimulated nuclear RNA polymerase activity.
PDH- and low-K, cAMP phosphodiesterase-stimulating and adenylate cy- clase- and phospholipid methyltransferase-inhibiting mediator activities have been extracted from rat liver by Macaulay et a1.54,58 (S. L. Macaulay, K. L. Kelly, and L. Jarett, unpublished observations). More activity was recovered from livers of rats treated with insulin than from control rats. On the basis of their behavior during purification, the PDH- and adenylate cyclase-modulating ac- tivities appear identical to those extracted from rat skeletal muscle. Parker et al.67 treated H4-11-EC3’ hepatoma cells with insulin and found increased amounts of PDH- and low-K, cAMP phosphodiesterase-stimulating activity. The activi- ties were heat- and acid-stable, and possessed apparent molecular weights of 1000-2000. Similarly to the material extracted from adipocyte membranes,36 the PDH-stimulating mediator extracted from H4-11-EC3’ cells bound reversibly to an anion-exchange resin (Dowex-1). This is in contrast to the behavior of the PDH- and adenylate cyclase-modulating mediators extracted from muscle and intact liver by Macaulay et al., which bound to cation but not anion ex- changers,% and may reflect tissue differences in the detailed structures of these mediators.
A strikingly unique feature of the H4-11-EC3‘ cell mediator is that the dose- response curve of insulin-promoted production of mediator activity increased linearly to a plateau level and did not exhibit biphasic behavior. This result has been confirmed by Esau and Koontz,68 who used Reuber H-35 hepatoma cells. In addition, these workers reported that cultures which were treated with dex- amethasone before incubation with insulin produced, in an insulin-dependent manner, both PDH-activating and PDH-inhibiting activities. Dexamethasone treatment alone did not result in generation of the inhibiting activity. The im- plications of these results on the mechanism of generation of mediator activity, and on the multiplicity of mediator activities elicited by insulin, will be discussed in section V.
Saltie140 has also reported the extraction of insulin-dependent PDH- and adenylate cyclase-modulating mediator activities from rat hepatocytes, but not enough information is available to allow a comparison between these materials and those reported by other workers.
A cAMP antagonist, which inhibits both CAMP-dependent protein kinase and adenylate cyclase and activates phosphoprotein phosphatase and PDH, has been identified in rat hepatocyte extracts by W a ~ n e r . ~ ~ The activities were elicited in response to treatment with either epinephrine or insulin, but were not present in extracts of control cells. Pretreatment of rats with indomethacin

GOTTSCHALK AND JARETT 239
resulted in diminished recoveries of the adenylate cyclase inhibitor after epi- nephrine stimulation of the isolated hepatocytes, but recoveries could be re- stored to normal levels by the addition of prostaglandin E l (PGE-1) during epinephrine treatment. PGE-1 itself was without effect on adenylate cyclase in these studies, and could be separated from the mediator activity by anion- exchange chromatography. It is possible that PGE-1 either forms an integral part of the mediator or is essential for the generation of the mediators. Was- ner has reported that partially purified mediator contains both prostaglandin and inositol phosphate.70 The active component(s) in Wasner’s preparation differs biologically from the CAMP-lowering activity described by Zhang et al.,45 since the activity described by Zhang et al. was apparent only when intact cells were employed for assay, while Wasner’s cAMP antagonist was report- edly active when either membranes or intact hepatocytes were used. The ac- tivity prepared by Wasner also differed chemically from the adenylate cyclase mediator reported by Jarett’s laboratory in that it was extractable into organic solvents whereas the activity prepared by the Jarett group was soluble only in aqueous buffers (unpublished results). Other properties of the cAMP antago- nist studied by Wasner differed significantly from those reported by other la- boratories in this field, and it appears that this prostaglandinlike material is, in fact, biochemically distinct from the other adenylate cyclase-inhibiting ac- tivities discussed in this review.
Human peripheral blood cells are richly endowed with insulin receptors. Since the generation of insulin mediators depends on ligand-receptor interac- tions, it was expected that these cells might provide an abundant source of insulin-dependent mediator. In addition, knowledge of the mediator generation system from human sources might have application in the study of abnormal conditions in man.
Insulin treatment of IM-9 lymphocytes resulted in decreased PDH-stimu- lating activity in the cell extract compared with the extracts of control cells.71 Although these results were unexpected, they are consistent with the fact that insulin has no detectable postbinding biological effects on these cells. The fur- ther implications of these results will be discussed in section V. In contrast, insulin treatment enhanced the amounts of PDH-stimulating activity that was extracted from human m o n ~ c y t e s . ~ ~ These cells also yielded increased amounts of glucose-6-phosphatase-inhibiting activity after insulin treatment. Both activ- ities are low-molecular-weight moieties (1000-2000 daltons), are heat- and acid- stable, and both bind reversibly to the anion-exchange resin Dowex-1 and to hydroxyapatite in ways reminiscent of the PDH-stimulating activity from adi- pocyte membrane^.^^
In summary, mediator activity has been extracted from a variety of sources, including plasma membranes, whole cells, and intact tissues, and in each case (with the exception of IM-9 cells) treatment with physiologic concentrations of insulin enhanced the amount or activity of the putative mediators that were extracted. These results substantiated the proposal that insulin promotes the generation and/or release of intracellular messengers, which appear to mediate many of insulin’s effects on metabolism. The physiologic and pathophysiologic importance of these activities are discussed in section VIII.
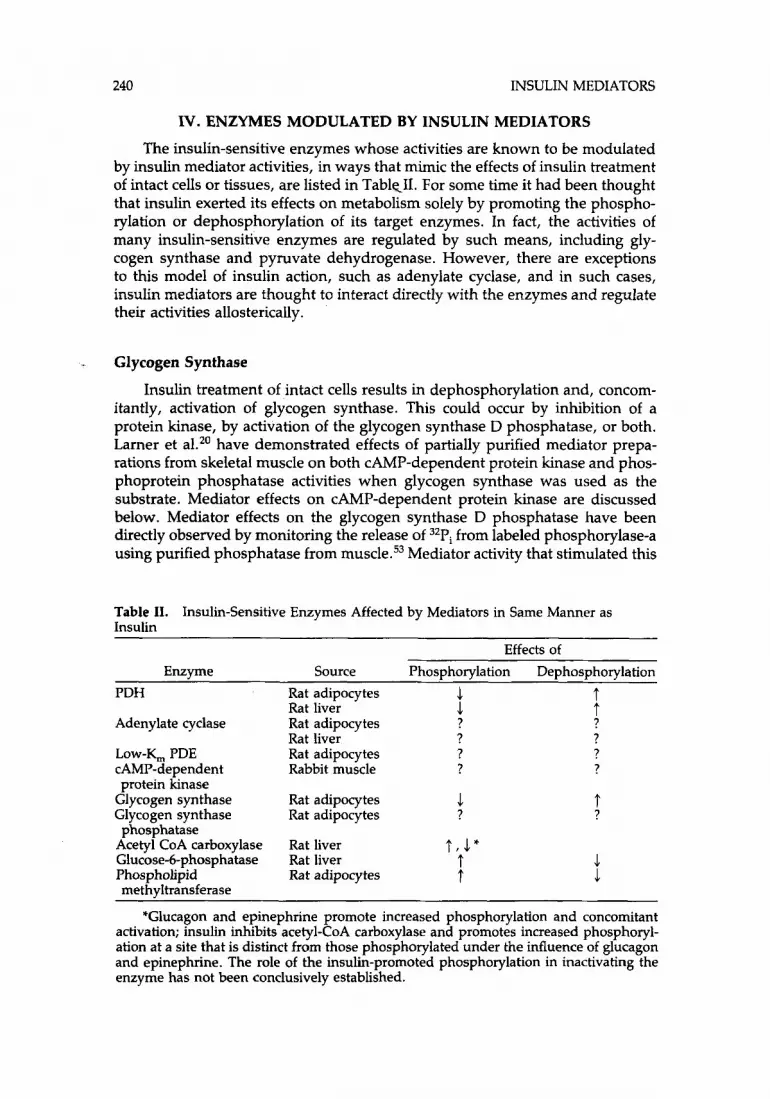
240 INSULIN MEDIATORS
IV. ENZYMES MODULATED BY INSULIN MEDIATORS
The insulin-sensitive enzymes whose activities are known to be modulated by insulin mediator activities, in ways that mimic the effects of insulin treatment of intact cells or tissues, are listed in TablCII. For some time it had been thought that insulin exerted its effects on metabolism solely by promoting the phospho- rylation or dephosphorylation of its target enzymes. In fact, the activities of many insulin-sensitive enzymes are regulated by such means, including gly- cogen synthase and pyruvate dehydrogenase. However, there are exceptions to this model of insulin action, such as adenylate cyclase, and in such cases, insulin mediators are thought to interact directly with the enzymes and regulate their activities allosterically.
Glycogen Synthase
Insulin treatment of intact cells results in dephosphorylation and, concom- itantly, activation of glycogen synthase. This could occur by inhibition of a protein kinase, by activation of the glycogen synthase D phosphatase, or both. Larner et aLZ0 have demonstrated effects of partially purified mediator prepa- rations from skeletal muscle on both CAMP-dependent protein kinase and phos- phoprotein phosphatase activities when glycogen synthase was used as the substrate. Mediator effects on CAMP-dependent protein kinase are discussed below. Mediator effects on the glycogen synthase D phosphatase have been directly observed by monitoring the release of 32Pi from labeled phosphorylase-a using purified phosphatase from muscle.53 Mediator activity that stimulated this
Table 11. Insulin
Insulin-Sensitive Enzymes Affected by Mediators in Same Manner as
Effects of Enzyme Source Phosphorylation Dephosphorylation
PDH Rat adipocytes 1 t .1 t Rat liver
Adenylate cyclase Rat adipocytes ? ? Rat liver ? ?
LOW-Y, PDE Rat adipocytes ? ? CAMP-dependent Rabbit muscle ? ? protein hnase
Glycogen synthase Rat adipocytes 1 Glycogen synthase Rat adipocytes ?
Acetyl CoA carboxylase Rat liver t , 1" Glucose-6-phosphatase Rat liver t Phospholipid Rat adipocytes t
phosphatase
methyltransferase
t ?
1 .1
*Glucagon and epinephrine promote increased phosphorylation and concomitant activation; insulin inhibits acetyl-CoA carboxylase and promotes increased phosphoryl- ation at a site that is distinct from those phosphorylated under the influence of glucagon and epinephrine. The role of the insulin-promoted phosphorylation in inactivating the enzyme has not been conclusively established.

GOTTSCHALK AND JARETT 241
phosphatase has been partially p~rified,’~ and by the criteria of gel filtration chromatography it appeared larger than, and therefore distinct from, either the PDH-stimulating or the protein kinase-inhibiting activities.
B. Pyruvate Dehydrogenase
PDH is a hormonally regulated enzyme which catalyzes one of the rate- limiting steps in fatty acid biosynthesis. Numerous laboratories have made ex- tensive use of pyruvate dehydrogenase as a routine assay for the identification and characterization of insulin mediator activity. PDH is a multi-enzyme com- plex composed of five enzymes: pyruvate decarboxylase, dihydrolipoyl acetyl- transferase, dihydrolipoyl dehydrogenase, a CAMP-independent protein kinase, and a phosphoprotein phosphatase. The activity of the complex is regulated by the extent of phosphorylation of the a-subunit of the decarboxylase, which is determined by the competing effects of the PDH-kinase and the PDH-phos- phatase. The dephosphorylated enzyme is the active form both in vitro and in vivo. The decarboxylase may be phosphorylated at any of three serine residues. Although only phosphorylation at the first residue inactivates the enzyme, more heavily phosphorylated forms are reactivated by the phosphatase more slowly than are more lightly phosphorylated forms. This shows that the phosphatase works on all three sites, and the first one phosphorylated is the one that controls the activity of the decarboxylase. The regulatory properties of the PDH complex have been reviewed recently by Reed” and Wieland.73
The insulin mediator could activate the a-subunit by either inhibiting the kinase or by activating the phosphatase. To decide between these possibilities, Popp et al.74 incubated plasma membrane/mitochondria mixtures with insulin or Con-A in the presence of sodium fluoride, a PDH-phosphatase inhibitor,75 or dichloroacetic acid, a PDH-kinase i n h i b i t ~ r . ~ ~ Sodium fluoride completely blocked the ability of either insulin or Con-A to activate PDH activity, whether or not ATP, a substrate for the kinase, was also present in the incubation mix- ture. Dichloroacetic acid (DCA) alone had no effect on enzyme activity in the absence of ATP, but stimulated activity, compared with controls without the kinase inhibitor, in the presence of ATP. In the absence of ATP, the addition of DCA had no effect on the stimulation of PDH activity by either insulin or Con-A. However, in the presence of added ATP the effects of DCA were addi- tive with those of either ligand. These data showed that stimulation of the a- subunit of pyruvate dehydrogenase by insulin mediator occurred via activation of the phosphoprotein phosphatase, and not by inhibiting the PDH-kinase. Similar studies, leading to the same conclusions, were performed with partially purified mediator preparations from a d i p ~ c y t e s , ~ ~ adipocyte plasma mem- b r a n e ~ , ~ ~ hepatoma cells,67 and human monocytes.44
Macaulay and Jarett77 have demonstrated this effect more directly by using mitochondria that had been prelabeled with [y-32P]ATP. As revealed by SDS- PAGE analysis, the a-subunit of the PDH complex was the only mitochondria1 protein detectably labeled. Insulin mediator, in the absence of added ATP, pro- moted the dephosphorylation of labeled mitochondria with a time course par- alleling the activation of pyruvate dehydrogenase measured on the same mitochondria. Both dephosphorylation and activation were abolished by so-

242 INSULIN MEDIATORS
dium fluoride. In addition, Newman et al.7R have reported that supernatant fluids from insulin-treated adipocyte membranes stimulated the activity of pur- ified PDH-phosphatase, but had no detectable effect on purified PDH-kinase.
Most routine assays of PDH activity in this and other laboratories have involved the use of broken mitochondria. One criticism of this practice was that such mitochondria did not accurately reflect physiologic conditions, and that the PDH complex was exposed to a number of agents that may artifactually modulate activity. Parker and Jarett54 addressed this problem by treating fully intact, respiring mitochondria with insulin mediators from liver membranes, hepatoma cells, IM-9 lymphocytes, and liver and muscle. In all cases except the IM-9 cells, mediators from insulin-treated sources stimulated PDH to greater extents than mediators from control sources. In addition, Jarett et a1.55,56 have shown that insulin activity from rat skeletal muscle stimulates pyruvate dehy- drogenase in intact adipocytes. These results showed that mediators can elicit physiologically relevant responses of PDH in intact systems.
C. Glucose-6-Phosphatase
Glucose-6-phosphatase catalyzes a key reaction in carbohydrate metabo- lism. Activation of the enzyme increases glucose levels while inhibition leads to increased glucose-6-phosphate levels, which may feed into lipogenesis, among other pathways. Insulin has been shown to inhibit the activity of glucose-6- phosphatase from rat liver micro~omes ,~~ perhaps by promoting its dephos- phorylation.80,s1 Suzuki et a1.42,44 have partially purified from human placenta and monocytes and from rat liver an activity that inhibits glucose-6-phos- phatase. This mediator activity co-eluted from gel filtration, ion-exchange (Dowex-1), and hydroxyapatite columns with PDH-stimulating mediator activ- ity. These results suggested that the same active principle may be responsible for both inhibition of glucose-6-phosphatase and stimulation of PDH, but this suggestion cannot be ascertained until both mediator substances have been purified and identified.
D. Phospholipid Methyltransferase
Phospholipid methyltransferse (PLMT) is a membrane-bound enzyme, which is ubiquitously distributed in mammalian tissues. It catalyzes the stepwise meth- ylation of phosphatidylethanolamine to phosphatidylcholine in response to re- ceptor-ligand interactions. PMLT appears to be important in the transduction of receptor-mediated signals and/or in mediating cellular responses, although the exact nature of its role in these processes is not understood (for review, see refs. 82 and 83). Glucagon, beta-adrenergic agonists, forskolin, human chorionic gonadotropin (hCG), and adrenocorticotropin (ACTH) stimulated PLMT activ- ity, apparently via a CAMP-dependent process.8446 Partially purified PLMT from rat liver has been phosphorylated and activated in parallel fashion by exoge- nously added CAMP-dependent protein k i n a ~ e . ~ ~ These data are consistent with the proposal that the activity of PLMT may be regulated by phosphorylation- dephosphorylation.

GOTTSCHALK AND JARETT 243
Hormones such as ACTH and epinephrine, whose actions on adipocytes oppose those of insulin, cause a stimulation of adipocyte PLMT in a manner that can be mimicked by added dibutryl CAMP or forskolin. Physiologic con- centrations of insulin antagonize the stimulatory effects of ACTH, epinephrine, and forskolin on PLMT, and insulin alone causes a concentration-dependent inhibition of PLMT (K. L. Kelly, E. H. A. Wong, and L. Jarett, submitted for publication). Partially purified preparations of insulin mediator from rat skeletal muscle and liver mimic this insulin effect on intact adipocytes, and more of this inhibitory activity is present in extracts from insulin-treated tissue than in ex- tracts from control tissue. Mediator extracted from skeletal muscle also inhibited isoproterenol-stimulated PLMT in rat adipocyte and liver plasma membranes (K. L. Kelly, S. L. Macaulay, and L. Jarett, unpublished results). These results suggest that the inhibitory effects of insulin on adipocyte PLMT may be via dephosphorylation, mediated by an as yet unidentified insulin mediator.
E. Acetyl-CoA Carboxylase
The activity of acetyl-CoA carboxylase, which catalyzes a rate-limiting step in fatty acid biosynthesis, is hormonally regulated. Glucagon and epinephrine promoted CAMP-dependent phosphorylations of the enzyme, which resulted in its inactivation.88 Insulin activated acetyl-CoA carboxylase and, paradoxically, also promoted the enzyme's phosphorylation, but at a site distinct from those sensitive to glucose or epinephrine and via a CAMP-independent protein ki- n a ~ e . * ~ - ~ l It has not yet been conclusively demonstrated that the phosphoryla- tion of acetyl-CoA carboxylase promoted by i n s u h is causally related to insulin's activation of this enzyme; the relationship between these events is currently under investigation in a number of laboratories.
Saltiel et al.40,41 recovered activities that modulated acetyl-CoA carboxylase in the supernatant fractions from insulin-treated liver particulate fractions. An acetyl-CoA carboxylase-stimulating activity was present in the insoluble residue after extraction of lyophilized supernatant fractions with ethanol, and an inhib- iting mediator was detected in the ethanol-soluble portion.
F. (Ca2+ + Mg2+)-ATPase
Treatment of intact adipocytes or isolated adipocyte plasma membranes with physiologic concentrations of insulin resulted in the dephosphorylation of a phosphoprotein of 110,000 MW. This protein has been identified as a high- affinity (Ca2+ + Mg2+)-ATPase,y2 and may be the enzymatic basis for a high- affinity, calmodulin-sensitive plasma membrane Ca2 + -transport ~ y s t e m . ~ ~ , ~ ~ Physiologic concentrations of insulin inhibited both the (Ca2+ + Mg*+)-ATPase and Ca2+-transport in intact adipocytes as well as in isolated adipocyte mem- b r a n e ~ . ~ ~ In contrast, incubation of adipocyte membranes with a crude insulin mediator preparation derived from adipocyte membranes stimulated (Ca2 + + Mg2+)-ATPase fourfold and stimulated Ca2+ transport more than twofold.y6 These opposing effects of insulin and of insulin mediator remain unexplained. They may have been due to contaminants present in the membrane extract, and the effect of insulin mediators on this ATPase needs reinvestigation using in- sulin mediator preparations of greater purity.

244 INSULIN MEDIATORS
G . CAMP-Dependent Protein Kinase
Larner’s laboratory demonstrated that insulin treatment of intact tissue in- hibited CAMP-dependent protein kinase by maintaining the enzyme in its in- active, holoenzyme form and desensitizing it to the effects of CAMP.'^,^^ Larner et al. l7 were also the first to show that extracts of insulin-treated tissue contained an active principle that inhibited this protein kinase, and decreased 13H]cAMP binding to it. They have used this enzyme extensively as the basis of an assay for insulin mediators, and, as discussed above, have substantially purified a mediator activity that appears to inhibit CAMP-dependent protein kinase spe- cifically. CAMP-dependent protein kinase has not been proven to be a phos- phoprotein.
H. Enzymes of cAMP Metabolism
It has been extensively documented that insulin treatment lowers hormon- ally elevated cAMP levels in intact a d i p ~ c y t e s ~ ~ , ~ ~ and other tissues.98 Insulin may bring about this effect by either inhibiting hormone-stimulated cAMP bio- synthesis or by enhancing the rate of cAMP degradation. Both adenylate cyclase and low-& cAMP phosphodiesterase activities are modulated by insulin me- diators in subcellular systems. In this regard, however, it is important to recall that the CAMP-lowering mediator reported by Zhang et al.45 had no detectable direct effects on either enzyme in cell homogenates.
1. Adenylate Cyclase
Insulin inhibits both basal and hormonally stimulated adenylate cyclase activities in adipocyte and hepatocyte plasma membranes ,99,100 No firm evidence has been provided that adenylate cyclase is regulated by phosphorylation or dephosphorylation.
Saltiel et al.38 identified an inhibitor of adenylate cyclase in lyophilized supernatant fractions from insulin-treated liver particles. The inhibitor was ethanol soluble, and was distinguishable on this basis from an ethanol-insoluble, PDH- stimulating activity that was also present in the crude extract.
Adenylate cyclase inhibitors have also been identified in extracts of insulin- treated rat skeletal m ~ s c l e , ~ ~ , ~ ~ rat liver,58 and rat heart.57 However, significant differences in the gel filtration characteristics of this mediator were reported by Macaulay et a1.57,58 and Cheng et al.53 Macaulay et al. reported that the inhibitor eluted after PDH-stimulating activity during chromatography on Sephadex G- 15, with an apparent molecular weight of 1500, whereas Cheng et al. claim this mediator activity elutes before PDH-stimulating activity, with an apparent mo- lecular weight of 3000-5000, during chromatography on Sephadex G-25. These discrepancies may reflect differences in the column buffers used67 and/or dif- ferences in the assay system. The cyclase-inhibiting activity isolated by Jarett’s laboratory also differed from that reported by Saltiel et al.3s in that the former was insoluble in 100% ethanol while the latter was soluble in ethanol.
As summarized above, both W a ~ n e r ~ ~ and Zhang et al.45 have detected activities that lowered hormone-elevated cAMP levels in whole cells. However,

GOTTSCHALK AND JARETT 245
the relationship of these activities with each other and with the adenylate cy- clase-inhibiting activities reported by other investigators will not be clear until the respective active principles have been purified and identified.
2. Low-K, C A M P Phosphodiesterase
Insulin treatment of hepa t~cy te s~~ and a d i p o c y t e ~ ~ ~ , ' ~ ~ increased the low- K, cAMP phosphodiesterase activity in isolated microsome fractions and in adipocyte plasma membranes."' A mediator activity that stimulated low-K, cAMP phosphodiesterase has been identified in extracts of rat ad ip~cy tes ,~~ H4-11-EC3' hepatoma cells,67 and rat liver and skeletal muscle,58 as well as the supernatant fractions from rat adipocyte plasma membranes.49 In each case, recoveries of the mediator activity were enhanced in samples from insulin- treated cells or tissue. Low-K, cAMP phosphodiesterase has not been proven to be regulated by phosphorylation or dephosphorylation.
V. EVIDENCE FOR A FAMILY OF MEDIATORS OF INSULIN ACTION
This section summarizes the evidence, often alluded to in the previous discussion, that insulin generates a family of related compounds that mediate many of its actions.
Indirect evidence that insulin elicited the generation of both stimulating and inhibiting mediators was provided by two kinds of experiments. First, the dose-response curves of insulin or of insulin-mimetic agents with regard to mediator production were biphasic when plasma membrans were used to gen- erate the mediators. 12,36*38,39 Second, dilution curves of the membrane super- natant fractions were also biphasic in the assays of PDH.36,39,44 acetyl-CoA carb~xylase,~~ and glucose-6-phosphatase.44 Larner et aL20 also showed that the insulin-dependent mediator from skeletal muscle produced a biphasic dose- response curve in the CAMP-dependent protein kinase assay. The mediator dose-response experiments suggested that the stimulating mediator was more active at low concentrations than was inhibiting activity, or that more stimulat- ing mediator was generatedheleased at lower concentrations, whereas at higher concentrations more of the inhibiting mediator was produced.
The insulin-promoted generation of multiple mediators has been demon- strated more directly by both biological and biophysical means. Cheng et al. were the first to report the separation of stimulating from inhibiting mediator activities.lo2 They fractioned a partially purified mediator preparation into two zones by high-voltage paper electrophoresis. Activity in one zone inhibited CAMP-dependent protein kinase and stimulated glycogen synthase phospha- tase, while activity in the other zone had the opposite effects. Larner's labora- tory has recently reported the preparation of both the CAMP-dependent protein kinase-inhibiting mediator and the PDH-stimulating mediator.53 In addition, they have reported that the adenylate cyclase-inhibiting and glycogen synthase phosphoprotein phosphatase-stimulating mediator activities can be separated from the PDH and the protein kinase mediators by gel filtration on Sephadex G-25 columns.

246 INSULIN MEDIATORS
Saltiel et al.3s were the first to resolve enzyme-stimulating from enzyme- inhibiting mediator activities by fractional extraction with ethanol. The ethanol- insoluble fraction from insulin-treated rat liver particles monotonically stim- ulated PDH and acetyl-CoA carboxylase activities, whereas the ethanol-soluble portion monotonically inhibited PDH, adenylate cyclase, and acetyl-CoA car- boxylase. When the mediator activities were resolved from each other in this manner, insulin was observed to elicit the generation of both the PDH-stimu- lating and -inhibiting activities in a dose-dependent manner.
Macaulay et a1.57,58 have separated PDH-stimulating activity from adenylate cyclase-inhibiting activity by gel filtration. However, the adenylate cyclase me- diator, whether extracted from rat liver or rat skeletal or heart muscle, behaved differently from those described by either Cheng et al. or Saltiel et al. First, both the PDH-stimulating and the adenylate cyclase-inhibiting activities were ethanol insoluble, in contradistinction to the ethanol-soluble adenylate cy- clase-inhibiting activity reported from liver particles by Saltiel et al.3s Second, in contrast to the material described by Cheng et al.,53 the PDH and adenylate cyclase mediators were separated from each other by gel filtration in 50 mM formic acid on Sephadex G-15 columns, with the adenylate cyclase-modulating activity eluting after the PDH m e d i a t ~ r , ~ ~ , ~ ’ but they were not separated by chromatography on Sephadex G-25 columns. The nature of these differences in solubility and chromatographic properties is not understood at the present time. The differences probably do not reflect species differences in the structures of these cyclase mediators, however, since all three groups used rat t i s s ~ e ~ ~ , ~ ~ , ~ ~ , ~ ~ for their work.
Hepatoma cells produce at least four biologically distinguishable insulin mediator activities: low-K, cAMP phosphodiesterase-stimulating, adenylate cy- clase-inhibiting, and PDH-stimulating and -inhibiting mediators. The produc- tion of the latter two mediators can be dissociated from one another by the conditions of cell culture. The promotion by insulin of the production of PDH- stimulating mediator activity from H4-11-EC3’ cells grown in standard media was not biphasic, but increased monotonically to a plateau at high insulin con- c e n t r a t i o n ~ . ~ ~ However, when the cells were first exposed to the glucocorticoid dexamethasone, they were able to produce a PDH-inhibiting mediator, in ad- dition to the PDH-stimulating activity, in response to insulin.68 H4-11-EC3’ cells did not require exposure to dexamethasone to produce the 1ow-Q cAMP phos- phodiesterase- or the adenylate cyclase-modulating activities in response to insulin. It is evident from these studies that the PDH-stimulating, PDH-inhib- iting, and adenylate cyclase-inhibiting activities produced by hepatoma cells all represent physically different substances. Since both the PDH-stimulating and the low-K, cAMP phosphodiesterase-stimulating mediator activities elute at the same position from Sephadex G-25 columns in acid buffer, more work needs to be done to determine whether they are due to the same or different sub- stances.
There are tissue differences in the structure of mediators that modulate the same enzyme in the same directions. This fact was shown by the contrasting behaviors of the H4-11-EC3’ and the rat muscle PDH-stimulating mediators in ethanol. The H4-11-EC3’ cell PDH-stimulating activity was completely soluble in 100% ethanol (Table III), in contrast to the ethanol insolubility of the PDH
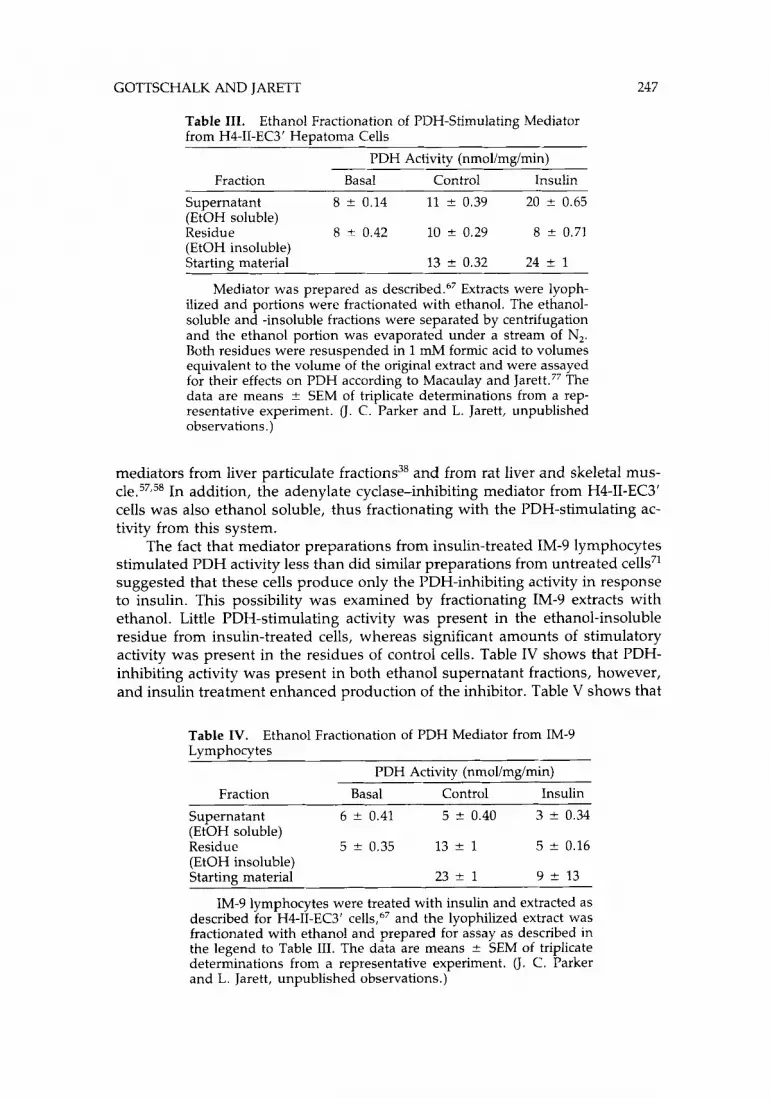
GOTTSCHALK AND JARETT 247
Table 111. from H4-11-EC3’ Heuatoma Cells
Ethanol Fractionation of PDH-Stimulating Mediator
PDH Activity (nmollmgimin) Fraction Basal Control Insulin
Supernatant 8 f 0.14 11 -+ 0.39 20 ? 0.65 (EtOH soluble) Residue 8 ? 0.42 10 f 0.29 8 f 0.71 (EtOH insoluble) Starting material 13 * 0.32 24 +- 1
~ ~~ ~
Mediator was prepared as d e ~ c r i b e d . ~ ~ Extracts were lyoph- ilized and portions were fractionated with ethanol. The ethanol- soluble and -insoluble fractions were separated by centrifugation and the ethanol portion was evaporated under a stream of N,. Both residues were resuspended in 1 mM formic acid to volumes equivalent to the volume of the original extract and were assayed for their effects on PDH according to Macaulay and Jarett.77 The data are means f SEM of triplicate determinations from a rep- resentative experiment. (J. C. Parker and L. Jarett, unpublished observations.)
mediators from liver particulate fractions38 and from rat liver and skeletal mus- c1e.57,58 In addition, the adenylate cyclase-inhibiting mediator from H4-11-EC3’ cells was also ethanol soluble, thus fractionating with the PDH-stimulating ac- tivity from this system.
The fact that mediator preparations from insulin-treated IM-9 lymphocytes stimulated PDH activity less than did similar preparations from untreated cells71 suggested that these cells produce only the PDH-inhibiting activity in response to insulin. This possibility was examined by fractionating IM-9 extracts with ethanol. Little PDH-stimulating activity was present in the ethanol-insoluble residue from insulin-treated cells, whereas significant amounts of stimulatory activity was present in the residues of control cells. Table IV shows that PDH- inhibiting activity was present in both ethanol supernatant fractions, however, and insulin treatment enhanced production of the inhibitor. Table V shows that
Table IV. Lymphocytes
Ethanol Fractionation of PDH Mediator from IM-9
PDH Activity (nmol/mg/min) Fraction Basal Control Insulin
Supernatant 6 f 0.41 5 f 0.40 3 -+ 0.34 (EtOH soluble) Residue 5 f 0.35 13 -+ 1 5 f 0.16 (EtOH insoluble) Starting material 23 f 1 9 ? 13
IM-9 lymphocytes were treated with insulin and extracted as described for H4-11-EC3‘ cells,67 and the lyophilized extract was fractionated with ethanol and prepared for assay as described in the legend to Table 111. The data are means ? SEM of triplicate determinations from a representative experiment. (J. C. Parker and L. Jarett, unpublished observations.)

248 INSULIN MEDIATORS
Table V. Mediator from IM-9 Lymphocytes
Ethanol Fractionation of Adenylate Cyclase-Inhibiting
Adenylate Cyclase Activity Starting (pmol/mg/min)
Sample Material Residue Supernatant Buffer blank 747 2 3 825 -+ 46 742 ? 18
Insulin extract 484 & 2 462 f 21 688 * 7 IM-9 lymphocytes were treated and extracted as described in
the legend to Table IV. A buffer blank was carried through the fractionation process for control purposes. The fractions were as- sayed for their effects on adenylate cyclase as described in Macau- lay and Jarett.77 The data are means ? SEM of triplicate determinations from a representative experiment. (J. C. Parker and L. Jarett, unpublished observations.)
Control extract 336 5 32 322 * 30 727 * 20
1M-9 cells also produced less of an adenylate cyclase inhibitor in response to insulin and that, in contrast to the PDH-inhibitor produced by these cells, this material was insoluble in ethanol.
These data provide evidence that, within a given tissue, a number of me- diator activities, which differ with respect to both their biological effects and, apparently, their chemical structures, were produced in response to the inter- action of insulin with its receptor. In addition, tissue differences are revealed in the structure of at least some mediator activities that regulate the same en- zyme, such as the PDH-modulating mediators from hepatoma cells and from intact rat tissue. The natures of these differences will be revealed only after the respective mediators have been purified to homogeneity and completely char- acterized. It is anticipated that these future studies will also help decide whether the mediator activities modulating acetyl-CoA carboxylase, glucose-6-phospha- tase, or phospholipid methyltransferase are represented by distinctive entities or by previously identified mediator substances.
VI. PHY SICOCHEMICAL PROPERTIES OF INSULIN MEDIATORS
The insulin mediators that have been reported to date share a number of common properties. All are water soluble and are low-molecular-weight moie- ties, as judged by gel filtration in acid b ~ f f e r s , ~ ~ , ~ , ~ ~ , ~ ~ ~ ~ r ~ ~ , ~ ~ , ~ ~ , ~ ~ resistant to boiling, and are stable at acid pH but only for short times at neutral or basic pH. They appear not to be aromatic or to contain substantial aromatic portions since they are not detectably adsorbed onto charcoal. The PDH mediator has an isoelectric point of 4-553 (J. C . Parker and L. Jarett, unpublished observa- tions); it adsorbs onto anion-exchange resins at pH 2 636,42t44,67*103 and onto cation exchangers at 5 pH 3.52,57,58
Information from several studies suggested that peptide bonds were essen- tial for the biological activity of insulin mediators. The evidence for this was twofold. First, a proteolytic process has been implicated in the generation and/ or release of mediators from the plasma The insulin-promoted generation of PDH-modulating activity from adipocyte membranes was blocked

GOTTSCHALK AND JARETT 249
by serine protease inhibitors and arginine ester protease substrate analogues (PMSF, TAME, DFB, NPGB), while trypsin, in the absence of either insulin or protease inhibitors, promoted the release of a PDH-mediator activity.33 Material with gycogen synthase-stimulating activity was also generated by trypsin treat- ment of intact adipocytes;lo4 however, in this latter study, no evidence for the generation of glycogen synthase-inhibiting activity was found, suggesting that trypsin did not faithfully reproduce all of insulin's actions at the cell surface with respect to mediator production. These results must be interpreted cau- tiously. Although the results showed that material with insulin mediatorlike activity can be produced by perturbing the structure of the cell membranes with proteolytic agents, including insulin, they do not speak to the chemical nature of the material that is released by such perturbations, specifically whether such material possesses amino acids or peptide bonds. For example, treatment of intact adipocytes with phospholipase-C mimicked insulin's actions,'05 as did trypsinlo6 treatment. Thus, a direct relationship between proteolytic events at the plasma membrane and the chemical nature of insulin mediators has not been established.
Second, various proteases have been reported to reduce the biochemical effects of several mediators. However, since none of these studies proved that the detected inactivation of mediator activity was specifically due to protease action by inclusion of all of the appropriate controls, such as treatment with inactivated protease, treatment with protease in the presence of excess alternate substrate, and so on, care must be taken to avoid overinterpretation of these experiments. The activity of the PDH-stimulating mediator was destroyed upon exposure to trypsin33,42,43,107 or ~ h y m o t r y p s i n . ~ ~ , ~ ~ Trypsin also abolished the activities of the glucose-6-phosphatase-inhibiting mediator42 and of the CAMP- lowering mediator prepared by Zhang et al.45 However, neither trypsin nor chymotrypsin had any measurable effect on PDH-inhibiting activity extracted from liver membranes.lo7
The activities of PDH-stimulating mediators from intact rat skeletal muscle, heart, or liver were reduced only about 40% by specific pro tease^.^^ Trypsin was without effect on any of these mediator activities. Chymotrypsin and sub- tilisin were the most effective proteases tested. Heat inactivation and incubation at 4°C prevented protease-mediated inactivation of the mediators. Although a number of known protease inhibitors failed to prevent inactivation, TLCK was effective. Since only partially purified preparations of the mediators were used in these studies, the failure to detect more complete inactivation could have been due to the presence of contaminating materials in the extracts, which were resistant to protease digestion, and therefore still stimulated PDH activity.
Amino acids have been detected in the acid hydrolyzates of partially pur- ified preparations of three mediators. Turakulov and Gainutdinov6' reported 43 amino acids after hydrolysis of an insulin-dependent mitochondria1 Ca2+ trans- port regulator. Twelve amino acids were detected after hydrolysis of a partially purified CAMP-dependent protein kinase-inhibiting mediator, and 14 amino acids were identified in the hydrolyzates of a partially purified preparation of the PDH-stimulating mediator.53 However, the biological activities of the prep- arations after hydrolysis were not reported in these studies, and it is not known if the mediators were, in fact, destroyed by these procedures. Also, although

250 INSULIN MEDIATORS
both groups claim to have purified the mediators to near homogeneity, no evidence supporting this claim was presented. For these reasons, the signifi- cance of the reported amino acids to the biological activities of these mediators awaits positive identification of the mediator substances.
Larner has suggested that, if proteases generate mediators from the external face of the cell surface, the mediators would most likely be g1y~opeptides.l~~ Evidence has been adduced in support of this proposal. The activities of both the PDH-stimulating and -inhibiting mediators from rat liver membranes were destroyed by exposure to either neuraminidase or P-galactosidase. lo8,109 Alter- nate substrates protected the mediators from inactivation in these experi- ments. 108,109 These results suggested that the PDH-modulating mediators possess carbohydrate residues that are essential for their biological activities. However, Jarett's laboratory was unable to confirm these findings using PDH-stimulating mediator from intact rat liver or skeletal muscle (Macaulay and L. Jarett, un- published results). This lack of inactivation could have been due to contami- nants in the mediator preparation that interfered with the digestion.
Phospholipids and their derivatives have also been suggested to either be part of or to constitute insulin mediators. Wasner70 claims that the cAMP an- tagonist substance he has extracted from hepatocytes contained prostaglandin El , phosphate, and inositol. It is important to recall (c.f. section III.B.3) that this substance is probably chemically distinct from the other adenylate cy- clase-inhibiting mediators discussed in this review. Begum et aI.lo9 have sug- gested that the PDH-modulating activity may be a prostaglandin derivative, since treatment of rats with indomethacin blocked the production of the PDH- stimulating mediator, and prostaglandin E2 overcame this block. In addition, they found that treatment of intact adipocytes with dexamethasone prevented PDH-stimulating mediator release from subsequently isolated plasma mem- b r a n e ~ . * ~ ~ Incubation of the adipocytes with arachidonic acid as well as dexa- methasone partially prevented the dexamethasone effect. lo9 Prostaglandin E2, however, did not overcome the ability of dexamethasone to prevent the release of PDH-stimulating mediators. The multiplicity of effects of indomethacin, dex- amethasone, prostaglandins, and arachidonic acids make it difficult to conclude that these insulin mediators are, in fact, prostaglandins or derivatives thereof.
Jarett's laboratory has shown that various phospholipids have effects on PDH and low-K, cAMP phosphodiesterase similar to those of several insulin mediators.'01,1*0,120 Farese et al. 111-114 have shown that insulin treatment of adi- pose tissue changed the concentrations of these same phospholipids. The dif- ferences in water solubility of the phospholipids and insulin mediators make it unlikely that the mediators are indeed phospholipid or phospholipid-like. How- ever, this does not rule out the possibility that phospholipids may be involved in regulating events related to insulin action.
In summary, circumstantial evidence suggests that mediators of insulin action may possess a variety of chemical entities, including peptides and car- bohydrates, which are essential for their biological activities. There is, however, no definitive proof that mediators are, for instance, peptides or glycopeptides, and no such proof will be possible until mediator activities have been purified to homogeneity and their complete chemical characterization accomplished.

GOTTSCHALK AND JARETT 25 1
VII. PHYSIOLOGIC AND PATHOPHYSIOLOGIC ROLES OF INSULIN MEDIATORS
The results summarized above clearly demonstrate that insulin mediators subserve physiologically relevant roles in regulating metabolism. Thus, the me- diators are generated from a variety of sources, including plasma membranes, cells, and intact tissues; their generationirelease is enhanced by insulin; they modulate the activities of a number of insulin-sensitive enzymes and metabolic systems in the same direction as insulin treatment; and they can affect both individual enzymes and metabolic systems in intact cells. Insulin mediators cannot mimic all of insulin's short-term effects on intact cells, however, since they have no detectable effects on glucose transport or oxidation, and their ability to mimic insulin's long-term effects has not yet been examined. The work summarized in this section provides additional evidence for the physiologic importance of these substances.
Diet clearly affects the generation or action of insulin mediators. Mem- branes from either adipocytes or liver taken from fat-fed rats released less PDH- stimulating activity, either spontaneously in Tris bufft!rs or in response to in- sulin, Con A, or trypsin, than membranes from carbohydrate-fed or control (chow-fed) rats. 108,115,116 Liver membranes from fat-fed rats also released less PDH-inhibiting activity.lo7 It is not known whether these results reflect reduced insulin binding to the membranes, which is known to occur as a result of high- fat diets, altered properties of the mediator or of its generationhelease mecha- nism, or all three.
Fasting, treatment with pharmacologic concentrations of glucocorticoids, and certain forms of diabetes all elicit insulin resistance. All have also been associated with the reduced generation of insulin mediators. Amatruda and Chang117 and Trowbridge et al."' showed that less PDH-stimulating mediator activity was released from the particulate fractions of livers from fasted rats than from control livers. Refeeding restored mediator release to control level^."^ Macaulay et al.57 also found that the recoveries of PDH- and adenylate cy- clase-modulating activities from heart muscle of insulin-treated rats that had been starved for 72 h were significantly reduced compared with the yields from control (24-h starved) rats.
The release of both PDH-stimulating and -inhibiting activities from mem- branes was attenuated by prior treatment of the donor animals with dexameth- a ~ o n e . ' ~ ' , ~ ~ ~ This result is consistent with this hormone's ability to induce insulin resistance in vivo. However, it is in sharp contrast with dexamethasone's effects on cultured hematoma cells. In that system, exposure to the hormone enables the cells to generate PDH-inhibiting mediator in response to insulin, without diminishing their abilities to generate the PDH-stimulating mediator. The rea- son(s) for these discrepant results may not be resolved until the mechanisms of mediator generationiaction have been elucidated.
Amatruda and Chang117 have shown that streptozocin-induced diabetes in rats reduced the amounts of PDH-stimulating mediator activity that was re- leased from liver membranes. Treatment of the diabetic rats with therapeutic concentrations of insulin restored mediator production to control levels.

252 INSULIN MEDIATORS
The observation that the production or action of insulin mediators is af- fected by the physiologic or the pathologic state of the source of mediators, in directions that are consistent with the responsiveness of the sources to insulin, argues forcibly that these substances play important roles in mediating insulin’s effects and in regulating metabolism.
VIII. CONCLUSION
The results summarized in this review support the conclusion that the interaction of insulin with its receptor on the plasma membrane results in the generation of a family of low-molecular-weight moieties that mediate many, but not all, of the short-term actions of insulin. These substances are produced in appropriate, insulin-sensitive subcellular systems, as well as in intact cells and tissue. They affect the activities of a number of insulin-sensitive enzymes and metabolic systems, both in cell homogenates and in intact cells. The quantities of the mediators produced are dependent on the physiologic and pathologic state of the organism, and are reduced under conditions in which the sensitivity of other metabolic parameters to insulin are also reduced. At present, the chem- ical natures of these substances are not known, but, in light of the currently available information, are likely to be novel, although they may contain amino acids.
The studies relating insulin resistance to the reduced production of insulin mediators suggests that the mediators have potential clinical importance, both for increased understanding of the nature of the postreceptor defects involved in cases of insulin resistance, particularly type I1 diabetes and obesity, and for their possible therapeutic value in such cases.
Future directions are clear. The active principles responsible for mediator action must be purified to homogeneity and identified; the mechanisms of their generation must be established; their spectrum of action, short-term effects, long-term effects, etc., must be precisely defined; and the molecular basis for their effects must be determined.
Acknowledgment The authors express their appreciation to S. L. Macaulay, J . 0. Macaulay, J. A.
Smith, E. H. A. Wong, J. C. Parker, and K. L. Kelly for their contributions to the work reported in this review and for their criticism of the review.
This work has been supported by NIH grants AM28144 and AM07409 and a grant from the Juvenile Diabetes Foundation.
References 1. Cori CF: Enzymatic reactions in carbohydrate metabolism. Harvey Led 41:253-272,
2. Stadie WC: Current concepts of the action of insulin. Physiol Rev 34:52-100, 1954. 3. Stadie WC, Haugaard N, and Vaugham M: Studies of insulin binding with isotop-
ically labeled insulin. J B i d Chem 199:792-795, 1952. 4. Levine R, Goldstein MS, Huddleston B, and Klein S: Action of insulin on the
”permeability” of cells to free hexoses as studied by its effect on the distribution of galactose. Am J Physiol 163:70-76, 1950.
1945.

GOTTSCHALK AND JARETT 253
5. Levine R, Goldstein MS, Klein S, and Huddleston B: The action of insulin on the distribution of galactose in eviscerated nephrectomized dogs. J Biol Chem 179985-986, 1949.
6. Goldstein MS, Henry WL, Huddleston B, and Levine R: Action of muscle work on transfer of sugars across cell barriers: comparison with action of insulin. A m f Physiol 173212-220, 1953.
7. Villar-Palasi C and Larner J: Insulin treatment and increased UDPG-glycogen trans- glucosylase activity in muscle. Arch Biochem Biophys 94:436442, 1961.
8. Freychet P, Roth J, and Neville DM, Jr: Insulin receptors in the liver: specific bind- ing of [125-1] insulin into the plasma membrane and its relation to insulin bioactivity. Proc Natl Acad Sci U S A 68:1833-1837, 1971.
9. Freychet P, Roth J, and Neville DM, Jr: Monoidoinsulin: demonstration of its bio- logical activity and binding to fat cells and liver membranes. Biochem Biophys Res Commun 43:400408, 1971.
10. Larner J: Insulin and glycogen synthase. Diabetes 22 (Suppl2):428438, 1972. 11. Turakulov YK, Gainutdinov MK, Lavina 11, and Akhmatov MS: Insulin dependent
cytoplasmic regulator of Ca+ + ion transport in the mitochondria of the liver. Proc Acad Sci USSR 234~1471-1473, 1977.
12. Seals JR and Jarett L: Activation of pyruvate dehydrogenase by direct addition of insulin to an isoltaed plasma membrane/mitochondria mixture: evidence for gen- eration of insulin's second messenger in a subcellular system. Proc Natl Acad Sci U S A 77:77-81, 1980.
13. Villar-Palasi C and Larner J: Insulin-mediated effect on the activity of UDPG-gly- cogen transglucosylase of muscle. Biochim Biophys Acta 39:171-173, 1960.
14. Larner J, Villar-Palasi C, and Richman T: Insulin-stimulated glycogen formation in rat diaphragm. Levels of tissue intermediates in short-time experiments. Arch Biochem Biophys 86:56-60, 1980.
15. Walkenbach RJ, Hazen R, and Larner J: Reversible inhibition of cyclic AMP-de- pendent protein kinase by insulin. Mol Cell Biochem 19:31-41, 1978.
16. Walkenbach RJ, Hazen R, and Larner J: Hormonal regulation of glycogen synthase. Insulin decreases protein kinase sensitivity to cyclic AMP. Biochem Biophys Acta 629:421-430, 1980.
17. Larner J, Takeda Y, Brewer HB, Brooker G, and Murad F: Studies on glycogen synthase and its control by hormones, in Metabolic Interconversion of Enzymes. Shatiel S , ed. Springer-Verlag, New York, pp 7, 1976.
18. Murad F: Techniques for the formation, partial purification, and assay of a cyclic AMP inhibitor, in Methods in Enzymology, Vol. 38, Part C. Hardman JG and OMally BW, eds. Academic Press, New York, pp 273-283, 1974.
19. Walsh DA, Ashby CD, Gonzalez C, Calkins D, Fischer EH, and Krebs EG: Purifi- cation and characterization of an inhibitor protein of cyclic AMP-dependent protein kinases. J Biol Chem 246:1977-1985, 1971.
20. Larner J, Galasko G, Cheng K, DePaoli-Roach AA, Huang L, Daggy P, and Kellog J: Generation by insulin of a chemical mediator that controls protein phosphoryl- ation and dephosphorylation. Science 206:140%1410, 1979.
21. Jarett L and Smith RM: The stimulation of adipocyte plasma membrane magnesium ion-stimulated adenosine triphosphatase by insulin and Concanavalin A. J B i d Chem 249:5195-5199, 1974.
22. Krebs EG and Beavo JA: Phosphorylation-dephosphorylation of enzymes. Ann Rev Biochem 48:923-959, 1979.
23. Seals JR, McDonald JM, and Jarett L: Direct effects of insulin on the labeling of isolated plasma membrane by [Y-~~PIATP. Biochem Biophys Res Commun 831365-1372, 1978.
24. Seals JR, McDonald JM, and Jarett L: Insulin effects on protein phosphorylation of plasma membranes and mitochondria in a subcellular system from rat adipocytes. I. Identification of insulin-sensitive phosphoproteins. f Biol Chem 254:6991-6996, 1979a.

254 INSULIN MEDIATORS
25. Seals JR, McDonald JM, and Jarett L: Insulin effects on protein phosphorylation of plasma membranes and mitochondria in a subcellular system from rat adipocytes. 11. Characterization of insulin-sensitive phosphoproteins and conditions for obser- vation of the insulin effect. J Biol Chem 254:6997-7001, 1979b.
26. Akers RF and Routtenberg A: Brain protein phosphorylation in vitro: selective substrate action of insulin. Life Sci 35:809--813, 1984.
27. Hughes WA, Brownsey RW, and Denton RM: Studies on the incorporation of ["-PI phosphate into pyruvate dehydrogenase in intact rat fat-cells: Effect of insulin. Biochem f 192:469481, 1980.
28. Denton RM and Hughes WA: Pyruvate dehydrogenase and the hormonal regula- tion of fat synthesis in mammalian tissues. Int J Biochem 9:454-552, 1978.
29. Denton RM, Randle PJ, Bridges BJ, Cooper RH, Kerbey AL, Pask HT, Severson DL, Stansbie D, and Whitehouse S: Regulation of mammalian pyruvate dehydro- genase. Ma1 Cell Biochem 9:27-53, 1975.
30. Fuller SJ and Randle PJ: Reversible phosphorylation of pyruvate dehydrogenase in rat skeletal-muscle mitochondria: effects of starvation and diabetes. Biochem J 219:635-646, 1984.
31. Linn TC, Pettit FH, and Reed L: a-keto dehydrogenase complexes. X. Regulation of the activity of the pyruvate dehydrogenase complexes from beef kidney mito- chondria by phosphorylation and dephosphorylation. Proc Nut1 Acad Sci U S A 62:234-241, 1969.
32. Loten EG, Assimacopoulos-Jeannett F, Exton JH, and Park CR: Stimulation of a low Km phosphodiesterase from liver by insulin and glucagon. J Biol Chem 253:746-757, 1978.
33. Seals JR and Czech MP: Evidence that insulin activates an intrinsic plasma mem- brane protease in generating a secondary chemical mediator. J Biol Chem 255:6529-6531, 1980.
34. Kiechle FL, Jarett L, Kotagal N, and Popp DA: Partial purification from rat adipocyte plasma membranes of a chemical mediator which stimulates the action of insulin on pyruvate dehydrogenase. J Biol Chem 256:2945-2951, 1981.
3.5. Jarett L and Seals JR: Pyruvate dehydrogenase activation in adipocyte mitochondria by an insulin-generated mediator from muscle. Science 206:1407-1408, 1979.
36. Seals JR and Czech MI': Characterization of a pyruvate dehydrogenase activator released by adipocyte plasma membrane in response to insulin. J Biol Chem 256:2894-2899, 1981.
37. Jarett L and Kiechle FL: Generation of a putative second messenger for insulin action from adipocyte and liver plasma membranes. 1st International Symposium on Insulin Receptors, Rome, 1980.
38. Saltiel AR, Siegel M, Jacobs S, and Cuatrecasas P: Putative mediators of insulin action: regulation of pyruvate dehydrogenase and adenylate cyclase activities. Proc Natl Acnd Sci U S A 79:3513-3517, 1982.
39. Saltiel AR, Jacobs S, Siegel M, and Cuatrecasas P: Insulin stimulates the release from liver plasma membranes of a chemical modulator of pyruvate dehydrogenase. Biochem Biophys Res Cornmuiz 102:1041-1047, 1981.
40. Saltiel A R Preparation and characterization of putative insulin mediators from liver, in Methods in Diabetes Research, Vol. 1. Laboratory Methods, Part B. Larner J and Pohl S, eds. John Wiley and Sons, Inc., New York, pp 73-79, 1984.
41. Saltiel AR, Doble A, Jacobs S, and Cuatrecasas P: Putative mediators of insulin action regulate hepatic acetyl-CoA carboxylase activity. Biochern Biophys Res Commun
42. Suzuki S, Toyota T, Suzuki H, and Goto Y: A putative second messenger of insulin action regulates hepatic microsomal glucose-6-phosphatase. Biochem Biophys Res Commun 118:40-46, 1984.
43. Sakamoto Y, Kuzuya T, and Sat0 J: Demonstration of a pyruvate dehydrogenase activator in insulin-treated human placental plasma membrane. Biomed Res 3599-605, 1982.
110:789-795, 1983.

GOTTSCHALK AND JARETT 255
44. Suzuki S, Toyota T, Suzuki H, and Goto Y: Partial purification from human mono- nuclear cells and placental plasma membranes of insulin mediator which stimulates pyruvate dehydrogenase and suppresses glucose-6-phosphatase. Arch Biochem Bio- phys 235:418426, 1984.
45. Zhang S-R, Shi Q-H, and Ho R: Cyclic AMP-lowering mediator of insulin action. J Biol Chem 258:6471-6476, 1983.
46. Car0 JF, Folli F, Cecchin F, and Sinha MK: Chemical mediator of insulin action stimulates lipid synthesis and down regulates the insulin receptors in primary cultures of rat hepatocytes. Bioclzem Biophys Res Commun 115:375-382, 1983.
47. Kiechle FL, Jarett L, Kotagal N, and Popp DA: Isolation from rat adipocytes of a chemical mediator for insulin activation of pyruvate dehydrogenase. Diabetes
48. Seals JR and Czech MP: Extraction of an insulin-dependent enzyme regulator from intact adipocytes. Fed Proc 41:1081, 1982.
49. Kiechle FL and Jarett L: The effect of an insulin-sensitive chemical mediator from rat adipocytes on low Km and high Km cyclic AMP phosphodiesterase. FEBS Lett 133:279-282, 1981.
50. Loten EG and Sneyd JGT: An effect of insulin on adipose-tissue adenosine 3':5'- cyclic monophosphate phosphodiesterase. Biochem J 120:187-193, 1970.
51. Czech MP and Seals J R Production by plasma membranes of a chemical mediator of insulin action. Fed Proc 41:2730-2735, 1982.
52. Thompson MP, Larner J, and Kilpatrick DL: Purification and partial characterization of a putative mediator of insulin action on cyclic AMP-dependent protein kinase. Mol Cell Biochem 62:67-75, 1984.
53. Cheng K, Thompson M, Schwartz C, Malchoff C, Tamura S, Craig J, Locher E, and Larner J: Multiple intracellular peptide mediators of insulin action, in Molecular Basis of lnsulin Action. Czech M, ed. Plenum Press, New York, 1985.
54. Parker JC and Jarett L: Insulin mediator stimulates pyruvate dehydrogenase of intact liver mitochondria. Diabetes 34:92-97, 1985.
55. Jarett L, Wong EHA, Smith JA, and Macaulay SL: Stimulation of pyruvate dehy- drogenase activity in. intact rat adipocytes by insulin mediator from rat skeletal muscle. Endocrinology 116:1011-1016, 1985.
56. Jarett L, Wong EHA, Macaulay SL, and Smith JA: Insulin mediators from rat skeletal muscle have differential effects on insulin-sensitive pathways of intact adipocytes. Science 227533435, 1985.
57. Macaulay SL, Macaulay JO, and Jarett L: Insulin stimulates generation of intracel- Mar mediators in rat heart. In press, Arch Biochem Biophys, 1985.
58. Macaulay SL, Macaulay JO, Kelly KL, and Jarett L: Characteristics and purification of putative insulin mediators from rat skeletal muscle and liver. Submitted for publication. 1985.
59. Turakulov YK and Gainutdinov MK: Insulin-dependent cytoplasmic regulator. Separation, identification, and investigation of insulin-like action, in Frontiers of Bioorganic Chemistry and Molecular Biology. Ananchenko SN, ed. Pergamon Press, Oxford, pp 115-121, 1978.
60. Linn TC, Pettit FH, Hucho F, and Reed L: a-keto acid dehydrogenase complexes. XI. Comparative properties of the pyruvate dehydrogenase complexes from kidney, heart, and liver mitochondria. Proc Nut1 Acad Sci U S A 64:227-234, 1969.
61. Turakulov YK, Gainutdinov MK, Lavina 11, and Akhmatov MS: Regulation of ox- idative phosphorylation by adrenalin and insulin: role of the insulin-dependent cytoplasmic regulator. Bull Exp Biol Med 88:301-304, 1979.
62. Gainutdinov MK, Abdullaev NK, Gizatullina ZZ, Akhmatov MS, and Turakulov Y K Hypoglycemic action of the insulin-dependent cytoplasmic factor. Proc Acad Sci USSR 243:1589-1592, 1978.
63. Purrello F, Vigneri R, Clawson GA, and Goldfine I: Insulin stimulation of nucleo- side triphosphatase activity in isolated nuclear envelopes. Science 216:1005-1007, 1982.
291852455, 1980.

256 INSULIN MEDIATORS
64. Purrello F, Burnshaw DB, and Goldfine I: Insulin receptor anti-serum and plant lectins mimic the direct effects of insulin on nuclear envelopes phosphorylation. Science 221:462-464, 1983.
65. Purrello F, Burnshaw DB, and Goldfine I: Insulin regulation of protein phospho- rylation in isolated rat liver nuclear envelopes: potential relationship to mRNA metabolism. Proc Nutl Acad Sci U S A 80:1193-1198, 1983.
66. Horvat A: Stimulation of RNA synthesis in isolated nuclei by an insulin-induced factor in liver. Nature 286:906-908, 1980.
67. Parker JC, Kiechle FL, and Jarett L: Partial purification from hepatoma cells of an intracellular substance which mediates the effects of insulin on pyruvate dehydro- genase and low Km cyclic AMP phosphodiesterase. Arch Biochem Biophys 215:339-344, 1982.
68. Esau B and Koontz J: Glucocorticoids enable H-35 hepatoma cells to produce an inhibitory mediator in response to insulin. Diabetes 33 (Suppl 1):141a, 1984.
69. Wasner HK Biosynthesis of cyclic AMP antagonist in hepatocytes from rats after adrenalin- or insulin-stimulation. F E B S Lett 133:260-264, 1981.
70. Wasner HK: Prostaglandyl-inositol cyclic phosphate, a second messenger of insulin action. 2nd International Symposium on Insulin Receptors, Rome, 1983.
71. Jarett L, Kiechle FL, and Parker JC: Chemical mediator or mediators of insulin action: response to insulin and mode of action. Fed Proc 41:2736-2741, 1982.
72. Reed LJ: Regulation of mammalian pyruvate dehydrogenase complex by a phos- phorylation-dephosphorylation cycle, in Current Topics in Cellular Regulation, Vol. 18. Estabrook RW and Srere P, eds. Academic Press, New York, pp 95-106, 1981.
73. Wieland OH: The mammalian pyruvate dehydrogenase complex: structure and regulation. Rev Physiol Biochem Pharmacol 96:123-170, 1983.
74. Popp DA, Kiechle FL, Kotagal N, and Jarett L: Insulin stimulation of pyruvate dehydrogenase in an isolated plasma membrane-mitochondria1 mixture occurs by activation of pyruvate dehydrogenase phosphatase. J Biol Chem 255:7540-7543, 1980.
75. Hucho F, Randall DD, Roche TE, Burgett MW, Pelley JW, and Reed LJ: a-keto acid dehydrogenase complexes. XVII. Kinetic and regulatory properties of pyruvate de- hydrogenase kinase and pyruvate dehydrogenase phosphatase from bovine kidney and heart. Arch Biochem Biophys 151:328-340, 1972.
76. Whitehouse S, Cooper RH, and Randle PJ: Mechanism of activation of pyruvate dehydrogenase by dichloroacetate and other halogenated carboxylic acids. Biochem J 141:761-774, 1974.
77. Macaulay SL and Jarett L: Insulin mediator causes dephosphorylation of the a- subunit of pyruvate dehydrogenase by stimulating phosphatase activity. Arch Biochm Biophys 237142-150, 1985.
78. Newman J, Armstrong JM, and Bornstein J: Assay of insulin mediator by activation of pyruvate dehydrogenase phosphatase. 2nd Annual Symposium on Insulin Re- ceptors, Rome, 1983.
79. Speth M and Schulze H-U: Hormone-induced effects on the rat liver microsomal glucose-6-phosphate system in vitro. Biochem Biopkys Res Commun 99:134-141, 1981.
80. Begley PJ and Craft JA: Evidence for protein phosphorylation as a regulatory mech- anism for heptic microsomal glucose-6-phosphatase. Biochem Biophys Res Commun
81. Burchell A and Burchell B: Stabilization of partially purified glucose-6-phosphatase by fluoride. Is enzyme inactivation caused by dephosphorylation? FEBS Lett
82. Hirata F and Axelrod J: Phospholipid methylation and biological signal transduc-
83. Mato JM and Alemany S: What is the function of phospholipid N-methylation?
103:1029-1034, 1981.
118:180-184, 1980.
tion. Science 209:1082-1090, 1980.
Biochem J 213:l-10, 1983.

GOTTSCHALK AND JARETT 257
84. Kelly KL, Wong EHA, and Jarett L: Adrenocorticotropic stimulation and insulin inhibition of adipocyte phospholipid methylation. J Biol Chem 260:3640-3644, 1985.
85. Nieto A and Catt KJ: Hormonal activation of phospholipid methyltransferase in the Leydig cell. Endocrinology 113:758-762, 1983.
86. Castano JG, Alemany S, Nieto A, and Mato JM: Activation of phospholipid meth- yltransferase by glucagon in rat hepatocytes. J Biol Chem 255:9041-9043, 1980.
87. Varela I, Merida I, Pajares M, Villalba M, and Mato JM: Activation of partially purified rat liver lipid methyltransferase by phosphorylation. Biochem Biophys Res Commun 122:1065-1070, 1984.
88. Witters LA, Lowaloff EM, and Avruch J: Glucagon regulation of protein phospho- rylation. Identification of acetyl CoA carboxylase as a substrate. J Biol Chem 254:245-248, 1979.
89. Witters LA: Insulin stimulates the phosphorylation of acetyl-CoA carboxylase. Biochem Biophys Res Commun 100:872-878, 1981.
90. Brownsey RW and Denton RM: Evidence that insulin activates fat-cell acetyl-CoA carboxylase by increased phosphorylation at a specific site. Biochem J 202:77-86, 1982.
91. Brownsey RW, Edge11 NJ, Hopkirk TJ, and Denton RM: Studies on insulin-stim- ulated phosphorylation of acetyl-CoA carboxylase, ATP citrate lyase and other pro- teins in rat epididymal adipose tissue: evidence for activation of a cyclic AMP- independent protein kinase. Biochem J 218:773-743, 1983.
92. Chan CM and McDonald TM: Identification of an insulin-sensitive calcium-stim- ulated phosphoprotein in ;at adipocyte plasma membranes. J Biol 257:7443-7448, 1982.
93. Pershadsingh HA, Landt M, and McDonald JM: Calmodulin-sensitive ATP-de- pendent Ca++ transport across adipocyte plasma membranes. Biol Chem
94. Pershadsingh HA and McDonald JM: A high affinity calcium-stimulated magne- sium-dependent adenosine triphosphatase in rat adipocyte plasma membranes. J Biol Chem 255:4087-4093, 1980.
95. Pershadsingh HA and McDonald JM: Direct addition of insulin inhibits a high affinity Ca + + -ATPase in isolated adipocyte plasma membrane. Nature 281:495497, 1979.
96. McDonald JM, Pershadsingh HA, Kiechle FL, and Jarett L: Parallel stimulation in adipocytes of the plasma membrane Ca+ + transport/(Ca+ + +Mg+ +)-ATPase system and mitochondria1 pyruvate dehydrogenase by a supernatant factor de- rived from isolated plasma membranes. Biochem Biophys Res Commun 100.857-864, 1981.
97. Kono T and Barham FW: Effects of insulin on the levels of adenosine 3':5'-mono- phosphate and lipolysis in isolated rat epididymal fat cells. f Bid Chem 248:7417-7426, 1973.
98. Exton JH, Harper SC, Tucker AL, and Ho R: Effects of insulin on gluconeogenesis and cyclic AMP levels in perfused livers from diabetic rats. Biochim Biophys Actu 329:2340, 1973.
99. Illiano T and Cuatrecasas P: Modulation of adenylate cyclase activity in liver and fat cell membranes by insulin. Science 175:906-908, 1972.
100. Torres HN, Flawia MM, Hernandez L, and Cuatrecasas P: Effects of insulin on the adenyl cyclase activity of isolated fat cell membranes. f Membr Biol 43:l-18, 1978.
101. Macaulay SL, Kiechle FL, and Jarett L: Phospholipid modulation of low Km cyclic AMP phosphodiesterase activity on rat adipocyte microsomes. Arch Biochem Biophys 225:130-136, 1983a.
102. Cheng K, Galasko G, Huang L, Kellogg J, and Larner J: Studies on the insulin mediator. 11. Separation of two antagonistic biologically active materials from frac- tion 11. Diabetes 29:659-661, 1980.
255~8983-8986, 1980.

258 INSULIN MEDIATORS
103. Cheng K, Thompson M, and Larner J: Isolation of insulin mediators from rat skeletal muscle, in Methods in Diabetes Research, Vol. 1. Laboratory Methods, Part B. Larner J and Pohl S, eds. John Wiley & Sons, Inc., New York, pp 81-89, 1984.
104. Larner J, Cheng K, Schwartz C, Kikuchi K, Tamura S, Creacy S, Bubler R, Galasko G, Pullin C, and Katz M: A proteolytic mechanism for the action of insulin via oligopeptide mediator formation. Fed Proc 41:2724-2729, 1982.
105. Rodbell M; Metabolism of isolated fat cells. 11. The similar effects of phospholipase C (Clostridium perfringens a-toxin) and of insulin on glucose and amino acid me- tabolism. Biol Chem 241:130-139, 1966.
106. Kikuchi K, Schwartz C, Creacy S, and Larner J; Independent control of selected insulin-sensitive cell membrane and intracellular functions-the linkage of cell membrane and intracellular events controlled by insulin. 111. The influence of tryp- sin on cell membrane hexose transport and on glycogen synthase. Mol Cell Biockern 37:125-130, 1981.
107. Begum N, Tepperman HM, and Tepperman J: Decreased insulin-generation of py- ruvate dehydrogenase inhibitor in insulin resistant states. Biochem Biopkys Res Corn- mun 117:456463, 1983a.
108. Begum, N, Tepperman HM, and Tepperman J: Effects of high fat and high carbo- hydrate diets on liver pyruvate dehydrogenase and its activation by a chemical mediator released from insulin-treated liver particulate fraction: effect of neuramin- idase treatment on the chemical mediator activity. Endocrinology 11250-59, 198313.
109. Begum N, Tepperman HM, and Tepperman J: Studies on the possible involvement of prostaglandins in insulin generation of pyruvate dehydrogenase activator. Biockern Biopkys Res Commun 34:29-37, 1985.
110. Kiechle FL and Jarett L: Phospholipids and the regulation of pyruvate dehydro- genase from rat adipocyte mitochondria. Mol Cell Endocrinol 55:99-105, 1983.
111. Farese RV, Barnes DE, Davis JS, Standaert ML, and Pollet RJ: Effects of insulin and protein synthesis inhibitors on phospholipid metabolism, diacylglycerol levels, and pyruvate dehydrogenase activity in BC3H-1 cultured myocytes. Biol Chem
112. Farese RV, Farese RV, Jr, Sabir MA, Larson RE, and Trudeau WL: The mechanism of action of insulin on phospholipid metabolism in rat adipose tissue. Requirement for protein synthesis and a carbohydrate source, and relationship to activation of pyruvate dehydrogenase. Diabetes 33:648-655, 1984.
113. Farese RV, Larson RE, and Sabir MA: Insulin acutely increases phospholipids in the phosphatidate-inositide cycle in rat adipose tissue. Biol Chem 257:4042-4045, 1982.
114. Farese RV, Sabir MA, Larson RE, and Trudeau WL: Insulin treatment acutely in- creases the concentration of phosphatidylserine in rat adipose tissue. Biockim Bio- phys Actu 750:200-202, 1983.
115. Begum N, Tepperman HM, and Tepperman J: Effects of high fat and high carbo- hydrate diets on adipose tissue pyruvate dehydrogenase and its activation by a plasma membrane-enriched fraction and insulin. Endocrinology 110:1914-1921, 1982.
116. Begum N, Tepperman HM, and Tepperman J: Effects of high carbohydrate and high fat diets on rat adipose tissue pyruvate dehydrogenase response to concana- valin A and spermine. Endocrinology 111:1491-1497, 1982.
117. Amatruda JM and Chang CL: Insulin resistance in the liver in fasting and diabetes mellitus: the failure of insulin to stimulate the release of a chemical modulator of pyruvate dehydrogenase. Biockem Biopkys Res Commun 112:3541, 1983.
118. Trowbridge M, Sussman A, Ferguson L, Draznin B, Neufeld N, Begum N , Tep- perman HM, and Tepperman J: Mechanisms of the fasting-induced dissociation of insulin binding from its action in isolated rat hepatocytes. Mol Cell Biockern 6225-36, 1984.
119. Begum N, Tepperman HM, and Tepperman J: Effect of dexamethasone on adipose tissue and liver pyruvate dehydrogenase and its stimulation by insulin generated chemical mediator. Endocrinology 114:99-107, 1984.
25917094-7100, 1984.

GOTTSCHALK AND JARETT 259
120. Macaulay SL, Kiechle FL, and Jarett L: Comparison of phospholipid effects on insulin-sensitive low-Km cyclic AMP phosphodiesterase in adipocyte plasma membranes and microsomes. Biochirn Biophys Actu 760:29%299, 1983b.
121. Makino H and Kono T: Characterization of insulin-sensitive phosphodiesterase in fat cells. 11. Comparison of enzyme activities stimulated by insulin and by isopro- terenol. J Bid Chem 255:785&7854, 1980.





























