Embed Size (px)
Citation preview

FEBS Letters 584 (2010) 4151–4156
journal homepage: www.FEBSLetters .org
Importin b3 mediates the nuclear import of human ribosomal protein L7 throughits interaction with the multifaceted basic clusters of L7
Chang-Wei Chou a, Lin-Ru Tai a, Ralph Kirby a, I.-Fang Lee a, Alan Lin a,b,⇑a Institute of Genome Sciences, School of Life Sciences, National Yang-Ming University, Shih-Pai, Taipei 112, Taiwan, ROCb Institute of Clinical Dentistry Science, School of Dentistry, National Yang-Ming University, Shih-Pai, Taipei 112, Taiwan, ROC
a r t i c l e i n f o
Article history:Received 17 May 2010Revised 29 July 2010Accepted 26 August 2010Available online 7 September 2010
Edited by Ulrike Kutay
Keywords:Eukaryotic ribosomal protein L7Importin b3Nuclear localization signal (NLS)BiosensorOverlay assayFluorescence recovery after photobleaching(FRAP) assay
0014-5793/$36.00 � 2010 Federation of European Biodoi:10.1016/j.febslet.2010.08.044
⇑ Corresponding author at: Institute of Genome SUniversity, Shih-Pai, Taipei 112, Taiwan, ROC. Fax: +8
E-mail address: [email protected] (A. Lin).
a b s t r a c t
We show that importin b3 is essential for the nuclear import of L7. The import is mediated via themultifaceted basic amino acid clusters present in the NH2-region of L7, and is RanGTP-dependent.Using a (EGFP)3 reporter system and a FRAP assay, the role the individual clusters play as a func-tional NLS has been characterized, and each cluster was found to exhibit a different rate of real timenuclear uptake. We assume that having such a multiple NLS may provide L7 with preferentialnuclear uptake.
Structured summary:MINT-7992735: Importin beta-3 (uniprotkb:O00410) binds (MI:0407) to L7 (uniprotkb:P18124) by bio-physical (MI:0013)MINT-7992687: L7 (uniprotkb:P18124) binds (MI:0407) to Importin beta-3 (uniprotkb:O00410) by filterbinding (MI:0049)MINT-7992699: L7 (uniprotkb:P18124) physically interacts (MI:0915) with Importin beta-3 (uni-protkb:O00410) by affinity chromatography technology (MI:0004)MINT-7992718: L7 (uniprotkb:P18124) physically interacts (MI:0915) with RAN (uniprotkb:P62826) bycompetition binding (MI:0405)MINT-7992671: L7 (uniprotkb:P18124) physically interacts (MI:0915) with Importin beta-3 (uni-protkb:O00410) by pull down (MI:0096)
� 2010 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
Biosynthesis of ribosomes requires highly efficient nuclear im-port of ribosomal proteins. The sequence of human large ribo-somal protein L7 carries multiple nuclear localization signals(NLS) [1,2], which are responsible for the nuclear import of L7and are required for ribosomal synthesis [2]. Interestingly, thesame region has also been reported to be the primary target ofauto-antibodies that are often elicited in nuclear-related auto-immunity disease [3,4]. Moreover, the L7 gene has been fre-quently reported to be up-regulated in cancer cells such as JurkatT-lymphoma [5], cervical cancer cells [6] and neoplastic colorectalcancer cells [7]. Moreover, L7 has been considered to be a regula-tor of cell proliferation [5,8] as well as a cold-responsive gene [9].
chemical Societies. Published by E
ciences, National Yang-Ming6 02 2826 4930.
In addition, L7 also displays an ER binding property [10]. Consid-ering the versatile roles that L7 plays in the cell, how L7 under-goes nuclear import, thereby participating in ribosome assemblyto maintain cell homeostasis, is an important issue. For this rea-son, the present study aims to characterize how L7 gains nuclearentry, including identification of the responsible carrier proteinfor L7 as well as elucidation of the roles of RanGTP and thoseof the basic clusters.
2. Materials and methods
2.1. Plasmids and protein purification
Plasmids carrying the genes for L7, N54 and mutants of DN1,DN1+2 and DN1+2+3 have been described previously [2]. Plasmidscontaining the individual basic clusters linked to (EGFP)3 wereindividually cloned into a CMV-driven eukaryotic expressionvector. Plasmid pET28a/Impb3 containing the human importin b3
lsevier B.V. All rights reserved.

4152 C.-W. Chou et al. / FEBS Letters 584 (2010) 4151–4156
coding sequence was obtained using PCR from a human liver cDNAlibrary.
The pET28a plasmids were expressed in Escherchia coli BL21cells and the recombinant protein purified using Ni++-affinitychromatography.
Fig. 1. The characterization of importin b3. (A) The preparation of GST-fused N54. SDSidentified by Western blotting using anti-b3 antibody. (C) The preparation of recombinoverlay assay. SDS–PAGE analysis of L7 and its mutant proteins (a); probed by recombinan(E) The effect of RanGTP. Unbound importin b3 from chromatographic competition was
2.2. GST-pull down assay
GST-conjugated N54 protein was prepared by expressingpGEX-4T-1/N54, encoding N54, with a thrombin-cleavable GSTtag in E. coli BL21 cells. GST-N54 protein was purified using
–PAGE (I); Western blot using anti-GST antibody (II). (B) GST-pull down productsant importin b3. SDS–PAGE (I); Western blot using anti-b3 antibody (II). (D) Thet [35S]-importin b3 (b). Human small ribosomal protein S20 was used as the control.identified using anti-b3 antibody.

C.-W. Chou et al. / FEBS Letters 584 (2010) 4151–4156 4153
Glutathione–Sepharose resin by elution with 50 mM Tris (pH 8.0)and 10 mM glutathione.
HeLa cell lysate was used for the GST-pull down assay. The pull-down products were analyzed using SDS–PAGE and Western blot-ting with an anti-human importin b3 (anti-b3) antibody (SantaCruz Co. USA).
2.3. Overlay assay with importin b3
The overlay assay [11] was carried out in buffer (20 mM HEPESpH 7.5, 110 mM KOAc, 2 mM MgCl2, 0.1% Tween 20 and 5% BSA)using in vivo-labeled [35S]-importin b3 probe.
2.4. RanGTP release experiments
The RanGTP release experiments were carried out using theprocedure of Chang and Tarn [12] with the Q69L mutant form ofRan protein (a kind gift from Dr. W.-Y. Tarn, Academia Sinica, Tai-wan). Bound and unbound products from chromatograph competi-tion with RanGTP or RanGDP were identified by Western blot usinganti-b3 antibody.
2.5. Kinetic binding assay using an IAsys biosensor
The kinetic binding assay was carried out using a resonant mir-ror biosensor (IAsys; Affinity Sensors, Cambridge, UK) according tothe manufacturer’s instructions. The binding and dissociationevents were measured as the arc second response in resonance an-gle arising as protein in free solution bound to or dissociated fromthe immobilized importin b3. The association and dissociation rateconstants, kass and kdiss, were calculated using non-linear curve-fit-ting FASTfit software (Affinity Sensors). The dissociation constant,KD, was calculated as kdiss/kass.
2.6. Nuclear import assay in digitonin-treated cells
Fluorescently labeled recombinant L7 was prepared by chemi-cally conjugating L7 with the fluorescent dye Alexa 555 (InvitrogenCo.).
The preparation of permeabilized cells and the import assaywere carried out as previously described [13]. The cells were incu-bated at room temperature for 15 min with 24 ll of import mix-ture containing excess Alexa 555-conjugated L7 with or withoutrecombinant importin b3 in an energy regenerating cocktail [13].After washing, the cells were observed under a confocalmicroscopy.
Fig. 2. The kinetics of importin b3 binds L7 determined by IAsys biosensor. (A) The concenplot of kon against ligand L7 concentration (r = 0.96). The derived KD value is 1.49 � 10�
The importin b3-depleted lysate was prepared by incubating500 ll lysate (5 � 106 HeLa cells) with anti-b3 antibody (50 lg)for 30 min at room temperature. The reacted antibodies were re-moved using protein A beads (Millipore Co.).
2.7. Effect of importin b3 silencing
The siRNA knock-down procedure was carried out as previouslydescribed [14]. The knock-down efficiency was checked at 48, 72and 96 h post-transfection by Western blot using an anti-b3antibody.
Cells at 72 h post siRNA-treatment were transfected with plas-mids carrying genes that encoded different basic cluster-(EGFP)3
cargo protein, and the cellular distribution of the (EGFP)3 cargoprotein was observed at 24 h post-transfection.
2.8. Fluorescence recovery after photobleaching (FRAP) assay
The procedure for the FRAP experiments was similar to Daele-mans et al. [15] with minor modifications. Transfected cells weregrown on a special microwell dish (MatTek Co.) which was directlymounted onto a confocal microscope equipped with a warm stagefor live observation at 37 �C. The complete area of the nucleus in acell was photobleached; bleaching was completed in 2 min, andrecovery images were acquired at a rate of one frame every 20 sfor 12 min. Next, a defined field of view in the bleached area wasscanned every frame, and used for the subsequent calculations.All FRAP experiments were carried out on cells at 8 h post-transfection.
The recovery rate of each (EGFP)3 chimeric cargo was quantifiedby assessing when the recovery fluorescent intensity reached 10%of the original intensity, termed t10%.
3. Results and discussion
3.1. Importin b3 is the carrier protein for L7
The N-terminal 54 amino acids of L7 contain three potential nu-clear localization signals, and we have previously shown that thisregion can direct nuclear localization [2]. Using the GST-conju-gated N54 peptide (Fig. 1A) as bait to pull down candidates fromHeLa cell extract, importin b3 was positively identified by anti-b3antibody (Fig. 1B). Next, the human importin b3 gene was clonedand expressed, allowing purification to homogeneity (Fig. 1C).The specificity of importin b3 towards L7 was assessed using anoverlay assay, and the result showed that importin b3 positively
tration-dependent binding of L7 to immobilized importin b3. (B) The corresponding7 M.
4154 C.-W. Chou et al. / FEBS Letters 584 (2010) 4151–4156
interacted with the N54 peptide, L7, GST-N54, as well as mutant L7proteins, DN1, DN1+2 and DN1+2+3, which contained variousnumber of clusters (Fig. 1D). This indicates that importin b3 inter-acted multifacetedly with L7 basic cluster, and allowed proteins toundergo nuclear entry as reported previously [2]. A RanGTP releaseexperiment [12] revealed that the binding between importin b3and the basic cluster is RanGTP dependent. This was evident fromthe fact that the binding of importin b3 to GST-N54 was uncoupled
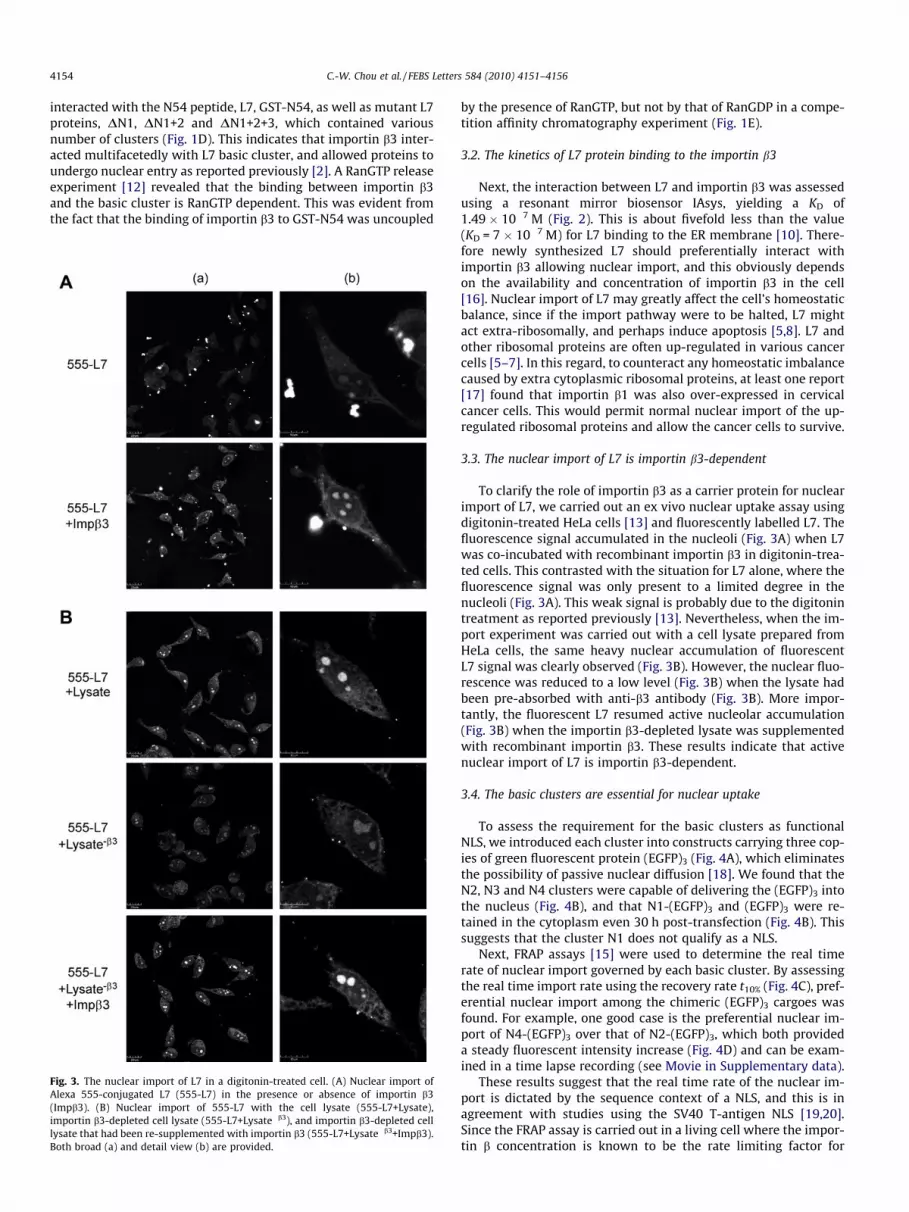
Fig. 3. The nuclear import of L7 in a digitonin-treated cell. (A) Nuclear import ofAlexa 555-conjugated L7 (555-L7) in the presence or absence of importin b3(Impb3). (B) Nuclear import of 555-L7 with the cell lysate (555-L7+Lysate),importin b3-depleted cell lysate (555-L7+Lysate�b3), and importin b3-depleted celllysate that had been re-supplemented with importin b3 (555-L7+Lysate�b3+Impb3).Both broad (a) and detail view (b) are provided.
by the presence of RanGTP, but not by that of RanGDP in a compe-tition affinity chromatography experiment (Fig. 1E).
3.2. The kinetics of L7 protein binding to the importin b3
Next, the interaction between L7 and importin b3 was assessedusing a resonant mirror biosensor IAsys, yielding a KD of1.49 � 10�7 M (Fig. 2). This is about fivefold less than the value(KD = 7 � 10�7 M) for L7 binding to the ER membrane [10]. There-fore newly synthesized L7 should preferentially interact withimportin b3 allowing nuclear import, and this obviously dependson the availability and concentration of importin b3 in the cell[16]. Nuclear import of L7 may greatly affect the cell’s homeostaticbalance, since if the import pathway were to be halted, L7 mightact extra-ribosomally, and perhaps induce apoptosis [5,8]. L7 andother ribosomal proteins are often up-regulated in various cancercells [5–7]. In this regard, to counteract any homeostatic imbalancecaused by extra cytoplasmic ribosomal proteins, at least one report[17] found that importin b1 was also over-expressed in cervicalcancer cells. This would permit normal nuclear import of the up-regulated ribosomal proteins and allow the cancer cells to survive.
3.3. The nuclear import of L7 is importin b3-dependent
To clarify the role of importin b3 as a carrier protein for nuclearimport of L7, we carried out an ex vivo nuclear uptake assay usingdigitonin-treated HeLa cells [13] and fluorescently labelled L7. Thefluorescence signal accumulated in the nucleoli (Fig. 3A) when L7was co-incubated with recombinant importin b3 in digitonin-trea-ted cells. This contrasted with the situation for L7 alone, where thefluorescence signal was only present to a limited degree in thenucleoli (Fig. 3A). This weak signal is probably due to the digitonintreatment as reported previously [13]. Nevertheless, when the im-port experiment was carried out with a cell lysate prepared fromHeLa cells, the same heavy nuclear accumulation of fluorescentL7 signal was clearly observed (Fig. 3B). However, the nuclear fluo-rescence was reduced to a low level (Fig. 3B) when the lysate hadbeen pre-absorbed with anti-b3 antibody (Fig. 3B). More impor-tantly, the fluorescent L7 resumed active nucleolar accumulation(Fig. 3B) when the importin b3-depleted lysate was supplementedwith recombinant importin b3. These results indicate that activenuclear import of L7 is importin b3-dependent.
3.4. The basic clusters are essential for nuclear uptake
To assess the requirement for the basic clusters as functionalNLS, we introduced each cluster into constructs carrying three cop-ies of green fluorescent protein (EGFP)3 (Fig. 4A), which eliminatesthe possibility of passive nuclear diffusion [18]. We found that theN2, N3 and N4 clusters were capable of delivering the (EGFP)3 intothe nucleus (Fig. 4B), and that N1-(EGFP)3 and (EGFP)3 were re-tained in the cytoplasm even 30 h post-transfection (Fig. 4B). Thissuggests that the cluster N1 does not qualify as a NLS.
Next, FRAP assays [15] were used to determine the real timerate of nuclear import governed by each basic cluster. By assessingthe real time import rate using the recovery rate t10% (Fig. 4C), pref-erential nuclear import among the chimeric (EGFP)3 cargoes wasfound. For example, one good case is the preferential nuclear im-port of N4-(EGFP)3 over that of N2-(EGFP)3, which both provideda steady fluorescent intensity increase (Fig. 4D) and can be exam-ined in a time lapse recording (see Movie in Supplementary data).
These results suggest that the real time rate of the nuclear im-port is dictated by the sequence context of a NLS, and this is inagreement with studies using the SV40 T-antigen NLS [19,20].Since the FRAP assay is carried out in a living cell where the impor-tin b concentration is known to be the rate limiting factor for

C.-W. Chou et al. / FEBS Letters 584 (2010) 4151–4156 4155
protein nuclear import [16], we therefore investigated the nuclearuptake behaviors of the cluster-(EGFP)3 cargoes when the expres-sion of importin b3 had been down regulated. This was carriedout using the siRNA knock-down approach. The results were ratherunexpected, since the import behavior of N2-, or the N3-(EGFP)3
cargo did not seem to be affected by the down regulation ofimportin b3 (Fig. 5). In contrast, the knock-down of importin b3
Fig. 4. Assessment of the role the basic clusters play for nuclear import of L7. (A) ConstruHeLa cell at 18, 24 and 30 h post-transfection. Bar = 10 lm. (C) The dynamic nuclear impimages of N2-(EGFP)3 and N4-(EGFP)3 during a 12-min time course.
had a great influence on the nuclear uptake of the N4-(EGFP)3 car-go (Fig. 5). The result suggests that the interaction between the N4cluster and importin b3 could play the major role in nuclear importof L7, notwithstanding the fact that the other basic clustersfunction as a NLS and each has a different rate of nuclear import.Nevertheless, the common existence of a multiple NLS as repre-sented by these clusters in L7 can not be ignored, and therefore
cts of chimeric (EGFP)3 cargoes. (B) Cellular localization observed in the transfectedort observed in FRAP assay (I) and their rate expressed as 1/t10% (II). (D) Post-bleach

Fig. 5. Effects of importin b3 silencing on the nuclear import of cluster-(EGFP)3
cargoes. (A) The importin b3 siRNA silencing effect detected by Western blottingusing anti-b3 antibody. GAPDH was used as the control. (B) The silencing effect onthe nuclear import of N2-, N3-, and N4-(EGFP)3 cargoes. Bar = 10 lm.
4156 C.-W. Chou et al. / FEBS Letters 584 (2010) 4151–4156
the importance of having such a cluster attached to L7 needs to beexplained. One possible answer is that these clusters are part of amultifaceted interaction that ensures highly efficient targeting ofL7 to the nucleus [2] and provide L7 with an advantage relativeto other proteins, namely preferentially nuclear import.
Acknowledgments
This work was supported in part by Grants from National Sci-ence Council, Taiwan (NSC93-2312-B010-004), and a Grant fromthe Ministry of Education, Aim for the Top University Plan.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.febslet.2010.08.044.
References
[1] Lin, A., Chan, Y.L., McNally, J., Peleg, D., Meyuhas, O. and Wool, I.G. (1987) Theprimary structure of rat ribosomal protein L7. The presence near the aminoterminus of L7 of five tandem repeats of a sequence of 12 amino acids. J. Biol.Chem. 262, 12665–12671.
[2] Ko, J.R., Wu, J.Y., Kirby, R., Li, I.F. and Lin, A. (2006) Mapping the essentialstructures of human ribosomal protein L7 for nuclear entry, ribosomeassembly and function. FEBS Lett. 580, 3804–3810.
[3] von Mikecz, A., Hemmerich, P., Peter, H.H. and Krawinkel, U. (1994)Characterization of eukaryotic protein L7 as a novel autoantigen whichfrequently elicits an immune response in patients suffering from systemicautoimmune disease. Immunobiology 192, 137–154.
[4] Neu, E., Hemmerich, P.H., Peter, H.H., Krawinkel, U. and von Mikecz, A.H.(1997) Characteristic epitope recognition pattern of autoantibodies againsteukaryotic ribosomal protein L7 in systemic autoimmune diseases. ArthriticRheum. 40, 661–671.
[5] Neumann, F. and Krawinkel, U. (1997) Constitutive expression of humanribosomal protein L7 arrests the cell cycle in G1 and induces apoptosis inJurkat T-lymphoma cells. Exp. Cell Res. 230, 252–261.
[6] Denko, N., Schindler, C., Koong, A., Laderoute, K., Green, C. and Giaccia, A.(2000) Epigenetic regulation of gene expression in cervical cancer cells by thetumor microenvironment. Clin. Cancer Res. 6, 480–487.
[7] Kasai, H., Nadano, D., Hidaka, E., Higuchi, K., Kawakubo, M., Sato, T.A. andNakayama, J. (2003) Differential expression of ribosomal proteins in humannormal and neoplastic colorectum. J. Histochem. Cytochem. 51, 567–574.
[8] Chen, F.W. and Ioannou, Y.A. (1999) Ribosomal proteins in cell proliferationand apoptosis. Int. Rev. Immunol. 18, 429–448.
[9] Wu, S., De Croos, J.N. and Storey, K.B. (2008) Cold acclimation-induced up-regulation of the ribosomal protein L7 gene in the freeze tolerant wood frog,Rana sylvatica. Gene 424, 48–55.
[10] Wu, W.C., Liu, H.W. and Lin, A. (2007) Human ribosomal protein L7 displays anER binding property and is involved in ribosome-ER association. FEBS Lett.581, 651–657.
[11] Aitchison, J.D., Blobel, G. and Rout, M.P. (1996) Kap104p: a karyopherininvolved in the nuclear transport of messenger RNA binding proteins. Science274, 624–627.
[12] Chang, W.L. and Tarn, W.Y. (2009) A role for transportin in deposition of TTP tocytoplasmic RNA granules and mRNA decay. Nucleic Acids Res. 37, 6600–6612.
[13] Jäkel, S. and Görlich, D. (1998) Importin beta, transportin, RanBP5 and RanBP7mediate nuclear import of ribosomal proteins in mammalian cells. EMBO J. 17,4491–4502.
[14] Deng, T., Engelhardt, O.G., Thomas, B., Akoulitchev, A.V., Brownlee, G.G. andFodor, E. (2006) Role of ran binding protein 5 in nuclear import andassembly of the influenza virus RNA polymerase complex. J. Virol. 80,11911–11919.
[15] Daelemans, D., Costes, S.V., Lockett, S. and Pavlakis, G.N. (2005) Kinetic andmolecular analysis of nuclear export factor CRM1 association with its cargoin vivo. Mol. Cell. Biol. 25, 728–739.
[16] Yang, W. and Musser, S.M. (2006) Nuclear import time and transportefficiency depend on importin beta concentration. J. Cell Biol. 174, 951–961.
[17] van der Watt, P.J., Maske, C.P., Hendricks, D.T., Parker, M.I., Denny, L.,Govender, D., Birrer, M.J. and Leaner, V.D. (2009) The Karyopherin proteins,Crm1 and Karyopherin beta1, are overexpressed in cervical cancer and arecritical for cancer cell survival and proliferation. Int. J. Cancer 124, 1829–1840.
[18] Beetz, C., Brodhun, M., Moutzouris, K., Kiehntopf, M., Berndt, A., Lehnert, D.,Deufel, T., Bastmeyer, M. and Schickel, J. (2004) Identification of nuclearlocalisation sequences in spastin (SPG4) using a novel Tetra-GFP reportersystem. Biochem. Biophys. Res. Commun. 318, 1079–1084.
[19] Hodel, M.R., Corbett, A.H. and Hodel, A.E. (2001) Dissection of a nuclearlocalization signal. J. Biol. Chem. 276, 1317–1325.
[20] Hodel, A.E., Harreman, M.T., Pulliam, K.F., Harben, M.E., Holmes, J.S., Hodel,M.R., Berland, K.M. and Corbett, A.H. (2006) Nuclear localization signalreceptor affinity correlates with in vivo localization in Saccharomycescerevisiae. J. Biol. Chem. 281, 23545–23556.





























