Embed Size (px)
Citation preview

Short communication
Identification of a novel herpesvirus from a California desert
tortoise (Gopherus agassizii)
April J. Johnson a,*, Allan P. Pessier b, James F. X. Wellehan a,Roseanne Brown c, Elliott R. Jacobson a
a Department of Small Animal Clinical Sciences, College of Veterinary Medicine, University of Florida, P.O. Box 100126,
Gainesville, FL 32610, USAb Department of Pathology, Center for Reproduction of Endangered Species, Zoological Society of San Diego,
San Diego, CA 92112-0551, USAc Rancho San Diego Animal Hospital, 2988 Jamacha Rd., Suite 176, El Cajon, CA 92013, USA
Received 12 May 2005; received in revised form 22 August 2005; accepted 14 September 2005
Abstract
Herpesviruses are significant pathogens of tortoises, causing upper respiratory tract disease and necrotizing stomatitis, with
infections often associated with high mortality rates. Herpesvirus infection in a captive California desert tortoise (Gopherus
agassizii) was detected by light microscopic observation of intranuclear inclusion bodies in various tissues followed by
transmission electron microscopic observation of herpesvirus-like particles, and amplification of herpesvirus nucleic acid
sequences using polymerase chain reaction. Using an indirect enzyme linked immunosorbent assay, anti-tortoise herpesvirus
antibodies were detected one month after initial onset of clinical signs. This novel herpesvirus is distinct from the previously
described tortoise herpesvirus (tortoise herpesvirus-1, THV-1) sharing 83% sequence identity of 60 amino acids of a portion of
the DNA polymerase gene and 79% sequence identity across 120 amino acids of a portion of the ribonucleotide reductase gene.
Similar to THV-1, this novel herpesvirus, tortoise herpesvirus-2 (THV-2), also clusters with the alphaherpesviruses.
# 2005 Elsevier B.V. All rights reserved.
Keywords: Desert tortoise; Gopherus agassizii; Herpesvirus; Reptiles
www.elsevier.com/locate/vetmic
Veterinary Microbiology 111 (2005) 107–116
1. Introduction
Herpesvirus infections were first described in
tortoises in the early 1980s (Harper et al., 1982).
* Corresponding author. Tel.: +1 352 392 4700x5256;
fax: +1 352 392 5464.
E-mail address: [email protected] (A.J. Johnson).
0378-1135/$ – see front matter # 2005 Elsevier B.V. All rights reserved
doi:10.1016/j.vetmic.2005.09.008
They have since been reported in many species with
varying clinical signs and degrees of severity
(Jacobson et al., 1985; Heldstab and Bestetti, 1989;
Drury et al., 1998; Muro et al., 1998). Infection is most
often associated with the oral cavity and respiratory
tract, with necrotizing stomatitis, glossitis, tracheitis,
pharyngitis, and rhinitis having been repeatedly
described (Jacobson et al., 1985; Muro et al., 1998;
.

A.J. Johnson et al. / Veterinary Microbiology 111 (2005) 107–116108
Fig. 1. Caseous yellow plaques in the oral cavity of the desert
tortoise on January 14, 2003.
Drury et al., 1998). Encephalitis (Heldstab and
Bestetti, 1989) and hepatitis (Hervas et al., 2002)
have also been observed.
Diagnosis of herpesvirus infections in tortoises
have been made by observation of intranuclear
inclusion bodies by light microscopy followed by
visualization of virus particles by transmission
electron microscopy (TEM) (Jacobson et al., 1985;
Muro et al., 1998), DNA in situ hybridization (Teifke
et al., 2000), and immunohistochemistry (Origgi et al.,
2003). Virus isolation on susceptible cell lines can be
performed (Marschang et al., 2001) followed by
negative staining electron microscopy. Amplification
of herpesvirus specific DNA segments can be
performed using consensus (VanDevanter et al.,
1996; Une et al., 2000) or tortoise herpesvirus specific
(Teifke et al., 2000; Origgi et al., 2004) polymerase
chain reaction (PCR) primers. Virus neutralization and
indirect enzyme linked immunosobent assay (ELISA)
have been used to detect anti-herpesvirus antibodies
(Marschang et al., 2001; Origgi et al., 2001). Three
tortoise herpesvirus sequences exist in GenBank for
three genes including the UL5 helicase primase
(Accession #AY188757), UL39 (Accession
#AY338245) and the DNA polymerase (Accession
#AB047545) genes.
The desert tortoise (Gopherus agassizii) has been
experiencing dramatic declines throughout parts of its
range and populations north and west of the Colorado
River are listed as threatened by the US federal
government (USFWS, 1994). Upper respiratory tract
disease (URTD) has been implicated as one of the
causes of declining numbers (USFWS, 1994). Captive
tortoises with URTD were first observed in the late
1970s (Fowler, 1980; Snipes and Biberstein, 1982)
and in wild desert tortoises in the late 1980s (Jacobson
et al., 1991). Mycoplasma agassizii was subsequently
identified and demonstrated to be a causative agent of
URTD (Brown et al., 1994). Clinical signs of
herpesvirus infection overlap with those of myco-
plasmosis including nasal and ocular discharge and
palpebral and periocular edema. The possibility that
herpesviruses may also cause URTD in desert tortoises
has not been thoroughly investigated. Using an
indirect ELISA and herpesvirus isolated from Med-
iterranean tortoises (Testudo graeca) as the coating
antigen, a recent survey of 109 captive desert tortoises
in southern California showed 26.6% to be seropo-
sitive for exposure to tortoise herpesvirus (Johnson
et al., In press). While there are currently three
published reports of herpesvirus-like particles being
detected by TEM in desert tortoises (Harper et al.,
1982; Pettan-Brewer et al., 1996; Martınez-Silvestre
et al., 1999), the phylogenetic relationship between
these viruses is unknown since no sequencing data is
available. Here we describe a novel herpesvirus from a
captive California desert tortoise with a necrotizing
stomatitis.
2. Materials and methods
2.1. Case summary
An adult female captive desert tortoise of unknown
age was presented to a veterinary clinic in El Cajon,
California on January 6, 2003, after an unusually early
exit from hibernation and exhibiting clinical signs of
anorexia and lethargy. She had been obtained through
an adoption agency in August of 2002 and placed in a
homewith three desert tortoises and one Texas tortoise
(Gopherus berlandieri). Physical examination
revealed yellow-white caseous plaques on the tongue
and palate (Fig. 1). A biopsy of the tongue was taken
on January 14 for light microscopic examination, PCR
and virus isolation. Plasma samples were collected on

A.J. Johnson et al. / Veterinary Microbiology 111 (2005) 107–116 109
Fig. 2. Caseous yellow and gray plaques in the oral cavity of the
desert tortoise at necropsy on February 11, 2003.
January 14, January 27 and February 6 for detection of
anti-herpesvirus antibodies by ELISA and plasma
samples were also collected for ELISA on February 4
from the four tortoises housedwith the ill desert tortoise.
Supportive therapy consisting of enrofloxacin, doxycy-
line, fluid therapy, and tube feeding was instituted. Oral
plaques progressed to cover the entire oral cavity (Fig. 2)
and the tortoise died on February 10. A necropsy was
performed and tissues from all major organs were
collected for histopathology, virus isolation and PCR.
2.2. Light and electron microscopy
A portion of the tongue biopsy and tissues collected
at necropsy were fixed in 10% neutral buffered
formalin and processed for histopathology. Paraffin
embedded tissues were cut into 6 mm sections and
stained with hematoxylin and eosin. Portions of the
paraffin embedded tissue from the tongue biopsy were
deparaffinized in xylene, embedded in Spurr’s resin,
and sectioned for TEM at the Electron Microscopy
Core Laboratory, University of Florida.
2.3. Virus isolation
A portion of the tongue biopsy and liver and tongue
collected at necropsy were homogenized and inocu-
lated onto 25 cm2 flasks (Costar, Corning, NY)
containing monolayers of turtle heart cells (TH1
cells; ATCC-CCL 50, American Type Culture
Collection, Rockville, MD) in Dulbecco’s Modified
Eagle Medium (DMEM, Gibco, Carlsbad, CA)
supplemented with 5% fetal bovine serum (Gibco),
gentamicin (60 mg/L; Sigma, St. Louis, MO),
penicillin G (120,000 U/L; Sigma), streptomycin
(120,000 U/L; Sigma) and amphotericin B (300 mg/
L; Sigma). Cells were incubated at 28 8C in the
presence of 5%CO2 and flasks were observed daily for
signs of cytopathic effects.
2.4. ELISA
Plasma samples were evaluated for the presence of
anti-herpesvirus antibodies by indirect enzyme linked
immunosorbent assay (ELISA). A previously
described method was used (Origgi et al., 2001) but
was adapted for a desert tortoise by using mouse-anti-
desert tortoise monoclonal biotinylated antibodies
(Schumacher et al., 1993) in place of mouse-anti-
Greek tortoise monoclonal biotinylated antibodies.
The plates were coated with THV-1 isolated and
purified from a Mediterranean tortoise, which was
used in the original ELISA; the positive optical density
cut off value used was that previously determined in
Mediterranean tortoises (Origgi et al., 2001).
2.5. PCR and DNA sequencing
DNA was extracted from a portion of the tongue
biopsy and brain, tongue, spleen, liver, kidney,
bladder, and adrenal gland collected at necropsy
using the Qiagen DNeasy kit (Qiagen, Valencia, CA).
Nested PCR amplification of partial sequence of the
herpesvirus DNA-dependent DNA polymerase gene
and direct PCR utilizing tortoise herpesvirus specific
primers targeting the ribonucleotide reductase gene
was performed using methods previously described
(VanDevanter et al., 1996; Origgi et al., 2004). The
PCR products were resolved in 1% agarose gels,
excised, and purified using the QIAquick gel
extraction kit (Qiagen). Products were sequenced
directly in both directions using the Big-Dye
Terminator Kit (Perkin-Elmer, Branchburg, NJ) and
analyzed on an ABI 377 automated DNA sequencer at
the University of Florida’s Sequencing Center.
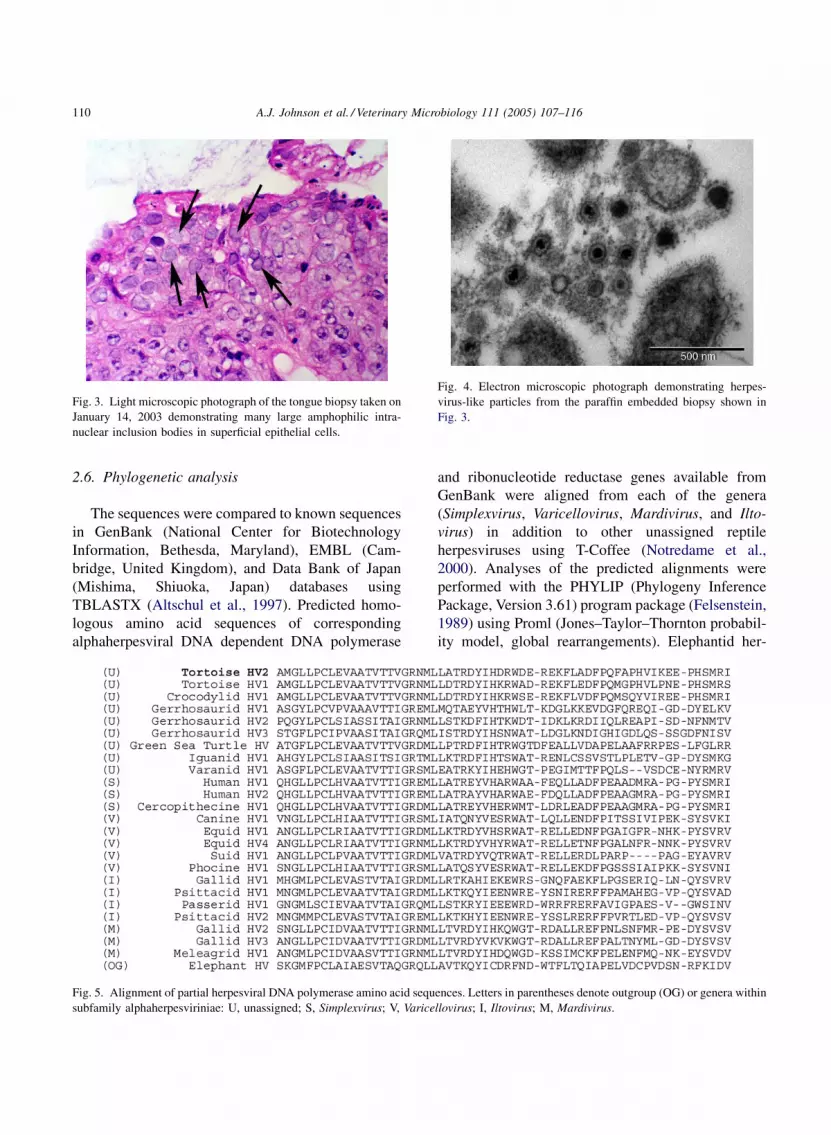
A.J. Johnson et al. / Veterinary Microbiology 111 (2005) 107–116110
Fig. 3. Light microscopic photograph of the tongue biopsy taken on
January 14, 2003 demonstrating many large amphophilic intra-
nuclear inclusion bodies in superficial epithelial cells.
Fig. 4. Electron microscopic photograph demonstrating herpes-
virus-like particles from the paraffin embedded biopsy shown in
Fig. 3.
2.6. Phylogenetic analysis
The sequences were compared to known sequences
in GenBank (National Center for Biotechnology
Information, Bethesda, Maryland), EMBL (Cam-
bridge, United Kingdom), and Data Bank of Japan
(Mishima, Shiuoka, Japan) databases using
TBLASTX (Altschul et al., 1997). Predicted homo-
logous amino acid sequences of corresponding
alphaherpesviral DNA dependent DNA polymerase
Fig. 5. Alignment of partial herpesviral DNA polymerase amino acid sequ
subfamily alphaherpesviriniae: U, unassigned; S, Simplexvirus; V, Varice
and ribonucleotide reductase genes available from
GenBank were aligned from each of the genera
(Simplexvirus, Varicellovirus, Mardivirus, and Ilto-
virus) in addition to other unassigned reptile
herpesviruses using T-Coffee (Notredame et al.,
2000). Analyses of the predicted alignments were
performed with the PHYLIP (Phylogeny Inference
Package, Version 3.61) program package (Felsenstein,
1989) using Proml (Jones–Taylor–Thornton probabil-
ity model, global rearrangements). Elephantid her-
ences. Letters in parentheses denote outgroup (OG) or genera within
llovirus; I, Iltovirus; M, Mardivirus.

A.J. Johnson et al. / Veterinary Microbiology 111 (2005) 107–116 111
p(O
G)orgenerawithin
subfamily
pesvirus-1 (GenBank Accession #AF322977), a
betaherpesvirus, was used as the outgroup for the
polymerase gene and human herpesvirus-4 (GenBank
Accession #NC_001345), a gammaherpesvirus, was
used as the outgroup for the ribonucleotide reductase
gene. The strength of both tree topologies obtained
were tested by bootstrap analysis (Felsenstein, 1985)
starting with Seqboot with 100 resamplings, followed
by maximum likelihood calculations. Consense was
used to calculate the bootstrap values.
Fig.6.Alignmentofpartial
herpesviral
ribonucleotidereductasegeneam
i no
acid
sequences.
Letters
inparentheses
denote
outgrou
alphaherpesviriniae:
U,unassigned;S,
Sim
ple
xvir
us;V,
Vari
cell
ovi
rus;
I,Il
tovi
rus;M,
Mar d
ivir
us.
3. Results
3.1. Light and electron microscopy
Histologic examination of the tongue biopsy
demonstrated many large amphophilic intranuclear
inclusion bodies in superficial epithelial cells (Fig. 3).
Histologic examination of tissues collected at
necropsy revealed a chronic and active necrotizing
stomatitis, pharyngitis, and tracheitis. Epithelial
surfaces in these areas were covered with a thick
coagulum composed of sloughed epithelial cells,
heterophils and bacterial colonies. Sloughed and
viable epithelial cells frequently had eosinophilic to
amphophilic intranuclear inclusion bodies. The sub-
mucosa in affected regions had variable mixed
infiltrates of heterophils, lymphocytes, plasma cells,
and macrophages. In some areas of the oropharynx,
there was complete epithelial loss and replacement by
granulation tissue. The only other significant histo-
logic finding was a granulomatous bronchopneumonia
with intralesional bacterial colonies. Sections of the
brain, liver, spleen, pancreas, small intestine, kidney,
urinary bladder, adrenal gland, thyroid gland, ovary,
and heart were unremarkable.
Transmission electron microscopy of the paraffin
embedded tongue biopsy revealed non-enveloped
virus particles within intranuclear inclusions in
epithelial cells. Virions were approximately 120 nm
in diameter and consistent in size, site of replication,
and morphology with herpesvirus (Fig. 4).
3.2. Virus isolation
All attempts at virus isolation failed to show any
cytopathic effects up to 30 days after inoculation.
A.J. Johnson et al. / Veterinary Microbiology 111 (2005) 107–116112
3.3. ELISA
ELISA performed on the desert tortoise plasma
samples were negative on January 14 and 27 but
positive on February 6. The ELISA performed on the
plasma samples collected from the other four
tortoises housed with the desert tortoise collected
on February 4 showed that two desert tortoises and
one Texas tortoise were positive for anti-tortoise
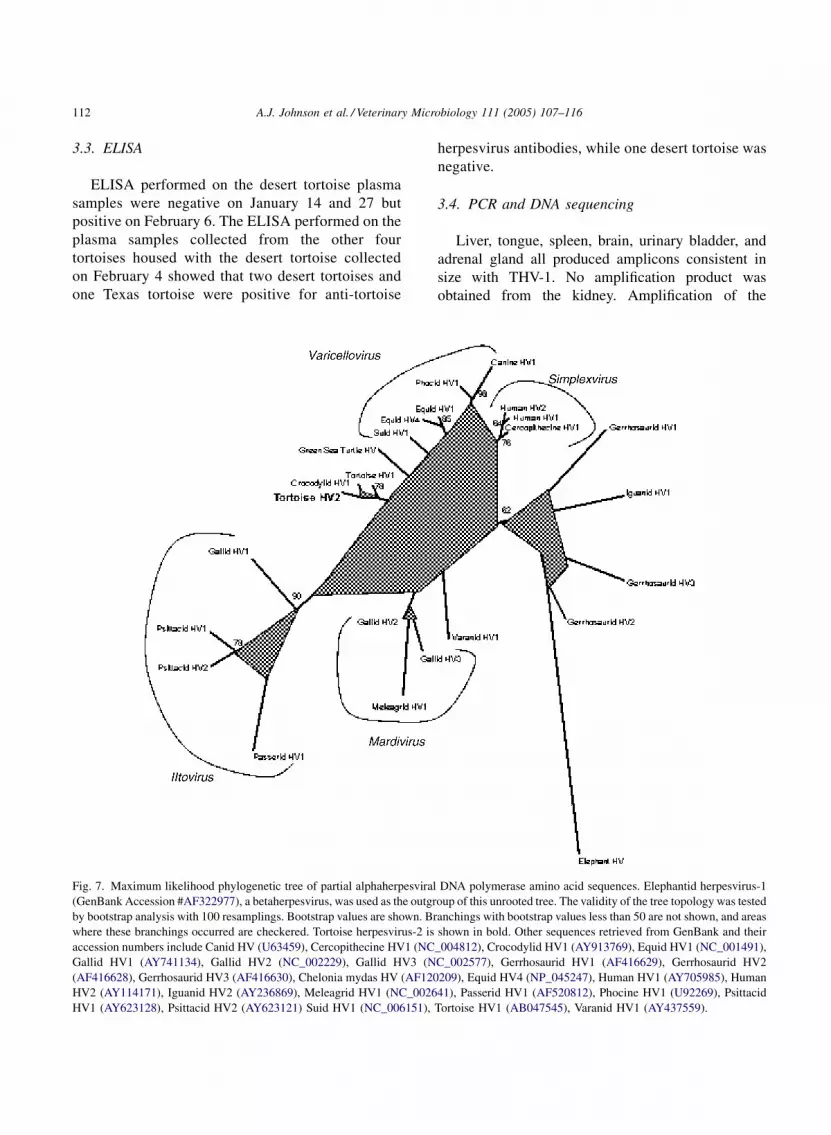
Fig. 7. Maximum likelihood phylogenetic tree of partial alphaherpesviral
(GenBank Accession #AF322977), a betaherpesvirus, was used as the outgr
by bootstrap analysis with 100 resamplings. Bootstrap values are shown. Br
where these branchings occurred are checkered. Tortoise herpesvirus-2 is
accession numbers include Canid HV (U63459), Cercopithecine HV1 (NC
Gallid HV1 (AY741134), Gallid HV2 (NC_002229), Gallid HV3 (N
(AF416628), Gerrhosaurid HV3 (AF416630), Chelonia mydas HV (AF120
HV2 (AY114171), Iguanid HV2 (AY236869), Meleagrid HV1 (NC_0026
HV1 (AY623128), Psittacid HV2 (AY623121) Suid HV1 (NC_006151),
herpesvirus antibodies, while one desert tortoise was
negative.
3.4. PCR and DNA sequencing
Liver, tongue, spleen, brain, urinary bladder, and
adrenal gland all produced amplicons consistent in
size with THV-1. No amplification product was
obtained from the kidney. Amplification of the
DNA polymerase amino acid sequences. Elephantid herpesvirus-1
oup of this unrooted tree. The validity of the tree topology was tested
anchings with bootstrap values less than 50 are not shown, and areas
shown in bold. Other sequences retrieved from GenBank and their
_004812), Crocodylid HV1 (AY913769), Equid HV1 (NC_001491),
C_002577), Gerrhosaurid HV1 (AF416629), Gerrhosaurid HV2
209), Equid HV4 (NP_045247), Human HV1 (AY705985), Human
41), Passerid HV1 (AF520812), Phocine HV1 (U92269), Psittacid
Tortoise HV1 (AB047545), Varanid HV1 (AY437559).

A.J. Johnson et al. / Veterinary Microbiology 111 (2005) 107–116 113
polymerase gene resulted in a product that was 181
base pairs when primer sequences were edited out and
amplification of the ribonucleotide reductase gene
resulted in a product that was 361 base pairs.
3.5. Phylogenetic analysis
TBLASTX results for the nested PCR sequence
showed the highest score with tortoise herpesvirus
Fig. 8. Maximum likelihood phylogenetic tree of partial alphaherpesvi
herpesvirus-4 (GenBank Accession #P03190), a gammaherpesvirus, was
topology was tested by bootstrap analysis with 100 resamplings. Bootstrap
not shown, and areas where these branchings occurred are checkered. Oth
include Bovine HV1 (CAA9029), Bovine HV2 (AAK56211), Bovine H
(YP_053066), Equid HV4 (CAA53100), Gallid HV1 (AAD56211), Ga
(NP_044509), Meleagrid HV1 (AAG30079), Psittacid HV1 (AAQ73718)
DNA polymerase (GenBank Accession #AB047545),
sharing 83% sequence identity while the direct PCR
showed highest score with the tortoise ribonucleotide
reductase gene (GenBank Accession #AY338245)
sharing 79% sequence identity. Novel sequence data
was submitted to GenBank; the accession number for
the partial polymerase sequence is AY916792 and for
the partial ribonucleotide reductase sequence is
DQ027825. The alignment of partial herpesvirus
ral ribonucleotide reductase gene amino acid sequences. Human
used as the outgroup of this unrooted tree. The validity of the tree
values are shown. Branchings with bootstrap values less than 50 are
er sequences retrieved from GenBank and their accession numbers
V5 (NP_954908), Cercopithecine HV1 (AAP41457), Equid HV1
llid HV2 (AAA80556), Human HV1 (NP_044641), Human HV2
, Suid HV1 (YP_068342), and Tortoise HV1 (AAQ73541).

A.J. Johnson et al. / Veterinary Microbiology 111 (2005) 107–116114
DNA polymerase and ribonucleotide reductase gene
amino acid sequences are shown in Figs. 5 and 6,
respectively. The phylogenetic tree comparing poly-
merase amino acid sequences is shown in Fig. 7 and
the ribonucleotide reductase gene sequences in Fig. 8.
4. Discussion
The lesions seen in the desert tortoise in this study
are consistent with previous reports of herpesvirus
infection in desert tortoises (Harper et al., 1982;
Pettan-Brewer et al., 1996; Martınez-Silvestre et al.,
1999). For the first time, sequencing data was
obtained for a desert tortoise herpesvirus. In a study
sequencing a portion of the gene encoding the DNA
polymerase gene in 5 strains of human herpesvirus-2,
17 strains of human herpesvirus-6 and 5 strains of
human herpesvirus-7, only single base variations that
did not result in alteration of the amino acid sequence
were seen within species (VanDevanter et al., 1996).
Tortoise herpesvirus-2 amino acid sequence differs
by 17% from the previously described tortoise
herpesvirus (Une et al., 2000), supporting the
interpretation that this is a novel virus, and not a
variant of THV-1. The designation THV-2 will
hereafter be used when referring to this virus. The
partial polymerase gene phylogenetic tree (Fig. 7)
demonstrates that human herpesvirus-1 and 2 are
more closely related to each other than THV-1 is
related to THV-2.
It is unknown whether THV-2 is native to North
American tortoises and whether it is affecting wild
populations of desert tortoises. Evidence of herpes-
virus exposure in wild desert tortoises based on
serology with indirect ELISA has been documented,
although until now, herpesvirus infection had not been
confirmed in a seropositive tortoise. Surveys have
shown a seroprevalence of herpesvirus exposure in
wild desert tortoises ranging from 30 to 51%
(Jacobson et al., 2001; Berry et al., 2003). This
suggests that there is a herpesvirus native to wild
desert tortoises or that wild desert tortoises have been
exposed to herpesvirus from non-native tortoises.
However, herpesvirus infection has not been demon-
strated in a wild desert tortoise and therefore cannot be
characterized as THV-1, THV-2, or another novel
herpesvirus. This desert tortoise was likely infected
during captivity because the tortoise did not exhibit
antibodies on ELISA initially, but seroconverted
within a month of clinical evidence of herpesvirus
infection. This tortoise was exposed to three other
desert tortoises and one Texas tortoise (Gopherus
berlandieri) for five months prior to developing
clinical signs of disease. On ELISA, two of the desert
tortoises and the one Texas tortoise were positive for
anti-tortoise herpesvirus antibodies. It is unknown
whether these tortoises had herpesvirus prior to being
taken into captivity, if they were exposed to other
tortoises at some point during captivity or if they had
the same point source as the infected tortoise.
This study shows that the previously described
ELISA (Origgi et al., 2001) can be adapted to detect
herpesvirus exposure in desert tortoises. While certain
herpesviruses are known to permanently infect their
hosts, we do not know if infected tortoises that recover
remain infected for life. In experimental infection
studies, herpesvirus was demonstrated in the central
nervous system of tortoises that were inoculated with
virus and recovered following development of clinical
signs of infection (Origgi et al., 2001).
The data strongly suggests that THV-1 and THV-2
are closely antigenically related, and the currently
used ELISA does not distinguish well between the
two tortoise herpesviruses. Antibodies to this her-
pesvirus cross-reacted to THV-1, the virus used as the
antigen in the indirect ELISA, indicating that
although differing in sequence, both herpesviruses
have similar antigenic sites to which the tortoise can
develop antibodies. A previous transmission study
with Greek tortoises (T. graeca) demonstrated
seroconversion within 4–7 weeks of experimental
infection (Origgi et al., 2001), which was consistent
with the time fromwhen lesions were first observed to
seroconversion.
5. Conclusions
A novel herpesvirus, tortoise herpesvirus-2, was
identified in a California desert tortoise. It is unknown
how this virus differs serologically and biologically to
tortoise herpesvirus-1, although antibodies to THV-2
cross-reacted to THV-1 on ELISA. The previously
described ELISA can be adapted to detect herpesvirus
in desert tortoises.

A.J. Johnson et al. / Veterinary Microbiology 111 (2005) 107–116 115
Acknowledgements
Funding was awarded by Department of Cultural
Natural Resources, Directorate of Public Works at the
National Training Center, Ft. Irwin, CA. We thank the
Cultural and Natural Resources Manager at Ft. Irwin,
Mr. Mickey Quillman, for support of this project,
Sylvia Tucker at the University of Florida and
Marianne Zeitz at the Rancho San Diego Animal
Hospital for their assistance.
References
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z.,
Miller, W., Lipman, D.J., 1997. Gapped BLAST and PSI-
BLAST: a new generation of protein database search programs.
Nucleic Acids Res. 25, 3389–3402.
Berry, K.H., Brown, M.B., Wendland, L., Origgi, F., Johnson, A.,
2003. Health assessments of captive and wild desert tortoises at
26 sites in the Mojave and Colorado deserts, California, in 2002.
In: 28th Annual Meeting and Symposium of the Desert Tortoise
Council, Las Vegas, pp. 6–7.
Brown, M.B., Schumacher, I.M., Klein, P.A., Harris, K., Correll, T.,
Jacobson, E.R., 1994. Mycoplasma agassizii causes upper
respiratory tract disease in the desert tortoise. Infect. Immun.
62, 4580–4586.
Drury, S.E.N., Gough, R.E., McArthur, S., Jessop, M., 1998. Detec-
tion of herpes-like and papilloma-like particles associated with
diseases of tortoises. Vet. Rec. 143, 639.
Felsenstein, J., 1985. Confidence limits on phylogenies: an approach
using the bootstrap. Evolution 39, 783–791.
Felsenstein, J., 1989. PHYLIP—phylogeny inference package. Cla-
distics 5, 164–166.
Fowler, M.E., 1980. Comparison of respiratory infection and hypo-
vitaminosis A in desert tortoises. In: Montali, R.J., Migake, G.
(Eds.), The Comparative Pathology of Zoo Animals. Smithso-
nian Institution Press, Washington, DC, pp. 93–97.
Harper, P.A.W., Hammond, D.C., Heuschele, W., 1982. A herpes-
virus-like agent associated with a pharyngeal abscess in a desert
tortoise. J. Wildl. Dis. 18, 491–494.
Heldstab, A., Bestetti, G., 1989. Herpesviridae causing glossitis and
meningoencephalitis in land tortoises (Testudo hermanii). Her-
petopathologia 1, 5–9.
Hervas, J., Sanchez-Cordon, P.J., Chacon de Lara, F., Carrasco, L.,
Gomez-Villamandos, J.C., 2002. Hepatitis associated with
herpes viral infection in the tortoise (Testudo horsfieldii). J.
Vet. Med. B 49, 111–114.
Jacobson, E.R., Clubb, S., Gaskin, J.M., Gardiner, C., 1985. Her-
pesvirus-like infection in Argentine tortoises. J. Am.Vet. Med.
Assoc. 187, 1227–1229.
Jacobson, E.R., Gaskin, J.M., Brown, M.B., Harris, R.K., Gardiner,
C.H., LaPointe, J.L., Adams, H.P., Reggiardo, C., 1991. Chronic
upper respiratory tract disease of free-ranging desert tortoises
(Xerobates agassizii). J. Wildl. Dis. 27, 296–316.
Jacobson, E.R., Klein, P.A., Romero, C., 2001. Evaluation of desert
tortoises in and around Fort Irwin for exposure to a tortoise
herpesvirus. US Department of the Interior Cooperative Agree-
ment No. 1434-HQ-97-RU-01544 RWO 196.
Johnson, A.J., Morafka, D.J., Jacobson, E.R. Seroprevalence
of Mycoplasma agassizii and tortoise herpesvirus in cap-
tive desert tortoises (Gopherus agassizii) from the greater
Barstow area, Mojave Desert, California. J. Arid Environ., in
press.
Marschang, R.E., Frost, J.W., Gravendyck, M., Kaleta, E.F., 2001.
Comparison of 16 chelonid herpesviruses by virus neutralization
tests and restriction endonuclease digestion of viral DNA. J. Vet.
Med. B 48, 393–399.
Martınez-Silvestre, A., Majo, N., Ramis, A., 1999. Caso clınico:
herpesvirosis en tortuga de desierto Americana (Gopherus
agassizii). Clınica Verinaria de Pequnos Animales 19, 99–
106.
Muro, J., Ramis, A., Pastor, J., Velarde, R., Tarres, J., Lavin, S.,
1998. Chronic rhinitis associated with herpesviral infection in
captive spur-thighed tortoises from Spain. J. Wildl. Dis. 34, 487–
495.
Notredame, C., Higgins, D., Heringa, J., 2000. T-Coffee: a novel
method for multiple sequence alignments. JMol. Biol. 302, 205–
217.
Origgi, F.C., Klein, P.A., Mathes, K., Blahak, S., Marschang, R.E.,
Tucker, S.J., Jacobson, E.R., 2001. Enzyme-linked immunosor-
bent assay for detecting herpesvirus exposure in Mediterranean
tortoises (spur-thighed tortoise [Testudo graeca] and Hermann’s
tortoise [Testudo hermanni]). J. Clin. Microbiol. 39, 3156–
3163.
Origgi, F.C., Klein, P.A., Tucker, S.J., Jacobson, E.R., 2003. Appli-
cation of immunoperoxidase-based techniques to detect herpes-
virus infection in tortoises. J. Vet. Diagn. Invest. 15, 133–
140.
Origgi, F.C., Romero, C.H., Bloom, D.C., Klein, P.A., Gaskin, J.M.,
Tucker, S.J., Jacobson, E.R., 2004. Experimental transmission of
a herpesvirus in Greek tortoises (Testudo graeca). Vet. Pathol.
41, 50–61.
Pettan-Brewer, K.C.B., Drew, M.L., Ramsay, E., Mohr, F.C., Low-
enstine, L.J., 1996. Herpesvirus particles associated with oral
and respiratory lesions in a California desert tortoise (Gopherus
agassizii). J. Wildl. Dis. 32, 521–526.
Schumacher, I.M., Brown, M.B., Jacobson, E.R., Collins, B.R.,
Klein, P.A., 1993. Detection of antibodies to a pathogenic
mycoplasma in desert tortoises (Gopherus agassizii) with
upper respiratory tract disease. J. Clin. Microbiol. 31, 1454–
1460.
Snipes, K.P., Biberstein, E.L., 1982.Pasteurella testudinis sp. nov.: a
parasite of desert tortoises. Int. J. Syst. Bacteriol. 32, 201–
210.
Teifke, J.P., Lohr, C.V., Marschang, R.E., Osterrieder, N., Posthaus,
H., 2000. Detection of chelonid herpesvirus DNA by nonra-
dioactive in situ hybridization in tissues from tortoises suffering
from stomatitis–rhinitis complex in Europe and North America.
Vet. Pathol. 37, 377–385.
Une, Y., Murakami, M., Uemura, K., Fugitani, H., Ishibashi, T.,
Nomura, Y., 2000. Polymerase chain reaction (PCR) for the

A.J. Johnson et al. / Veterinary Microbiology 111 (2005) 107–116116
detection of herpesvirus in tortoises. J. Vet. Med. Sci. 62, 905–
907.
United States Fish and Wildlife Service, 1994. Desert Tortoise
(Mojave Population) Recovery Plan. U.S. Fish and Wildlife
Service. Portland, Oregon, 73 pp.
VanDevanter, D.R., Warrener, P., Bennett, L., Schultz, E.R., Coulter,
S., Garber, R.L., Rose, T.M., 1996. Detection and analysis of
diverse herpesviral species by consensus primer PCR. J. Clin.
Microbiol. 34, 1666–1671.





























