Embed Size (px)
Citation preview

Oropharyngeal Morphology in the Basal TortoiseManouria emys emys With Comments on Form andFunction of the Testudinid Tongue
Egon Heiss,1* Nikolay Natchev,1 Thomas Schwaha,1 Dietmar Salaberger,2 Patrick Lemell,1
Christian Beisser1, and Josef Weisgram1
1Department of Theoretical Biology, Morphology Section, University of Vienna, Vienna, Austria2Upper Austria University of Applied Sciences, Campus Wels, Wels, Austria
ABSTRACT In tetrapods, the ability to ingest food onland is based on certain morphological features of theoropharynx in general and the feeding apparatus in par-ticular. Recent paleoecological studies imply that terres-trial feeding has evolved secondarily in turtles, so theyhad to meet the morphological oropharyngeal require-ments independently to other amniotes. This study isdesigned to improve our limited knowledge about theoropharyngeal morphology of tortoises by analyzing indetail the oropharynx in Manouria emys emys. Specialemphasis is placed on the form and function of thetongue. Even if Manouria is considered a basal memberof the only terrestrial turtle clade and was hypothesizedto have retained some features reflecting an aquaticancestry, Manouria shows oropharyngeal characteristicsfound in more derived testudinids. Accordingly, the oro-pharyngeal cavity in Manouria is richly structured andthe glands are large and complexly organized. Thetongue is large and fleshy and bears numerous slenderpapillae lacking lingual muscles. The hyolingual skele-ton is mainly cartilaginous, and the enlarged anteriorelements support the tongue and provide insertion sidesfor the well-developed lingual muscles, which showstriking differences to other reptiles. We conclude thatthe oropharyngeal design in Manouria differs clearlyfrom semiaquatic and aquatic turtles, as well as fromother reptilian sauropsids. J. Morphol. 272:1217–1229,2011. � 2011 Wiley-Liss, Inc.
KEY WORDS: turtle; Testudines; reptiles; oral cavity;tongue
INTRODUCTION
The oropharyngeal cavity in tetrapods isinvolved in a variety of functions, comprising ther-moregulation, oropharyngeal respiration, defense,mating, and perhaps most importantly, feeding(Druzisky and Brainerd, 2001). Feeding is crucialfor survival, and the contribution of feeding sys-tems to fitness is without any doubt substantial(Schwenk and Wagner, 2001). Studies on aquaticand terrestrial turtles showed a correlationbetween phylogeny, feeding mode, and oropharyn-geal morphology (Winokur, 1973; Weisgram et al.,1989; Iwasaki et al., 1996a,b; Lemell et al., 2000,
2002; Beisser et al., 2004; Richter et al., 2007;Heiss et al., 2008; Natchev et al., 2009). Purelyaquatic (nonmarine) turtles have a smooth oralsurface, poorly developed oropharyngeal glands, aflat palate, and a tiny tongue (Bramble and Wake,1985; Winokur, 1988; Weisgram et al., 1989;Beisser et al., 1995, 1998, 2001; Lemell and Weis-gram, 1997; Lemell et al., 2000, 2002, 2010). Sucha design appears advantageous for the hydrody-namic feeding mechanisms used by these turtles.In contrast, fully terrestrial turtles (‘‘tortoises’’:family Testudinidae) have an oral cavity rich instructural complexity (e.g. Winokur, 1988; Weis-gram et al., 1989), with epithelial flaps and ridgesaround the choanae, well-developed oropharyngealglands, and a large, papillose tongue ventrally(Wocheslander et al., 1999, 2000). All testudinidsstudied so far use their tongue as a preliminaryfood uptake organ (see Bels et al., 2008 for over-view). However, little information is available onthe morphology of the tortoise oropharynx in gen-eral and on form and function of the tongue in par-ticular. The first goal of the present study is there-fore to study the oropharynx of a testudinid turtlein detail, with special emphasis on the tongue.This is of particular interest if we accept that ter-restrial feeding evolved secondarily and independ-ently compared to other terrestrial amniotes.Accordingly, paleoecological studies indicate thatall extant turtle clades (Li et al., 2008; Anquetinet al., 2009), including the only terrestrial family,had an aquatic ancestor (Joyce and Gauthier,
Contract grant sponsor: Austrian Science Fund; Contract grantnumber: FWF P20094-B17.
*Correspondence to: Egon Heiss, Department of Theoretical Biol-ogy, Morphology Section, University of Vienna, Althanstr. 14, 1090Vienna, Austria. E-mail: [email protected]
Received 11 May 2010; Revised 17 March 2011;Accepted 21 March 2011
Published online 19 May 2011 inWiley Online Library (wileyonlinelibrary.com)DOI: 10.1002/jmor.10978
JOURNAL OF MORPHOLOGY 272:1217–1229 (2011)
� 2011 WILEY-LISS, INC.

2004). The genus Manouria is suggested to repre-sent the most ancestral (according to Gaffney andMeylan, 1988; Meylan and Sterrer, 2000; Takaha-shi et al., 2003; Spinks et al., 2004), or at least oneof the most basal (according to Le et al., 2006;Fritz and Bininda-Emonds, 2007), recent tortoises.The oropharyngeal design of this genus thereforemight retain certain ancestral features. The sec-ond goal of our study is to test this by comparingour results with those available for aquatic andsemiaquatic turtles.
MATERIALS AND METHODS
Manouria emys emys (Schlegel and Muller, 1844), the AsianForest Tortoise, ranges from southern Thailand through Malay-sia, Sumatra, and Borneo, where it preferably inhabits tropicalevergreen woodlands (Ernst and Barbour, 1989). M. emys emysprefers moist situations (Ernst and Barbour, 1989; Bonin et al.,2006) and is described as having a strong association to water(Høybye-Mortensen, 2004). The diet of these tortoises consistsof not only land and water plants, mushrooms, and insects, butalso amphibians and carrion (Bonin et al., 2006).We studied one adult and six juvenile M. emys emys. The adult
individual was provided by Mr. Richard Gemel, Museum of Nat-ural History Vienna and had been preserved in 70% ethanol(stock nr. 30.830, straight carapace length: 330 mm). The six cap-tive-bred juvenile animals were obtained commercially andranged in size (straight carapace length) between 109 and 135mm, and in weight between 234 and 265 g. They were kept intwo terraria with 150 cm 3 100 cm ground area at a 12-h darkand 12-h light cycle. The terraria contained bark mulch (5 cmhigh) as substrate, cork bark pieces as hiding places, and a lowbasin (40 cm 3 100 cm 3 7 cm) filled with water which was per-manently filtered by an external aquarium filter. The animalswere fed with a variety of vegetables, commercially available tor-toise pellets, fresh cattle heart, and dead mice. For morphologi-cal investigations, all six animals were anesthetized by intraper-itoneal injection of sodium pentobarbital and, after deep narco-sis, decapitated. The heads were immersed immediately in thefixation solutions described below.For scanning electron microscopy (SEM), the heads of two
juveniles were immersed for 24 h at room temperature inmodified Karnovsky solution (2.5% glutaraldehyde and 2%formaldehyde in 0.1 mol l21 cacodylate buffer; Karnovsky,1965). After rinsing in 0.1 mol l21 cacodylate buffer, the lowerjaw with the tongue was removed from the head to betterview both the ventral and dorsal surfaces of the oropharyngealcavity. Then, samples were postfixed in buffered 1% osmiumtetroxide for 2 h at 378C, washed in distilled water, andtreated with 25% HCl at 408C for 30 min to remove the mucusfrom the surface. After repeated rinsing in distilled water, thesamples were dehydrated in a graded ethanol and acetone se-ries and immersed in HMDS (hexamethyldisilazane) for 7days. Then, the HMDS was evaporated (see Oshel, 1997) andthe samples were mounted on aluminum stubs for SEM andcoated with gold in an AGAR B7340 Sputtercoater (Agar Sci-entific, Stansted, UK). Observations and digital photographicdocumentations were made using a Philips XL-20 scanningelectron microscope (Philips, Eindhoven, NL) and a PhilipsXL-30 environmental scanning electron microscope (Philips,Eindhoven, NL).To analyze the ‘‘in situ’’ 3D structure of the hyolingual skel-
eton, lCT (Micro Computed Tomography) scans were per-formed on two juvenile (two-year-old) and one adult (ca. 30-year-old) individuals. The juveniles were anesthetized asdescribed above, decapitated, and the heads were immersed inbuffered 4% formaldehyde solution for 3 weeks, changing thesolution once a week. The heads of the juvenile individualswere scanned using a Sub-lm-device Nanotom (Phonix|x-ray,
Wunstorf, GER). During measurement, projection images weregrabbed using an amorphous Silicon matrix detector at severalangular positions. After a full 3608 rotation, 1,500 images weregenerated. A mathematical algorithm calculated a 3D datasetusing the projection images. Gray values corresponding to thetissue density were assigned to each spatial element (voxel).The size of each voxel was 6 lm3. The adult specimen wasscanned by focusing on the head only, using a Viscom X8060NDT x-ray system (Viscom, Hanover, GER). After a full 3608scan, 610 images were generated and gray values correspondingto the tissue density were assigned to each voxel. The size ofeach voxel was 90 lm3.
For 3D reconstruction and visualization, the resulting gray-scale image stacks were imported into Amira 4.1 software (Mer-cury Computer Systems, Chelmsford, MA). For isolating thebony structures of the hyoid apparatus from the remainingskull, they were first roughly labeled. Afterward, gray-scale in-formation within the labels were separated from the originalgray-scale image stack with the Amira arithmetic tool [expres-sion: (a 55 1) * b, where ‘‘a’’ represents the Amira label fileand ‘‘1’’ is the Id of the labeled material; b is the original imagestack file]. Separating the skull from the hyoid was done in thesame way with labeling all but the hyoid bones. Surfaces of thebony structures were created with the IsoSurface tool of Amira.The cartilaginous parts of the hyoid were surface reconstructedby labeling them manually with the brush tool of the Amirasegmentation editor, followed by the generation of a surface.Surface optimizations were performed by iterated steps of trian-gle reduction and smoothing. Snapshots of the reconstructionswere taken with the Amira software.
For paraffin histology, two juvenile animals were used. Theheads were immersed in Bouin’s fixative (Romeis, 1989) for 30days, changing the solution twice a week. After complete fixa-tion and decalcification, the upper jaw was removed from therest of the head and the cornified rhamphothecae were cut off.Then, the samples were dehydrated in a graded ethanol–iso-propanol series and embedded in paraffin. After polymeriza-tion, 7 lm serial-sections were made on a Reichert-Jung 2030rotatory microtome (Reichert-Jung, Bensheim, GER). These se-ries included about 2,000 sections of the upper and 2,000 sec-tions of the lower jaw per animal. The sections were mountedon glass slides and, after deparaffination, stained with eitherhematoxylin (H) and eosin (E), periodic acid Schiff (PAS), Alcianblue (AB), or Coomassie Brilliant blue (after Romeis, 1989;Kiernan, 2003). As controls for the Coomassie Brilliant bluestaining, we used sections from Pleurodeles skin (for details,see Heiss et al., 2009). Histological slides were documented bydigital photography using a Nikon Eclipse E800 light micro-scope (Nikon, Tokyo, Japan).
RESULTSScanning Electron Microscopy
The ventral part of the oral cavity of M. emysemys is largely occupied by the elongated and well-developed tongue. The tongue is somewhat trian-gular with blunt apex and rounded posterolateralends (Figs. 1 and 2A). The glottis is positionedbetween the two posterolateral lingual ends (Figs.1 and 2A). Two elevations are present on the floorof the mouth, one on each side of the tongue; theseelevations contain slot-shaped openings of glands(Fig. 2A). The tongue itself bears numerous tallpapillae (Fig. 2A). These lingual papillae areshaped differently at different positions of thetongue. In the dorsal-most, anterior and middleportions of the tongue, they are slender, sharplypointed with a rough surface in the dorsal-most,
1218 E. HEISS ET AL.
Journal of Morphology

but, in the lateral and more posterior parts, tallwith a blunt apex and a smooth surface (Fig. 2A–C). Higher magnification reveals that the roughsurface of the slender papillae is due to numerouskeratinocytes; such keratinocytes are lacking inthe blunt-tipped papillae (compare Fig. 2B,C). Thekeratinized, sharply pointed papillae tend to pointposteriorly, an orientation that becomes graduallystronger toward the median line of the tongue(Fig. 2A).
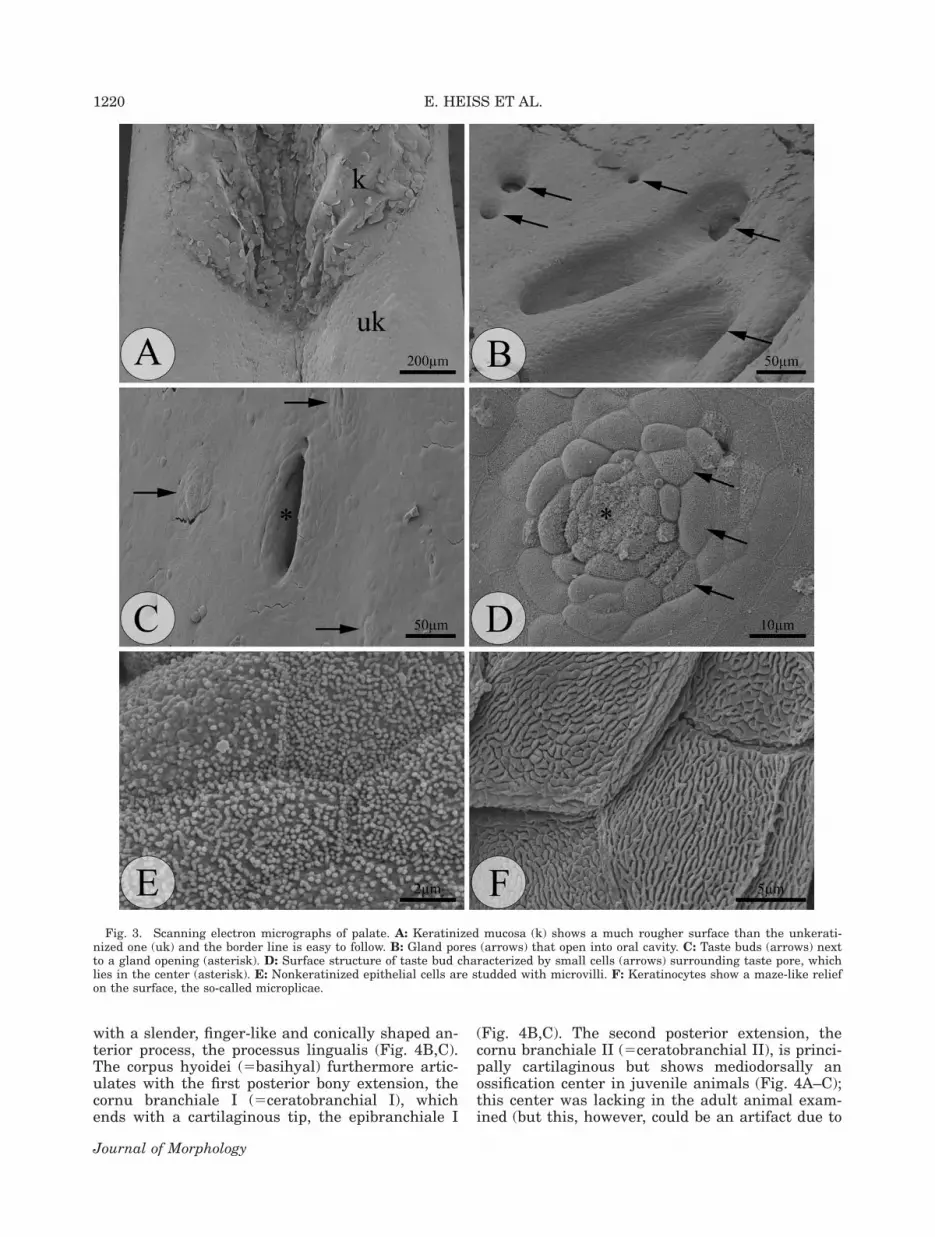
The surface of the palate is moderately struc-tured and bears the two large openings of thechoanae (see schematic drawing in Fig. 1). Highermagnification reveals that the whole anterior pala-
tal epithelium is keratinized (Figs. 1 and 3A). Ker-atinized epithelial cells exhibit maze-like micropli-cae on their surface (Fig. 3F), whereas nonkerati-nized cells are studded with microvilli (Fig. 3E).
Taste buds are detected on the oropharyngealsurface based on their typical concentricallyarranged small epithelial cells (Fig. 3D). The tastepore, with its characteristic field of microvilli, liesin the center of the taste bud (Fig. 3D). These mi-crovilli are larger and more numerous than those ofother epithelial cells. Taste buds are not homogene-ously distributed throughout the oropharyngealcavity. Most taste buds occur in the anterior-mostfloor of the mouth, but were also found on a smalllateral band along the tongue’s dorsolateral surface,as well as adjacent to and behind the tongue (seeFig. 1). On the palate, taste buds are present poste-riorly and on a small strip lateral to the choanae,in the very posterior interchoanal and postchoanalregions of the palate (Fig. 1). No taste buds werefound in the anterior-most, prechoanal palate, andon the anterior interchoanal part.
Computed Tomography
The hyolingual skeleton in M. emys emys is posi-tioned in the floor of the oropharyngeal cavity withtwo posterior extensions (branchial horns) thatrun dorsolaterally to the pharyngo-esophagealregion (Fig. 4A). The hyolingual skeleton consistsof a cartilaginous central body (corpus hyoidei)
Fig. 1. Schematic drawing showing the oral topography inM. emys emys. Top: palate, bottom: floor of mouth. Strongly ker-atinized regions of anterior palate and anterior half of tongueare visualized gray speckled. Oral glands are tagged in blue.Sublingual gland (asterisks) and glandula anguli oris (arrow-heads) are the largest oral secretory entities in M. emys emys.Smaller gland distribution is indicated by blue dots. Taste buds(small red dots) are not homogeneously distributed but re-stricted to certain regions. Horizontal lines indicate where his-tological sections (Figs. 6 and 7) were taken. Ad, adductor man-dibulae; Ch, choanae; Et, eustachian tube; G, glottis; Jj, jawjoint; Rh, rhamphothecae (horny beak); To, tongue.
Fig. 2. Scanning electron micrographs of floor of mouth.A: Overview showing large tongue (To), glottis (G), and bulgelateral to tongue bearing openings of sublingual gland (arrows).B: Detail of strongly keratinized lingual papillae. Note therough surface and pointed apex. C: Detail of nonkeratinized lin-gual papillae showing smooth surface and blunt apex.
OROPHARYNGEAL MORPHOLOGY IN M. emys 1219
Journal of Morphology
with a slender, finger-like and conically shaped an-terior process, the processus lingualis (Fig. 4B,C).The corpus hyoidei (5basihyal) furthermore artic-ulates with the first posterior bony extension, thecornu branchiale I (5ceratobranchial I), whichends with a cartilaginous tip, the epibranchiale I
(Fig. 4B,C). The second posterior extension, thecornu branchiale II (5ceratobranchial II), is princi-pally cartilaginous but shows mediodorsally anossification center in juvenile animals (Fig. 4A–C);this center was lacking in the adult animal exam-ined (but this, however, could be an artifact due to
Fig. 3. Scanning electron micrographs of palate. A: Keratinized mucosa (k) shows a much rougher surface than the unkerati-nized one (uk) and the border line is easy to follow. B: Gland pores (arrows) that open into oral cavity. C: Taste buds (arrows) nextto a gland opening (asterisk). D: Surface structure of taste bud characterized by small cells (arrows) surrounding taste pore, whichlies in the center (asterisk). E: Nonkeratinized epithelial cells are studded with microvilli. F: Keratinocytes show a maze-like reliefon the surface, the so-called microplicae.
1220 E. HEISS ET AL.
Journal of Morphology

the long preservation time). Anteriorly, immedi-ately ventral to the corpus hyoidei in the region ofthe processus lingualis lies the hypoglossum: a car-tilaginous element with one slender, finger-like an-terior and posterior process (Fig. 4A–C).
Light MicroscopyFloor of mouth. The floor of the mouth in M.
emys emys mainly consists of the oral mucosa, sup-ported by musculature and the hyolingual skele-ton. Here, some of the most important lingual
muscles will be described, mainly using the termi-nology proposed by Werneburg (2010). The mostimportant muscles supporting the floor of mouthdetected by histological serial sections are the M.geniohyoideus pars lateralis and the M. interman-dibularis. The M. geniohyoideus pars lateralis orig-inates laterally on the mandibular symphysis andruns rostromedially to finally insert on the proxi-mal one-third of the cornu branchiale I to thearticulation of cornu branchiale I and corpus hyoi-dei (Fig. 5A,B). The M. geniohyoideus pars lateralislies just dorsally to the M. intermandibularis (Fig.
Fig. 4. 3D reconstructions of the hyolingual skeleton in M. emys emys, based on lCT scans. Bone is colored golden, cartilage isvisualized blue and slightly transparent. A: Ventral view shows the ‘‘in situ’’ configuration of the hyolingual skeleton in relation toskull and lower jaw (De, dentale). The hyolingual skeleton consists of a central body, the corpus hyoidei (Chy), which articulatesposteriorly with the cornu branchiale I (CbI) and the cornu branchiale II (CbII). Anteroventrally to the corpus hyoidei lies thehypoglossum (Hy). B: Dorsal view on the hyolingual skeleton after virtually removing skull and lower jaw. The corpus hyoidei(Chy) shows anteriorly an elongated process, the processus lingualis (Pl), which is hidden ventrally by the hypoglossum (Hy). Notethe ossified cornu branchiale I (CbI) and the ossification center (O) on the cornu branchiale II (CbII). Such ossification centers werepresent only in juvenile individuals and were lacking in the adult. C: Lateral view; anterior is left. Processus lingualis (Pl) andhypoglossum (Hy) are prominent elements and support the tongue. CbI, cornu branchiale I; CbII, cornu branchiale II; Chy, corpushyoidei; Ep, epibranchiale I.
OROPHARYNGEAL MORPHOLOGY IN M. emys 1221
Journal of Morphology

6C–E). The M. intermandibularis runs transverselybetween the halves of the lower jaw just beneaththe integument of the throat. It originates mediallyfrom the dentary, and both controlateral parts areconnected by a medial aponeurosis (raphe medialis),which represents the insertion side.
One of the most striking structures in the floorof the mouth is the sublingual gland. It is fused to-gether to one functional entity immediately poste-rior to the mandibular symphysis opening into theoral cavity through a common duct (Fig. 6I). Moreposteriorly, the sublingual gland is divided intoone left and one right part that enclose the tongue
laterally (Fig. 6J). Both left and right partsslightly decrease in size posteriorly, finally disap-pearing as distinct glandular entities in the regionof the posterior end of the tongue. The sublingualgland consists of a variety of deep and complexlybranched and folded tubular glands with multipleslit-like openings to the floor of the mouth. Histo-chemically, the sublingual gland in M. emys emysshowed positive reaction to AB at pH 2.5 (for acidmucosubstances), PAS (for neutral mucosubstan-ces) or both AB and PAS (see Fig. 6I,J) but nega-tive reaction to Coomassie Brilliant blue (for pro-teins, see Fig. 6K) (controls reacted positively).
Fig. 5. Simplified schematic drawing illustrating the main lingual muscles in M. emys emys. The hyolingual skeleton is 3Dreconstructed based on lCT scans, and the muscle courses are based on the analysis of histological serial sections. A: Dorsal andB: lateral view; left 5 anterior; right 5 posterior. The M. genioglossus (1, white) originates from the dentaries (De), close to thesymphysis, and runs dorsally into the anterior tongue (To) as well as posterodorsally on the flank of the tongue. The M. hypoglosso-glossus (2, red) originates laterally on the very anterior to the middle portion of the hypoglossum (Hy). Anteriorly, the muscle fibersof the right and left portion of the M. hypoglossoglossus are separated and run straight upward to finally insert on the dorsal lin-gual mucosa. More posteriorly, the fibers of the right and left portion are curved to the midline, meet each other, and form an arch-like myostructure. The M. hypoglossohyoideus (3, green) originates laterally on the distal portion of the processus lingualis of thecorpus hyoidei (Chy) and runs caudoventrally to finally insert dorsally on the posterior hypoglossum. The M. hyoglossus (4, blue)originates on the very proximal part of the cornu branchiale I (CbI), lateral to the articulation to corpus hyoidei and runs antero-dorsally, dorsal to the hypoglossum, and is embedded between M. hypoglossoglossus, M. genioglossus, and dorsal lingual mucosa.The M. geniohyoideus pars lateralis (5, gray) originates laterally on the mandibular symphysis and runs rostromediad to finallyinsert on the proximal one-third of the cornu branchiale I to the articulation of cornu branchiale I and corpus hyoidei. CbII, cornubranchiale II.
1222 E. HEISS ET AL.
Journal of Morphology

Fig. 6. Light micrographs showing histological cross sections through different parts of the ventral oropharynx in M. emys emys.A–E: Sections through the tongue from the tip (A) to the hindtongue (E). For better orientation, the black horizontal lines in the sche-matic drawing of Figure 1 indicate where the sections were taken. Note the gradual differences in general tongue shape, structure ofthe lingual papillae (slender and keratinized anteriorly and broader and unkeratinized posteriorly), lingual glands, hyolingual skele-ton, and lingual muscles. Hy, hypoglossum; Lg, lingual glands; Pl, processus lingualis; Se, vertical septum; Sg, sublingual gland.Muscles: (1) M. genioglossus; (2) M. hypoglossoglossus; (3) M. hypoglossohyoideus; (4) M. hyoglossus; (5) M. geniohyoideus pars later-alis; (6) M. intermandibularis. F-G: Details showing structure of anterior, keratinized (F) and posterior, unkeratinized (G) lingualpapillae. H: Detail of the dorsolateral anterior tongue showing the connective tissue layer (Ct) that separates the lingual musclefibers (M) from the proximal-most portion of the lingual papillae. I: Anterior to the tongue, left and right portions of the sublingualgland are fused together and open through a common opening (asterisk). J: More posteriorly, left and right portions of the sublingualgland are separated and follow the course of the tongue laterally. The lingual glands (Lg) are relatively simple dorsally, and coiledand branched laterally. K: Lingual (Lg) and sublingual glands (Sg), just like all orophryngeal glands in M. emys emys, showed no pos-itive reaction when tested for proteinaceous content. L: Well-developed mucous glands were also present posterior to the tongue, onthe glottis. Ar, Cartilago arytaenoidea. Histological stains: (A–H) Azan; (I, J, and L) AB-PAS; (K) Coomassie Brilliant blue.
OROPHARYNGEAL MORPHOLOGY IN M. emys 1223
Journal of Morphology

When stained with both AB and PAS, the glandshowed a certain dichotomy, with both AB- andPAS-positive cells proximally and AB-positive cellsdistally (Fig. 6I,J).
Apart from the very anterior and lateral edges,close to the ventral rhamphotheca, no keratiniza-tion in the mucosa of the floor of mouth was foundin this species.
Tongue. The tongue shows a well-developedmusculature. The M. genioglossus originates fromthe dentary (lateral to the mandibular symphysis)and runs posterolaterally for almost the entirelength of the tongue. Anteriorly, the fibers of theM. genioglossus fan vertically and insert on thedorsal lingual mucosa of the tongue tip. Posteri-orly, the fibers become oriented more longitudi-nally and form the muscular sides of the tongue(Figs. 5A,B and 6A–E). The M. hypoglossoglossusoriginates laterally on the anterior to the middleportion of the hypoglossum. Anteriorly, the musclefibers of the right and left portion of the M. hypo-glossoglossus are separated by a median septumand run straight upward to finally insert on the dor-sal mucosa of the tongue (Figs. 5A,B and 6A). Moreposteriorly, the fibers of the right and left portionare curved to the midline, where they meet andform an arch-like myostructure. Some of the dorsal-most fibers of the M. hypoglossoglossus still inserton the dorsal mucosa of the tongue (Figs. 5A,B and6C,D). The M. hypoglossohyoideus originates later-ally on the distal portion of the processus lingualisof the corpus hyoidei and runs caudoventrally tofinally insert dorsally on the proximal hypoglossum(Figs. 5A,B and 6A–E). The M. hyoglossus originateson the very proximal part of the cornu branchiale-I,lateral to the articulation to the corpus hyoidei andruns anterodorsally, dorsal to the hypoglossum, lyingbetween M. hypoglossoglossus, M. genioglossus anddorsal lingual mucosa (Figs. 5A,B and 6A–E). In thearea of the distal-most part of the processus lingua-lis of the corpus hyoidei, the M. hyoglossus insertson the dorsal lingual mucosa (Fig. 6B). Even if thelingual musculature and muscles associated withthe tongue are well developed in M. emys emys, nomuscle fibers running into the lingual papillae couldbe observed (see Fig. 6F–H).
Lingual glands are well developed in M. emysemys. Secretory cells are mostly organized intoglandular entities, and single goblet cells are rare,but present. Simple straight and slightly branchedtubular glands were typically found on the dorsaltongue surface area, in the interface between thelingual papillae (Fig. 6A–E,J). Coiled and morecomplexly branched tubular glands are present onthe lateral parts of the tongue (Fig. 6A–E,J). His-tochemically, lingual glands in M. emys emysshowed the same reaction as the sublingual gland(see results above).
The lingual epithelium is keratinized only onthe lingual papillae in the anterior one-third of the
dorsal lingual surface area (see Figs. 1 and 6A–C,F). This keratin layer decreases in thicknessfrom 5–7 flattened keratinocytes anteriorly to 1–2keratinocytes more posteriorly (compare Fig.6F,G). Lingual taste buds in M. emys emys arerare and were found only on a narrow band later-ally (see Fig. 1). No taste buds are present on theanterior one-fourth of the tongue.
Palate. The palate in M. emys emys is structur-ally divided into prechoanal (i.e. anterior to thechoanae), choanal, and postchoanal (i.e. posteriorto the choanae) regions. The prechoanal palateconsists of a mucosal layer with prominent laminapropria that stabilizes the roof of the mouth andseparates the oral cavity from the nasal cavities(Fig. 7A,B). These nasal cavities open into the oro-pharyngeal cavity through the choanae (Fig. 7C).Posterior to the choanae, the extension of the lam-ina propria decreases distinctly, and the mucosa asa whole is supported by the bony roof of themouth. The prechoanal mucosa is keratinized,bearing a keratin layer consisting of 6–8 keratino-cytes anteriorly. This keratin layer graduallydecreases in extension posteriorly, finally disap-pearing in the choanal and postchoanal region,where keratinization is found only very laterally:in the interface between palate and dorsal rham-photheca.
Oral glands are completely absent in the ante-rior 90% of the prechoanal palate and first appearas relatively small, branched, and coiled tubularentities just anterolaterally to the choanae (Fig.7B). From there, glands increase in size and num-ber lateral to both choanae, finally forming a broadglandular field in the postchoanal region. The post-choanal glandular field is rather sparse just imme-diately behind the choanae, but increases in den-sity posteriorly (Fig. 7F,G). On the level of the pos-terior one-third of the choanae, a proper and verylarge glandular entity, the glandula anguli oris(Gao), was found (Fig. 7D,E). The Gao is deeplyembedded between the adductor muscle, eye, andthe connective tissue of the oral roof; it opensthrough a single duct into the oral cavity (Fig.7D). Histochemically, all palatine glands showedthe same reaction as the sublingual and lingualglands (see respective sections above).
Taste buds are completely absent from the pre-choanal palate, but appear lateral to the choanae,in the posterior one-fourth of the interchoanalregion and posterior to the choanae (Fig. 1).
DISCUSSION
Turtles show a great diversity regarding theirecology, they span the range from fully aquatic tofully terrestrial, with a rich range of intermediatevariations. This diversity is largely congruent withphylogeny and is reflected in structural specializa-tions of the oropharynx (Winokur, 1988; Weisgram
1224 E. HEISS ET AL.
Journal of Morphology

et al., 1989; Iwasaki, 2002; Iwasaki et al., 1996a,b;Beisser et al., 1995, 1998, 2001, 2004; Heiss et al.,2008). The family Testudinidae is the only turtleclade known to exclusively contain true terrestrialforms (Pritchard, 1979; Bonin et al., 2006; Belset al., 2008). Terrestriality in testudinids, however,is believed to be secondarily derived. According toJoyce and Gauthier (2004) the ancestor of all tes-tudinoids was an aquatic turtle. This would meanthat any adaptation to a terrestrial life, includingmanaging the biophysical demands of terrestrialfeeding, is secondarily and independently evolvedcompared to other terrestrial amniotes. Manouriais suggested to be basal to all other recent tor-toises (according to Gaffney and Meylan, 1988;
Meylan and Sterrer, 2000; Takahashi et al., 2003;Spinks et al., 2004), or to share this basal-mostposition with the sister genus Gopherus (Le et al.,2006; Fritz and Bininda-Emonds, 2007). Due to itsbasal position, it was hypothesized that oropharyn-geal morphology of Manouria might represent alink between semiaquatic and terrestrial turtles.However, the oropharyngeal morphology of thisturtle is well adapted to terrestrial food uptakeand shows significant similarities to other, morederived testudinids. The tongue is large and fleshy,studded with slender, and tall papillae that arekeratinized and sharply pointed anteriorly butunkeratinized with blunt apex posteriorly. Such arough anterior dorsal tongue surface increases fric-
Fig. 7. Light micrographs showing histological cross sections through different parts of the dorsal oropharynx in M. emys emys.A: The anterior prechoanal palate consists of lamina propria (Lp) and keratinized epithelium (Ep), which separate the oral fromthe nasal cavities (Nc). Orc, oral cavity. B: More posteriorly, close to the choanal openings, the thick keratin layer is graduallyreduced to a single keratinocyte-layer and the first glands appear laterally (arrows). Note the absence of a bony palate in the pre-choanal region (as shown in A and B). C: Both nasal cavities open to the oral cavity through the choanae (Choa). Glands wereabsent from the interchoanal region but were abundant lateral to the choanae (arrows). Bo, bony roof of oral cavity; Lp, laminapropria; Orc, oral cavity. D: Comparable region as in (C) but now showing the huge glandula anguli oris (Gao). The Gao is embed-ded between the eye (Eye), M. adductor mandibulae (Ad), and the connective tissue of the oral roof and opens to the oral cavity(Orc) through one single duct (arrowhead). E: Higher magnification of Gao. Note the large, branched glands opening into the com-mon duct (asterisk). The epithelium of the duct itself is covered by single AB-positive goblet cells. F: Posterior to the choanae, thenumber of oral glands increases to finally occupy most of the space between oropharyngeal epithelium and the bony roof (shown inG). The oropharyngeal cavity (Orphc) is here completely supported by the bony roof. Histological stainings: (A) hematoxylin–eosin;(B, C, E–G) AB–PAS–hematoxylin; (D) AB–hematoxylin.
OROPHARYNGEAL MORPHOLOGY IN M. emys 1225
Journal of Morphology

tion forces between tongue and food item, which isadvantageous when the tongue is used for preycapture and/or intraoral food transport on land(Bramble and Wake, 1985; Schwenk, 2000a). Theblunt and unkeratinized papillae posteriorly mayplay an important role in pharyngeal packing (i.e.,pushing the food bolus posteriorly to the pharynxfor swallowing; Smith, 1984; Schwenk, 2000a,b).Winokur (1988) reported lingual muscle fibers toenter the lingual papillae in the more derived tor-toises Xerobates and Geochelone. Such muscularpapillae could actively perform simple movements,improving the fine motor manipulation of thetongue as a whole. Manouria, however, lackedsuch muscular papillae. The dorsal lingual muscu-lature was clearly separated from the papillae bythick connective tissue. Such a feature is alsoknown for all aquatic and semiaquatic turtlesexamined. Further studies on the tongues of othertortoises are necessary to clarify if the lack of mus-cular papillae in Manouria represents an ancestraltrait shared with aquatic and semiaquatic turtles.Muscular papillae could represent a specializationto functional demands in Xerobates and Geoche-lone. The fact that muscle fibers entering lingualpapillae were found in Sphenodon (Schwenk, 1985,1988) and most lizards (Schwenk, 1988, 2000b;Herrel et al., 1998) suggests a possible connectionwith terrestrial lingual feeding.
The tongue in tortoises needs to be highly mova-ble and, in fact, it can be retracted backward orextended out of the mouth: an important abilityfor efficient food uptake and intraoral transport onland (Wocheslander et al., 1999, 2000). Accord-ingly, the hyolingual skeleton and associatedmuscles (including tongue muscles) show some dif-ferences to semiaquatic and especially, aquatic tur-tles. Although the principal components of the hyo-lingual skeleton are more or less the same in allturtles studied so far, extension and composition ofthe individual elements can vary considerably. Themain components are the central hyoid body,which articulates with the paired first and secondbranchial horns. Anteriorly, immediately beneaththe hyoid, lies the hypoglossum. The hypoglossumis an autapomorphic structure of turtles. To man-age the biophysical demands of food uptake under-water (i.e., compensatory suction feeding; see VanDamme and Aerts, 1997), aquatic turtles showenlarged posterior hyolingual elements, and ossifi-cation of the hyolingual skeleton as a whole is pre-dominant. This probably helps to handle the highforces induced by the massively developed muscu-lature which retracts the hyolingual complex. Ter-restrial turtles, in contrast, need a flexible supportfor the tongue and, generally, the anterior hyoidcomponents are enlarged and the hyolingual skele-ton remains mainly cartilaginous. Besides support,the hyolingual skeleton offers important insertionsides for the lingual musculature. The lingual
muscles are apparently present in most turtles (fora possible exception see Lemell et al., 2010), butlittle is known so far about their contribution tolingual movements.
The mobility of the tongue as a unit is widelycoupled to the movement of the hyolingual skele-ton as a whole (Schwenk, 2000a). The first step intongue protraction probably involves protraction ofthe entire hyolingual skeleton by contraction ofthe M. geniohyoideus which runs between hyolin-gual skeleton and dentaries. In a second step, wesuggest that contraction of the M. hypoglossohyoi-deus, which runs between anterior part of proces-sus lingualis and posterior hypoglossum, com-presses these skeletal structures relative to eachothers and causes rostral sliding of the hypoglos-sum relative to the processus lingualis of thehyoid, resulting in further tongue elongation. Thismovement is probably stabilized and further sup-ported by contraction of the medial and posteriorM. hypoglossoglossus. The M. hypoglossoglossusoriginates on the lateral parts of the hypoglossumand its fibers run vertically and are curved to themidline to form a muscular arch which encirclesboth processus lingualis and M. hypoglossohyoi-deus. If the arch-shaped hypoglossoglossus musclecontracts, apart from stabilizing hypoglossal slid-ing caused by the M. hyoglossus, it supports thismovement by pressurizing the lingual process tocause the hypoglossum to slide anteriorly alongthe processus lingualis. In a last step, intrinsictongue elongation might be caused by a hydro-static mechanism (Kier and Smith, 1985) involvinganterior fibers of both M. hypoglossoglossus andM. genioglossus.
For tongue retraction, we suggest that contrac-tion of the hyoglossus muscle, which runs betweentongue tip and proximal cornu branchiale I, causesretraction of the hypoglossum relative to processuslingualis and therefore increases tongue diameterand decreases tongue length. Further tongue re-traction is probably caused by retraction of thewhole hyolingual skeleton by the coracohyoideusmuscle which runs between hyolingual skeletonand shoulder girdle (not shown in this study).
The complicated spatial orientation of the lin-gual muscles in Manouria indicates that these ani-mals can reshape the form of the tongue in a com-plex manner. Manouria (and probably all testudi-nids) might use hydrostatic deformation not onlyto elongate the anterior tongue (as describedabove) but also to make localized shape changes(e.g., cupping or bulging).
Musculo-skeletal function in semiaquatic turtlesmay be similar to that in Manouria, resulting incomparable movement capabilities. The system as awhole, however, may be less adapted for terrestrialfeeding in semiaquatic forms. Such amphibious tur-tles are able to feed underwater, an ability lost inall testudinids (Bels et al., 2008; Natchev et al.,
1226 E. HEISS ET AL.
Journal of Morphology

2010). Fully aquatic turtles (at least those few stud-ied so far) seem to show all major lingual musclecomponents as well (for an overview see Werne-burg, 2010), but the usually very small and weaktongues in these forms seem to play a subordinaterole in their feeding mechanism, and tongue move-ments may therefore be limited (see Bramble andWake, 1985; Iwasaki et al., 1996b; Lemell et al.,2000, Beisser et al., 2001; Heiss et al., 2010).
Comparing the musculo-skeletal system in Man-ouria with that in Sphenodon (Schwenk, 1986)and iguanian lizards (e.g. Gnanamuthu, 1937;Schwenk, 1988, 2000b) reveals that some compo-nents are similar, whereas others are completelydifferent. As in turtles, the hyoid body in theselepidosaurians bears an anterior process and artic-ulates with branchial horns. Also, the courses ofthe M. genioglossus and M. hyoglossus are compa-rable in turtles, Sphenodon, and iguanian lizards.However, the hypoglossum and its associated mus-culature is a unique feature of turtles, makingdirect comparison of tongue structure and functionwith other tetrapods difficult.
Beside the hyolingual musculo-skeletal system,the design of oropharyngeal glands in turtlesappears to correlate highly with phylogeny, habitat(aquatic vs. semiaquatic vs. terrestrial) and diet(herbivory vs. carnivory; Fahrenholz, 1937; Kochva,1978; Winokur, 1988; Iwasaki, 2002; Beisser et al.,2004; Heiss et al., 2008; Natchev et al., 2009).Aquatic turtles (which are usually carnivorous toomnivorous) show simple single-celled glands intheir oral cavity (Nalavade and Varute, 1976; Wino-kur, 1988; Weisgram et al., 1989; Iwasaki, 1992;Iwasaki et al., 1996b; Beisser et al., 1995, 1998,2001). Terrestrial turtles (typically herbivorous toopportunistic) have prominent oral glands (Fahren-holz, 1937; Kochva, 1978; Winokur, 1988; Weisgramet al., 1989; Wocheslander et al., 1999). Theseglands were reported from the floor of the mouth(sublingual glands), the palate (palatal glands), thecorner of the mouth (Gao), the area around the glot-tis (laryngeal glands), and from the dorsal and lat-eral tongue, mostly in between the lingual papillae(lingual glands). The oropharyngeal glands in Man-ouria are very well developed and are similar inextent and complexity to those of more derived tes-tudinids (see Fahrenholz, 1937; Kochva, 1978,Winokur, 1988; Weisgram, 1989: Wocheslanderet al., 1999). In Manouria, these glands produceand store mucosubstances, mainly neutral andacidic mucopolysaccharides. Proteinaceous mate-rial, mainly in the form of proteins associated withcarbohydrates (i.e., proteoglycans and glycopro-teins), could also be present in some amount, butwere not detected by the test used in this study.From a functional point of view, the lubricating na-ture of mucus helps avoid dehydration of the oro-pharyngeal epithelium in a terrestrial environment(Fahrenholz, 1937). Mucus also improves terres-
trial food ingestion and oropharyngeal transport,because this viscous medium induces wet adhesionbetween tongue and food surfaces (i.e., makes thetongue ‘‘sticky’’; see Schwenk 2000b). Finally, mu-cus as a lubricating medium is fundamental forswallowing in a terrestrial environment (Schwenk,2000a). Aquatic turtles, in contrast, do not needlubrication or lingual adhesion and consequentlytheir oropharyngeal glands are poorly developed(Fahrenholz, 1937; Kochva, 1978; Iwasaki, 1992;Iwasaki et al., 1996b: Beisser, 1995, 1998, 2001).
Taste buds are the organs for taste receptionand are structurally very similar in all reptileorders, but their occurrence in the oropharynx canvary considerably. In contrast to snakes and vara-nid lizards, which lack lingual taste buds and haverelatively few even in the oral mucosa (seeSchwenk, 1985, 2000b; Young, 1997; Berkhoudtet al., 2001), or crocodiles who show taste budsscattered throughout the oropharynx (Bath, 1905;Putterill and Soley, 2003, 2004), most lepidosaursand turtles possess numerous taste buds, some-times in very high densities (Sphenodon: Schwenk,1986; iguanian squamates: Schwenk, 1985; turtles:Korte, 1980; Uchida, 1980; Beisser et al., 1998;Heiss et al., 2008). The taste buds are not ran-domly distributed in the oropharynx of Manouria,and earlier studies point to a correlation betweentaste bud distribution and feeding mode in turtles(Heiss et al., 2008). Our knowledge on the feedingmode and taste bud distribution in Manouria,however, does not allow a definitive interpretationof a correlation. In Manouria, only few taste budswere present on the tongue, and no taste budswere found on the foretongue. A kinematical study(Natchev et al., unpublished data) shows that,unlike all other testudinids studied so far, Manou-ria does not use its tongue to ingest food (tongueprehension), but obligatorily uses its jaws (jaw pre-hension). During tongue prehension, a mechanismobligatory in all other testudinids studied to date(see Weisgram et al., 1989; Wocheslander et al.,1999; Bels et al., 2008; Heiss et al., 2008), thetongue tip and anterior one-fourth of the dorsaltongue surface are the first areas to contact thepotential food item. A high taste bud density therecould have advantages in allowing a fast chemo-sensory feedback, helping to avoid further contactwith unpalatable, inedible or harmful food. Thelacking taste buds on the anterior, dorsal tonguein Manouria could therefore reflect the absence oftongue prehension in this species and represent aremnant of its aquatic ancestor. This, however, is ahypothesis that needs to be tested. This willrequire much more comparative data on feedingkinematics and taste bud distribution in other tes-tudinids and turtles in general.
In summary, even if M. emys emys is considereda very basal tortoise, this species shows morpho-logical characteristics also found in more derived
OROPHARYNGEAL MORPHOLOGY IN M. emys 1227
Journal of Morphology

members of the family. These characteristicsreflect the ecomorphological adaptation to a terres-trial trophic ecology. Consequently, the tongue hasgained a central role in feeding, similar to morederived tortoises, but also similar to Sphenodonand iguanian lizards. The morphofunctional designof the tongue in Manouria and more derived tor-toises, however, clearly differs from that in Sphe-nodon, iguanian lizards, or other reptiles.
ACKNOWLEDGMENTS
The authors acknowledge Ms. Daniela Gruberand Ms. Ulrike Hormann (both CIUS, Universityof Vienna) for friendly assistance in electron mi-croscopy, Dr. Michaela Gumpenberger (VeterinaryUniversity of Vienna) for conducting CT-experi-ments on our material, Mr. Richard Gemel (Mu-seum of Natural History, Vienna) for many impor-tant discussions on turtles and for kindly provid-ing a preserved adult M. emys emys for a CT-scankindly conducted by Prof. Gerhard Weber and Mr.Martin Dockner (both Department of Anthropol-ogy, University of Vienna), Prof. Vincent Bels andDr. Stephane Montuelle (both Museum of NaturalHistory, Paris) for friendly help in procuring thejuvenile animals, Dr. Michael Stachowitsch(Department of Marine Biology, University ofVienna) for improving our English, and Dr. IngmarWerneburg (Institution of Paleontology, Universityof Zurich), along with two anonymous reviewersfor helpful comments on the manuscript.
LITERATURE CITED
Anquetin J, Barrett PM, Jones MEH, Moore-Fay S, Evans SE.2009. A new stem turtle from the Middle Jurassic of Scotland:New insights into the evolution and palaeoecology of basalturtles. Proc R Soc B 276:879–886.
Bath W. 1905. Uber das Vorkommen von Geschmacksorganenin der Mundhohle von Crocodylus niloticus Laur. Zool Anz29:352–353.
Beisser CJ, Weisgram J, Splechtna H. 1995. Dorsal lingual epi-thelium of Platemys pallidipectoris (Pleurodira, Chelidae).J Morphol 226:267–276.
Beisser CJ, Weisgram J, Hilgers H, Splechtna H. 1998. Finestructure of the dorsal lingual epithelium of Trachemysscripta elegans (Chelonia: Emydidae). Anat Rec 250:127–135.
Beisser CJ, Lemell P, Weisgram J. 2001. Light and transmissionelectron microscopy of the dorsal lingual epithelium of Pelu-sios castaneus (Pleurodira, Chelidae) with special respect toits feeding mechanics. Tissue Cell 33:63–71.
Beisser CJ, Lemell P, Weisgram J. 2004. The dorsal lingual epi-thelium of Rhinoclemmys pulcherrima incisa (Chelonia, Cryp-todira). Anat Rec 277A:227–235.
Bels V, Baussart S, Davenport J, Shorten M, O’Riordan RM,Renous S, Davenport JL. 2008. Functional evolution of feedingbehavior in turtles. In: Wyneken J, Godfrey MH, Bels V, editors.Biology of Turtles. Boca Raton: CRC Press. pp 187–212.
Berkhoudt H, Wilson P, Young B. 2001. Taste buds in the pala-tal mucosa of snakes. Afr Zool 36:185–188.
Bonin F, Devaux B, Dupre A. 2006. Turtles of the world. Lon-don: A & C Black Publishers. 416 p.
Bramble DM, Wake DB. 1985. Feeding mechanisms of lowertetrapods. In: Hildebrand M, Bramble DM, Liem KF, Wake
DB, editors. Functional Vertebrate Morphology. Cambridge,MA: Harvard University Press. pp 230–261.
Druzisky KA, Brainerd EL. 2001. Buccal oscillation and lungventilation in a semi-aquatic turtle, Platysternon megacepha-lum. Zoology 104:143–152.
Ernst CH, Barbour RW. 1989. Turtles of the world. Washington,DC: Smithsonian Institution Press. 313 p.
Fahrenholz C. 1937. Drusen der Mundhohle. In: Bolk L, Gop-pert E, Kallius E, Lobosch W, editors. Handbuch der verglei-chenden Anatomie der Wirbeltiere. Berlin: Urban u. Schwar-zenberg. pp 115–155.
Fritz U, Bininda-Emonds ORP. 2007. When genes meet nomen-clature: Tortoise phylogeny and the shifting generic conceptsof Testudo and Geochelone. Zoology 110:298–307.
Gaffney ES, Meylan PA. 1988. A phylogeny of turtles. In: BentonNJ, editor. The Phylogeny and Classification of the Tetrapods,Vol. 1: Amphibian, Reptiles, Birds Systematics Association Spe-cial Volume No. 35A. Oxford: Clarendon Press. pp 157–219.
Gnanamuthu CP. 1937. Comparative study of the hyoid andtongue of some typical genera of reptiles. Proc Zool Soc Lond137B:1–63.
Heiss E, Plenk HJ, Weisgram J. 2008. Microanatomy of the palatalmucosa of the semiaquatic Malayan box turtle, Cuora amboinen-sis, and functional implications. Anat Rec 291:876–885.
Heiss E, Natchev N, Rabanser A, Weisgram J, Hilgers H. 2009.Three types of cutaneous glands in the skin of the salaman-drid Pleurodeles waltl. A histological and ultrastructuralstudy. J Morphol 270:892–902.
Heiss E, Natchev N, Beisser C, Lemell P, Weisgram J. 2010.The fish in the turtle: On the functionality of the oropharynxin the common musk turtle Sternotherus odoratus (Chelonia,Kinosternidae) concerning feeding and underwater respira-tion. Anat Rec 239:1416–1424.
Herrel A, Timmermans JP, De Vree F. 1998. Tongue flicking inagamid lizards: Morphology, kinematics, and muscle activitypatterns. Anat Rec 252:102–116.
Høybye-Mortensen K. 2004. The tortoise Manouria emys emys:Behaviour and habitat in the wild. [M.sc.]. Odense: Univer-sity of Southern Denmark. 100 p.
Iwasaki SI. 1992. Fine-structure of the dorsal epithelium of thetongue of the fresh-water turtle, Geoclemys reevesii (Chelonia,Emydinae). J Morphol 211:125–135.
Iwasaki SI, Asami T, Wanichanon C. 1996a. Fine structure ofthe dorsal lingual epithelium of the juvenile Hawksbill turtle,Eretmochelys imbricata bissa. Anat Rec 244:437–443.
Iwasaki SI, Asami T, Wanichanon C. 1996b. Ultrastructuralstudy of the dorsal lingual epithelium of the soft-shell turtle,Trionyx cartilagineus (Chelonia, Trionychidae). Anat Rec 246:305–316.
Iwasaki S. 2002. Evolution of the structure and function of thevertebrate tongue. J Anat 201:1–13.
Joyce WG, Gauthier JA. 2004. Palaeoecology of Triassic stemturtles sheds new light on turtle origins. Proc R Soc LondonSer B Biol 271:1–5.
Karnovsky MJ. 1965. A formaldehyde–glutaraldehyde fixativeof high osmolarity for use in electron microscopy. J Cell Biol27:137–138.
Kiernan JA. 2003. Histological and Histochemical Methods: Theoryand Practice. New York: Oxford University Press. 502 p.
Kier WM, Smith KK. 1985. Tongues, tentacles and trunks: Thebiomechanics of movement in muscular-hydrostats. Zool JLinn Soc 83:307–324.
Kochva E. 1978. Oral glands of the reptilia. In: Gans C, GansKA, editors. Biology of the Reptilia. London: Academic Press.pp 43–161.
Korte GE. 1980. Ultrastructure of the tastebuds of the red-earedturtle, Crysemys scripta elegans. J Morphol 163:231–252.
Le M, Raxworthy CJ, McCord WP, Mertz L. 2006. A molecu-lar phylogeny of tortoises (Testudines: Testudinidae) basedon mitochondrial and nuclear genes. Mol Phylogenet Evol40:517–531.
Lemell P, Weisgram J. 1997. Feeding patterns of Pelusios casta-neus (Chelonia: Pleurodira). Neth J Zool 47:429–441.
1228 E. HEISS ET AL.
Journal of Morphology

Lemell P, Beisser CJ, Weisgram J. 2000. Morphology and func-tion of the feeding apparatus of Pelusios castaneus (Chelonia;Pleurodira). J Morphol 244:127–135.
Lemell P, Lemell C, Snelderwaard P, Gumpenberger M, Woches-lander R, Weisgram J. 2002. Feeding patterns of Chelus fim-briatus (Pleurodira: Chelidae). J Exp Biol 205:1495–1506.
Lemell P, Beisser CJ, Gumpenberger M, Snelderwaard P, GemelR, Weisgram J. 2010. The feeding apparatus of Chelus fim-briatus (Pleurodira, Chelidae)—Adaptation perfected?Amphibia-Reptilia 31:97–107.
Li C, Wu XC, Rieppel O, Wang LT, Zhao LJ. 2008. An ancestralturtle from the Late Triassic of southwestern China. Nature456:497–501.
Meylan PA, Sterrer W. 2000. Hesperotestudo (Testudines: Testudi-nidae) from the Pleistocene of Bermuda, with comments on thephylogenetic position of the genus. Zool J Linn Soc 128:51–76.
Nalavade MN, Varute AT. 1976. Histochemical studies onmucins of vertebrate tongues. VIII. Histochemical analysis ofmucosubstances in the tongue of the turtle. Folia HistochemCytochem 14:123–134.
Natchev N, Heiss E, Lemell P, Stratev D, Weisgram J. 2009.Analysis of prey capture and food transport kinematics in twoAsian box turtles, Cuora amboinensis and Cuora flavomargi-nata (Chelonia, Geoemydidae), with emphasis on terrestrialfeeding patterns. Zoology 112:113–127.
Natchev N, Lemell P, Heiss E, Beisser C, Weisgram J. 2010.Aquatic feeding in a terrestrial turtle: A functional–morpho-logical study of the feeding apparatus in the Indochinese boxturtle Cuora galbinifrons (Testudines, Geoemydidae). Zoomor-phology 129:111–119.
Oshel P. 1997. HMDS and specimen drying for SEM. MicroscToday 5(4):16.
Pritchard PCH. 1979. Encyclopedia of turtles. Neptune, NJ:TFH Publications, Inc.895 p.
Putterill JF, Soley JT. 2003. General morphology of the oralcavity of the Nile crocodile, Crocodylus niloticus (Laurenti,1768). I. Palate and gingivae. Onderstepoort J Vet Res 70:281–297.
Putterill JF, Soley JT. 2004. General morphology of the oralcavity of the Nile crocodile, Crocodylus niloticus (Laurenti,1768). II. The tongue. Onderstepoort J Vet Res 71:263–277.
Richter S, Auer M, Fritz U. 2007. Variation of hyoid morphologyin geoemydid terrapins. Amphibia-Reptilia 28:148–153.
Romeis B. 1989. Mikroskopische Technik. In: Bock P, editor.Munchen, Wien, Baltimore: Urban u. Schwarzenberg.697 p.
Schlegel H, Muller S. 1844. Over de Schildpadden van den Indis-chen Archipel. In: Temminck V, editor. Verhandelingen over denatuurlijke geschiedenis der Nederlandsche overzeesche bezittin-gen, door de leden der Natuurkundige Commisie in Oost-Indie enandere schrijvers. Leiden: Leuchtmans u. Hoeck. pp 29–36.
Schwenk K. 1985. Occurrence, distribution and functional sig-nificance of taste buds in lizards. Copeia 1985:91–101.
Schwenk K. 1986. Morphology of the tongue in the tuatara,Sphenodon punctatus (Reptilia: Lepidosauria), with com-ments on function and phylogeny. J Morphol 188:129–156.
Schwenk K. 1988. Comparative morphology of the lepidosaurtongue and its relevance to squamate phylogeny. In: Estes R,Pregill G, editors. Phylogenetic Relationships of the LizardFamilies. Stanford: Stanford University Press. pp 569–598.
Schwenk K. 2000a. An introduction to tetrapod feeding. In:Schwenk K, editor. Feeding—Form, Function and Evolution inTetrapod Vertebrates. San Diego: Academic Press. pp 21–61.
Schwenk K. 2000b. Feeding in lepidosaurs. In: Schwenk K, edi-tor. Feeding—Form, Function and Evolution of Tetrapod Ver-tebrates. San Diego: Academic Press. pp 175–291.
Schwenk K, Wagner GP. 2001. Function and the evolution ofphenotypic stability: Connecting pattern to process. Am Zool41:552–563.
Smith KK. 1984. The use of the tongue and hyoid apparatusduring feeding in lizards (Ctenosaura similis and Tupinambisnigropunctatus). J Zool Lond 202:115–143.
Spinks PQ, Shaffer HB, Iverson JB, McCord WP. 2004. Phyloge-netic hypotheses for the turtle family Geoemydidae. Mol Phy-logenet Evol 32:164–182.
Takahashi A, Otsuka H, Hirayama R. 2003. A new species of thegenus Manouria (Testudines: Testudinidae) from the UpperPleistocene of the Ryukyu Islands. Jpn Pal Res 7:195–217.
Uchida T. 1980. Ultrastructural and histochemical studies onthe taste buds in some reptiles. Arch Histol Jpn 43:459–478.
Van Damme J, Aerts P. 1997. Kinematics and functional mor-phology of aquatic feeding in Australian snake-necked turtles(Pleurodira; Chelodina). J Morphol 233:113–125.
Weisgram J, Ditrich H, Splechtna H. 1989. Comparative func-tional anatomical study of the oral cavity in two turtle spe-cies. Plzen lek Sborn Suppl 59:117–122.
Werneburg I. 2010. Evolution and development of turtles: Orga-nogenesis and cranial musculature. Zurich: Scidinge Hall Ver-lag. 393 p.
Winokur RM. 1973. Adaptive modifications of buccal mucosaein turtles. Am Zool 13:1347–1348.
Winokur RM. 1988. The buccopharyngeal mucosa of the turtles(Testudines). J Morphol 196:33–52.
Wocheslander R, Hilgers H, Weisgram J. 1999. Feeding mecha-nism of Testudo hermanni boettgeri (Chelonia, Cryptodira).Neth J Zool 49:1–13.
Wocheslander R, Gumpenberger M, Weisgram J. 2000. Intraoralfood transport in Testudo hermanni (Chelonia, cryptodira)—Aradiographic video analysis. Neth J Zool 50:445–454.
Young BA. 1997. On the absence of taste buds in monitor liz-ards (Varanus) and snakes. J Herpetol 31:130–137.
OROPHARYNGEAL MORPHOLOGY IN M. emys 1229
Journal of Morphology





























