Embed Size (px)
Citation preview

P.S.Z.N.: Marine Ecology, 18 (3): 21 1-226 (1997) ( ' I 1997 Blackwell Wissenschafts-Verlag, Berlin ISSN 0 173-9565
Accepted: December 18. 1996
Horizontal Transfer of Matter by a Cave- Dwelling Mysid R . COMA', M . CAROLA', T. RIERA' & M . ZABALA'
' lnstitut de Ciencies del Mar (CSIC). Passeig Joan de Borbo sjn, E-08039 Barcelona.
'Laboratoire Arago U.R.A. C.N.R.S. 117, F-66650 Banyuls sur Mer, France. 1 Departament d'Ecologia, Focultat de Biologia. Universitat de Barcelona, Avda.
Spain (E-mail: coma(rr'icnl.csic.es).
Diagonal 645. E-08028 Barcelona, Spain.
With 4 figures and 7 tables
Key words: I imi in i~~ . s i . s .s/~chr~co/u. Mysidacea. marine caves. fecal pellet, egesta. N W Medi- terranean.
Abstract. The role of a cave-dwelling mysid population as a matter carrier was evaluated in a cave of the Medes Is. (NW Mediterranean) during 1988-89. Ilenrini~. .s is .spi~/rmco/i~ (LEINYER. 1963) is a greg- arious mysid whose swarms migrate daily from the inner end of the cave ~ where they remain during the day-to the exterior where they feed during the night. Fecal pellet composition. pellet egestion and pellet decomposition were measured in order to evaluate the transfer of matter by mysids. Based on the strong daily behavioural rhythms of these mysids. special importance was attached to the sampling frequency (every 2 h). In order to assess seasonal variability, four daily cycles were evaluated within a year. Iri situ incubations were carried out to determine changes on the egestion rates. morphology. weight and composition (AA. C : N ratios) of pellets in the course of a day. The number of pellets deposited inside the cave was estimated using containers distributed along the cave bottom by SCUBA divers.
Fecal pellets showed ;in amorphous composition. whereby diatom frustules, dinoflagellate loricae and coccolithoporids were very scarce. About 25% of the body weight were estimated to be egested daily as fecal pellets, suggesting a detritivorous feeding habit. The population oscillated seasonally between I and 12 millions. Individuals egested between 1.6 and 3.5 pellets a day into the cave. each averaging from 9.4 to 11.9 pg DW, 0.5 to 1.0 pg C and 0.046 to 0.27 pg N . Therefore, the population carried daily about 2 W 0 7 g DW POM. 2-21 g C and 0.5 2.7 g N from outside to inside the cave. Pellets decomposed very quickly; between 20 to 50% of both C and N were released from pellets in less than 2 h after egestion. Oxidation of pellets theoretically consumes an amount of oxygen which agrees with the high BOD values previously reported for this cave. Marine caves are generally viewed as strictly oligotrophic systems; dense mysid populations, however, could strongly modify the trophic relationships i n marine caves.
Problem
Marine caves are generally regarded as trophic-limited systems where reduced water turnover decimates planktonic communities (PALAU rt nl., 1991); this leads to low values of POM, POC, PON and Chl a, which, in turn, explains the striking
U. S. Copyright Clearance Center Code Statement: 0 173-9565/97/ 1 803-02 1 1 $14.00/0

112 C'OMA, C'AR0I .A. RII:RA & z A l l A l A
paucity of benthic fauna (Rmwi(;, 1981; F i c t i w 1989, 1990a,b, 1991u.b). Sur- prisingly, previous studies on a cave from the Medes Islands (Spain, NW Medi- terruneun), where both planktonic communities and benthic fauna exhibit the same pattern of disappearance (GILI c ~ t id.. 1986; PALACJ c t d.. 1991), showed that v;iIucs o f POM, POC, PON and Biological Oxygen Demand (BOD) are similar or eveti higher inside than outside the cave (ZABALA o f ( I / . , 1989). To date this phenomenon has not been satisfactorily explained.
In this cave, a dense population of the tnysid H ~ ~ ~ i i / i i j ~ . ~ i . s .sp/unc*okr (Lriuovti~. 1963) migrates horizontally in a predictable nycthemeral rhythm ( RIERA ot ( r / . , I99 I ) . During daylight hours the population remains densely packed i n the darkest areas of the cave. When night falls. a column of niysids nioves along the walls to the exterior (PASSI;LAIWE & BOCJKDILLON, 1985; R ~ E K A (it ( I / . . 1991 ). All night they search for food near the entrance, usually no farther than 100 in away (MACQIJAKT- MOIILIN & PATKITI, 1966). Before sunrise, the mysids return to the darkest areas of the cave.
As an undefined amount of matter is picked up a s food outside the c;ive and released a s fecal pellets inside. niysids act as carriers of matter from the outside t o the inside of the cave. Although laboratory studies of some components of I / . . s / w / i r n c d r r metabolism, such as respiration, excretion (GAUDY PI ( I / . , 1980) atid growth (GAUDY & GiJriKIN. 1979) have been reported. egesta has received less attention. However. CAKOLA Pt d. ( I 993) proposed and succcssl'ully tested ii fecal pellet collection method using H . .vpc/unc*o/rr to estimate egesta of gregarious. densely packed crustiiceans.
The remaining question is: is thc m o u n t ofmysid egesta high enough to account for the increase of POM. POC. PON and BOD measured inside the cave'? To solvc this problem we need to iinswer three questions: ( i ) what is the content of fecul pellets, ( i i ) how inany pellets can a mysid produce daily and ( i i i ) how many tnysids inhabit the cave?
High mobility, extreme gregariousness and acute rheotropism ;ire the three main features of H . . s / w / i r n c d r r behaviour which prevent the use of standard censusing methods (JONI'S, 1979; CHAPMAN, 1980). Marking and resanipling methods ( BAYLXY, 1951; MARGALIII:. 1983) are unrealistic for populations con- sisting of several million individuals. Catches with nets or oceanographic bottles hardly provide quantifiable saniples because of the extremely patchy distribution of H . .spr/u~ic~o/tr populations in time and space. Recently, R i m ci r r l . (1990) con1 pa red t wo methods t o es t i ma t c /I. 4 s p d i m ~ o / u pop ti I at i on s: ha ti Is wit h plankton nets towed by divers and collection of fecal pellets. They concluded that i n a closed site (a cave) inhabited by a dense, nionospecific population, net-based estimates were less reliable than the fecal pellet collection method. So. among indirect methods based on estimates of activity, fecal deposition activity seems to be the most adequate method when egesta is the object of the
In this paper we examined fecal pellet content. deposition and decomposition rate. Furthermore, we measured egesta seasonally over an annual cycle i n order to quantify pellet production and mysid population size. These processes allowed us to estimate the transf'er of matter by the mysids from outside to inside the cave and the importance of this apport to trophic-limited systems such as marine ca ve s .
study.

Mysid's transfer ol'matter 213
Material and Methods
1. The study site
The study site is located i n ii NW Mediterranean cave in Medes Islands (Catalonia. Spain). Sampling was done between l i l l 19x8 and summer 1989. The cave has been described in previous works (CilI.1 PI
d, . I9Xh; ZAEAIA c / d.. 19x4). I t ends in an air-lilled chamber (4 x 4 x 3 m) where mysid manipulations were conducted (Fig. I ) . The total volume of the cave has been estimated at around 7.56 m' (ZAHAI.A PI
d.. 19x4).
2. Pellet composition. C and N pellet release rate
Cryofractured pellets were checked for the presence of diatoms. coccolithophorids. remains of crus- tacean carapaces. o r any other morphologically identifiable liagnient with a Cambridge mod. 2000 Scanning Electron Microscope (SEM) a s used for qualitative feeding studies by HONJO & ROMAU (1978) and TLIHNI.K (1986).
To estimate both the C and N content of pellets and the c' and N release rates. groups of pellets were separated from inysids after a 2-h incubation and were reincubated irr .r i / ir for periods of I 2. I. 2. 3.4. X. 12. 24, 56 and 144 h. After each reincubation. over 70 pellets were selected with an M-X WILD stereomicroscope. placed on a calcinated G F C lilter and immediately submerged in liquid nitrogen. C and N contents of pellets were analysed with a Carlo Erba NA IS00 Analyser. using the standard contiguration for these determinations (KRISTENSI:N & ANIXHSEN. 1987).
3. Biometric survey
A biornetric survey was carried out at each incubation: several hundred individuals and pellets o f each class were recorded. The following dimensions were measured: ( I ) gut length, ( 2 ) gut diameter. ( 3 ) cephalothorax Icngth, (4) body area. (5) body perimeter. (6) niysid dry weight. (7) pellet length. (8) pellet diamcter. (9) pellet weight.
Cephalothoras length and diameter. gut length B S well iis length and diameter of pellets were mensured with i i n M-X WILD stereomicroscope equipped with a micrometric lens. Body area and perimeter were estimated by an Image Analyser System (IRAS).
Fig. I . Study area and methods of sampling. (a ~ b ) Location of the Meda Xica cave (arrcws). (c ) Collector distribution along the cave bottom. (d) Cross section o f the cave showing where collectors were placed (full star). where incubations were manipulated (full triangle) and carried o u t (empty star). (e) Detail of the incubation and (f) collection devices.

214 COMA. CAKOLA. RIFKA & ZAIIAI.A
Mysid size class frequency was obtained by sorting all individuals of the live subsamples from each season into six demographic cl~sses: juveniles. immature males, immature females. adult males, mature non-ovigerous females and ovigerous females. Pellets were classified into six discrete size classes by using length and diameter a s descriptors: large-thick. short-thick, thick-fragments. large-thin, short- thin and thin-fragments. Pellets and mysids were dried at 70 C for 48 h. then weighed with a precision balance (accuracy 1 jig). Length, diameter and weight for a standard pellet was estimated. Biometric parameters for mysids and pellets classes were published in CAROLA c / ol. (1993).
4. Daily rhythms of fecal pellet deposition inside the cave
The incubation procedure for pellet collection was carried out using the niethod described by CAKOLA ( ’I NI. (1993). Five replicates of each incubation were repeated every 2 h during the niysids’ stay i n the cave (from 0600 to 21 : O K in summer). Daily cycles of incubations were seasonally perfortned between fall 1988 and summer 19x0. Data from the summer daily cycle was previously published in a test o f the method (CAKOLA r i d., 1993). After incubation. niysids and pellets were classified as above and counted with an M-X WILD stereomicroscope.
5. Quantifying the mysid population
The number of niysids inhabiting the cave could be indirectly estimated hy counting the pellets collected from the cave during a selected period of time if the fecal deposition rate during this time is also known. Sixty-live collectors, identical in size to those used by CAKOLA (’I ul. (1993), were placed along the bottom of the cave. Five collectors were placed equidistantly (mean distance: 1.5 111) in 13 parallel transects from the entrance to the end of the cave (Fig. Ic). The highest collector density was i n the inner parts. where mysids remain throughout the day (Fig. Id). Closed collectors were placed the previous day at sunsct in order to avoid the resuspension of sediments by SCUBA divers during the operations. The following day. 2 h after sunrise. the covers of the collectors were carefully opened by ii
diver who pulled OR the cover by means of a I-m-long nylon string which was kept buoyant by a small piece of cork (Fig. I t ) . This procedure. which started at the inner end of the cave and ended at the exit, lasted 20 iiiin. Two hours later, the collectors were closed again by divers who moved slowly from the entrance t o the end of the cave. Pellets from the collectors were retained with pieces of plankton net ( S O pin aperture mesh) and immediately placed in 50 cm’ of prefiltered seawater with S‘Yn formalin solution. At the laboratory, pellets were counted with an M-X WILD stereomicroscope.
For calculations we assumed that 3 fragments correspond to one entire pellet. The number of inysids per collector was calculated by multiplying the number of pellets counted in the collector by the inverse of the calculated deposition rate. Then. mysid density per square meter was calculated by multiplying the number of inysids per collector by the inverse of the area of the collector opening (s = 56.7 cm’). Finally. these values were multiplied by the total area.
1. Biometric survey
Seasonal changes in the size-structure pattern of the mysid population are shown in Fig. 2 (see also Table I ) . Ovigerous females were present in every season, suggesting continuous reproduction throughout the year. Nevertheless, an impor- tant recruitment event in late spring is suggested by both the strong increase in the frequency of the juvenile class from spring (43%) to summer (71 YO) and by the seasonal change in the juvenile class weight due to the low summer value.

Mysid's transfer of matter 215
Fig. 2. Seasonal fluctuation of the population size structure of Hmiimysis ,spe/uni~olu. Body area ranges (IBAS measurements) of all individuals measured during this study (n = Il0,300).
Table 2 shows that thick-fragments were always the most frequent class (63 YO), followed by short-thick (27 YO) and large-thick pellets (6%). Large-thin pellets were found to be below 3 YO of total number. Short-thin pellets and thin-fragments were rare.
The characteristics of a standard pellet and mysid for each season were estimated based on the frequency and weight of each class (Tables 1 and 2).
2. Fecal pellet composition
The fecal pellets of H. speluncolu normally have a very thin peritrophic membrane (Fig 3a). Despite the high number of observations, this membrane was rarely found, suggesting that decomposition is rapid.
Broken fecal pellets, when observed with SEM, showed an amorphous com- position in which diatom frustules, dinoflagellate loricae and coccolithoporids were very scarce (Fig. 3 b 4 ) .
Amino acid analysis revealed that at least 14 different amino acids were present in pellets. The more abundant ones, in pmol AA*mg pellet-', were: SER (0.227). ASP (0. I ) , GLY (0.097) and GLU (0.092).
3. C and N release rates
The annual mean carbon and nitrogen content of pellets ranged from 0.40 to 1.44 p g C - pellet and 0.046 to 0.27 pg N - pellet-'. Values were always higher in winter than in summer (Fig. 4).
In winter, a very fast POC and PON release from pellets was detected, with

Tabl
e 1.
Sea
sona
l cha
nges
of p
aram
eter
s of
thre
e ag
e cl
asse
s of H
mii
ni.s
is sp
elut
icol
u an
d of
a s
tand
ard
mys
id: a
rea
[mm
']. d
ry w
eigh
t bg]
. fre
quen
cy a
nd g
ut le
ngth
km
]. S
tand
ard
devi
atio
n (S
D).
juve
nile
im
mat
ure
mat
ure
stan
dard
mys
id
acg
1.1
25.6
50
.3
fall
SD
0.27
6.
3 n avg
1.2
29.4
47
.5
win
ter
SD
0.3
7.3
n avg
1.1
25.6
42
.8
sprin
g SD
0.
3 6.
9 n av
g 0.
57
8.9
71.2
su
mm
er
SD
0.37
5.
8 n
2.03
68
.3
32.2
0.
36
12.1
2.1
72.1
32
.2
0.3
10.3
2.1
72.1
39
.5
0.3
10.3
73
77
.7
15.3
0.
37
13.1
4.52
24
5.8
17.5
1.
12
60.9
4.7
261.
7 20
.2
1.6
89.1
4.7
261.
7 17
.6
1.9
105.
8
5.2
307.
6 13
.5
1.95
11
5.3
~~~
7 - 77
.2
1758
5775
2.2
90.1
17
97
4682
2.1
85.6
18
45
2846
1.4
59.8
15
00
5771
s P

Mysid's transfer of matter 217
m m I=
s 0 N
0
9
m 01
142 m m
m v , 2 2 N
2 v, m
$9 =

218 ( 'OMA. ('AKOI A. KII KA & Z A I i A I A
Fig. 3 . Fecal pellets of / f w i i i i i r . s i , s . s / d r n i c d t r : (a ) Pellet with peritrophic niemhrane. Contents of a I'ecal pellet: ( h ) niaiiily amorphous material. (c) coccolitophorid and ( d ) dinol1agell;itc. Scale is 2.5 pin in :I
and h: 10 imi in c and d .
values ranging between 20 and 51 O h ) in 2 h , and 75-80% in 12 11 (Fig. 4). The C and N content showed two differentiated patterns depending on season. While i n winter the concentrittion o f C and N decreased with time, a surprising rise i n both C and N was detected in summer pellets (Fig. 4a-b). A similar pattern was described by FIctiEz (1989, 1991b) when studying glucide release from POM sinking in ;I
ciive. However, even though relative enrichment of organic matter by bacteria- mediated processes has been reported in marine pellets (J0HANNf:S & SATOMI. 1966), the observed absolute enrichment remains dificult to explain.
4. Daily cycles of fecal pellet deposition rates
In fall, winter and spring we found the same pattern of fecal pellet deposition rate with time (slope of all daily cycles were between -0.17 and -0.2X; Table 3 ) that was previously observed and described in summer by CAROLA et cil. (1993). After a short initial increase, deposition rates decrease with time to nearly zero.
Season affects the absolute rate of pellet production and also the timing of pellet production. Therefore, the accumulated number of pellets deposited daily inside the cave varied among seasons. The lowest accumulated pellet production per mysid and day was I .62 in fall iind the highest was 3.53 in spring (Table 3 ) .

Mysid's transfer o f matter 219
b 2l
E C
301
25-
20-
0 15- z
10-
5-
1.8-
1.6- - 1.4- - a, - 1.2-
Cj, 0.8- a 0 OA-
a) Q 1-
I
0.4-
0.24
H
time [min]
Fig. 4. Release of C and N and C : N ratio, of Hemin7j,si.s .spe/uttco/u fecal pellets during an in sit ir aerobic degradation on 29 December 1988 (13 'C, full triangle) and on 2 June 1989 (22 C, empty square). ( a ) N release in december (two trends, K = -0.01 and K = -0.00026) and June. (b) C release in December (two trends. K = -0.01 and K = - 0.0009) and June. (c) Evolution of the C : N ratio versus time.
S& ldw 1400~-0
time [rnin]
5. How many mysids inhabit the cave?
Mysid densities throughout the cave for each season are summarized in Table 4. This table is an idealized representation of the cave bottom, with mysid density being estimated from the number of pellets in the collectors and the deposition rate during that period (daily cycle). The collector experiments confirm visual observations on the number and position of mysid swarms. In fall and winter, the mysid population was concentrated in two semi-separated holes at the deepest end of the cave (maximurn density in both holes: 272 x lo3 mysids.m-2 = 68% total population). Conversely, in spring and summer, mysids occupied nearly the entire volume of the inner end of the cave and their number increased strongly (maximum density in both holes: 642 x 10' mysids.m-2 = 1.2% total population). The total number of mysids estimated to be inhabiting the cave were (in millions): one in fall, five in winter, twelve in spring and seven in summer.
Fewer pellets were found in collectors at the outer part of the cave. SCUBA- divers repeatedly stated that there were no mysids at the entrance during collections. The pellets found could have been egested during the preceding migrational move- ment (the return had terminated just 4 h before). Therefore, we assumed that such pellets were 'old' fragments resuspended by divers and omitted them from subsequent calculations.
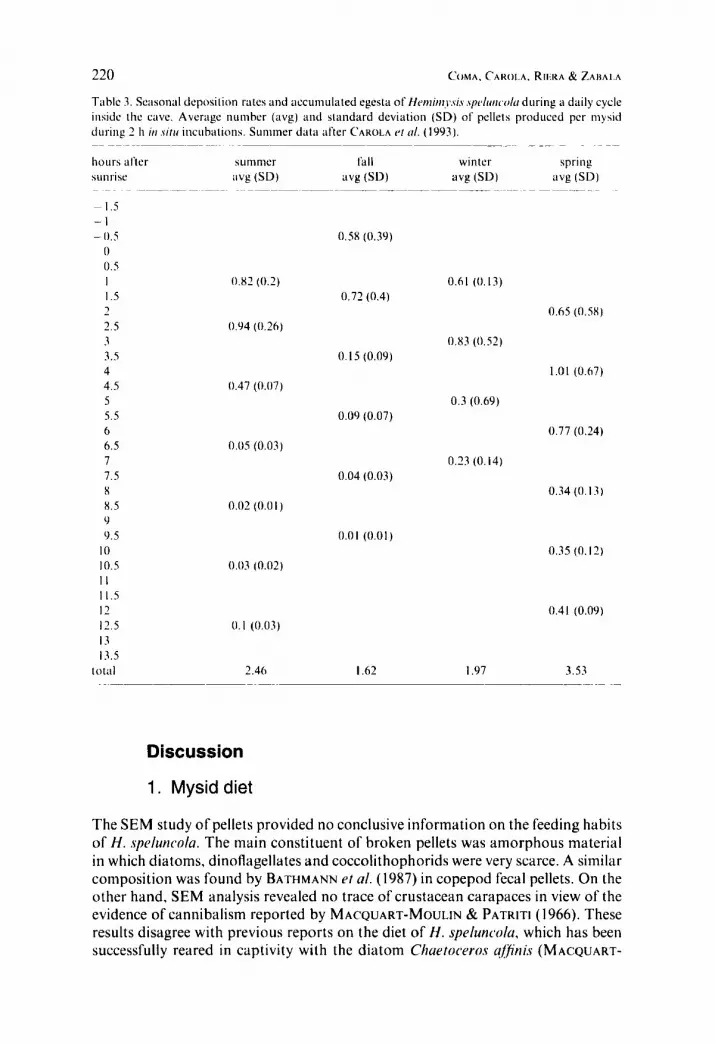
220 C(JMA. CAKOL.A. R i I ~ . i u & ZAHALA
Table 3. Seasonal deposition rates and accumulated egesta of H m b y ~ ~ s i . s . s p c h r r c ~ o / u d tiring a daily cycle inside the cave. Average number (avg) and standard deviation (SD) of pellets produced pcr niysid during 2 h i r i .vim incubations. Summer data after CAHOLA ('I c d . (1993).
hours after sumnier fall winter spring
~~ ~~ ~ _ _ ~ ~ _ _ _ _ _ ~ ~ ~~~
sunrise iivg (SD) avg (SD) avg (SD) avg (SD) ~ ~~~ ~ ~~ ~
~ 1,s -- I - 0.5 0.58 (0.39)
0 0 . S I 0.82 (0.2) 0.61 (0.13) I .5 0.72 (0.4) - 0.65 (0.M) 2.5 0.94 (0.26) 3 0.x3 (0.52) 3.5 0.15 (0.09) 4 1.01 (0.67) 4.5 0.47 (0.07) 5 0.3 (0.69) 5.5 0.09 (0.07) 6 0.77 (0.24) 6.5 0.0s (0.03) 7 0.23 (0.14) 7.5 0.04 (0.03) 8 0.34 (0.13) x.5 0.02 (0.0 I ) 9 9.5 0.01 (0.01)
I 0 0.35 (0.12) 10.5 0.03 (0.02) I I 11.5 I2 0.41 (0.09) 12.5 0.1 (0.03) 13 13.5
7
t o t d 2.46 I .62 I .97 3.53 ~ ~~ ~-
Discussion
1. Mysid diet
The SEM study of pellets provided no conclusive information on the feeding habits of H . speluncolu. The main constituent of broken pellets was amorphous material in which diatoms, dinoflagellates and coccolithophorids were very scarce. A similar composition was found by BATHMANN e f al. (1987) in copepod fecal pellets. On the other hand, SEM analysis revealed no trace of crustacean carapaces in view of the evidence of cannibalism reported by MACQUART-MOULIN & PATRITI (1 966). These results disagree with previous reports on the diet of H . speluncdu, which has been successfully reared in captivity with the diatom Chuetoceros u1fini.s (MACQUART-

3
5 2 2 9 2 2 2 Y 2.
c
Tab
le 4
. Sea
sona
l evo
lutio
n of
den
sity
and
spa
tial d
istr
ibut
ion
of th
e H
emir
nrsi
s spe
lunc
ola
popu
lati
on in
side
the
cave
. Thi
s ta
ble
is a
n id
ealiz
ed re
pres
enta
tion
of th
e ca
ve b
otto
m, w
here
the
colle
ctor
s w
ere
posi
tione
d. C
olle
ctor
s fr
om th
e ou
ter
half
(0-2
0 m
) of t
he c
ave
are
omitt
ed b
ecau
se th
ey n
ever
trap
ped
pelle
ts. (
d =
den
sity
of
Hen
iiniy
sis
spel
imco
lu in
10'
indi
v.. m
-' de
duce
d fr
om p
elle
t col
lect
ors.
s =
sur
face
[m']
of e
ach
colle
ctor
).
dist
ance
of
mou
th
fall
win
ter
spri
ng
sum
mer
[m
l
45
d:
24
32
89
63
63
12
78
58
-
12
34
5
12
34
5
12
34
5
12
34
5
-
S:
1.7
2.1
2.5
1.7
1.7
2.1
3.2
2.5
44
d:
9 20
4 13
60
48
14
48
56
40
28
52
41
d:
8 10
80
21
51
1 16
67
12
9 12
1 49
10
3
39
d:
272
135
23
26
78
210
147
133
487
321
73
67
94
72
36
d:
65
413
10
104
359
5 48
7 32
1 21
8 44
273
203
160
19
35
d:
1 23
1 19
36
2 35
34
4 78
16
3 I9
0
30
d:
10
12
27
71
61
181
176
25
128
25
d:
1 8
28
13
S:
2.8
2.2
2.2
2.8
2.6
2.x
2.2
2.2
2.8
2.6
2.8
S:
2.2
2.6
2.6
3.4
3.4
2.2
2.6
3.4
3.4
3.4
2.2
s:
2.6
3.1
2.6
4.2
2.6
4.2
3.1
2.8
1.4
2.6
4.2
3.1
2.8
1.4
s: 2.
2 2.
8 2.
4 2.
8 0.
8 2.
2 2.
8 2.
4 2.
8 0.
8 2.
2 2.
8 2.
4 2.
8 0.
8
S:
1.3
2.8
2.8
2.8
2.9
1.3
2.8
2.8
1.3
2.8
2.8
2.8
2.9
1.3
S:
9.3
15
7.4
3.0
15
7.4
3.0
13.4
15
9.
3 7.
4 3.
0
S:
6.9
6.5
3.1
3.7
3.7
6.9
6.5
3.1
1.9
1 22
3 00
0 5
120 0
00
12 2
72 0
00
7 28
0 00
0

222 COMA. CAKOLA. RIERA & ZAIMLA
MOULIN & PATRITI, 1966); and with a mixed diet composed ofcommercial Tetrainin and copepods of genus Tishe (GAUDY & GUERIN, 1979).
A mature mysid with an average weight of 270 pg DW (Table 1 ) egests 6 pellets per day, 3 inside and 3 outside the cave. If one pellet weighs 10.6 pg DW (Table 2), niysids move about 25% of their own weight daily, suggesting detritivorous feeding as observed in other niysids species (MAUCHLINE, 1980). The same con- clusion is suggested by changes in both the C and N content and the C : N ratio of pellets between summer and winter, which showed strong seasonal changes in pellet composition. The C content, N content and the C : N ratio in pellets showed the same trends as the C content, N content and C : N ratio of organic matter from caves of the Marseille region ( FICHEZ, 1989).
The fast POC and PON release from pellets (between 20%) and 50% in the first 2 h) agrees with fragmentation rates reported by CAROLA rf ul. (1993). suggesting a pellet half-life of about 2 h .
As amino acids and amino sugars represent the more labile fraction of organic N present in pellets, it is generally accepted that pellet decomposition raises the g1ucidic:amino acid ratio, and also the C : N ratio (MULLER & SUESS, 1979). However, these assumptions seem to be an oversimplification of several more complex processes (HARRISON & MANN, 1975; HARRISON, 1989). Differences i n the evolution of the C : N ratio with time in winter and summer can be due not only to the different water temperature, which accelerates the release of N and C in summer, but also due to the amount of N relative to that of C, the former being lower in summer than in winter.
The increase in the rnysid population from winter to spring was sudden. To what extent this spring bloom is due to reproduction or to immigration is difficult to answer. However, this pattern repeated in 1990 (RIBES rt a/.. 1996) and 1991 (pers. observ.), and literature data give many examples for similar patterns of spring recruitment in other mysids species (MAUCHLINE, 1980; SORBE, 1984; SAN VICENIX & SORRE. 1993).
2. Role of mysids in transporting matter
Various factors suggest that the rates of transport of matter by mysids we are going to evaluate here represent conservative estimates. On one hand, a portion of the small pellets produced by juvenile mysids were probably lost because of their short degradation time (CAROLA et d., 1993). Moreover, the pellets egested during migrational movements across the cave were not considered ( R l E R A rf d., I99 I ). On the other hand, C and N content per pellet could be underestimated because the time elapsed between incubation and the freezing of pellets implies carbon and nitrogen losses. This release during the first minutes after pellet egestion should be the most important for the cave metabolism because i t represents the most reactive fraction.
If, as summarized in Table 5, each one of the 1 to 12 millions H . spe/uncolr inhabiting the cave egested 1.6 to 3.5 pellets a day, each pellet averaging 9.4 to 1 I .9 pg DW, 0.5 to 1 p g carbon and 0.046 to 0.270 pg nitrogen. then about 20-407 g of DW POM. 2-21 g of carbon and 0.5-2.7 g of nitrogen were transferred daily from outer waters to the interior of the cave by the mysids. More than 50% of this POM

Mysid's transfer of matter 223
Table 5. Calculation of total POM. POC and PON carried daily into the cave by H ~ ~ n i n i I ~ . s i . s . s p h ~ i u ~ / ~ / . ( * ) Winter and summer values have been considered for spring and fall calculations. respectively.
~- ~ -~
winter spring summer fall
individuals [ x IO6] pellets. mysid ~ I .d ' POM [nig DW -pellet '1 carbon [mg. pellet '1 nitrogen [mg. pellet '1 POM carried daily into the cave [g DW] carbon carried daily into the cave [g DW] nitrogen carried daily into the cave [g] YO C and N content released within 12 h carbon released daily into the cave [g], within 12 h nitrogen released daily into the cave [g]. within 12 h -~ -. ~
5 12 2 3.5 9.4 9.7 I I* 0.27 0.27*
94 407 10 71 2.7 2. I
80 X0* 8 I6.X 2.2 I .h8
7.2 2.5
11.9 0.5 0.05
214 9 0.9
75 7.2 0.72
I .2 I .6
10.3 0.5* 0.05*
20 '7 - 0.5
7s* I .s 0.4
was finely fragmented, and between 20 and 50% of both C and N was released in less than 2 h after egestion.
The POC consumption in a marine cave from the adjoining region of Marseille was estimated by FICHEZ (1989). In that cave, where the mysid population is small (with respect to cave size) about 43 g POC and 7 g POC.m-"a-' are consumed by sediments from semi-dark and dark areas of the cave, respectively; about 5.7 g POC and 0.14 g POC.rn- '*a- ' are consumed by the water bodies confined in semi-dark and dark regions of the cave, respectively. As the accumulation of POM is negligible (FICHEZ, 1989), this consumption should be considered as a good estimate of 'passive' ( i .e . , other pathways than the mysid carrier system) import, which may also be assumed to be roughly proportional to the bed-surface and volume of the cave.
If values from FICHEZ ( 1 989) are applicable to our cave (756 in' volume; roughly 10% the size of that reported by FICHEZ, 1989), an estimate of the 'passive' import of organic matter from the outer waters could be made (Table 6). Comparing these estimates with the mysid related organic matter import, the latter accounts for 5- 52% of the passive POC input available for metabolic purposes. If related to the dark area only - where pellets are effectively deposited - this represents 134-1400 % of the expected passive POC consumption (Table 6 ) .
These results could also explain the strong disagreement between previous data from ZABALA et al. (1989) and data from FICHEZ (1989) on BOD measurements. FICHEZ (1989) suggested that the BOD estimates of waters from the inner cave (where mysids accumulate) were large overestimates, which surpassed by 1-2 orders of magnitude the values reported in the literature. Nevertheless, if 1.87 1 oxygen is required to oxidize 1 g carbon, and 80% of the egested carbon is assumed to be mineralized inside the cave (no POM accumulation was detected on the cave bottom), then 15 to 31.5 1 oxygen should be consumed daily in pellet oxidation (Table 7). Even if respiration of microbiota is neglected and if this consumption is distributed over the 120 m' total volume of the dark area, a daily demand of 0.125 to 0.26 ml 02.1-' (5.2 to 10.9 p1 02.1-' - h-') would result.
If BOD values measured from the outer waters of the cave, which range from 8.3 (winter) to 20.8 (spring) pI O2-l - l .h - ' (ZABALA et ul., 1989) are considered as

224 COMA, CAROLA, RIERA & ZAHALA
Table 6. (a) 'Passive' POC has been estimated from FICHEZ (1989). (b) Comparison between POC carried by mysids into the Meda Xica cave and POC metabolized ('passive' POC) by other causes in the cave. Mysid-mediated POC represents the average value measured in this study. (*) Estimation based on POC rates from Tremies cave
(a) Treinies cave (FICHE%. 1989)
P O C [ k g C * a '1 volume [m'] surface [m']
'passive' P O C [ g C . d - ' . m '1 POC [g Cad ' a m '1
(b) Meda Xica cave
volumc [ni'] surface [m']
'passive' [g C * d ~ '1 POC estimated*
semi-dark sediment water mass
36. I 37.6 ~ 6600
- 840
~ 0.015 0.118 ~
semi-dark sediment water mass
~~ 636 - 248
29.3 9.5
POC [g C - d '1 egested by mysids
dark sediment water mass
4.6 0.4
650 ~
- 2800
- 0.0004 0.019 -
dark sediment water mass - 120 76 ~
1.44 0.048
2-2 1
total
79.2 9400 I490
total
756 324
40.28
13
Table 7. Oxygen demand of the inner water of the cave-where mysids aggregate-as deduced from BOD measurements and combustion of pellets (*). Volume of inner cave water body: 120 000. (**) = 0,: C ratio in OM (organic matter) combustion = 1; O?: C ratio organic matter combustion = I . 87 l a g I .
__ -_
winter spring
averaged BOD5 in inner water [ml o,*l-'] 2.25 4.5 averaged BOD5 in exterior water 1 .o 2.5
daily excess of BOD of inner water body [I OZ]* daily excess of BOD 0.25 0.4
daily oxygen consumption for pellet combustion ( I 0,; only 80% of total C)** 31.5 30 48 15
'baseline', and they are compared with those from inner waters (from 9.3 pl O2. l - ' - h - ' in winter to 38.7 in spring; ZABALA cf al., 1989), a total daily BOD excess of 3 0 4 8 I O2 arises. Our estimates showed that pellet oxidation ( 1 5-3 1.5 I 0 2 * d - ' ) accounts for an important percentage of the daily excess BOD detected by ZABALA et al. (1989). The BOD values of the inner cave waters are high, on the order of those reported for some euphotic areas (POMEROY & JOHANNES, 1968; PACKARD rt al., 1971; PACKARD, 1979). Our results therefore show the importance of the transfer of matter inside the cave by the mysid population and the key role they play in the metabolism of the inhabited caves. If extreme oligotrophic conditions were the general rule in marine caves, a very different pattern can arise for those caves inhabited by large populations of mysids.

Mysid’s transfer of matter 225
Summary
Marine caves are generally regarded as trophic-limited systems in which reduced water turnover and light attenuation decimate the planktonic community. Under extreme oligotrophic conditions, the behaviour of certain organisms can play an important role in the general metabolism of the caves. This is the case in those caves inhabited by dense populations of mysids, which, act as active transfers of matter. This could strongly modify our understanding of the trophic relationships in marine caves.
Acknowledgements
We would like to acknowledge our colleagues in the Department of Ecology, University of Barcelona, for their help during this study, and particularly to M. RIBES, A. GARCIA, C. SAN V i c m m and I. PIBERNAT; to authorities from the ‘Direccio General de Pesca’ of the ‘Generalitat de Catalunya’ for giving us permission and facilities to work in the Medas Islands Marine Reserve; to Mr. R. ALGARRA, the guard of the reserve, and Mr. J. M. LLENAS for their help in our field work; to Dr. R. FONTARNAU and his colleagues at the ‘Servei de Microscopia Electronica’ for offering us all their expertise in SEM surveys and Mr. I. CASALS and Ms. P. FERNANDEZ for carrying out the analytical work. We are also grateful to J. SORBE. J. J. HARMELIN and two other anonymous reviewers for their help in improving this manuscript. This work has been supported by a grant from ‘La Caixa de Barcelona’.
References
BATHMANN, U. V., T. T. NOJI, M. Voss & R. PEINERT, 1987: Copepod fecal pellets: abundance, sedimentation and content at a permanent station in the Norwegian Sea in May/June 1986. Mar. Ecol. Prog. Ser., 38: 45-51,
BAYLEY, N. T. J., 1951: On estimating the size of mobile population from recapture data. Biometrika,
CAROLA, M., R. COMA, T. RIERA & M. ZABALA, 1993: Fecal pellet collection as a method for assessing egesta of the marine cave-dwelling mysid Hemimysis speluncolu. Sci. Mar., 57(1): 51-63.
CHAPMAN, C. J . , 1980: Ecology of juvenile and adult Nephrops. In: J. S. COBB & B. F. PHILLIPS (Eds.), The biology and management of lobsters, Vol. 2. Academic Press, New York: 143-180.
FICHEZ, R., 1989: Conditions d’oligotrophie en milieu aphotique, etude des grottes sous-marines, comparaison avec les milieux profonds et bilans energetiques. These de Doctorat de I’Universite d’Aix-Marseille 11; 251 pp.
- -, 1990 a: Les pigments chlorophylliens: indices d’oligotrophie dans les grottes sous-marines. C. R. Acad. Sci. Paris, 310(3): 155-161.
- -, 1990 b: Absence of redox potential discontinuity in dark submarine cave sediments as evidence of oligotrophic conditions. Estuarine Coastal Shelf Sci., 31(6): 875-881.
--, 1991 a: Benthic oxygen uptake and carbon cycling under aphotic and resource-limiting conditions in a submarine cave. Mar. Biol., llO(1): 137-143.
--, 1991 b: Suspended particulate organic matter in a Mediterranean submarine cave. Mar. Biol.,
GAUDY, R. & J. P. GUERiN, 1979: Ecophysiologie comparee des mysidaces Hemimjuis speluncolu LEDOYER (cavernicole) et Leptomysis lingoura G. 0. SARS (non cavernicole). Action de la tem- perature sur la croissance en elevage. J. Exp. Mar. Biol. Ecol., 38: 101-1 19.
--, J. P. GUERIN & M. PAGANO, 1980: Ecophysiologie comparee des mysidaces Hemimysis speluncolu LEDOYER (cavernicole) et Leptomysis linguuru G. 0. SARS (non cavernicole). Respiration et excretion. J. Exp. Mar. Biol. Ecol., 44: 2 9 4 6 .
GiLi, J. M., T. RIERA & M. ZABALA, 1986: Physical and biological gradients in a submarine cave on the Western Mediterranean coast (North East Spain). Mar. Biol., 90: 291-297.
HARRISON, P. G., 1989: Detrital processing in seagrass systems: a review of factors affecting decay rates. remineralization and detritivory. Aquatic Botany, 23: 263-288.
38: 293-306.
108(I): 167-174.

226 COMA, CAHOLA, RIIXA & ZA1lAI.A
~- & K . H. MANN. 1975: Chemical changes during the seasonal cycle of growth and decay in eelgrass
HONJO. S. & M. R. ROMAN. 1978: Marine copepod fecal pellets: production. preservation and sedi-
JOHANNES. R. E. & M. SATOMI, 1966: Composition and nutritive value of fecal pellets of a marine
JoNIIS. R.. 1979: Material and methods used in marking experiments in fishery research. F.A.O. Fishcries Technical Paper, 190; 134 pp.
KKISTBNSEN, E. & F. 0. ANIIERSIiN, 1987: Determination of organic carbon in marine sediments: il comparison of two CHN-analyzer methods. J . Exp. Mar. Biol. Ecol.. 109 15 23.
MACQUART-MOULIN. C. & G . PATRITI. 1966: Reniarque sur la biologie d'tlcniim,v.vis . s p c ~ / w r i ~ o / i r , LBDOYER. Mysidace sciaphile des grottes sous-niarines obscures de la region de Marseille. Recl Trav. Stn Mar. Endoume. 40(56): 253 258.
(Zo .y / iw murintr) on the Atlantic coast of Canada. J. Fish. Res. Board Can.. 32: 615-621.
mentation. J . Mar. Res., 3 6 45-57.
n. Limnol. Oceanogr., 2: 191-197.
MARGALEF. R.. 1983: Lininologia. Ed. Omega. Barcelona; 1010 pp. MAIICIILINE, J.. 1980: The Biology of Mysids and Euphausiids. Adv. Mar. Biol., I& I 681. MUIUEH, P. J . & J . P. SUESS, 1979: Productivity, sedimentation rate and sedimentary organic inattcr in
PACKAHI), T. T., 1979: Respiration and respiratory electron transport activity in plankton from the
, M. L. HEALY & F. A. RIC'IIAKIX, 1971: Vertical distribution of the activity of the respiralory
PALAU. M.. C. CORNET. T. RIERA & M. ZAFIALA. 1991: Planktonic gradients along a Mediterranean sea
PASSEI.AIGUI%. F. & A. BOUKI)II.LON, 1985: Les migrations nycthe-nieralcs du mysidacc cavernicole
POMI:HOY. H. M. & R. E. JOIIANNES, 1968: Occurrence and respiration of ultraplankton i n the upper
Rtiswic;, H. M.. 1981: Particulate organic carbon of bottom boundary and submarine cavern waters
R1~1.s. M.. R. COMA, M. Z A I ~ A L A & J . M. GILI. 1996: Small-scale spatial heterogeneity and se;imii;iI
thc oceans. 1. Organic carbon preservation. Deep-sea Res., 2 6 1347- 1369.
Northwest African upwelling area. J . Mar. Res.. 37(4): 71 I--742.
electron transport system in marine plankton. Limnol. Oceanogr.. 16: 6C70.
cave. Oecol. Aquat.. 10: 299-316.
I f iwi i t i j* . s i ,u . S / J P / U ~ I ~ ~ N LI~DOYER. C.I.E.S.M. 29(5): 157- 158.
500 m ol'the ocean. Deep-Sea Res., IS: 381 39 I .
of tropical coral reef. Mar. Ecol. Prog. Ser.. 5: 129 133.
variation in a population of a cave-dwelling Mediterranean niysid. J . Plankton Res.. l X ( 5 ) : 659 671.
RIERA. T., M. ZAIIALA & J . P s i W e ~ ~ s . 1991: Mysids from a submarine cave emerge each night to Iced. Sci. Mar.. SS(4): 605-609.
SAN VICINTI , C. & J. C. SOHIIC, 1993: Biologic du Mysidace suprabenthique littoral . S i , / i / , s / f ~ ~ i i ~ , . s ; . \ / ~ u i k c ~ i NOHMAN. 1892 dan le sud du golle de Gascogne (plage d'Hendaye). In: P. N o i i ~ (coord.). A C I L L ~ dc I? Premiere Conference Europcenne sur les Crustaces, Paris, 3 I aodt ~ 5 Septemhre, 1992 (r6sumL;s). Editions du Museum national d'Histoire naturelle, Paris: 137.
SOKHI.., J . C., 1984: Contribution d la connaisance des peuplcrnents suprabenthiclues ni.riiiques d u sud Gascogne. These d'Etat. Univ. Bordeaux; 265 pp.
TURNER. J . T., 1986: Zooplankton feeding ecology: contents oTfecal pellets of the Cyclopoid copepodus Oiiiuiw i'ctiu.s/u. Corj*i.uiw cr111iizonii~14.s. Oi//iotiu p / u t t i i / i w . and 0. . s i t t ip / i~ \ - from the northern C i t i l l of Mexico. P.S.Z.N.1: Marine Ecology, 7(4): 289-302.
ZAHALA, M., J. M. GII.I. T. RIEKA & M. F. HUELLIN. 1984: Estudio de 10s (actores Fisicos y biol6gico.; de una cueva submarina del litoral cataldn. I . Metodologia y resultados prcliminares. Proc. Acr;is I Simp. Iber. Est. Bentos Mar. I: 109- 121. . T. RIIIKA, J. M. GILI. M. BAKANGI~. A. LORO & J . PIRuL:L.As, 1989: Water flow. trophic dcpletion. and benthic macrofauna impoverishment in ii suhmarinc cave from the Western Mcditerranenn. P.S.Z.N.1: Marine Ecology. lO(3): 271 -287.























![Alpine Cave Techniques [EN]](https://img.dokumen.tips/doc/110x75/6352300816edf97a790cd5da/alpine-cave-techniques-en.jpg)





