Embed Size (px)
Citation preview

F3
JAa
3b
g
A3glatincbtcrslssngtrirsobtsdnnOvasrr
*EAB3sTpbp
Neuroscience 138 (2006) 1245–1263
0d
OREBRAIN PROJECTIONS OF TUBEROINFUNDIBULAR PEPTIDE OF
9 RESIDUES (TIP39)-CONTAINING SUBPARAFASCICULAR NEURONSKfa
TpoPe(tsac2fiTCstgstritcuian
cc(ltvccrgcrc
nt1
. WANG,a M. PALKOVITS,a,b T. B. USDINa*ND A. DOBOLYIa,b
Laboratory of Genetics, National Institute of Mental Health, Building5, Room 1B215, 35 Convent Drive, Bethesda, MD 20892-3728, USA
Laboratory of Neuromorphology, Semmelweis University and Hun-arian Academy of Sciences, Budapest, 1094, Tuzalto U. 58, Hungary
bstract—Neurons containing tuberoinfundibular peptide of9 residues (TIP39) constitute a rostro-caudally elongatedroup of cells in the posterior thalamus. These neurons are
ocated in the rostral part of the subparafascicular nucleusnd in the subparafascicular area, caudally. Projections ofhe caudally located TIP39 neurons have been previouslydentified by their disappearance following lesions. We haveow mapped the projections of the rat rostral subparafas-icular neurons using injections of the anterograde traceriotinylated dextran amine and the retrograde tracer choleraoxin B subunit, and confirmed the projections from moreaudal areas previously inferred from lesion studies. Neu-ons from both the rostral subparafascicular nucleus and theubparafascicular area project to the medial prefrontal, insu-ar, ecto- and perirhinal cortex, nucleus of the diagonal band,eptum, central and basomedial amygdaloid nuclei, fundustriati, basal forebrain, midline and intralaminar thalamicuclei, hypothalamus, subthalamus and the periaqueductalray. The subparafascicular area projects more densely tohe amygdala and the hypothalamus. In contrast, only theostral part of the subparafascicular nucleus projects signif-cantly to the superficial layers of prefrontal, insular, ecto-hinal and somatosensory cortical areas. Double labelinghowed that anterogradely labeled fibers from the rostral partf the subparafascicular nucleus contain TIP39 in many fore-rain areas, but do not in hypothalamic areas. Injections ofhe retrograde tracer cholera toxin B subunit into the lateraleptum and the fundus striati confirmed that they were in-eed target regions of both the rostral subparafascicularucleus and the subparafascicular area. In contrast, TIP39eurons did not project to the anterior hypothalamic nucleus.ur data provide an anatomical basis for the potential in-olvement of rostral subparafascicular neurons in limbicnd autonomic regulation, with TIP39 cells being majorubparafascicular output neurons projecting to forebrainegions. © 2005 IBRO. Published by Elsevier Ltd. All rightseserved.
Corresponding author. Tel: �1-301-402-6976; fax: �1-301-402-0245.-mail address: [email protected] (T. B. Usdin).bbreviations: ABC, avidin–biotin–horseradish peroxidase complex;DA, biotinylated dextran amine; CTB, cholera toxin B subunit; DAB,,3-diaminobenzidine; ir, immunoreactive; PBS, phosphate-bufferedaline at pH�7.4; PBS/T, phosphate-buffered saline containing 0.5%riton X-100; rSPF, rostral subparafascicular nucleus; SPA, sub-arafascicular area; SPF, subparafascicular nucleus; TB, 0.1 M Tris
buffer at pH�8.0; TH, tyrosine hydroxylase; TIP39, tuberoinfundibulareptide of 39 residues.
306-4522/06$30.00�0.00 © 2005 IBRO. Published by Elsevier Ltd. All rights reseroi:10.1016/j.neuroscience.2005.12.022
1245
ey words: immunohistochemistry, neuropeptide, tuberoin-undibular peptide of 39 residues, subparafascicular, neuro-natomical tract tracing, biotinylated dextran amine.
uberoinfundibular peptide of 39 residues (TIP39) wasurified from bovine hypothalamus based on its activationf the parathyroid hormone 2 receptor (Usdin et al., 1999).harmacological data support assignment of TIP39 as thendogenous ligand of the parathyroid hormone 2 receptorUsdin, 2000), which has a relatively widespread distribu-ion in the CNS (Wang et al., 2000). Functional studiesuggest that TIP39 has anxiolytic- and antidepressant-likections (LaBuda et al., 2004) and also that it affects someomponents of the hypothalamo-pituitary axis (Ward et al.,001; Sugimura et al., 2003). The distribution of TIP39bers has been described in detail (Dobolyi et al., 2003b).hey have a relatively widespread distribution in the ratNS including a high density of fibers in the limbic cortex,eptal, and amygdaloid regions, some midline and in-ralaminar thalamic nuclei, hypothalamus, periaqueductalray, parabrachial nuclei, brainstem auditory nuclei, andpinal cord (Dobolyi et al., 2003b). In contrast, TIP39 syn-hesizing cell bodies are almost entirely restricted to threeegions of the rat brain (Dobolyi et al., 2003b): 1) an arean the medial part of the posterior thalamus that includeshe subparafascicular nucleus (SPF) and the subparafas-icular area (SPA); 2) an area medial to the medial genic-late body in the lateral part of the posterior thalamus that
ncludes the posterior intralaminar nucleus of the thalamusnd the parvicellular SPF; and 3) the medial paralemniscalucleus in the lateral pons.
In the posterior thalamus TIP39 neurons form a rostro-audally elongated cell group. Anteriorly, these cells oc-upy the rostral half of the subparafascicular nucleusrSPF) and posteriorly, a portion of the SPA. The rSPF isocated within the thalamus, dorsal to the posterior hypo-halamic nucleus and medial to the parvicellular part of theentral posterior thalamic nucleus. The SPA includes theaudal part of the SPF and an area between the mostaudal part of the third ventricle and the fasciculus ret-oflexus. It is continuous caudally with the periaqueductalray of the midbrain (Moriizumi and Hattori, 1991). Inontrast to the rSPF, the caudal part of the SPF contains aelatively low number of TIP39 cells. At this level, TIP39ells are located more dorsomedially in the SPA.
The SPA received major attention when A11 dopami-ergic cells projecting to the spinal cord were identified inhe area (Bjorklund and Skagerberg, 1979; Hökfelt et al.,979; Skagerberg and Lindvall, 1985) and analgesia could
e elicited by stimulating the region (Yeung et al., 1977;ved.

R1Ltcea1igBv1et(inj
etfpTptTSdpftas
aaorttatgottdfiwtatntt
A
AtwA(Lpa
T
TE(rbwidlcLstcUfpw
lvsatfhga
H
SpPBtma
fnunmb
V
EptcSo
J. Wang et al. / Neuroscience 138 (2006) 1245–12631246
hodes and Liebeskind, 1978; Boivie and Meyerson,982; Hosobuchi, 1983; Fleetwood-Walker et al., 1988).abeled cells in the SPA were found following retrograderacer injections into several regions of the forebrain, in-luding the frontal cortex (Musil and Olson, 1988; Takadat al., 1988; Takada, 1993), the auditory cortex (Moriizumind Hattori, 1991), the amygdala (Ottersen and Ben-Ari,979; Peschanski and Mantyh, 1983; Takada, 1990; Mori-
zumi and Leduc-Cross, 1992; Takada, 1993), striatal re-ions (Peschanski and Mantyh, 1983; Beckstead, 1984;entivoglio and Molinari, 1984; Takada, 1993), and theentromedial nucleus of the hypothalamus (LeDoux et al.,985). The few studies using anterograde tract tracing toxamine the projections of this area only described projec-ions to the brainstem (Yasui et al., 1992) and amygdalaTurner and Herkenham, 1991). A lesioning study aimed atdentifying the projections of the recently discovered TIP39eurons in this area revealed much more extensive pro-
ections (Dobolyi et al., 2003a).Large lesions encompassing both the medial and lat-
ral areas at the diencephalon-midbrain junction that con-ain TIP39 cell bodies suggest that TIP39 fibers in theorebrain mostly originate in neurons of the SPA and thearvicellular SPF. These lesion studies also show thatIP39 fibers in the hindbrain mostly originate in the medialaralemniscal nucleus (Dobolyi et al., 2003a). The projec-
ions of the rSPF, in which there are a fairly high number ofIP39-containing neurons, have not yet been mapped.ince a number of axons from TIP39 neurons in the cau-ally located SPA as well as ascending sensory fibersass through the rSPF, electrolytic lesions are not optimalor identifying the projections from this area. Because ofhe proximity to the third ventricle excitotoxic lesions arelso problematic. Tract tracing methods may be betteruited for defining the projections from this area.
Investigation of the physiological functions of the SPFnd SPA is limited by the difficult identification of thenatomical borders of these regions, and by the absencef detailed anterograde studies of the projections of neu-ons located there. To better define the region of interest inhis study, we first examined the distribution of TIP39- andyrosine hydroxylase (TH) -containing neurons in the SPFnd SPA, because the A13 dopaminergic cell group is in
he proximity of the rSPF and the A11 dopaminergic cellroup is within the SPA. Next, we examined the projectionsf neurons in the rSPF using injections of the anterograde
racer biotinylated dextran amine (BDA) and comparedhem to projections of the SPA, with special attention to theistribution of TIP39-immunoreactive (ir) fibers. We per-
ormed double-labeling for BDA and TIP39 following BDAnjections into the SPF to identify which of the projectionsere from TIP39 neurons. We then injected retrograde
racer into the lateral septum, the fundus striati, and thenterior hypothalamic nucleus to confirm that they are
arget areas of the SPF and area, and that projectioneurons contain TIP39. The final goal of this study was
o complete the map of the TIP39 innervation pattern
hroughout the forebrain in rat. LEXPERIMENTAL PROCEDURES
nimals
nimal procedures were performed according to U.S. National Insti-utes of Health and National Institute of Mental Health guidelines andere approved by the National Institute of Mental Health Institutionalnimal Care and Use Committee. Adult male Sprague–Dawley rats
250–290 g in body weight) were purchased from Taconic Breedingaboratories and housed under standard laboratory conditions. Allossible efforts were made to minimize the number of animals usednd their suffering.
racer injections
he anterograde tracer BDA (10,000 MW, Molecular Probes,ugene, OR, USA) was targeted to the rSPF (n�25) and the SPA
n�8). Injections immediately dorsal, ventral and caudal to theSPF (n�10) served as controls. Rats anesthetized with pento-arbital (50 mg/kg, i.p.) were positioned in a stereotaxic apparatusith the incisor bar set at �3.3 mm. Glass micropipettes (10 �m
.d. for the rSPF and 15 �m i.d. for the SPA) filled with 10% BDAissolved in phosphate-buffered saline at pH�7.4 (PBS) were
owered to the targets of interest using the following stereotaxicoordinates (Paxinos and Watson, 1998): AP��3.8 mm,�0.5 mm from the midline, V�6.9 mm from the surface of thekull for the rSPF and AP��4.4 mm, L�0.4 mm, V�6.4 mm forhe SPA. BDA was injected by iontophoresis using a constanturrent source (BAB-350, Kation Scientific, Minneapolis, MN,SA) that delivered a current (�10 �A for the rSPF and �15 �A
or the SPA) which pulsed for 5 s on and 5 s off for 15 min. Theipette was left in place for 5 min with no current and thenithdrawn under negative current.
The retrograde tracer cholera toxin B subunit (CTB; List Bio-ogical Laboratories, Campbell, CA, USA) was injected into theentral part of the lateral septal nucleus (two animals), the fundustriati (two animals), and the anterior hypothalamic area (twonimals). The coordinates were: AP�0.0, L�1.0, V�7.4 mm forhe ventral lateral septum, AP��2.1, L�4.7, V�8.6 mm for theundus striati, and AP��1.6, L�0.9, V�9.8 mm for the anteriorypothalamic nucleus. CTB (0.25% in PBS) was delivered from alass micropipette (20 �m I.D.) by iontophoresis (�15 �A, 5 s onnd 5 s off for 20 min).
istology
even days after tracer injection, rats deeply anesthetized withentobarbital (80 mg/kg) were perfused transcardially with 200 mlBS followed by 300 ml of ice-cold 4% paraformaldehyde in PBS.rains were post-fixed in 4% paraformaldehyde for 24 h and then
ransferred to PBS. Coronal sections cut at 50 �m on a vibratingicrotome (Leica S700) were collected in PBS/0.1% sodium azidend stored at 4 °C.
Seven rats, which did not receive tracer injections, were per-used, and their brains were collected as described above. Coro-al sections from one animal were Nissl-stained with Thionin andsed as a reference for the identification and demonstration of theuclear groups. Coronal (three animals) and sagittal (three ani-als) brain sections were collected for TIP39 and TH immunola-eling and TIP39/TH dual labeling.
isualization of BDA
very fourth section was stained for BDA using either immuno-eroxidase or immunofluorescence procedures with or withoutyramide-mediated amplification. Sections were pretreated in PBSontaining 0.5% Triton X-100 (PBS/T) for 1 h at room temperature.ections were then incubated in avidin–biotin–horseradish per-xidase complex (ABC) at 1:500 (Vectastain ABC Elite kit, Vector
aboratories, Burlingame, CA, USA) for 2 h. Then the BDA-fibers
wSdc0spTFmpFf(DosUr
V
Ebidf
T
Afibhtacipcp
a(argnals
D
TsAP
tfs(wasi(t
n6tf
iiltS(eP
I
Tfphlb
D
TiwaawIuumP
cCBa
mtp3
T
Tds(thsjtbcsaia
J. Wang et al. / Neuroscience 138 (2006) 1245–1263 1247
ere visualized using 0.02% 3,3-diaminobenzidine (DAB; Sigma,t. Louis, MO, USA), 0.08% nickel (II) sulfate and 0.0012% hy-rogen peroxide in PBS. Alternatively, for biotin tyramide amplifi-ation, sections were rinsed extensively first in PBS and then in.1 M Tris at pH�8.0 (TB) after the first ABC incubation. Next,ections were placed in biotin–tyramide solution (1:1000) pre-ared in TB containing 0.0012% hydrogen peroxide for 20 min.hen a second ABC incubation was conducted at 1:1000 for 1 h.inally, the reaction product was visualized by incubation for 10in in DAB solution as described above. For FITC-tyramide am-lification, after the ABC reaction sections were incubated inITC-tyramide (1:10,000) and 0.0012% hydrogen peroxide in TB
or 10 min. All sections were mounted on positively charged slidesSuperfrost Plus; Fisher Scientific, Pittsburgh, PA, USA). MostAB sections were counterstained with Nuclear Red (Vector Lab-ratories), and all DAB sections were dehydrated and cover-lipped with Cytoseal 60 (Stephens Scientific, Riverdale, NJ,SA). Fluorescent sections were mounted using Prolong Antifade
eagent (Molecular Probes).
isualization of CTB
very second brain section from CTB-injected animals was incu-ated overnight in goat anti-CTB antibody (1:30,000; List Biolog-
cal Laboratories) at room temperature and then in Alexa 594onkey anti-goat secondary antibody (1:400; Molecular Probes)or 1 h.
IP39 and TH immunocytochemistry
previously characterized (Dobolyi et al., 2003a,b) affinity-puri-ed rabbit polyclonal antibody to rat TIP39, which labels cellodies with exactly the same distribution as observed by in situybridization histochemistry and which can be absorbed with syn-hetic TIP39 (Dobolyi et al., 2003b) was used in this study. Thentibody against TH (mouse monoclonal anti-TH, Novus Biologi-als, Littleton, CO, USA) provided the distribution well establishedn the literature (Hökfelt et al., 1976) in this study and also inrevious studies (Guo et al., 2002; Nocjar et al., 2002). Immuno-ytochemical controls were conducted with the omission of therimary antibody throughout the study.
Brain sections were pretreated in PBS/T/1% bovine serumlbumin for 1 h followed by incubation with TIP39 primary antibody1:2000 for tyramide amplification and 1:400 for DAB reaction) ornti-TH (1:1000) in PBS/T/1% bovine serum albumin for 48 h atoom temperature. Sections were then incubated in biotin-conju-ated donkey anti-rabbit secondary IgG at 1:600 (Jackson Immu-oresearch, West Grove, PA, USA) or biotin-conjugated donkeynti-mouse IgG at 1:600 (Jackson Immunoresearch) for 1 h, fol-
owed by incubation in ABC and DAB or FITC-tyramide as de-cribed above for the visualization of BDA.
ouble labeling
IP39/TH double labeling on sagittal sections was done as foringle labeling except that TIP39 was visualized by incubation inlexa Fluor 594 anti-rabbit secondary antibody (1:400; Molecularrobes) for 2 h after the labeling of TH with FITC-tyramide.
For BDA/TIP39 and BDA/TH colocalization every fourth sec-ion was labeled. TIP39 or TH immunocytochemistry was per-ormed first and then followed by visualization of BDA. Briefly,ections were pretreated in PBS/T/3% normal donkey serumVector Laboratories) for 1 h. The sections were then incubatedith primary antibody against TIP39 (1:1000) or with primaryntibody against TH (1:800) for 48 h in PBS/T/3% normal donkeyerum. Following thorough rinses in PBS, sections were incubatedn polymerized peroxidase anti-rabbit or anti-mouse antibodyImmPress Reagent, Vector Laboratories) for 30 min, then rinsed
hree times in PBS and three times in TB. TIP39-, or TH-immu- poreactivity was visualized using FITC-tyramide at 1:10,000 formin. Subsequently, BDA was visualized by incubating the sec-
ions in Alexa Fluor 594 streptavidin at 1:500 (Molecular Probes)or 2 h at 37 °C.
For CTB/TIP39 and CTB/TH double labeling, TIP39 or THmmunocytochemistry was performed first. TIP39 immunoreactiv-ty was visualized with FITC-tyramide as described for singleabeling. TH immunoreactivity was visualized by a 2-hour incuba-ion of sections in Alexa Fluro 488 (1:400; Molecular Probes).ubsequently, sections were incubated in goat anti-CTB antibody
1:30,000; List Biological Laboratories) overnight and then in Al-xa Fluor 594 donkey anti-goat antibody (1:400; Molecularrobes) for 1 h.
n situ hybridization histochemistry
wo rat brains were removed, and the fresh tissue was quicklyrozen on dry ice. Coronal sections were cut at 12 �m, mounted onositively charged slides, dried, and stored at �80 °C. In situybridization was performed as previously described with a 35S-
abeled antisense riboprobe to rat TIP39 that labels the same cellodies as the antibody to TIP39 (Dobolyi et al., 2003a,b).
ata analysis
he locations of the injection sites were plotted onto correspond-ng regions of a rat atlas (Paxinos and Watson, 1998). Animalsith injection sites centered in the rSPF or the SPA, or that haddjacent but non-overlapping injection sites were included in thenalysis. Images were captured at 1300�1030 pixel resolutionith a Photometrix CoolSnap Fx digital camera on an Olympus
X70 light microscope equipped with fluorescent epi-illuminationsing 4� objective. The individual pictures were then montagedsing Photoshop CS 8.0. Drawings were prepared by aligning theontaged pictures with corresponding schematics adapted fromaxinos and Watson (2005).
Confocal images were acquired with a Zeiss LSM 510 confo-al microscope using a 10� objective for the CTB/TIP39 andTB/TH studies and a 63� objective for the BDA/TIP39 andDA/TH studies. The presented confocal images are of single 10nd 0.8 �m optical layers, respectively.
Contrast was adjusted using the “levels” and the “unsharpask” commands in Adobe Photoshop. Full resolution was main-
ained until the micrographs were cropped and assembled forrinting, at which point images were adjusted to a resolution of00 dpi.
RESULTS
opography of the SPF and the SPA
he SPF is a cytoarchitectonically well-defined rostro-cau-ally elongated nucleus, with a circular profile in coronalections, in the ventromedial part of the posterior thalamusFig. 1). The rostral part, which comprises almost half ofhe nucleus, is embedded in the posterior thalamic andypothalamic nuclei (Fig. 1A, B), while the caudal part isituated within the SPA at the diencephalon–midbrain
unction (Fig. 1C, D). No obvious landmarks separatehese two parts. The rostral part of the nucleus is borderedy the posterior hypothalamic nucleus ventrally, the parvi-ellular part of the ventral posterior thalamic nucleus, apecific gustatory relay thalamic nucleus, dorsolaterallynd the central medial thalamic nucleus dorsally. Rostrally,
t is clearly separated from the reuniens thalamic nucleusnd the compact A13 dopaminergic cell group. The caudal
art of the nucleus extends into the SPA and occupies a
srcvs
utvr
Fm oronal sA 00 �m.
AaAAAAABcCCCDffFGHIIIILLL
J. Wang et al. / Neuroscience 138 (2006) 1245–12631248
mall region just ventral and ventromedial to the fasciculusetroflexus. Here, the SPF is connected to a laterally andaudally extending group of small neurons, called the par-icellular SPF. Caudally, the SPF is bordered by the inter-titial nucleus of Cajal.
ig. 1. Coronal view of the rostral part of the subparafascicular thalodified from Paxinos and Watson (1998). (B, D) Thionin-stained crrowheads indicate the SPF, with its caudal end in D. Scale bars�2
Abbreviation
A anterior amygdaloid areac anterior commissurecbS nucleus accumbens, shell portionH anterior hypothalamic nucleusMY amygdala11 A11 dopaminergic cell group13 A13 dopaminergic cell groupST bed nucleus of the stria terminalisc corpus callosume central nucleus of the amygdalaM central medial thalamic nucleusP caudate-putamenM dorsomedial nucleus of the hypothalamus
fornixr fasciculus retroflexusS fundus striatiP globus pallidus
hippocampusC inferior colliculusL infralimbic cortexns insular cortexPF interpeduncular fossaH lateral hypothalamic areaS lateral septal nucleus
V lateral ventricleBoth parts of the SPF contain heterogeneous cell pop-lations. We found TIP39- as well as TH-ir cell bodies inhe rSPF (Fig. 2A–C), as well as in the SPA along the thirdentricle, an area between the midline and the fasciculusetroflexus (Fig. 2D–F), but relatively few TIP39 cells were
leus (A and B) and the SPA (C and D). (A, C) Schematic drawingsections illustrate the topography of the rSPF (B) and the SPA (D).
the figures
medial amygdaloid nucleusmedial lemniscus
A medial preoptic areamamillothalamic tractoptic tract
G periaqueductal grayposterior commissureparafascicular nucleusposterior hypothalamic nucleusposterior intralaminar thalamic nucleuspontine nucleiprelimbic cortex
N paraventricular nucleus of the hypothalamusT paraventricular nucleus of the thalamus
pyramidal tractinterstitial nucleus of Cajalred nucleussuperior colliculussubstantia innominata
FPC parvicellular subparafascicular nucleusPC parvicellular part of the ventral posterior thalamic nucleus
zona incertathird ventriclefourth ventricle
amic nuc
s used in
MAmlMPmtotPApcPFPHPILPnPrLPVPVpyRIRNSCSISPVPZI3V4V

lTtcecawtAlsaW
P
TsSalTi
I
Stiulatophtn
C
Brrl(at
FTd black cei 50 �m in
J. Wang et al. / Neuroscience 138 (2006) 1245–1263 1249
ocated in the caudal part of the SPF (arrows on Fig. 2D).IP39-ir neurons in the rSPF appeared somewhat smaller
han those in the SPA. On sagittal sections, the neuro-hemical markers revealed a characteristic rostro-caudallyxpanded sigmoid shape of a group of cells that wasontinuously distributed throughout the periventricularrea of the caudal thalamus (Fig. 3). Rostrally, TIP39 cellsere located within the SPF, but caudally occupied a
erritory dorsal to it, in the upper part of the SPA (Fig. 3C).lthough, the distributions of TIP39-ir and TH-ir cell bodies
argely overlapped, the TH-ir cell bodies were situatedomewhat more laterally than the TIP39 cells in the rSPFnd more laterally and ventrally in the SPA (Fig. 3B, C, D).e found no double-labeled cells (Fig. 3D).
rojections from the rostral part of the SPF
he distribution of BDA-labeled fibers was relatively wide-pread following BDA injections into either the rSPF or thePA (Table 1). The great majority of labeled fibers werelways ipsilateral to the injections, and only a low or very
ow density of labeled fibers was present contralaterally.he distribution of labeled fibers was similar for the two
ig. 2. Coronal sections through the rostral part of the subparafascicuhe TIP39-ir cell bodies are more difficult to see at low magnificationetected by in situ hybridization (high silver grain density causes the
ndicate the caudal part of the SPF. Scale bars�500 �m in A and D,
njection sites with a few notable exceptions (Table 1). r
njection sites
mall injections of the anterograde tracer BDA were placed inhe rSPF and dorsocaudally to it, in the SPA (Fig. 4A–F). Annjection site is shown in Fig. 4G and H to illustrate the tracerptake by neurons in the rSPF. Similarly, an injection site
ocated in the SPA is shown in Fig. 4I and J. Injections in brainreas that border the SPF and area were analyzed as con-
rols. The control injection sites did not overlap with the SPFr area (Fig. 4A–F) and include parts, in various sizes, of thearvicellular ventral posterior thalamic nucleus, the posteriorypothalamic nucleus, the caudal part of the central medial
halamic nucleus, or the interstitial nucleus of Cajal and theucleus of Darkschewitsch.
erebral cortex
DA fibers from the rSPF and the SPA were present inestricted parts of the cerebral cortex. Neurons from bothegions projected to limbic cortical areas including cingu-ate, prelimbic, infralimbic, and dorsal peduncular corticesFig. 5A, B, H and J), and to deeper layers of frontalssociation, including somatosensory, orbital, insular, ec-orhinal, perirhinal and entorhinal cortices. In contrast, a
ic nucleus (A–C) and the SPA (D–F). (A, D) TIP39 immunolabeling.PF; the boxed area in A is enlarged in the inset. (B, E) TIP39 mRNAnters in this darkfield image). (C, F) TH immunolabeling; the arrowsinset, 400 �m in B and E, and 200 �m in C and F.
lar thalamin the rS
emarkable difference was seen between the cortical pro-

FbFob
J. Wang et al. / Neuroscience 138 (2006) 1245–12631250
ig. 3. Sagittal view of the rostro-caudally elongated group of TIP39- and TH-ir cells in the SPF and SPA. (A) Parasagittal drawing of the ratrain modified from Paxinos and Watson (1998). (B) Immunolabeling demonstrates the distribution of the TH-immunoreactivity in the framed area inig. A. (C) Immunolabeling demonstrates the distribution of the TIP39-immunoreactivity in the SPF and SPA. (D) Dual immunofluorescence labelingf TIP39 (red) and TH (green). Note that the distribution of TIP39-immunoreactivity overlaps with that of TH, but they are not co-localized. Scale
ars�800 �m in B, and 200 �m in C and D.
Tt
A
F
T
A
D
J. Wang et al. / Neuroscience 138 (2006) 1245–1263 1251
able 1. Distribution of BDA-labeled fibers following BDA injections inhe rSPF and the SPA compared to that of TIP39 fibers
rea BDAfibers(rSPF)
BDAfibers(SPA)
TIP39fibers
orebrainCerebral cortex
Frontal association cortex � � �
Superficial layer ��� 0 0Cingulate cortex � � 0Prelimbic cortex � � �
Infralimbic cortex � � �
Dorsal peduncular cortex �� �� ��
Tenia tccta �� �� ��
Orbital cortex � � 0Superficial layer �� 0 0
Insular cortex �� � �
Superficial layer ��� 0 0Somatomotor cortex 0 � 0Somatosensory cortex � � 0Visual cortex 0 0 0Auditory cortex 0 0 0
Superficial layer � 0 0Ectorhinal cortex �� �� ��
Superficial layer �� 0 0Perirhinal cortex �� � �
Superficial layer �� 0 0Entorhinal cortex � � 0
Olfactory bulb 0 0 0Ant. olfact. nucleus 0 � �
Olfactory tubercle � � �
Nucleus of the diagonal band �� �� 0Hippocampus
CA1 0 0 0CA3 0 0 0Dentate gyrus 0 0 0Subiculum 0 � �
Parasubiculum � � �
SeptumMedial septal nucleus � �� 0Lateral septal nucleus
Dorsal 0 0 0Intermediate �� ��� ���
Ventral � �� ��
Bed nucleus of the stria terminalis � �� ��
AmygdalaCentral nucleus � �� ��
Basolateral nucleus 0 � 0Basomedial nucleus � �� �
Lateral nucleus 0 � �
Medial nucleus 0 � ��
Cortical nucleus � � �
Anterior amygdaloid area 0 � �
“Fundus striati” � �� ��
Amygdalo-hippocamp. trans. area 0 � �
Amygdalo-piriform trans. area 0 � �
Basal nucleiCaudate-putamen 0 � 0
Ventromedial part 0 �� 0Globus pallidus � �� 0Endopiriform nucleus � � �
Claustrum � � 0
able 1. Continued
rea BDAfibers(rSPF)
BDAfibers(SPA)
TIP39fibers
Nucleus accumbens
Shell � �� ��Core 0 �� �
Ventral pallidum � �� �Substantia innominala � �� ��
iencephalonThalamus
Anterodorsal nucleus 0 �� 0Anteroventral nucleus 0 � 0Paratenial nucleus � � �Paraventricular nucleus � �� ��Reuniens nucleus ��� ��� �Laterodorsal thalamic nucleus � �� 0Central medial thalamic nucleus 0 �� �Mediodorsal nucleus � �� 0Parafascicular nucleus � �� 0SPF Site �� PerikaryaSPA �� Site PerikaryaParvicell. subparafascicular nu. �� �� PerikaryaVentral nuclei � � 0Reticular nucleus � � 0Medial habenular nucleus 0 0 0Lateral habenular nucleus � �� 0Posterior thalamic nucleus � �� �Lateral posterior nucleus �� �� �Post. intralaminar thalamic nu. �� ��� PerikaryaPeripeduncular area � � �Suprageniculate thalamic nu. � � ��Medial geniculate body
Ventral nucleus � � �Medial nucleus � � ��
Lateral geniculate body � � 0Hypothalamus
OVLT 0 0 0Medial preoptic area � ��� �Lateral preoptic area �� ��� �Supraoptic nucleus � � �Supraoptic decussations �� �� ���Suprachiasmatic nucleus 0 � 0Retrochiasmatic area � � �Anterior hypothalamic nucleus �� ��� �Paraventricular nucleus � � ��Periparaventricular area �� �� �Periventricular nucleus � � �Arcuate nucleus � � ��Median eminence 0 0 0Lateral hypothalamic area �� ��� �Perifornical nucleus �� ��� �Ventromedial nucleus � �� 0Dorsomedial nucleus � �� ���Posterior hypothalamic nucleus �� ��� ��Tuberomamillary nucleus �� �� 0Ventral premamillary nucleus � � �Dorsal premamillary nucleus � � 0Supramamillary nucleus � �� �Mamillary body
Medial mamillary nucleus 0 0 0Lateral mamillary nucleus � � 0
Subthalamic nucleus � � �
Forel’s fields �� ��� ��
Fd(##tmt
J. Wang et al. / Neuroscience 138 (2006) 1245–12631252
ig. 4. Illustrations of the locations and sizes of BDA injections into the rSPF, the SPA and in several surrounding control sites. (A–F) Schematicrawings show BDA injections restricted to the rSPF (injection #63, #28, #32, #41, and #62), centered in the rSPF but including surrounding areas#39), involving SPF and some surrounding areas (#40, #34), restricted to the SPA (#72, #129), involving the SPA and some surrounding areas (#84,103, #127), as well as control injections into the PH (#57, #37, #105), the VPPC (#35), the central median thalamic nucleus (#128), the PAG (#104,126), and an area that includes the RI and the nucleus of Darkschewitsch (#124). (G) The coronal section shows an injection site (#28) centered inhe rSPF. (H) Higher magnification of the injection site in G. (I) The coronal section shows an injection site (#129) centered in the SPA. (J) Higheragnification of the injection site in I. Scale bars�800 �m in G and I, and 200 �m in H and J. The schematic drawings are adapted and modified from
he atlas of Paxinos and Watson (1998).

Fc
J. Wang et al. / Neuroscience 138 (2006) 1245–1263 1253
ig. 5. (A–N) Schematic illustrations of the terminal patterns of labeled fibers after BDA injections into the rSPF (left column: A–G) and the SPA (right
olumn: H–N). Maps were modified from the atlas of Paxinos and Watson, 1998.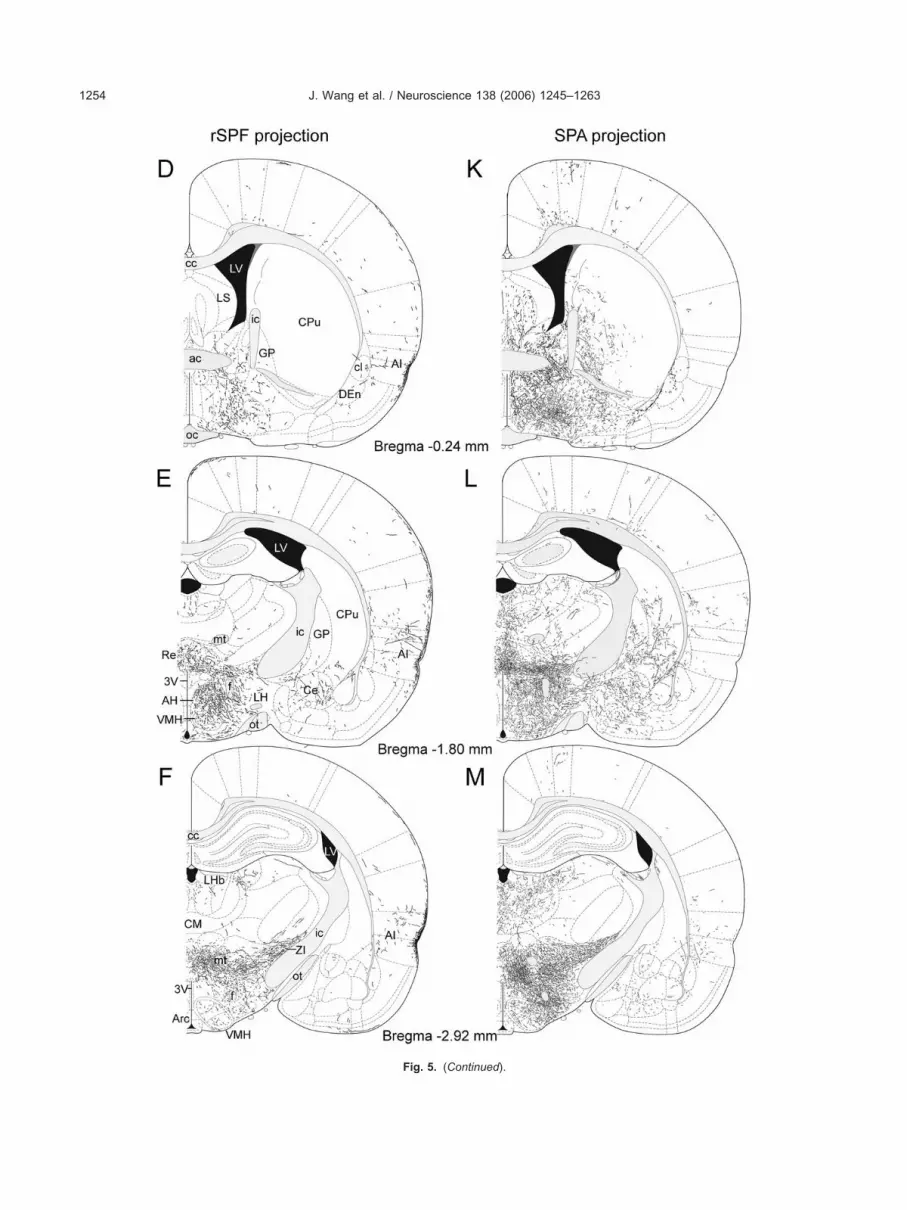
J. Wang et al. / Neuroscience 138 (2006) 1245–12631254
Fig. 5. (Continued).

jtphbctfi
H
Fbr5
S
T5twtwa
A
Taaw
. (Contin
FdB
J. Wang et al. / Neuroscience 138 (2006) 1245–1263 1255
ections of the rSPF and the SPA in the superficial layers ofhe frontal association, orbital, insular, ectorhinal, anderirhinal cortices. These areas contained a very high toigh density of fibers following BDA injection into the rSPFut not the SPA (Fig. 5A–N). The olfactory bulb and someortical areas, including the anterior olfactory nucleus andhe visual and auditory cortices did not contain labeledbers (Table 1).
ippocampus
ew BDA-fibers were found in the subiculum or parasu-iculum. No BDA-fibers were visualized in the CA1–CA3egions of the hippocampus or the dentate gyrus (Fig.E–G and L–N).
Fig. 5
ig. 6. Fluorescence photomicrographs illustrate the distribution of th
istribution of TIP39-ir fibers (B) in the septum. Note the high density of BDA-conDA fibers are absent in the adjacent shell part of the nucleus accumbens. Aleptum
he septum was a major target of both the rSPF (Figs. 5C,D and 6A) and the SPA (Figs. 5J, 5K and 6B). Injections inhe two sites led to similar labeling patterns. Labeled fibersere present in the intermediate and ventral subdivisions of
he lateral septal nucleus but not in its dorsal part. BDA-fibersere moderately dense in the medial septal nucleus, as wells in the nucleus of the diagonal band.
mygdala and the bed nucleus of the stria terminalis
he distribution patterns of BDA-containing fibers in themygdala were different following injections into the rSPFnd the SPA (Table 1). Moderate or low labeling densitiesere seen in all of the amygdaloid nuclei and areas after
ued).
radely labeled fibers following BDA injections into rSPF (A), and the
e anterog taining fibers in the septum ipsilateral to the injection site. In contrast,so note the similar distribution of TIP39 fibers. Scale bars�1 mm.

inrfttacbt
ao
ts
B
TwgtsStd
T
Bwrdcasppfiapwfii
H
Thrsht
Fldwwb
Fth
J. Wang et al. / Neuroscience 138 (2006) 1245–12631256
njections into the SPA (Fig. 5L and 5M), but only a limitedumber of labeled fibers appeared after injections into theSPF (Fig. 5E and 5F). After injections into the SPA, theundus striati (or amygdalo-striatal transitional zone) showedhe highest density of BDA-containing fibers (Fig. 7A), whilehe distribution of the fibers was relatively uniform in othermygdaloid nuclei. Only the central and basomedial nucleiontained a somewhat higher density of BDA-containing fi-ers than other amygdaloid nuclei. Following injections intohe rSPF we found labeled fibers in the central, basomedial
ig. 7. Fluorescence photomicrographs illustrate the distribution ofabeled fibers following BDA injections into the SPA (A), and theistribution of TIP39-ir fibers (B) in the AMY. Arrows point to the FShere the density of BDA containing fibers as well as TIP39-ir fibersas high. Also note the higher density of TIP39 fibers in the MA. Scalears�500 �m.
ig. 8. Fluorescence photomicrographs demonstrate the distribution
he rSPF (A) and the SPA (B). The distribution of TIP39-ir fibers is shown for comypothalamic nucleus, which is devoid of BDA-containing fibers. Scale bars�5nd cortical nuclei, as well as in the fundus striati, but not inther amygdaloid nuclei or areas (Table 1).
All of the divisions of the bed nucleus of the striaerminalis had labeled fibers following injection into eitherite (Fig. 5B–E and 5I–L).
asal ganglia
he rSPF did not project to the caudate-putamen. Thereere large differences in the density of labeled fibers in thelobus pallidus, the endopiriform nucleus, the claustrum,he nucleus accumbens, the ventral pallidum and the sub-tantia nigra after injections into the rSPF and the SPA.PA injections resulted in a high density of BDA fibers in
hese areas whereas rSPF injections resulted in only a lowensity (Fig. 5D, E, K and L; Table 1).
halamus
DA-labeled fibers were present in several thalamic nucleiith a similar distribution pattern after injections into the
SPF and the SPA (Fig. 5E–G and L–N). The highestensity of labeled fibers was present in the reuniens nu-leus and the posterior intralaminar thalamic nucleus. Welso observed a very high density of fibers in the laterodor-al thalamic nucleus, the lateral posterior nucleus, and thearvicellular SPF. In addition, a high density of fibers wasresent in the midline and intralaminar thalamic nucleiollowing SPA, but only at very low density after rSPF,njections. A low density of fibers was also present in thenterodorsal, anteroventral, parataenial, ventral, reticular,eripeduncular, and suprageniculate thalamic nuclei asell as in the medial and lateral geniculate bodies. BDAbers were absent from the medial habenula while present
n the lateral habenula.
ypothalamus
he density of labeled fibers was high in many regions of theypothalamus (Fig. 5D–G, K–N and Fig. 8) following bothSPF and SPA injections. There was a particularly high den-ity of labeled fibers in the lateral preoptic area, the anteriorypothalamic nucleus, around the paraventricular nucleus,
he lateral hypothalamic area, the perifornical nucleus, the
ontaining fibers in the anterior hypothalamus following injections into
of BDA-c parison (C). Note that TIP39-ir fibers are present in the paraventricular00 �m.
pmfiifpmlltptfitn
Pa
IttsaasoIl(t(dcpitlltbl
Twesrh
Tr
FvncstcgHactTash
Ri
Ttsit
Ff(pt
TB
F
D
J. Wang et al. / Neuroscience 138 (2006) 1245–1263 1257
osterior hypothalamic nucleus, and the fields of Forel. Theedial preoptic area contained a very high density of labeled
bers following injection into the SPA but not following rSPFnjection. Labeled fibers were present, at only low densityollowing rSPF injections, in the supraoptic, suprachiasmatic,araventricular, periventricular, arcuate, ventromedial, dorso-edial, premamillary, tuberomamillary (Fig. 5D–G), and the
ateral mamillary nucleus. The low density of anterogradelyabeled fibers in the paraventricular nucleus stands out, par-icularly when compared with the high density of fibers in theeriparaventricular area or the high density of TIP39 fibers inhe paraventricular nucleus (Fig. 8). Anterogradely labeledbers were absent from the organum vasculosum laminaeerminalis, the median eminence, and the medial mamillaryucleus.
rojections following BDA injections into regionsdjacent to the rSPF or the SPA
njections into the parvicellular part of the ventral posteriorhalamic nucleus, the central medial thalamic nucleus, andhe interstitial nucleus of Cajal and the nucleus of Dark-chewitsch resulted in a dramatically different pattern ofnterograde labeling than did the injections into the SPF orrea. Injections into these three sites did not result in aignificant density of anterogradely labeled fibers in mostf the regions receiving projections from the SPF or area.nstead, the parvicellular part of the ventral posterior tha-amic nucleus projected heavily to the insular cortexmostly lamina V but also lamina I); the central medialhalamic nucleus projected heavily to the insular cortexlamina I and also lamina V), the ventral part of the cau-ate-putamen, the substantia innominata (Fig. 9), and theentral and lateral amygdaloid nuclei. Differences are alsoronounced between the distribution of BDA fibers follow-
ng BDA injection into the rostral periaqueductal gray andhe rSPF and SPA. BDA injections into the rSPF and SPAabeled prelimbic, infralimbic, and cingulate cortices, theateral septum, the bed nucleus of the stria terminalis, andhe medial preoptic area, whereas there was not apprecia-le anterograde labeling present in these regions following
arge BDA injections into the rostral periaqueductal gray.
ig. 9. Distribution of anterogradely labeled BDA-containing fibersollowing a control injection of BDA into the caudal part of the CMinjection #128). Dense labeling is present in the Ins, the caudal ventral
sart of the CP, and the SI, but BDA-containing fibers are absent fromhe hypothalamus. Scale bar�1 mm.
he projections of the posterior hypothalamic nucleusere relatively similar to those of the rSPF and the SPA,xcept only the rSPF projected to the superficial layer ofome cortical areas. Another difference was that neu-ons in the posterior hypothalamus projected to theippocampus.
IP39 content of the efferent projections of theSPF and SPA
ollowing injections into the rSPF, as well as the SPA,arying numbers of the BDA containing fibers were immu-opositive for TIP39 in several forebrain areas includingortical, basal forebrain, septal, extended amygdalar, andome diencephalic regions (Table 2). Double-labeled fibersypically demonstrated several double-labeled varicositiesonnected by regions with BDA content only (Fig. 10), sug-esting that TIP39-ir is concentrated in the varicosities.ypothalamic nuclei, including the paraventricular and thenterior hypothalamic nuclei, as well as hindbrain regionsontained both BDA- and TIP39-labeled fibers. However,hese areas did not contain double-labeled fibers (Table 2).he distribution of double-labeled fibers following rSPFnd SPA tracer injections was generally similar, howeverome differences were observed in the amygdala andypothalamus (Table 2).
etrograde labeling in the SPF and area followingnjections into their terminal fields
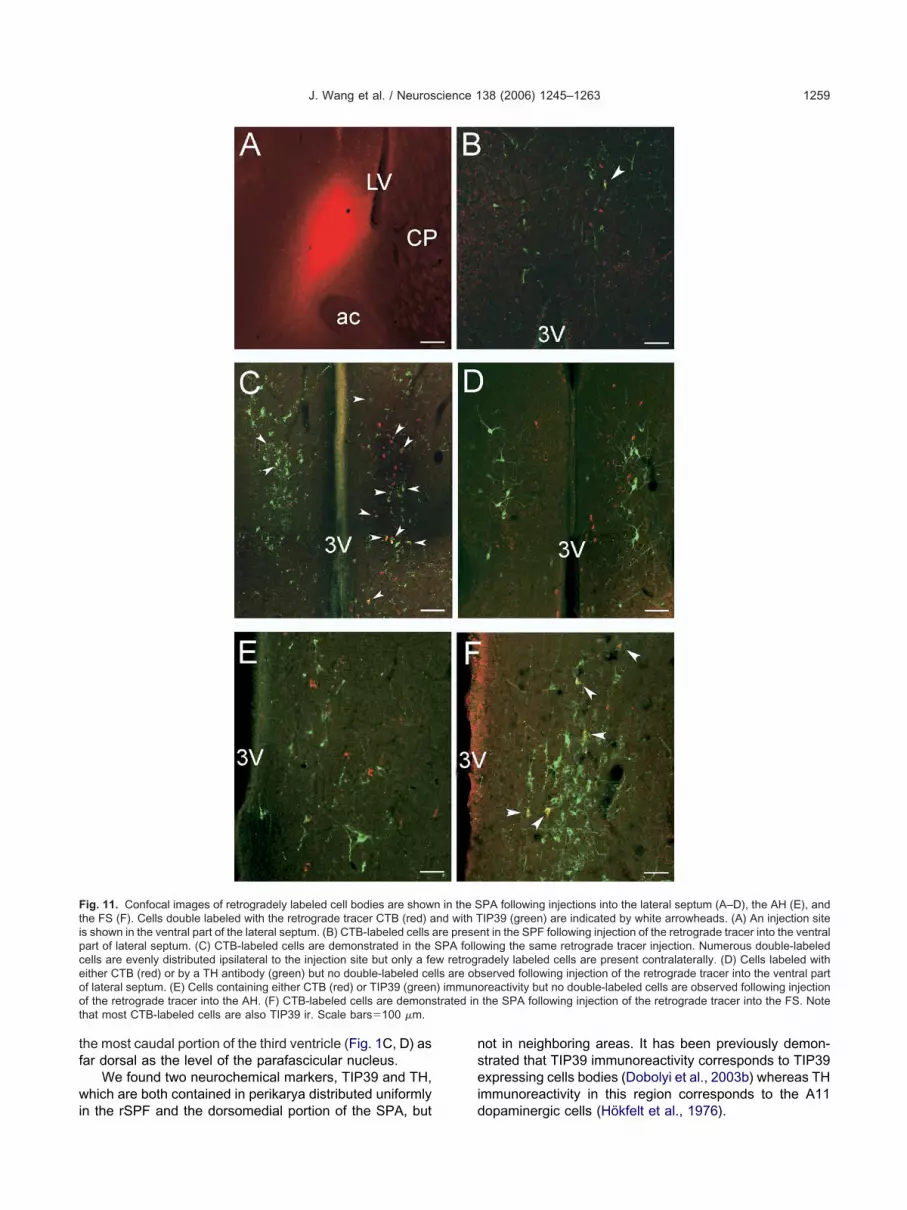
he retrograde tracer CTB was injected into three areas:he ventral part of the lateral septal nucleus, the fundustriati, and the anterior hypothalamic nucleus. Although the
njection sites were not very small, they were restricted tohe target areas, as shown for the ventral part of the lateral
able 2. Presence of BDA and TIP39-ir double-labeled fibers followingDA injections into the rSPF and the SPA
rSPF SPA
orebrainLimbic cortex � �
Insular cortex � �
Ectorhinal cortex � �
Olfactory tubercle � �
Nucleus accumbens. shell part � �
Lateral septal nucleus � �
Bed nucleus of the stria terminalis � �
Substantia innominata � �
Fundus striati � �
Central amygdaloid nucleus � �
iencephalonParaventricular thalamic nucleus � �
Medial preoptic area � �
Anterior hypothalamic nucleus � �
Supraoptic decussations � �
Paraventricular hypothalamic nucleus � �
Arcuate nucleus � �
Dorsomedial hypothalamic nucleus � �
Posterior intralaminar thalamic nucleus � �
eptal nucleus (Fig. 11A). All three injections resulted in
diobtst
mfhTsct
rtpofn
T
TuHnMmglao
swcaomt1dcTtdTc1mg1sMtcbow
iwlcftcc
Fi rs and teo well as d
J. Wang et al. / Neuroscience 138 (2006) 1245–12631258
ominantly ipsilateral labeling of cells distributed uniformlyn the rSPF (Fig. 11B) and the SPA (Fig. 11C). The numberf retrogradely labeled cells was comparable to the num-er of TIP39-labeled cells following injection into the ven-ral part of the lateral septal nucleus (Fig. 11B, C), andomewhat less following injections into the anterior hypo-halamic nucleus (Fig. 11E) and fundus striati (Fig. 11F).
Double-labeling of CTB with TIP39 revealed that al-ost all retrogradely labeled cells were positive for TIP39
ollowing injection into the fundus striati (Fig. 11F), aboutalf of the retrogradely labeled cells were positive forIP39 following injection into the ventral part of the lateraleptal nucleus (Fig. 11B, C), and very few double-labeledells were found following injection into the anterior hypo-halamic nucleus (Fig. 11E).
We found no double-labeling of CTB and TH immuno-eactivity in the SPA following injection of CTB into any ofhe three projection areas, as demonstrated for the ventralart of the lateral septum (Fig. 11D). A uniform distributionf double-labeled cells in the SPF and SPA was observedollowing injections into the ventral part of the lateral septalucleus and the fundus striati (Fig. 11B, C, E).
DISCUSSION
he SPF and the SPA
he SPF has been considered a thalamic nucleus withnknown function (Faul and Mehler, 1985; Turner anderkenham, 1991), or a member of the posterior intralami-ar group of nuclei (LeDoux et al., 1985; Rub et al., 2002).ost authors, however, do not consider it to be one of theidline or intralaminar nuclei (Berendse and Groenewe-en, 1991; Van der Werf et al., 2002). Modern brain at-
ases are consistent in their definition of the SPF, a cyto-rchitectonically distinct structure with circular appearance
ig. 10. Confocal microscope image demonstrates the co-localizationnjection of BDA into the rSPF. Arrowheads indicate double-labeled fiber terminals containing only either TIP39 immunoreactivity or BDA as
n coronal sections (Paxinos and Watson, 1998; Swan- t
on, 2004; Paxinos and Watson, 2005). This terminologyas followed in this manuscript. This is a confined use asompared with the original definition of the SPF as a largerea below the fasciculus retroflexus (Rioch, 1929). More-ver, influential comprehensive descriptions of the thala-us do not mention the SPF or the area corresponding to
he rostral central gray as part of the thalamus (Jones,985; Price, 1995). The complex shape and the difficulty inefining the area without neurochemical markers probablyontributed to the introduction of different terminologies.he term “SPA” and its “compartments” corresponding to
he SPF and the rostral central gray were introduced toescribe connectional data (Moriizumi and Hattori, 1992).he area caudal and dorsal to the SPF has also beenalled “subfascicular area” (Peschanski and Mantyh,983). The term “periventricular gray of the caudal thala-us” was used to describe the location of A11 dopaminer-ic cells (Hökfelt et al., 1979; Skagerberg and Lindvall,985), and to describe sites for stimulus-induced anesthe-ia in the region (Rhodes and Liebeskind, 1978; Boivie andeyerson, 1982). Another classification included the SPF
ogether with the interstitial nucleus of Cajal and the nu-leus of Darkschewitsch as the “perifascicular region”ased on their locations and their projections to the inferiorlive (Saint-Cyr, 1987). However, the inclusion of the SPFas later debated (Onodera and Hicks, 1995).
In our definition, the SPF is a continuous group of cellsn the posterior thalamus. The rostral part of the nucleus,hich contains TIP39- and TH-immunopositive neurons is
ocated rostral to the level of the fasciculus retroflexus. Theaudal part can be followed caudally just ventral to theasciculus retroflexus at the diencephalic–midbrain junc-ion and it is topographically connected to the laterally andaudally located parvicellular SPF. This part of the SPF isonsidered a component of the SPA. The major portion of
-ir (green) with BDA-containing fibers (red) in the lateral septum afterrminals that contain both BDA and TIP39-ir. Note that there are fibersouble labeled ones. Scale bar�10 �m.
of TIP39
his area is located between the fasciculus retroflexus and
tf
wi
nsei
Ftipceoot
J. Wang et al. / Neuroscience 138 (2006) 1245–1263 1259
he most caudal portion of the third ventricle (Fig. 1C, D) asar dorsal as the level of the parafascicular nucleus.
We found two neurochemical markers, TIP39 and TH,hich are both contained in perikarya distributed uniformly
ig. 11. Confocal images of retrogradely labeled cell bodies are showhe FS (F). Cells double labeled with the retrograde tracer CTB (red) as shown in the ventral part of the lateral septum. (B) CTB-labeled cells aart of lateral septum. (C) CTB-labeled cells are demonstrated in theells are evenly distributed ipsilateral to the injection site but only a feither CTB (red) or by a TH antibody (green) but no double-labeled cef lateral septum. (E) Cells containing either CTB (red) or TIP39 (greenf the retrograde tracer into the AH. (F) CTB-labeled cells are demonshat most CTB-labeled cells are also TIP39 ir. Scale bars�100 �m.
n the rSPF and the dorsomedial portion of the SPA, but d
ot in neighboring areas. It has been previously demon-trated that TIP39 immunoreactivity corresponds to TIP39xpressing cells bodies (Dobolyi et al., 2003b) whereas TH
mmunoreactivity in this region corresponds to the A11
PA following injections into the lateral septum (A–D), the AH (E), andIP39 (green) are indicated by white arrowheads. (A) An injection sitent in the SPF following injection of the retrograde tracer into the ventralwing the same retrograde tracer injection. Numerous double-labeledadely labeled cells are present contralaterally. (D) Cells labeled withserved following injection of the retrograde tracer into the ventral partreactivity but no double-labeled cells are observed following injectionthe SPA following injection of the retrograde tracer into the FS. Note
n in the Snd with Tre prese
SPA follow retrogrlls are ob) immunotrated in
opaminergic cells (Hökfelt et al., 1976).

E
NtpdttfabcdtmftT
lt2jNpnDmidTtaatga(rca
tiditpvlicpbtfh
B
imb1dtn
Tm
Tjd(ottnstttzlattofica
bsTtlrt
adMTsTcls(d(stp
b
J. Wang et al. / Neuroscience 138 (2006) 1245–12631260
fferent projections of the rSPF and the SPA
eurons in the rSPF send a large number of efferent fiberso forebrain areas with a somewhat different projectionattern than neurons located in the SPA. In general, theensity of anterogradely labeled fibers observed in most ofhe forebrain regions is lower for rostral SPF injectionshan for the caudal ones. The superficial layers of therontal association, orbital, insular, auditory, ectorhinal,nd perirhinal cortices receive many fibers from the rSPFut not from the SPA, or areas around the SPF. In turn, theore portion of the nucleus accumbens, the globus palli-us, the anterodorsal, mediodorsal, and central medialhalamic nuclei, the medial preoptic area, as well as theidbrain raphe nuclei receive no, or only solitary, fibers
rom the rSPF. Furthermore, differences in projections tohe amygdala from the rSPF and the SPA are significant.he innervation of the amygdala by the rSPF is minor.
The specificity of the projections to the injection sites isikely, because high molecular weight BDA as anterograderacer is not taken up by passing fibers (Reiner et al.,000), and because we observed profoundly different pro-
ection patterns following injections into neighboring nuclei.otably, injections into the parvicellular part of the ventralosterior thalamic nucleus, the central medial thalamicucleus, the interstitial nucleus of Cajal, and the nucleus ofarkschewitsch resulted in restricted projections that wereost significant in deep layers of the insular cortex, the
nsular cortex–ventral caudate-putamen–central amyg-ala regions, and in the oculomotor nuclei, respectively.hese projections are in agreement with published data for
hese sites (Berendse and Groenewegen, 1991; Onoderand Hicks, 1995; Wang and Spencer, 1996; Nakashima etl., 2000; Van der Werf et al., 2002). We also observedhat the ascending projections of the rostral periaqueductalray are also much more restricted than those of the SPAnd rSPF, which is in agreement with published dataCameron et al., 1995). In addition, the projections fromSPF to the lateral septum and the fundus striati wereonfirmed by injecting retrograde tracer into these threereas and mapping labeled cell bodies in the SPF.
There are many brain areas where more fibers con-ained the anterograde tracer than TIP39 immunoreact-vity, including the superficial layers of the cortex, the me-ial septal nucleus, the caudate-putamen, several midline,
ntralaminar and ventral thalamic nuclei, the lateral preop-ic and lateral hypothalamic areas, the area around thearaventricular nucleus of the hypothalamus, the anterior,entromedial hypothalamic nuclei and some of the mamil-
ary nuclei. The absence or very low density of TIP39 fibersn these regions suggests that the rSPF and the SPAontain neurons other than TIP39 synthesizing ones thatroject to these regions. This finding is further supportedy the very low percentage of TIP39-positive cells among
he retrogradely labeled cells in the rSPF and the SPAollowing injection of retrograde tracer into the anteriorypothalamic nucleus.
We found no fibers in the brain double labeled with
DA and TH. This finding is consistent with previous stud- les demonstrating that the major target of the A11 dopa-inergic cells is the spinal cord (Bjorklund and Skager-erg, 1979; Hökfelt et al., 1979; Skagerberg and Lindvall,985). Since there were brain regions with high BDA fiberensity that did not contain either TIP39 or TH, it is likelyhat there are projection neurons in the SPA-rSPF that doot contain either of these markers.
IP39 innervation of the forebrain: TIP39 cells asajor output neurons of the posterior thalamus
he patterns of anterogradely labeled fibers following in-ections into the rSPF and the SPA were similar to theistribution of TIP39-ir fibers in many areas of the brainTable 1). There was a high density and similar distributionf both anterogradely labeled fibers and TIP39-ir fibers inhe limbic and ectorhinal cortices, the olfactory tubercle,he shell part of the nucleus accumbens, the lateral septalucleus and the bed nucleus of the stria terminalis, theubstantia innominata, the central amygdaloid nucleus andhe fundus striati, the paraventricular and posterior in-ralaminar nuclei of the thalamus, the medial preoptic area,he posterior hypothalamic nucleus, the fields of Forel, theona incerta and the periaqueductal gray. Fibers double-
abeled with BDA and TIP39 were present in all of thesereas. Together these data suggest that TIP39-ir fibers inhese regions originate in the medial part of the posteriorhalamus. This finding was further confirmed by injectionsf retrograde tracers into the lateral septal nucleus and theundus striati that resulted in labeled TIP39 perikarya bothn the SPF and SPA. The percentage of double-labeledells was high, supporting the idea that TIP39 neurons aremajor type of output neuron in both the SPF and SPA.
The fact that many brain areas contained fibers dou-le-labeled with BDA and TIP39 following injections in bothubparafascicular regions, and the finding that labeledIP39 cell bodies were evenly distributed in the SPF and
he SPA following injections of retrograde tracer into theateral septum and the fundus striati suggest that TIP39 neu-ons in the rSPF and the SPA have similar projection pat-erns, and that TIP39 neurons constitute a single population.
Some of the hypothalamic cell areas including thenterior, paraventricular, arcuate and dorsomedial nucleiid not contain fibers double-labeled with TIP39 and BDA.ost of these nuclei actually contain a higher density ofIP39-ir than BDA-labeled fibers. Together these datauggest that TIP39 fibers in these regions originate inIP39 cells other than those in the rSPF or the SPA. TIP39ell bodies are present in the parvicellular subparafascicu-
ar and the posterior intralaminar thalamic nuclei and theurrounding area medial to the medial geniculate bodyDobolyi et al., 2003b). Lesions of this area markedly re-uced the density of TIP39-ir fibers in the hypothalamusDobolyi et al., 2003a). In addition, these lesions clearlyhowed that the majority of TIP39-ir fibers and terminals inhe amygdala emanate from neurons in the parvicellular sub-arafascicular and the posterior intralaminar thalamic nuclei.
Part of the trajectory of anterogradely labeled fibers cane followed from the rSPF to the forebrain. A group of fibers
eaves the nucleus dorsolaterally, then after a hook-shaped

tpfilAotacts
P
LScsaariaMavspact
nSpmptScsncrvisagc(sSpt
Iad
F area. Prof FS, the A
J. Wang et al. / Neuroscience 138 (2006) 1245–1263 1261
urn they enter the epithalamus and run rostralwards in thearaventricular nucleus of the thalamus (Fig. 12). Some otherbers run rostrally from the nucleus, just dorsal to the medialemniscus and terminate, most probably in thalamic nuclei.nother group, which comprises the most substantial numberf fibers, joins the zona incerta and the supraoptic decussa-ions and enters the basal forebrain/substantia innominatarea. From here, the fibers divide into branches to limbicortical, septal and amygdaloid areas (Fig. 12). The fineopography and the terminal pattern of these fibers will be theubjects of further studies.
ossible functions of the SPF and area
ittle functional data are available regarding the SPF andPA. Some evidence suggests a role of the SPA in noci-eptive modulation. Cells in the area are activated in re-ponse to noxious stimuli (Dong et al., 1978; Sugiyama etl., 1992) and analgesia can be induced by stimulating therea. In addition to the activation of cells in the SPA inesponse to nociceptive stimuli (Dong et al., 1978; Sug-yama et al., 1992), cells in the area also exhibit c-fosctivation following exposure to cold (Kiyohara et al., 1995;iyata et al., 1995) and high intensity noise (Palkovits etl., 2004). The resulting corticosterone response might in-olve projections to the medial prefrontal cortex, the lateraleptum, the bed nucleus of the stria terminalis, the medialreoptic area, the periparaventricular region of the hypothal-mus, the amygdala, and the paraventricular thalamic nu-leus, areas which have all been shown to be able to affect
ig. 12. Summary of major projections of neurons in the SPF andorebrain, the lateral septum, the BST, the MPA, the AH, the PH, the
he corticosterone response driven by the paraventricular r
ucleus (Herman et al., 2003). Other projections of thePF and the SPA reach regions, such as the intermediateart of the lateral septum, the anteromedial division of theedial amygdaloid nucleus ventromedial and anterior hy-othalamic nuclei, that are parts of a neural network con-rolling defensive behavior (Fuchs et al., 1985; Risold andwanson, 1997; Canteras, 2002). Amygdaloid projectionsould affect emotional responses including the fear re-ponse (LeDoux, 2003). Projections to midline thalamicuclei and to cholinergic areas of the basal forebrain couldontribute to awareness (Smythies, 1997). Several areaseceiving projections from the SPA are involved in cardio-ascular and respiratory regulation. Such target regions
nclude the amygdala, the A5 noradrenergic cell group, theubcoeruleus area, and the pontine raphe nucleus (Loewynd McKellar, 1980). Based on all of the projections to-ether, we hypothesize that the SPA is involved in theentral regulation of the generalized stress responseChrousos and Gold, 1992). Although it is premature topeculate about functions of subgroups of cells within thePF and the SPA, it is worth mentioning that TIP39 cellsreferentially project to limbic areas that potentially affecthe corticosterone response.
CONCLUSION
n conclusion, our results allowed us to identify the SPFnd SPA neurochemically using TIP39 and tyrosine-hy-roxylase as markers distributed throughout the elongated
jections to the infralimbic, prelimbic and insular cortices, the basalMY, and the paraventricular thalamic nucleus are indicated.
ostrocaudal axis of the region. This study is the first to

stan
AtfaF
B
B
B
B
B
C
C
C
D
D
D
F
F
F
G
H
H
H
H
JK
L
L
L
L
M
M
M
M
M
N
N
O
O
P
P
P
P
P
J. Wang et al. / Neuroscience 138 (2006) 1245–12631262
ystematically describe the efferent connections of bothhe SPF and the SPA, and to demonstrate dense limbicnd autonomic projections. In addition we identified TIP39eurons as major output neurons of the rSPF and the SPA.
cknowledgments—Support was provided by the National Insti-ute of Mental Health Intramural Research Program. We are grate-ul to Dr. Éva Mezey for advice and assistance. We also appreci-te help from Dr. Carolyn Smith and the NINDS Light Imagingacility with confocal microscopy.
REFERENCES
eckstead RM (1984) The thalamostriatal projection in the cat.J Comp Neurol 223(3):313–346.
entivoglio M, Molinari M (1984) The interrelations between cellgroups in the caudal diencephalon of the rat projecting to thestriatum and to the medulla oblongata. Exp Brain Res 54(1):57–65.
erendse HW, Groenewegen HJ (1991) Restricted cortical terminationfields of the midline and intralaminar thalamic nuclei in the rat.Neuroscience 42:73–102.
jorklund A, Skagerberg G (1979) Evidence for a major spinal cordprojection from the diencephalic A11 dopamine cell group in the ratusing transmitter-specific fluorescent retrograde tracing. Brain Res177:170–175.
oivie J, Meyerson BA (1982) A correlative anatomical and clinicalstudy of pain suppression by deep brain stimulation. Pain 13:113–126.
ameron AA, Khan IA, Westlund KN, Cliffer KD, Willis WD (1995) Theefferent projections of the periaqueductal gray in the rat: a Phaseo-lus vulgaris-leucoagglutinin study. I. Ascending projections. J CompNeurol 351:568–584.
anteras NS (2002) The medial hypothalamic defensive system: hod-ological organization and functional implications. Pharmacol Bio-chem Behav 71:481–491.
hrousos GP, Gold PW (1992) The concepts of stress and stresssystem disorders. Overview of physical and behavioral homeosta-sis. JAMA 267:1244–1252.
obolyi A, Palkovits M, Bodnar I, Usdin TB (2003a) Neurons contain-ing tuberoinfundibular peptide of 39 residues project to limbic,endocrine, auditory and spinal areas in rat. Neuroscience 122:1093–1105.
obolyi A, Palkovits M, Usdin TB (2003b) Expression and distributionof tuberoinfundibular peptide of 39 residues in the rat centralnervous system. J Comp Neurol 455:547–566.
ong WK, Ryu H, Wagman IH (1978) Nociceptive responses of neu-rons in medial thalamus and their relationship to spinothalamicpathways. J Neurophysiol 41:1592–1613.
aul RLM, Mehler WR (1985) Thalamus. In: The rat nervous system,Vol. I (Paxinos G, ed), pp 129–168. Sydney: Academic Press.
leetwood-Walker SM, Hope PJ, Mitchell R (1988) Antinociceptiveactions of descending dopaminergic tracts on cat and rat dorsalhorn somatosensory neurones. J Physiol 399:335–348.
uchs SA, Edinger HM, Siegel A (1985) The organization of thehypothalamic pathways mediating affective defense behavior inthe cat. Brain Res 330:77–92.
uo ZL, Li P, Longhurst JC (2002) Central pathways in the pons andmidbrain involved in cardiac sympathoexcitatory reflexes in cats.Neuroscience 113:435–447.
erman JP, Figueiredo H, Mueller NK, Ulrich-Lai Y, Ostrander MM,Choi DC, Cullinan WE (2003) Central mechanisms of stressintegration: hierarchical circuitry controlling hypothalamo-pituitary-adrenocortical responsiveness. Front Neuroendocrinol 24:151–180.
ökfelt T, Johansson O, Fuxe K, Goldstein M, Park D (1976) Immu-
nohistochemical studies on the localization and distribution ofmonoamine neuron systems in the rat brain. I. Tyrosinehydroxylase in the mes- and diencephalon. Med Biol 54:427–453.
ökfelt T, Phillipson O, Goldstein M (1979) Evidence for a dopaminer-gic pathway in the rat descending from the A11 cell group to thespinal cord. Acta Physiol Scand 107:393–395.
osobuchi Y (1983) Combined electrical stimulation of the periaque-ductal gray matter and sensory thalamus. Appl Neurophysiol46:112–115.
ones EG (1985) The thalamus. New York: Plenum.iyohara T, Miyata S, Nakamura T, Shido O, Nakashima T, Shibata M
(1995) Differences in Fos expression in the rat brains between coldand warm ambient exposures. Brain Res Bull 38:193–201.
aBuda CJ, Dobolyi A, Usdin TB (2004) Tuberoinfundibular peptide of39 residues produces anxiolytic and antidepressant actions. Neu-roreport 15:881–885.
eDoux J (2003) The emotional brain, fear, and the amygdala. CellMol Neurobiol 23:727–738.
eDoux JE, Ruggiero DA, Reis DJ (1985) Projections to the subcor-tical forebrain from anatomically defined regions of the medialgeniculate body in the rat. J Comp Neurol 242:182–213.
oewy AD, McKellar S (1980) The neuroanatomical basis of centralcardiovascular control. Fed Proc 39:2495–2503.
iyata S, Ishiyama M, Shido O, Nakashima T, Shibata M, Kiyohara T(1995) Central mechanism of neural activation with cold acclima-tion of rats using Fos immunohistochemistry. Neurosci Res22:209–218.
oriizumi T, Hattori T (1991) Non-dopaminergic projection from thesubparafascicular area to the temporal cortex in the rat. NeurosciLett 129:127–130.
oriizumi T, Hattori T (1992) Anatomical and functional compartmen-talization of the subparafascicular thalamic nucleus in the rat. ExpBrain Res 90:175–179.
oriizumi T, Leduc-Cross B (1992) Is there a dopaminergic projectionfrom the A11 catecholamine cell group to the amygdala? Exp BrainRes 88(2):451–454.
usil SY, Olson CR (1988) Organization of cortical and subcorticalprojections to anterior cingulate cortex in the cat. J Comp Neurol272(2):203–218.
akashima M, Uemura M, Yasui K, Ozaki HS, Tabata S, Taen A(2000) An anterograde and retrograde tract-tracing study on theprojections from the thalamic gustatory area in the rat: distributionof neurons projecting to the insular cortex and amygdaloid com-plex. Neurosci Res 36:297–309.
ocjar C, Roth BL, Pehek EA (2002) Localization of 5-HT(2A) recep-tors on dopamine cells in subnuclei of the midbrain A10 cell group.Neuroscience 111:163–176.
nodera S, Hicks TP (1995) Patterns of transmitter labelling andconnectivity of the cat’s nucleus of Darkschewitsch: a wheat germagglutinin-horseradish peroxidase and immunocytochemical studyat light and electron microscopical levels. J Comp Neurol 361:553–573.
ttersen OP, Ben-Ari Y (1979) Afferent connections to the amygdaloidcomplex of the rat and cat. I. Projections from the thalamus.J Comp Neurol 187(2):401–424.
alkovits M, Dobolyi A, Helfferich F, Usdin TB (2004) Localization andchemical characterization of the audiogenic stress pathway. AnnN Y Acad Sci 1018:16–24.
axinos G, Watson C (1998) The rat brain in stereotaxic coordinates.San Diego: Academic Press.
axinos G, Watson C (2005) The rat brain in stereotaxic coordinates.San Diego: Academic Press.
eschanski M, Mantyh PW (1983) Efferent connections of the subfas-cicular area of the mesodiencephalic junction and its possibleinvolvement in stimulation-produced analgesia. Brain Res 263:181–190.
rice JL (1995) Thalamus. In: The rat nervous system (Paxinos G, ed),
pp 629–648. San Diego: Academic Press.
R
R
R
R
R
S
S
S
S
S
S
T
T
T
T
U
U
V
W
W
W
Y
Y
J. Wang et al. / Neuroscience 138 (2006) 1245–1263 1263
einer A, Veenman CL, Medina L, Jiao Y, Del Mar N, Honig MG (2000)Pathway tracing using biotinylated dextran amines. J NeurosciMethods 103:23–37.
hodes DL, Liebeskind JC (1978) Analgesia from rostral brain stemstimulation in the rat. Brain Res 143:521–532.
ioch DK (1929) Studies on the diencephalon of carnivora. Part I. Thenuclear configuration of the thalamus, epithalamus, and hypothal-amus of the dog and cat. J Comp Neurol 49:1–120.
isold PY, Swanson LW (1997) Connections of the rat lateral septalcomplex. Brain Res Brain Res Rev 24:115–195.
ub U, Del Tredici K, Del Turco D, Braak H (2002) The intralaminarnuclei assigned to the medial pain system and other componentsof this system are early and progressively affected by the Alzhei-mer’s disease-related cytoskeletal pathology. J Chem Neuroanat23:279–290.
aint-Cyr JA (1987) Anatomical organization of cortico-mesen-cephalo-olivary pathways in the cat as demonstrated by axonaltransport techniques. J Comp Neurol 257:39–59.
kagerberg G, Lindvall O (1985) Organization of diencephalic dopa-mine neurones projecting to the spinal cord in the rat. Brain Res342:340–351.
mythies J (1997) The functional neuroanatomy of awareness: with afocus on the role of various anatomical systems in the control ofintermodal attention. Conscious Cogn 6:455–481.
ugimura Y, Murase T, Ishizaki S, Tachikawa K, Arima H, Miura Y,Usdin TB, Oiso Y (2003) Centrally administered tuberoinfundibularpeptide of 39 residues inhibits arginine vasopressin release inconscious rats. Endocrinology 144:2791–2796.
ugiyama K, Ryu H, Uemura K (1992) Identification of nociceptiveneurons in the medial thalamus: morphological studies of nocicep-tive neurons with intracellular injection of horseradish peroxidase.Brain Res 586:36–43.
wanson LW (2004) Brain maps: structure of the rat brain. San Diego:Academic Press.
akada M (1990) The A11 catecholamine cell group: another origin ofthe dopaminergic innervation of the amygdala. Neurosci Lett
118(1):132–135.akada M (1993) Widespread dopaminergic projections of the sub-parafascicular thalamic nucleus in the rat. Brain Res Bull32(3):301–309.
akada M, Li ZK, Hattori T (1988) Single thalamic dopaminergicneurons project to both the neocortex and spinal cord. Brain Res455(2):346–352.
urner BH, Herkenham M (1991) Thalamoamygdaloid projections inthe rat: a test of the amygdala’s role in sensory processing. J CompNeurol 313:295–325.
sdin TB (2000) The PTH2 receptor and TIP39: a new peptide-receptor system. Trends Pharmacol Sci 21:128–130.
sdin TB, Hoare SR, Wang T, Mezey E, Kowalak JA (1999) TIP39: anew neuropeptide and PTH2-receptor agonist from hypothalamus.Nat Neurosci 2:941–943.
an der Werf YD, Witter MP, Groenewegen HJ (2002) The intralami-nar and midline nuclei of the thalamus. Anatomical and functionalevidence for participation in processes of arousal and awareness.Brain Res Brain Res Rev 39:107–140.
ang SF, Spencer RF (1996) Spatial organization of premotor neu-rons related to vertical upward and downward saccadic eye move-ments in the rostral interstitial nucleus of the medial longitudinalfasciculus (riMLF) in the cat. J Comp Neurol 366:163–180.
ang T, Palkovits M, Rusnak M, Mezey E, Usdin TB (2000) Distribu-tion of parathyroid hormone-2 receptor-like immunoreactivity andmessenger RNA in the rat nervous system. Neuroscience 100:629–649.
ard HL, Small CJ, Murphy KG, Kennedy AR, Ghatei MA, Bloom SR(2001) The actions of tuberoinfundibular peptide on the hypo-thalamo-pituitary axes. Endocrinology 142:3451–3456.
asui Y, Nakano K, Mizuno N (1992) Descending projections from thesubparafascicular thalamic nucleus to the lower brain stem in therat. Exp Brain Res 90:508–518.
eung JC, Yaksh TL, Rudy TA (1977) Concurrent mapping of brainsites for sensitivity to the direct application of morphine and focalelectrical stimulation in the production of antinociception in the rat.
Pain 4:23–40.(Accepted 1 December 2005)(Available online 3 February 2006)





























