Embed Size (px)
Citation preview

gy and Ecology 344 (2007) 88–100www.elsevier.com/locate/jembe
Journal of Experimental Marine Biolo
Environmental factors influencing urchin spatial distributions ondisturbed coral reefs (New Caledonia, South Pacific)
P. Dumas a,⁎, M. Kulbicki b, S. Chifflet c, R. Fichez c, J. Ferraris b
a UR 128 CoReUs, Centre IRD de Nouméa, BP A5, 98848 Nouméa Cedex, New Caledonia, Franceb UR 128, EPHE, 52 Av. P. Alduy, 66860 Perpignan Cedex, France
c Centre d'Océanologie de Marseille, Station Marine d'Endoume, Rue de la Batterie des Lions, 13007 Marseille, France
Received 2 June 2006; received in revised form 5 October 2006; accepted 28 December 2006
Abstract
Few works have examined the relative contributions of habitat variables to the distribution of coral reef urchins. In the presentstudy, the spatial distribution of two common urchin species (Diadema setosum and Echinometra mathaei) was studied in thefringing reefs of two urban bays in New Caledonia (South Pacific). Urchins were surveyed at 105 stations with contrasted habitatstructure/anthropic disturbance levels; 32 environmental variables (water/sediment characteristics, reef structuring species) wereconsidered. Moderate densities were generally observed at station scale (mean 0.5 individuals m−2). The combination of univariateand multivariate techniques highlighted patchy distributions for Diadema as well as Echinometra, with distinct species/habitatassociations; environmental gradients occurring within the bays did not seem to influence the species patterns. For Diadema, thespatial variability was better explained by sediment type than by biotic cover; increasing densities occurred across habitats withlarger sediment sizes and decreasing coral complexity/macrophytes cover. In contrast, the distribution of E. mathaei exhibitedweak relationships with habitat variables. In coral reefs, small-scale heterogeneity may thus be responsible for most of urchinsspatial variability.© 2007 Elsevier B.V. All rights reserved.
Keywords: Coral reefs; New Caledonian; Sea urchins; Spatial distribution
1. Introduction
As a consequence of increasing human pressure,coastal ecosystems are facing a wide range of threats interms of resource exploitation, habitat modification and/or pollution (Hodgson, 1999; Vitousek et al., 1997;Jackson et al., 2001; Healy and Wang, 2004; Raschet al., 2005). Recent evaluations thus reported that 30%of the world's coastal land area is severely impacted byanthropogenic activities including urban settlement,
⁎ Corresponding author.E-mail address: [email protected] (P. Dumas).
0022-0981/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.jembe.2006.12.015
development of fisheries and aquaculture, or thegrowing demand for recreational activities (e.g. Burkeet al., 2001). Yet, as coastal and inland populationscontinue to grow, impacts can be expected to grow aswell (Richmond, 1993; Wilkinson, 1999). In thiscontext, there is an urgent need for better informationregarding the ecological processes that influencepopulations and assemblages, in order to assess thevulnerability of aquatic communities to human-inducedenvironmental changes and to derive adequate manage-ment policies (Grigg and Dollar, 1990).
The study of species/habitat associations has a longhistory in aquatic ecology. An abundant literature

89P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
addresses the influence of environmental factors on thedistribution patterns of benthic organisms in diverseecosystems such as streams, lakes/ponds, ground waters,littoral zones including mangroves, estuaries, saltmarshes, and marine open-waters (e.g. see reviews inVandel, 1964; Gray, 1974; Hildrew and Townsend,1987; Alongi, 1990; Alongi and Sasekumar, 1992;Balvay, 1993; Gibert et al., 1994; Herman et al., 1999;Laine, 2003; Nanami et al., 2005a). For coral reefs,studies on species/habitat associations began morerecently in the mid 70s (e.g. works of Salvat et al.,1972; Risk, 1972; Loya, 1976; Luckhurst and Luckhurst,1978; Sale, 1978, 1980), and understanding the factorsthat influence species composition and communitystructure still remains a challenge (Hughes and Connell,1999; Chiappone et al., 2002; Schiel et al., 2004).
While results tend to be species-specific and stronglyscale-dependant, recent works suggested that spatialdistributions of coral-associated organisms are influ-enced by a rather diverse set of environmental factorsrelative to both substrate and water column, whoserespective contributions may vary (McClanahan, 1992;Chabanet et al., 1997; Zuschin et al., 2001; Lecchiniet al., 2003; Nanami et al., 2005b; Cleary et al., 2005;Bozec et al., 2005). The intensity of species/habitatassociations may also be mediated through biological orbehavioral processes, e.g. when considering sedentary/territorial vs. wandering species (Chabanet et al., 1997).
In comparison with fishes, available data on thefactors structuring reef invertebrates is scarce. Forurchins, species exhibit close linkage with substrataderiving from their life habits (feeding strategies,locomotory behavior, substrate relations etc.) and spatialdistributions are generally expected to reflect similartrends (Lawrence, 2001). Yet, a high variability is oftenobserved as the result of complex interactions betweenhabitat variables including depth, wave exposure, water/sediment composition and the presence/absence of reef-building or covering species (Nishihira et al., 1991;Flammang et al., 1997; Adjeroud, 1997; Adjeroud,2000; Chiappone et al., 2002; King and Riddle, 2001;Cleary et al., 2005). As the influence of physical/chemical factors is mediated by the biological nature ofreef substrate, structural species such as corals and algaeconstitute a critical dimension of habitats (Done et al.,1996) that should be encompassed in studies addressingreef urchin distribution patterns. Yet, as physicalcomplexity in coral reefs makes habitat descriptionand sampling uneasy, most studies focused on arestricted set of variables or stations, making it difficultto discern the contributions of environmental factors tothe observed urchin distributions.
The aim of this study was to investigate the factorsresponsible for urchin spatial distributions in thefringing reefs of two urban bays in New Caledonia.Common, widely-distributed reef urchins Diademasetosum and Echinometra mathaei were surveyed in alarge set of stations with contrasted habitat structure andanthropic disturbance levels. 105 stations correspondingto 210 transects and 32 environmental variables (water/sediment characteristics, reef structuring species) wereconsidered 1) to investigate the relationships betweenurchins spatial distributions and gradients in environ-mental variables; 2) to elucidate the impacts of anthropicdisturbances on urchins spatial gradients, and 3) toassess the relative contribution of environmental vari-ables to the distributions observed.
2. Materials and methods
2.1. Study area
All stations were located on the fringing reefs ofGrande Rade and Sainte Marie bays, on the Noumeapeninsula, New Caledonia (Fig. 1). While they are verysimilar in size, the two bays exhibit different exposure totrade winds and water circulation (Douillet et al., 2001)and are submitted to different types of humandisturbances. Grande Rade mainly receives industrialpollutants from the surrounding industrial zones (nickelsmelter, commercial harbor) whereas Sainte Marie bayis mainly submitted to domestic wastes. As a result ofurban influences, anthropogenic eutrophication alsooccurs in the inshore parts of the bays (Fichez et al.,submitted for publication). In the bays, fringing reefs aregenerally composed of three different zones in closesuccession: a) a reef flat (depth range 0.5–2 m) withpoor coral cover; b) a reef crest (depth range 1–3 m)with flourishing corals; and c) a slight slope with similarcorals cover connecting to the sandy bottom of the bays,up to 6 m depth (Bozec et al., 2005).
2.2. Urchin sampling
The surveys occurred in the bays between December,1997 and August, 1998 at 105 reef stations (47 inGrande Rade, 58 in Sainte Marie). On each station, two50 m-long transects were defined, parallel to oneanother. They were laid within the same reef habitat(reef flat, reef crest or reef slope); distance between thetransects was approximately 3 m. Each transect wasdivided into 5 sectors of 10 m; all urchins which weredetectable without disturbing the substrate were countedon a 1 m band for each sector along the transects. Two

Fig. 1. Study area — location of the sampling stations in the bays of Grande Rade and Sainte Marie, New Caledonia (South Pacific). ● Urchin/sediment sampling stations ▴ water sampling stations.
90 P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
species of urchins were then taken into account:D. setosum and E. mathaei.
2.3. Environmental factors
Environmental variables were composed of threedata sets referred hereafter as sediment type, substratumcoverage and water characteristics (Table 1).
Sediment type and substratum coverage variableswere estimated using the linear point intercept method(Ohlhorst et al., 1988). Every meter along each transect,the various sediment types (from mud to hard bottoms,10 variables) and substratum coverage (large, sessilecovering organisms, 8 variables) were recorded, thenpercentages were calculated (cf. Table 1). For eachstation, the exposure to dominant trade winds was alsorecorded (i.e. windward/leeward). Water physicochem-
ical characteristics were sampled with a different spatialresolution: 3 recording stations were sampled in SainteMarie, 5 stations in Grande Rade. These stations wereequally distributed along an axis spanning from thebottom to the entrance of each bay (“longitudinal axis”,hereafter). Temperature, turbidity, salinity, nitrates,phosphates, ammonium, particulate and dissolvedorganic matter concentrations (PON, POP, POC,DON, DOP) were measured monthly using CTD pro-filer and standard analytical techniques (Fichez et al.,submitted for publication).
2.4. Data analysis
Bay/wind exposure effects on urchin densities wereinvestigated using factorial analysis of variance(ANOVA). Relationships between urchin patterns and
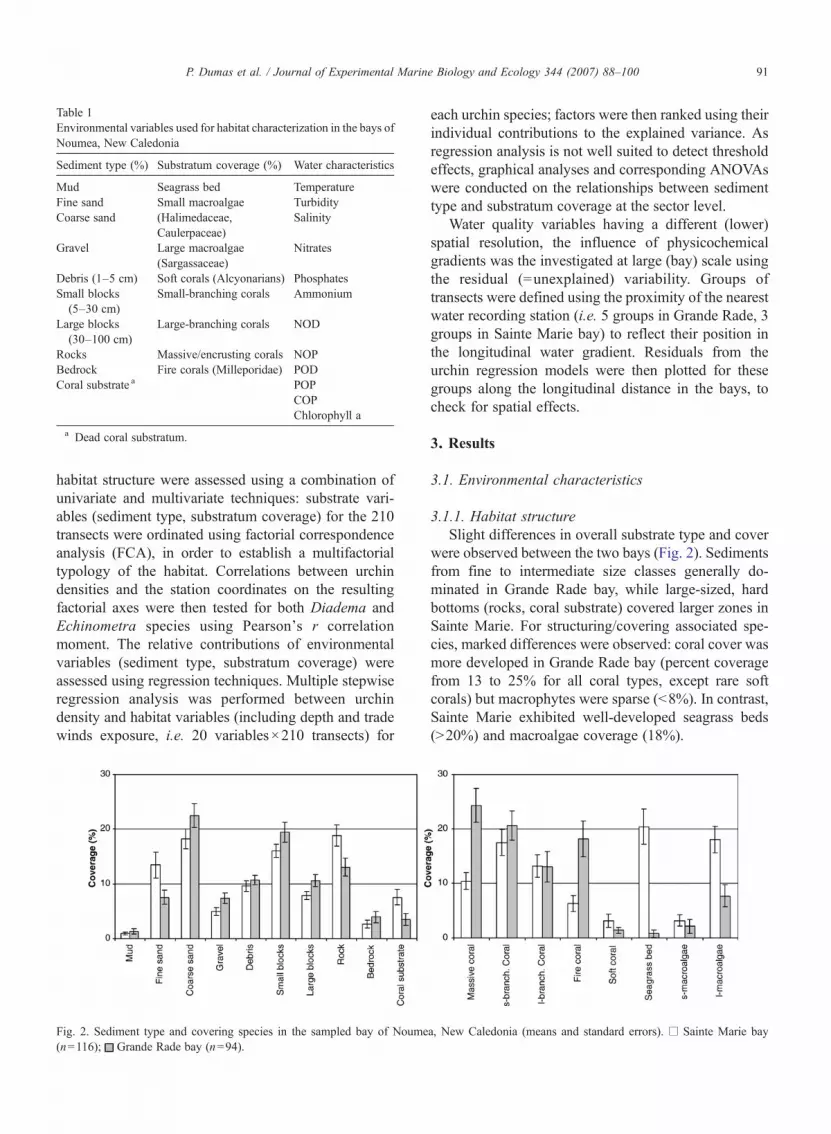
Table 1Environmental variables used for habitat characterization in the bays ofNoumea, New Caledonia
Sediment type (%) Substratum coverage (%) Water characteristics
Mud Seagrass bed TemperatureFine sand Small macroalgae TurbidityCoarse sand (Halimedaceae,
Caulerpaceae)Salinity
Gravel Large macroalgae(Sargassaceae)
Nitrates
Debris (1–5 cm) Soft corals (Alcyonarians) PhosphatesSmall blocks
(5–30 cm)Small-branching corals Ammonium
Large blocks(30–100 cm)
Large-branching corals NOD
Rocks Massive/encrusting corals NOPBedrock Fire corals (Milleporidae) PODCoral substrate a POP
COPChlorophyll a
a Dead coral substratum.
91P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
habitat structure were assessed using a combination ofunivariate and multivariate techniques: substrate vari-ables (sediment type, substratum coverage) for the 210transects were ordinated using factorial correspondenceanalysis (FCA), in order to establish a multifactorialtypology of the habitat. Correlations between urchindensities and the station coordinates on the resultingfactorial axes were then tested for both Diadema andEchinometra species using Pearson's r correlationmoment. The relative contributions of environmentalvariables (sediment type, substratum coverage) wereassessed using regression techniques. Multiple stepwiseregression analysis was performed between urchindensity and habitat variables (including depth and tradewinds exposure, i.e. 20 variables×210 transects) for
Fig. 2. Sediment type and covering species in the sampled bay of Noume(n=116); Grande Rade bay (n=94).
each urchin species; factors were then ranked using theirindividual contributions to the explained variance. Asregression analysis is not well suited to detect thresholdeffects, graphical analyses and corresponding ANOVAswere conducted on the relationships between sedimenttype and substratum coverage at the sector level.
Water quality variables having a different (lower)spatial resolution, the influence of physicochemicalgradients was the investigated at large (bay) scale usingthe residual (=unexplained) variability. Groups oftransects were defined using the proximity of the nearestwater recording station (i.e. 5 groups in Grande Rade, 3groups in Sainte Marie bay) to reflect their position inthe longitudinal water gradient. Residuals from theurchin regression models were then plotted for thesegroups along the longitudinal distance in the bays, tocheck for spatial effects.
3. Results
3.1. Environmental characteristics
3.1.1. Habitat structureSlight differences in overall substrate type and cover
were observed between the two bays (Fig. 2). Sedimentsfrom fine to intermediate size classes generally do-minated in Grande Rade bay, while large-sized, hardbottoms (rocks, coral substrate) covered larger zones inSainte Marie. For structuring/covering associated spe-cies, marked differences were observed: coral cover wasmore developed in Grande Rade bay (percent coveragefrom 13 to 25% for all coral types, except rare softcorals) but macrophytes were sparse (b8%). In contrast,Sainte Marie exhibited well-developed seagrass beds(N20%) and macroalgae coverage (18%).
a, New Caledonia (means and standard errors). □ Sainte Marie bay
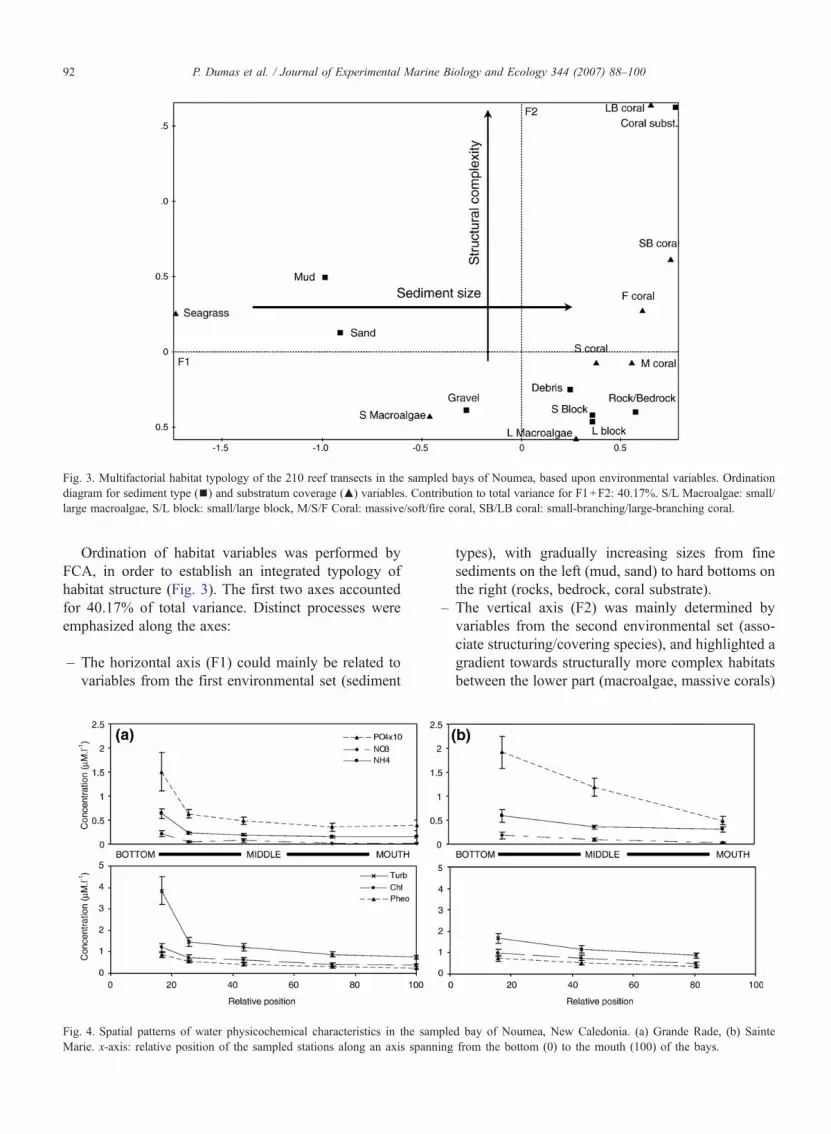
Fig. 3. Multifactorial habitat typology of the 210 reef transects in the sampled bays of Noumea, based upon environmental variables. Ordinationdiagram for sediment type (▪) and substratum coverage (▴) variables. Contribution to total variance for F1+F2: 40.17%. S/L Macroalgae: small/large macroalgae, S/L block: small/large block, M/S/F Coral: massive/soft/fire coral, SB/LB coral: small-branching/large-branching coral.
92 P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
Ordination of habitat variables was performed byFCA, in order to establish an integrated typology ofhabitat structure (Fig. 3). The first two axes accountedfor 40.17% of total variance. Distinct processes wereemphasized along the axes:
– The horizontal axis (F1) could mainly be related tovariables from the first environmental set (sediment
Fig. 4. Spatial patterns of water physicochemical characteristics in the samMarie. x-axis: relative position of the sampled stations along an axis spann
types), with gradually increasing sizes from finesediments on the left (mud, sand) to hard bottoms onthe right (rocks, bedrock, coral substrate).
– The vertical axis (F2) was mainly determined byvariables from the second environmental set (asso-ciate structuring/covering species), and highlighted agradient towards structurally more complex habitatsbetween the lower part (macroalgae, massive corals)
pled bay of Noumea, New Caledonia. (a) Grande Rade, (b) Sainteing from the bottom (0) to the mouth (100) of the bays.
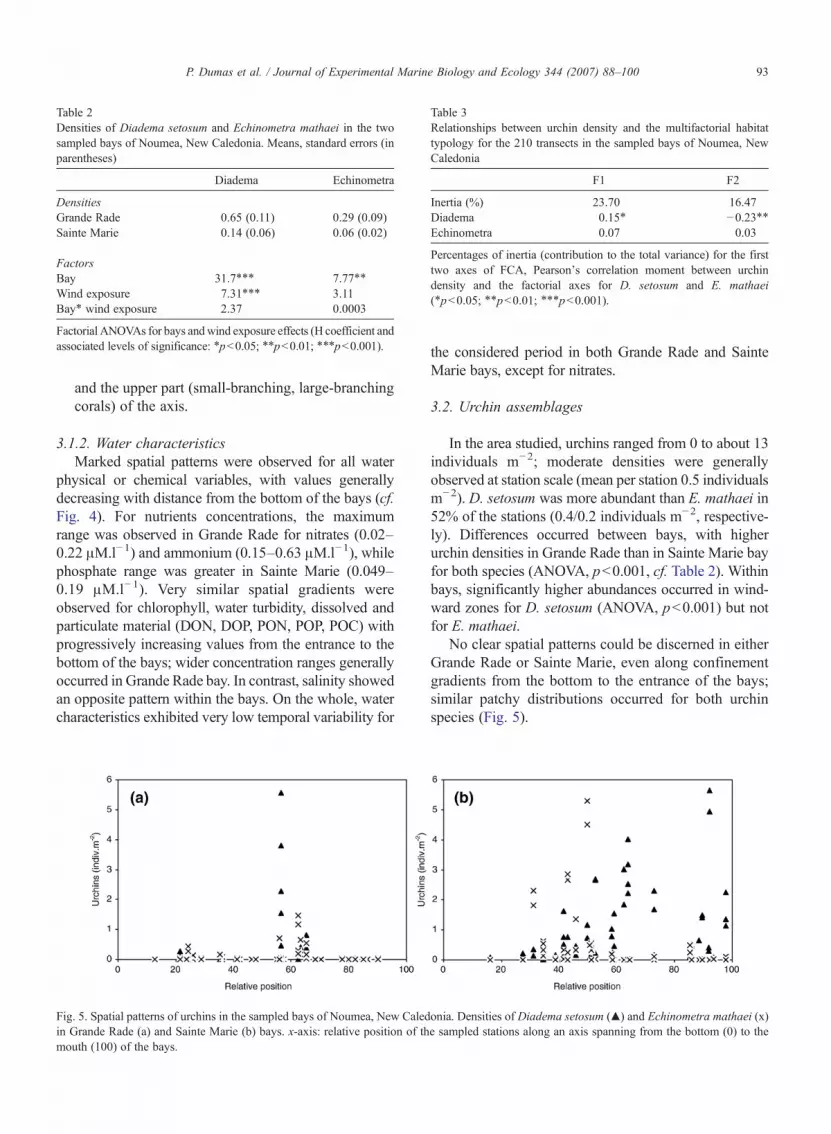
Table 3Relationships between urchin density and the multifactorial habitattypology for the 210 transects in the sampled bays of Noumea, NewCaledonia
F1 F2
Inertia (%) 23.70 16.47Diadema 0.15⁎ −0.23⁎⁎Echinometra 0.07 0.03
Percentages of inertia (contribution to the total variance) for the firsttwo axes of FCA, Pearson's correlation moment between urchindensity and the factorial axes for D. setosum and E. mathaei(⁎pb0.05; ⁎⁎pb0.01; ⁎⁎⁎pb0.001).
Table 2Densities of Diadema setosum and Echinometra mathaei in the twosampled bays of Noumea, New Caledonia. Means, standard errors (inparentheses)
Diadema Echinometra
DensitiesGrande Rade 0.65 (0.11) 0.29 (0.09)Sainte Marie 0.14 (0.06) 0.06 (0.02)
FactorsBay 31.7⁎⁎⁎ 7.77⁎⁎
Wind exposure 7.31⁎⁎⁎ 3.11Bay⁎ wind exposure 2.37 0.0003
Factorial ANOVAs for bays andwind exposure effects (H coefficient andassociated levels of significance: ⁎pb0.05; ⁎⁎pb0.01; ⁎⁎⁎pb0.001).
93P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
and the upper part (small-branching, large-branchingcorals) of the axis.
3.1.2. Water characteristicsMarked spatial patterns were observed for all water
physical or chemical variables, with values generallydecreasing with distance from the bottom of the bays (cf.Fig. 4). For nutrients concentrations, the maximumrange was observed in Grande Rade for nitrates (0.02–0.22 μM.l−1) and ammonium (0.15–0.63 μM.l−1), whilephosphate range was greater in Sainte Marie (0.049–0.19 μM.l−1). Very similar spatial gradients wereobserved for chlorophyll, water turbidity, dissolved andparticulate material (DON, DOP, PON, POP, POC) withprogressively increasing values from the entrance to thebottom of the bays; wider concentration ranges generallyoccurred in Grande Rade bay. In contrast, salinity showedan opposite pattern within the bays. On the whole, watercharacteristics exhibited very low temporal variability for
Fig. 5. Spatial patterns of urchins in the sampled bays of Noumea, New Caledin Grande Rade (a) and Sainte Marie (b) bays. x-axis: relative position of thmouth (100) of the bays.
the considered period in both Grande Rade and SainteMarie bays, except for nitrates.
3.2. Urchin assemblages
In the area studied, urchins ranged from 0 to about 13individuals m−2; moderate densities were generallyobserved at station scale (mean per station 0.5 individualsm−2). D. setosum was more abundant than E. mathaei in52% of the stations (0.4/0.2 individuals m−2, respective-ly). Differences occurred between bays, with higherurchin densities in Grande Rade than in Sainte Marie bayfor both species (ANOVA, pb0.001, cf. Table 2). Withinbays, significantly higher abundances occurred in wind-ward zones for D. setosum (ANOVA, pb0.001) but notfor E. mathaei.
No clear spatial patterns could be discerned in eitherGrande Rade or Sainte Marie, even along confinementgradients from the bottom to the entrance of the bays;similar patchy distributions occurred for both urchinspecies (Fig. 5).
onia. Densities of Diadema setosum (▴) and Echinometra mathaei (x)e sampled stations along an axis spanning from the bottom (0) to the
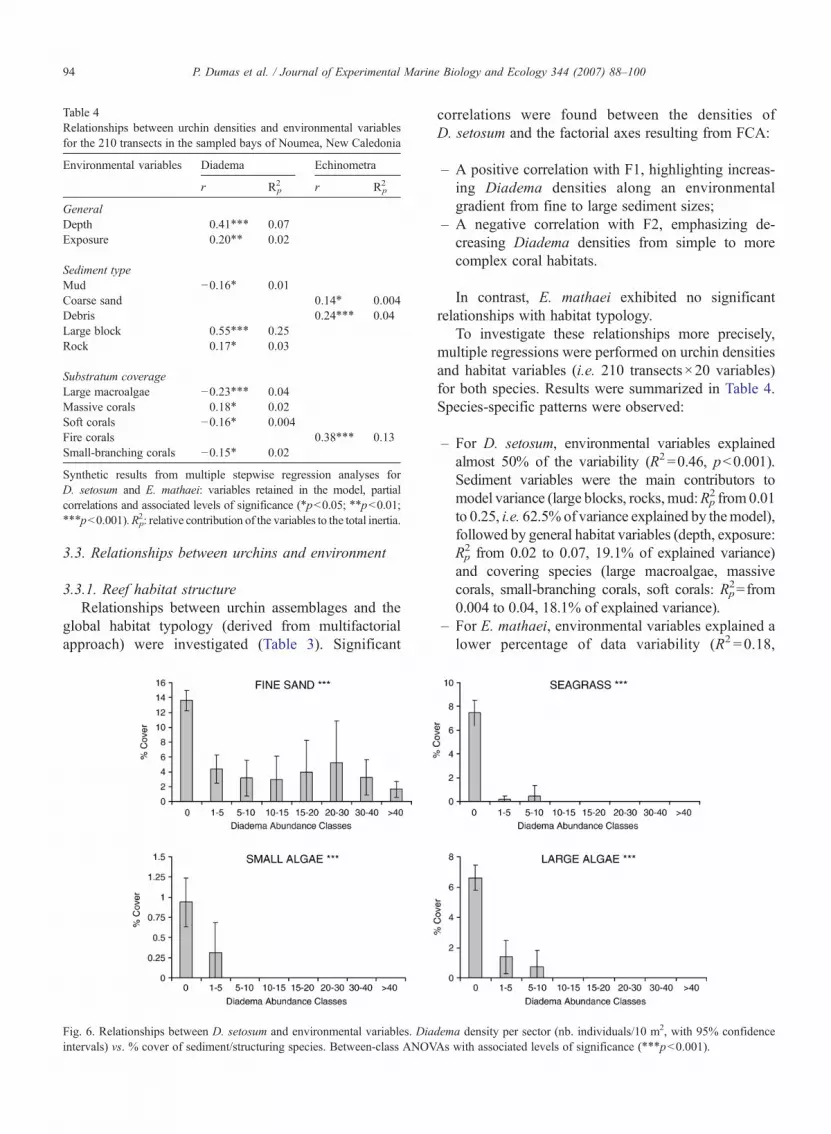
Table 4Relationships between urchin densities and environmental variablesfor the 210 transects in the sampled bays of Noumea, New Caledonia
Environmental variables Diadema Echinometra
r R2p r R2
p
GeneralDepth 0.41⁎⁎⁎ 0.07Exposure 0.20⁎⁎ 0.02
Sediment typeMud −0.16⁎ 0.01Coarse sand 0.14⁎ 0.004Debris 0.24⁎⁎⁎ 0.04Large block 0.55⁎⁎⁎ 0.25Rock 0.17⁎ 0.03
Substratum coverageLarge macroalgae −0.23⁎⁎⁎ 0.04Massive corals 0.18⁎ 0.02Soft corals −0.16⁎ 0.004Fire corals 0.38⁎⁎⁎ 0.13Small-branching corals −0.15⁎ 0.02
Synthetic results from multiple stepwise regression analyses forD. setosum and E. mathaei: variables retained in the model, partialcorrelations and associated levels of significance (⁎pb0.05; ⁎⁎pb0.01;⁎⁎⁎pb0.001).R2p: relative contribution of the variables to the total inertia.
94 P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
3.3. Relationships between urchins and environment
3.3.1. Reef habitat structureRelationships between urchin assemblages and the
global habitat typology (derived from multifactorialapproach) were investigated (Table 3). Significant
Fig. 6. Relationships between D. setosum and environmental variables. Diaintervals) vs. % cover of sediment/structuring species. Between-class ANOV
correlations were found between the densities ofD. setosum and the factorial axes resulting from FCA:
– A positive correlation with F1, highlighting increas-ing Diadema densities along an environmentalgradient from fine to large sediment sizes;
– A negative correlation with F2, emphasizing de-creasing Diadema densities from simple to morecomplex coral habitats.
In contrast, E. mathaei exhibited no significantrelationships with habitat typology.
To investigate these relationships more precisely,multiple regressions were performed on urchin densitiesand habitat variables (i.e. 210 transects×20 variables)for both species. Results were summarized in Table 4.Species-specific patterns were observed:
– For D. setosum, environmental variables explainedalmost 50% of the variability (R2=0.46, pb0.001).Sediment variables were the main contributors tomodel variance (large blocks, rocks,mud:R2
p from 0.01to 0.25, i.e. 62.5% of variance explained by themodel),followed by general habitat variables (depth, exposure:R2p from 0.02 to 0.07, 19.1% of explained variance)
and covering species (large macroalgae, massivecorals, small-branching corals, soft corals: R2
p=from0.004 to 0.04, 18.1% of explained variance).
– For E. mathaei, environmental variables explained alower percentage of data variability (R2 =0.18,
dema density per sector (nb. individuals/10 m2, with 95% confidenceAs with associated levels of significance (⁎⁎⁎pb0.001).
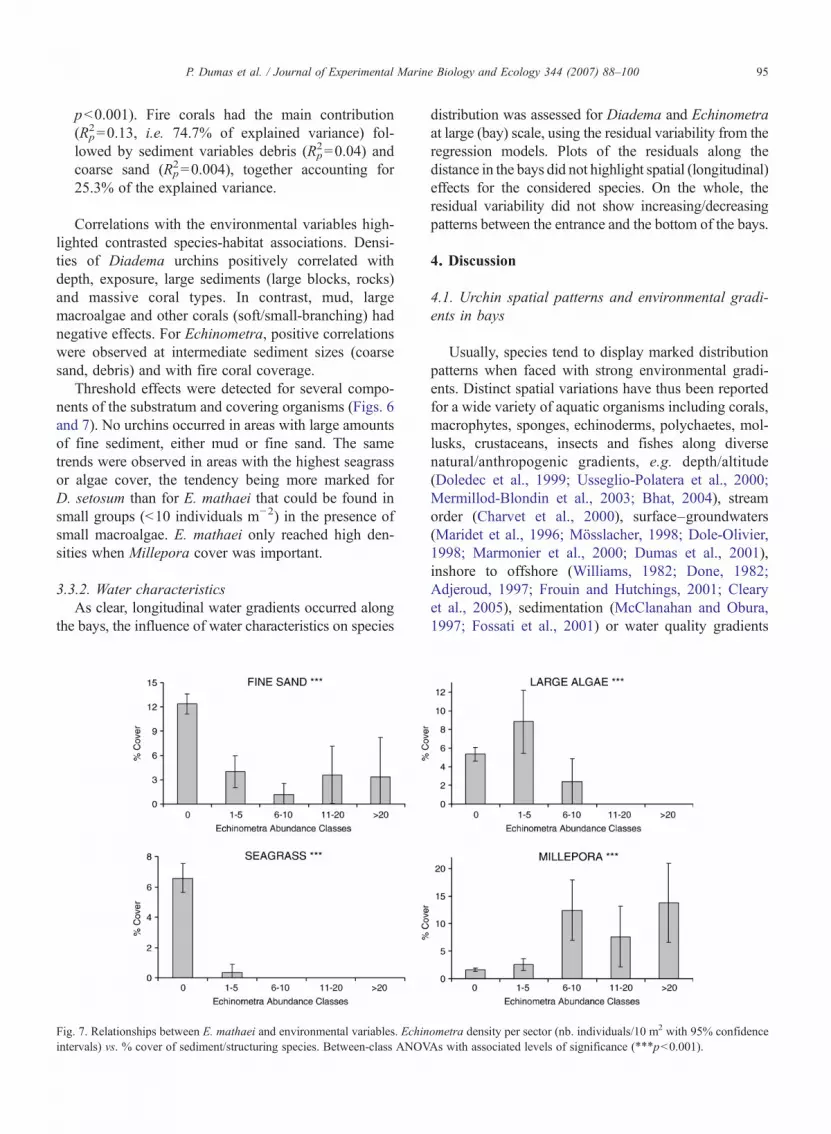
95P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
pb0.001). Fire corals had the main contribution(R2
p=0.13, i.e. 74.7% of explained variance) fol-lowed by sediment variables debris (R2
p=0.04) andcoarse sand (R2
p=0.004), together accounting for25.3% of the explained variance.
Correlations with the environmental variables high-lighted contrasted species-habitat associations. Densi-ties of Diadema urchins positively correlated withdepth, exposure, large sediments (large blocks, rocks)and massive coral types. In contrast, mud, largemacroalgae and other corals (soft/small-branching) hadnegative effects. For Echinometra, positive correlationswere observed at intermediate sediment sizes (coarsesand, debris) and with fire coral coverage.
Threshold effects were detected for several compo-nents of the substratum and covering organisms (Figs. 6and 7). No urchins occurred in areas with large amountsof fine sediment, either mud or fine sand. The sametrends were observed in areas with the highest seagrassor algae cover, the tendency being more marked forD. setosum than for E. mathaei that could be found insmall groups (b10 individuals m−2) in the presence ofsmall macroalgae. E. mathaei only reached high den-sities when Millepora cover was important.
3.3.2. Water characteristicsAs clear, longitudinal water gradients occurred along
the bays, the influence of water characteristics on species
Fig. 7. Relationships between E. mathaei and environmental variables. Echinintervals) vs. % cover of sediment/structuring species. Between-class ANOV
distribution was assessed for Diadema and Echinometraat large (bay) scale, using the residual variability from theregression models. Plots of the residuals along thedistance in the bays did not highlight spatial (longitudinal)effects for the considered species. On the whole, theresidual variability did not show increasing/decreasingpatterns between the entrance and the bottom of the bays.
4. Discussion
4.1. Urchin spatial patterns and environmental gradi-ents in bays
Usually, species tend to display marked distributionpatterns when faced with strong environmental gradi-ents. Distinct spatial variations have thus been reportedfor a wide variety of aquatic organisms including corals,macrophytes, sponges, echinoderms, polychaetes, mol-lusks, crustaceans, insects and fishes along diversenatural/anthropogenic gradients, e.g. depth/altitude(Doledec et al., 1999; Usseglio-Polatera et al., 2000;Mermillod-Blondin et al., 2003; Bhat, 2004), streamorder (Charvet et al., 2000), surface–groundwaters(Maridet et al., 1996; Mösslacher, 1998; Dole-Olivier,1998; Marmonier et al., 2000; Dumas et al., 2001),inshore to offshore (Williams, 1982; Done, 1982;Adjeroud, 1997; Frouin and Hutchings, 2001; Clearyet al., 2005), sedimentation (McClanahan and Obura,1997; Fossati et al., 2001) or water quality gradients
ometra density per sector (nb. individuals/10 m2 with 95% confidenceAs with associated levels of significance (⁎⁎⁎pb0.001).

96 P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
(Gray, 1979; Thorne and Williams, 1997; Rees et al.,1999; Cardell et al., 1999; Arana et al., 2005; Houket al., 2005; Habit et al., 2006).
In the area studied, two main environmental gradientswere observed along a longitudinal axis spanning fromthe entrance to the bottom of the bays, as a result ofdistinct physicochemical processes:
a) a confinement gradient reflecting oceanic vs. terres-trial influences, from clear/saline lagoon waters tomore turbid, slightly desalinated inshore waters, alsocharacterized by higher water residence times (Jouonet al., 2006);
b) a disturbance gradient reflecting anthropic influences,with increasing nutrient/organic matter/chlorophyllconcentrations emphasizing a progressive degrada-tion in water quality, in particular anthropogeniceutrophication.
On the whole, patchy distributions were observed forreef urchins that could not easily be related to the lattergradients. In particular, the densities of Diadema andEchinometra did not show clear increasing or decreasingpatterns between the entrance and the bottom of the bays,either for Grande Rade or Sainte Marie. While no urchinswere observed in the very bottom of both bays, highdensities punctually occurred in the middle of SainteMarie at a very limited number of stations while GrandeRade showed more scattered longitudinal distributions.Similar trends were also reported along two Polynesianbays, where the abundance of reef urchins exhibitedcomplex spatial patterns that could not easily be related toenvironmental gradients (Adjeroud, 1997, 2000). Thesefindings raise the problem of multiple environmentalfactors occurring as inter-related gradients in reef habitats,making it difficult to elucidate the resulting speciesdistributions. This is of particular importance for urchins,whose densities are known to vary over short distancesand patchiness to increase with the considered spatialscales: McClanahan and Muthiga (2001) thus reportedspatial variability of Echinometra from less than 50%within reef sites to more than 150% between reefs.
4.2. Habitat characteristics influencing reef urchindensities
Among the factors influencing species distributions incoral reefs, prevalence of habitat structure may be ex-pected for benthic species exhibiting close linkage withsubstrate, e.g. urchins. Ranking based upon contributionscores allowed us to assess the relative importance ofhabitat-related variables to the urchin spatial variability.
Our results emphasized contrasted, species-specificpatterns for Diadema and Echinometra. For Diadema, asignificant part (46%) of the spatial variability wasexplained by habitat variables, suggesting that thephysical nature of substrate exerts a prevailing influenceon spatial distributions. From the initial data set (20variables), 9 variables exhibited significant contribu-tions to the regression model; among the latter, only 4had major contributions: large blocks (30–100 cm) androck (N100 cm) coverage, depth, and large macroalgaecover, together accounting for 39% of the totalvariability. On the whole, the spatial variability ofDiadema was better explained by sediment type (i.e.substrate grain sizes) than by biotic cover (i.e. presenceof structuring/covering species). Bearing in mind thatcorrelation may not necessarily implies direct causalrelationships, these results highlight the close physi-cal linkages between reef urchins and substratum, as aprobable consequence of their particular life-historytraits, e.g. feeding/locomotory behavior or shelteravailability (McClanahan and Kurtis, 1991; Sala et al.,1998; McClanahan and Muthiga, 2001). They alsoprovide interesting insights into habitat preferences forDiadema. Individuals were thus preferentially foundon exposed, rocky bottoms exhibiting moderate spatialheterogeneity (30–100 cm size-class blocks) andtypically avoided areas combining coral reefs and finesediments beds (fine sand/mud N10%). Lower urchindensity was also observed in zones with complex coralcover (branching corals), suggesting that Diadema mayavoid reef habitats exhibiting high architectural com-plexity. Opposite distributions were found betweenDiadema and macrophytes: no urchins were observed inzones with developed macrophyte coverage (seagrass/large macroalgae N6%, small macroalgae N1%);conversely, stations with average density of DiademaN1/m2 had no macrophytes. The mechanisms underly-ing these patterns remain uncertain in the context of thisstudy; yet, conflicting hypotheses may be proposed: a)reef zones dominated by macrophytes are unsuitablebiotopes for urchins; b) benthic over-grazing by urchinsleading to macrophytes decline. The latter hypothesis,however, appears hazardous at least for large macro-algae (Sargassaceae): previous studies on urchin grazingpreferences showed that large, Sargassum algae werelow preference foods due to chemical compoundssuch as tannins and phenols that act as repellents forherbivores (Haley and Solandt, 2001; Solandt andCampbell, 2001).
For Echinometra, only a restricted part of the spatialvariability was explained by habitat variables (18%).These resultsmay have important ecological implications,

97P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
since they suggest that substrate characteristics may be oflow predictive value for reef urchin distributions. Incontrast with Diadema, sediment types had the lowestcontributions (cumulated contributions b5%) whereascoral cover had the main contribution through a singlevariable (coverage of fire corals, 13%). Inferringecological processes from these results may be hazardousat this point, butMillepora could provide shelters againstpredation for Echinometra urchins as: a) cavities are largeenough for urchins but too small for predators; b) firecorals may repulse urchin predators, in particular fishes.
4.3. Spatial distribution patterns of reef urchins
While a certain part of urchin distributions could belinked to environmental variables, most of the spatialvariability remained unaccounted for, in particular forEchinometra. Similar trends were also highlighted byother authors, with high spatial patchiness reported forsea urchins as the result of diverse factors interacting atmultiple scales (e.g. Sanchez-Jerez et al., 2001).Biological processes that also influence urchins'patterns were not considered in this study that focusedon environmental habitat factors. Sea urchin recruitmentoften exhibits significant differences across space, thuspatchiness of settling larvae may occur at very shortspatial scales (e.g. hundreds of meters, Sala and Zabala,1996). Biotic interactions are also considered as majordriving forces for urchin assemblages, and spatialheterogeneity was positively related to predation levelsor the degree of agonistic/aggregating behavior in somestudies (Tsuchiya and Nishihira, 1985; Tsuchiya et al.,1991; Sala and Zabala, 1996; McClanahan, 1998;Pinnegar et al., 2000; McClanahan and Muthiga,2001). Inter-specific competition for space and/or foodmay also occur; yet, in the context of our results, therewas no evidence for it, at least at the stations level: thetwo species coexisted in some stations, while a singlespecies was observed in others. Introducing urchindensities in the model did not allow us to detect density-dependence effects. Other, post-settlement environmen-tal factors such as food and shelter availability are alsoevoked in the literature (McClanahan and Muthiga,2001; Sala et al., 1998) but were not formally testedhere. Yet, the strong affinities of Diadema for reef zoneswith medium-sized blocks that arise from our resultsprobably relates to the latter factor, as well as theiravoidance for biotopes with developed branching coralsor macroalgae.
Water quality is an essential point, as urchins aregenerally considered sensitive bioindicators of variouscontaminants (Flammang et al., 1997, Breau, 2003).
Acute toxicity of ammonium/heavy metals was inparticular reported for echinoid species, mainly at eggand larval stages (e.g. Cesar et al., 2002; Arizzi Novelliet al., 2003). The high levels of ammonium punctu-ally observed in both bays (N1 μM.l−1) might theninfluence urchins distributions, at least partly. Yet, whilemarked, longitudinal water quality gradients wereobserved along the bays, they did not seem to influenceDiadema or Echinometra distributions. Nevertheless,these results have to be considered with respect to thespatial scales, as physicochemical data were obtainedfrom a restricted number of stations (3 in Sainte Marie, 5in Grande Rade). Local inputs/physicochemical pro-cesses may thus influence invertebrates at smallerscales, below the resolution of water quality variables(e.g. station/transect/sector scales). Threshold responseto contaminants may then occur but may not be detectedin the bays, with most of the study area below thethreshold.
5. Conclusion
As a matter of fact, the spatial patchiness of reefurchins is usually not easy to explain (McClanahan andMuthiga, 2001). Conclusions from this quantitativestudy emphasized the importance of substrate character-istics in shaping the density distributions, with respect tospecies characteristics and environmental gradients.Investigating the densities of two common, widely-distributed urchin species over a wide range of stationsin contrasted reef habitats allowed us to highlightcontrasted species-habitat associations, and determinethe relative contributions of environmental habitatvariables to the observed spatial variability. In contrastwith D. setosum, it is apparent that the distribution ofE. mathaei was poorly explained by habitat character-istics, at least those considered in this study. Eventhough it may indicate the prevalence of additional (e.g.biological) factors structuring the populations, it mayalso raise the problem of taxonomic status for theconsidered species. Because of their wide distributionand spinal color variation, Echinometra have alwaysbeen a subject of controversy (Araraki and Uehara,1991). Recent morphological, biochemical, ecologicaland reproductive studies thus revealed that at least fourdistinct species of Echinometra (referred to as speciesA, B, C and D) exist in the Indo-West Pacific, withEchinometra sp. B being now recognized as E. mathaei(de Blainville, 1825) sensu stricto (see Rahman andUehara, 2004a,b). Yet, species confusion may occur asfield identification is particularly difficult, henceencompassing assemblages with distinct species-habitat

98 P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
relationships may explain the high amount of unex-plained variability observed for Echinometra.
Finally, the question of study scales is of particularimportance in this context. Our results suggest that strongvariations in urchin densities occur across spatial scales(e.g. between sector, 10 m2/station, 50 m2/reef scale), atrend that complicates the interpretation of abundancepatterns. In complex systems such as coral reefs, small-scale heterogeneity may thus be responsible for most ofthe observed variability and adequate study scales aretherefore required to avoid misinterpretations of what isobserved at larger scales. Temporal variability should alsobe considered, as short-scale variations of urchin densitiesmay have occurred within the study period, thereforeadding complexity to the resulting patterns. Reef urchinscan achieve high population densities, hence playing amajor role in coral ecosystems through biotic processesincluding bioerosion and benthic grazing. Understandingthe mechanisms that control their populations is achallenging task with important consequences in termsof reef habitat management, in particular in the face ofincreasing environmental changes.
Acknowledgements
This work is part of the UR 128-CoReUs program.Thanks to the anonymous reviewers for their usefulcomments. [SS]
References
Adjeroud, M., 1997. Factors influencing spatial patterns on coral reefsaroundMoorea, FrenchPolynesia.Mar. Ecol. Prog. Ser. 159, 105–119.
Adjeroud, M., 2000. Zonation of macrobenthic communities alongtwo bays in an insular coral reef ecosystem (Moorea, FrenchPolynesia). C. R. Acad. Sci. 323 (3), 305–313.
Alongi, D.M., 1990. The ecology of tropical soft-bottom benthicecosystems. Oceanogr. Mar. Biol. 28, 381–496.
Alongi, D.M., Sasekumar, A., 1992. Benthic communities. In: Robertson,A.I., Alongi, D.M. (Eds.), Tropical Mangrove Ecosystems. AmericanGeophysical Union, Washington, D.C., pp. 137–171.
Arana, H.A.H., Warwick, R.M., Attrill, M.J., Rowden, A.A., Gold-Bouchot, G., 2005. Assessing the impact of oil-related activities onbenthic macroinfauna assemblages of the Campeche Shelf,Southern Gulf of Mexico. Mar. Ecol. Prog. Ser. 289, 89–107.
Araraki,Y.,Uehara, T., 1991. Physiological adaptation and reproduction ofthe four types of Echinometra mathaei (Blainville). In: Yanagisawa,T., Yasumasu, I., Oguro, C., Suzuki, N., Motokawa, T. (Eds.),Biology of Echinodermata. Balkema, Rotterdam, pp. 105–111.
Arizzi Novelli, A., Picone, M., Losso, C., Volpi Ghirardini, A., 2003.Ammonia as confounding factor in toxicity tests with the sea urchinParacentrotus lividus (Lmk). Toxicol. Environ. Chem. 85 (4–6),183–191.
Balvay,G., 1993. Space partitionwithin aquatic ecosystems. Proceedingsof the Second International Congress of Limnology and Oceanog-raphy. Kluwer Academic Publishers, Evian, France.
Bhat, A., 2004. Patterns in the distribution of freshwater fishes in riversof central western Ghats, India and their associations withenvironmental gradients. Hydrobiologia 529 (1), 83–97.
Bozec, Y.M., Doledec, S., Kulbicki, M., 2005. An analysis of fish-habitat associations on disturbed coral reefs: chaetodontid fishes inNew Caledonia. J. Fish Biol. 66 (4), 966–982.
Breau, L. 2003. Etude de la bioaccumulation des métaux dansquelques espèces marines tropicales: Recherche de bioindicateursde contamination et application à la surveillance de l'environne-ment côtier dans le lagon sud ouest de la Nouvelle-Calédonie. Phd-thesis, University of la Rochelle, France, 366 pp.
Burke, L., Kura, Y., Kassem, K., Revenga, C., Spalding, M.,McAllister, D., 2001. Pilot Analysis of Global Ecosystems:Coastal Ecosystems. World Resources Institute, Washington, D.C.
Cardell, M.J., Sarda, R., Romero, J., 1999. Spatial changes insublittoral soft-bottom polychaete assemblages due to river inputsand sewage discharges. Acta Oecol. 20 (4), 343–351.
Cesar, A., Marin-Guirao, L., Vita, R., Marin, A., 2002. Sensitivity ofMediterranean amphipods and sea urchins to reference toxicants.Cienc. Mar. 28 (4), 407–417.
Chabanet, P., Ralambondrainy, H., Amanieu, M., Faure, G., Galzin, R.,1997. Relationships between coral reef substrata and fish. CoralReefs 16 (2), 93–102.
Charvet, S., Statzner, B., Usseglio-Polatera, P., Dumont, B., 2000.Traits of benthic macroinvertebrates in semi-natural Frenchstreams: an initial application to biomonitoring in Europe. Freshw.Biol. 43 (2), 277–296.
Chiappone, M., Swanson, D.W., Miller, S.L., 2002. Density, spatialdistribution and size structure of sea urchins in Florida Keys coralreef and hard-bottom habitats. Mar. Ecol. Prog. Ser. 235, 117–126.
Cleary, D.F.R., Becking, L.E., De Voogd, N.J., Renema, W., De Beer,M., Van Soest, R.W.M., Hoeksema, B.W., 2005. Variation in thediversity and composition of benthic taxa as a function of distanceoffshore, depth and exposure in the Spermonde archipelago,Indonesia. Estuar. Coast. Shelf Sci. 65 (3), 557–570.
Dole-Olivier, M.J., 1998. Surface water–groundwater exchanges inthree dimensions on a backwater of the Rhône River. Freshw. Biol.93–109.
Doledec, S., Statzner, B., Bournard, M., 1999. Species traits for futurebiomonitoring across ecoregions: patterns along a human-impactedriver. Freshw. Biol. 42 (4), 737–758.
Done, T.J., 1982. Patterns in the distribution of coral communitiesacross the central Great Barrier reef. Coral Reefs 1, 95–107.
Done, T., Ogden, J.,Wiebe,W., Rosen,B., 1996. Biodiversity and ecosystemfunction in coral reefs. In: Mooney, H., Cushman, H., Medina, E., Sala,O., Schulze, E.-D. (Eds.), Functional Roles of Biodiversity: a GlobalPerspective. John Wiley & Sons Inc., pp. 394–429.
Douillet, P., Ouillon, S., Cordier, E., 2001. A numerical model for finesuspended sediment transport in the southwest lagoon of NewCaledonia. Coral Reefs 20 (4), 361–372.
Dumas, P., Bou, C., Gibert, J., 2001. Groundwater macrocrustaceansas natural indicators of the Ariege alluvial aquifer. Int. Rev.Hydrobiol. 86 (6), 619–633.
Fichez, R., Chifflet, S., Douillet, P., Gérard, P., Ouillon, S., submittedfor publication. Biogeochemical typology and temporal variabilityof lagoon waters in a coral reef ecosystem subject to terrigeneousand anthropogenic inputs (New Caledonia). Mar. Pollut. Bull.
Flammang, P., Warnau, M., Temara, A., Lane, D.J.W., Jangoux, M.,1997. Heavy metals in Diadema setosum (Echinodermata, Echinoi-dea) from Singapore coral reefs. J. Sea Res. 38 (1–2), 35–45.
Fossati, O., Wasson, J.G., Hery, C., Salinas, G., Marin, R., 2001. Impactof sediment releases on water chemistry and macroinvertebrate

99P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
communities in clear water Andean streams (Bolivia). Arch.Hydrobiol. 151 (1), 33–50.
Frouin, P., Hutchings, P., 2001. Macrobenthic communities in atropical lagoon (Tahiti, French Polynesia, central Pacific). CoralReefs 19 (3), 277–285.
Gibert, J., Danielopol, D.L., Stanford, J., 1994. Groundwater Ecology.Academic Press, San Diego.
Gray, J.S., 1974. Animal–sediment relationships. Oceanogr. Mar. Biol.12, 223–261.
Gray, J.S., 1979. Pollution-induced changes in populations. Philos.Trans. R. Soc. Lond., B 286, 545–561.
Grigg, R.W., Dollar, S.J., 1990. Natural and anthropogenic disturbanceon coral reefs. In: Dubinsky, Z. (Ed.), Ecosystems of the World:Coral Reefs. Elsevier, pp. 439–453.
Habit, E., Belk, M.C., Tuckfield, R.C., Parra, O., 2006. Response ofthe fish community to human-induced changes in the Biobio riverin Chile. Freshw. Biol. 51 (1), 1–11.
Haley, M.P., Solandt, J.L., 2001. Population fluctuations of the seaurchins Diadema antillarum and Tripneustes ventricosus atDiscovery Bay, Jamaica: a case of biological succession? Caribb.J. Sci. 37 (3–4), 239–245.
Healy, T., Wang, Y., 2004. Integrated coastal zone management forsustainable development—with comment on ICZM applicabilityto muddy coasts. J. Coast. Res. 229–242.
Herman, P.M.J., Middelburg, J.J., VanDeKoppel, J., Heip, C.H.R., 1999.Ecology of estuarine macrobenthos. Adv. Ecol. Res. 29, 195–240.
Hildrew, A.G., Townsend, C.R., 1987. Organization in freshwaterbenthic communities. In: Gee, J.H.R., Giller, P.S. (Eds.), Organisa-tion of Communities: Past and Present. Blackwell Scientific, Oxford,pp. 347–371.
Hodgson, G., 1999. A global assessment of human effects on coralreefs. Mar. Pollut. Bull. 38, 345–355.
Houk, P., Didonato, G., Iguel, J., Van Woesik, R., 2005. Assessing theeffects of non-point source pollution on American Samoa's coralreef communities. Environ. Monit. Assess. 107 (1–3), 11–27.
Hughes, T.P., Connell, J.H., 1999. Multiple stressors on coral reefs:a long-term perspective. Limnol. Oceanogr. 44 (3), 932–940.
Jackson, J.B.C., Kirby, M.X., Berger, W.H., Bjorndal, K.A., Botsford,L.W., Bourque, B.J., Bradbury, R.H., Cooke, R., Erlandson, J.,Estes, J.A., Hughes, T.P., Kidwell, S., Lange, C.B., Lenihan, H.S.,Pandolfi, J.M., Peterson, C.H., Steneck, R.S., Tegner, M.J.,Warner, R.R., 2001. Historical overfishing and the recent collapseof coastal ecosystems. Science 293 (5530), 629–638.
Jouon, A., Douillet, P., Ouillon, S., Fraunié, P., 2006. Calculations ofhydrodynamic time parameters in a semi-opened coastal zoneusing a 3D hydrodynamic model. Cont. Shelf Res. 26 (12–13),1395–1415.
King, C.K., Riddle, M.J., 2001. Effects of metal contaminants on thedevelopment of the common Antarctic sea urchin Sterechinusneumayeri and comparisons of sensitivity with tropical andtemperate echinoids. Mar. Ecol. Prog. Ser. 215, 143–154.
Laine, A.O., 2003. Distribution of soft-bottom macrofauna in the deepopen Baltic sea in relation to environmental variability. Estuar.Coast. Shelf Sci. 57, 87–97.
Lawrence, J.M., 2001. Edible Sea Urchins: Biology and Ecology.Elsevier.
Lecchini, D., Adjeroud, M., Pratchett, M.S., Cadoret, L., Galzin, R.,2003. Spatial structure of coral reef fish communities in the RyukyuIslands, southern Japan. Oceanol. Acta 26 (5–6), 537–547.
Loya, Y., 1976. Effects of water turbidity and sedimentation on thecommunity structure of Puerto Rican corals. Bull. Mar. Sci. 26,450–466.
Luckhurst, B.E., Luckhurst, K., 1978. Analysis of the influence ofsubstrate variable on coral reef fish communities. Mar. Biol. 49,317–323.
Maridet, L., Philippe, M., Wasson, J.G., Mathieu, J., 1996. Spatial andtemporal distribution of macroinvertebrates and trophic variableswithin the bed sediment of three streams differing by their mor-phology and riparian vegetation. Arch. Hydrobiol. 136 (1), 41–64.
Marmonier, P., Claret, C., Dole-Olivier, M.J., 2000. Interstitial fauna innewly-created floodplain canals of a large regulated river. Regul.Rivers, Res. Manag. 16 (1), 23–36.
McClanahan, T.R., 1992. Epibenthic gastropods of the Middle Floridakeys: the role of habitat and environmental stress on assemblagecomposition. J. Exp. Mar. Biol. Ecol. 160, 169–190.
McClanahan, T.R., 1998. Predation and the distribution and abundanceof tropical sea urchin populations. J. Exp. Mar. Biol. Ecol. 221 (2),231–255.
McClanahan, T.R., Kurtis, J.D., 1991. Population regulation of therock-boring sea-urchin Echinometra mathaei (Deblainville).J. Exp. Mar. Biol. Ecol. 147 (1), 121–146.
McClanahan, T.R., Muthiga, N.A., 2001. The ecology of Echinome-tra. In: Lawrence, J.M. (Ed.), Edible Sea Urchins. Elsevier,Amsterdam, pp. 225–243.
McClanahan, T.R., Obura, D., 1997. Sedimentation effects on shallowcoral communities in Kenya. J. Exp. Mar. Biol. Ecol. 209 (1–2),103–122.
Mermillod-Blondin, F., Marie, S., Desrosiers, G., Long, B., DeMontety, L., Michaud, E., Stora, G., 2003. Assessment of thespatial variability of intertidal benthic communities by axialtomodensitometry: importance of fine-scale heterogeneity. J. Exp.Mar. Biol. Ecol. 287 (2), 193–208.
Mösslacher, F., 1998. Subsurface dwelling crustaceans as indicators ofhydrological conditions, oxygen concentration and sedimentstructure in an alluvial aquifer. Int. Rev. Hydrobiol. 83, 349–364.
Nanami, A., Saito, H., Akita, T., Motomatsu, K., Kuwahara, H., 2005a.Spatial distribution and assemblage structure of macrobenthicinvertebrates in a brackish lake in relation to environmentalvariables. Estuar. Coast. Shelf Sci. 63 (1–2), 167–176.
Nanami, A., Nishihira, M., Suzuki, T., Yokochi, H., 2005b. Species-specific habitat distribution of coral reef fish assemblages inrelation to habitat characteristics in an Okinawan coral reef.Environ. Biol. Fishes 72 (1), 55–65.
Nishihira, M., Sato, Y., Arakaki, Y., Tsuchiya, M., 1991. Ecologicaldistribution and habitat preference of four types of the sea urchinEchinometra mathaei on the Okinawan coral reefs. In: Yanagisawa,T., Yasumasu, I., Oguro, C., Suzuki, N., Motokawa, T. (Eds.),Biology of Echinodermata: Proceedings of the 7th InternationalEchinoderm Conference. Balkema, Rotterdam, pp. 91–104.
Ohlhorst, S.L., Liddell, W.D., Taylor, R.J., Taylor, J.M., 1988.Evaluation of reef census techniques. Proceedings of the 6thInternational Coral Reefs Symposium.
Pinnegar, J.K., Polunin, N.V.C., Francour, P., Badalamenti, F.,Chemello, R., Harmelin-Vivien, M., Hereu, B., Milazzo, M.,Zabala, M., D'Anna, G., Pipitone, C., 2000. Trophic cascades inbenthic marine ecosystems: lessons for fisheries and protected-areamanagement. Environ. Conserv. 27 (2), 179–200.
Rahman, M.A., Uehara, T., 2004a. Interspecific hybridization andbackcrosses between two sibling species of pacific sea urchins(genus Echinometra) on Okinawan intertidal reefs. Zool. Stud.43 (1), 93–111.
Rahman, S.M., Uehara, T., 2004b. Interspecific and intraspecificvariations in sibling species of sea urchin Echinometra. Comp.Biochem. Physiol., Part A Mol. Integr. Physiol. 139 (4), 469–478.

100 P. Dumas et al. / Journal of Experimental Marine Biology and Ecology 344 (2007) 88–100
Rasch, P.S., Ipsen, N., Malmgren-Hansen, A., Mogensen, B., 2005.Linking integrated water resources management and integratedcoastal zone management. Water Sci. Technol. 51 (11), 221–229.
Rees, J.G., Setiapermana, D., Sharp, V.A., Weeks, J.M., Williams, T.M.,1999. Evaluation of the impacts of land-based contaminants on thebenthic faunas of Jakarta Bay, Indonesia. Oceanol. Acta 22 (6),627–640.
Richmond, R.H., 1993. Coral reefs: present problems and futureconcerns resulting from anthropogenic disturbances. Am. Zool. 33,524–536.
Risk, M.J., 1972. Fish diversity on a coral reef in the Virgin Islands.Atoll Res. Bull. 193, 1–6.
Sala, E., Zabala, M., 1996. Fish predation and the structure of the seaurchin Paracentrotus lividus populations in the new Mediterra-nean. Mar. Ecol. Prog. Ser. 140 (1–3), 71–81.
Sala, E., Ribes, M., Hereu, B., Zabala, M., Alva, V., Coma, R.,Garrabou, J., 1998. Temporal variability in abundance of the seaurchins Paracentrotus lividus and Arbacia lixula in the northwest-ern Mediterranean: comparison between a marine reserve and anunprotected area. Mar. Ecol. Prog. Ser. 168, 135–145.
Sale, P.F., 1978. Coexistence of coral reef fishes a lottery for livingspace. Environ. Biol. Fishes 3, 85–102.
Sale, P.F., 1980. The ecology of fishes on coral reefs. Oceanogr. Mar.Biol. 18, 376–421.
Salvat, B., Richard, G., Salvat, F., Berigaud, R., Antoine, L., Berigaud,M.C., Plessis, Y., 1972. Moorea-Tiahura: Etude des peuplements dulagon et du récif. Ecole Pratique desHautes Etudes, Perpignan, France.
Sanchez-Jerez, P., Cesar, A., Cortez, F.S., Pereira, C.D.S., Silva, S.L.R.,2001. Spatial distribution of the most abundant sea urchinpopulations on the southeast coast of Sao Paulo (Brazil). Cienc.Mar. 27 (1), 139–153.
Schiel, D.R., Steinbeck, J.R., Foster, M.S., 2004. Ten years of inducedocean warming causes comprehensive changes in marine benthiccommunities. Ecology 85 (7), 1833–1839.
Solandt, J.L., Campbell, A.C., 2001. Macroalgal feeding character-istics of the sea urchin Diadema antillarum Philippi at DiscoveryBay, Jamaica. Caribb. J. Sci. 37 (3–4), 227–238.
Thorne, R.J., Williams, P., 1997. The response of benthic macro-invertebrates to pollution in developing countries: a multimetricsystem of bioassessment. Freshw. Biol. 37, 671–686.
Tsuchiya, M., Nishihira, M., 1985. Agonistic behavior and its effect onthe dispersion pattern in two types of the sea urchin, Echinometramathaei (Blainville). Galaxea 4, 37–48.
Tsuchiya, M., Nishihira, M., Sato, Y., 1991. Behavioral characteristicsof the sea urchin Echinometra mathaei (Blainville): agonisticbehavior and recolonization process on the depopulated reef flat.Biology of Echinodermata: Proceedings of the 7th InternationalEchinoderm Conference. Balkema, Rotterdam, pp. 113–118.
Usseglio-Polatera, P., Bournaud, M., Richoux, P., Tachet, H., 2000.Biological and ecological traits of benthic freshwater macroinver-tebrates: relationships and definition of groups with similar traits.Freshw. Biol. 43 (2), 175–205.
Vandel, A., 1964. Biospéologie: la biologie des animaux cavernicoles.Gauthier-Villars, Paris.
Vitousek, P.M., Mooney, H.A., Lubchenco, J., Melillo, J.M., 1997.Human domination of earth's ecosystems. Science 277 (5325),494–499.
Wilkinson, C.R., 1999. Global and local threats to coral reeffunctioning and existence: review and predictions. Mar. Freshw.Res. 50 (8), 867–878.
Williams, D.M., 1982. Patterns in the distribution of fish communitiesacross the central Great Barrier reef. Coral Reefs 1, 35–43.
Zuschin, M., Hohenegger, J., Steininger, F.F., 2001. Molluscanassemblages on coral reefs and associated hard substrata in thenorthern Red Sea. Coral Reefs 20 (2), 107–116.





























