Embed Size (px)
Citation preview

Published OnlineFirst on June 8, 2010 as 10.1158/0008-5472.CAN-09-4238
Tumor and Stem Cell Biology
CancerResearch
Effects of Ionizing Radiation on Self-Renewal andPluripotency of Human Embryonic Stem Cells
Kitchener D. Wilson1,2,3, Ning Sun2,3, Mei Huang2,3, Wendy Y. Zhang2,3, Andrew S. Lee2,3,Zongjin Li2,3, Shan X. Wang4, and Joseph C. Wu2,3,5
Abstract
Authors' A3MedicineEngineerinMedicine, S
Note: SupResearch
CorresponDrive, GranFax: 650-7
doi: 10.115
©2010 Am
www.aacr
Human embryonic stem cells (hESC) present a novel platform for in vitro investigation of the early embry-onic cellular response to ionizing radiation. Thus far, no study has analyzed the genome-wide transcriptionalresponse to ionizing radiation in hESCs, nor has any study assessed their ability to form teratomas, the de-finitive test of pluripotency. In this study, we use microarrays to analyze the global gene expression changes inhESCs after low-dose (0.4 Gy), medium-dose (2 Gy), and high-dose (4 Gy) irradiation. We identify genes andpathways at each radiation dose that are involved in cell death, p53 signaling, cell cycling, cancer, embryonicand organ development, and others. Using Gene Set Enrichment Analysis, we also show that the expression ofa comprehensive set of core embryonic transcription factors is not altered by radiation at any dose. Trans-plantation of irradiated hESCs to immune-deficient mice results in teratoma formation from hESCs irradiatedat all doses, definitive proof of pluripotency. Further, using a bioluminescence imaging technique, we havefound that irradiation causes hESCs to initially die after transplantation, but the surviving cells quickly recoverby 2 weeks to levels similar to control. To conclude, we show that similar to somatic cells, irradiated hESCssuffer significant death and apoptosis after irradiation. However, they continue to remain pluripotent and areable to form all three embryonic germ layers. Studies such as this will help define the limits for radiationexposure for pregnant women and also radiotracer reporter probes for tracking cellular regenerative therapies.Cancer Res; 70(13); OF1–10. ©2010 AACR.
Introduction
Ionizing radiation is a form of electromagnetic radiationproduced by X-ray machines, fluoroscopy, radioactive iso-topes, as well as nuclear environmental catastrophe. Inpregnant mothers undergoing diagnostic or therapeutic pro-cedures involving ionizing radiation, or who may be exposedto environmental radiation, there is a great potential fordamage to the early embryo (1, 2). The current consensusis that exposure to radiation of <0.05 Gy during pregnancyis not related to an elevated risk of malformation, and manydiagnostic procedures remain below this threshold (note thatGy is a unit of absorbed dose and reflects the amount ofenergy deposited into a mass of tissue; refs. 1, 3). However,these data are based on limited human data or on animalmodels and so may not accurately reflect the human embry-onic response to radiation.
ffiliations: Departments of 1Bioengineering, 2Radiology,, and 4Electrical Engineering and Materials Science andg and 5Institute for Stem Cell Biology and Regenerativetanford University, Stanford, California
plementary data for this article are available at CancerOnline (http://cancerres.aacrjournals.org/).
ding Author: Joseph C. Wu, Stanford University, 300 Pasteurt Building S140, Stanford, CA 94305. Phone: 650-736-2246;36-0234; E-mail: [email protected].
8/0008-5472.CAN-09-4238
erican Association for Cancer Research.
journals.org
With the growing number of imaging procedures thatuse ionizing radiation, such as X-rays, computed tomo-graphic (CT) scans (4–6), and positron emission tomogra-phy (PET) or single-photon emission CT, reporter probesthat monitor stem cell transplantation for regenerativeand antioncogenic therapies (7, 8), as well as concernsover terrorist attacks involving radioactive materials (9),there is a need to better understand the effects on humanembryonic stem cells (hESC). A number of reports havestudied both UV- and γ-irradiated mouse (10–12) and hu-man (13–17) embryonic stem cells, and have primarily fo-cused on the DNA damage response such as cell cycling,p53 signaling, and apoptosis. Only two (14, 17) have at-tempted to characterize the effects of radiation on the de-fining feature of hESCs: pluripotency, or ability to form allthree embryonic germ layers. However, these studies fo-cused on the expression of just two embryonic genes(Oct4 and Nanog) after irradiation, and none have per-formed teratoma studies to prove pluripotency.To address this lack of knowledge, we perform gene ex-
pression profiling of irradiated hESCs at three differentdoses, allowing us to analyze global pluripotency programsthat may be affected by radiation. Taking advantage ofnovel molecular imaging techniques to track hESC prolifer-ation, we also inject irradiated hESCs into mice and showthat despite a transient decrease in cellular proliferation
OF1

Wilson et al.
OF2
with the highest dose used in this study (4 Gy), these cellsare still able to ultimately form teratomas. Taken together,we present definitive proof that hESCs that survive irradi-ation up to 4 Gy are pluripotent.
Materials and Methods
hESC cultureUndifferentiated hESCs (H9 line from Wicell, passages
45–55) were grown on Matrigel-coated plates in mTeSR1 me-dium (Stem Cell Technologies) as previously described (18).Cell medium was changed daily and passaged approximatelyevery 4 to 6 days using collagenase IV. For cell counting,hESC colonies were digested to single cells with 0.05% trypsinEDTA and counted with a Countess Automated Cell Counter(Invitrogen).
IrradiationhESCs were irradiated with 0.4, 2, or 4 Gy of γ-radiation
using a cesium-137 irradiator. Immediately after irradiation,cells were returned to the incubator for recovery until theappropriate time point.
Reverse transcription-PCR18S was used as housekeeping gene control. The primer
sets used in the amplification reaction are as follows: humanCXCL10(IP10), 5′-CTGATTTGCTGCCTTATCTTTCT-3′ (for-ward) and 5′-ACATTTCCTTGCTAACTGCTTTC-3′ (reverse);human GADD45, 5′-TGGAGGAAGTGCTCAGCAAAGCC-3′(forward) and 5′-ACGCCTGGATCAGGGTGAAGTGG-3′ (re-verse); and human 18S, 5′-ACACGGACAGGATTGACAGA-3′(forward) and 5 ′-GGACATCTAAGGGCATCACAG-3′(reverse).
Immunohistochemical analysisForty-eight hours after irradiation, hESCs were fixed
with 2% formaldehyde in PBS for 2 minutes, permeabi-lized with 0.5% Triton X-100 in PBS for 10 minutes, andblocked with 5% bovine serum albumin in PBS for anhour. Cells were then stained with appropriate primaryantibodies and Alexa Fluor–conjugated secondary antibo-dies (Invitrogen). The primary antibodies for Oct4 (SantaCruz Biotechnology), Sox2 (Biolegend), Nanog (SantaCruz Biotechnology), SSEA4 (Chemicon), TRA-1-60 (Che-micon), and TRA-1-81 (Chemicon) were used in thestaining.
Annexin V flow cytometry analysisForty-eight hours after irradiation, hESCs were har-
vested and resuspended in binding buffer and stainedwith 5 μL of Annexin V–FITC and 5 μL of propidiumiodide (PI) using the Annexin V-FITC Apoptosis DetectionKit II (BD Pharmingen). The cell suspension was incubat-ed for 15 minutes at room temperature and analyzed ona FACScan flow cytometer (BD Biosciences). Flow cytome-try data were analyzed with FlowJo (Treestar) analysissoftware.
Cancer Res; 70(13) July 1, 2010
Microarray hybridization and data acquisitionTotal RNA samples were isolated in Trizol (Invitrogen) fol-
lowed by purification over an RNeasy column (Qiagen) fromhESCs 48 hours after irradiation. Three independent experi-ments for each radiation group plus control (for a total of12 unique samples) were harvested for RNA isolation. UsingAgilent Low RNA Input Fluorescent Linear Amplificationkits, cDNA was reverse transcribed from each of 12 RNAsamples representing four biological triplicates, as well asthe pooled reference control, and cRNA was then transcribedand fluorescently labeled with Cy5/Cy3. cRNA was purifiedusing an RNeasy kit. Cy3- and Cy5-labeled and amplifiedcRNA (825 ng) was hybridized to Agilent 4x44K whole humangenome microarrays (G4112F) and processed according tothe manufacturer's instructions. The array was scanned usingAgilent G2505B DNA microarray scanner. The image fileswere extracted using Agilent Feature Extraction software ver-sion 9.5.1 applying LOWESS background subtraction and dyenormalization.The data were analyzed using GeneSpring GX 10.0 (Agilent
Technologies) to identify genes that had statistically signifi-cantly changed expression between groups. Genes were con-sidered significantly differentially regulated with P value of<0.05 and fold change of ≥1.4. For hierarchical clustering,we used Pearson correlation for similarity measure and aver-age linkage clustering. A heat map was generated using Pear-son correlation clustering of a significant gene list afterone-way ANOVA of the raw data from all four groups.
Gene Set Enrichment AnalysisGene Set Enrichment Analysis (GSEA) was performed us-
ing the GeneSpring GX software and gene sets downloadedfrom Molecular Signatures Database (MSigDB; Broad Insti-tute, Massachusetts Institute of Technology); a custom listof 26 pluripotency genes was also created based on literaturereview. Gene sets were considered significant with Q value of<0.25, as recommended (19). Briefly, the primary result ofGSEA is the enrichment score (ES), which reflects the degreeto which a gene set is overrepresented at the top or bottomof a ranked list of genes. The normalized ES (NES) is the pri-mary statistic for examining gene set enrichment results. Bynormalizing the ES, GSEA accounts for differences in geneset size and in correlations between gene sets and the expres-sion data set.
Ingenuity Pathway AnalysisSignificant gene lists were generated from the GeneSpring
software and uploaded to Ingenuity Pathway Analysis (IPA)for analysis. IPA assigns biological functions to genes usingthe Ingenuity Pathways Knowledge Base (Ingenuity Systems,Inc.). This information is used to form networks to create an“interactome” of genes that are involved in specific biologicalprocesses.Functional analysis. The Functional Analysis identified
the biological functions and/or diseases that were most sig-nificant to the data set. Molecules from the data set that metthe P value cutoff of 0.05 and fold change cutoff of 1.4 werethen associated with biological functions and/or diseases in
Cancer Research

Irradiation of hESCs
Ingenuity's Knowledge Base. Right-tailed Fisher's exact testwas used to calculate a P value determining the probabilitythat each biological function and/or disease assigned to thatdata set is due to chance alone.Canonical pathway analysis. Canonical pathway analysis
identified the pathways from the Ingenuity Pathways Analy-sis library of canonical pathways that were most significantto the data set. The significance of the association between
www.aacrjournals.org
the data set and the canonical pathway was measured in twoways. (a) A ratio of the number of molecules from the dataset that map to the pathway divided by the total number ofmolecules that map to the canonical pathway is displayed.(b) Fisher's exact test was used to calculate a P value deter-mining the probability that the association between thegenes in the data set and the canonical pathway is explainedby chance alone.
Figure 1. A, schematic of experimental setup. B and C, immunostaining of pluripotency markers in control (B) and irradiated (C) hESCs shows maintenanceof marker expression 48 h after irradiation. D, flow cytometry of FITC Annexin V and PI double-stained hESCs 48 h after the indicated radiation dose(one representative experiment of three is shown).
Cancer Res; 70(13) July 1, 2010 OF3

Wilson et al.
OF4
Generation of stable reporter gene hESC linesEnhanced green fluorescent protein (eGFP) and firefly lu-
ciferase (Fluc) double-fusion reporter gene–positive hESCs(Fluc+/eGFP+hESC) have been previously described (20, 21).Briefly, SIN lentivirus was prepared by transient transfectionof 293T cells. H9 hESCs were stably transduced with LV-pUB-Fluc-eGFP at a multiplicity of infection of 10. The infectivitywas determined by eGFP expression as analyzed on a FACS-can. eGFP-positive cell populations were isolated by FACS-Vantage SE cell sorter (Becton Dickinson), followed byplating for long-term culture.
Subcutaneous transplantation of hESCsAnimal protocols were approved by the Stanford Universi-
ty Animal Care and Use Committee guidelines. All proce-dures were performed on 8- to 10-week-old female severecombined immunodeficient (SCID) Beige mice (Charles RiverLaboratories). Following induction with inhaled isoflurane(2–3%), anesthesia was then maintained with 1% to 2.5% iso-
Cancer Res; 70(13) July 1, 2010
flurane. Two hundred thousand Fluc+/eGFP+hESCs were sus-pended in a 50 μL volume of a 1:1 mixture of growthfactor–reduced Matrigel and DMEM and then irradiated atthe appropriate dosage (0.4, 2, or 4 Gy). Irradiated cellsuspensions were each injected s.c. into the dorsum of eightSCID mice; injections were performed within 2 hours ofirradiation.
Bioluminescence imaging of transplanted cell survivalBioluminescence imaging (BLI) was performed using the
Xenogen IVIS 200 system (Caliper Life Sciences). After i.p. in-jection of the reporter probe D-luciferin (375 mg luciferin/kgbody weight), animals were imaged for 1 to 10 minutes. Thesame mice were imaged for 6 weeks. Regions of interest weredrawn over the signals using the Igor image analysis software(Wavemetrics). BLI signal was standardized for acquisitiontime and quantified in units of maximum photons per sec-ond per square centimeter per steridian (photons/s/cm2/sr), as described (22).
Figure 2. Bioluminescence reporter gene imaging of irradiated hESCs in living animals. A, double-fusion reporter gene construct carrying Fluc and eGFP.B, diagram of the subcutaneous injection sites for each radiation group plus control, as well as representative bioluminescent images for three micethrough day 42. C, plot of longitudinal bioluminescent signal intensities for each group. *, P < 0.05, 0 and 0.4 Gy versus 2 and 4 Gy. Points, mean; bars, SE.D, H&E staining of teratoma section from representative 4 Gy group showing three embryonic germ layers.
Cancer Research
Irradiation of hESCs
Postmortem immunohistochemical stainingAnimals were sacrificed according to protocols approved
by the Stanford Animal Research Committee after the dura-tion of the study. Teratomas were explanted and processedfor H&E staining. Slides were interpreted by an expert pa-thologist (A.J.C.).
Statistical analysisNonmicroarray data are presented as mean ± SD. Data
were compared using standard or repeated-measures ANO-VA, where appropriate. Differences were considered signifi-cant for P < 0.05.
Results
Figure 1A gives a schematic of our experimental design.We first confirmed that low-dose irradiation (<1 Gy) ofhESCs was capable of upregulating known stress-responsivegenes: GADD45, which mediates activation of the p38/c-JunNH2-terminal kinase pathway via MTK1/MEKK4 kinase, andCXCL10, a chemokine for receptor CXCR3 that is involved inrecruitment of inflammatory cells (Supplementary Fig. S1).At a higher dose of 4 Gy, we observed massive cell death thatwas concurrent with the development of holes and patchyregions in hESC colonies at 48 hours (Fig. 1B and C); holeformation has also been reported in colonies 6 hours after5 Gy irradiation (14). However, the surviving hESCs contin-ued to express common pluripotency markers, such asTRA-1-81, SSEA4, and TRA-1-60, and embryonic transcriptionfactors, such as Oct4, Sox2, and Nanog, which are key regu-lators of pluripotency and self-renewal (Fig. 1C).We were curious about the relative extent of apoptosis and
cell death after irradiation at the different dosages, and so wedouble stained hESCs with Annexin V (early apoptosis) and
www.aacrjournals.org
PI (cell death) 48 hours after irradiation and analyzed thecells with flow cytometry (Fig. 1D). Clearly, there is a trendtoward increasing apoptosis and cell death at the higher ra-diation doses (2 and 4 Gy) compared with low dose (0.4 Gy)and control. The majority (>70%) of hESCs are dead after4 Gy irradiation, although an apoptotic minority (<30%)seems to survive at 48 hours. This latter population likely re-presents the surviving cells that continue to express pluripo-tency markers, as seen in Fig. 1C. However, the definitive testof pluripotency of human cells is the ability to form a tera-toma, which we performed next.To confirm that surviving hESCs are pluripotent, we in-
jected irradiated cells into immunocompromised mice andmonitored for teratoma formation. We tracked their growthkinetics in vivo by using hESCs that constitutively express aFluc-eGFP double-fusion reporter gene (Fig. 2A), enablinglongitudinal monitoring of cellular photon emission, and byextension their proliferation, as described (20, 21). After irra-diation and injection of Fluc+/eGFP+hESCs, we found that pho-ton emission from the 2 and 4 Gy groups reached a statisticalminimum at 7 days that was less than photon emission fromcells exposed to 0 and 0.4 Gy, suggesting massive cell death(P < 0.05; n = 8 per group; see Fig. 2B and C). Based on thephoton intensities, we estimated that 38 ± 30%, 63 ± 20%,80 ± 9%, and 83 ± 7% (mean ± SE) of the 0 Gy–, 0.4 Gy–, 2Gy–, and 4 Gy–irradiated cells, respectively, had died at day 7.We expected that the 2 and 4 Gy groups would continue to
die, but surprisingly, all four groups emitted similar levels ofphotons by day 21, indicating that surviving hESCs had re-covered from high-dose irradiation. We confirmed this resultby studying long-term in vitro cultures of irradiated hESCsand found that cell proliferation was inhibited in the firstweek after high-dose irradiation, but thereafter, all groups ex-hibited similar growth kinetics (Supplementary Fig. S2). Note
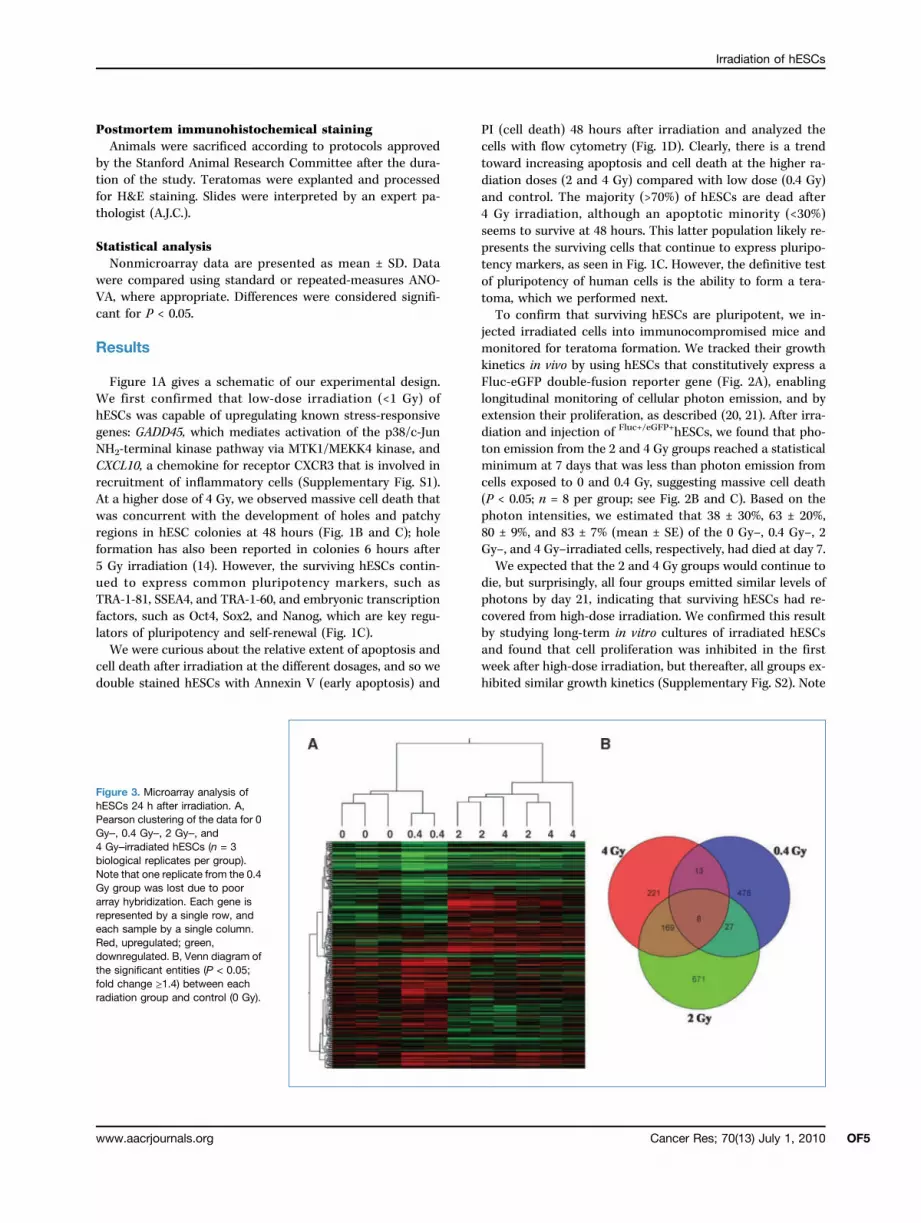
Figure 3. Microarray analysis ofhESCs 24 h after irradiation. A,Pearson clustering of the data for 0Gy–, 0.4 Gy–, 2 Gy–, and4 Gy–irradiated hESCs (n = 3biological replicates per group).Note that one replicate from the 0.4Gy group was lost due to poorarray hybridization. Each gene isrepresented by a single row, andeach sample by a single column.Red, upregulated; green,downregulated. B, Venn diagram ofthe significant entities (P < 0.05;fold change ≥1.4) between eachradiation group and control (0 Gy).
Cancer Res; 70(13) July 1, 2010 OF5

Wilson et al.
OF6
that after the postirradiation “recovery period,” we did notobserve any compensatory increase in cell proliferation inthe high-dose groups. Finally, of the eight mice used in thisstudy, five developed teratomas in the 4 Gy group by thesixth week (see Fig. 2D for representative H&E images andSupplementary Fig. S3 for a representative gross image offour teratomas from a single mouse). The three mice thatfailed to form teratomas in the 4 Gy group likely experiencedsignificant apoptosis and cell death and not loss of pluripo-tency. To confirm this, we performed a careful microarraystudy of the core set of pluripotency genes to determinewhether there are any detectable changes in pluripotencyprograms, however subtle, in response to ionizing radiation.For the transcriptomic analysis of irradiated hESCs, RNA
was isolated from cells 24 hours after irradiation and thenlabeled and hybridized to microarrays (raw data files havebeen uploaded to Gene Expression Omnibus under accessionnumber GSE20951). When analyzing microarray data, it is of-ten informative to start from a system-wide rather than indi-vidual-gene view of the resulting data, especially when theoverall gene fold changes are no more than 7-fold (Supple-mentary Table S1). An overview of the gene profiles can beseen in the heat map of Fig. 3A. Most apparent is the coclus-tering of the control and low-dose samples (0 and 0.4 Gy),which were distinct from the coclustering of the high-dosesamples (2 and 4 Gy). This pattern is also evident in Table 1,in which global Pearson correlation shows 95% correlationbetween the 2 and 4 Gy groups but only 86% correlationwith the low-dose 0.4 Gy group. A Venn diagram of theentities that are significantly different (P < 0.05; foldchange ≥ 1.4) between radiation dosage and control furtherillustrates this pattern (Fig. 3B; note that microarrays oftencontain multiple probes, or “entities,” for a given gene).Again, we observe the same grouping as in the Pearsoncorrelation: the 2 Gy– and 4 Gy–irradiated samples exhibita higher degree of overlap between themselves than theydo with the 0.4 Gy group.We next used IPA (Ingenuity Systems, http://www.ingenuity.
com) and GSEA (19) to further analyze the microarray data.Selected canonical pathways and functions that are disruptedafter 4 Gy irradiation (versus control) are summarized inTable 2; full data sets can be found in Supplementary TablesS2 and S3. After 4 Gy irradiation, canonical pathways, such asVDR/retinoid X response (RXR) activation, p53 signaling, andaryl hydrocarbon signaling, and functions, such as cancer, celldeath, cell cycle, growth and proliferation, and embryonic de-
Cancer Res; 70(13) July 1, 2010
velopment, are significantly affected in hESCs. Specifically,several tumor protein p53–associated genes such as TP53INP1(up 2.6-fold) and target genes such as CDKN1A (up 2-fold) andMDM2 (up 1.7-fold; ref. 23), as well as several tumor necrosisfactor receptor superfamily members, were disregulated afterirradiation. A small group of genes associated with develop-ment also exhibited differential expression, including HES1(down 1.8-fold; ref. 24), RUNX1 (up 1.5 fold), and PBX1 (down1.8-fold); note thatmany of these genes are also associatedwithcancer (Table 2). Supporting the observation that genes relatedto cancer are disregulated with radiation, GSEA, a methodfor analyzing a priori gene sets within microarray data,revealed upregulation of gene sets that have also been reportedin cells after treatment with chemotherapeutic drugs (Table 3;refs. 25–27).We have also analyzed the progression of gene and path-
way changes that occur in hESCs at each increasing radiationdose: between 0 and 0.4 Gy (Supplementary Tables S4–S6),0.4 and 2 Gy (Supplementary Tables S7–S9), and 2 and 4Gy (Supplementary Tables S10–S12). Similar to 4 Gy radia-tion, 0.4 Gy irradiation affects cellular functions such as celldeath, cancer, and signaling pathways such as p53, althoughnot important p53 downstream target genes such asCDKN1A and MDM2. Because CDKN1A is an important neg-ative regulator of cell cycling (28), the lack of upregulationof CDKN1A by 0.4 Gy irradiation could partly explain whywe did not observe a similar reduction in cell proliferationas in the 2 and 4 Gy groups. Relative to 0.4 Gy irradiation,2 Gy irradiation affects canonical transforming growth fac-tor-β and Wnt/β-catenin signaling, including the genesTgfbr2 (up 1.4-fold), Wnt1 (up 1.4-fold), Wnt10A (up 2.1-fold),and Wnt9a (up 1.8-fold); notably, Wnt proteins play impor-tant and diverse roles in embryonic stem cells (29). Irradi-ation (2 Gy) also induces CDKN1A upregulation by 2.3-foldbut not MDM2. Interestingly, many genes involved in func-tions such as cellular compromise, amino acid metabolism,molecular transport, and cell morphology, in addition tocancer and cell death, were significantly disrupted by2 Gy of radiation, including a number of solute carrier fam-ily proteins such as Slc6a13 (up 2-fold) and Slc25a13 (down2.2-fold). Clearly, there is an overall significant increase incellular dysfunction after 2 Gy irradiation, which explainsthe results from our in vivo and in vitro studies (Figs. 1and 2). Finally, in the 2 Gy versus 4 Gy group, the overallgene changes were not large, but a small group of genesrelated to organ and tissue development did have alteredexpression, such as Tnfsf11 (up 1.6-fold), Otx1 (down 1.6-fold), B4galt1 (down 1.4-fold), and Mef2c (up 1.9-fold). Pre-sumably, there are subtle development and differentiationprocesses that are activated with 4 Gy irradiation but arenot robust enough to cause loss of pluripotency, asevidenced by successful formation of teratomas from4 Gy-irradiated hESCs. Furthermore, the gene expressionprofiles of irradiated cells showed no increased correlationwith previously reported data for differentiated hESCs andprimary cell types (20, 30, 31), giving additional evidencethat differentiation is not significantly increased with ion-izing radiation (Supplementary Fig. S4).
Table 1. Global Pearson correlation of the mi-croarray data from irradiated hESCs
Gy
0 0.4 2 40
1 0.4 0.91 1 2 0.87 0.86 1 4 0.89 0.86 0.95 1Cancer Research

Irradiation of hESCs
With these observations, we were particularly interested tounderstand whether core pluripotency genes were also af-fected by radiation. Importantly, well-known embryonic tran-scription factors such as Oct4 (Pou5f1), Sox2, and Nanog,which are expressed exclusively or predominantly in hESCsand are critical for maintaining pluripotency and self-renewal,were not present in any of our significant gene lists across allradiation dosages. Moving beyond individual genes, we creat-ed a gene set of 26 known factors that are well known to bespecific to hESCs (bottom row of Table 3). GSEA revealed nosignificant upregulation or downregulation of this set in themicroarray data for the highest radiation dose, 4 Gy (P = 0.4;
www.aacrjournals.org
Q = 0.5; NES = −1.0). This finding agrees with a previous reportthat showed normalization of Oct4 and Nanog after only24 hours in 2 Gy–irradiated hESCs (14). We therefore con-clude that pluripotency gene programs are not significantlyaffected by high-dose radiation, and this accounts for the ob-servation that surviving hESCs are still capable of forming allthree embryonic germ layers.
Discussion
In pregnant mothers undergoing diagnostic or therapeuticprocedures involving ionizing radiation, or who may be
Table 2. Selected genes and biological processes affected by 4 Gy irradiation of hESCs
Canonical pathways
GenesVDR/RXR activation
IGFBP6, CDKN1A, CSNK2A1, HES1, RXRB, PRKCB p53 signaling TP53INP1, CDKN1A, TNFRSF10B, C12ORF5, MDM2, HIPK2 Aryl hydrocarbon receptor signaling TFF1, NQO1, CDKN1A, MDM2, DHFR, RXRB, AHR Platelet-derived growth factor signaling CSNK2A1, INPP5D, PRKCB Notch signaling DTX1, HES1 Cell cycle: G2-M DNA damage: check-point regulationCDKN1A, MDM2
Molecular mechanisms of cancer
HHAT, RALA, CDKN1A, MDM2, HIPK2, GLI1, FAS, PRKCBFunctions
GenesCancer
DPYD, PLK3, PBX1, EIF4A2, ATP4A, HES1, DDB2. LATS2, TTC22, FHL2, TFF1,MPHOSPH8, NEK6, CSNK2A1, TNFRSF10C, CBFB, TUBA1C, TTC5, HIPK2, CDKN1C,AHR, MYO6, IGFBP6, IFIT3, TUBB3, TP53INPL, NKTR, PPP1R1B, TNFRSF10B, LlCAM,UBE2S, RASD1, CD40, MIB1, LRP8, RUNX1, DHFR, HELB, EB13, RPL13A, KIAA1370,POLE3, FBXL7, PIAS1, GDF15, FAS, STK17A, GBP2, RXRB, VASP, CALR, ATP1B1,CKM, NQO1, TOB1, MDM2, SLC7A8, FDXR, INPP5D, DNM1, TRPM6, HP, GMPS,CDKN1A, MEF2C, GLI1, COBLL1, POLH, PRKCBCell death
QKI, PLK3, PBX1, DDB2, HES1, TREM2, LATS2, FHL2, BLOC1S2, TFF1, MTF1, NEK6,CSNK2A1, CBFB, TTC5, TNFRSF10C, CDKN1C, HIPK2, AHR, MYO6, SHISA5, IGFBP6,TUBB3, TP53INP1, SGCG, PPP1R1B, TNFRSF10B, L1CAM, UBE2S, LAX1, RASD1,NCF1, MNT, CD40, MIB1, RUNX1, GDF15, PIAS1, FAS, STK17A, RPS3, SOX5, CALR,THG1L, NQO1, CDC42EP3, MDM2, INPP5D, FDXR, DNM1, CDKN1A, MEF2C, GLI1,PRKCB, POLHCell morphology
RALA, PLK3, GDF15, ATP4A, HES1, LATS2, FAS, ANK1, MACF1, CSNK2A1, CBFB,CDKN1C, RXRB, VASP, VASP, AHR, PRICLE2, TNFRSF10B, MDM2, LICAM, DNM1,PIP5K1A, CD40, MIB1, CDKN1A, FOXJ1, B3GAT1, LRP8, NDST2, GLI1Cellular assembly and organization
RALA, TUBGCP3, NQO1, TNFRSF10B, PLEC1, MDM2, FAS, ANK1, DNM1, TUBGCP5,RAB11FIP4, AMPH, CDKN1A, TTL, NDST2, CDKN1C, VASP, AHREmbryonic development
TP53INP1, PIAS1, TNFRSF10B, PBX1, LICAM, HES1, FAS, DNM1, MTF1, RUNX1,HIPK2, GLI1, VASP, AHRCellular growth and proliferation
GDF15, PIAS1, PBX1, HES1, FAS, FHL2, HS6ST2, CBFB, CDKN1C, HIPK2, VASP, AHR,CALR, IGFBP6, TUBB3, TP53INP1, TNFRSF10B, TOB1, PTP4A1, MDM2, INPP5D,TRPM6, NCF1, MNT, CD40, CDKN1A, UHMK1, DHFR, RUNX1, GLI1, PRKCBCell cycle
CALR, TP53INP1, GDF15, PIAS1, MDM2, HES1, LATS2, INPP5D, FAS, DNM1, SESN1,MNT, CD40, CDKN1A, NEK6, CSNK2A1, UHMK1, RUNX1, CDKN1C, HIPK2, AHRCellular development
QK1, PBX1, HES1, TREM2, FAS, ANK1, FOXN4, FHL2, DTX1, TFF1, CBFB, CDKN1C,RXRB, AHR, VASP, SOX5 (includes EG:6660), MYO6, IGFBP6, CALR, CTSK, TUBB3,PPP1R1B, DNER, TNFRSF10B, TOB1, L1CAM, MDM2, INPP5D, NCF1, MNT, CD40,MIB1, CDKN1A, MEF2C, RUNX1, GLI1, PRKCBCancer Res; 70(13) July 1, 2010 OF7

Table 3. GSEA of 4 Gy–irradiated versus 0 Gy–irradiated hESCs
Gene set name P Q ES NES Description Genes in set
METHOTREXATE_PROBCELL_UP <0.0001 0.1447 0.6866 1.8669 Upregulated in pro-Bcells (FL5.12) followingtreatment with metho-trexate (Brachat et al.)
ABI1, ABLIM1, CARHSP1,CASP4 , CDKN1A , E I 2 4 ,H2AFJ, LPIN1, LRRC2, MA-LAT1, MDM2, PVRI4, SLC7A14,TOB1, TP53INP1, TRAFD1,TXNIP, UCHL5
BLEO_HUMAN_LYMPH_HIGH_4HRS_UP <0.0001 0.1929 0.7732 1.8725 Upregulated at 4 h follow-ing treatment of humanlymphocytes (TK6) witha high dose of bleomycin(Islaih et al.)
BBC3, BTG1, BTG2, CDKN1A,CLK1, DDB2, DDIT4, DUSP14,ENC1, FAS, FDXR, GADD45A,GDF 1 5 , HNRPA1 , I E R 3 ,PLXNB2, PMAIP1, PPM1D,TNFRSF10B, TNFSF9, XPC
CAMPTOTHECIN_PROBCELL_UP <0.0001 0.2290 0.6617 1.8162 Upregulated in pro-Bcells (FL5.12) followingtreatment with camp-tothecin (Brachat et al.)
AB I 1 , C 1 2ORF 2 2 , C 1R ,CARHSP1, CBS, CDKN1A,EI24, GRB10, H2AFJ, LPIN1,LRRC2, NUDCD2, PMM1,PVRL4, SERTAD1, SLC7A14,YTOB1, TP53INP1, TPP1,TRAFD1, TXNIP, UCHL5, ULK1
OXSTRESS_BREASTCA_UP <0.0001 0.2314 0.7095 1.8060 Upregulated by H2O2,menadione, and t-butylhydroperoxide in breastcancer cells (Chuang et al.)
C1ORF107, CTAGE5, CYP1B1,DDTT3, DKK1, EDN1, EGR1,EPS8L2, FAS, FDXR, GDF15,H19, HMOX1, HSPA1B, JUN,L IF , LRP6 , MT1H, MT1X,NQO1, PLA2R1, PPP2CB,PSITPTE22, PSMD12, PSMD3,RCBTB1, RPL38, SLC35B3,ZBTB4
Human Embryonic Cell Markers 0.4000 0.5000 −0.3177 −1.0009 Common hESC genes(based on l i t e ra tu research)
POU5F1, NANOG, KLF2, KLF5,SOX1, SOX2, SOX5, LIN28,DNMT3B , GDF3 , DPPA4 ,DPPA5 , ESRRG , SALL4 ,NR6A1, TDGF1, TBX3, FOXD3,FGF4, ZFP42, LEFTY1, LEFTY2,ERAS, PODXL, TERT, UTF1
NOTE: GSEA of the same microarray data.
Wilso
net
al.
Cancer
Res;
70(13)July
1,2010
CancerRese
arc
hOF8

Irradiation of hESCs
exposed to environmental radiation, there is a great po-tential for damage to the early embryo. Although the em-bryo is somewhat protected by the uterus, it is particularlysensitive to ionizing radiation, and the developmentalconsequences can be quite serious (2). Data about the po-tential biological effects on the embryo after in utero ir-radiation are based on the results of animal studies (32–35)and a limited number of human exposures such as the1945 atomic bomb survivors from Hiroshima and Naga-saki. Based on these collective data, it has been well es-tablished that the dose of ionizing radiation and thedevelopmental stage of the embryo are the determiningfactors for reproductive toxicity in embryonic develop-ment (1). Above poorly defined threshold doses, the majoreffects of ionizing radiation are lethality during the preim-plantation-preorganogenetic period and malformationsand growth retardation during organogenesis (1, 3). Othersequelae later in life may include severe mental retarda-tion, a reduced intelligence quotient, and childhood can-cer (1, 3, 36). Unfortunately, the absolute incidence andradiation dose at which these changes occur, as well asthe mechanisms of damage, remain unclear.Because the in utero embryonic response to ionizing radi-
ation is not well understood due to the obvious ethical con-cerns of exposing pregnant mothers to radiation, hESCspresent a novel in vitro platform for studying the human em-bryonic response to irradiation. These cells are derived fromthe inner cell mass of the blastocyst during embryonic devel-opment and are therefore closely related to the early-stagehuman embryo. Admittedly, hESCs are still different fromthe early embryo in that they lack the complex and dynamicsignaling environment of the uterus and are instead main-tained long term in relatively simple in vitro cultures. How-ever, the advantage is that we can begin to tease out theembryonic response to irradiation in a human rather thanmurine system. Furthermore, because radiotracers and PETreporter genes that monitor cellular transplantation foremerging regenerative and antioncogenic therapies are beingincreasingly used in laboratory research (8), and one day mayeven achieve routine clinical application (7), it will be impor-tant to determine whether such radioactive probes can di-rectly affect the viability and function of the transplantedcells. For these reasons, we decided that a broad survey ofthe functional and global molecular response of hESCs to ir-radiation, and in particular the effect of radiation on pluripo-tency, was a critical area of investigation.Not surprisingly, our results show that high doses of radi-
ation cause massive cell death, with a trend toward increas-
www.aacrjournals.org
ing apoptosis and death at the higher radiation doses (2 and4 Gy) compared with low dose (0.4 Gy) and control. Interest-ingly, an apoptotic minority does seem to survive at 48 hours.Using a BLI technique, we confirmed that the higher doses ofradiation cause hESCs to initially die after transplantation,but the surviving cells recover by 2 weeks to levels similarto control. Regardless of the radiation dose used in our study,all groups of irradiated hESCs were able to form teratomas,the definitive test of pluripotency. Our genome-wide analysisof gene expression revealed genes and pathways at each ra-diation dose that are involved in cell death, p53 signaling, cellcycling, cancer, embryonic and organ development, andothers. Importantly, GSEA showed that the expression of acomprehensive set of core embryonic transcription factorsis not significantly altered by radiation at any dose and helpsexplain how irradiated hESCs are still able to form teratomas.In summary, this is the first study of hESC genome-wide
transcriptional changes induced by ionizing radiation andis a preliminary step toward a better understanding of notonly hESC molecular changes but also the in utero embryonicgene expression response. We have shown that, similar to so-matic cells, irradiated hESCs suffer significant death and ap-optosis after irradiation. Although some gene programsinvolved in developmental pathways are altered with high-dose radiation, the expression of pluripotency genes is unaf-fected, and these cells can still form teratomas. Studies suchas this may help define the limits for radiation exposure forpregnant women and also radiotracer reporter probes fortracking cellular regenerative therapies.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Pauline Chu and Andrew J. Connolly for assistance with tissueprocessing and histologic analysis.
Grant Support
Stanford Bio-X Fellowship (K.D. Wilson), R21 HL091453 ( J.C. Wu), and R33HL089027 ( J.C. Wu).
The costs of publication of this article were defrayed in part by the paymentof page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
Received 11/24/2009; revised 03/25/2010; accepted 04/13/2010; publishedOnlineFirst 06/08/2010.
References
1. Brent RL. Utilization of developmental basic science principlesin the evaluation of reproductive risks from pre- and postcon-ception environmental radiation exposures. Teratology 1999;59:182–204.
2. McCollough CH, Schueler BA, Atwell TD, et al. Radiation exposureand pregnancy: when should we be concerned? Radiographics2007;27:909–17.
3. De Santis M, Cesari E, Nobili E, Straface G, Cavaliere AF, Caruso A.
Radiation effects on development. Birth Defects Res C EmbryoToday 2007;81:177–82.
4. Einstein AJ, Moser KW, Thompson RC, Cerqueira MD, Hanzlova MJ.Radiation dose to patients from cardiac diagnostic imaging. Circula-tion 2007;11:1290–305.
5. Fazel R, Krumholz HM, Wang Y, et al. Exposure to low-dose ionizingradiation from medical imaging procedures. N Engl J Med 2009;361:849–57.
Cancer Res; 70(13) July 1, 2010 OF9

Wilson et al.
OF10
6. Smith-Bindman R, Lipson J, Marcus R, et al. Radiation dose associ-ated with common computed tomography examinations and the as-sociated lifetime attributable risk of cancer. Arch Intern Med 2009;169:2078–86.
7. Yaghoubi S, Jensen M, Satyamurthy N, et al. Noninvasive detectionof therapeutic cytolytic T cells with 18F-FHBG PET in a patient withglioma. Nat Clin Pract Oncol 2009;6:53–8.
8. Narsinh K, Cao F, Wu J. Molecular imaging of human embryonicstem cells. Methods Mol Biol 2009;515:13–32.
9. Hogan D, Kellison T. Nuclear terrorism. Am J Med Sci 2002;323:341–9.
10. Aladjem MI, Spike BT, Rodewald LW, et al. ES cells do not activatep53-dependent stress responses and undergo p53-independent ap-optosis in response to DNA damage. Curr Biol 1998;8:145–55.
11. Hong Y, Stambrook P. Restoration of an absent G1 arrest and pro-tection from apoptosis in embryonic stem cells after ionizing radia-tion. Proc Natl Acad Sci U S A 2004;101:14443–8.
12. Solozobova V, Rolletschek A, Blattner C. Nuclear accumulation andactivation of p53 in embryonic stem cells after DNA damage. BMCCell Biol 2009;10:46.
13. Filion T, Qiao M, Ghule P, et al. Survival responses of human embry-onic stem cells to DNA damage. J Cell Physiol 2009;220:586–92.
14. Momcilovic O, Choi S, Varum S, Bakkenist C, Schatten G, Navara C.Ionizing radiation induces ataxia telangiectasia mutated-dependentcheckpoint signaling and G(2) but not G(1) cell cycle arrest in plurip-otent human embryonic stem cells. Stem Cells 2009;27:1822–35.
15. Qin H, Yu T, Qing T, et al. Regulation of apoptosis and differentiationby p53 in human embryonic stem cells. J Biol Chem 2007;282:5842–52.
16. Becker K, Stein J, Lian J, van Wijnen A, Stein G. Establishment ofhistone gene regulation and cell cycle checkpoint control in humanembryonic stem cells. J Cell Physiol 2007;210:517–26.
17. Sokolov M, Panyutin I, Onyshchenko M, Panyutin I, Neumann R. Ex-pression of pluripotency-associated genes in the surviving fraction ofcultured human embryonic stem cells is not significantly affected byionizing radiation. Gene 2010;455:8–15.
18. Ludwig TE, Levenstein ME, Jones JM, et al. Derivation of human em-bryonic stem cells in defined conditions. Nat Biotechnol 2006;24:185–7.
19. Subramanian A, Tamayo P, Mootha V, et al. Gene set enrichmentanalysis: a knowledge-based approach for interpreting genome-wideexpression profiles. Proc Natl Acad Sci U S A 2005;102:15545–50.
20. Cao F, Wagner RA, Wilson KD, et al. Transcriptional and functionalprofiling of human embryonic stem cell-derived cardiomyocytes.PloS One 2008;3:e3474.
Cancer Res; 70(13) July 1, 2010
21. Wilson KD, Huang M, Wu JC. Bioluminescence reporter gene imag-ing of human embryonic stem cell survival, proliferation, and fate.Methods Mol Biol 2009;574:87–103.
22. Li Z, Wu JC, Sheikh AY, et al. Differentiation, survival, and function ofembryonic stem cell derived endothelial cells for ischemic heart dis-ease. Circulation 2007;116:I46–54.
23. Shu K-X, Li B, Wu L-X. The p53 network: p53 and its downstreamgenes. Colloids Surf B Biointerfaces 2007;55:10–8.
24. Kobayashi T, Mizuno H, Imayoshi I, Furusawa C, Shirahige K,Kageyama R. The cyclic gene Hes1 contributes to diverse differen-tiation responses of embryonic stem cells. Genes Dev 2009;23:1870–5.
25. Brachat A, Pierrat B, Xynos A, et al. A microarray-based, integratedapproach to identify novel regulators of cancer drug response andapoptosis. Oncogene 2002;21:8361–71.
26. Chuang Y-YE, Chen Y, Chandramouli GVR, et al. Gene expressionafter treatment with hydrogen peroxide, menadione, or t-butyl hydro-peroxide in breast cancer cells. Cancer Res 2002;62:6246–54.
27. Islaih M, Li B, Kadura I, et al. Comparison of gene expressionchanges induced in mouse and human cells treated with direct-acting mutagens. Environ Mol Mutagen 2004;44:401–19.
28. Besson A, Dowdy S, Roberts J. CDK inhibitors: cell cycle regulatorsand beyond. Dev Cell 2008;14:159–69.
29. Nusse R, Fuerer C, Ching W, et al. Wnt signaling and stem cell con-trol. Cold Spring Harb Symp Quant Biol 2008;73:59–66.
30. Li Z, Wilson KD, Smith B, et al. Functional and transcriptional char-acterization of human embryonic stem cell-derived endothelial cellsfor treatment of myocardial infarction. PloS One 2009;4:e8443.
31. Jia F, Wilson KD, Sun N, et al. A nonviral minicircle vector for derivinghuman iPS cells. Nat Methods 2010;7:197–9.
32. Russell WL. Mutation frequencies in female mice and the estimationof genetic hazards of radiation in women. Proc Natl Acad Sci U S A1977;74:3523–7.
33. Jacquet P. Sensitivity of germ cells and embryos to ionizing radia-tion. J Biol Regul Homeost Agents 2004;18:106–14.
34. Brent RL, Bolden BT. The indirect effect of irradiation on embryonicdevelopment. 3. The contribution of ovarian irradiation, uterine irra-diation, oviduct irradiation, and zygote irradiation to fetal mortalityand growth retardation in the rat. Radiat Res 1967;30:759–73.
35. Jacquet P, de Saint-Georges L, Vankerkom J, Baugnet-Mahieu L.Embryonic death, dwarfism and fetal malformations after irradiationof embryos at the zygote stage: studies on two mouse strains. MutatRes 1995;332:73–87.
36. Wakeford R, Little MP. Risk coefficients for childhood cancer afterintrauterine irradiation: a review. Int J Radiat Biol 2003;79:293–309.
Cancer Research





























