Embed Size (px)
Citation preview

Brain Research, 531 (1990) 117-126 117 Elsevier
BRES 15961
Dye-induced 'photo-degeneration' and 'photo-permeabilization' of mammalian neurons in vivo
S. Picaud 1'2, L. Peichl 1 a n d N. F r a n c e s c h i n i 2
IMax-Planck lnstitut far Hirnforschung, Frankfurt (ER.G.) and 2C.N.R.S., Laboratoire de Neurobiologie, E3, Marseille (France)
(Accepted 24 April 1990)
Key words: Dye; Photosensitization; Neuronal degeneration; Cell permeabilization; Lesion; Vertebrate retina; Rat
Dyes are known to induce neuronal 'photo-degeneration' and 'photo-permeabilization' in fly photoreceptor cells in vivo. In the present study, we attempted to generalize this photodynamic damage to vertebrate neurons, using the rat retina, a brain part which is optically accessible in vivo. After intravitreal injection of the photosensitizing dye Rose Bengal (RB), irradiation of the retina of a living rat with a T-shaped microbeam was found to induce striking 'optograms' which could be observed on the excised retina. The T-shaped pattern which was to be seen in the translucent retina under transmitted light was attributed to neuronal degeneration of the neurons irradiated in the presence of RB, as attested by classical degenerative features such as a cytoplasmic darkening or a drastic swelling. The T-shaped pattern could also be observed on adding the dye Lucifer yellow to the extracellular space of the retina either in vitro or in vivo, showing that the cells irradiated in the presence of RB became permeable. These structural reactions were observed in the cells in the inner nuclear layer (INL) and ganglion cell layer (GCL), in the processes in both plexiform layers, and in the ganglion cell axons crossing this area, whereas the photoreceptors in the outer retina appeared to be undamaged. From these reactions, due to photo-degeneration and photo-permeabilization, it was possible to identify the photodynamic damage to the nervous system histologically at the macroscopic, cellular and ultrastructural levels. In view of its accuracy and reproducibility, the photo-lesion technique holds great potential as a tool for investigating various nervous systems.
INTRODUCTION
In studies on living biological tissue, dyes are increas-
ingly used as markers to label cell populations, cell
organelles or even molecules. While some dyes are
themselves target specific, others can be coupled to vectors such as antibodies or toxins. Some dyes are
convenient indicators of pH, CaZ+-concentration or
membrane potential and can be used to obtain kinetic
measurements of these parameters, even at the single cell level 4. However , a major drawback encountered when
using dyes in living tissue is the cell photosensitization 7"22
that may lead to photodynamic damage and cell death.
Photooxidation, the underlying mechanism responsible
for the dye-induced photosensitization, is known to produce protein alteration and lipid peroxidation 7'22. In
cancer therapy, this photodynamic effect of dyes has been turned to an advantage to eradicate tumor cells 5.
In nervous tissue, photodynamic damage has mainly been attested using electrophysiological techniques 3,1°, 13.17. Irradiation in the presence of a photosensitizing dye has also been found to stimulate dye uptake into excitable cells in culture 6. Dyes have become so widely
used in neurobiology that there now exists a need to
determine the histological symptoms of photodynamic damage in nervous tissue.
The photodynamic effect of an intracellularly applied
dye has been reported to result in the immediate disintegration of the dye-filled neuron 13. Recently, the
structural modifications occurring in neurons due to the
photodynamic effect of an extracellularly applied dye were investigated in vivo on individual fly ph0toreceptor cells 18'19. Photodynamic action was found to trigger
neuronal degeneration and cell permeabilization, which were termed 'photo-degenerat ion '19 and 'photo-perme-
abilization 'is, respectively. The photodynamic damage
was subsequently assessed in fly photoreceptor cells,
either on the basis of classical neuronal degenerative
features, such as an increase in the electron density of the
cytoplasm, or on the basis of dye permeation into altered cells. Both the dark degeneration and dye permeation observed after this photolesion provided adequate cyto- plasmic labeling for anatomical investigations 18'19.
Since the photodynamic damage due to dye-induced photosensitization is known to affect components which are common to all cells, all neurons should theoretically
Abbreviations: RB, Rose Bengal; LY, Lucifer yellow; ONL, outer nuclear layer; OPL, outer plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer; GCL, ganglion cell layer Correspondence: S. Picaud, CNRS - LNB 3, 31 Chemin Joseph Aiguier, 13402 Marseille cedex 09, France.
0006-8993/90/$03.50 © 1990 Elsevier Science Publishers B.V. (Biomedical Division)

118
be subjec t to p h o t o - d e g e n e r a t i o n and pho to -pe rmeab i l i -
za t ion. T h e p resen t s tudy was an a t t em p t to set t le this
ques t i on as far as v e r t e b r a t e neura l t issue is conce rned .
T h e v e r t e b r a t e re t ina was used because it conta ins
we l l -de f ined n e u r o n a l popu la t ions and affords easy op-
tical access in v ivo th rough the pupi l of the eye. H e r e we
descr ibe the h is to logica l modi f ica t ions resul t ing f rom a
prec ise i r rad ia t ion of the rat re t ina in v ivo af ter infusion
of the dye R o s e Benga l ( R B ) into the v i t reous body. R B
was chosen in these e x p e r i m e n t s because it is he ld to be
the p r ime pho tosens i t i ze r 9 and was p rev ious ly obse rved
to t r igger p h o t o - d e g e n e r a t i o n and pho to -pe rmeab i l i z a -
t ion in fly p h o t o r e c e p t o r s at a physio logica l l ight intensi ty
level .
MATERIALS AND METHODS
In vivo procedure Albino rats of either sex were anesthetized by an intraperitoneal
injection of choral hydrate (40 rag/100 g body weight, dissolved in distilled water). A small scleral incision was made into one eyeball near the retinal periphery, and 5/~! of a Rose Bengal (RB) solution were injected into the vitreous body with a 10 ktl Hamilton syringe. The solution contained 0.1% RB (w/v) in an Ames and Nesbett medium 1 (Flow Laboratories, UK). The pupil was dilated with Neosynephrin POS 5%. The anesthetized animal was then mounted on a plate. To minimize eye movements, threads were pulled through the conjunctiva of the injected eye at four cardinal points and attached to the head.
The animal on its plate was placed on the stage of an epifluo- rescence microscope (ACM, Zeiss) and the injected eye was aligned with the optical axis of the microscope. In order to produce a well-defined image on the retina, the epifluorescence irradiation unit was modified as shown in Fig. 1. Light from a high-pressure mercury lamp (HBO 50 W) was focused onto an iris diaphragm (aperture stop D1) conjugated with the pupil of the eye. A photographically produced T-shaped diaphragm D2 (bar width: 0.1 ram; vertical bar length: 1.1 mm) was imaged onto the back focal plane of the objective (Nikon M Plan 40x/0.5 ELWD) and hence onto the retina. The image on the retina could be observed on-line by substituting a 'centering telescope' for one of the eyepieces. Using the green excitation unit, the eye of the animal was exposed for 30 min to green light (essentially the Hg green line, the wavelength of which lies close to the maximal absorbance of RB, ~'max : 548 rim; ref. 9). The illuminance of the T imaged onto the retina was 2000 lux, as measured with a calibrated silicon photo- diode (BPY 11; Siemens, F.R.G.) placed under the microscope objective. Note that this kind of 'irradiation' lies within the physiological intensity range.
Cell permeability assay The fluorescent dye Lucifer yellow (LY) was used to assess the
cell permeability of irradiated cells in an in vitro assay. Immediately after the green irradiation in the presence of RB, the animal was sacrificed by an overdose of chloral hydrate and the irradiated eye enucleated. The eyeball was cut open around the cornea and the anterior part including the lens and cornea was removed. The posterior eye cup, with the retina attached, was immersed for 3 min in a 0.05% Lucifer yellow solution in Ames and Nesbett medium osmotically balanced with 0.9 g/1 saccharose and quickly rinsed in LY-free medium prior to fixation.
Histological procedures The posterior eye cup, dissected as described above, was
immersion-fixed in 4% paraformaldehyde in phosphate buffer (0.1
M, pH 7.4) overnight at 4 °C. After fixation, the retina was dissected free and mounted vitreal side up in phosphate buffer. To assess the general effect of the treatment, the retinal whole mount was examined under normal transmitted light and with epifluores- cence illumination for RB (interference filter: 546 nm; dichroic mirror: 589 nm; barrier filter: >590 nm) or LY (broadband excitation: 400-440 nm; dichroic mirror: 460 nm; barrier filter: >470 nm) with a low-power objective (Zeiss, Plan 1 ×/0.04). Only retinae with a clear 'biological print' of the letter 'T' previously imaged onto the retina in vivo were further processed histologically. Pieces of tissue containing the 'T'-area were dehydrated and embedded in araldite. Semithin vertical sections (2/am) were cut in a plane perpendicular to one of the crossbars of the "T'. These sections were first examined for fluorescence under blue excitation and later counterstained with Toluidine blue (1%) for observation under transmitted white light.
For EM observation, the eye was enucleated and opened as described above, and immediately immersed in a fixative containing 2% paraformaldehyde and 2.5% glutaraldehyde in phosphate buffer (0.1 M PB, pH 7.4) overnight at 4 °C. Pieces of tissue containing the irradiated area were postfixed in 1% osmium tetroxide in 0.1 M PB (pH 7.4) and conventionally embedded in araldite. Ultrathin sections were counterstained with uranyl acetate and lead citrate.
RESULTS
Structural changes
W h e n , af ter in t rav i t rea l app l ica t ion of t he dye R o s e
Benga l ( R B ) , a T - s h a p e d l ight pa t t e rn was p r o j e c t e d
on to the rat re t ina in v ivo for 30 min (Fig. 1), a T - shaped
' o p t o g r a m ' was subsequen t ly o b s e r v e d u n d e r t r ansmi t t ed
l ight, seeming ly i m p r i n t e d in the t rans lucen t t issue o f the
excised re t ina (Figs. 2A , 3 A , C ) . This p h e n o m e n o n was
consis tent ly induced (n = 30) u n d e r g r een l ight , i .e. in a
L 1 L 2 L 3 , ~
Fig. 1. Diagram of the irradiation set-up used to project a T-shaped pattern in vivo onto the retina of an anesthetized rat (not to scale). HBO 50 W mercury lamp; L = lenses; L 6 = cornea + crystalline lens of the animal eye; M = dichromatic mirror; D 1 = aperture stop (iris diaphragm) imaged onto the pupil of the rat eye; D 2 = T-shaped diaphragm imaged onto the rat retina; Obs = observer.

119
spectral range corresponding to the absorption maximum of RB, and the rat retinae were usually fixed immediately after irradiation. The fact that the appearance of the retina outside the irradiated area was normal showed that RB by itself had no effect unless photo-excited. In control experiments with the same green light, omitting the application of RB (n = 7), no such effect was observed (Fig. 3E). These controls demonstrated that production of the pattern on the retina resulted from an interaction between the irradiating green light and the green absorbing dye RB that had diffused from the vitreous body into the retina.
Vertical, semithin sections of the retina across a bar of the T revealed the existence of striking differences between the irradiated retinal region and adjacent re- gions (Fig. 4). The inner nuclear layer (INL) and outer plexiform layer (OPL) were severely affected. When the section was counterstained with Toluidine blue, the INL somata showed an intense staining within the irradiated part, whereas the photoreceptor somata in the outer nuclear layer (ONL) showed normal staining (Fig. 4B). The cells in the ganglion cell layer (GCL) showed normal, pale staining at the light microscopic level, although they were altered at the ultrastructural level (see below). A sharp boundary separated the damaged region from the unaffected tissue, in which all the retinal layers showed normal staining. At the border of the irradiated area some cells in the GCL were also intensely stained. All the cells showing this increased stainability
were found to be sharply contrasted under phase-contrast microscopy before the section was stained with Toluidine blue. As shown at the ultrastructural level (see below), the increased stainability with Toluidine blue coincided with the occurrence of the dark state of degeneration 16.
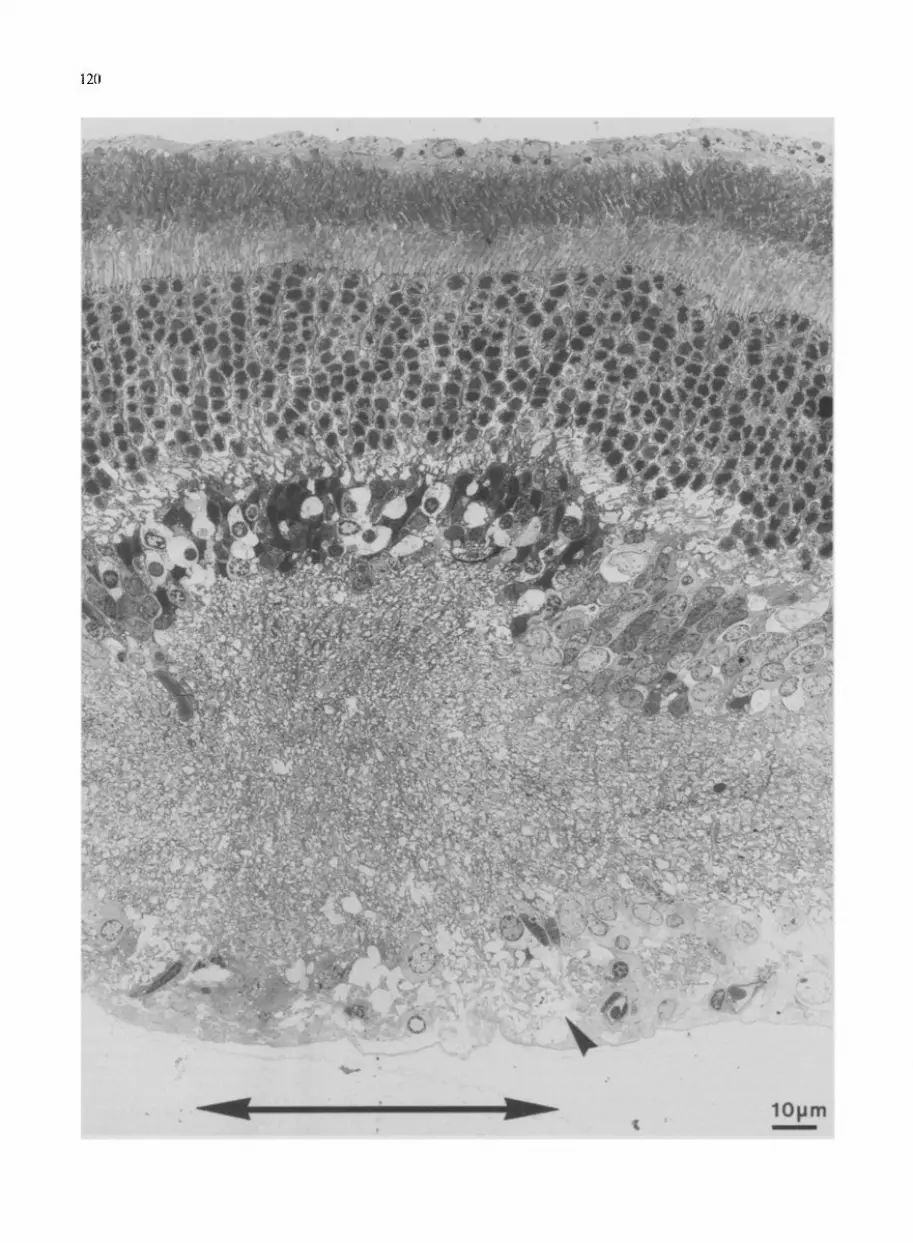
Fig. 5 shows the ultrastructural changes observed in the lesion area, which extends between the two arrows indicated beneath. With respect to the neighboring tissue, the most striking differences are the darkening of the INL and the fold in this layer associated with a widening of the IPL. The photoreceptors again appeared to be normal, whereas the cells in the INL and GCL were in a state of degeneration. The cells in the INL became either electron-dense or swollen with an empty cyto- plasm, features which are characteristic of dark and watery degeneration 16, respectively. The somata in the GCL showed evidence of degenerative changes, such as an increased vesieulation, and an increase in the electron density of their cytoplasm and chromatin. The ax0n bundles in the irradiated area were disintegrated, show- ing a few remaining axons scattered in the GCL (arrow- head). The processes in both plexiform layers (OPL and IPL) were either heavily swollen or electron dense. At the border of the lesion area, a number of somata and fibers also displayed altered structures, the occurrence of which is discussed below.
In short, application of RB to the vitreous body and subsequent irradiation of the retina within the absorption band of RB gave rise to degenerative features in the cells
Fig. 2. Coiour photomicrographs of a fiat-mounted whole retina showing 'optograms' resulting from photo-degeneration and photo- permeabilization. A special T-shaped pattern was projected with green light for 30 min after application of Rose Bengal (RB), a green absorbing dye, to the vitreous body. In this particular experiment, the blue absorbing, yellow-green fluorescing dye Lucifer yellow (LY) was added to the solution of RB at the same concentration (0.1% w/v). The T-shaped pattern imaged onto the retina in vivo gave rise to both a darkening of the retinal tissue under transmitted light (A) and a bright yellow-green fluorescent labeling under blue excitation (B). The latter stemmed from the intracellular staining of neurons, which became permeable to dyes within the irradiated area. This retina was fixed 1 h after the in vivo irradiation.
121
O N L
O P L I N L
I P L
G C L
O N L
O P L I N L
I P L
G e L
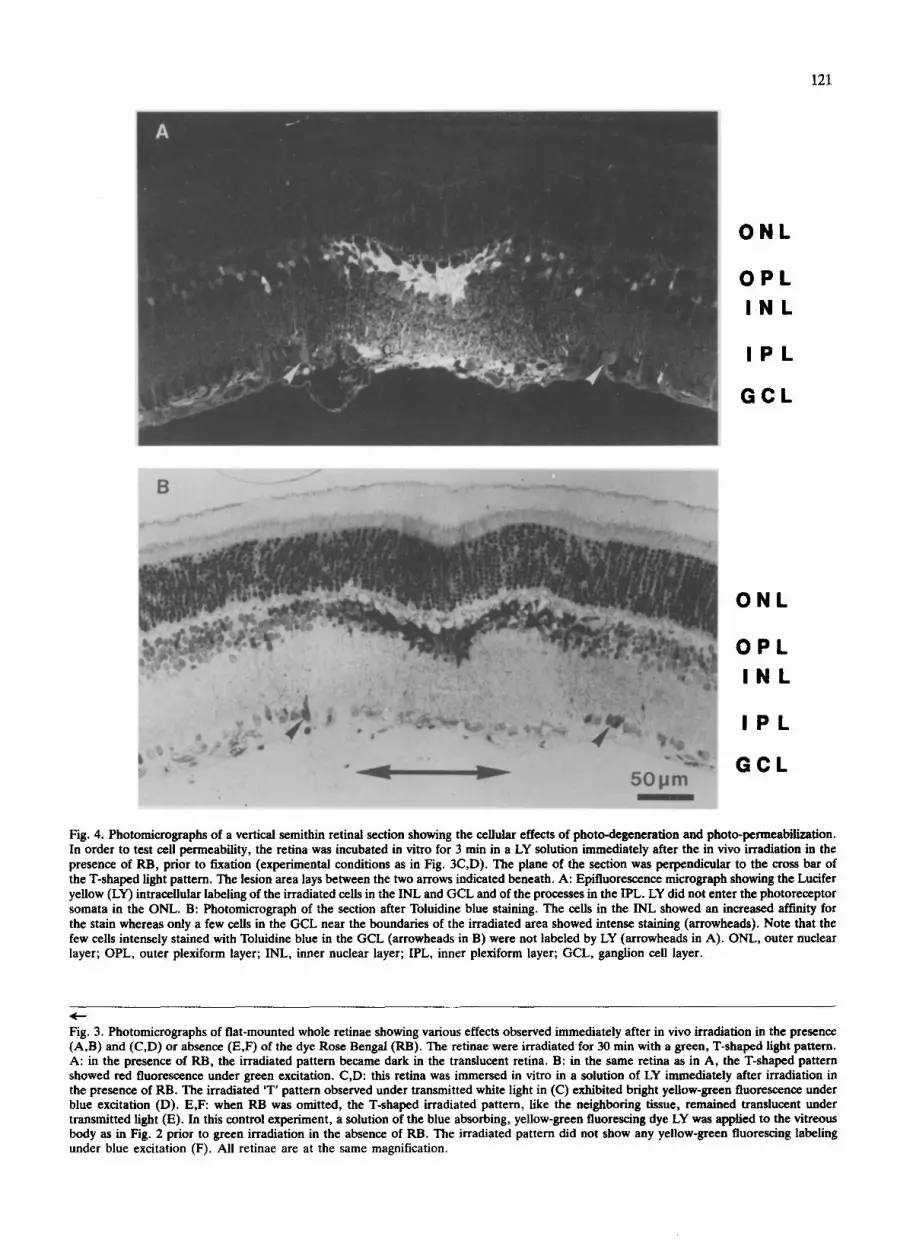
Fig. 4. Photomicrographs of a vertical semithin retinal section showing the cellular effects of photo-degeneration and photo-permeabilization. In order to test cell permeability, the retina was incubated in vitro for 3 min in a LY solution immediately after the in vivo irradiation in the presence of RB, prior to fixation (experimental conditions as in Fig. 3C,D). The plane of the section was perpendicular to the cross bar of the T-shaped light pattern. The lesion area lays between the two arrows indicated beneath. A: Epifluorescence micrograph showing the Lucifer yellow (LY) intracellular labeling of the irradiated cells in the INL and GCL and of the processes in the IPL. LY did not enter the photoreceptor somata in the ONL. B: Photomicrograph of the section after Toluidine blue staining. The cells in the INL showed an increased affinity for the stain whereas only a few cells in the GCL near the boundaries of the irradiated area showed intense staining (arrowheads). Note that the few cells intensely stained with Toluidine blue in the GCL (arrowheads in B) were not labeled by LY (arrowheads in A). ONL, outer nuclear layer; OPL, outer plexiform layer; INL, inner nuclear layer; IPL, inner plexiform layer; GCL, ganglion cell layer.
Fig. 3. Photomicrographs of fiat-mounted whole retinae showing various effects observed immediately after in vivo irradiation in the presence (A,B) and (C,D) or absence (E,F) of the dye Rose Bengal (RB). The retinae were irradiated for 30 rain with a green, T-shaped light pattern. A: in the presence of RB, the irradiated pattern became dark in the translucent retina. B: in the same retina as in A, the T-shaped pattern showed red fluorescence under green excitation. C,D: this retina was immersed in vitro in a solution of LY immediately after irradiation in the presence of RB. The irradiated 'T' pattern observed under transmitted white light in (C) exhibited bright yellow-green fluorescence under blue excitation (D). E,F: when RB was omitted, the T-shaped irradiated pattern, like the neighboring tissue, remained translucent under transmitted light (E). In this control experiment, a solution of the blue absorbing, yellow-green fluorescing dye LY was applied to the vitreous body as in Fig. 2 prior to green irradiation in the absence of RB. The irradiated pattern did not show any yellow-green fluorescing labeling under blue excitation (F). All retinae are at the same magnification.


123
in the INL and GCL, in the processes of the plexiform layers, and in the ganglion cell axons passing through the irradiated area.
Increased cell permeability Retinae irradiated in vivo in the presence of RB and
fixed immediately afterwards showed a red fluorescing T-shaped pattern when examined with the fluorescence microscope under green excitation (Fig. 3B). This pattern was an exact replica of the dark one observed under transmitted light (Fig. 3A). This fluorescent pattern was suggestive of a RB uptake into the irradiated cells.
To determine whether a genuine dye uptake occurred upon green irradiation in the presence of RB, we used a blue absorbing dye, Lucifer yellow (LY). This dye was selected because it affords a yellow-green fluorescence which can easily be distinguished from the red fluores- cence of RB. LY was added to the solution of RB applied to the vitreous body prior to the irradiation. After irradiation, the T-shaped pattern observable under trans- mitted light (Fig. 2A) now showed the characteristic bright yellow-green fluorescence of LY under blue excitation (Fig. 2B) (n = 7). Since in the absence of LY, hardly any fluorescence of the 'T' could ever be detected under blue excitation in similar experiments, the yellow- green labeling illustrated in Fig. 2B shows that LY uptake did occur in the irradiated area. In control experiments without RB application (n = 5), the same green irradi- ation in the presence of LY did not yield any yellow-green labeling of the irradiated area (Fig. 3F). It can therefore be concluded that dye uptake in the irradiated area resulted from the photosensitizing effect of RB on the retinal tissue.
To assess the hypothesis that the cells may have simply been made permeable to dyes, we again used LY as a test substance, but this time in an in vitro assay. Immediately after irradiation in the presence of RB alone, the eye was enucleated, and the eyecup was dissected and incubated in vitro for 3 min in a LY solution prior to fixation. After this in vitro procedure, the T-shaped pattern which first showed up under transmitted white light (Fig. 3C) also displayed a bright yellow-green LY fluorescence under blue excitation (Fig. 3D) (n -- 17). Control retinae (n = 2) with no RB application prior to the green irradiation, further processed with the Lucifer yellow permeability
assay, showed that green light alone did not give rise to the T-shaped LY labeling. The yellow-green labeling of the irradiated area therefore actually served to reveal a latent effect induced by the interaction of light and RB. Note that the various stages in the treatment were strongly reminiscent of the photographic process: expo- sure of the retina impregnated with RB first produced what might be called a 'latent image' which was subse- quently 'developed' in a LY 'bath' prior to 'fixation'.
Vertical sections of an experimental retina incubated in vitro with LY after in vivo irradiation showed the localization of LY in greater detail (Fig. 4A). In the irradiated region, the somata in the INL and GCL as well as the processes in both plexiform layers (ONL, INL) were labeled with LY. Neither the photoreceptors in the ONL nor any of the cells in the neighboring, unirradiated tissue were labeled. These results confirm that the LY labeling observed on the retinal wholemounts resulted from a genuine intracellular staining of the irradiated neurons. As the intraceUular labeling was readily ob- served after a short in vitro incubation of the retina (3 min) in the dye solution, the LY incorporation attested simply dye diffusion into the neurons made permeable by the in vivo irradiation in the presence of RB.
Fig. 4A is in fact a fluorescence micrograph taken prior to the Toluidine blue staining of the section (Fig. 4B). It shows clearly that the increase in Toluidine blue stain- ability mainly occurred in the INL previously labeled with LY. Hence those retinal structures that became permeable to LY also showed signs of dark degeneration. However, the 'dark degeneration' and permeabilization to LY, both indicating photodynamic damage, were not exactly coextensive. Most of the cells in the GCL were labeled with LY but showed normal staining with Tolui- dine blue. Conversely, some cells found to be unlabeled with LY showed the intense Toluidine blue staining indicative of dark degeneration. This typically occurred near the border of the irradiated area in the GCL (arrowheads in Fig. 4A,B). The fact that these cell populations did not completely overlap suggested that permeabilization and dark degeneration may in fact constitute different degrees of damage (see Discussion). They therefore provide two complementary features with which photodynamic damage at the LM level can easily be identified.
Fig. 5. Low-power (250x) electron micrograph of a vertical retinal section showing the ultrastructurai alterations of the tissue irradiated in the presence of RB, contrasting with the neighboring unirradiated tissue. The lesion area lay between the two arrows indicated beneath as in Fig. 4. Photoreceptors do not seem to have been affected by the treatment. Fibers in the OPL- and IPL-like cells in the INL became either electron-dense, or swollen with a loss of cytoplasm. Cells in the GCL exhibited less striking symptoms of injury, such as high vesiculation, a slightly increased cytoplasmic electron density, or clumped chromatin. The axon bundles were disintegrated (arrowhead) and the axonal fibers showed an increased axoplasmic electron density or a swelling associated with the complete loss of the axoplasm.

124
DISCUSSION
Photodynamic damage due to in vivo irradiation in the presence of an extracellular dye has been reported to produce neuronal 'photo-degeneration '19 and 'photo-
permeabilization as in fly photoreceptor neurons. The present study, conducted on the rat retina, demonstrates that various kinds of neurons display similar reactions in vertebrates. The occurrence of cell permeabilization was evidenced by the observed permeation of LY into the photo-treated neurons. The occurrence of neuronal degeneration was attested by classical neuronal degener- ative features, such as the cellular darkening observed in the photo-treated neurons and their subsequent pha- gocytosis by macrophages. A separate paper (Picaud et al., in preparation) will deal more specifically with the removal of degenerating neurons and the sequence of events which accompany healing of the retinal wound.
Evidence for dye-induced photosensitization On the one hand, the low illumination level (2000 lux)
at which these phenomena occurred rules out the possibility that either photothermolysis 2 or photo- coagulation 2° may have been involved. On the other hand, the irradiation period (30 min) was far too short to produce photoreceptor degeneration, as confirmed in the control experiments where the dye was omitted. This is consistent with the finding that at green light illumination levels as low as this (2000 lux, i.e. 185 foot-candles), rat photoreceptor degeneration usually occurs after a 24 h exposure 23. In fact, the presence of RB was necessary, but other highly potent photosensitizers such as Mero- cyanine 540 and Hematoporphyrin were found to pro- duce similar effects, provided the irradiation wavelength was tuned to the absorption band of the dye. The fact that the presence of a dye was required demonstrates the involvement of dye-induced photosensitization 7'22 in the
described phenomena. Contrary to what was observed with photothrombosis,
where RB was applied to the circulating blood 15, the photosensitizing dye was injected here into the vitreous body, where it diffused into the extracellular space of the retinal tissue. The specific experimental conditions and the absence of both haemorrhage and blood vessel occlusion suggest that the kind of retinal injury illustrated here resulted rather from direct photodynamic effects on the retinal neurons, as was found to be the case in the fly retina ~9. The fact that the RB was applied to the extracellular space just prior to irradiation suggests that the actual site of damage was the cell membrane. This putative cellular site of photooxidation is in agreement with the finding that RB phototoxicity can affect neuro- nal membranes from either side 17. As observed in the
latter study, dye-induced photosensitization caused alter- ations in ionic channels but membrane lipids too are known to undergo photooxidation in the presence of a dye, which has adverse effects upon cell integrity and viabilityT,22,
Permeability of neurons after photodynamic damage Here we demonstrated with the rat retina that in vivo
irradiation in the presence of a photosensitizing dye can induce permeabilization of the irradiated cells. This finding extends to vertebrate neurons the phenomena originally observed in fly photoreceptors TM. Since retinal
cells incubated in RB were not permeable to LY unless irradiated, the in vitro cell permeability assay can be taken to constitute a valid test of cell viability. Hence 'photo-permeabilization' indicates a loss of cell viability and provides the first evidence that photodynamic dam- age occurred in these vertebrate neurons. Cell permea- bilization following a similar dye-induced photosensitiza- tion was previously reported on cell cultures 6'21 and
isolated membranes 8 on the basis of the leakage of
intracellular molecules such as ions, sugars and even proteins s'21 or the permeation of the dye Trypan blue 6.
Here we used the dye LY to assess cell permeability in the retinal tissue because it exhibits an intense fluorescence which can easily be distinguished from the RB fluores- cence and because it has proven to be a very effective means of assessing the permeability of fly photoreceptor cells in vivo TM. The use of a fluorescent dye such as LY
enabled us to detect the damaged cells even at the macroscopic level (Figs. 2B, 3D, 4A). This 'development ' of the treated area was usually achieved either by subsequently incubating the retinal tissue in vitro in the LY solution (e.g. Fig. 3D), or by simply adding LY to the photosensitizer solution applied in vivo to the vitreous body (e.g. Fig. 2B). The achieved cell-permeabilization to LY therefore results in specific cellular labeling and thus provides an efficient means of detecting photody- namic damage in situ or in vitro at macroscopic and cellular levels.
The T-shaped pattern which showed up after irradia- tion in the presence of RB exhibited a red fluorescent labeling under green excitation (Fig. 3B). The very fact that a concomitant photo-permeabilization took place suggested that the RB photosensitizer itself may have diffused into the damaged cells. However, the relatively long irradiation period (30 min) may have stimulated a dye uptake via endocytosis as has been suggested in other studies 14'24. Although its molecular basis should be interpreted with caution, the fluorescent labeling ob- served upon excitation within the absorption range of the photosensitizer is a convenient means of locating pho- todynamic damage at the macroscopic level.

Structural changes following photodynamic damage After irradiation in the presence of RB, most of the
cells in the INL and a few cells in the GCL became electron dense in the irradiated area (Fig. 5). This increase in the electron density of the cytoplasm is known to be the most striking histological symptom of degen- erating neurons ('dark degeneration') 16. The increased electron opaqueness of 'dark neurons' was paralleled by their increased stainability with Toluidine blue in semi- thin sections. This process clearly affected various types of neurons in the irradiated area. The results obtained here considerably extend the original report dealing exclusively with one class of (photoreceptive) neurons 19. The photodynamic effect of dyes therefore provides a novel and powerful means of triggering dark degenera- tion in neurons.
When the correlation between dark degeneration and cell permeabilization was analysed, a number of cells at the edges of the irradiated area in the GCL showed dark degeneration without any concomitant permeabilization to LY. The fact that these neurons lay at the periphery of the irradiated area suggests that they were less severely impaired by the treatment. Hence the range of damage leading to dark degeneration might be gradual, the injured neurons becoming either simply dark neurons or permeable dark neurons. The gradual transition observed at the edges of the irradiated area (Fig. 5) may be attributable to stray light and/or to retrograde/antero- grade degeneration of those neurones having neurites within the lesion area. Moreover, permeabilized cells may have released intracellular molecules such as ions and glutamate, thereby inducing neurotoxicity n in the adjoining tissue, which would account for the apparent 'spillover' of the degeneration.
Although most of the cells in the GCL did not show a genuine dark degeneration, their degenerative state was attested by their ultrastructural appearance and their permeability to dyes. In agreement with previous obser- vations on fly photoreceptor neurons 18'19, the occurrence of these degenerative features suggests that permeable cells in the GCL were more severely affected by the procedure than the injured cells in the INL (which were mostly in a state of 'dark degeneration'). Hence the procedure resulted in different degrees of damage across the retinal layers. The photoreceptor cell bodies in the ONL were apparently unaffected, most of the somata in the INL showed a conspicuous dark degeneration, and the somata in the GCL showed other features indicative of degeneration which were not so striking in the light microscopic picture but indicated heavier damage. Both grades of damage, however, included photo-permeabili- zation. The vertical gradient of photodynamic damage in the rat retina may reflect the graded diffusion of RB from
125
the vitreous body into the retinal layers as well as a gradual attenuation of the light through the successive retinal layers (the absorbance of which was not negligible since the retina was impregnated with RB). This suggests that photodynamic action may trigger dark degeneration in a neuron only when the local product of 'light dose x dye concentration' reaches a given value. This hypothe- sis, which it is far beyond the scope of this paper to test, is in agreement with some preliminary results obtained on individual fly photoreceptors 19.
At the macroscopic level, the structural alterations induced in the irradiated cells resulted in a darkening of the tissue, which was very reproducible and convenient for recovering the irradiated retinal area (Figs. 2A, 3A,C). At the cellular level, the degenerative effects in the INL were sometimes less extensive than in the case illustrated in Figs. 4 and 5. This variability may be due to variations in the diffusion gradient of RB into the retina prior to irradiation, a gradient which itself may account for the absence of photoreceptor damage, as mentioned above.
Concluding remarks Photodynamic effects are usually thought to be a
major drawback when dyes are used to study living nervous tissue. They may, however, be a potentially useful tool for producing specific lesions in the nervous system. For instance, photodynamic action was reported to trigger dark degeneration and to induce cell permea- bilization and cell deletion in fly photoreceptor neu- rons 18'19. The present study has extended to vertebrate neural tissue these original findings on neuronal photo- degeneration and photo-permeabilization. In particular, we have shown that these phenomena can also occur in vertebrate non-photoreceptive neurons, where photody- namic damage can be identified at the macroscopic, cellular and ultrastructural levels. It now appears likely that these phenomena are common to all kinds of neurons. The procedure requires application of a dye to the extracellular space and irradiation of the living tissue. Light intensity in a physiological range can be used, as illustrated here. Due to its accuracy and reproducibility, the photo-lesion technique holds great potential as a tool for investigating various nervous systems.
A most interesting way of extending these findings would be to couple a photosensitizing dye to molecules such as antibodies targeted to the outer surface of specific neurons. Such molecules would home in on the requisite neurons and equip them with a photosensitizing tag. Target neurons, the sole absorbing entities in an appro- priate beam of light, would undergo photo-degeneration and photo-permeabilization and could possibly be erad- icated from the tissue in a similar way to what occurs with

126
t u m o r cells in ' p h o t o i m m u n o t h e r a p y '~2. With this meth-
od the spatial and spectral select ivi ty of i r rad ia t ion wou ld
be added to the chemical select ivi ty o f the h o m i n g
molecu le . P rocedu re s o f this k ind may he lp to d isentan-
gle n e u r o n a l n e t w o r k s by causing a chemica l ly prese lec-
ted popu l a t i on o f neu rons to p h o t o - d e g e n e r a t e on com-
mand .
Acknowledgements. We thank H. W~issle for his generous support
throughout this study and J. Blanc, M:H. Chun, A. Karschin, B. Patel and J. Schnitzer for their contributions and comments. The technical assistance of H. Ahmed, G.S. Nam, W. Hofer and A. Totin is gratefully acknowledged. S. Picaud received a 'Procope' fellowship from the Minist~re des Affaires Etrang6res during his National Service, a post-doctoral fellowship from the Max-Planck- Gesellschaft, financial support from the University of Marseilles and a fellowship from the Fondation de la Vocation. This study was supported by the Max-Planck-Gesellschaft (F.R.G.), C.N.R.S., D.R.E.T. and Fondation pour la Recherche Medicale (France) and by the Committee for the Development of European Science and Technology (CODEST, E.E.C.).
REFERENCES
1 Ames III, A. and Nesbett, F.B., In vitro retina as an experi- mental model of the central nervous system, J. Neurochem., 37 (1981) 867-877.
2 Anderson, R.R. and Parrish, J.A., Selective photothermolysis: precise microsurgery by selective absorption of pulsed radiation, Science, 220 (1983) 524-527.
3 Chalazonitis, N. and Chagneux, R., Photopotentials of the Sepia giant axons sensitized to light, Bull. Inst. Ocean. Monaco, 1223 (1961) 1-21.
4 De Weer, E and Salzberg, B.M., Optical Methods in Cell Physiology, Society of General Physiologists Ser., Vol. 40, Wiley, New York, 1986, 480 pp.
5 Dougherty, T.J., Photodynamic therapy of cancer. In T. Hasan (Ed.), Advances in Photochemotherapy, Proceeding of the International Society for Optical Engineering, Vol. 997, SPIE, Bellingham, U.S.A., 1988, pp. 2-7.
6 Easton, T.G., Valinsky, I.E. and Reich, E., Merocyanine 540 as a fluorescent probe of membranes: staining of electrically excitable cells, Cell, 13 (1978) 475-486.
7 Girotti, A.W., Mechanisms of photosensitization, Photochem. Photobiol., 38 (1983) 745-751.
8 Girotti, A.W. and Deziel, M.R., Photodynamic action of protoporphyrin on resealed erythrocyte membranes: mecha- nisms of release of trapped markers. In D. Kessel and T.J. Dougherty (Eds.), Porphyrin Photosensitization, Advances in Experimental Medicine and Biology Ser., Vol. 160, Plenum, New York, 1983, pp. 213-225.
9 Lamberts, J.J.M. and Neckers, D.C., Rose bengal derivatives as singlet oxygen sensitizers, Tetrahedron, 41 (1985) 2183-2190.
10 Lippay, E, Ueber Wirkungen des Lichtes auf den quergestreif- ten Muskel. I. Mitteilung, Versuche mit sichtbarem Licht an sensibilisierten Kaltbluetermusketn, Pfluegers Arch. Ges. Phys- iol., 222 (1929) 616-639.
11 Mayer, M.L. and Westbrook, G.L., Cellular mechanisms un- derlying excitotoxicity, Trends Neurosci., 10 (1987) 59-61.
12 Mew, D., Wat, C., Towers, N.G.H. and Levy, J.G., Photoim- munotherapy: treatment of animal tumors with tumor-specific monoclonal antibody-Hematoporphyrin conjugates, J. lmmu-
nol., 130 (1983) 1473-1477. 13 Miller, J.P. and Selverston, A.I., Rapid killing of single neurons
by irradiation of intracellularly injected dye, Science, 206 (t979) 702-704.
14 Moan, I., Johannessen, J.V., Christensen, T., Espevik, T. and McGhie, J.B., Porphyrin-sensitized photoinactivation of human cells in vitro, Am. J. Pathol., 109 (1982) 184-192.
15 Mosinger, J.L. and Olney, J.W., Photothrombosis-induced ischemic neuronal degeneration in the rat retina, Exp. Neurol., 105 (1989) 110-113.
16 Mugnaini, E. and Friedrich Jr., V.L., Electron microscopy: identification and study of normal and degenerating neural elements by electron microscopy. In L. Heimer and M.J. Robards (Eds.), Neuroanatomical Tract-Tracing Methods, Ple- num, New York, 1981, pp. 377-406.
17 Oxford, G.S., Pooler, J.P. and Narahashi, T., Internal and external application of photodynamic sensitizers on squid giant axons, J. Mernbr. Biol., 36 (1977) 159-173.
18 Picaud, S., Wunderer, H.J. and Franceschini, N., Dye-induced photopermeabilization and photodegeneration: a lesion tech- nique useful for neuronal tracing, J. Neurosci. Methods, in press.
19 Picaud, S., Wunderer, H.J. and Franceschini, N., 'Photo- degeneration' of neurones after extracellular dye application, Neurosci. Lea., 95 (1988) 24-30.
20 Pomerantzeff, O., Wang, G.J., Pankratov, M. and Schneider, J., Time and location analysis of lesion formation in photocoa- gulation, Arch. Ophthalmol., 101 (1983) 954-957.
21 Sandberg, S., Glette, J., Hopen, G., Solberg, C.O. and Romslo, I., Porphyrin-induced photodamage to isolated human neutro- phils, Photochem. Photobiol., 34 (1981) 471-475.
22 Spikes, I.D., Photosensitization. In K.C. Smith (Ed.), The Science of Photobiology, 2nd edn., Plenum, 1989, pp. 79-110.
23 Tso, M.O.M., Experiments on visual cells by nature and man: in search of treatment for photoreceptor degeneration, Frie- denwald lecture, Invest. Ophthalmol. Vis. Sci., 30 (1989) 2430-2454.
24 Wilcox, M. and Franceschini, N., Illumination induces dye incorporation in photoreceptor cells, Science, 225 (1984) 851- 854.





























