Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Florence Guillemin Æ Fabienne Guillon Æ Estelle Bonnin
Marie-Francoise Devaux Æ Therese Chevalier
J Paul Knox Æ Francoise Liners Æ Jean-Francois Thibault
Distribution of pectic epitopes in cell walls of the sugar beet root
Received: 6 January 2005 / Accepted: 28 February 2005 / Published online: 11 May 2005� Springer-Verlag 2005
Abstract Immunolabelling techniques with antibodiesspecific to partially methyl-esterified homogalacturonan(JIM5: unesterified residues flanked by methylesterifiedresidues. JIM7: methyl-esterified residues flanked byunesterified residues), a blockwise de-esterified homo-galacturonan (2F4), 1,4-galactan (LM5) and 1,5-arab-inan (LM6) were used to map the distribution ofpectin motifs in cell walls of sugar beet root (Betavulgaris). PME and alkali treatments of sections wereused in conjunction with JIM5-7 and 2F4. The JIM7epitope was abundant and equally distributed in allcells. In storage parenchyma, the JIM5 epitope wasrestricted to some cell junctions and the lining ofintercellular spaces while in vascular tissues it occurredat cell junctions in some phloem walls and in xylemderivatives. After secondary wall formation, the JIM5epitope was restricted to inner cell wall regions be-tween secondary thickenings. The 2F4 epitope was notdetected without de-esterification treatment. PMEtreatments prior to the use of 2F4 indicated that HGat cell corners was not acetylated. The LM5 epitopewas mainly present in the cambial zone and whenpresent in storage parenchyma, it was restricted to the
wall region closest to the plasma membrane. The LM6epitope was widely distributed throughout primarywalls but was more abundant in bundles than in me-dullar ray tissue and storage parenchyma. These datashow that the occurrence of oligosaccharide motifs ofpectic polysaccharides are spatially regulated in sugarbeet root cell walls and that the spatial patterns varybetween cell types suggesting that structural variants ofpectic polymers are involved in the modulation of cellwall properties.
Keywords Cell wall Æ Pectin (immunolocalization) ÆCambium Æ Differentiation Æ Sugar beet root (pectin)
Abbreviations GalA: Galacturonic acid Æ Ara:Arabinose Æ Gal: Galactose Æ Rha: Rhamnose Æ HG:Homogalacturonan Æ RGI: Rhamnogalacturonan I ÆPME: Pectin methylesterase Æ PAE: Pectinacetylesterase
Introduction
Pectins are the major matrix components of dicotyle-donous cell walls. They are thought to contribute tocell adhesion and play a role in the mechanical andtextural properties of plant organs. The term pectinsrefers to a complex and heterogeneous family ofpolysaccharides in which galacturonic acid (GalA) andrhamnose (Rha) are the common features (see reviewsby Willats et al. 2001a). Different structural domainsare generally described: contiguous (1–4)-a-linked GalA(homogalacturonan, HG), (1–4)-a-linked GalA inter-rupted by the insertion of (1–2) linked a-L-Rha (type Irhamnogalacturonan, RGI) and a substituted galactu-ronan (type II rhamnogalacturonan, RGII), whichexhibits a very complex structure with a diversity ofsugars and linkages. HG can contain 100–200 residues(Bonnin et al. 2002) in which the GalA can be methyl-esterified at position 6, acetylated at position 2 and/or
F. Guillemin Æ F. Guillon (&) Æ E. Bonnin Æ M.-F. DevauxT. Chevalier Æ J.-F. ThibaultInstitut National de la Recherche Agronomique,Unite de Recherche sur les Polysaccharides,leurs Organisations et Interactions,rue de la Geraudiere, BP 71627,44316 Nantes cedex 03, FranceE-mail: [email protected].: +33-24-0675016Fax: +33-24-0675066
J. P. KnoxCentre for Plant Sciences,University of Leeds,Leeds, LS2 9JT, UK
F. LinersUnite de Recherche en Biologie Cellulaire et Moleculaire Vegetale,Facultes Universitaires Notre-Dame de la Paix of Namur,rue de Bruxelles 61, 5000 Namur, Belgium
Planta (2005) 222: 355–371DOI 10.1007/s00425-005-1535-3

3 (Quemener et al. 2003) or substituted on O-2 and/orO-3 by xylose or apiose unit or short 2-linked xyloseside chains (Le Goff et al. 2001). In muro, the molarratio of methanol to GalA (DM) and the distributionof methyl groups strongly depend on the activities ofendogenous pectin methylesterases. These features areimportant as they determine the gelling capacity ofpectins and can influence cell wall stiffening (Jarviset al. 2003). In many cell walls, acetyl groups appear tobe concentrated in the backbone of RGI (Schols andVoragen 1994; Ishii 1995, 1997). However, abundantacetylation of HG has been reported for sugar beet(Rombouts and Thibault 1986) and for potato tubercell walls (Ishii 1997). The role of acetylation is still notunderstood even though it is known than it can de-crease the gelling of pectins (Renard and Jarvis 1999a;Ralet et al. 2003), the degradation by endopolygalac-turonases (Bonnin et al. 2003; Buccholt et al. 2004) andcell adhesion (Liners et al. 1994).
Neutral sugar side chains composed predominantlyof (1–4)-b-D-galactan and/or (1–5)-a-L-arabinan aregenerally attached onto O-4 and sometimes on O-3 ofthe Rha units of RGI. Occurence of type II arabino-galactans, which consist of (1–3)-b-D-galactan carryingat position O-6 (1–6)-b-D-galactan side chains were alsoreported (Guillon and Thibault 1989). In some pectins,for example sugar beet, neutral sugars can be esterifiedby small quantities of ferulic acid (Rombouts andThibault 1986; Guillon and Thibault 1989; Levigneet al. 2004a). The proportion of the different sidechains, the degree of branching and the chain lengthvary according to the pectin source (see review by2001a).
To increase the understanding of the structure/function relationships of pectins in planta, the distri-bution of the structural domains of pectins have beeninvestigated using immunocytochemical approaches(see Willats et al. 2001a). Taken together, observationsindicate that low methyl-esterified or de-esterified HGare often localized to middle lamellae, cell corners andaround air spaces whereas esterified pectins are dis-tributed throughout the cell wall (Liners and VanCutsem 1992; Liners et al. 1994; Guglielmino et al.1997; Bush and McCann 1999; Willats et al. 1998,1999b; Ermel et al. 2000; Willats et al. 2001b). RGIIoccurs mainly in all primary cell walls but is not de-tected in middle lamellae (Matoh et al. 1998). Thepresence and location of galactan and arabinan sidechains are extensively developmentally regulated (Joneset al. 1997; Willats et al. 1998, 1996b; Bush andMcCann 1999; Ermel et al. 2000; Willats et al. 2001b).For example, an appearance of galactan and a decreasein arabinan in carrot cell walls were correlated with thetransition from cell division to cell elongation (Willatset al. 1996b). In hybrid aspen shots, during the sec-ondary growth, the LM5 galactan epitope is present inthe meristematic cells of cambial zone and enlargingxylem cells and is lost through the course of derivativesentering the phloem pathway (Ermel et al. 2000).
Arabinan and galactan are generally absent from themiddle lamella, and when present in the same cell, theycan have distinct localizations within the cell wall. Inplant systems in which the galactan epitope appearsduring cell differentiation, it is localized to a zone closeto the plasma membrane (Jones et al. 1997; Bush andMcCann 1999; McCartney et al. 2000) while arabinanis detected throughout the primary cell wall (Bush andMcCann 1999; McCartney et al. 2000).
The differential occurrence of pectin epitopes sug-gests that the different structural features of pectinshave diverse roles in cell development. The purpose ofthis study was to investigate the modulations of pectinstructure in a species with distinctive pectin chemistry.Sugar beet roots contain a large amount of pectins (upto 40–50% of the cell wall dry matter), which havebeen extensively studied (Rombout and Thibault 1986;Guillon and Thibault 1989; Guillon et al. 1989; Oo-sterveld et al. 2000). Sugar beet pectins do not res-semble commonly described pectins in term of HGacetylation and biochemistry of pectic side chains andso the the capacity of anti-pectin antibodies to rec-ognize sugar beet pectins was first assessed by im-muno-dot assays with a series of sugar beet pectinsamples with different chemical compositions andsubstitutions.
Materials and methods
Plant material
Sugar beet roots (Beta vulgaris, cv Roberta) were kindlyprovided by INRA d’Estree Mons (France).
Monoclonal antibodies
The antibodies used in this study were the rat mono-clonal antibodies JIM7, JIM5, LM5 and LM6 and themouse monoclonal antibody 2F4. Primary references forthe generation of these antibodies and epitopes recog-nized are shown in Table 1.
Pectins
A series of beet pectins with different DM and DA(molar ratio acetic acid/ GalA) and varied neutral sidechain compositions were prepared according to Levigneet al. (2002a).
Enzymatic treatments
Two pectin methyl-esterases, one from orange peel(oPME) (Sigma) and one from Aspergillus aculeatus(fPME) (Novozymes, Denmark) and a pectin acetyl-esterase (PAE) from A. aculeatus (Bonnin et al. 2005,submitted for publication) were used.
356

Pectins (10 mg ml�1) were dissolved in 0.1 M citratebuffer (pH between 4.5 and 6.0 according to the enzymeused). Enzymes were added and incubations were car-ried out at 30 �C. The final concentration of enzymesnecessary to reach the target composition was calculatedfrom the known enzyme activities and the initial com-position of pectins. The time of incubation was 4 h forall the enzymes except PAE, for which it was 40 h. Afterthe degradation, the solutions were dialysed againstdeionized water and retentates were freeze-dried. In thecase of the sequential degradations, the pH between twotreatments was adjusted to the right value by either0.2 M sodium citrate or 0.2 M citric acid.
Chemical de-esterification treaments
Extensive de-esterification of Gal A were performed bybase (NaOH and Na2CO3) catalysed de-esterification.Samples of fresh aqueous solution (10 mg ml�1) ofpectins were prepared and cold 1-M NaOH was addedto the pectin solution in order to reach a pH of 12.5. Thesolution was left under agitation for 30 min at 4 �C.Then, the pH was adjusted to 4.5 with 1-M HCl and thesolution dialysed to remove the released methanol, ace-tic acid and salts. The retentate was freeze-dried. For lessextensive de-esterification, Na2CO3 (1 M, pH 12) waschosen. The reaction was carried out as described above.
Analytical methods
Uronic acid contents were determined by the m-hy-droxybiphenyl assay (Thibault 1979). DM and DA weredetermined according to Levigne et al. 2002b.
Immuno-dot assays
Pectic samples (10 mg ml�1) were dissolved in water(containing 0.05% sodium azide) and applied by hand as1 ll aliquots to nitrocellulose in a five- or ten-folddilution series. The nitrocellulose membranes were leftto dry for 1 h. Assays with JIM5, JIM7, LM5 and LM6were carried out as described by Willats et al. (2000).Assays with 2F4 were carried out as described byRenard et al. (1993).
Preparation of specimens for microscopy
Fresh sections
Cross-sections (300-lm thick) were hand-cut from freshsugar beet roots. The sections were left overnight at 4 �Cin deionized water containing 0.01% sodium azide inorder to remove sucrose. Then, the sections were furtherprocessed for immunolabelling.
Embedded sections
Pieces (1 mm3 for light microscopy and <1 mm3 forelectron microscopy) were sampled from fresh sugarbeet roots. Two types of samples were taken, the firsttype containing storage parenchyma and the second typecontaining bundles. The samples were fixed in 2.5% (w/v) glutaraldehyde in 0.1-M Na phosphate buffer(pH 5.9) overnight at room temperature. Samples weremaintained under vacuum for the first 30 min of thefixation step. Samples were washed with the same bufferfollowed with washes with deionized water. They werethen dehydrated in a graded aqueous ethanol series.Dehydrated cubes were progressively infiltrated withLondon Resin White (LRW) acrylic resin and thenembedded in gelatine capsules. The resin was polymer-ized for 4 days at 60 �C. The blocks were labelled eitheras parenchyma blocks or bundle blocks. Semi-thin sec-tions (1-lm thick) were prepared and mounted onmultiwell glass sides pre-treated with VectaBond sectionadhesive (British BioCell International, UK). Ultrathinsections for electron microscopy were collected on car-bon coated nickel grids.
Immunolabelling of pectin epitopes
Prior to incubation with JIM5, JIM7, LM5, LM6, sec-tions were blocked with 0.1 M phosphate buffered saline(pH 7.2) containing 3% (w/v) bovine serum albumin(PBS/BSA) for 1 h at room temperature. Sections to beincubated with 2F4 were blocked with 3% (w/v) skim-med milk in 20 mM Tris–HCl, pH 8.2, 0.5 mM CaCl2,150 mM NaCl (TcaS buffer) for 1 h at room tempera-ture. The Abs was applied to the sections 1 h at roomtemperature. JIM5, JIM7, LM5, LM6 were diluted 1:10(v/v) in PBS/BSA while 2F4 was diluted two-fold in 1%
Table 1 Monoclonal antibodies used to map the distribution of pectic epitopes in the cell walls of the sugar beet root
Mabs Antigen/epitope References
JIM5 Partially methyl-esterified HG epitope: unesterified residuesadjacent to or flanked by residues with methylester groups
Clausen et al. (2003)
JIM7 Partially methyl-esterified HG epitope: methyl-esterified residueswith adjacent or flanking unesterified GalA / alternatingmethyl-esterified GalA residues
Clausen et al. (2003)
2F4 De-esterified HG epitope: calcium requiring configuration of HG Liners et al. (1989, 1992)LM5 (1 fi 4)-b-D-galactan Jones et al. (1997)LM6 (1 fi 5)-a-L-arabinan Willats et al. (1998)
357

skimmed milk in TcaS buffer. The sections were thenwashed with several changes in an excess of the bufferused for diluting the Abs and then incubated for 1 h atroom temperature with the secondary Abs (anti-rat oranti-mouse IgG according to the animal species fromwhich the primary Abs were raised) coupled to Alexa-Fluor 546 diluted 1:100 (v/v) in the respective bufferused for diluting primary Abs. Sections were washed inbuffer as described above and finally rinsed twice indistilled water. The semi-thin embedded sections wereexamined on a LEICA DMRD microscope equippedwith epifluorescence irradiation (light microscopy LM).A band-pass filter 515–560 nm was used as excitationfilters and fluorescence detected at >570 nm. Immu-nolabelling with JIM5 on fresh hand cut sections wascarried out as described above except for the incubationtime (4 h) and temperature (at 4 �C) with antibodies(primary and secondary). Gentle rocking was main-tained during immunolabelling. The sections wereexamined on a laser scanning confocal microscope(ZEISS LSM 410, (LSCM)). Excitation was at 546 nmand fluorescence was detected at >570 nm. For electronmicroscopy (TEM), sections were treated following thesame procedure than that of resin embedded semi-thinsections, using as secondary Abs, gold conjugates (anti-rat-IgG or anti-mouse-IgG conjugated with 10 nm or1 nm colloidal gold complexes, Aurion, NL), diluted1:20 (v/v) in the respective buffer used for diluting theprimary Abs. After washing, the grids were stained with1% uranyl acetate.
Labelling with 1 nm gold conjugates was intensifiedwith the silver enhancement kits (Aurion, NL) accordingto the manufacturer’s instructions.
In some cases, thin sections were submitted to enzy-matic and chemical treatments, before immunolabelling,for in situ de-esterification of pectins; enzymatic de-esterification was carried out by incubated the sectionswith 50 ll of oPME at 1 mg ml�1 in 0.1 M citrate buf-fer, pH 6 for 30 min at 30 �C. For chemical de-esterifi-cation, the sections were incubated with 50 ll of 0.05 MNaOH (pH 12.6) or 0.1 M Na2CO3 (pH 11.8) for30 min at 4 �C. Sections were rinsed thoroughly withdeionized water. Sections were examined with a JEOL100S transmission with an accelerating voltage of 80 kV.Control sections were treated in parallel but with theomission of the primary Abs.
Results
Binding of monoclonal antibodies to sugar beet pectinsand derived fractions
Sugar beet pectins with different chemical compositions(Table 2), different extents (DM) and patterns ofmethyl-esterification and different degrees of acetylation(DA) were used to assess Ab binding to sugar beetpectins in immunodot assays as shown in Fig. 1. TheGalA content of the pectins varied from 29/ to 46%, and
that of Rha from 1.4/ to 5.4%. The amount of Araexhibited the largest variation from 0.6% to 35.7%, dueto the acid-labile nature of the linkage between arabi-nosyl residues. The Gal content varied between 3.2/ and9.5% and the DM varied between 43/ and 77% and DAbetween 10/ and 43%. PME, PAE and alkali treamentswere applied on the reference pectin C to vary the degreeand pattern of esterification. Pectin with a blockwisedistribution of de-esterified GalA residues was preparedusing oPME, while another with a random distributionof esters was obtained using fPME. The DM decreaseddown to 34 and 29 for oPME and fPME, respectively.Using PAE, the DA decreased from 26 to 16. Sequentialtreatments involving PAE and oPME were applied tofurther decrease the DM (DE 19). Extensive de-esterifi-cation of GalA was performed by base (NaOH orNa2CO3) catalysed de-esterification. The NaOH andNa2CO3 acted to the same extent. The DM was de-creased from 53 to 14–16 and the DA from 26 to 16–17(Table 2).
JIM7 bound strongly to pectins A to SP1 detectingthem down to 10 ng but to a lesser extent with SP3, SP4,SP5, SP6. JIM5 bound more strongly to all pectinsamples than JIM7. DM or DA alone could not explainthe variations observed in the JIM5 binding. Forexample, JIM5 bound more strongly to SP3 than to SP4,although these pectins have similar DM (51). Bothantibodies equally bound to pectin C and SP1, pectinswith same DM (65) and different DA (28 and 10). ThefPME-treated pectins (DM 29) exhibited the lowestbinding whilst base de-esterified pectins in spite of theirlow degree of esterification (DM 14–16; DA 17–16) stillreacted with JIM5.
2F4 bound weakly to pectins A to SP1 and did notbind to SP3–SP6. This lack of binding was expected asthese pectins have high degrees of esterification and nota blockwise distribution of de-esterified GalA. Indeed,strong binding was observed after all the de-methyl-esterification treatments. De-acetylation of C (DA, 17,19) with PAE when associated with oPME did not in-crease 2F4 binding, indicating that DA up to 26 is notcritical for 2F4 binding.
LM5 bound equally to samples with Gal contentfrom 9.5 to 3.2 indicating that these pectins contain atleast four consecutive units of 1,4-b-linked Gal (Joneset al. 1997). LM6 did not bind well to the pectins havingthe lowest arabinose content (<1.5%) (G and SP1), butbound equally to all other pectins apparently in-dependently of the Ara content (3–36.5%) indicatingthat unbranched sequences of 5–6 1,5-a-linked Ara re-sidues were present.
Distribution of pectic epitopes in cell wallsof the sugar beet root
Sections through storage parenchyma and vascular re-gions of the sugar beet root were examined by LM tolocate pectic epitopes in tissues and cells. TEM was
358

used to determine the distribution of pectic epitopeswithin cell walls. Enzymatic and alkali de-esterificationtreatments were applied to sections to gain insight in toany spatial regulation of the methyl- and acetyl-esteri-fication patterns of pectins in sugar beet root cell walls.All cell walls remained unlabelled when the primarymAbs were omitted from control sections in both LMand TEM.
Distribution of de-esterified, methyl-esterifiedand acetylated homogalacturonan
The JIM7 epitope was abundant and equally distrib-uted in all parenchyma and vascular tissue cell walls(Fig. 2a, b). After NaOH treatment, no labelling was
found in cell walls whatever the tissue origin, whilelabelling remained intense after de-esterification withNa2CO3 (not shown). This suggests that de-methy-lesterification with NaOH was more extensive thanwith Na2CO3, so that the epitope recognized by JIM7was no longer present. In contrast, the JIM5 epitopewas less abundant and not evenly distributed throughtthe walls of bundles and parenchyma cells (Fig. 2c, d).We first checked that the low level of JIM5 bindingwas not due to instability of the pectins duringsample preparation for microscopy. Immunolabellingperformed on hand cut sections and then imaged withLSCM confirmed the observations made with resin-embedded sections (Fig. 3). In medullar ray andstorage parenchyma cell walls, labelling was faint(Figs. 2c, d, 3a, c) and more intense at cell corners. In
Fig. 1 Immuno-dot assays ofmonoclonal antibodies JIM7(a, b), JIM5 (c, d), 2F4 (e, f),LM5 (g), LM6 (h) binding tosugar beet pectin samples.a, c, e, g, h Pectin series wereprovided by Levigne et al.(2002a); b, d, f pectin C wasde-esterified by enzymatic orbase-catalysed treatments
359

vascular tissue, phloem and xylem parenchyma cellwalls were bound strongly by JIM5 while the cam-bium zone exhibited a lower level of labelling(Figs. 2c, 3b). The JIM5 epitope was more abundantin radial walls of the cambial zone and at cell junc-tions between some phloem cell walls and in still-expanding xylem derivatives. After de-esterificationwith Na2CO3 (Fig. 2e, f) or with NaOH (not shown),JIM5 labelled all cell walls. Cell walls in sectionswithout treatment did not label with 2F4 (not shown).After oPME treatment, labelling was mainly found atcell junctions (Fig. 2g, h). The cell corners of thephloem and xylem tissues were more intensivelylabelled than the surrounding parenchyma cell walls.After de-esterification with Na2CO3, cell junctions inphloem tissues were labelled moderately (not shown).After NaOH saponification of sections, the 2F4labelled the entire primary cell walls of all cells(Fig. 2i, j). This indicates that NaOH saponification ofsections was more efficient in removing both acetyland methyl ester groups than Na2CO3 saponification.This strongly suggest that methyl esters are lessresistant to saponification than acetyl ester linkages.This observation is also supported by a study, whichdemonstrated that longer incubation times in thepresence of cold dilute alkali was necessary for acomplete de-acetylation of pectins (Buchholt et al.2004).
At the ultrastructural level, all cell walls except thoseof vessels and vessel adjacent cells were labelled acrosstheir width and entire length by JIM7 (Fig. 4a, e, f).This labelling was still present in primary walls of
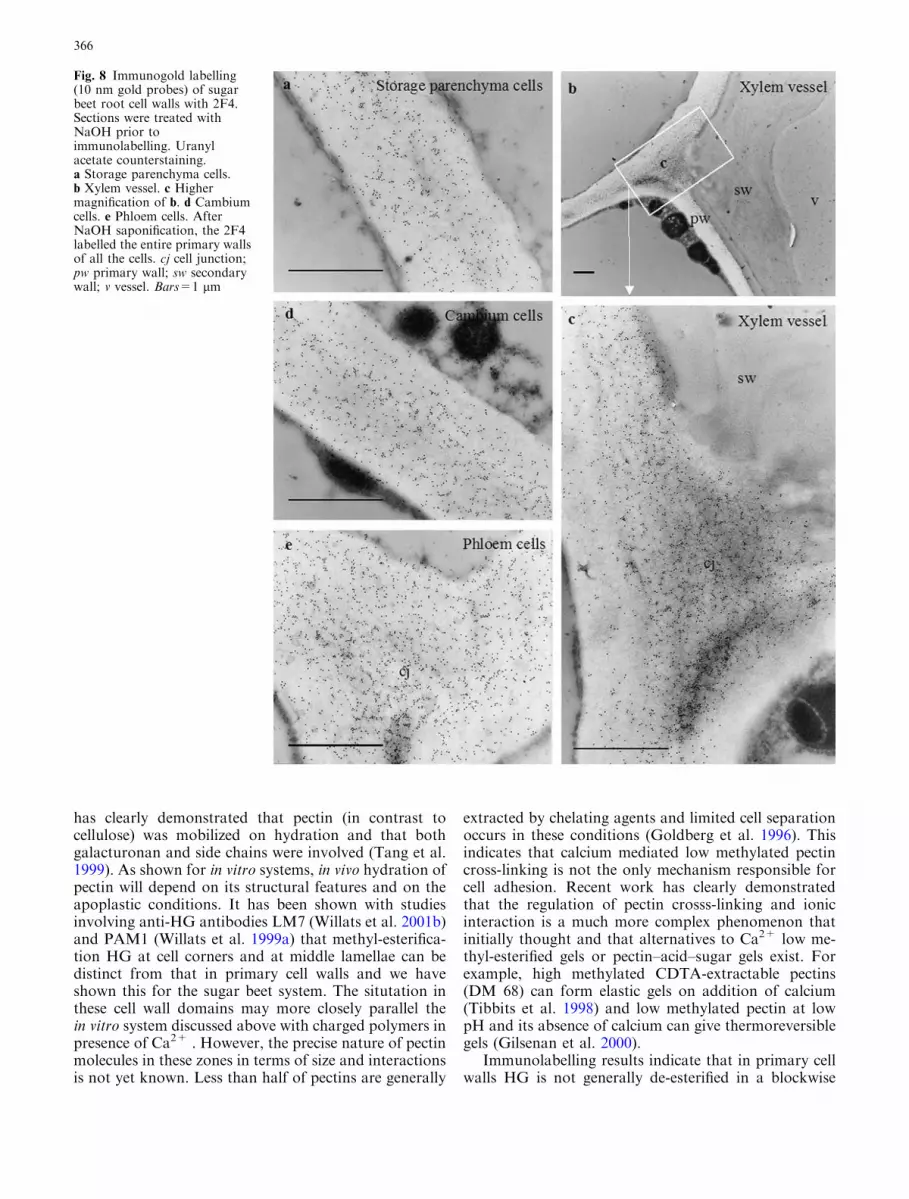
vessel cells (Fig. 4b, d). In vessel adjacent cells, some-times, the middle lamella was lignified and only pri-mary walls were labelled (Fig. 4c). In storageparenchyma cells, labelling with JIM5 was very faintand restricted to cell junctions and lining of intercel-lular spaces (Fig. 5c). In cambial cells, no significantlabelling was detected (Fig. 5a). In phloem, some cellswere not labelled. When present, the gold particles wereirregularly distributed and formed dense patches at celljunctions (Fig. 5d). On the xylem side, labelling withJIM5 was observed at cell junctions of cambial deriv-atives (Fig. 5b). After secondary wall formation, theJIM5 epitope was restricted to inner regions of the cellwall between secondary thickenings (Fig. 5e, f).Na2CO3 treated sections showed an increased labellingwith JIM5 (Fig. 6) with a labelling pattern ressemblingthat obtained with JIM7. Cell walls were not labelledwith 2F4 in absence of de-methyl-esterification treat-ments (not shown). After oPME treatment, labellingwas detected at cell junctions (Fig. 7) and at the liningof intercellular spaces. Phloem cell junctions (Fig. 7e)were labelled abundantly and those in other cells werelabelled more moderately (Fig. 7a, b, c, d). AfterNa2CO3 saponification, the labelling pattern was thesame as the one observed after PMEo treatment butthe density of gold particles was less (not shown),whereas, treatment with NaOH led to sections beingheavily labelled (Fig. 8). In parenchyma, phloem andcambium cells, the gold particles were uniformly dis-tributed throughout the wall (Fig. 8a, e, d). In xylemcells, the labelling was uneven and most abundant inprimary walls at cell junctions (Fig. 8b, c).
Table 2 Sugar composition and degrees of esterification of sugar beet pectins
Conditions of pectin isolation ortreatment applied to isolated pectins
Rhaa Ara Gal Gal A DMb DAc
A pH 1, 75 �C, 30 min, HCl 2.8 29.4 6.8 29.5 34 37C pH 1, 95 �C, 30 min, HCl 4.1 3.1 8.5 45.5 65 28E pH1, 75 �C, 90 min, HCl 3.5 12.7 8.0 40.2 68 28G pH 1, 95 �C, 90 min, HCL 4.3 0.6 6.5 46.0 55 15I pH 1, 75 �C, 30 min, HNO3 3.0 27.3 6.8 38.0 54 28M pH 1, 75 �C, 90 min, HNO3 3.0 3.2 8.1 46.0 63 19SP1 pH 1, 85 �C, 60 min, HCl 5.4 1.3 9.5 34.7 65 10SP3 pH 2, 75 �C, 60 min, HCl 1.6 26.5 3.2 40.7 51 43SP4 pH 2, 95 �C, 60 min, HCl 2.1 36.5 4.4 39.2 51 32SP5 pH 2, 85 �C, 30 min, HCl 1.4 26.8 3.2 30.4 77 43SP6 pH 2, 85 �C, 90 min, HCl 1.9 35.7 4.0 37.7 46 28P. oPME C treated with 10 PME 4.5 1.4 7.7 48.9 34 26P. fPME C treated with 18 PME 4.6 1.4 7.5 49.7 29 25P. PAE C treated with PAE 4.5 1.4 7.9 46.9 49 16P. oPME+PAE C sequentially treated with oPME and PAE 4.8 1.6 8.4 52.0 27 17P. PAE+oPME C sequentially treated with PAE and oPME 4.1 1.3 8.0 50.9 19 19P. NaOH C treated with NaOH, pH 12 4.2 1.3 7.4 49.1 16 17P. Na2CO3 C treated with Na2CO3, pH 12 4.5 1.2 7.5 52.2 14 16
Data on samples A to SP6 courtesy of Levigne et al. (2002a)oPME, Pectin methyl esterase from orange peel; fPME, Pectin methyl esterase from Aspergillus aculeatus; PAE, Pectin acetyl-esterasefrom Aspergillus aculeatusaExpressed as weight percentage of the pectin fractionbMoles of methyl groups/100 moles of anhydrogalacturonic acid residuescMoles of acetyl groups/100 moles of anhydrogalacturonic acid residues
360

Distribution of arabinan and galactan
With anti-galactan LM5, strong labelling was found inthe cambial zone with a decreasing gradient of epitopeabundance on both sides of the cambial layer (Fig. 9a).Cell walls of storage parenchyma (Fig. 9b) and phloemand xylem zones (Fig. 9a) exhibited a low-level andirregular immunofluorescence. No staining was detectedin xylem parenchyma cells close to vessels.
The pattern of LM5 labelling seen by light micros-copy was confirmed at the ultrastructural level (Fig. 10).Immunogold label was absent from medial zone and thecell junctions. The labelling density was high in the wallsof cambium (Fig. 10a, b) and phloem (Fig. 10c, d), lowand non-uniform in the walls of vessel cells and of vas-cular parenchyma (Fig. 10f). For example, dense pat-ches of gold particles were seen in the primary wallsunderlying secondary thickenings (Fig. 10f). In storage
Fig. 2 Immunofluorescencelabelling with anti HGantibodies of LRW-embeddedsugar beet root bundle (a, c, e,g, i) and storage parenchyma (b,d, f, h, j) cell walls. a and b withJIM7. All the cell walls werelabelled. c and d with JIM5. Theepitope was not evenlydistributed throughout bundleand storage parenchyma. e andf with JIM5, sections weretreated with Na2CO3 prior toimmunolabelling. All the cellwalls were labelled. g and h with2F4, sections were treated withoPME prior toimmunolabelling. Labelling wasmainly found at cell junctions.i and j with 2F4, sections weretreated with NaOH prior toimmunolabelling. All the cellwalls were labelled. cbcambium; mr medullar ray; phphloem; v vessel. Bars=100 lm
361
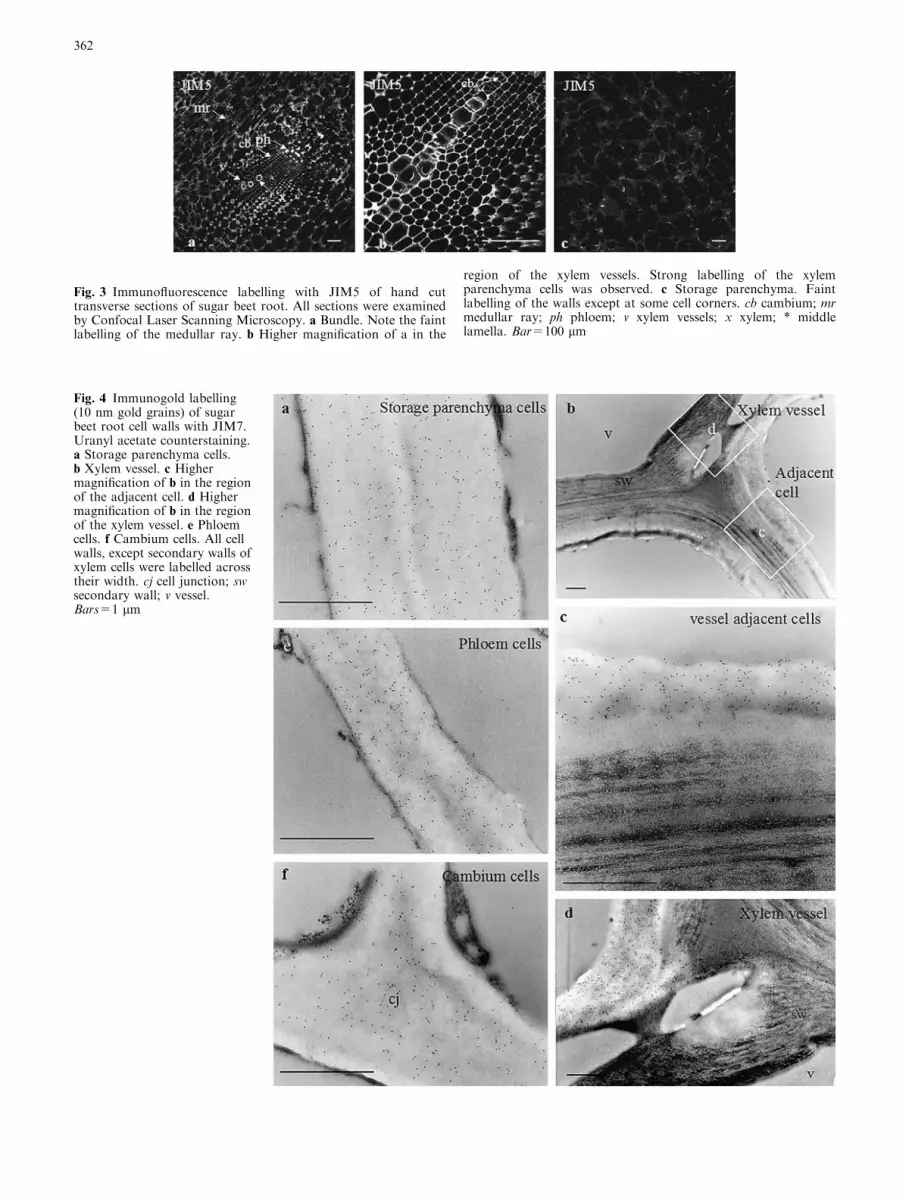
Fig. 4 Immunogold labelling(10 nm gold grains) of sugarbeet root cell walls with JIM7.Uranyl acetate counterstaining.a Storage parenchyma cells.b Xylem vessel. c Highermagnification of b in the regionof the adjacent cell. d Highermagnification of b in the regionof the xylem vessel. e Phloemcells. f Cambium cells. All cellwalls, except secondary walls ofxylem cells were labelled acrosstheir width. cj cell junction; swsecondary wall; v vessel.Bars=1 lm
Fig. 3 Immunofluorescence labelling with JIM5 of hand cuttransverse sections of sugar beet root. All sections were examinedby Confocal Laser Scanning Microscopy. a Bundle. Note the faintlabelling of the medullar ray. b Higher magnification of a in the
region of the xylem vessels. Strong labelling of the xylemparenchyma cells was observed. c Storage parenchyma. Faintlabelling of the walls except at some cell corners. cb cambium; mrmedullar ray; ph phloem; v xylem vessels; x xylem; * middlelamella. Bar=100 lm
362

parenchyma, gold label was mainly localized to a zoneclose to the plasma membrane (Fig. 10e).
All cell walls were labelled by anti-arabinan LM6with the cell walls of bundles labelled more abundantlythan medullar ray and storage parenchyma cells(Fig. 9c, d). The LM6 epitope appeared to occur evenlyin each cell type and this was confirmed by the analysisat the ultrastructural level, which indicated that the LM6epitope occurred throughout primary walls with medianregions (including middle lamella) and cell junctionsbeing lightly labelled or unlabelled. (Fig. 11).
Discussion
In sugar beet root cell walls, the JIM7 HG epitope isabundant while the JIM5 HG epitope is restricted tosome cells and cell wall domains. This observation does
not fit with the immuno-dot assays that indicate that theJIM5 epitope is more abundant than the JIM7 epitope inacid extracted sugar beet pectins. We clearly demon-strate that DA at least in the range 10–30 does notinfluence JIM7 and JIM5 binding to sugar-beet pectins.Possibly, in acid extracted sugar beet pectins, the patternof distribution of methyl groups along the backbone ismore favourable for JIM5 binding. The apparent dis-crepancy between immuno-dot and immunolabellingresults with JIM5 could arise from the fact that either,the pectic acid extraction modifies to some extent thedegree and pattern of polymer esterification, or that aciddoes not extract all the pectins and those extracted differfrom those remaining.
The restricted occurence of the JIM5 epitope hasbeen already described in other plant systems (see reviewWillats et al. 2001a, b). Its distribution pattern in sugarbeet root bundles fits quite well the one described by
Fig. 5 Immunogold labelling(10 nm gold grains) of sugarbeet root cell walls with JIM5.Uranyl acetate counterstaining.a Cambium cells. b Parenchymacells close to vessel. c Storageparenchyma cells. d Phloemcells. e Xylem vessel. f Highermagnification of e. Not all thecell walls were labelled withJIM5 and the labelling wasrestricted to some regions: celljunction and lining ofintercellular spaces in storageparenchyma and phloem cells,inner region between secondarythickenings in xylem vessel. cjcell junction; is intercellularspace; pw primary wall; swsecondary wall; v vessel.Bars=1 lm
363

Guglielmino et al. (1997) in poplar cambium, whereJIM5 epitope occurrence varies according to the celldifferenciation either on the xylem or the phloem path-way.
Immuno-dot assays and immunocytochemical inves-tigations with 2F4 confirm that extensive de-methy-lesterification and de-acetylesterification or blockwisedistribution of GalA units are required for 2F4 binding(Liners et al. 1992). In plant tissues, the 2F4 confor-mational epitope of HG is much less widespread that theJIM7 or JIM5 epitopes. In sugar beet root, it is notdetected in any of the cell types. In potato tubers (Bushand Mc Cann 1999) only cortex cells contain the 2F4epitope. Labelling is restricted to cell junctions, middlelamella close to the junction zones, and the lining ofintercellular spaces. In suspension-cultured carrot cells,a similar pattern of the 2F4 distribution is observed
(Liners and Van Cutsem 1992). In sugar beet compactcalli, the epitope is present in fibrillar material expandedin areas of cell separation, while it is not detected infriable callus. The cells corners have been also demon-strated to be rich in Ca2+ (Guglielmino et al. 1997). HGin these zones is thought to contribute to the resistanceto the stress that tends to separate cells through formingstrong gel via calcium bridges (Jarvis et al. 2003). Thefact that this epitope is not detected in non-treated sec-tions suggests that other mechanisms must be involvedin the resistance to high cell separation stresses at tri-cellular junctions and the corners of intercellular spacesinduced by the high turgor pressure generated by sucrosecontent in storage parenchyma.
A feature of sugar beet pectins is the presence of highamounts of acetyl groups on HG. The results reportedhere demonstrate that HGs were less acetylated at cell
Fig. 6 Immunogold labelling(10 gold grains) of sugar beetroot cell walls with JIM5.Sections were treated withNa2CO3 prior toimmunolabelling. Uranylacetate counterstaining.a Storage parenchyma cells.b Xylem vessel. c Highermagnification of b in the regionbetween secondary thickenings.d Cambium cells. e Phloemcells. Note that afterde-esterification treatment withNa2CO3, JIM5 labelled all theprimary cell walls throughouttheir width. pw primary wall; swsecondary wall; v vessel.Bars=1 lm
364
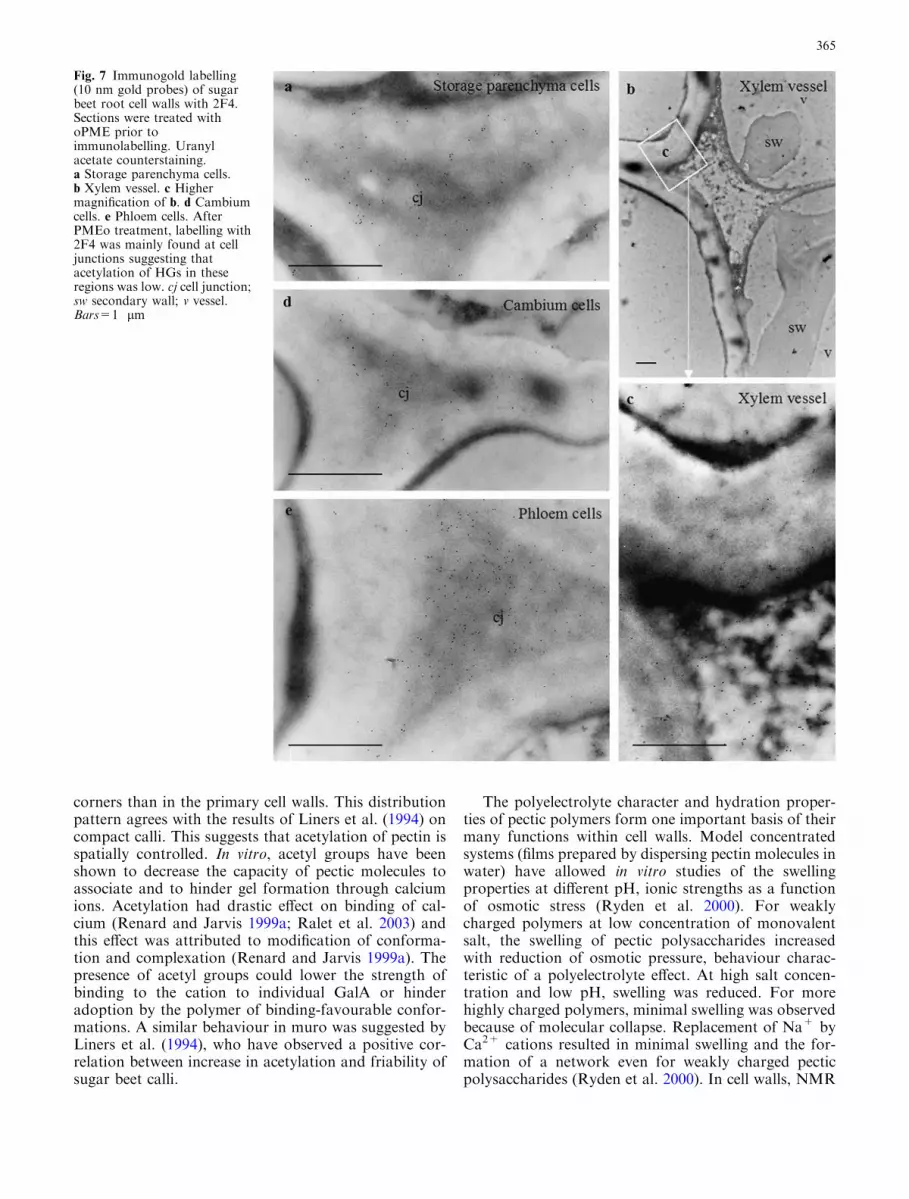
corners than in the primary cell walls. This distributionpattern agrees with the results of Liners et al. (1994) oncompact calli. This suggests that acetylation of pectin isspatially controlled. In vitro, acetyl groups have beenshown to decrease the capacity of pectic molecules toassociate and to hinder gel formation through calciumions. Acetylation had drastic effect on binding of cal-cium (Renard and Jarvis 1999a; Ralet et al. 2003) andthis effect was attributed to modification of conforma-tion and complexation (Renard and Jarvis 1999a). Thepresence of acetyl groups could lower the strength ofbinding to the cation to individual GalA or hinderadoption by the polymer of binding-favourable confor-mations. A similar behaviour in muro was suggested byLiners et al. (1994), who have observed a positive cor-relation between increase in acetylation and friability ofsugar beet calli.
The polyelectrolyte character and hydration proper-ties of pectic polymers form one important basis of theirmany functions within cell walls. Model concentratedsystems (films prepared by dispersing pectin molecules inwater) have allowed in vitro studies of the swellingproperties at different pH, ionic strengths as a functionof osmotic stress (Ryden et al. 2000). For weaklycharged polymers at low concentration of monovalentsalt, the swelling of pectic polysaccharides increasedwith reduction of osmotic pressure, behaviour charac-teristic of a polyelectrolyte effect. At high salt concen-tration and low pH, swelling was reduced. For morehighly charged polymers, minimal swelling was observedbecause of molecular collapse. Replacement of Na+ byCa2+ cations resulted in minimal swelling and the for-mation of a network even for weakly charged pecticpolysaccharides (Ryden et al. 2000). In cell walls, NMR
Fig. 7 Immunogold labelling(10 nm gold probes) of sugarbeet root cell walls with 2F4.Sections were treated withoPME prior toimmunolabelling. Uranylacetate counterstaining.a Storage parenchyma cells.b Xylem vessel. c Highermagnification of b. d Cambiumcells. e Phloem cells. AfterPMEo treatment, labelling with2F4 was mainly found at celljunctions suggesting thatacetylation of HGs in theseregions was low. cj cell junction;sw secondary wall; v vessel.Bars=1 lm
365
has clearly demonstrated that pectin (in contrast tocellulose) was mobilized on hydration and that bothgalacturonan and side chains were involved (Tang et al.1999). As shown for in vitro systems, in vivo hydration ofpectin will depend on its structural features and on theapoplastic conditions. It has been shown with studiesinvolving anti-HG antibodies LM7 (Willats et al. 2001b)and PAM1 (Willats et al. 1999a) that methyl-esterifica-tion HG at cell corners and at middle lamellae can bedistinct from that in primary cell walls and we haveshown this for the sugar beet system. The situtation inthese cell wall domains may more closely parallel thein vitro system discussed above with charged polymers inpresence of Ca2+ . However, the precise nature of pectinmolecules in these zones in terms of size and interactionsis not yet known. Less than half of pectins are generally
extracted by chelating agents and limited cell separationoccurs in these conditions (Goldberg et al. 1996). Thisindicates that calcium mediated low methylated pectincross-linking is not the only mechanism responsible forcell adhesion. Recent work has clearly demonstratedthat the regulation of pectin crosss-linking and ionicinteraction is a much more complex phenomenon thatinitially thought and that alternatives to Ca2+ low me-thyl-esterified gels or pectin–acid–sugar gels exist. Forexample, high methylated CDTA-extractable pectins(DM 68) can form elastic gels on addition of calcium(Tibbits et al. 1998) and low methylated pectin at lowpH and its absence of calcium can give thermoreversiblegels (Gilsenan et al. 2000).
Immunolabelling results indicate that in primary cellwalls HG is not generally de-esterified in a blockwise
Fig. 8 Immunogold labelling(10 nm gold probes) of sugarbeet root cell walls with 2F4.Sections were treated withNaOH prior toimmunolabelling. Uranylacetate counterstaining.a Storage parenchyma cells.b Xylem vessel. c Highermagnification of b. d Cambiumcells. e Phloem cells. AfterNaOH saponification, the 2F4labelled the entire primary wallsof all the cells. cj cell junction;pw primary wall; sw secondarywall; v vessel. Bars=1 lm
366

manner, but the degree and pattern of methyl-esterifi-cation is still not precisely known. However, in case ofsugar beet pectin, the fact that acetyl groups are presenton HG throughtout the primary wall would suggest thatgelation, whatever the mechanisms, may be reduced andthat strong gels are not formed. In this situation, it ispossible that the swelling behaviour of HG influencesprimary cell wall properties and that side chains play arole in the establisment of a pectic network.
Immuno-dot results with LM5 and LM6 are inagreement with chemical data according to which sugarbeet pectins contain both galactan and arabinan sidechains (Guillon and Thibault 1989; Guillon et al. 1989;Levigne et al. 2002a). However, the extent of LM5 andLM6 binding to extracted pectins does not increase withtheir arabinose and galactose content; this suggests theoccurence of branching or of substitution of the sidechains (Guillon and Thibault 1989; Guillon et al. 1989;Oosterveld et al. 2000) that prevent increased binding.From the immuno-dot assays on isolated pectins, anequivalent occurrence of the LM5 and LM6 epitopes incell walls was expected. However, as already reported byother authors, the distribution of galactan and arabinanwas different according to cell wall domains and cell andtissue types. Immuno-dot assays and physico-chemicalanalyses provide average information on populations ofpectins and are poorly sensitive to the effect of pectinheterogeneity in muro. Most of the extractions andseparations of pectin populations are based on the pecticbackbone properties and little information is availableon the pectin heterogeneity with respect to side chains.
Galactan was mainly detected in the cambial zoneand partially lost during tissue differentiation. Similarresults have been reported on active cambial derivativesof aspen (Ermel et al. 2000). Disappearance could beexplained either by a decrease of the epitope accessibilitydue to deposition of new material in the cell wall, or by aremodelling of the galactan side chains through addition
of some decorations on the main chain, or by in muroenzymatic degradation of the galactan. Arabinan ismore widespread amonge tissues than galactan, butagain differences in the labelling intensity could be seenbetween cells that may reflect differences in the degree ofbranching.
Our results are in agreement with the previous dataon other organs showing that 1,5-arabinan and 1,4-ga-lactan are developmentally regulated. The distributionof branched pectins within cell walls and in particulartheir absence in middle lamellae or abundance in par-ticular wall domains could reflect both developmentalregulation and molecular interactions. In vitro, it hasbeen shown that in the absence of calcium ions, a con-centrated binary mixture of esterified, relatively un-branched pectic polysaccharides and highly branchedde-esterified pectic polysacccharides, both extractedfrom unripe tomato cell walls exhibit immiscibility,resolving into separate phases, each of which is enrichedin one polymer (MacDougall et al. 1997). The gelationof pectic polysaccharides or the restricted mobility of thedifferent polymers under high concentrated conditionsof the cell walls could inhibit or hinder phase separation.MacDougall et al. (1997) suggest that the phase sepa-ration phenomenom could occur during the formationof the middle lamella or during the period before enzy-matic de-esterification when pectins are not capable offorming ionic-cross links with calcium. The confinementof neutral sugar rich pectic polymers to some cell walldomains (e.g. absent from middlle lamellae and celljunctions) may be of importance in the modulation ofcell wall properties. Side chains are thought to contrib-ute to the functional properties of pectins, involvinginteractions between molecules, especially gel formationand rheological phenomena, but their contribution orrole is far from clear. For example, contradictoryarguments have been issued for their role on gelling.According to Selvendran (1985), arabinose and galac-
Fig. 9 Immunofluorescencelabelling of LRW-embeddedsugar beet root bundle(a, c) and storage parenchyma(b, d) cell walls. a and b withLM5. c and d with LM6 . Notethe high labelling of thecambium walls with LM5. LM6labelled all the cell walls. cbcambium; ph phloem; xp xylemparenchyma close to vessels;v vessel; * non-labelled area.Bars=100 lm
367

tose side chains may contribute to gelling by holdingwater molecules within a gel framework. In contrast,BeMiller (1986) argued that side chains might limit theextent of interchain associations. This was consistentwith the results of Berth and Dahme (1991), who re-ported that increasing the degree of branching reducedthe strength of the pectin gel. This could be particularlytrue for high methylated pectins, which require inter-molecular associations by hydrogen bonding for gela-tion. It has been also suggested that side chains throughtheir involvement in entanglements of pectins in con-centrated solution could contribute to elastic propertiesof the cell walls (Hwang and Kokini 1992) and recentwork on guard cells indicate that arabinans are requiredto maintain cell wall flexibility (Jones et al. 2003). Insugar beet cell walls, arabinan has been shown to behighly mobile (Renard and Jarvis 1999b). The authors
suggested that rather than participating in cross-links,these side chains, by filling the pore of the cell wallnetwork may act as plasticizers or water binding agents.
However, it cannot be ruled out that arabinan insugar beet walls can participate in the pectin cross-linkvia oxidative coupling of ferulic acid esterifying thearabinose and thus, playing a direct role in the control ofcell wall extensibility and mechanical properties. Feru-loyl groups are mainly esterified to O-2 of arabinoseresidues of the main core of a (1–5)-a-linked arabinanside chains and to O-6 of galactose residues of the maincore of (1–4)-b-linked type I galactan chains (Colqu-houn et al. 1994; Clausen et al. 2004). Recently minoramounts of feruloyl groups were assumed to be esterlinked to O-5 of arabinose, indicating peripheral loca-tion of feruloyl groups on pectic hairy regions (Levigneet al 2004a). In beet root, over 10% of the ferulic acid
Fig. 10 Immunogold labelling(1 nm gold probes, silverenhancement) of sugar beetroot cell walls with LM5.Uranyl acetate counterstaining.a Cambium cells, radial walls.b Cambium cells, tangentialwalls. c Phloem cells, celljunction. d Phloem cell wall.e Storage parenchyma cells.f Xylem vessels. Labellingdensity was high in thecambium and in phloem cellwalls. In vessel cells, the epitopewas detected in the primarywalls underlying the secondarythickenings. No labelling wasobserved at cell junctions, andin storage parenchyma, it wasrestricted to wall regions closeto plasmalemma. cj celljunction; pw primary wall; swsecondary wall; v vessel.Bars=1 lm
368

was found in dimer form with the 8-O-4¢ and 8,5¢ di-ferulate benzofuran forms being the most prominant(Waldron et al. 1997). The evidence that pectic arabin-ans in sugar beet cell walls are covalently cross-linkedthrough diferulate was recently demonstrated throughthe isolation a dehydrodiferuloylated oligoarabinan(Levigne et al. 2004b). In planta, autofluorescence(Waldron et al. 1997) and immunofluorescence (mono-clonal antibody against feruloylated (1–4)-b-D-galactan,Clausen et al. 2004) studies suggest that esterified ferulicacid is an ubiquitous feature of sugar beet pectins. Theferulic acid dimers are thought to be involved in cell tocell adhesion (Waldron et al. 1997). However, the factwe did not find RGI side chains at tricellular junctionsuggests that cross-linking via ferulic acid dimers is un-likely in these wall domains.
No information on the location, distribution anddynamics of ferulic acid dimers is avalaible, which willbe crucial to further understand in vivo the role of ferulicacid in the control of the cell to cell adhesion and cellwall mechanical properties.
Acknowledgements The authors would like to thank Dr S Levigneand Dr M-C Ralet, both from Unite de Recherche sur les Polysac-charides, leurs Organisations et Interactions, Institut National de laRecherche Agronomique, Nantes, France, for they kind gifts ofseries of sugar beet pectins. The authors thank Prof. P. Van Cutsemfrom Unite de Recherche en Biologie Cellulaire, Facultes Universi-taires Notre Dame de la Paix, Namur, Belgium, for the generous giftof 2F4 monoclonal antibody. They are grateful to C. Nicolas,Institut National de la Recherche Agronomique, Nantes, France forphotographic assistance. F. Guillemin gratefully acknowledges theInstitut National de la Recherche Agronomique and the RegionPays de Loire for their support through a doctoral fellowship.
Fig. 11 Immunogold labelling(1 nm gold probes, silverenhancement) with LM6.Uranyl acetate counterstaining.a Cambium cells, cell junction.b Cambium cells, radial walls.c Phloem cells, cell junction.d Phloem cell walls. e Storageparenchyma cells. f Xylemvessel. All the primary cell wallswere labelled with LM6. Notethat the labelling was lighter orabsent at cell junctions andmiddle lamella. cj cell junction;is intercellular space; pwprimary wall; sw secondarywall; v vessel; *middle lamella.Bars=1 lm
369

References
BeMiller JN (1986) An introduction to pectins: structure andproperties. In: Fishman ML, Jen JJ (eds) Chemistry and func-tion of pectins. American Chemical Society, Washington DC,pp 2–12
Berth G, Dahme A (1991) Macromolecular structural features offruit pectins and their relation to gel strength. Food Hydrocoll5:101–104
Bonnin E, Dolo E, Le Goff A, Thibault J-F (2002) Characterisationof pectin subunits released by an optimised combinaison ofenzymes. Carbohydr Res 337:1687–1696
Bonnin E, Le Goff A, van Alebeek GJWM, Voragen AGJ, Thi-bault J-F (2003) Mode of action of Fusarium monoliformeendopolygalacturonase towards acetylated pectin. CarbohydrPolym 52:381–388
Bonnin E, Daniel S, Clavurier K, Mikkelsen JD, Thibault J-F(2005) A new enzyme from from Aspergillus aculeatus to de-acetylate pectins. Biochim Biophys Acta (submitted)
Buchholt HC, Christensen TMIE, Fallesen B, Ralet M-C, ThibaultJ-F (2004) Preparation and properties of enzymatically andchemically modified sugar beet pectins. Carbohydr Polym58:149–161
Bush MS, McCann MC (1999) Pectic epitopes are differentiallydistributed in the cell walls of potato (Solanum tuberosum) tu-bers. Physiol Plant 107:201–213
Clausen MH, Willats WGT, Knox JP (2003) Synthetic methylhexagalacturonate hapten inhibitors of anti homogalacturonanmonoclonal antibodies LM7, JIM5 and JIM7. Carbohydr Res338:1797–1800
Clausen MH, Ralet M-C, Willats WGT, McCartney L, Marcus SE,Thibault J-F, Knox JP (2004) A monoclonal antibody to fer-uoylated (1–4)-b-D-galactan. Planta 210:1036–1041
Colquhoun IJ, Ralet M-C, Thibault J-F, Faulds CB, Williamson G(1994) Structure and indentification of feruloylated oligosac-charides from sugar beet pulp by NMR spectroscopy. Carbo-hydr Res 263:243–256
Ermel FF, Follet-Gueye M-L, Cibert C, Vian B, Morvan C,Catesson AN, Goldberg R (2000) Differential localization ofarabinan and galactan side chains of rhamnogalacturonan 1 incambial derivatives. Planta 210:732–740
Gilsenan PM, Richardson RK, Morris ER (2000) Thermallyreversible acid induced gelation of low methoxy pectin. Car-bohydr Polym 41:339–349
Goldberg R, Morvan C, Jauneau A, Jarvis MC (1996) Methy-lesterification, de-esterification and gelation of pectins in theprimary cell wall. In: Visser J, Voragen AGJ (eds) Pectins andpectinases. Elservier, Amsterdam, pp 151–172
Guglielmino N, Liberman M, Jauneau A, Vian B, Catesson A-M,Goldberg R (1997) Pectin immunolocalization and calciumvisualization in differentiating derivatives from poplar cam-bium. Protoplasma 199:151–160
Guillon F, Thibault J-F (1989) Methylation analysis and mild acidhydrolysis of the ‘‘hairy’’ fragments of sugar-beet pectins.Carbohydr Res 190:85–96
Guillon F, Thibault J-F, Rombout FM, Voragen AGJ, Pilnik W(1989) Enzymatic hydrolysis of the hairy fragments of sugarbeet pectins. Carbohydr Res 190:97–108
Hwang J, Kokini JL (1992) Contribution of the side branches torheological properties of pectins. Carbohydr Polym 19:41–50
Ishii T (1995) Isolation and characterization of acetylatedrhamnogalacturonan oligomers liberated from bamboo shootcell-walls by Driselase. Mokuzai Gakkaishi 41:561–572
Ishii T (1997) O-acetylated oligosaccharides from pectins of potatotuber cell walls. Plant Physiol 116:337–347
Jarvis MC, Briggs SPH, Knox JP (2003) Intercellular adhesion andcell separation in plants. Plant Cell Environ 26:977–989
Jones L, Seymour GB, Knox JP (1997) Localization of pectic ga-lactan tomato cell walls using a monoclonal antibody specific to(1–4)-b-D-galactan. Plant Physiol 113:1405–1412
Jones L, Milne JL, Ashford D, McQueen-Mason SJ (2003) Cellwall arabinan is essential for guard cell function. Proc NatlAcad Sci USA 100:11783–11788
Le Goff A, Renar CMGC, Bonnin E, Thibault J-F (2001)Extraction, purification and chemical characterisation ofxylogalacturonans from pea hulls. Carbohydr Polym 45:325–334
Levigne S, Ralet M-C, Thibault J-F (2002a) Characterization ofpectins extracted from fresh sugar beet under differentconditions using experimental design. Carbohydr Polym49:145–53
Levigne S, Thomas M, Ralet M-C, Quemener B, Thibault J-F(2002b) Determination of the degrees of methylation andacetylation of pectins using C18 column internal standard.Food Hydrocolloid 16:547 – 550
Levigne SV, Ralet M-CJ, Quemener BC, Pollet BM-L, Lapierre C,Thibault J-F (2004a) Isolation from sugar beet cell walls ofarabinan oligosaccharides esterified by two ferulic acid mono-mers. Plant Physiol 134:1173–1180
Levigne S, Ralet M-C, Quemener B, Thilbault J-F (2004b) Isola-tion of diferulic bridges ester-linked to arabinan in sugar beetwell walls. Carbohydr Res 339:2313–2319
Liners F, Letesson JJ, Didembourg C, Van Cutsem P (1989)Monoclonal antibodies against pectin. Recognition of a con-formation induced by calcium. Plant Physiol 94:1419–1424
Liners F, Van Cutsem P (1992) Distribution of pectic polysaccha-rides throughout walls of suspension-cultured carrot cells. Animmunocytochemical study. Protoplasma 170:10–21
Liners F, Thibault J-F, Van Cutsem P (1992) Influence of the de-gree of polymerization of oligogalacturonates and of esterifi-cation pattern of pectin on their recognition by monoclonalantibodies. Plant Physiol 99:1099–1104
Liners F, Gaspar T, Van Cutsem P (1994) Acetyl- and methylesterification of pectins of friable and compact sugar-beet calli:consequences for intercellular adhesion. Planta 192:545–556
MacDougall AJ, Rigby NM, Ring S (1997) Phase separation ofplant cell wall polysaccharides and its implication for cell wallassembly. Plant Pysiol 114:353–362
Matoh T, Takasaki M, Takabe K, Kobayashi M (1998). Immu-nocytochemistry of rhamnogalacturonan II in cell walls ofhigher plants. Plant Cell Physiol 39:483–491
McCartney L, Ormerod AP, Gidley MJ, Knox JP (2000) Temporaland spatial regulation of pectic (1 fi 4)-b-D-galactan in cellwalls of developping pea cotyledons: implications for mechan-ical properties. Plant J 22:105–113
Oosterveld A, Beldman G, schols HA, Voragen AGJ (2000)Characterization of arabinose and ferulic acid rich pecticpolysaccharides and hemicelluloses from sugar beet pulp. Car-bohydr Res 328:185–197
Quemener B, Cabrera Pino JC, Ralet MC, Bonnin E, Thibault JF(2003) Assignment of acetyl groups to O-2 and/or O-3 of pecticoligogalacturonides using negative Electrospray-Ion Trap MassSpectrometry. J Mass Spectrom 38:641–658
Ralet M-C, Crepeau M-J, Buchholt H-C, Thibault J-F (2003)Polyelectrolyte behaviour and calcium binding properties ofsugar beet pectin differing in their degrees of methylation andacetylation. Biochem Eng J 16:191–201
Renard CMGC, Thibault JF, Liners F, Van Cutsem P (1993)Immunological probing of pectins isolated or in situ. ActaBotanica Neerlandica 42:199–204
Renard CMGC, Jarvis MC (1999a) Acetylation and methylation ofhomogalacturonans 2: effect on ion-binding properties andconformation. Carbohydr Polym 39:209–216
Renard CMGC, Jarvis MC (1999b) A cross-polarization, magic-angle-spinning, 13C-nuclear-magnetic-resonance study ofpolysacchrides in sugar beet cell walls. Plant Physiol 119:1315–1322
Rombout FM, Thibault J-F (1986) Feruloylated pectic substancesfrom sugar beet pulp. Carbohydr Res 154:177–188
Ryden P, MacDougall, Tibbits CW, Ring SG (2000) Hydration ofpectic polysaccharides. Biopolymers 54:398–405
370

Schols HA, Voragen AGJ (1994) Occurence of pectic hairy regionin various plant cell wall materials and their degradability byrhamnogalacturonase. Carbohydr Res 256:83–96
Selvendran RR (1985) Developments in the chemistry and bio-chemistry of pectic and hemicelluloic polymers. J Cell Sci Suppl2:51–58
Tang H, Belton PD, Ng A, Ryden P (1999) 13C MAS NMRstudies of the effects of hydration on the cell walls of potatoesand chines water chestnuts. J Agric Food Chem 47:510–517
Thibault J-F (1979) Automatisation du dosage des substancespectiques par la methode au metahydroxydiphenyle. LebensmWissenschaft Technol 12:247–251
Tibbits CW, MacDougall AJ, Ring SG (1998) Calcium binding andswelling behaviour of a high methoxyl pectin gel. CarbohydrRes 310:101–107
Waldron KW, Ng A, Parker ML, Parr A (1997) Ferulic acid de-hydromers in the cell walls of beta vulagris and their possiblerole in texture. J Sci Food Agric 74:221–228
Willats WGT, Marcus SE, Knox JP (1998) Generation of mono-clonal antibody specific to (1–5)-a-L-arabinan. Carbohydr Res308:149–152
Willats WGT, Gilmartin PM, Mikkelsen JD, Knox JP (1999a) Cellwall antibodies without immunization: generation and use ofdesesterified homogalacturonan block-specific antibodies froma naive phage display library. Plant J 18:57–65
Willats WGT, Steele-King CG, Marcus SE, Knox JP (1999b) Sidechains of pectic polysaccharides are regulated in relation to cellproliferation and cell differentiation. Plant J 20:619–628
Willats WGT, Limberg G, Bucholt HC, van Alebeck GJ, Benen J,Christensen TMIE, Visser J, Voragen A, Mikkelsen JD, KnoxJP (2000) Analysis of pectic epitope recognized by hybridomaand phage display monoclonal antibodies using defined oligo-saccharides, polysaccharides, and enzymatic degradation. Car-bohydr Res 327:309–320
Willats WGT, McCartney L, Mackie W, Knox JP (2001a) Pectin:cell biology and prospects for functional analysis. Plant MolBiol 47:9–27
Willats WGT, Orfila C, Limberg G, Bucholt HC, van Alebeck GJ,Voragen A, Marcus SE, Christensen TMIE, Mikkelsen JD,Murray BS, Knox JP (2001b) Modulation of the degree andpattern of methyl-esterification of pectic homogalacturonan inplant cell walls. J Biol Chem 276:19404–19413
371





























