Embed Size (px)
Citation preview

Molecular Ecology (2004)
13
, 2393–2404 doi: 10.1111/j.1365-294X.2004.02223.x
© 2004 Blackwell Publishing Ltd
Blackwell Publishing, Ltd.
Differences in mycorrhizal preferences between two tropical orchids
J . TUPAC OTERO,
*
JAMES D. ACKERMAN and PAUL BAYMAN
Departamento de Biología, Universidad de Puerto Rico — Río Piedras, PO Box 23360, San Juan, Puerto Rico 00931–3360
Abstract
Orchids parasitize their mycorrhizal fungi and are dependent on them for seed germination.Controversy reigns over how specific the mycorrhizal association is in tropical species.Although there is little experimental evidence to support any viewpoint, some variation isknown to exist. We compared mycorrhizal specificity and performance in two phylogeneticallyrelated epiphytic orchids from Puerto Rico,
Tolumnia variegata
and
Ionopsis utricularioides
(Oncidiinae) by integrating two techniques: phylogenetic analysis of mycorrhizal fungibased on nuclear ribosomal internal transcribed spacer (ITS) sequences, and symbioticseed germination experiments. Most of the mycorrhizal isolates from
T. variegata
fell intofour different clades of
Ceratobasidium
, while most of those from
I. utricularioides
wererestricted to a single clade of the same genus. Seeds of
T. variegata
germinated equally wellwith fungi from both
T. variegata
and
I. utricularioides
, but seeds of
I. utricularioides
ger-minated significantly better with its own isolates. Seeds of
I. utricularioides
germinatedand developed faster than those of
T. variegata
. Both the molecular phylogeny and the seedgermination experiments showed that
T. variegata
is a generalist in its association withfungal symbionts. In contrast,
I. utricularioides
is more specialized and more effective atexploiting a specific fungal clade. Our data are consistent with the theoretical trade-offsbetween specialized and generalized interactions.
Keywords
:
Ceratobasidium
,
Orchidaceae
, phylogeny,
Rhizoctonia
, specificity, symbiotic seed germination,trade-offs
Received 1 November 2003; revision received 18 March 2004; accepted 7 April 2004
Introduction
Specificity is unexpected in ecological interactions becausethose individuals that are able to interact with manypartners should have more choices than those that interactwith a narrow range of partners (Timms and Read 1999).Nevertheless, this prediction assumes that all potentialpartners are ecologically equivalent and homogeneouslydistributed. A species is specific for a given interactionif it interacts with a limited number of partner species(Thompson 1994). This definition is not precise enough, soseveral different methods are used to assess specificity.One method involves phylogenetic relationships among
possible partners for a given species. If all partners forma single clade, then one may argue that specificity existsregardless of how many species are involved (Johnsonand Steiner 2000; Taylor
et al
. 2002). Functional tests ofinteractions, such as reciprocal transfer experiments, mayindicate whether or not the cause of specificity is limiteddispersal or limited adaptation (Timms and Read 1999).Geographic distributions of interacting species are alsouseful because limited distributions may impose specificity(Ackerman 1983; Timms and Read 1999). Each of theseapproaches to specificity is best considered in a relativecontext. Here we apply all these approaches to an inter-action that has puzzled biologists for a century: orchid–fungus mycorrhizal relationships.
Much of the diversity in the orchid family has beenattributed to selection for pollination mechanisms that pro-mote cross-pollination (van der Pijl & Dodson 1966) and toa lesser degree selection for vegetative characteristics that
Correspondence: J. Tupac Otero. *Present address: CSIRO PlantIndustry, Australian National Herbarium, GPO Box 1600,Canberra ACT 2601, Australia; Fax: + 61-2-6246-5249; E-mail:[email protected]

2394
J . T . O T E R O , J . D . A C K E R M A N and P . B A Y M A N
© 2004 Blackwell Publishing Ltd,
Molecular Ecology
, 13, 2393–2404
enhance survival in the epiphytic habitat (Benzing 1986).Remarkable specializations in both pollinator relationshipsand vegetative features have long been a testament forthose hypotheses (Darwin 1877). Yet another possible factorthat may have accelerated or promoted diversification isspecialization in the orchid–mycorrhizal relationship.
The specificity of the interaction between orchids andtheir mycorrhizal fungi has long been controversial, withsome authors claiming that orchids are generalists (Knudson1922; Burgeff 1932; Curtis 1939; Hadley 1970; Masuhara &Katsuya 1989, 1991; Masuhara
et al
. 1993), but others arguingthat they are specialists (Bernard 1909; Clements 1987, 1988;Smreciu & Currah 1989; Taylor & Bruns 1997; McKendrick
et al
. 2002; Selosse
et al
. 2002). Arditti
et al
. (1990) speculatedthat terrestrial temperate orchids should be specialists intheir mycorrhizal relationships because they are difficultto germinate on nutritive media under aseptic conditions,whereas epiphytic, tropical orchids should be generalistsin their relationships with mycorrhizal fungi because theirseeds are easily germinated
in vitro
. Rasmussen (1995)suggested that temperate orchids are generalists in theirmycorrhizal interactions, based on the morphology of fungiisolated from different orchids, whereas Taylor & Bruns(1997) showed that temperate achlorophyllous orchids arespecialized on different families of ectomycorrhizal fungi.Still less is known about specificity in epiphytic orchids.The only empirical data available suggest that specificityoccurs in at least some species (Clements 1987, 1988).Nevertheless, a subtropical epiphytic orchid was able togerminate with a fungus isolated from another species(Zettler
et al
. 1999) and in a previous study we found thatthe specificity of orchid mycorrhizal fungi varies amongtropical epiphytic orchids (Otero
et al
. 2002). A single fungalclade (clade B) was isolated from several unrelated orchidspecies (Otero
et al
. 2002), but given the size of the family,we certainly expect a broad spectrum of specificities(Clements 1988).
To address the question of specificity in orchid–fungalrelationships it is necessary to understand the phylogeneticsof both groups. Orchid floras of many parts of the world arewell known (Dressler 1981) and their phylogenetic treat-ments are improving with the use of DNA sequence data(e.g. Freudenstein
et al
. 2004). In contrast, the systematicsof orchid mycorrhizal fungi are poorly understood (Otero
et al
. 2002). Most of the mycorrhizal fungi are classified inthe form-genus
Rhizoctonia
, a very diverse and artificialgroup (Harley & Smith 1983; Vilgalys & Cubeta 1994; Cubeta& Vilgalys 1997). Fungal systematics is based on the mor-phology of sexual structures (teleomorphs), but it isdifficult to induce
Rhizoctonia
-like fungi to produce suchstructures in the laboratory (Warcup & Talbot 1966, 1971;Sneh
et al
. 1991; Roberts 1999). To resolve this problem,molecular systematics is now being applied successfully to
Rhizoctonia
-like fungi (Vilgalys & González 1990; Vilgalys
& Cubeta 1994; Kuninaga
et al
. 1997; Taylor & Bruns 1997,1999; González
et al
. 2001; Pope & Carter 2001). In thepresent study we used two complimentary approaches tocompare mycorrhizal preferences and performances in twoepiphytic orchids from Puerto Rico: fungal phylogeny andsymbiotic seed germination. We tested the null hypothesisthat the two orchid species are generalists in their associa-tions with mycorrhizal fungi. If this is true, then we havetwo expectations. Fungi isolated from different orchidspecies collected from the same site should be more closelyrelated phylogenetically than fungi isolated from a singleorchid species in different sites. Using DNA sequences weconstructed a phylogenetic tree for fungi isolated from thetwo epiphytic orchids,
Tolumnia variegata
(Swartz) Braemand
Ionopsis utricularioides
(Swartz) Reichenbach f., growingsympatrically in two of the sites. We chose those two speciesbecause preliminary data indicated contrasting levelsof specificity (Otero
et al
. 2002). Our second expectation isthat seeds of each species will germinate and developequally well with fungi isolated from the other speciesas with fungi from itself. This was tested in a symbioticgermination experiment with seeds from
T. variegata
and
I.utricularioides
and fungi isolated from both orchids. If theamplitude of preferences proved to be contrasting, then wealso expected that growth rates would be higher for themore specific relationship.
Methods
Orchid species and study sites
Tolumnia variegata
occurs in the Greater Antilles, exceptJamaica (Ackerman 1995), whereas
Ionopsis utricularioides
(Swartz) Lindley has a more extensive range, including theAntilles, Florida (USA), Mexico, Central America, tropicalSouth America and the Galapagos Islands. These twospecies were chosen because they are abundant in PuertoRico, and many aspects of their biology are well studied(Montalvo & Ackerman 1987; Meléndez & Ackerman 1993,1994; Ackerman
et al
. 1996; Sabat & Ackerman 1996; Otero
et al
. 2002). The two species are related and ecologicallysimilar, facilitating comparative studies. Both are twigepiphytes in the subtribe Oncidiinae (Williams
et al
. 2001)and grow on small trees or shrubs such as guava (
Psidiumguajava
L.) and tintillo (
Randia aculeata
L.). They differ eco-logically in that
I. utricularioides
occurs in moist to wetbroadleaf forests whereas
T. variegata
inhabitats wet todry forests. Orchid roots were collected from five sites inPuerto Rico: Tortuguero, Cambalache, Dorado, Guánicaand San Cristóbal Canyon Three study sites had sympatricpopulations of the two orchid species, often growing on thesame host tree (Otero 2002), which allowed us to control forenvironmental variation. Voucher specimens are depositedat Universidad de Puerto Rico — Rio Piedras.

M Y C O R R H I Z A L S P E C I F I C I T Y I N T R O P I C A L O R C H I D S
2395
© 2004 Blackwell Publishing Ltd,
Molecular Ecology
, 13, 2393–2404
Fungal isolation
The most efficient way to obtain orchid mycorrhizaeis to remove pelotons (hyphal coils in the root cortex)and culture them. This has proven effective for temperatespecies with massive infections (Rasmussen 1995), butpelotons are scarce in many tropical epiphytic orchids,including
T. variegata
and
I. utricularioides
, and very dif-ficult to isolate. Consequently, the following methodwas used. Four roots per plant were surface-sterilized bya 2-min wash with tap water and 0.1% Tween-20, a 1 minwash with 70% ethanol, a 30-s wash with 2.5% sodiumhypochlorite (50% Clorox), and a final 1-min rinse in 70%ethanol (Otero
et al
. 2002). Three 0.5–1-cm pieces of rootwere incubated in a Petri dish with potato dextroseagar (Difco Inc.) and incubated at 25
°
C with light : darkcycles of 12 : 12 h for 1–2 weeks. The cultures were exam-ined by light microscopy at 400
×
magnification, the
Rhizoctonia
-like fungi were characterized according to theconcept of Sneh
et al
. (1991), and they were transferred topure culture.
Sequencing of internal transcribed spacer (ITS) region and phylogenetic analysis
The DNA extraction used the Lee & Taylor (1990) miniprepmethod. The ITS region of nuclear ribosomal DNA wasamplified by polymerase chain reaction (PCR) using theprimers ITS1 and ITS4 (White
et al
. 1990). PCR productswere purified using the QIAquick PCR Purification Kit(Qiagen) and sequenced using the BigDye cycle sequencingkit with the automatic sequencer ABI Prism 377 (AppliedBiosystems) according to the manufacturer’s instructions.Sequences were edited using the
sequencher
3.0 program(Gene Codes Corp.) and compared with sequences availablein GenBank for taxonomic confirmation. Sequences werealigned by the
clustal
method using the program
soap
v1.05a (Löytynoja & Milinkovitch 2001). Twenty-four
clustal
W (Thomson
et al
. 1994) alignments were produced, varyingthe gap-opening penalty between 15 and 25 in steps of two,and the gap-extension penalty between 8 and 14 also in stepsof two. The resulting consensus alignments were adjustedby eye. The alignments were subjected to a
modeltest
3.06 to determine the mode of evolution to be used forphylogenetic analysis (Posada & Crandall 1998). The mostlikely model of sequence evolution was GTR + G + I. Phylo-genetic relationships were estimated using both neighbour-joining and a heuristic search with maximum likelihoodmethods in
paup
(Version 40b8, Swofford 1998). Multinucleate
Rhizoctonia
fungi were used as the outgroup. Because thedata generated the same topology using both methods,we present the maximum likelihood tree with neighbour-joining bootstrap values (1000 replications) indicating supportfor the nodes.
Seed collection and symbiotic germination experiments
Ten fruits each of
I. utricularioides
and
T. variegata
werecollected in the Tortuguero Natural Reserve (Manatí, PR)on August, 2001, one fruit per plant. Fruits were surfacesterilized as described above. Seeds were transferred tosterile water and the viability was measured with triphenyltetrazolium chloride after staining for 24 h (Vujanovic
et al
.2000). Seed viability was 50–96% for each fruit. Seeds of thefive fruits with highest viability for each species werepooled for germination experiments.
Four fungal isolates from each orchid species werechosen for germination experiments. Two fungi from eachspecies came from the sympatric populations in Tortu-guero from Dorado
.
Fungi from
T. variegata
were chosenbecause preliminary tests showed that they stimulatedorchid seed germination; fungi from
I. utricularioides
werechosen at random. The phylogenetic position of the fungiwas not known at the time the germination experimentbegan. Fungal isolates were inoculated in Petri dishescontaining cellulose agar, a modification of Clements’isolation medium with sucrose replaced by 10 g/L cellulosepowder (Clements 1982). Cellulose is a carbon source thatis available to the fungus, but not to the plant. Three daysafter inoculation, 100
µ
L of a suspension of the pooledseeds (
x
= 346
±
86 seeds,
n
= 13) in sterile water wastransferred to each fungal culture. Each fungus–orchidcombination was replicated five times. Controls werecellulose agar plates with seeds but without fungi; forsome fruits it was necessary to add fungicide (benomyl,35 ppm) to inhibit the growth of endophytic fungi in thefruits. The fungicide did not inhibit seed germination. Itis known that orchid seeds do not grow on culture mediawithout either sugars or mycorrhizal fungi (Rasmussen1995). For
I. utricularioides
, fruits for control plates werecollected separately from the same populations because ofendophyte contamination of the original plates; in bothcases pooled seeds were used. The plates were incubatedin the laboratory with natural and fluorescent light at24
°
C.Data were collected by inspecting each plate at 40
×
magnification three times: 6, 10 and 13 weeks after sowing.In each plate, 30 or more randomly selected seeds werecensused. Each seed or seedling observed was assigneda score (0–6) on the germination scale (Table 1; Stenberg& Kane 1998). The percentage of seed germination wasestimated by dividing the number of seeds censusedthat had germinated (with score 1–6) by the total numberof seeds censused. A standardized Growth Index (GI)modified from Spoerl (1948) was calculated by the follow-ing formula:
GI
=
(
N
1
+
N
2
* 2
+
N
3
* 3
+
N
4
* 4
+
N
5
* 5
+
N
6
* 6)/(
N
0
+
N
1
+
N
2
+
N
3
+
N
4
+
N
5
+
N
6
)
2396
J . T . O T E R O , J . D . A C K E R M A N and P . B A Y M A N
© 2004 Blackwell Publishing Ltd,
Molecular Ecology
, 13, 2393–2404
where
N
0
is the number of seeds in stage zero,
N
1
is thenumber of seeds in stage one and so on. The GI can rangefrom zero (when no seeds germinated) to six (when allseeds reached seedling stage)
.
Statistical analysis
To determine whether or not there were significant dif-ferences in seed germination and GI, we analysed the datausing a factorial analysis of variance (
anova
). The factorswere source of the seeds (
T. variegata vs. I. utricularioides),fungal source (T. variegata vs. I. utricularioides), site (Tortuguerovs. South Dorado), and fungal strain (the eight isolates
chosen). For this particular experiment, fungal clade wasnot a factor because clade B was over-represented. The GIdata were standardized by dividing each value by six. Thegermination data were per cent values, and the standardizedGI did not have a normal distribution, so both data setswere normalized with an arcsine square root transformation(Zar 1999). Site and fungal strain were considered randomeffects (Zar 1999). The analysis was performed using theprogram statistica (StatSoft, Inc.). To compare the diversityof Rhizoctonia communities of T. variegata to those of I.utricularioides the Shannon–Weiner Index (H′) was calculatedusing the number of fungi of each clade isolated in eachsite. These indices were compared with Student’s t-tests (Zar1999). To compare the distribution of clades of Rhizoctoniaamong sites, observed and expected values were comparedwith a goodness of fit G-test (Zar 1999). Zero values werereplaced with 0.1 to allow calculation of logarithms.
Results
Phylogenetic analysis of preferences
Sixty-three uni- and binucleate Rhizoctonia-like fungiwere isolated from the roots of Ionopsis utricularioides andTolumnia variegata (Table 2). Almost all fungi belongedto the A, B, C and D clades of Ceratobasidium previously
Table 1 Germination stages of Tolumnia variegata seeds underlaboratory conditions
Stage Description
0 No germination1 Swelling of the embryo2 Development of radical hairs3 Development of a leaf projection4 Development of the first leaf5 Development of the second leaf6 Development of roots
Table 2 Isolates of Rhizoctonia-like fungi used in this study
Isolate Origin Site* Host orchid Genbank accession no.
JTO-001 Puerto Rico C T. variegata AF503958JTO-002 Puerto Rico C T. variegata AF472278JTO-004 Puerto Rico T T. variegata AF472279JTO-006 Puerto Rico T T. variegata AF503959JTO-008 Puerto Rico T T. variegata AY443531JTO-010 Puerto Rico T I. utricularioides AF472280JTO-014 Puerto Rico T T. variegata AF503960JTO-015 Puerto Rico T T. variegata AF503961JTO-016 Puerto Rico C T. variegata AF503962JTO-017 Puerto Rico T T. variegata AF503963JTO-018 Puerto Rico T T. variegata AF503964JTO-019 Puerto Rico T T. variegata AF503965JTO-020 Puerto Rico T T. variegata AF503966JTO-021 Puerto Rico C T. variegata AF503967JTO-022 Puerto Rico C T. variegata AF503968JTO-023 Puerto Rico C T. variegata AF503969JTO-024 Puerto Rico T I. utricularioides AF472281JTO-031 Puerto Rico C T. variegata AF503970JTO-032 Puerto Rico C I. utricularioides AF472282JTO-033 Puerto Rico T T. variegata AF503971JTO-037 Puerto Rico T T. variegata AF503972JTO-041 Puerto Rico T T. variegata AF503973JTO-042 Puerto Rico T T. variegata AF503974JTO-043 Puerto Rico T I. utricularioides AF472283JTO-044 Puerto Rico T T. variegata AF503975JTO-047 Puerto Rico T I. utricularioides AF472284
M Y C O R R H I Z A L S P E C I F I C I T Y I N T R O P I C A L O R C H I D S 2397
© 2004 Blackwell Publishing Ltd, Molecular Ecology, 13, 2393–2404
Table 2 Continued
Isolate Origin Site* Host orchid Genbank accession no.
JTO-049 Puerto Rico C T. variegata AF503976JTO-050 Puerto Rico C T. variegata AF503977JTO-058 Puerto Rico SC T. variegata AF472287JTO-060 Puerto Rico SC I. utricularioides AF503978JTO-063 Puerto Rico SC I. utricularioides AF503979JTO-089 Puerto Rico C T. variegata AF503980JTO-092 Puerto Rico T I. utricularioides AF503981JTO-112 Puerto Rico SC I. utricularioides AF503982JTO-119 Puerto Rico SC I. utricularioides AF503983JTO-126 Puerto Rico D T. variegata AF503984JTO-127 Puerto Rico D I. utricularioides AF503985JTO-128 Puerto Rico D T. variegata AF503986JTO-129 Puerto Rico D I. utricularioides AF503987JTO-131 Puerto Rico D I. utricularioides AF503988JTO-132 Puerto Rico D I. utricularioides AF503989JTO-133 Puerto Rico D I. utricularioides AF503990JTO-134 Puerto Rico D T. variegata AF503991JTO-135 Puerto Rico D T. variegata AF503992JTO-138 Puerto Rico D T. variegata AF503993JTO-141 Puerto Rico D T. variegata AF503994JTO-142 Puerto Rico D T. variegata AF503995JTO-143 Puerto Rico D T. variegata AF503996JTO-144 Puerto Rico D T. variegata AF503997JTO-145 Puerto Rico D T. variegata AF503998JTO-147 Puerto Rico D T. variegata AF503999JTO-148 Puerto Rico D T. variegata AF504000JTO-149 Puerto Rico D T. variegata AF504002JTO-153 Puerto Rico D T. variegata AF504001JTO-155 Puerto Rico D I. utricularioides AF504004JTO-157 Puerto Rico D I. utricularioides AF504005JTO-158 Puerto Rico D I. utricularioides AF504003JTO-159 Puerto Rico D I. utricularioides AF504006JTO-160 Puerto Rico D I. utricularioides AF504007JTO-161 Puerto Rico G T. variegata AF504008JTO-162 Puerto Rico G T. variegata AF472302JTO-171 Puerto Rico D T. variegata AF504009JTO-173 Puerto Rico D T. variegata AF504010C. AG-Q Japan Soil AF354095C. CAG1 US Turf grass AF354086C. AGD Japan ? AF354090C. bicorne ? Moss AF200514R. 83 Finland Pinus sylvestris AF200516R. 85 Finland Pinus sylvestris AF200519R 248–1 Finland Pinus sylvestris AF200517C. AG-O Japan Soil AF354094C. AG-L Japan Soil AF354093C. AG-Bo Japan Sweet potato AF354091C.AG-A Japan Soil AF354092R. AG-A ? ? AB000040ROTH-25 Japan? AB019027Puerto Rico-1 Puerto Rico Phaseolus vulgaris AF153805Puerto Rico-5 Puerto Rico Phaseolus vulgaris AF308629
The first 63 were isolated from Ionpsis utricularioides and Tolumnia variegata; the others are isolates and sequences included for comparison in the phylogenetic analysis.*Sites: C = Cambalache, D = Dorado, G = Guánica, SC = San Cristóbal, T = Tortuguero.

2398 J . T . O T E R O , J . D . A C K E R M A N and P . B A Y M A N
© 2004 Blackwell Publishing Ltd, Molecular Ecology, 13, 2393–2404
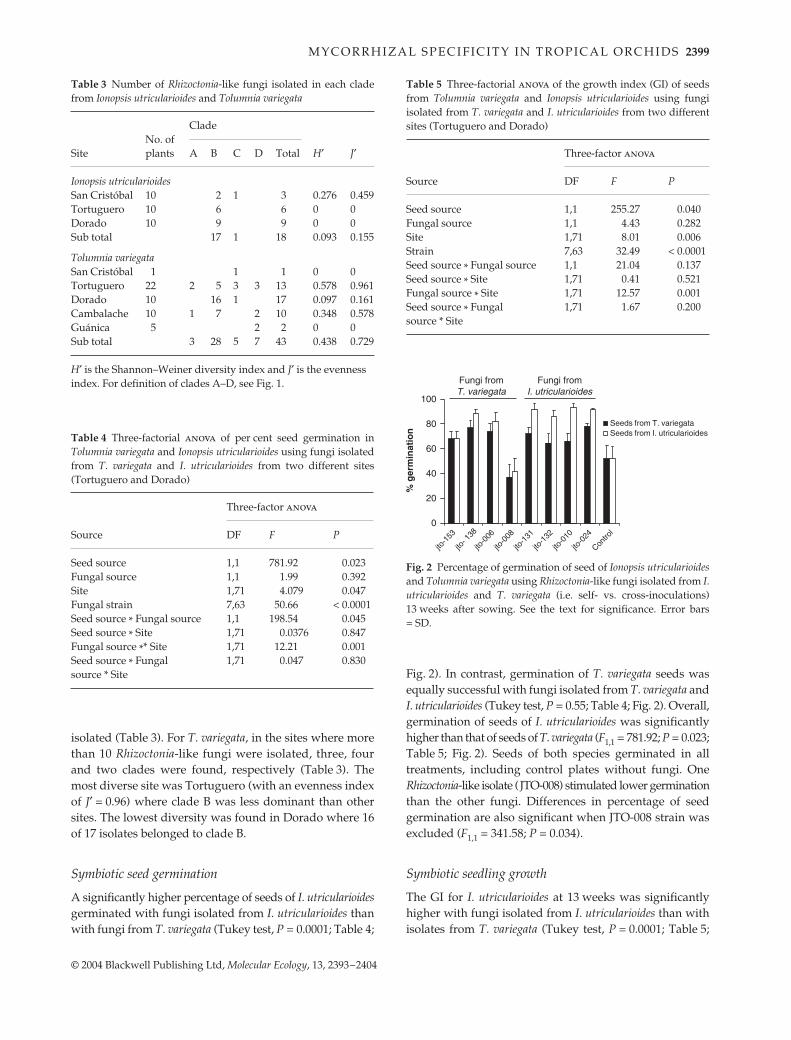
described (Otero et al. 2002; Fig. 1(a,b). These clades had100, 82, 88 and 100% bootstrap support, respectively. CladeA was closely related to Ceratobasidium group Ag-Q, isolatedfrom soil in Japan (González et al. 2001). Clades B and Cwere sister groups to Clade A. Clades A, B and C andCeratobasidium Ag-Q formed a monophyletic group with98% bootstrap support. Clade D formed a separate cladeand was related to Ceratobasidium Ag-H. One sequence(JTO008) could not be aligned with the rest. Consequently,it is not included in the cladogram. It was similar toBjerkandera adusta [AB096737, identities = 543/579 (93%);Sato et al. 2003].
All fungi isolated from I. utriculariodes belonged to cladeB, except one in clade C isolated from San Cristóbal. Fungi
from T. variegata belonged to all clades (Fig. 1). Rhizoctonia-likeendophytes isolated from T. variegata were significantlymore diverse than those from I. utricularioides (Student’st = 4.03, d.f. = 42, P = 0.0002, Table 3).
Geographic variation
Differences among Tortugero, Dorado and Cambalache indistribution of Rhizoctonia clades were significant for T.variegata (G-test, G = 14.9, d.f. = 6, P = 0.02). (Sites with < 10isolates were not included; this test was not appropriate forfungi from I. utricularioides because all but one belonged toClade B.) In two of three sampling sites for I. utricularioides,fungi from clade B were the only Rhizoctonia-like fungi
Fig. 1 Maximum likelihood trees of ITS sequences of Ceratobasidium fungi isolated from Tolumnia variegata and Ionopsis utricularioides. (a)relationships among clades A, B, C and D; (b) clade B in detail. Fungi in bold font were used in the germination experiment. Numbers showthe bootstrapping percentage that supports the branches. C = Cambalache, D = Dorado, G = Guánica, SC = San Cristóbal, T = Tortuguero.
M Y C O R R H I Z A L S P E C I F I C I T Y I N T R O P I C A L O R C H I D S 2399
© 2004 Blackwell Publishing Ltd, Molecular Ecology, 13, 2393–2404
isolated (Table 3). For T. variegata, in the sites where morethan 10 Rhizoctonia-like fungi were isolated, three, fourand two clades were found, respectively (Table 3). Themost diverse site was Tortuguero (with an evenness indexof J′ = 0.96) where clade B was less dominant than othersites. The lowest diversity was found in Dorado where 16of 17 isolates belonged to clade B.
Symbiotic seed germination
A significantly higher percentage of seeds of I. utricularioidesgerminated with fungi isolated from I. utricularioides thanwith fungi from T. variegata (Tukey test, P = 0.0001; Table 4;
Fig. 2). In contrast, germination of T. variegata seeds wasequally successful with fungi isolated from T. variegata andI. utricularioides (Tukey test, P = 0.55; Table 4; Fig. 2). Overall,germination of seeds of I. utricularioides was significantlyhigher than that of seeds of T. variegata (F1,1 = 781.92; P = 0.023;Table 5; Fig. 2). Seeds of both species germinated in alltreatments, including control plates without fungi. OneRhizoctonia-like isolate (JTO-008) stimulated lower germinationthan the other fungi. Differences in percentage of seedgermination are also significant when JTO-008 strain wasexcluded (F1,1 = 341.58; P = 0.034).
Symbiotic seedling growth
The GI for I. utricularioides at 13 weeks was significantlyhigher with fungi isolated from I. utricularioides than withisolates from T. variegata (Tukey test, P = 0.0001; Table 5;
Table 3 Number of Rhizoctonia-like fungi isolated in each cladefrom Ionopsis utricularioides and Tolumnia variegata
SiteNo. of plants
Clade
H′ J′A B C D Total
Ionopsis utricularioidesSan Cristóbal 10 2 1 3 0.276 0.459Tortuguero 10 6 6 0 0Dorado 10 9 9 0 0Sub total 17 1 18 0.093 0.155
Tolumnia variegataSan Cristóbal 1 1 1 0 0Tortuguero 22 2 5 3 3 13 0.578 0.961Dorado 10 16 1 17 0.097 0.161Cambalache 10 1 7 2 10 0.348 0.578Guánica 5 2 2 0 0Sub total 3 28 5 7 43 0.438 0.729
H′ is the Shannon–Weiner diversity index and J′ is the evenness index. For definition of clades A–D, see Fig. 1.
Table 4 Three-factorial anova of per cent seed germination inTolumnia variegata and Ionopsis utricularioides using fungi isolatedfrom T. variegata and I. utricularioides from two different sites(Tortuguero and Dorado)
Source
Three-factor anova
DF F P
Seed source 1,1 781.92 0.023Fungal source 1,1 1.99 0.392Site 1,71 4.079 0.047Fungal strain 7,63 50.66 < 0.0001Seed source * Fungal source 1,1 198.54 0.045Seed source * Site 1,71 0.0376 0.847Fungal source ** Site 1,71 12.21 0.001Seed source * Fungal 1,71 0.047 0.830source * Site
Table 5 Three-factorial anova of the growth index (GI) of seedsfrom Tolumnia variegata and Ionopsis utricularioides using fungiisolated from T. variegata and I. utricularioides from two differentsites (Tortuguero and Dorado)
Source
Three-factor anova
DF F P
Seed source 1,1 255.27 0.040Fungal source 1,1 4.43 0.282Site 1,71 8.01 0.006Strain 7,63 32.49 < 0.0001Seed source * Fungal source 1,1 21.04 0.137Seed source * Site 1,71 0.41 0.521Fungal source * Site 1,71 12.57 0.001Seed source * Fungal 1,71 1.67 0.200source * Site
Fig. 2 Percentage of germination of seed of Ionopsis utricularioidesand Tolumnia variegata using Rhizoctonia-like fungi isolated from I.utricularioides and T. variegata (i.e. self- vs. cross-inoculations)13 weeks after sowing. See the text for significance. Error bars= SD.

2400 J . T . O T E R O , J . D . A C K E R M A N and P . B A Y M A N
© 2004 Blackwell Publishing Ltd, Molecular Ecology, 13, 2393–2404
Fig. 3). In contrast, the GI for T. variegata seeds did notdiffer significantly between fungi from T. variegata andfrom I. utricularioides (Tukey test, P = 0.701; Table 5; Fig. 3).In general, seedlings of I. utricularioides grew significantlymore than those of T. variegata (F1,1 = 255.27; P = 0.040;Table 5; Fig. 3). Seeds of I. utricularioides had higher GIwhen inoculated with six of the eight fungal strains thanuninoculated control seeds [Fig. 3; Tukey–Kramer HonestlySignificant Difference (HSD) tests]. Four of the eight fungistimulated significantly higher GI in T. variegata seedscompared to uninoculated controls.
One Rhizoctonia-like isolate ( JTO-008) did not stimulategrowth and was not closely related to the others. It ispossible that JTO-008 was not a mycorrhizal fungus, andbiased the results of the anova. For this reason we repeatedthe anovas without both JTO-008 and JTO-132 (chosen atrandom), an isolate from I. utricularioides eliminated tobalance the anova. For the seed germination analyses, thereremained significant differences (F1,1 = 341.58; P = 0.034).Likewise, differences in the effect on GI of seeds of I. utric-ularioides between fungi isolated from T. variegata and I.utricularioides remained significant (F1,1 = 17.2, P = 0.0001).
There was a site effect on the development of seeds.Fungi from Dorado induced a higher GI than those fromTortuguero (F1,71 = 8.01, P = 0.006), and the differences wereindependent of the seed-source orchid (F1,71 = 0.41, P = 0.521).In addition, the effects of fungal isolates were significant(F1,71 = 12.57, P = 0.0007), but not the interactions amongsource of seeds, fungal host and site (F1,71 = 1.67, P = 0.20).
In general, fungi that stimulated seed germination alsostimulated seedling development. This correlation was strongfor I. utricularioides (r2 = 0.78, n = 8, P = 0.0006) and marginal forT. variegata (r2 = 0.67, n = 8, P = 0.0665). The fungi that inducedhighest seed germination, seedling development and rateof change in the GI (JTO-010, JTO-024, JTO-131, JTO-132, andJTO-138) all belonged to Ceratobasidium clade B (Fig. 1b).
Discussion
Preference in orchid mycorrhizal relationships
Both fungal phylogeny and symbiotic seed germinationexperiments showed that in Puerto Rico Ionopsis utri-cularioides was more specific than Tolumnia variegata in itsmycorrhizal relationships. Fungi from T. variegata belongedto four clades within Ceratobasidium, whereas fungi from I.utricularioides belonged to one clade (with one exception)(Fig. 1b). The germination experiments also showed thatI. utricularioides is more specific than that of T. variegata.In I. utricularioides, both seed germination and GI weresignificantly higher with fungi isolated from plants ofthe same species than from plants of T. variegata (Figs 2, 3).In contrast, T. variegata showed no preference for fungiisolated from itself. Thus species with a similar habit(twig epiphytes, sensu Chase 1987) that are phylogeneticallyrelated and have overlapping habitat preferences can bequite different in the amplitude of their mycorrhizal pre-ferences. In a previous study we compared mycorrhizalfungi and mycorrhizal specificity among nine species ofepiphytic orchids in Puerto Rico (Otero et al. 2002). A rangeof specificity was observed, with I. utricularioides being themost specific. The present study combines a more extensivesampling of I. utricularioides and T. variegata with symbioticseed germination tests that test the ability of the fungito stimulate plant growth. These data shed new lighton the debate regarding specificity of orchid mycorrhizalrelationships (Dressler 1981, 1993; Taylor & Bruns 1997;Otero et al. 2002). Unlike previous studies we found thatboth generalist and specific interactions occur in orchidsthat are phylogenetically related, and also in species ofsimilar habit—both are twig epiphytes (sensu Chase 1987)often growing side by side. The fact that both specieswere sampled on the same host plants removes orchiddistribution and host preference as a complicating factor.
Obligately mycorrhizal, nonphotosynthetic plants aredependent on their mycorrhizal fungi for carbon through-out their lives (Leake 1994). These myco-trophic plants areexpected to have higher specificity than their photo-synthetic counterparts (Taylor et al. 2002). Indeed, they allexploit specific fungal genera. Surprisingly, the photo-synthetic T. variegata and I. utricularioides do the same byexploiting only clades of Ceratobasidium. Furthermore, thebreadth of mycorrhizal preferences in myco-heterotrophicspecies of various families, including orchids, is no lessnarrow than that of T. variegata of Puerto Rico (Taylor &Bruns 1997, 1999; Kretzer et al. 2000; McKendrick et al. 2000,2002; Bidartondo & Bruns 2001, 2002; Bidartondo et al. 2002).Consequently, I. utricularioides appears to be more specificthan any of the nonphotosynthetic plants studied.
Mycorrhizal specificity is evolutionarily relevant at thespecies level only in the context that preferences have a
Fig. 3 Growth index (GI) of seeds from Ionopsis utricularioides andTolumnia variegata using Rhizoctonia-like fungi isolated from I. utri-cularioides and T. variegata (i.e. self- vs. cross-inoculations) 13 weeksafter sowing. See the text for significance. Error bars = SD.

M Y C O R R H I Z A L S P E C I F I C I T Y I N T R O P I C A L O R C H I D S 2401
© 2004 Blackwell Publishing Ltd, Molecular Ecology, 13, 2393–2404
genetic component and that gene flow among populationsis significant. Although we have no data on the geneticbasis for mycorrhizal preferences, we do know that thegenetic variation among populations of T. variegata isconsistent with high levels of gene flow among PuertoRican populations and reduced gene flow among islands(Ackerman & Ward 1999). It is thus appropriate to focuson mycorrhizal associations within an island. Any othermycorrhizal fungal associates that may be found for thesespecies outside Puerto Rico may indicate geographicalmosaics in the preferences of the orchids.
The difference in mycorrhizal preferences between ourtwo orchid species is probably robust. We sampled fivedifferent locations scattered throughout the island. Dis-tributions of fungi among sites showed that the narrowpreferences of I. utricularioides was not imposed by limiteddistributions of potential partners. The association of fungiwith the two orchids was statistically different from ran-dom. A Fisher exact test carried out using the number offungi from clade B, and number of fungi from otherclades (A, C and D) from I. utricularioides and T. variegata:(17, 1; 28, 13); showed that the association was not random(P = 0.044). Furthermore, mycorrhizal isolates of I. utri-cularioides from Costa Rica and Panama also belong to cladeB (Otero, unpublished data). Although we expect to findmore mycorrhizal isolates if more populations were sam-pled, we do not expect that the contrast between the twospecies will change.
Methods for assessing specificity in orchid–fungus relationships
To determine specificity between orchids and their mycorrhi-zal fungi, some studies have used symbiotic germinationexperiments (Hadley 1970; Clements 1987, 1988; Masuhara& Katsuya 1989, 1991, 1994; Muir 1989; Smreciu & Currah1989; Masuhara et al. 1993; Zettler et al. 1999; McKendricket al. 2000, 2002), and others have used identification offungi (Warcup & Talbot 1971; Currah et al. 1988, 1997;Masuhara et al. 1988; Currah 1991; Rasmussen 1995; Taylor& Bruns 1997, 1999; Pope & Carter 2001; Otero 2002; Oteroet al. 2002). Here we have integrated both techniques. It isimportant to have both perspectives because the isolationof a fungus from an orchid root does not necessarilymean that it is mycorrhizal (Masuhara & Katsuya 1994).Orchid roots also contain a variety of endophytes, fungithat live in plant tissues without causing symptoms ofdisease (Richardson et al. 1993; Richardson & Currah 1995;Bayman et al. 1997; Currah et al. 1997). These endophytesform a continuum of interactions with plants, from mut-ualists to potential pathogens (Saikkonen et al. 1998). It islikely that some fungi isolated or amplified from roots areendophytic and not mycorrhizal which is why germina-tion experiments are critical. Because some mycorrhizal
fungi may not be culturable, it is possible that other my-corrhizal fungi were also present in the roots of T. variegataand I. utricularioides. Direct PCR amplification from pelotonscan address this problem (Kristiansen et al. 2001; Tayloret al. 2003), though the use of taxon-specific PCR primerscan also bias sampling.
Orchid seeds are tiny and dust-like; they lack endo-sperm and consist of an undifferentiated embryo with fewstored nutrients. They depend on their mycorrhizal fungifor germination and growth, but can be independent asjuveniles or adults in photosynthetic species (Dressler 1981;Arditti 1992; Rasmussen 1995). However, their small sizemakes orchid seed germination very difficult to study inthe field. Techniques exist for retrieving seeds for study(Rasmussen & Whigham 1993), but they have not yet beenapplied to epiphytic orchids. Because of this difficulty, itremains unclear whether these tests of seed germinationon nutritive media under aseptic conditions are truly rep-resentative of the process under natural conditions.
Masuhara & Katsuya (1994) showed that in the temperateterrestrial orchid, lower levels of specificity occur underin vitro conditions than under in situ conditions. Thus,isolation of endophytic fungi from adult plants followedby in vitro seed germination experiments suggest thatthose fungi are potentially mycorrhizal in the field, but onemight expect to measure higher levels of specificity with insitu conditions. If this bias holds true, then I. utricularioidesmay have even narrower preferences than those detectedin this study.
Implications of differences in mycorrhizal preferences
High mycorrhizal specificity suggests that mycorrhizaecould play a role in orchid speciation and diversification(Rasmussen 1995; Taylor et al. 2002, 2004). Our data showthat distribution of mycorrhizal fungal clades differedsignificantly among sites (Table 3). For orchids with relativelynarrow mycorrhizal preferences, the irregular occurrenceof the fungi could produce patchy distributions of orchidpopulations, adversely affecting gene flow, leading to repro-ductive isolation of populations (Ackerman & Zimmerman1994; Tremblay & Ackerman 2001). Since T. variegata isassociated with four Ceratobasidium clades, patchy distributionof a single clade is less likely to affect the distribution of theorchid. The fact that T. variegata has greater ecological ampli-tude than I. utricularioides is consistent with this hypothesis.
Where there is variation in performance among mycor-rhizal fungal symbionts, we expect orchid populations toexperience selection for specialization on the fungi thatinduces the best germination and seedling development.Specialized relationships should be the derived condition(Taylor & Bruns 1999). However, when populations of fun-gal symbionts vary in time and space, a generalist strategymay be advantageous, just as it can be for plant–pollinator

2402 J . T . O T E R O , J . D . A C K E R M A N and P . B A Y M A N
© 2004 Blackwell Publishing Ltd, Molecular Ecology, 13, 2393–2404
interactions (Schemske & Horvitz 1984; Waser et al. 1996).Unfortunately, distribution, temporal variation and naturalhistory of orchid mycorrhizal fungi are almost completelyunknown.
Orchids are considered to be parasites of their my-corrhizal fungi (Smith 1966, 1967; Alexander et al. 1984;Hadley 1984; Alexander & Hadley 1985). Specificity andefficient exploitation are often associated in parasites (Price1980), and our data are consistent with these observations.Seed germination and seedling development are better forI. utricularioides but only occur with a narrow range of Cer-atobasidium. On the other hand, germination and develop-ment are not as rapid for T. variegata but it has a muchbroader choice of fungi. Thus, contrasting trade-offs existas one would predict for the specialists and generalists.
Orchids are dependent on mycorrhizal fungi for ger-mination, and many of the unique characteristics of orchidsare associated with the mycorrhizal relationship (Rasmussen1995). Here we show that related orchid species with over-lapping habitat preferences may differ in mycorrhizalspecificity or preferences and that the distribution of theirfungi is patchy. We found that narrow preferences areassociated with efficient exploitation but may restrict theecological/or geographical amplitude of the orchids. Ourresults indicate that mycorrhizal relationships may beshaped by selection, may affect the distribution of orchids,and ultimately may influence orchid diversification.
Acknowledgements
We thank Amauri Castro, Angelica Carrillo, Jessyka García, LauraFidalgo Paola Pabón, Andrea Porras and Sergio Rocafort forassistance in the laboratory, Tugrul Giray for valuable ideas, W.Owen McMillan for use of facilities, and Nicola S. Flanagan forsupport, all at UPR-Río Piedras. Alberto Sabat, Nicola S. Flanagan,Kelli Gowland and anonymous reviewers commented on earlyversions of the manuscript. Jess Zimmerman and Todd Crowl pro-vided statistical advice. Nyeri and Inti gave us extra stimulus forfinishing this research. This research was supported by an NSF-EPSCoR scholarship (NSF grant EPS-9874782), an Organizationfor Tropical Studies post-course fellowship (OTS-2000–13), and aNASA-IRA grant to the University of Puerto Rico to J. T. Otero.
References
Ackerman JD (1983) Diversity and seasonality of male euglossine bees(Hymenoptera: Apidae) in central Panama. Ecology, 64, 274–283.
Ackerman JD (1995) An orchid flora of Puerto Rico and the VirginIslands. Memoirs of the New York Botanical Garden, 73, 1–203.
Ackerman JD, Zimmerman JK (1994) Bottlenecks in the life historiesof orchids: resources, pollination, population structure andseedling establishment. In: Proceedings of the 14th World OrchidConference (ed. Pridgeon A), pp. 125–129. Scientific Books BaloghInternational Inc, Champaign, IL.
Ackerman JD, Ward S (1999) Genetic variation in a widespread,epiphytic orchid: where is the evolutionary potential? SystematicBotany, 24, 282–291.
Ackerman JD, Sabat A, Zimmerman JK (1996) Seedling establish-ment in an epiphytic orchid: an experimental study of seed lim-itation. Oecologia, 106, 192–198.
Alexander C, Alexander IJ, Hadley G (1984) Phosphate uptake byGoodyera repens in relation to mycorrhizal infection. New Phytol-ogist, 97, 391–400.
Alexander C, Hadley G (1985) Carbon movement between hostand mycorrhizal endophyte during the development of theorchid Goodyera repens. New Phytologist, 101, 657–665.
Arditti J (1992) Fundamentals of Orchid Biology. John Wiley andSons, New York.
Arditti J, Ernst R, Yam TW, Glabe C (1990) The contributions oforchid mycorrhizal fungi to seed germination: a speculativereview. Lindleyana, 5, 249–255.
Bayman P, Lebrón LL, Tremblay RL, Lodge DJ (1997) Variationin endophytic fungi from roots and leaves of Lepanthes(Orchidaceae). New Phytologist, 135, 143–149.
Benzing DH (1986) The vegetative basis of vascular epiphytism.Selbyana, 9, 23–43.
Bernard N (1909) L’evolution dans la symbiose. Annales Des SciencesNaturelles; Botanique, Paris, 9, 1–196.
Bidartondo MI, Bruns TD (2001) Extreme specificity in epiparasiticMonotropoideae (Ericaceae): widespread phylogenetic andgeographical structure. Molecular Ecology, 10, 2285–2295.
Bidartondo MI, Bruns TD (2002) Fine-level mycorrhizal specificityin the Monotropoideae (Ericaceae): specificity for fungal speciesgroups. Molecular Ecology, 11, 557–569.
Bidartondo MI, Redecker D, Hijri I, et al. (2002) Epiparasitic plantsspecialized on arbuscular mycorrhizal fungi. Nature, 419, 389–392.
Burgeff H (1932) Saprophytismus und Symbiose. Fischer Verlag,Jena.
Chase MW (1987) Obligate twig epiphytes: a distinct subset ofneotropical orchidaceous epiphytes. Selbyana, 10, 24–30.
Clements MA (1982) Developments in the symbiotic germinationof Australian terrestrial orchids. In: Proceedings of the 10th WorldOrchid Conference (eds Stewart J, van der Merwe CN), pp. 269–273. South African Orchid Council, Johannesburg.
Clements MA (1987) Orchid–fungus–host associations of epi-phytic orchids. In: Proceedings of the 12th World Orchid Conference(eds Saito K, Tanaka R), pp. 80–83. Japanese Orchid Council,Tokyo.
Clements MA (1988) Orchid mycorrhizal associations. Lindleyana,3, 73–86.
Cubeta MA, Vilgalys R (1997) Population biology of the Rhizoctoniasolani complex. Phytopathology, 87, 480–484.
Currah RS (1991) Taxonomic and developmental aspects of thefungal endophytes of terrestrial orchid mycorrhizae. Lindleyana,6, 211–213.
Currah RS, Hambleton S, Smreciu A (1988) Mycorrhizae andmycorrhizal fungi of Calypso bulbosa. American Journal of Botany,75, 739–752.
Currah RS, Zettler LW, Hambleton S, Richardson KA (1997) Fungifrom orchid mycorrhizae. In: Orchid Biology: Reviews and Perspec-tives, Vol. VII (eds Arditti J, Pridgeon AM), pp. 117–170. KluwerAcademic Publishers, Dordrecht.
Curtis JT (1939) The relationship of specificity of orchid mycorrhizalfungi to the problem of symbiosis. American Journal of Botany, 26,390–399.
Darwin C (1877) The Various Contrivances by Which Orchids AreFertilized by Insects, 2nd edn. William Clowes and Sons, Limited,London.

M Y C O R R H I Z A L S P E C I F I C I T Y I N T R O P I C A L O R C H I D S 2403
© 2004 Blackwell Publishing Ltd, Molecular Ecology, 13, 2393–2404
Dressler RL (1981) The Orchids: Natural History and Classification.Harvard University. Press, Cambridge.
Dressler RL (1993) Phylogeny and Classification of the Orchid Family.Dioscorides Press, Portland, OR.
Freudenstein JV, van den Berg C, Goldman DH, Kores PJ, MolvrayM, Chase MW (2004) An expanded plastid DNA phylogeny ofOrchidaceae and analysis of jackknife branch support strategy.American Journal of Botany, 91, 149–157.
González D, Carling DE, Kuninaga S, Vilgalys R, Cubeta M(2001) Ribosomal DNA systematics of Ceratobasidium andThanatephorus with Rhizoctonia anamorphs. Mycologia, 93, 1138–1150.
Hadley G (1970) Non-specificity of symbiotic infection in orchidmycorrhizae. New Phytologist, 69, 1015–1023.
Hadley G (1984) Uptake of [14C] Glucose by asymbiotic andmycorrhizal orchid protocorms. New Phytologist, 96, 263–273.
Harley JL, Smith SE (1983) Mycorrhizal Symbiosis. Academic Press,London.
Johnson SJ, Steiner KE (2000) Generalization versus specializationin plant pollination systems. Trends in Ecology and Evolution, 15,140–143.
Knudson L (1922) Non-symbiotic germination of orchid seeds.Botanical Gazette, 77, 212–219.
Kretzer AM, Bidartondo MI, Grubisha L, Spatafora JW, Szaro TM,Bruns TD (2000) Regional specialization of Sarcodes sanguinea(Ericaceae) on a single fungal symbiont from the Rhizopogonellenae (Rhizopogonaceae) species complex. American Journal ofBotany, 87, 1778–1782.
Kristiansen KA, Taylor DL, Kjoller R, Rasmussen HN, Rosendahl S(2001) Identification of mycorrhizal fungi from single pelotonsof Dactylorhiza majalis (Orchidaceae) using single-strand con-formation polymorphism and mitochondrial ribosomal largesubunit DNA sequences. Molecular Ecology, 10, 2089–2093.
Kuninaga S, Natsuaki T, Takeuchi T, Yokosawa R (1997) Sequencevariation of the rDNA ITS regions within and between anas-tomosis groups in Rhizoctonia solani. Current Genetics, 32, 237–243.
Leake JR (1994) Tansley Review, 69: the biology of myco-heterotrophic(‘saprophytic’) plants. New Phytologist, 127, 171–216.
Lee SB, Taylor JW (1990) Isolation of DNA from fungal myceliaand single spores. In: PCR Protocols: a Guide to Methods andApplications (eds Innis MA, Gelfand DH, Snisnsky JJ, White TJ),pp. 282–287. Academic Press, San Diego CA.
Löytynoja A, Milinkovitch MC (2001) SOAP, cleaning multiplealignments for unstable blocks. Bioinformatics, 17, 573–574.
Masuhara G, Katsuya K (1989) Effects of mycorrhizal fungi onseed germination and early growth of three Japanese terrestrialorchids. Scitia Horticulturae, 37, 331–337.
Masuhara G, Katsuya K (1991) Fungal coil formation of Rhizoctoniarepens in seedlings of Galeola septentrionalis (Orchidaceae). Botan-ical Magazine of Tokyo, 104, 275–281.
Masuhara G, Katsuya K (1994) In situ and in vitro specificitybetween Rhizoctonia spp. and Spiranthes sinensis (Persoon) Ames.var. amoena (M. Bieberstein) Hara (Orchidaceae). New Phytologist,127, 711–718.
Masuhara G, Katsuya K, Yamaguchi K (1993) Potential for symbi-osis of Rhizoctonia solani and binucleate Rhizoctonia with seeds ofSpiranthes amoena var. amoena in vitro. Mycological Research, 97,746–752.
Masuhara G, Kimura S, Katsuya K (1988) Seasonal changes inmycorrhizae of Bletia striata (Orchidaceae). Transactions of theMycological Society of Japan, 29, 25–31.
McKendrick SL, Leake JL, Taylor DL, Read DJ (2000) Symbioticgermination and development of myco-heterotrophic plants innature: ontogeny of Corallorhiza trifida and characterization of itsmycorrhizal fungi. New Phytologist, 145, 523–537.
McKendrick SL, Leake JL, Taylor DL, Read DJ (2002) Symbioticgermination and development of the myco-heterotrophic orchidNeottia nidus-avis in nature and its requirements for locallydistributed Sebacina spp. New Phytologist, 154, 233–247.
Meléndez EJ, Ackerman JD (1993) The effects of a rust infection onfitness components in a natural population of Tolumnia variegata(Orchidaceae). Oecologia, 94, 361–367.
Meléndez EJ, Ackerman JD (1994) Factors associated with rustinfection (Sphenosphora saphaena) in an epiphytic orchid (Tolumniavariegata). American Journal of Botany, 81, 287–293.
Montalvo AM, Ackerman JD (1987) Limitation of fruit productionin Ionopsis utricularioides (Orchidaceae). Biotropica, 19, 24–31.
Muir HJ (1989) Germination and mycorrhizal fungus compatibilityin European orchids. In: Modern Methods in Orchid Conservation:the Role of Physiology, Ecology and Management (ed. PrichardHW), pp. 39–56. Cambridge University Press, Cambridge.
Otero JT (2002) Specificity of orchid mycorrhizae: their role in orchidevolution. PhD Thesis, University of Puerto Rico.
Otero JT, Ackerman JD, Bayman P (2002) Diversity and hostspecificity of endophytic Rhizoctonia-like fungi from tropicalorchids. American Journal of Botany, 89, 1852–1858.
van der Pijl L, Dodson CH (1966) Orchid Flowers: Their Pollinationand Evolution. University of Miami Press, Coral Gables, Florida.
Pope EJ, Carter DA (2001) Phylogenetic placement and host spe-cificity of mycorrhizal isolates AG-6 and AG-12 in the Rhizocto-nia solani species complex. Mycologia, 93, 712–719.
Posada D, Crandall KA (1998) Model test: testing the model ofDNA substitution. Bioinformatics, 14, 817–818.
Price PW (1980) Evolutionary Biology of Parasites. Princeton Univer-sity Press, Princeton NJ.
Rasmussen HN (1995) Terrestrial Orchids: from Seeds to MycotrophicPlants. Cambridge University Press, Cambridge.
Rasmussen HN, Whigham DF (1993) Seed ecology of dust seed insitu: a new technique and its applications in terrestrial orchids.American Journal of Botany, 80, 1374–1378.
Richardson KA, Currah RS (1995) The fungal community associ-ated with the roots of some rainforest epiphytes of Costa Rica.Selbyana, 16, 49–73.
Richardson KA, Currah RS, Hambleton S (1993) Basidiomycetousendophytes from the roots of neotropical epiphytic Orchidaceae.Lindleyana, 8, 127–137.
Roberts P (1999) Rhizoctonia-Forming Fungi: a Taxonomic Guide.Royal Botanical Garden, Kew, London.
Sabat AM, Ackerman JD (1996) Fruit set in a deceptive orchid: theeffect of flowering phenology, display size, and local floralabundance. American Journal of Botany, 83, 1181–1186.
Saikkonen K, Faeth SH, Helander M, Sullivan TJ (1998) Fungalendophytes: a continuum of interactions with host plants. AnnualReview of Ecology and Systematics, 29, 319–343.
Sato A, Watanabe Y, Nugroho NB, Chrisnayanti E, Natusion UJ,Koesnandar Nishida H (2003) Screening for dioxin-degradingbasidiomycetes from temperate and tropical forest. World Journalof Microbiology and Biotechnology, 19, 763–766.
Schemske DW, Horvitz CC (1984) Variation among floral visitorsin pollination: a precondition for mutualism specialization.Science, 225, 519–521.
Selosse MA, Weiß M, Jany JL, Tiller A (2002) Communities andpopulations of sebacinoid basidiomycetes associated with the

2404 J . T . O T E R O , J . D . A C K E R M A N and P . B A Y M A N
© 2004 Blackwell Publishing Ltd, Molecular Ecology, 13, 2393–2404
achlorophyllous orchid Neottia nidus-avis (L.) L.C.M. Rich. andneighbouring tree ectomycorrhizae. Molecular Ecology, 11, 1831–1844.
Smith S (1966) Physiology and ecology of orchid mycorrhizal fungiwith reference to seedling nutrition. New Phytologist, 65, 488–499.
Smith S (1967) Carbohydrate translocation in orchid mycorrhizae.New Phytologist, 67, 371–378.
Smreciu EA, Currah RS (1989) Symbiotic germination of seeds ofterrestrial orchids of North America and Europe. Lindleyana, 4,6–15.
Sneh B, Burpee L, Ogoshi A (1991) Identification of Rhizoctonia Species.American Phytopathological Society, St Paul MN.
Spoerl E (1948) Amino acids as source of nitrogen for orchidembryos. American Journal of Botany, 35, 88–95.
Stenberg ML, Kane ME (1998) In vitro seed germination and green-house cultivation of Encyclia boothiana var. erthyronioides, anendangered Florida orchid. Lindleyana, 13, 101–112.
Swofford DL (1998) PAUP: Phylogenetic Analysis Using Parsimonyand other methods. Test Version 4.0.0. D54, D59. Laboratory ofMolecular Systematics, Smithsonian Institution, Washington D.C.
Taylor DL, Bruns TD (1997) Independent, specialized invasion ofectomycorrhizal mutualism by two nonphotosynthetic orchids.Proceedings of the National Academy of Sciences of USA, 94, 5410–5415.
Taylor DL, Bruns TD (1999) Population, habitat and geneticcorrelates of mycorrhizal specialization in the ‘cheating’ orchidsCorallorhiza maculata and C. mertensiana. Molecular Ecology, 8,1719–1732.
Taylor DL, Bruns TD, Hodges SA (2004) Evidence for mycorrhizalraces in a cheating orchid. Proceedings of the Royal Society of LondonB, 271, 35–43.
Taylor DL, Bruns TD, Leake JR, Read DJ (2002) Mycorrhizalspecificity and function in myco-heterotrophic plants. In:Mycorrhizal Ecology (eds van der Heijden MGA, Sanders I),pp. 375–414. Springer-Verlag, Berlin.
Taylor DL, Bruns TD, Szaro TM, Hodges SA (2003) Divergence inmycorrhizal specialization within Hexalectris spicata (Orchidaceae),a nonphotosynthetic desert orchid. American Journal of Botany,90, 1168–1179.
Thompson JN (1994) The Coevolutionary Process. University of Chi-cago Press, Chicago.
Thomson JD, Higgins DG, Gibbons TJ (1994) CLUSTAL W: improvingthe sensitivity of progressive multiple sequence alignment throughsequence weighting, position-specific gap penalties and weightmatrix choice. Nucleic Acids Research, 22, 4673–4680.
Timms R, Read AF (1999) What makes a specialist special? Trendsin Ecology and Evolution, 14, 333–334.
Tremblay R, Ackerman JD (2001) Gene flow and effective popula-tion size in Lepanthes (Orchidaceae): a case for genetic drift.Biology Journal of the Linnaean Society, 72, 47–62.
Vilgalys R, Cubeta MA (1994) Molecular systematics and popula-tion biology of Rhizoctonia. Annual Review of Phytopathology, 32,135–155.
Vilgalys R, González D (1990) Ribosomal DNA restriction fragmentlength polymorphisms in Rhizoctonia solani. Phytopathology, 80,151–158.
Vujanovic V, St-Arnaud M, Barabé D, Thibeault G (2000) Viabilitytesting of orchid seed and promotion of coloration and germi-nation. Annals of Botany, 86, 79–86.
Warcup JH, Talbot PHB (1966) Perfect states of some Rhizoctonia.Transaction of the British Mycological Society, 49, 427–435.
Warcup JH, Talbot PHB (1971) Perfect states of Rhizoctonia associ-ated with orchids III. New Phytologist, 70, 35–40.
Waser NM, Chittka L, Price MV, Williams NM, Ollerton J (1996)Generalization in pollination systems, and why it matters.Ecology, 77, 1043–1060.
White TJ, Bruns T, Lee S, Taylor JW (1990) Amplification anddirect sequencing of fungal ribosomal RNA genes for phylo-genetics. In: PCR Protocols: a Guide to Methods and Applications(eds Innis MA, Gelfand DH, Snisnsky JJ, White TJ), pp. 315–322.Academic Press, San Diego CA.
Williams NH, Chase MW, Fulcher T, Whitten WM (2001) Molecularsystematics of the Oncidiinae based on evidence from four DNAsequence regions: expanded circumscriptions of Cyrtochilum,Erycina, Otoglossum, and Trichocentrum and a new genus(Orchidaceae). Lindleyana, 16, 113–139.
Zar JH (1999) Biostatistical Analysis, 4th edn. Prentice Hall, UpperSaddle River, NJ.
Zettler LW, Burkhead JC, Marshall JA (1999) Use of a mycorrhizalfungus from Epidendrum conoseum to germinate seed of Encycliatampensis in vitro. Lindleyana, 14, 102–105.
Dr. Tupac Otero was graduated fro Puerto Rico Universitystudding mycorrhizal interaction of orchids, actually he is a post-doctoral fellow at CSIRO Plant Industry in Australia studdingmycorrhizal interactions of Pterostylidinae orchids. Dr. James D.Ackerman is Professor at University of Puerto Rico and Curator ofthe UPRRP Herbarium. He is interested on plant reproductivebiology, pollination ecology, plant systematics, Epiphyte Biologyand Orchidology. Dr. Paul Bayman is Associate Professor atUniversity of Puerto Rico. He is studding orchid mycorrhizalfungi of tropical orchids with focus on Vanilla Spp.





























