Embed Size (px)
Citation preview

UKV '^'
Р О С С И Й С К А Я А К А Д Е М И Я НАУК БОТАНИЧЕСКИЙ ИНСТИТУТ им. В. Л. КОМАРОВА
УДК 582.594.2 : 502.753 Па правах рукописи
Н А З А Р О В ВЛАДИМИР ВИКТОРОВИЧ
РЕПРОДУКТИВНАЯ БИОЛОГИЯ ОРХИДНЫХ КРЫМА
Специальность: 03.00.05 — Б о т а н и к а
А В Т О Р Е Ф Е Р А Т диссертации на соискание ученой степени
кандидата биологических наук
САНКТ-ПЕТЕРБУРГ 1 9 9 5

Работа выполиииа в Симферопольском государствеипом университете имени М. В. Фруизе и в Ботаническом инсти
туте имени В. Л. Комарова РАН.
Н а у ч н ы й р у к о в о д и т е л ь :
профессор, доктор биологических паук В. Г. М И Ш Н Ё В .
Н а у ч н ы й к о н с у л ь т а н т :
засл. деятель науки РФ, профессор, доктор биологических паук Т. Б. БАТЫГИНА.
О ф и ц и а л ь н ы е о п п о н е н т ы : профессор, доктор биологических наук Л. В. АВЕРЬЯ
НОВ; доцент, кандидат биологических наук М. П. БАРАНОВ.
В е д у щ е е у ч р е ж д е н и е — Всероссийский институт растениеводства им. Н. И. Вавилова.
Защита состоится « » апреля 1995 года в /Г часов на заседании специализированного совета К.002.46.01 по присуждению ученой степени кандидата биологических наук при Ботаническом институте имени В. Л. Комарова РАН по адресу: 197376, Санкт-Петербург, ул. проф. Попова, 2, БИН, зал Ученого совета.
С диссертацией можно ознакомиться в библиотеке Ботанического института им. В. Л. Комарова РАН.
Отзывы на автореферат просьба присылать в двух экземплярах по адресу специализированного совета.
Автореферат разослан ч. о i> марта 1995 г.
VM. Ученый секретарь специализированного совета \^Х,Л|ЛМ О- С. ЮДИНА

- 1 -ОВаДЯ ХАРАКТЕРИСПИКА РАБОТЫ
Актуальность темы Все возрастающее антропогенное влияние на природные экосистекы остро, ставит проблему сохранения бкологичес-гсого разнообразия. На грани исчезновения сегодня находятся уяе не отдельные виды, а целке роды и дата семейства растений. Крупней-lee семейство цветковых растений - Orchldaceae целиком включено 30 II Прилохекие Конвенции по международной торговле видами дикой фауны и флоры, что находятся пол угрозой исчезновения. Ьйогие виды включены в Красные книги УССР (1980) и СССР (19843.
Орхидные издавна были об'ъектом массового сбора Все представители семейства орхидных высоко декоративны Многие виды являются цекней!глм лекарственным сырьем (Ломагина, 11Ьгова,1994). В силу особенностей репродуктивной биологии чис.тенность природных популяций орхидных не высока. Поэтому целенаправленный сбор наносит сильный урон популяциям этих растений. Тесная зависимость семенного воспроизведения и возобновления в популяциях орхидных от наличия в биоценозах специфических опылителей и микоризных грибов делает эти растения оссбекко чувствительным-и к нарупениям слошв-сихся консортизных связей. Выпадение из биоценоза хотя бы одного из этих консортов сводит на нет репродуктивное усилие растений и неизбежно ведет к исчезновению данной популяции орхидных.
Горный Крым является вторы-м центром видового разнообразия орхидных ка территории бывшего СССР. Его флора сегодня насчитывает 44 вида орхидей. Ситуация в Кры>лу с орхидными в настоящее время складывается крайне неблагоприятно. В результате увлечения нетрадиционными методами лечения резко возрос сбор клубней. Массовое выделение участков под застройку без соотвествующей экологической экспертизы привело к уничтожению сотен популяций многих видов.
Успешная орга.чизация охраны и воотановление численности популяций орхидных в Крыму невозможны без всестороннего комплексного исследования репродука'ивной биологии этих растений. Вместе с тем о биологии и экологии размножения орхидных Крыма известно крайне кало, что и побудило нас к проведению настоящего исследования.
Цель и задачи исследования. Пелью настоящей работы явилась структурно-функциональное и количественное исследование основных этапов семенного размножения, воспроизведения и возобновления в популяциях у некоторых Крымских орхидей.
В ходе исследования предстояло решить следующие задачи:

- 2 -- установить потенциальную и реальную семенную продуктивность
цветка, генеративного побега и особи в целом; - установить продуктивность андроцея; - дать кол;1чественную оценку сни.тения уровня семенной продук
тивности 3 ходе цветения, при опылении и созревании се.мян; - установить видовой состав опылителей, способы их привлечения
цветками и результативность опыления ими отдельных цветков и соцветий;
- установить видовой состав фитофагов и степень их влияния на семенную продуктивность орхидей;
- изучить характер диссеминации на примере отдельных видов; - изучить в популяциях некоторых видов эффективность возобнов
ления. Научная новизна работа Впервые разработаны методики, псзволя-
юш'.е непосредственно определять число семязачатков (или их при-мордиев) в завязи, число зрелых неповрекденкых семян в коробочке и число пыльцевых зерен в поллинии. Методики по опредению первых двух показателей защищены авторским свидетельством N 1476506 с приоритетом от 26 октября 1987 г.
Впервые установлена потенциальная семенная продуктивность (ПСП) цветка, генеративного побега и особи у 29 видов орхидей Крыма. Реальная семенная продуктивность (РСП) плода, генеративного побега и особи установлены у 17 видов. Впервые определено число пыльцевых зерен в андроцее у 11 видов.
Приводится ранее не изученный состав опылителей для 10 видов крымских орхидей. Изучены особенности привлечения опылителей в Крыму. Впервые описан способ привлечения опылителей цветка\м Steveniella satyrioides. Сушественно дополнены сведения о способе привлечения опылителей цветками Cephalanthera rubra, Anacamptis pyramidalis, Dactylorhlza incarnata, D. romana
Впервые установлен видовой состав насекомых-фитофагов крымских орхидей и определена роль вредителей в сниягении се.менной продуктивности.
Теоретическая и практическая ценность работи Разработанные для орхидей Крыма методики определения се.менно*. продуктивности, продуктивности пыльцы открывают новую возможность в изучении семенной продуктивности и эффективности опыления и у тропических представителей орхидей. В настоящее время эти методики опробиро-заны еше на 24 европейских и 7 тропических видах в дополнение к

- 3 -вида!.!, приведенным в диссертации (неопубликованные данные). Эти методик;', пригодны и для изучения семенной продуктивности у других видов растений с ме.ткими семена.м;!. В частности, хорошие результаты пол^^:екы при изучении некоторых видов Orcbanchaceae и Scrophulariaceae (неопубликова.чные дачные), хозяйственная значимость которых не вызывает сомнений.
^?иое теоретическое значение имеют данные о ПСП цветка, отко-гении числа пыльцевых зерен к числу семязачатков в цветке, степени диффереяцированнссти семлзачатка во время распускгшия цветка. Зти показатели тесно связаны со многими морфологическими призна-кл.ми, которые лез«ат в основе филогенетической системы орхидных.
Данные о семенной продуктивности и количестве пыльцы в андро-цее обработаны статистически, что позволяет применять их при проведении исс.тедований в об.1аоти репродуктивной биологии и в других частях ареала изученых видов орхидей. Разработанный способ оценки ?СП п.тада яо его размерам позволяет достаточно точно определять семенную продуктивность, не прибегая к вскрытию коробочек. Этот метод оценки семенной продуктивности незаменим при изучении дио-сеьжнации и проведении мониторинга попу.тяиий исчезаюшх видов.
Апробация работы Основные теоретические пста^ения и результаты настоящей работы докладывались на Всесоюзном совещании по охране и культивированию орхидей (Москва,1987). наXI Международном симпоэиу!.!е по эмбриологии и семенной продуктивности (Санкт-Петербург, 1990) и на семинаре по эмбриологии и репродуктивной биологии 1989.1995 гг. (БИН АН РОССИЯ, Санкт-Петербург). По теме диссертации опубликовано 16 работ и две находятся в печати.
Объем работы Диссертация изложена на 123 страницах машинописного текста и состоит из введения, 7 глав и выводов. Список литературы содержит 45 работ отечественных и 179 - зарубе.чных авторов. Приложение включает 76 страниц машинописного текста (3 главы) 10 таблиц и 18 рисунков.
ГЛАВА I. ОСОБЕННОСТИ РЕПРОДУКТИВНОЯ БИОЛОГШ сем. ORCHIDACEAE (по данным литературы)
По числу семян в отдельном плоде орхидные являются бесспорными лидерами среди цветковых растений. В каждой коробочке у них со-дер.жится от 400 до 6000000 семян (Ard i t t i , 1967,1992). Еще выше репродуктивная способность у генеративного победа (Salisbury,

- 4 -1942; Работнов.1950; Терёхин,1977).
Эта особенность репродуктивной биологии орхидных неоднократно обсузкдалась исследователями в связи с различными аспектами биологии и филогении • семейства (Darwin,1862; Burgeff,1935 Salisbury. 1942; van der Fiji. Dodscn, 1966; Arditti. 1957.1992 Терёхин.1977; Dressier. • 1981,1993; Robinzon. Eurr£-Balogh,1982 Benzig, Atvood, 1984; Benyig, 1987; Аверьянов. 1991 a и др.). Хотя перечисленные выше авторы иногда придержвагася противоположных взглядов по отдельным вопросам, все они сходятся во мнении. что высокая семенная продуктивность у орхидных коррелирует с ме.т-кими (пылевидными) семенами, 'отсутствием в них эндосперма и редукцией зародышей. Очевидна связь семенной прсду)':тквности орхидных и с агрегацией их пыльцы в особые ср1'ктуры - полл1:кии (Darwin, 1862; van der Fiji, 1966; van der Fiji, Dodson, 1965).
Несмотря на бесспорную з.чачимосгь данных о семенной продуктивности для изучения репродуктивной биологии, систематики и филогении орхидных эта сторона жизни этих растений изучена крайне слабо.
ГЛАВА 2. МАТЕРИАЛ К МЕТОДЫ ИССЛЕЛОВАНИ.Ч
7i2 44 ВИДОВ орхидей Крыма исследованы 29. Они представляют главнейше роды, экологические и биоморфологические группы крьмс-ких орхидей, относявдхся к самым различным флорогенетическим зле-MeHTairf (табл. 1,2). Иссследования проводились на территории Горного Крыма в.течении 1982-1994 гг.
Семенную продуктивность цветка и продуктивность андроцея определяли по специально разработанным методикам. Морфологические измерения в зависимости от величины объекта проводили при помощи штангенциркуля, мерной линейки бинокулярного микроскопа КЕС-10 и ЫБИ-15. Для измерения объема коробочек был сконструирован прибор, который позволял вести измерения с точностью до 0.02 куб. мм. Щзоективную площадь семенной кожуры определяли на масштабных фотографиях при помощи планиметра.
Строение семени изучалось на постоянных препаратах, изготовленных по общепринятой цитозмбриологической методике (Пауше-ва.1974). Материл фиксировали в смеси FAA состава: 70%-ный спирт, 40%-ный формалин и ледяная уксусная кислота в пропорции 10: 3:1. Толгина срезов соста:.ляла 12 мкм. Щ>алараты окрашивались метилг-
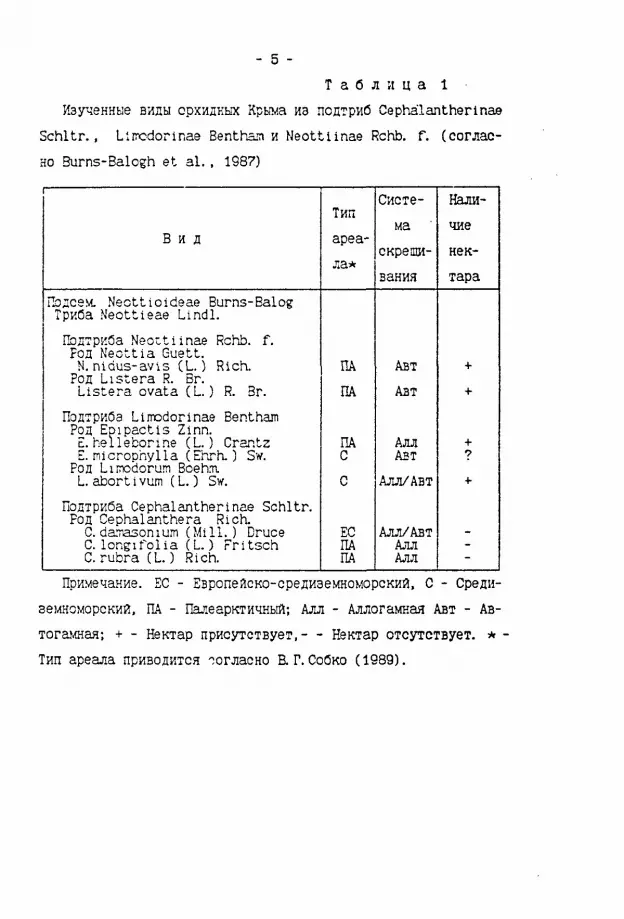
- 5 -Т а б л и ц а 1
Изученные виды орхидных Крыма из псдтриб Cephalantherinae Schltr., Llmcdorinae Benthan и Neottlinae Rchb. f. (согласно Burns-Balogh et al. . 1987) 1
Тип Систе Налима чие В и д ареа
ла* скрещи неквания тара
ГЬдсем. Necttloideae Burns-Balog Триба Neottieae Lindl. ПЬдтриба Neottlinae Rchb. f. Род Neottia Guett. N. nidus-avis (L.) Rich. ПА Авт + Род Listera R. Br. Listera ovata (L ) R. Br. ПА Авт +
ПодтркОа Liirodorinae Bentham Род Ер1 pact is Zinn. E. hellebori.ne (L ) Crantz Род Ер1 pact is Zinn. E. hellebori.ne (L ) Crantz ПА Алл + E. microphylla (Ehrh.) Sw. С Авт 7 Род Limodorum Boehm. L. abort 1 vum (L.) Sw. С Алл/Авт +
Подтриба Cephalantherinae Schltr. Род Cephalanthera Rich. С darrasonium (Mill. ) Druce ЕС Алл/Авт -С longifolia (L.) Fritsch ПА Алл -С rubra (L.) Rich. ПА Алл -
Примечание. ЕС - Европейско-средиэемноморский. С - Средиземноморский. ПА - Палеарктичный; Алл - Аллогамкая Авт - Автогамная-, + - Нектар присутствует,— Нектар отсутствует. * Тип ареала приводится ';огласно R Г. Собко (1989).

- 6 -Т а б л и ц а 2
Изученные виды орхидных Крыма иэ подтрибы Orchidlnae Dressier (сс-ласно Dressier. 1933)
Тип Систе Налима чие В и д ареа
ла* скрещи неквания тара
Гюдсем. Orchidoideae Dressier Триба Orchidoeae Dressier Псдтриба Orchidlnae Dressier ?сд Anaca-nptis Rich. A. Dyra-midaiis (L.) R1 ch. EC Алл - (+) Род Ccrperia C.Koch C. ccnperiana (Stev.) Aschers. et С Алл -Graebn. Род Dactylorhiza Nevski D. 1 ncarrata (L.) Soo ПА Алл -D. rorrana (Seb. et Mauri) Soo С Алл -Род Gymadenia R. Br. G. conopsea (L.) R. Br. Г Алл + Род Kirantoflossum Koch H. caprinum (Bleb.) Koch С Алл -Род Ophrys L. 0. oestrifera Bleb. с Алл -0. taurica (Agg.) Nevski эк Алл -Род Orchis L. 0. corlcDhora L. ЕС Алл + 3. rrascula (L.) L. ЕС Алл -0. nilitaris L ПА Алл -0. picta Loisel. С Алл -0. provmc talis Balb. С Алл -0. pur.ctulata Stev. ex Lindl. СП Алл -0. purpurea Huds. с Алл -0. SI mi a Lani ЕС Алл -0.tridentata Scop. С Алл -Род Platanthera Rich. P. bifolia (L.) Rich. ПА Алл + .P. chlorantha (Gust.) Reichenb. ЕС Алл + Род Stevemella Schlechter
i S. satyrioides (Stev.) Schlechter СП Алл -Примечание. Г - Голарктичный, СП - Средиземноморско-переднеа-
зиатский, ЭК - Зндем Крыма. Остальные условные обозначения приведены а таблице 1.

- 7 -рюяпирокнном с подкраской алциансвым синим.
Степень дифференциации семязачатка на момент распускания цветка изучалась ка давленных препаратах, изготовленных с применением просвет.таюЕэй методики Герра (Джори,Амб9гаоклр,1990).
Видовой состав опьиителей устанавливался в ходе наблюдений за цветением орхидей. Все насекомые, посекавсие цветки, отлавливались для ясследуюшего определения и морфометрии. Параллельно проводился отлов насекомых с пыльцой орхидей на окружающих растениях. Насекомое считалось истинным опылителем лишь при нахолденки на нем пол.такиез или поллинариев и соответствии морфологии насекомого морфологии цветка.
Л;1на>/ика цветения и опыления изучалась на этикетированных особях в ходе периодических наблюдений. При этом у каждого цветка помимо трех фенологических фаз - бутонизащги, цветения и завяда-ния определял;! наличие и.чи отсутсвке гемиполлинаркев в гнездах пыльника и массул на рыльце.
Семена в ходе изучения ближней диссе!Д!нации у 0. purpurea улавливались при помощи клейких полос.
Статистяческля оценка .показателей семенной продуктивности, а тгкув других изученных параметров производилась согласно методике ;L аВайкагия (1974).
РЕЗУЛЬТАТЫ ИССЛЕДСШЮШ
ГЛАЗк 3 . П0ТЕНШ1АЛЬНАЯ СЕМЕННАЯ ПРОДУКТИБНОСТЬ ( Ш П )
Число семязачатков в завязи (ПСП цветка). ПСП цветка оценивали по числу примордиев семязачатков в завязи во время распускания цветка. Этот показатель определен у 586 завязей 29 видов, относя-щихояся к 13 родам из четырех подтриб. Среднее значение 1ЮП цветка по некоторым изученным видам приведено на табл. 3.
ПСП цветка и его фитомасса Фитомасса цветка жестко связана с его, ПСП. Корреляция между этими характеристиками цветка обнаружена на уровне видов не только в пределах отдельных подтриб (г-0.76 для Orchidinae), но и среди всех изученных видов (г-0.56). Интересно отметить, что корреляция ПСП о фитомассой згшязи оказалась слабее, чем о общей фитомассой цветка. Вероятно, •это обусловлено

- 8 -тем. что у MHorta видов завязи и семязачатки сильно недоразвиты ко времени распускания цветка. Нормального развития они достигают только через несколько недель после опыления.
У Orcnis, Ophrys, Dactylorhiza ка единицу фитсмассы колонки и цветка приходится гораздо больше примордиев семязачатков, чем у видов Listera и Epipactis. Увеличение этого показателя, вероятно, происходило по мере усиления руминированности плаценты и задердаи в развитии семязачатков.
Из-за сильных различий между видами в степени р^тйширсвакности плаценты и д/.фференцированности семязачатков во время распускания цветка связь между ПСП цветка и длиной завязи на уровне видов не улавливается, хотя эти показатели .ггст'ко связаны меу^у собой .в пределах отдельного генеративного побега.
ПСП цветка и пдодадь рыльца. Установлена тесная связь между ГЮП цветка и площадью зосприкимаюагей поверхности рыльца. У восьми видов ка популяционном уровне' коэффициент корреляции между ШШ цветка к площадью рыльца колебался от 0.77 до 0.90 Эта связь, вероятно, опосредована через количество пыльцы, поступающей на рыльце. Хорошей иллюстрацией этому служит симпатрические виды -Platanthera chlorantha и Р. bi fol ia Рыльце у первого вида вследствие расхождения прилипалец занимает на колонке значительно боль-Ею плошадь, чем у второго. Зто, вероятно, и обусловило разницу между этими орхидными в ШП цветка примерно в 1700 семязачатков. (табл. 3).
Связь между ПСП цветка и ПЛОЕЗДЬЮ рыльца улавливается и на видовом уровне в пределах отдельных подтриб (г-0.47 для видов Orchidinae). Однако, на эту зависимость оказывают влияние и другие факторы, у видов подтрибы Orchidinae на отношение числа семяпочек в завязи к плокади рыльца существенное влияние оказывает размер массул. Например, у Ophrys oestrifera на 1 кз. мм плошадк рыльца приходится 5463 семязачатка, тогда как у Orchis pieta -только 2126. Это обусловлено скорее всего тем, что у 0. oestrifera апикальные массулы значительно крупнее и содержат в 3-3.5 раза больше пьАяьцы. чем у О. picta С увеличением размера пыльцевого агрегата на единице площади рыльца размеоается гораздо больше пыльцы. Зта закономерность отмечена и у представителей из других семейств (например, сем. Asclepiadaceae) (Cruden,1977).
На отношение числа семяпочек к площади рыльца у ' видов Orchidinae оказывают также влияние и число гемиполлинариев ка на-
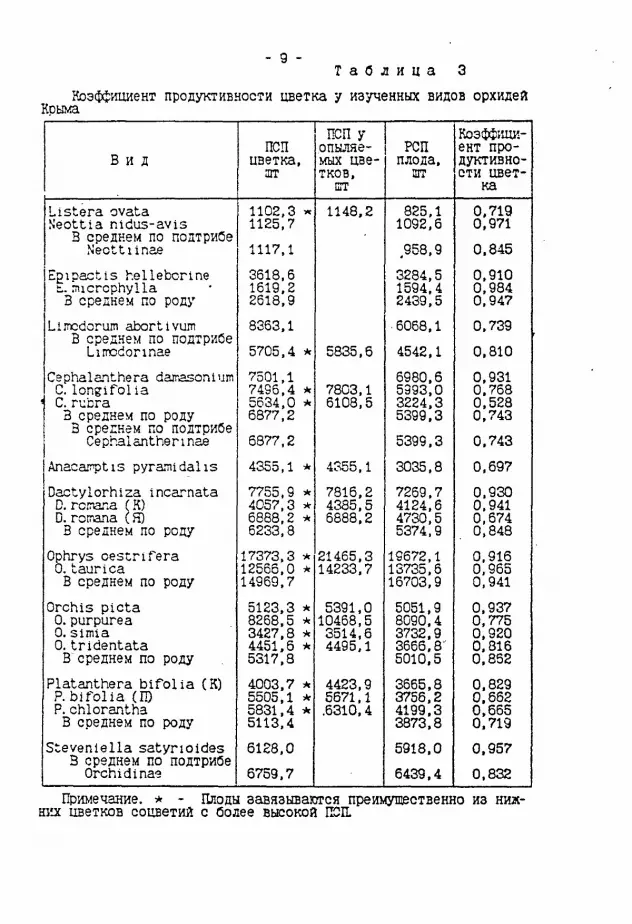
Т а б л и ц а Коэффициент продуктивности цветка у изученных видов орхидей
Крыма
В и д ПСП
цветка, агг
ПСП у опыляемых цветков. шт
РСП плода, шт
Коэффициент продуктивности цвет
ка Listera ovata Neottta nidus-avis
В среднем по подтрибе N'eottiinae
1102,3 ^ 1125.7 1117,1
1148.2 825.1 1092,6 ,958.9
0.719 0,971 0.845
Egipactts hellebcrine ь. microphylla 3 среднем по роду
3618,6 1619.2 2618,9
3284,5 1594.4 2439.5
0,910 0.984 0.947 Limcdorum abortivum
В среднем по подтрибе Limodorinae
8363.1 5705,4 * 5835.6
6068.1 4542.1
0,739 0.810
Cephalanthera darasonium С.longifolia С. rubra 3 среднем по роду 3 среднем по подтрибе Cephalanthennae
7501.1 7496.4 * 5634,0 * 6877,2 6877,2
7803,1 6108,5
6980,6 5993,0 3224,3 5399.3 5399,3
0.931 0.768 0,528 0.743 0.743
Anaca'nptiS pyraiiidalis 4355,1 * 4355,1 3035,8 0.697 Dactylorhiza incarnata D. rcrana (K) D. romana (Я) В среднем по роду
7755,9 * 4057,3 * 6888,2 * 6233,В
7816,2 4385,5 6888.2
7269.7 4124.6 4730,5 5374,9
0.930 0.941 0,674 0.848
Ophrys oestrlfera 0. taurica В среднем по роду
17373.3 * 12566,0 * 14969,7
21465,3 14233,7 19672,1 13735,6 16703,9
0,916 0,965 0,941 Orchis picta 0.purpurea 0.Simla 0. tridentata B'среднем no роду
5123.3 * 8268.5 * 3427.8 * 4451.6 * 5317,8
5391,0 10468,5 3514,6 4495.1
5051,9 8090.4 3732.9 3666,8' 5010.5
0.937 0,775 0.920 0.816 0.862
Platanthera bifolia (K) P. Ыfolia (П) P.chlorantha В среднем по роду
4003.7 А 5505,1 * 5831,4 * 5113,4
4423,9 5671.1 .6310,4
3665,8 3756,2 4199.3 3873.8
0,829 0.662 0.665 0.719 Stevenlella satyrioides 3 среднем по подтрибе Orchidinae
6128,0 6759.7
5918,0 6439.4
0.957 0.832
Примечание. * - Плоды завязываются преимущественно из ниж-ш'л цветков соцветий с более высокой ШШ.

- 10 -секомых-переносчиках и мор(|ологическое разннсбразие опылителей. что подробнее рассматривается в основном тексте диссертации. Отметим лишь то, что при низком числе гемиполлинариев на спьиителе и зысскоЯ степени морфологического соотвествия опылителей цветку число семяпочек ка единицу рыльца возрастает.
ПС31 генеративного побега Этот показатель семенной продуктивности рассчитывался исходя из среднего числа семязачатков в завязи (ПСП цветка) и среднего числа цветков на генеративном побеге. Среднее значение числа семязачаток на генеративном побеге (ПСП генеративного побега) по некоторым изученном видам приведено на табл. 4.
Число цветков на соцветии и способ привлечения опылителей. Число цветков на генеративном побеге очень изменчиво. У бо.тьаинс-тва видов коэффициент вариации по этому признаку оказался больсе, чем в отношении ПСП цветка. Однако при сразкекии средних значений числа цветков на генеративном побеге метеу Еида.ми выявляется четкая таксономическая специфичность этого показателя. Число цветков, наряду с плотностью их расположения на оси соцветия и окр^-кой венчика, придает соцветию характерный вке!лкий облик. Число цветков у изученных орхидей сбнаруиявает' отчетливЗ'Тс связь со способом привлечения опылителей. Яркий пример тому виды Cphrys. Кх ма.10- и редкоцветковые соцветия с последовательным распусканием к завяданием цветков точно соотвествуют способу привлечения саул;ов одиночных пчел за счет им.митации их самок.
Организация соцветий у безнектарных Dactylcriza rcra.ia. Compena comperiana, Orchis rascula, 0. provincial is , 0. рюгаи видов Cephalanthera, Anaca^ptis pyramidal is сходна с соцветиями нектаросодер.жашкх модельных растений из других семейств. Эта зависимость от.мечалась ранее и для других видов орхидей (Dafni, Ivri,1981; Da^ni,1987).
Соцветия у некоторых безнектарных видов рода Orchis (0. tridentata, 0. mili taris, ©.purpurea, 0. punctulata) и Dactylo-rhiza incarnata эволюционировали в направлении увеличения числа цветков в соцветии, вероятно, и под воздействием способа привлечения опылите.-:зй. Эти орхидеи имеют многоцветковые плотные соцветия на высоком цветоносе. Как установлено JL Нильссоном (Nilsson,1980), такой тип соцветий характерен для видов, которые опыляются главным образом "неопытными" насекомыми. Эти насекомые привлекаются цветкам;: орхвдей главным образом визуальными атрак-
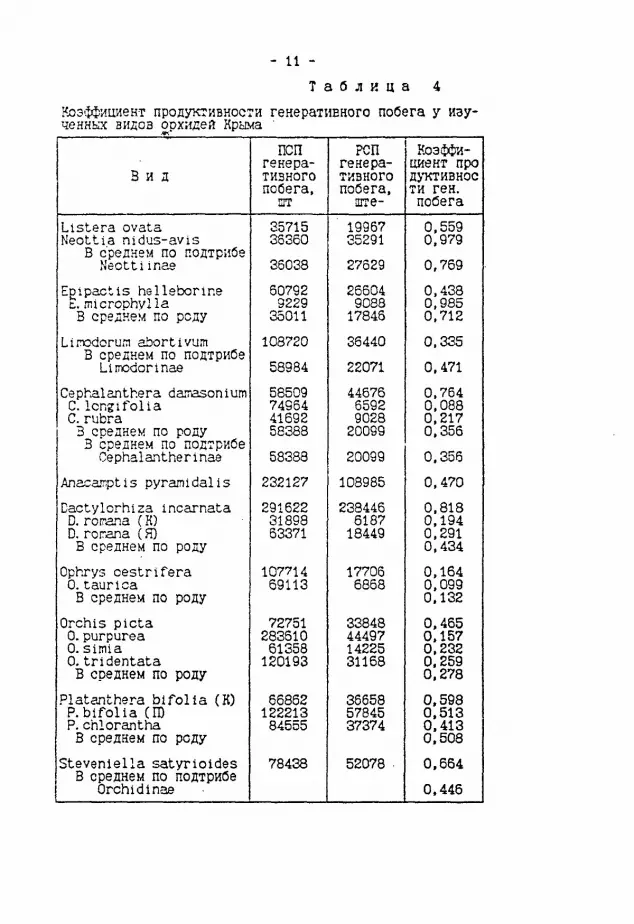
- 11 -Т а б л и ц а 4
Коэ'ффициент продуктивности генеративного побега у изученных видов орхидей Крыма '
В и д ПСП
генеративного побега, зт
РСП генеративного побега.
ште-
Коэффициент про дуктивнос ти ген. побега Llstera ovata Neottia nidus-avis
В среднем по подтрибе Neotti inae
35715 36360 36038
19967 35291 27629
0.559 0.979 0,769
Enipactis helleborine L. rracrophvlla В среднем по роду
60792 9229 35011
26604 9088 17845 0.438 0,985 0,712
Linodorum abortivum В среднем по подтрибе LiiTWdorinae
108720 58984
36440 22071
0.335 0,471
Cephalanthera daTasonlum С. long i foil a С rubra В соеднем по роду 3 среднем по подтрибе Cephalantherinae
58509 74964 41692 58388 58383
44676 6592 9028 20099 20099
0,764 0,088 0.217 0.356 0,356
Anacamptis pyramidal is 232127 108985 0.470 Dactylorhiza incarnata D. rorrana (Ю D. rorana (Я) В среднем по роду
291622 31898 63371 238446 6187 18449
0.818 0.194 0.291 0,434
Ophxys cestrifera 0. taurica В среднем по роду
107714 69113
17706 6868
0.164 0.099 0.132
Orchis picta 0.purpurea 0. simia 0.tridentata В среднем по роду
72751 283610 61358 120193
33848 44497 14225 31168
0.465 0.157 0,232 0.259 0,278 Platanthera bifolla (К) P. bifolia (ID P.chlorantha В среднем по роду
66862 122213 84555
36658 57845 37374
0,598 0.513 0.413 0.508 Stevenlella satyrioides В среднем по подтрибе
Orchidinae 78438 52078 0.664
0.446

- 12 -тактами. Процент опыленных цветков на соцветии у этих видов коррелирует с величиной соцветий и числом цветков на оси соцветия.
Число цветков на соцветии и ПСП цветка. Несмотря на сильную изменчивость как числа цветков на соцветии, так и ПСП цветка, ПСП генеративного побега у изученных видов колеблется в относительно узких пределах - от 40 до 120 тысяч. Это обусловлено тем. что в силу ограниченности ресурсов, которые особь может затратить на репродукцию, у большинства изученных видов число цветков на генеративном побеге находится в обратной зависимости от ПСП цветка. Только у Epipactia mlcrophylla низкие значения ПСП сочета.ются с малоцветковостью соцветий, а у Dactylorhiza incarnata и Hirrantog-Icssum caprinur, наблюдаются .относительно высокие значения обоих показателей. В результате ПСП генеративного побега значительно отклоняется от указанных выше пределов (табл. 4).
ПСЯ1 особи. У изученных видов подтрибы Orchidinae ПСП особи за один вегетационный период тожественно ПСП генеративного побега. У видов из других изученных псдтриб особи могут образовывать несколько генеративных побегов в течении одного вегетационного периода. Наиболее сильно это вырахеко у видов Neottiinae.
ГЛАВА 4. ПРОДУКТИВНОСТЬ^АНДРОЦЕЯ (ПА)
Изученные виды принадлежат к однстычинксвыу. орхидеям. Продуктивность андроцея у_ них соотвествует продуктивности пыльника единственной ферти^ньной тычинки. Вся пыль'ца этой тычинки агреги-рованна в четьре поллиния. У видов подтрибы Orchidmae, чьи no.v линии из одной теки объединены попарно в гемиполлинарий и состоят из массул, число пыльцевых зерен в пьшьнике (П. ) расчитывалось исходя КЗ средних значений количества пыльцы в одной массуле к количетва массул в поллиниях одного гемиполлинария. У видов из остальных подтриб, имеюшх неструктурированные поллинии, ПА определяли непосредственно'- по' числу пыльцевых зерен в поллинии.
Чз1слЬ пыльцевьк зерен в массуле. Этот показатель определен у 14 видов из подтрибы Orchidinae. При этом изучено 442 массулы. Пределы изме.ччизости и средние значения этого показателя приведены на табл. 5.
Степень дифференциации массул в полдинии. У изученных видов подтрибы Orchidinae массулы одного поллиния сильно различаются по размерам. Эти различи., обусловлены числом пыльцы в них. Пыльцевые

Число пыльцевых зёрен в андроцее и П/С-отношение Т а б л и
у видов подтриО
В и д Число пыльцевых зёрен в массуле,
шт
Число массул в геми-поллинарии. шт
Число пыльцевых зерен в гемнпол-линарии, шт
Числ пыль вых рен андр цее шт
В и д
min-max X
Число массул в геми-поллинарии. шт
Число пыльцевых зерен в гемнпол-линарии, шт
Числ пыль вых рен андр цее шт Anacamptls pyramidal is 62- 501 237,4 124,3 29509 5901 Comperla comperlana 518-2593 1311,8 137,8 180766 3615 Dactylorhlza Incarnata 71- 723 423,0 127,5 53933 1078 Ophrys oestrifera 123-3476 1265,6 • 99,1 125421 2608 0. taurioa 98-2347 1082,9 87,3 94537 1890
Orchis corlophora 61- 442 223.8 111,2 24887 497 0. mllltarls 102- 803 516,3 113,9 58807 1176 0. plcta 60-1008 503,1 113,8 57253 1145 0.provincialIs 68-1303 477,8 123,2 80758 1615 0. purpurea 49-1882 621,3 131,7 81825 1636 0. Simla 42-1004 382,2 128,3 49036 980 0.tridentata 73-1068 429.4 81,9 35168 703 Platanthera bifolia (П) 53- 307 220.0 292,3 64306 1286 Stevenlella satyrloides 83- 761 318,3 134,1 42684 853

- 14 -зерна в массулё однородны по размеру. Самые крупные массулы с кая большим количеством пыльцы располагаются в апикальной части пол-линия, самые мелкие - в его основании. В степени дифференциации массул отдельного поллиния по числу пыльцевых зерен у изученных видов набл-юдаются сильные колебания. На:'.более отчетливо эта дифференциация выражена у "видов Ophrys. Массулы у Platanthera bifolia и Orchis coriophora более однородны по числу пыльцы в них. Среднее чисто пыльцы в массуле тесно связяяо со степенью дифференциации поллиния на апикальные и базальные массулы (г-0.79).
Степень дифференциации массул в поллинии и частота поседения-цветков одним опылителем. Размер массул и число пьш цевых зерен в поллинии гемиполлинария у видов подтрибы Opchidinae в оначите.-Ц:-ной степени обусловлено частотой посещения цветков одним опылителем. Самцы' Еисега nigra Lep. редко псрецают цветки Cphrys ces tn fe ra К концу цветения они оставляют на рыльце менее половины массул- поллиния. Это обусловлено способом привлечения опылителей и в.ызвало сильное ук-рупнекие апикальных массул.
Самые мелкде массулы с низким числом пыльцевых зерен в них обнаружены у Platanthera bifolia и Orchis coriophora Эти кектаркые орхидеи посещаются опылителями значительно чаще, чем виды Ophrys. При таком способе привлечения опылителей вероятность попадания базальных улссул на рыльце значительно высе, чем у Cp!-j-ys..
Число массул в поллинии Самое большое чис.10 массул в гемипо линарии отмечено у Platanthera bifolia, которая и.меет самые мелкие массулы и самое низкое количество пыльцы в них. С другой стороны, одно из самых низких значений числа массул в гемипсллинарии отмечены у видов Ophrys, которые имеют самые крупные массулы с наибольшим числом пыльцы з них (табл.5). Однако связь между числом массул в поллинии и средним числом пыльцевых зерен в одной массуле 'у изученных видов перекрывается другими корреляциями.
Продуктивность андроцея. Цродуктивность андроцея у каждого отдельного вида неразрывно связана с ПСП цветка. Отчетливая зависимость мевду этими параметрами прослежена у видов подтрибы Orchidinae. Коэффициент корреляции между ПА и ПСП у них составил 0.83. Он мог бы быть еще выше, но изученные виды значительно раз-•личались между собой по отношению числа пыльцы к числу семяпочек .Б цветке (П/С-отношение) (табл. 5).
П/0-отнотение. П/С и система скрещивания. Известно, что П/С в

- 15 -первую очередь отражает особенности системы скреслвания. Установлено, что у видов с неагрегированнсй пыльцой П/С возрастает в логарифмической зависимости от клейтогашых к ксеногамным видам (Cruden,1977). У изученных орхидных наблюдалась обратная картина у аллогамных видов IVC оказалось ниже, чем у автогамных (табл.5).
П/С и агрегация пыльцы в поллинии. Низкие значения П/С у изученных орхидных обусловлены агрегацией пыльцы. Резкое снижение П/С у видов с агрегированной пыльцой установлено ранее и для представителей семейств Asclepladaceae, Mlmosaceaa и Onagraceae (Cruden, 1977; Cruden, Jensen,1979). Виды Asclepiadaceae имеют почти такие УЗ значения П/С, что и орхидные. Пыльца у представителей обоих семейств агрегирсУвана в поллинии.
П/С и способ восприятия пыльцы рыльцем. На примере изученных видов орхидей, которые имеют "мягкие" поллинии, установлена зависимость П/С-отноЕзния от спобоба восприятия пыльцы рыльцем. У изученных видов орхидных установлено два способа восприятия пыльцы рыльцем - "соскребаю21ий" (когда поллиний оскребается с насеко-лого ростеллумом) и "контактный" (когда поллиний вдавливается на-секомьм в рьшьце) (табл. 6). При "соскребающем" способе восприятия пы.1ьцы рыльцем, который установлен у видов Cephalanthera, на pbLibue остается больная часть пыльцы "мягких" поллиниев. Это обуслозливет низкое значение П/С. Низкое значения П/С (3,8-3,9) установлено ранее и у двух видов Isotria (Mehrhofr,1983). Судя по морфологии колонки этим орхидным так же присущ "соскребающий" способ Ессприяти пылцы.
При "контактном" способе восприятия пыльцы на рыльце попадает лишь небольшая часть пыльцы "мягкого" поллиния. Поэтому значения П/С у видов с таким способом восприятия пыльцы значительно выше, чем у предьщущх видов. Эволюция в направлении совершенствования механизмов "порционного" расходования пыльцы одного поллиния в ходе опыления нескольких цветков привела к разъединению поллиния на массулы у видов подтрибы Orchidinae (Darwin,1862).
П/С и некоторые аспекты филогении орхидных. Как было рассмотрено Bbmie. орхидные в целом сильно отклоняются от правила Крудена из-за агрегации пыльцы в поллинии. Однако у изученных видов из подтриб Neottilnae и Llmodorinae это отклонение не столь велико, как у представителей подтриб Cephalantherinae и Orchidinae. Зна-' чения П/С у видов Neottla и Epipactis, согласно Р. Крудену (Cruden,1977), соотвествуют видам с облигатно аутотогамной систе-
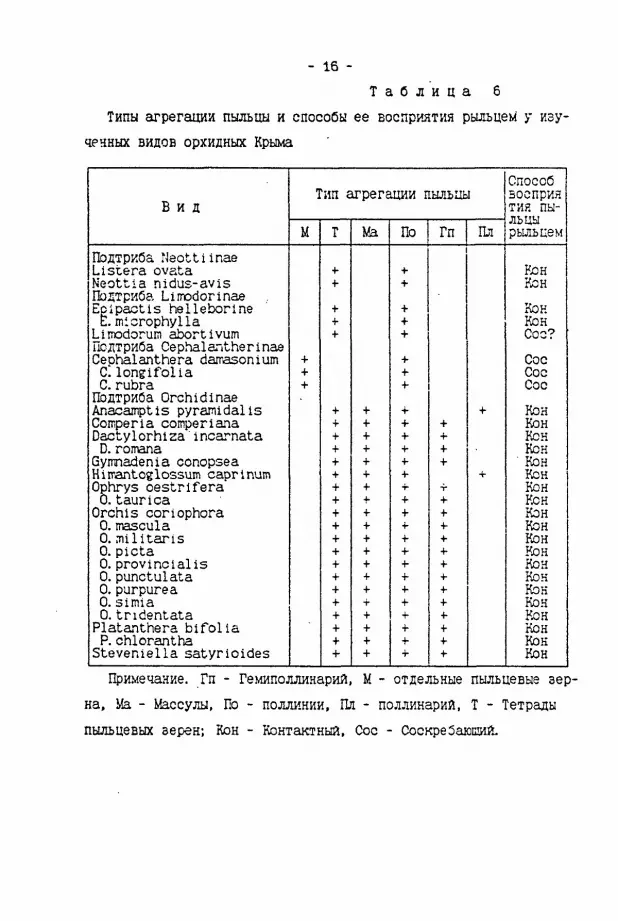
- 16 -Т а б л и ц а 6
Типы агрегации пыльцы и способы ее восприятия рыльцем у изученных видов орхидных Крыма
Способ Тип агрегации пыльцы аосприя
В и д тия пыльцы М т Ma По Гп Пл рыльцем
Подтриба Naottiinae Listera ovata • ( - + Кон Neottia nidus-avis + + Кон Подтриба Limodorinae Eplpactis helleborine + + Кон E. microphylla + + Кон Limodorun abortivum + + Сое? Подтриба Cephalantherinae Cephalanthera dan-asonium + + Сое С lonffifolla + + Сое С rubra + + Сое Подтриба Orchidinae Anacamptis pyramidal is + + •i- + Кон Comperia comperiana Dactylorhiza'incarnata
+ + +
+ + + Кон
Кен D. romana + + + + Кок Gymnadenia conopsea + + + + " Кон Himantoelossum caprlnum + + +• Ч- Кок Ophrys oestrlfera + + +• т- Кок 0. taurlca + 4- + + Кок Orchis coriophora + + + + Кон 0. mascula + + + + Кон 0. milltaris + + + Ч- Кон 0. picta + + + + Кок 0.provincial is + + + + Кон 0.punctulata + + + + Кок 0.purpurea + + + • ( - Кок 0. Simla + Т + 4- Кон 0.tridentata + + + +• Кон Platanthera bifolia + + + + Кон P. chlorantha + + + + Кок Steveniella satyrioides + + + •t- Клн
Примечание. Гп - Гемиполлинарий, М - отдельные пыльцевые зерна, Ь!а - Массулы, По - поллинии, Пл - поллинарий, Т - Тетрады пыльцевых зерен; Кон - Контактный, Сое - СоекреЗаюшй.

- 17 -Т а б л и ц а 7
Связь системы скрепивания и отношгния числа пыльцевых зерен к
числу сеуяпочек в цветке (П/С-отношение) у некоторых видов покры-
тосеуянных с неагрегированной пыльцой (согласно Р.- Крудену (Сги-
den,1977)) и у изученных видов сем. Orchidaceae
П/С-отношение у некоторых видов покрытосемянных с неагрегированной пыльцой (по Cruden, 1Q77)
Тип скревд-вания
П/С-отношение у некоторых видов сем, Orchidaceae
5859,2 (1062-16S07)
1. Ксено-гамные виды Comperla comperlana 39.2 Llstera ovata 31,1
Orchis Simla 28,6 0. purpurea 19,8 0. provincial is 17.0 0. tridentata 15,8 0. niilltaris 14.5 Ophrys taurica 15,0 0. oestrifera . 14,4 Steveniella satyrioldes 13,9
1 Cephalanthera long!foil a 8,4 1 В среднем 19,8
796.6 (161- 2559)
1 1 1 2, Факуль- > тативно 1 ксено-гамные виды
Comperla comperlana 39.2 Llstera ovata 31,1 Orchis Simla 28,6 0. purpurea 19,8 0. provincial is 17.0 0. tridentata 15,8 0. niilltaris 14.5 Ophrys taurica 15,0 0. oestrifera . 14,4 Steveniella satyrioldes 13,9
1 Cephalanthera long!foil a 8,4 1 В среднем 19,8
796.6 (161- 2559)
1 1 1 2, Факуль- > тативно 1 ксено-гамные виды
1 ."Platanthera blfolla 23,4 \ /Orchis picta 22.4 1 ' 0. coriophora 20.7 1 ,'Epipactls helliborine 18,9 I't I Dactylorhiza incarnata 13,9 \i ' Isotria verticulata 3,9* i', / В среднем 17,2
168.5 (32-397)
3. Факуль- \ татиБно > авто- ' гамные виды
1 ."Platanthera blfolla 23,4 \ /Orchis picta 22.4 1 ' 0. coriophora 20.7 1 ,'Epipactls helliborine 18,9 I't I Dactylorhiza incarnata 13,9 \i ' Isotria verticulata 3,9* i', / В среднем 17,2
27.7 (18-39)
4, Об ли- \ г^тно > автогамные виды
1 ."Platanthera blfolla 23,4 \ /Orchis picta 22.4 1 ' 0. coriophora 20.7 1 ,'Epipactls helliborine 18,9 I't I Dactylorhiza incarnata 13,9 \i ' Isotria verticulata 3,9* i', / В среднем 17,2
27.7 (18-39)
4, Об ли- \ г^тно > автогамные виды
ii ( / П 1 /Cpphalanthera damasonlum 7,9 ( \--'/ Isotria medeololdes 3,8* \'- В среднем 5,9
27.7 (18-39)
4, Об ли- \ г^тно > автогамные виды \ /
• J. / Epipactls mlcrophylla 14,3 4,7 (3-7)
5, Кпейсто-гамные виды
\ / • J. / Epipactls mlcrophylla 14,3 4,7
(3-7) 5, Кпейсто-
гамные виды
L./ Примечание. * - Ш данным Л. Мерхоффа (Kfehrhoff,1983).

- 18 -МОЙ скревдзания (табл. 7). Автогалжя широко распространена среди этих орхидных, чему способствует слабо дифференцированный рсстел-люм (Burns-Balogh e t a l . . 1987; Cat l ing. 1990). В этой сзлзи особый интерес представляют Diplandrorchis s in ica Chen и Archineottla gaudissa r t i i (Hand.-Mzt.) Chen, которые Бкличактся в трибу Neottieae и рассматриваются больсинсгЕом исследователей з качестве наиболее примитивных представителей'отднотычикковьэс орхидных (Chen,' Ta.'ig, 1982; Rasmussnen. 1982; Burns-Balcgh e t a l . . 1987; Аверьянов, 1991 a.6 и др.) . У этих орхидних ростеллюм восб-ке недифферьнцирован, и поэтому нигиая часть пыльки.ка "лежит" на горизонтальном рыльце (Chen.Tang.1982). При такой организации репродуктивных органов сблигаткое автоопыление неизбе^зю приводит к облигатной аутогамии. Из всего этого следует, что гипотетические предки однотычинковых орхидей имели облигатно автогамного (вероятно, весьма близкую к клейстогамной) систему скрез1Еаемости. которая реализовывалась через автоопыление. Автсопь^текие, очевидно. было исходным типом опыления для современных ОДКОТЫЧ;1НКОЕЫ:< орхидных. Такие видоизменения в строении их цветка, как срастание тычиночных нитей со столбиком, вероятно, следует рассматривать как приспособление к автоопылению, что неизбежно сопрово.^лалось редукцией числа фертильных тычинок до одной.
ГЛАВА 6. РЕАЛЬНАЯ СЕМЕННАЯ ПРОДУКТИВНОСТЬ (РСП)
реп плода. Реальная семенная продуктивность плода определялась по числу зрелых неповрежденных семян в коробочке. Число семян подсчитано в 470 коробочках от 21 вида, ОТНОСЯЕКХСЯСЯ К 12 родам из четырех подтриб ( табл. 3).
РСП плода и козта^ициент продуктивности цветка. Реальная семенная продуктивность (РСГО плода является производным показателем от потенциальной семенной продуктивности (ПСП) цветка и связана с ней через коэффициент продуктивности цветка (Левина,1981). Коэффициент продуктивности (КП) цветка принято находить по следующей формуле: РСП
КП . ПСП
где РСП - среднее число зрелых неповреддекных семян в плоде. ГЮП - среднее число семязачатков в завязи.

- 19 -У подаЕЛЯ!С!!:9го больЕинства цветковых растений коэффициент про-
луктизнссти всегда мэньсе единицы, так как в ходе семенного восп-рсиэзедекия не происходит увеличения числа семязачатков (Левина, 1981). Однако при определении КП цветка у изученных орхидных )&; столкну.тась с тем фактом, что у некоторых из 'чекных видов в коробочке сбразовывалссь в среднем больше семян, чем закладывалось семязачатов в завязи. Это было особенно заметно у Dactylorhiza гсга,^а (К), Cphrys cestrifera, 0. taurica Orchis sL-nia и 0. tridentata (табл. 3). В ходе дополнительных исследований было устаксвлено, что во всех случаях превышение числа семян в плоде над числом семлзачатков в завязи обусловлено характером распределения плодов на побеге. У этих орхидных плоды зааяэыва-л>'.сь пре'.эд' эстненно в ни.таей части соцветий, где располагаются самые крупные цвет.чи с каиболыгим числом семязачатков в завязи. Поэтому при определении FJI цветка у изученных видов орхидных вместо ПСП цветка в формулу подставляли среднее число се.мязачат-коз в завязи у опыленных цветков. Зтот показатель расчитывата исходя из местополо.тания коробочек на побеге, на основании тесной корреляции ме.уду числом семязачатков в завязи и местоположением цветка и их числом в соцветии. Ч! .сло семязачатков в завязи у опыленных цветков, если таковое отличаюсь от ПСП цветка, приведены на табл. 3.
КП цзетка и система скресивания. /{звестно, что коэффициент продуктивности (КП) цветка у автогамных растений значительно вы-Е9, чем у ксенога}.шых (Wisr.s, 1984). Эта закономерность просле.тена у видов Cephantherinae, Llnodorinae и Neottiinae. Самые высокие значения КП цветка среди представителей этих подтриб обнаружены у Neottia nidus-avis (0.97) и Epipactis microphylla (0.98), которые в Крыму, вероятно, являются облигатно автогамными видами. У факультативно автогамных Limodorum abortivum и Cephalanthera daiiasonium КП был несколько меньше (0.74 и 0.93. соотвественно). У факультативно ксеногамного Epipactis helleborine и облигатно ксеногамных Cephanthera rubra, С. longifolia и Llstera ovata КП цветка в среднем составил всего 0.67.
КП цветка и агрегация пыльцы в подлинии. Высокие значения КП цветка установлены у представителей подтрибы Orchidinae, которые были преимущественно ксеяогамными видами. В среднем по подтрибе этот показатель составл 0.83. что значительно выше, чем у коено-гa 2 ыx видов из других подтриб. Более высокие зна''ения КП у видов

- 20 -Crchldlnae обусловлены, вероятно, типом агрегации пыльцы и более высокой эффективностью адресного переноса пыльцевых агрегатов этих орхидных при помощи насекомых. Это подтвервдается е^е и тем. что 41 цветка у изученных видов орхидных в целом значительно превысил аналогичный показатель, установленный для многолетних тря-вянитстых растений с неагрегированкой пыльцой из других семейств покрытосемянных (0.50 согласно Д. Винсу (WieriS,1984)).
РСП плода и объем коробочки. Установлена прямая зависимость числа семян в коробочке от ее внешнего объема. Особенно отчетливо ока проявляется у видов с высоким значением коэффициента продуктивности цветка. Для некоторых видов рассчитаны формулы, на основании которых можно определить объем' коробочки по ее линейным размерам.
РСП генеративного побега. Реальная семенная продуктивность генеративного побега расчитывалась исходя из РСП плода и среднего числа зрелых неповредденных плодов на на побеге. В отличие от реальной семенной продуктивности плода РСП генеративного побега у многих изученных орхидных резко сни.жается от исходного уровня. Особенно заметно это у ксеногамкых безнектарных видов. Коэффициент продуктивности (КПЗ цветка у этой группы составил в среднем 0,347. Самые высокие значения КЛ генеративного побега обнаружены у автогамных автоопыляюшхся видов (табл. 4).
ГЛАВА 5. ФАКТОРЫ, СНШАЩИЕ СЕМЕННУЮ ПРОДУКТЙВНХТЬ
Как показано в предыдущей главе, болытнство изученных видов орхидных образуют в плоде и на генеративном побеге значительно меньше семян, чем закладывается семязачатков в завязи или на соцветии. Это происходит в ходе опыления, оплодотворения и созревания семян. Была произведена количественная оценка снижения семенной продуктивности в результате недоопыления отдельного цветка и неопыления некоторых цветков на соцветии, а также нарушений в ходе Эмбриогенеза и повреждения репродуктивных оргсшов фитофагами.
Снижение семенной продуктивности в ходе опыления. Снижение семенной продуктивности в ходе опыления определено у 17 видов как для генеративного побега в целом, так и для отдельного цветка
Эффективность опыления цветка. Эффективность опыления цветка определяли двумя способами - по количеству пыльцы, поступившей на рыльце и по разнице между ДСП цветка и РСП плода за вычетом числа

- 21 -абортивных семян.
Количество пыльцы, поступившей на рыльце, определено у 12 видов КЗ подтркбы Crchidir.ae исходя из среднего числа массул на рыльце и среднего числа пыльцевых зерен в одной массуле. Опылители оставляли на рыльце у этих орхидей в среднем в 1.5-3 раза больше пыльцы, чем закладывалось семязачатков в завязи. Число •.:асоул на рыльце значительно варьировало у изученных видов. У определенной части опыленных цвет.чов на рыльце поступало недостаточное количество пыльцы Значительный процент недоопыленных цветков отмечен у Апаса.трс1з pyramidalis. Orchis picta, Daci-ylcrhiza гоггла (близ г.Ялта), Platanthera bifolia (близ с. Передовое) и Plata."thera chiorantha. 3 результате коэффициент эффективности цветка у этих видов был сувдственно ниже, чем у ос-тальньзс предотазктелеЯ подтрибы (табл. 3).
Недсопылекие цветков обусловлено особенностями поведения и мсрфслсгии опылителей. Например, у Dactylorhiza romana цветки не-лсопыл-плиоь э основном самцами Anthophora acervorurri L. Эти пчелы "патрулировали" среди соцветий орхидеи, лишь изредка обследуя со-дергамое т-оры При этом они никогда не садились на губу, а лишь зависали над цветком. Поэтому они едва касались гемиполлинариями поверхности рыльца, оставляя на нем 4-12 массул.
З'ффективнссть опьи:ения цветков генеративного побега. У ксено-гамных видов орхидей многие цветки на соцветиях остаются неопыле-ным;1 3 конце цветения. У бо.льЕикства безнектарных видов опыляется только 9-30 X цветков. При этом уровень семенной продуктивности этих орхидей многократно снизхается. Осбенно сильно он падает у Orchis purpurea, 0. slir.ia и 0. punctulata, которые привлекают опы-лите.-ей на свои цветки только благодаря яркому облику их соцветий. Значительное снижение семенной продуктивности присуще и видам Ophrys, опьшение у которых основано на псевдокопуляции.
Мимикрия, цветков орхидей с цветками "вознагракдающих" растений более результативна с точки зрения репродукции. Даже при отдаленном сходстве по окраске и морфологии цветков опыление составляет 27-43% (Orchis picta. 0. tridentata, 0. provincial is, Dactylorhlza ron-ana, Comperla coperiana). При значительном сходстве с цветками "B03Harpa.YaaioiiiHx" растений, что было установлено у Anacaitiptis pyramidalis, Cephalanthera rubra, Dactylorhiza romana и D. incarnata, на соцветии опылялось до 90% цветков.
Сншгекиэ семенной продуктивности фитофагами Три вида фитофа-

- 22 -гов повреждали репродуктивные органы крымских орхидей - гусеницы листовертки Lobesia спптеа Flkv. (Lepidoptera, Tor t r ic idae) , личинки мухи Chyliza v i t t a t a f»feig. (Dipthera, Psil idae) и сачсж жука-майки Melee prcscarabaeus L. (Coleopthera, Melcidae).
Lobesia Crimea повреждала 11 видов орхидей. Ее гусеницы питались развивающимися семенами незрелых коробочек. Сна повреждала коробочки только в попуяциях с высоким регулярным плодоношением и большой численностью генеративных особей. Средний процент поврежденных коробочек в популяциях измекался от О до 217..
Chylisa v i t t a t a повреждали репродуктивные органы только у Limodorum abort1УШП и Cephalanthera damasonium. Личинки этой мухи проникают в завязь еще до опыления цветка и питаются вначаче семязачатками. В силу этой особенности биологии этого фитофага он, вероятно, способен повреждать коробочки только у автогамных орхидей. В отдельные годы С. v i t t a t a повреждает более Б07. завязей к незрелых коробочек у L. abort i vuir,.
Численность обоих фитофагов контролируется паразитическими наездниками из рода Вгасоп.
Наименьшее влияние на семенную продуктивность крымских орхидея среди изученных фитофагов о.чазывали самки Kfeloe. Они повреждает репродуктивные органы только у Dactylorhlza гоггапа, поедая верхушки генеративных побегов задолго до распускания цветков.
Снижение семенной продуктивности в ходе оплодотворения и эмбриогенеза. У некоторых видов в плодах paspiiBaiocb значительное количество абортивных семян, которые резко отличались от сстачь-ных по своей форме и по размеру зародыша. Самый высокий процент абортивных семян отмечен у Orchis purpurea и видов Platanthera, которые имели преимуществе-чно ксеногамную систему скревдвания. В коробочках у О. purpurea, образовавшихся после искуственкого гей-теногамного опыления, число абортивных семян было в несколько раз выше по сравнению с коробочкаш от ксеногамного опыления. У автогамных видов в коробочках образовывалось менее 0.1 7. абортивных семян (табл. 3). Более высокий процент абортивных семян у фач^'ль-тативно Kcenoraifflbix видов, вероятно, обусловлен частичной самоке-совместимостью.
ГЛАВА 7. ОСОБЕННОСТИ ДИССЕМИНАЦШ
Диссеиинационные х.4>актеристики семян. Определены следуюше

- 23 -дкссеминаштакные характеристики семени: а) откокение объема тесты (семенной кояуры) к об'ьему эародьша (у 15 видов), б) отнесение проективной п.1о:зди тесты к объему эародьпга Су 15 видов), в) отнесение проективной плопади тесты к весу семени (у 13 видов) и г) скорость свободного паления семени (у б видов). Отношение объема тесты к объему зарсдьпиа коррелирует со скоростью свободного падения семени слабо. Использование этого показателя для оценки способности орхидных к рассеиванию семян при помощи ветра, по-видимому, нецелесообразно. Остальные изученные характеристики семени вполне пригодны д.-л этой цели.
Особенности диссекопяции и возобновления у Orchis purpurea Изучение бли?:-:ей диссеминации у 0. purpurea показало, что наивысшая плотность cnaaa-OEix семян наблюдается на расстоянии от плодоносящего растения, равном, примерно, его полуторной высоте. У 0. purpurea этот псказаьель колеблется ог 50 до 150 см.
Многолетние наблюдения за группа.ми растений семенного происхождения показали, что больЕинство всходов появляются в зоне распрсстра1:екия корневой системы особей более старших возрастных групп (2-5 см от центра листовой розетки).
Выводы. 1. у изученных видов орхидньтс взаимосвязь между уровнем семен
ной продуктивности и редукцией зародыза вырахена не столь явно клк у других микотрофных паразитных растений. Обратная связь между числом семязачатков и степенью общей редукции зародыша наиболее заметна в пределах отдельных родов из относительно примитивных подтриб Cephaiantherinae и Limcdorinae. У более продвинутых родов из подтрибы Orchidinae эта зависимость на фоне общей сильной редукции зародыша значительно ослабляется.
2. Потенциальная семенная продуктивность цветка у изученных видов в значительной степени формируется под влиянием таких особенностей системы опыления, как размер масоул, площадь рыльца, число поллиниев на переносчике, степень морфологического и поведенческого разнообразия опылителей и способ их аттрактации цвет-ка.ми. Сильные различия по этим показателям между отдельными представителями подтрибы Orchidinae, по-видимому, ослабляют связь между семенной продуктивностью и редукцией зародыша.
3. Потенциальная семенная продуктивность цветка на видовом уровне жестко коррелирует с общей фитомассой цветк-г. С фитомассой

- 24 -завязи семенная продуктивность связана менее тесно главным образом из-за сильных видовых различий в степени дифференцированности семязачатклБ во время распускания цветка и степени ру1.!инации плаценту.
4. Орхидные в следствие агрегации пыльцы в поллинии являются исключением из правила, которое установлено Р. Круденом (Cruden, 1977) для изменения отношения числа пыльцевых зерен в'андроцее к числу сешпочек в завязи (П/С-отношение) в зависимости от системы скрещивания вида. Наиболее сильно выпадают из правила Крудена ксеногамные виды. В то время, как у облигатно автогамных видов орхидных П/С-отношение.почти соответствует, правилу Крудена.
5. Величина коэффициента семенной продуктивности цветка и генеративного побега определяются главным образом двумя факторами: насекомыми-опылителями и насекомыми- фитофагами. Уровень семенной продуктивности подвержен наиболытм флюктуациям у безнектарных видов орхидей, которые привлекают опылителей за счет обманных способов атракталии. Фитофаги влияют на семенную продуктивность только в популяциях с регулярно высоким уровнем плодоношения.
6. Щзи автогамном способе скреаквалия у перекрестно опыляемых видов уровень семенной продуктивности цветка может заметно снижаться вследствие частичной самонесовместимссти.
7. Высокое значение коэффициента продуктивности цветка у ксе-ногамных видов подтрибы Orchidinae по сравнению с ксеногамныуд видами Cephalantherinae, Llmodorlnae и Nectinae в значительной степени определяется характером агрегации пыльца
8. Переход орхидей на обманные способы аттрактации прежде всего обусловлен высокой эффективностью адресного переноса пыльцы насеомыми. Переход к безнектарности мокко, вероятно, рассматривать и как эволюционный ответ орхидей на сильное и постоянное давление со стороны фитофагов.
9. Наблюдения за.группами особей семенного происхоадения показали, что вокруг плодоносящего растения 0. purpurea зона с наилуч-ш'лй'. услсЕ;!я^с! длл прорастания се:.шК не сивпадает с зоной наивысшей плотности опадаюазлх семян. Это противоречие между условиями диссеминации и прорастания, по-видимому, резко ограничивает способность этой орхидеи' к семенному возобновлению в популяциях. Его можно устранить за счет оптимизации пространствнной структуры популяций.

- 25 -Работы опубликованные по теме диссертации
1. О критериях выделения возрастных групп цекопопуляций ятрып-ника пурпурного / / 3 сб.: Природные комплексы Крыма, их оптимизация и охрана. Симферополь: СГУ, -1984. С. 74-75.
2. О зозникновекии дочерних групп у ятрышника пурпурного / / В сб.; Природоохра.чные исстедования экосистем Горного Крыма. Симферополь: СГУ. -1986. С. 23-25.
3. Семенная продуктивность некоторых орхидей, произрастагоших в Крь:>лу / / 3 сб.: Тез. докл. 111 Зсес. совещ. "Охрана и культивирование орхидей" У. , февраль 1987. М.: АН СССР. -1986. -С. 38-39.
4. О повреждении плодов орхидных гусеница.ми листовертки Lobesia сп-пеа Flkv. (Lepldcptera, Tortricidae) в Крыму / / Энтог мол. сбозре.чие. -1987. -Т. 66. N. 3. -С. 519-520.
5. Определение реальной семенной продуктивности у Dactylorhiza гога.пз и D. incarnata (Orchidaceae) / / Ботан. журн. -1988. -Т. 73, N. 2. -С. 231-233.
с. Методика подсчета мелких семян и семяпочек (на примере сем. Orchidaceas) / / Ботан. «урн. -1989. -Т. 74. N 8. -С. 1194-1196.
7. Участие комаров из рода Dactylolabis Sav. в опылении тайника овального / / В сб.: Экологические аспекты охраны природы Крыма. Киев: yiffi 30, -1991. С. 87-91.
8. Reproductive system of orchids / / Abstr. of the papers XI Intern. Symp. " Embryology and seed reproduction" (Leningrad. July 3-7, 1990b -Lenigrad, 1990. - P. I l l (совм. с Т. Б. Батыги-ной).
9. Reproductive system of orchids / / Proc. of the papers XI Intern. Symp. " Embryology and seed reproduction" (Leningrad. July 3-7. 1990). -St. Petersdurg: Nauka. -1992. - P. 395-396 (coBM. с T. Б. Батыгиной).
10. Участие рода Chelostoma Latr. (Hymenoptera, Megachiiidae) в опылении мимикрируюш1{Х видов Cephalanthera rubra (Z.) Rich, и Campanula taurica Juz. в Крыму / / Энтомол- обозрение. -1990. -Т. 69. N. 3. -С. 534-537 (совм. с С. II Ивановым).
11. О состоянии крымской популяции ятрышика пурпурного / / в сб.: Шпуляции растений: принципы организации и проблемы охраны природы (Материалы конфер., Йошкар-Ола. 4-8 февраля 1991), Йошкар-Ола. -1991. -С 42-43 (совм. с Л. В. Денисовой. С. Е Никитиной).
12. О репродуктивной способности ятрышника пурпурного / / В сб.:

- 26 -Популяции растений: принципы организации и проблемы охраны природы (Материалы конфер., Яошкар-Ола, 4-8 февраля 1991), Йошкар-Ола, -1991. •-С 43-44 (совм. с С. В. Никитиной, Л. R Ленисовой).
13. Возрастная сруктура ценопопуляциЯ Orchis picta Loisel в Горном Крыму / / В сб.: Рациональное использование и охрана экосистем Крыма. Киев: УМК ВО, -1992. -С. 28-32 (совм. с Л. П. Вахру-шевой).
14. Участие пестрянок (Lepidopthera, Zygaenidae) 1фыма з опылении орхидеи Anacanptls pyramidal is (Orchidaceae) / / Зоол. зкурн. -1993. -Т. 72, N 10. -С. 54-67 (совм. с К. А. Ефетовым).
15. Экология опыления Dactylorhiza гоггапа Soo (Orchidaceae) / / Ук?. ботак. журн. -1993. -Т. 50. N 1. -С. 107-110 (совм. с О. И. Лагутовой).
15. А. е.: Способ подсчета семян: 1476506 СССР - 2 с . : ил.

![The Russian Catholic Work: Roman Catholic Church and Russian émigrés in 1917-1991 [in Russian]](https://img.dokumen.tips/doc/110x75/6352ca8df036fabe550ada2d/the-russian-catholic-work-roman-catholic-church-and-russian-emigres-in-1917-1991.jpg)



























