Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Development of the human fetal insular cortex: studyof the gyration from 13 to 28 gestational weeks
A. Afif Æ R. Bouvier Æ A. Buenerd Æ J. Trouillas ÆP. Mertens
Received: 5 April 2007 / Accepted: 14 September 2007 / Published online: 26 October 2007
� Springer-Verlag 2007
Abstract To describe the morphological stages of insular
sulci and gyri development we carried out a macroscopical
study on 21 human fetal brains, showing no anomalies,
from 13 to 28 gestational weeks (GWs). Particular focus
was given to morphological appearance during the devel-
opment of insular and periinsular structures, especially the
gyration and sulcation of the insula, central cerebral region
and opercula, as well as the vascularization of these
regions. The periinsular sulci and the central (insular and
cerebral) sulci were the first macroscopical structures
identified on the lateral surface of the human fetal cerebral
hemisphere with earlier development on the right hemi-
sphere. Here we describe five stages of insular gyral and
sulcal development closely related to gestational age: stage
1: appearance of the first sulcus at 13-17 GWs, stage 2:
development of the periinsular sulci at 18–19 GWs, stage
3: central sulci and opercularization of the insula at 20–22
GWs, stage 4: covering of the posterior insula at 24–26
GWs, stage 5: closure of the sylvian fissure at 27–28 GWs.
We provide evidence that cortical maturation (sulcation
and gyration) and vascularization of the lateral surface of
the brain starts with the insular region, suggesting that this
region is a central area of cortical development.
Keywords Insular cortex development � Morphology �Insular sulci � Insular gyri � Periinsular sulci
Introduction
The human insular lobe was initially described by the
anatomist Reil JC (Reil 1809) and named the Island of
Reil. Several later anatomical studies focused on this fifth
cerebral lobe and its development (Guldberg 1887; Cunn-
ingham 1891; Clark 1896; Retzius 1896). The first studies
on the cytoarchitectonical development of the human cor-
tex report that the insula is the first cortex to differentiate
and develop in the fetus beginning from 6 weeks of the
fetal life in a cortical region that will later become the
limen insula (Streeter 1912; Kodam 1926). At the first
stages of telencephalic development, the insula is localized
on the lateral surface of each hemisphere (Lockard 1948).
Nevertheless, its surface expansion is more limited than for
any other cerebral lobe. After becoming buried in the
depths of the sylvian fissure during fetal growth, the insula
is progressively covered by temporoparietal and then
frontal opercula (Dorovini-Zis and Dolman 1977; Feess-
Higgins and Larroche 1987). Anatomical data have shown
a left–right asymmetry of gyral and sulcal development,
with the precession of the right cerebral hemisphere gyri
and sulci (Cunningham 1891; Chi et al. 1977; Dorovini-Zis
and Dolman 1977).
The use of more recent techniques, including macro-
scopical, ultrasonographical, and magnetic resonance
A. Afif � P. Mertens (&)
Department of Anatomy, Lyon-1 University,
Inserm U 879, 8 avenue Rockefeller, Lyon 69003, France
e-mail: [email protected]
A. Afif � P. Mertens
Department of Neurosurgery, Neurological Hospital,
Hospices civils de Lyon, Lyon 69003, France
e-mail: [email protected]
R. Bouvier � A. Buenerd
Department of Pathology, Edouard Herriot Hospital,
Hospices civils de Lyon, Lyon 69003, France
J. Trouillas
Laboratory of Histology and Embryology,
Lyon-1 University, Inserm U 842,
Lyon 69003, France
123
Brain Struct Funct (2007) 212:335–346
DOI 10.1007/s00429-007-0161-1

imaging, has led to interesting findings concerning fetal
brain development. In all cases however, the structural
appearance of the insular cortex and its development have
rarely been investigated with no precise description of
insular sulcal development.
The insula constitutes part of the paralimbic structures.
The transitional insular cortex progressively develops from
an allo- to iso-type cortex. Three cytoarchitectonical areas
can be described in primates and man within the insular
cortex (Mesulam and Mufson 1985): (1) the more anterior
agranular area made up of three layers of cells; (2) the
dysgranular area located behind of the agranular area,
occupying the pericentral region; and (3) the posterior
granular area. The topographic distribution of efferent and
afferent projections between the insular cortex and other
cortical regions (Mesulam and Mufson 1982) are related to
different functional systems within the insula. The pos-
terodorsal part of the insula (the granular area) connects
with the retroinsular area, temporal cortex, supplementary
motor area, and both primary and secondary somatosensery
cortex, and specializes in auditory, somesthetic and skel-
etomotor functions. However, the anteroventral agranular
part connects with the entorhinal cortex, cingulated area and
periamygdaloid cortex, and specializes in olfactory, gusta-
tory and viscero-autonomic functions. Limits between these
three insular architectonic areas have never been identified
with respect to insular sulci and gyri organization.
There are a number of recent functional studies that
suggest an important role of the insular cortex especially in
language production and grammatical processing (McCar-
thy et al. 1993; Isnard et al. 2004; Friederici et al. 2006),
pain processing (Peyron et al. 2004; Brooks et al. 2005;
Schreckenberger et al. 2005), viscerosensitive manifesta-
tions (Ostrowsky et al. 2000) and auditory processing
(Lewis et al. 2000; Bamiou et al. 2003).
This study aimed to describe the morphological stages
of insular sulcal and gyral formation by direct anatomical
observation of the human fetal brain between 13 and 28
gestational weeks (GWs). The present study expected to
demonstrate an earlier development of insular sulci than
had previously been suggested by modern imaging tech-
niques (Cohen-Sacher et al. 2006; Ruiz et al. 2006; Garel
et al. 2001). We studied the dynamics of development of
the insular lobe in relation to the opercularization process
and to the development of its arterial supply. Special
interest was devoted to the relationship between the
development of the central cerebral sulcus (CCS) and the
central insular sulcus (CIS).
As embryology enables a better understanding of gross
anatomy, this study also aimed to provide a better com-
prehension of insular morphometry and development. The
results of this study could be useful in: (1) estimating fetal
development at different stages of pregnancy and (2)
describing the sulcal pattern of the insula, thereby helping
to understand the cytoarchitectonical and functional orga-
nization of the gyri and sulci.
Materials and methods
Macroscopical examinations were performed under bin-
ocular vision with low magnification (94) and each
specimen was photographed. Twenty-one human fetal
brains were studied: 42 (30 males and 12 females) for-
malin-fixed hemispheres at different GWs (2–8
hemispheres per GW) ranging from 13 to 28 GWs, except
the 23rd GW. GW in this study was estimated according to
the mother’s last menstrual period and later corrected by
Echography data. All fetal brains came from spontaneous
miscarriage or rarely from medical interruption of preg-
nancy. Fetuses with cerebral or chromosomic anomalies
were excluded. In either case parental consent was obtained
for the study of the fetus.
The first appearance of each of the periinsular and the
insular sulcus was macroscopically identified, as well as
sulci in the central cerebral region. The relationship between
both central (cerebral and insular) sulci was observed. The
dynamics of opercular development was described in the
different stages. The following measurements were taken
(Fig. 1): (1) the length of each periinsular sulcus: the ante-
rior periinsular sulcus separating the insula from the fronto-
orbital operculum, the superior periinsular sulcus separating
the insula from the frontoparietal operculum, and the pos-
teroinferior periinsular sulcus separating the insula from the
Heschl gyrus and the temporal operculum. The length of the
CIS separating the insula in two parts anteroinferior to
posterosuperior, was also studied, (2) the distance between
the projections of the superior extremity of CIS and the
inferior extremity of CCS, (3) the two distances (A and B)
between the superior extremity of CIS and both insular
angles: anterior (the angle between the superior periinsular
sulcus and the anterior periinsular sulcus), and posterior (the
angle between the superior periinsular sulcus and the pos-
teroinferior periinsular sulcus).
We described the chronological development of: (1) the
lateral aspect of the brain hemisphere and the periinsular
sulci, (2) the temporo-parieto-frontal opercular develop-
ment, (3) the insular gyration and sulcation, (4) the sulci in
the central cerebral region (Rolandic), and (5) the devel-
opment of branches of the middle cerebral artery (MCA).
Results
The insular region and the Sylvian fossa were first mac-
roscopically identifiable at 18 GWs by the appearance of
336 Brain Struct Funct (2007) 212:335–346
123

periinsular shallow depressions or periinsular sulci, and the
development of adjacent structures. This development was
always earlier on the right side.
We recorded the mean increase in length of periinsular
sulci and CIS between stage 2 (18–19 GWs) and stage 5
(27–28 GWs) (Table 1): At stage 2, the mean length of the
anterior periinsular sulcus was 8.35 mm (95% confidence
intervals (CI): 7.74–8.96) becoming 14.8 mm (95% CI:
12.10–17.49) at stage 5, with a mean increase equal to
77.24%. The mean length of the superior periinsular sulcus
increased from 16.1 mm (95% CI: 15.27–16.92) to
27.2 mm (95% CI: 22.28–32.12) with a mean increase
equal to 68.9%. The mean length of postero-inferior peri-
insular sulcus increased from 13.3 mm (95% CI: 12.36–
14.24) to 21.1 mm (95% CI: 18.52–23.67) with a mean
increase equal to 58.64%.
The mean length of CIS increased from 10.16 mm (95%
CI: 9.31–11.02) to 17.6 mm (95% CI: 15.71–19.48) with a
mean increase equal to 60.66%.
From 18 and 28 GWs, the mean distance between the
superior extremity of CIS and the anterior insular angle or
the posterior insular angle increased from 11.75 mm (95%
CI: 11.31–12.18) to 19.6 mm (95% CI: 16.70–22.49) with
a mean increase equal to 66.80%, and from 4.91 mm (95%
CI: 4.07–5.75) to 7.6 mm (95% CI: 5.06–10.13) with a
mean increase equal to 54.78%, respectively.
These data show a greater range of development in the
anterior than in the posterior part of the insula. Data
detailing the different stages of development is presented in
Table 1.
Data obtained from study concerning the development
of the insula, the opercula and the central cerebral region
(Table 2) allowed us to identify five stages in the devel-
opment of insula and periinsular structures. Stages of this
original classification are closely related to gestational age
(Table 3).
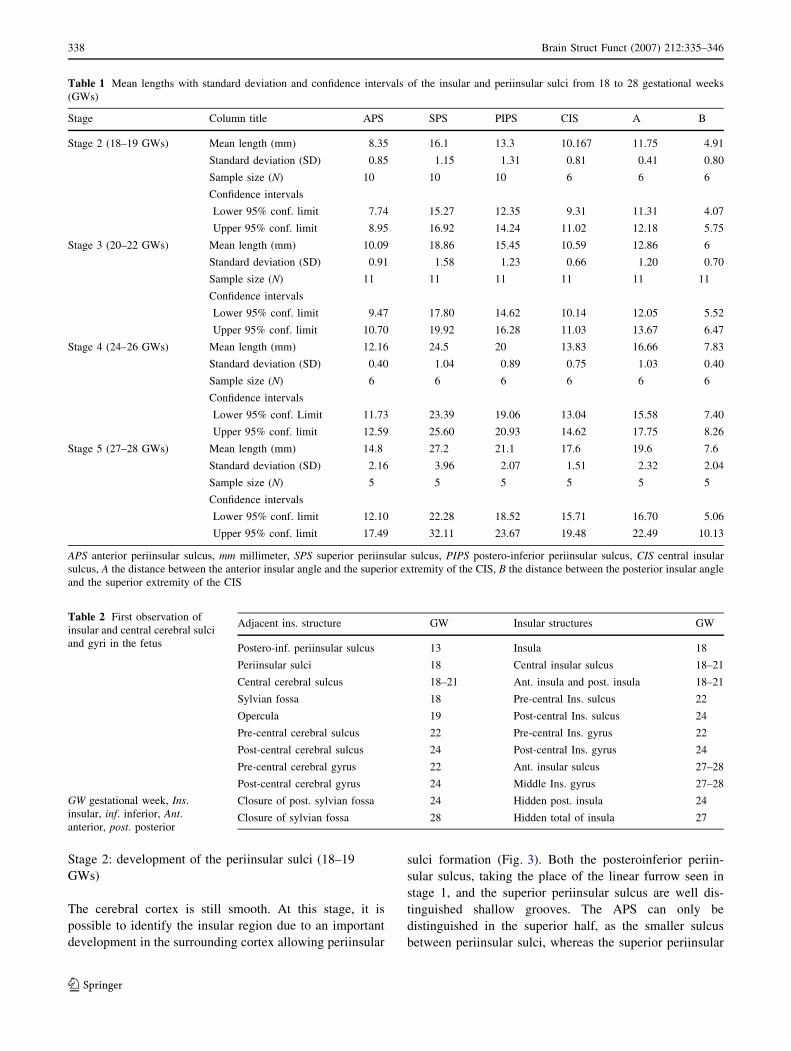
Stage 1: appearance of the first sulcus (13–17 GWs)
The surface of the brain is smooth (Fig. 2a, c). It is not
possible at this stage to macroscopically differentiate the
insular cortex from other cortical structures. At the 13th
GW, a linear furrow (groove), almost perpendicular to the
ventral border of the hemisphere is visible on the lateral
surface at the level of the pterion (Fig. 2a). This linear
furrow can be considered as the limit between the temporal
lobe below, and the insula, frontal and parietal lobes above.
At 15 GWs this limit becomes oblique from anteroinferior
to posterosuperior (Fig. 2b) due to the important develop-
ment of the anterosuperior part of the brain hemisphere.
This first groove, visible on the lateral surface of the
hemisphere at 13 GWs, will later become the posteroin-
ferior periinsular sulcus. It is important to note that all later
development of the insular cortex is above of this groove
(Fig. 2b, d). At 16–17 GWs the inferior and posterosupe-
rior limits of the insula and sylvian fossa are
macroscopically identifiable above of the temporal lobe
(Fig. 2d). The MCA divides into two branches (superior
and inferior), with the predominant superior branche sup-
plying the insular cortex. The MCA and its proximal M1
and M2 branches are present as early as in the 13th GW
(Fig. 2a), at which stage, they are more developed on the
right side than on the left side.
Fig. 1 Insular schematic
showing the different gyri and
sulci. Gyri: I Anterior short
gyrus; II middle short gyrus; IIIprecentral gyrus; IV postcentral
gyrus; V posterior long gyrus;
VI insular pole. Sulci: APSanterior periinsular sulcus; SPSsuperior periinsular sulcus;
PIPS posteroinferior periinsular
sulcus; 1 Anterior insular
sulcus; 2 precentral sulcus; 3central insular sulcus (CIS); 4postcentral sulcus; A distance
between the anterior insular
angle and the superior extremity
of the CIS; B distance between
the posterosuperior insular
angle and the superior extremity
of the CIS
Brain Struct Funct (2007) 212:335–346 337
123
Stage 2: development of the periinsular sulci (18–19
GWs)
The cerebral cortex is still smooth. At this stage, it is
possible to identify the insular region due to an important
development in the surrounding cortex allowing periinsular
sulci formation (Fig. 3). Both the posteroinferior periin-
sular sulcus, taking the place of the linear furrow seen in
stage 1, and the superior periinsular sulcus are well dis-
tinguished shallow grooves. The APS can only be
distinguished in the superior half, as the smaller sulcus
between periinsular sulci, whereas the superior periinsular
Table 1 Mean lengths with standard deviation and confidence intervals of the insular and periinsular sulci from 18 to 28 gestational weeks
(GWs)
Stage Column title APS SPS PIPS CIS A B
Stage 2 (18–19 GWs) Mean length (mm) 8.35 16.1 13.3 10.167 11.75 4.91
Standard deviation (SD) 0.85 1.15 1.31 0.81 0.41 0.80
Sample size (N) 10 10 10 6 6 6
Confidence intervals
Lower 95% conf. limit 7.74 15.27 12.35 9.31 11.31 4.07
Upper 95% conf. limit 8.95 16.92 14.24 11.02 12.18 5.75
Stage 3 (20–22 GWs) Mean length (mm) 10.09 18.86 15.45 10.59 12.86 6
Standard deviation (SD) 0.91 1.58 1.23 0.66 1.20 0.70
Sample size (N) 11 11 11 11 11 11
Confidence intervals
Lower 95% conf. limit 9.47 17.80 14.62 10.14 12.05 5.52
Upper 95% conf. limit 10.70 19.92 16.28 11.03 13.67 6.47
Stage 4 (24–26 GWs) Mean length (mm) 12.16 24.5 20 13.83 16.66 7.83
Standard deviation (SD) 0.40 1.04 0.89 0.75 1.03 0.40
Sample size (N) 6 6 6 6 6 6
Confidence intervals
Lower 95% conf. Limit 11.73 23.39 19.06 13.04 15.58 7.40
Upper 95% conf. limit 12.59 25.60 20.93 14.62 17.75 8.26
Stage 5 (27–28 GWs) Mean length (mm) 14.8 27.2 21.1 17.6 19.6 7.6
Standard deviation (SD) 2.16 3.96 2.07 1.51 2.32 2.04
Sample size (N) 5 5 5 5 5 5
Confidence intervals
Lower 95% conf. limit 12.10 22.28 18.52 15.71 16.70 5.06
Upper 95% conf. limit 17.49 32.11 23.67 19.48 22.49 10.13
APS anterior periinsular sulcus, mm millimeter, SPS superior periinsular sulcus, PIPS postero-inferior periinsular sulcus, CIS central insular
sulcus, A the distance between the anterior insular angle and the superior extremity of the CIS, B the distance between the posterior insular angle
and the superior extremity of the CIS
Table 2 First observation of
insular and central cerebral sulci
and gyri in the fetus
GW gestational week, Ins.
insular, inf. inferior, Ant.anterior, post. posterior
Adjacent ins. structure GW Insular structures GW
Postero-inf. periinsular sulcus 13 Insula 18
Periinsular sulci 18 Central insular sulcus 18–21
Central cerebral sulcus 18–21 Ant. insula and post. insula 18–21
Sylvian fossa 18 Pre-central Ins. sulcus 22
Opercula 19 Post-central Ins. sulcus 24
Pre-central cerebral sulcus 22 Pre-central Ins. gyrus 22
Post-central cerebral sulcus 24 Post-central Ins. gyrus 24
Pre-central cerebral gyrus 22 Ant. insular sulcus 27–28
Post-central cerebral gyrus 24 Middle Ins. gyrus 27–28
Closure of post. sylvian fossa 24 Hidden post. insula 24
Closure of sylvian fossa 28 Hidden total of insula 27
338 Brain Struct Funct (2007) 212:335–346
123
sulcus is longer and consistently bigger on the left side in
both sexes. At this stage, opercularization has not yet
begun. Anterior and posterior insular regions are separated
by a very fine notch at the surface that will become the CIS.
This notch, better identified in its inferior part (Fig. 3b, d),
crosses the insula in an oblique axis, anteroinferior to
posterosuperior, to join the superior periinsular sulcus.
Dorsally the CCS becomes macroscopically identifiable,
especially in its inferior part, limiting the posterior part of
the frontal cortex (Fig. 3b, d). The inferior extremity of
CCS is localized 3–4 mm anterior to the superior extremity
of the CIS. Both CIS and CCS are consistently better
identified on the right side.
Middle cerebral artery branch development over the
surface of the insula is more important compared with other
regions of the lateral cerebral cortex. Anastomosis between
the MCA and anterior cerebral artery can be observed in the
parasagittal region of the lateral cortex and is more devel-
oped within the peri-central (peri-Rolandic) branches. The
diameter of the vessels associated with the central insular
notch progressively increases, with more rapidity on the
right side, representing the rise of the vascular supply to the
future pericentral cerebral region (Fig. 3a, c).
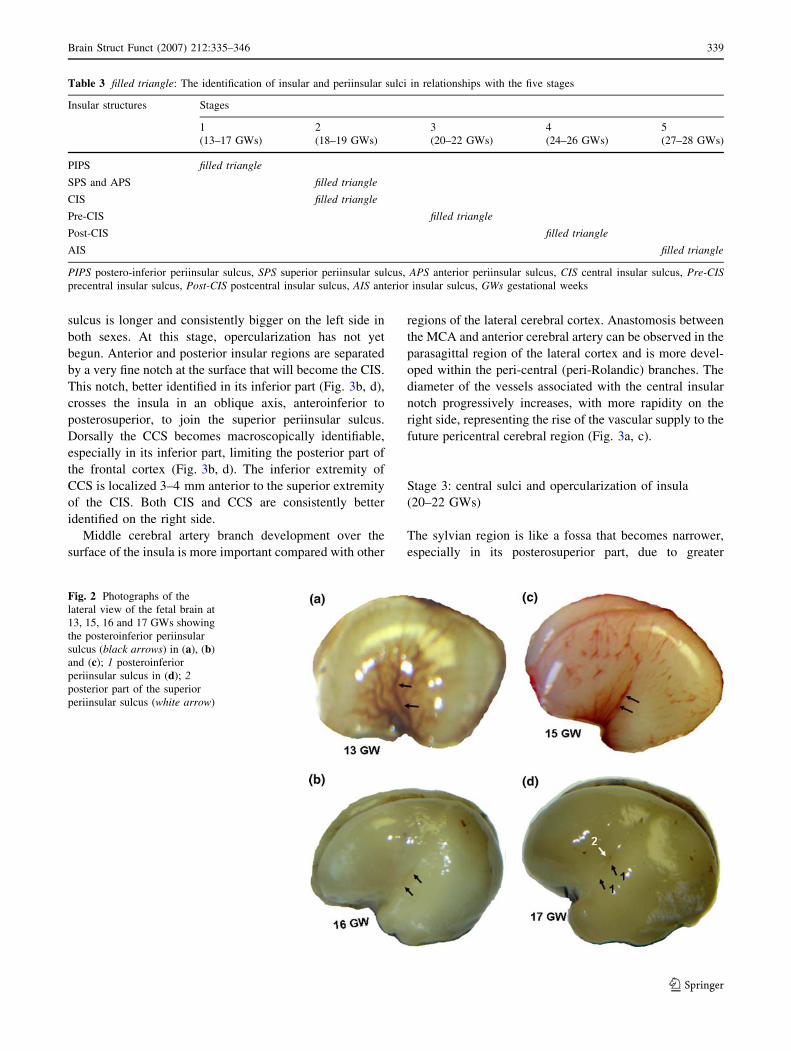
Stage 3: central sulci and opercularization of insula
(20–22 GWs)
The sylvian region is like a fossa that becomes narrower,
especially in its posterosuperior part, due to greater
Table 3 filled triangle: The identification of insular and periinsular sulci in relationships with the five stages
Insular structures Stages
1 2 3 4 5
(13–17 GWs) (18–19 GWs) (20–22 GWs) (24–26 GWs) (27–28 GWs)
PIPS filled triangle
SPS and APS filled triangle
CIS filled triangle
Pre-CIS filled triangle
Post-CIS filled triangle
AIS filled triangle
PIPS postero-inferior periinsular sulcus, SPS superior periinsular sulcus, APS anterior periinsular sulcus, CIS central insular sulcus, Pre-CISprecentral insular sulcus, Post-CIS postcentral insular sulcus, AIS anterior insular sulcus, GWs gestational weeks
Fig. 2 Photographs of the
lateral view of the fetal brain at
13, 15, 16 and 17 GWs showing
the posteroinferior periinsular
sulcus (black arrows) in (a), (b)
and (c); 1 posteroinferior
periinsular sulcus in (d); 2posterior part of the superior
periinsular sulcus (white arrow)
Brain Struct Funct (2007) 212:335–346 339
123
development of the temporal and parietal opercula com-
pared with the frontal opercula (Fig. 4). Moreover,
opercularization takes place more rapidly on the right side.
The periinsular sulci (especially posteroinferior periinsular
sulcus and superior periinsular sulcus) and the majority of
the posterior insula are, at this stage, hidden by operculi
(Fig. 4c, d). The inferior part of the anterior periinsular
sulcus becomes macroscopically identifiable. Both CCS
and CIS become more visible ‘‘shallow grooves’’. Projec-
tion of the lower extremity of the CCS is localized 1–2 mm
anterior to the superior extremity of CIS. At this stage of
development, it becomes possible to identify the notches of
precentral sulci (insular and cerebral) by the 20th GW, and
postcentral sulci (insular and cerebral) by the 22nd GW. It
is therefore possible to distinguish the pre- and postcentral
(insular and cerebral) gyri (Fig. 7).
Fig. 3 Photographs of the
lateral view of the fetal brain aat 18 GWs showing the middle
cerebral artery and its branches
‘‘M2, M3, M4 and M5’’; A the
artery associated with the
central insular notch
representing the future
pericentral region arteries (Barrows). b At 18 GWs showing
the insular region (blackarrows) due to development of
adjacent insular structures; 1central cerebral sulcus; 2 central
insular sulcus. c At 19 GWs
showing the vascularization of
insular region; A central insular
artery and the central cerebral
‘‘Rolandic’’ artery (B). d At 19
GWs showing the central
insular sulcus (2 arrowheads);
and the central cerebral sulcus
(2 arrows). Both sulci are better
identified in the inferior part
Fig. 4 Photographs of the
lateral view of the fetal brain aat 21 GWs showing the sylvian
fossa which has become narrow
at its posterior part following
temporal and parietal opercula
development (arrows). b At 21
GWs showing the central
cerebral sulcus (1) and the
central insular sulcus (2). c At
22 GWs showing the artery
which corresponds to the central
insular sulcus (‘a’ arrow) giving
the central cerebral artery (‘b’
arrow). A large area of the
posterior insula is hidden by the
temporal and parietal opercula
(white arrows). d At 22 GWs
showing the inferior extremity
of the central cerebral sulcus
(1), the superior extremity of the
central insular sulcus (2) and the
precentral cerebral sulcus (3)
340 Brain Struct Funct (2007) 212:335–346
123
The diameter of MCA and its branches M2 and M3 are
increased, in particular the vessels which correspond to the
CIS. The superior division and its branches supply the
anterior part of the insula. The central cerebral artery arises
from the central insular artery.
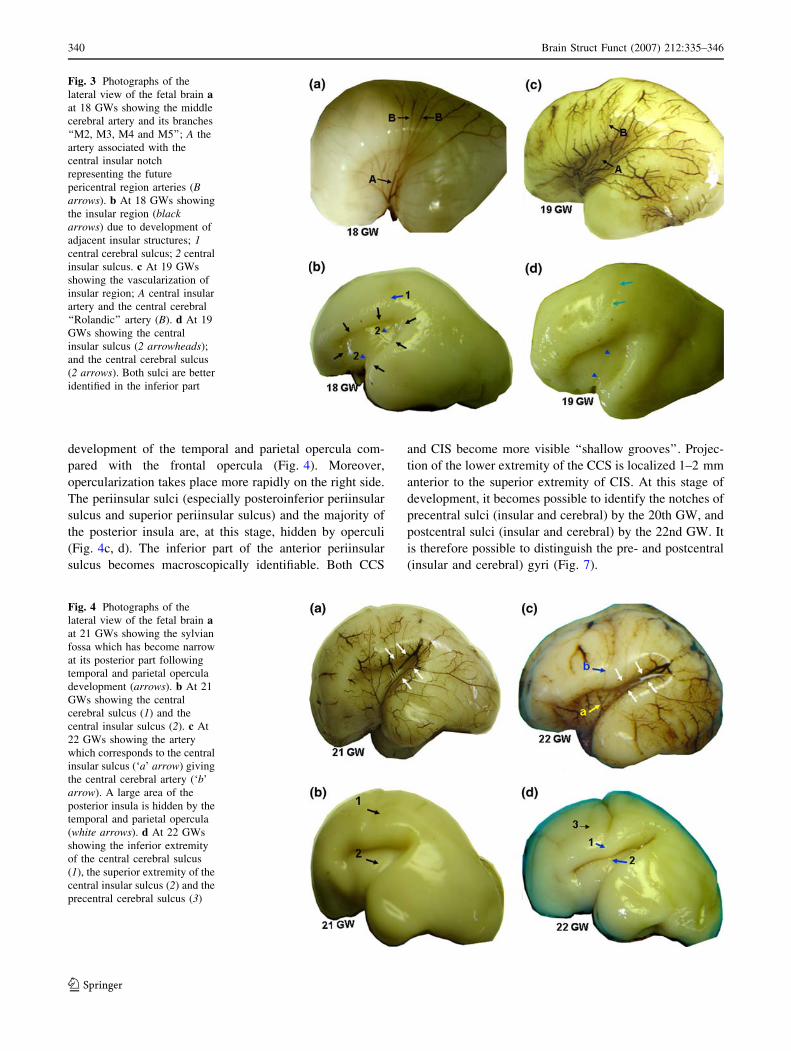
Stage 4: covering of the posterior insula (24–26 GWs)
The surrounding cortex overlaps the posterior part of the
sylvian region at this stage, with only partial covering of
the anterior tip of the anterior insula and the anterior
periinsular sulcus (Fig. 5c, d). The superior part of the
anterior insula gradually becomes hidden along with the
development of the frontoorbital opercula. The lower
extremity of CCS is orientated in the same axis to the
superior extremity of the CIS (Fig. 5b). The superior
extremity of CIS separates the superior periinsular sulcus
into two interior thirds and one posterior third (Table 1). At
this stage, the postCIS can clearly be seen, and the insular
gyri (posterior, postcentral, and precentral) are macro-
scopically identifiable (Fig. 5c, d).
The distal branches (M4 and M5) of the MCA and their
anastomoses with the branches of the anterior cerebral
artery are more easily identifiable. The central cerebral
artery arises from the central insular artery (collateral of
the superior division of the MCA) (Fig. 5a).
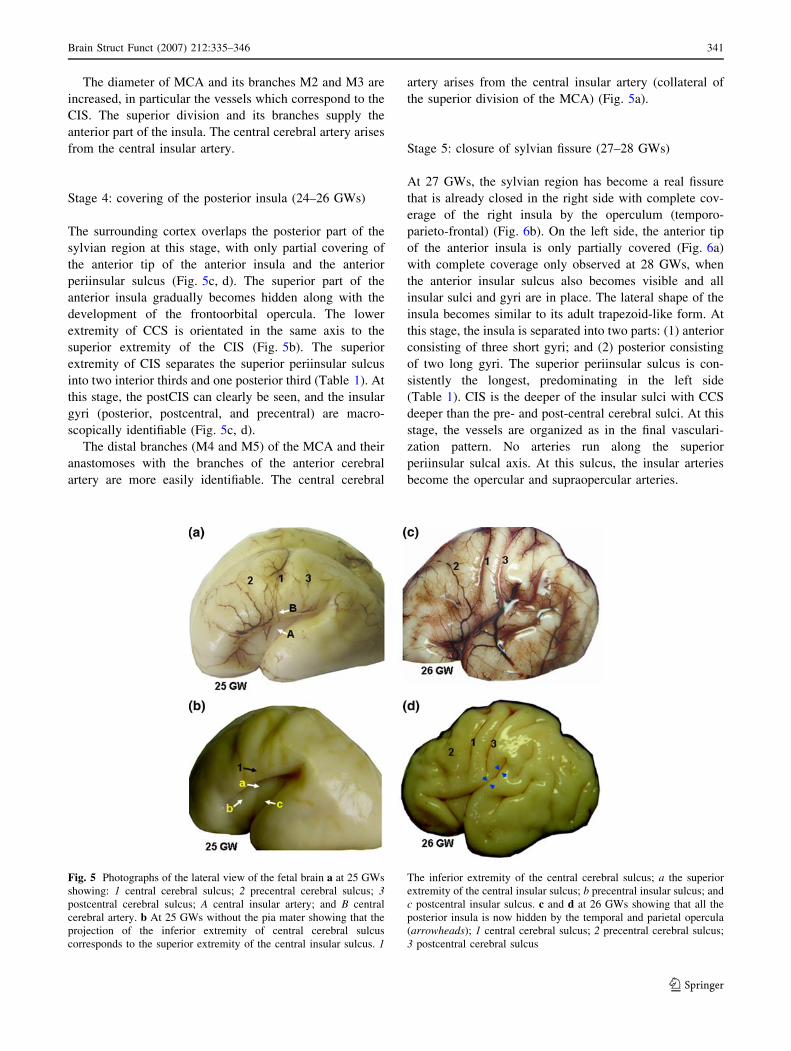
Stage 5: closure of sylvian fissure (27–28 GWs)
At 27 GWs, the sylvian region has become a real fissure
that is already closed in the right side with complete cov-
erage of the right insula by the operculum (temporo-
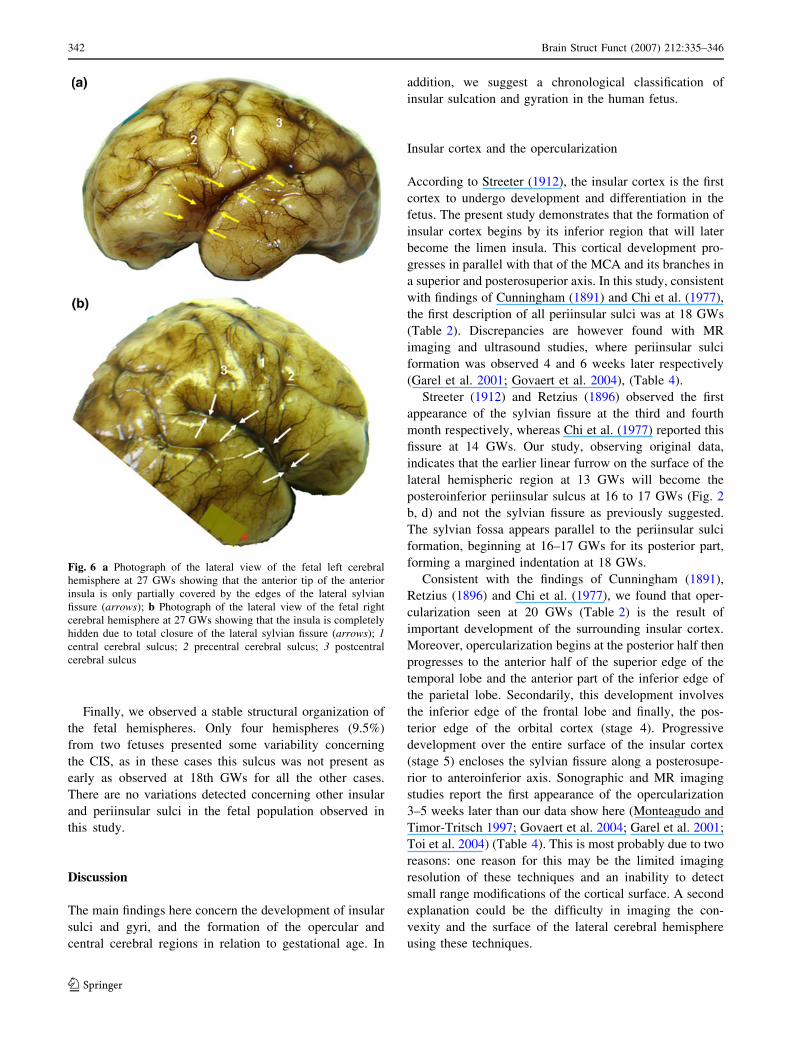
parieto-frontal) (Fig. 6b). On the left side, the anterior tip
of the anterior insula is only partially covered (Fig. 6a)
with complete coverage only observed at 28 GWs, when
the anterior insular sulcus also becomes visible and all
insular sulci and gyri are in place. The lateral shape of the
insula becomes similar to its adult trapezoid-like form. At
this stage, the insula is separated into two parts: (1) anterior
consisting of three short gyri; and (2) posterior consisting
of two long gyri. The superior periinsular sulcus is con-
sistently the longest, predominating in the left side
(Table 1). CIS is the deeper of the insular sulci with CCS
deeper than the pre- and post-central cerebral sulci. At this
stage, the vessels are organized as in the final vasculari-
zation pattern. No arteries run along the superior
periinsular sulcal axis. At this sulcus, the insular arteries
become the opercular and supraopercular arteries.
Fig. 5 Photographs of the lateral view of the fetal brain a at 25 GWs
showing: 1 central cerebral sulcus; 2 precentral cerebral sulcus; 3postcentral cerebral sulcus; A central insular artery; and B central
cerebral artery. b At 25 GWs without the pia mater showing that the
projection of the inferior extremity of central cerebral sulcus
corresponds to the superior extremity of the central insular sulcus. 1
The inferior extremity of the central cerebral sulcus; a the superior
extremity of the central insular sulcus; b precentral insular sulcus; and
c postcentral insular sulcus. c and d at 26 GWs showing that all the
posterior insula is now hidden by the temporal and parietal opercula
(arrowheads); 1 central cerebral sulcus; 2 precentral cerebral sulcus;
3 postcentral cerebral sulcus
Brain Struct Funct (2007) 212:335–346 341
123
Finally, we observed a stable structural organization of
the fetal hemispheres. Only four hemispheres (9.5%)
from two fetuses presented some variability concerning
the CIS, as in these cases this sulcus was not present as
early as observed at 18th GWs for all the other cases.
There are no variations detected concerning other insular
and periinsular sulci in the fetal population observed in
this study.
Discussion
The main findings here concern the development of insular
sulci and gyri, and the formation of the opercular and
central cerebral regions in relation to gestational age. In
addition, we suggest a chronological classification of
insular sulcation and gyration in the human fetus.
Insular cortex and the opercularization
According to Streeter (1912), the insular cortex is the first
cortex to undergo development and differentiation in the
fetus. The present study demonstrates that the formation of
insular cortex begins by its inferior region that will later
become the limen insula. This cortical development pro-
gresses in parallel with that of the MCA and its branches in
a superior and posterosuperior axis. In this study, consistent
with findings of Cunningham (1891) and Chi et al. (1977),
the first description of all periinsular sulci was at 18 GWs
(Table 2). Discrepancies are however found with MR
imaging and ultrasound studies, where periinsular sulci
formation was observed 4 and 6 weeks later respectively
(Garel et al. 2001; Govaert et al. 2004), (Table 4).
Streeter (1912) and Retzius (1896) observed the first
appearance of the sylvian fissure at the third and fourth
month respectively, whereas Chi et al. (1977) reported this
fissure at 14 GWs. Our study, observing original data,
indicates that the earlier linear furrow on the surface of the
lateral hemispheric region at 13 GWs will become the
posteroinferior periinsular sulcus at 16 to 17 GWs (Fig. 2
b, d) and not the sylvian fissure as previously suggested.
The sylvian fossa appears parallel to the periinsular sulci
formation, beginning at 16–17 GWs for its posterior part,
forming a margined indentation at 18 GWs.
Consistent with the findings of Cunningham (1891),
Retzius (1896) and Chi et al. (1977), we found that oper-
cularization seen at 20 GWs (Table 2) is the result of
important development of the surrounding insular cortex.
Moreover, opercularization begins at the posterior half then
progresses to the anterior half of the superior edge of the
temporal lobe and the anterior part of the inferior edge of
the parietal lobe. Secondarily, this development involves
the inferior edge of the frontal lobe and finally, the pos-
terior edge of the orbital cortex (stage 4). Progressive
development over the entire surface of the insular cortex
(stage 5) encloses the sylvian fissure along a posterosupe-
rior to anteroinferior axis. Sonographic and MR imaging
studies report the first appearance of the opercularization
3–5 weeks later than our data show here (Monteagudo and
Timor-Tritsch 1997; Govaert et al. 2004; Garel et al. 2001;
Toi et al. 2004) (Table 4). This is most probably due to two
reasons: one reason for this may be the limited imaging
resolution of these techniques and an inability to detect
small range modifications of the cortical surface. A second
explanation could be the difficulty in imaging the con-
vexity and the surface of the lateral cerebral hemisphere
using these techniques.
Fig. 6 a Photograph of the lateral view of the fetal left cerebral
hemisphere at 27 GWs showing that the anterior tip of the anterior
insula is only partially covered by the edges of the lateral sylvian
fissure (arrows); b Photograph of the lateral view of the fetal right
cerebral hemisphere at 27 GWs showing that the insula is completely
hidden due to total closure of the lateral sylvian fissure (arrows); 1central cerebral sulcus; 2 precentral cerebral sulcus; 3 postcentral
cerebral sulcus
342 Brain Struct Funct (2007) 212:335–346
123
Insular and central cerebral region sulcation and
gyration
Central sulci
In agreement with studies by Cunningham (1891) and
Retzius (1896), we first observed both CCS and CIS at 18
GWs (stage 2), with the right side appearing before the left.
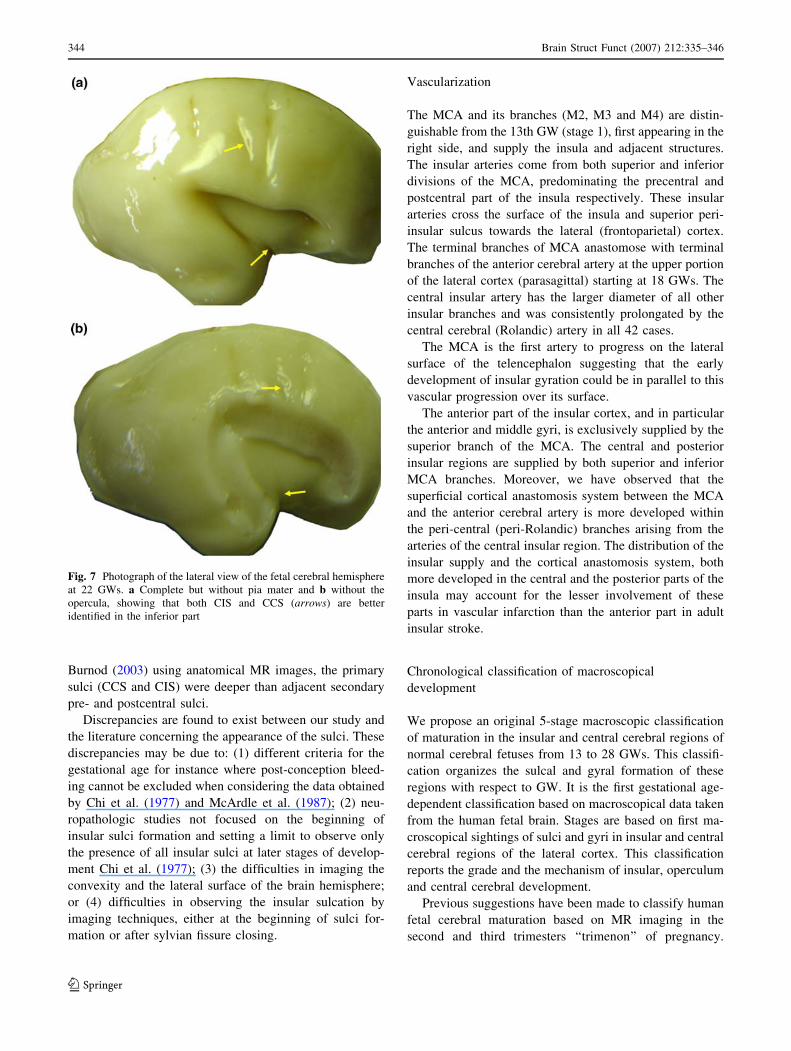
In the present study, the initial formation of the CCS was
identified in the inferior part (Fig. 7) and not in the para-
sagittal part as suggested by Chi et al. (1977). No data
concerning the maturation of CIS exists from imaging
technique studies, which report the appearance of CCS 6–8
weeks later (Cohen-Sacher et al. 2006; Ruiz et al. 2006;
Garel et al. 2001) (Table 4).
The CCS and the CIS were found to develop inde-
pendently suggesting that the CCS is not strictly
continuous with the CIS. Original data from our study
show that at early development stages, the inferior
extremity of CCS is located anterior to the superior
extremity of CIS. The two extremities correspond at stage
4. This evolution may be due to two reasons: (1) the
posterior displacement of CCS, especially in its inferior
part, related to the development of the frontal lobe (2) the
anterior displacement of the superior portion of CIS, due
to postero-anterior growth of the upper posterior area of
the insular cortex. The CIS, straight in the early stages 2
and 3, becomes curved in its superior part at stage 4 of
development.
Pre- and postcentral sulci
Consistent with a study by Cunningham (1891), we
observed the precentral (insular and cerebral) sulci at 20–
22 GWs, and the postcentral (insular and cerebral) sulci at
22–24 GWs. Chi et al. (1977) observed the insular sulci
later at 32–35 GWs. To our knowledge no radiological
studies have reported the first appearance of the insular
sulci. Under MRI, the pre- and postcentral cerebral sulci
appeared only 6–7 weeks later (Garel et al. 2001, 2003)
(see Table 4). According to a geometric study by Toro and
Table 4 Chronology of sulcation and gyration in the present study in comparison with anatomical, RM imaging, and ultrasound studies
Structure Anatomical studies (GWs) RMI studies (GWs) Ultrasound studies (GWs) Present study (GWs)
Periinsular sulci 16–19 (1) After 17 (5) 17–18
CCS 20–23 (1) 24–25 (2) 26 (3) 18–19
Sylvian fissure 14 (1) 19 (6) 18 (4) 18
Opercula 16–19 (1) 22–23 (2) After 17 (5) 20
Pre-CCS 24 (1) 26 (2) 27 (3) 22
Post-CCS 25 (1) 27 (2) 28 (3) 24
Pre-central cerebral Gy. 24 (1) 26 (2) 27 (3) 22
Post-central cerebral Gy. 25 (1) 27 (2) 28 (3) 24
Closure post. part of Syl. 26–30 (1) After 23 (2) 25 (3) 24
Closure total of Syl. After 30 (1) 34 (3) 28
Insula 18 (1) 22–23 (2) After 17 (5) 18
CIS 18–19
Ant. insula and post. Ins. 18–9
Pre-CIS 5.5–6 GMs (7) 22
Post-CIS 5.5–6 GMs (7) 24
AIS 27–28
Pre-central Ins. Gy. 22
Post-central Ins. Gy. 24
Middle Ins. Gy. 27–28
Hidden post. Ins. 26–30 (1) After 23 (2) 25 (3) 24
Hidden total of Ins. 32 (3) 27–28
Total Ins. sulci and gyri 32–35 (1) 33–34 (2) 33–34 (3) 28
GWs gestational weeks, GMs gestational months, (1) Chi, (2) Garel, (3) Ruiz, (4) Monteagudo, (5) Toi, (6) Abe, (7) Cunningham, CCS central
cerebral sulcus, Pre-CCS pre-central cerebral sulcus, Post-CCS post-central cerebral sulcus, Gy. gyrus, Syl. sylvian, Ant. anterior, Post posterior,
CIS central insular sulcus, Pre-CIS precentral insular sulcus, Post-CIS postcentral insular sulcus, AIS anterior insular sulcus, Ins. insular
Brain Struct Funct (2007) 212:335–346 343
123
Burnod (2003) using anatomical MR images, the primary
sulci (CCS and CIS) were deeper than adjacent secondary
pre- and postcentral sulci.
Discrepancies are found to exist between our study and
the literature concerning the appearance of the sulci. These
discrepancies may be due to: (1) different criteria for the
gestational age for instance where post-conception bleed-
ing cannot be excluded when considering the data obtained
by Chi et al. (1977) and McArdle et al. (1987); (2) neu-
ropathologic studies not focused on the beginning of
insular sulci formation and setting a limit to observe only
the presence of all insular sulci at later stages of develop-
ment Chi et al. (1977); (3) the difficulties in imaging the
convexity and the lateral surface of the brain hemisphere;
or (4) difficulties in observing the insular sulcation by
imaging techniques, either at the beginning of sulci for-
mation or after sylvian fissure closing.
Vascularization
The MCA and its branches (M2, M3 and M4) are distin-
guishable from the 13th GW (stage 1), first appearing in the
right side, and supply the insula and adjacent structures.
The insular arteries come from both superior and inferior
divisions of the MCA, predominating the precentral and
postcentral part of the insula respectively. These insular
arteries cross the surface of the insula and superior peri-
insular sulcus towards the lateral (frontoparietal) cortex.
The terminal branches of MCA anastomose with terminal
branches of the anterior cerebral artery at the upper portion
of the lateral cortex (parasagittal) starting at 18 GWs. The
central insular artery has the larger diameter of all other
insular branches and was consistently prolongated by the
central cerebral (Rolandic) artery in all 42 cases.
The MCA is the first artery to progress on the lateral
surface of the telencephalon suggesting that the early
development of insular gyration could be in parallel to this
vascular progression over its surface.
The anterior part of the insular cortex, and in particular
the anterior and middle gyri, is exclusively supplied by the
superior branch of the MCA. The central and posterior
insular regions are supplied by both superior and inferior
MCA branches. Moreover, we have observed that the
superficial cortical anastomosis system between the MCA
and the anterior cerebral artery is more developed within
the peri-central (peri-Rolandic) branches arising from the
arteries of the central insular region. The distribution of the
insular supply and the cortical anastomosis system, both
more developed in the central and the posterior parts of the
insula may account for the lesser involvement of these
parts in vascular infarction than the anterior part in adult
insular stroke.
Chronological classification of macroscopical
development
We propose an original 5-stage macroscopic classification
of maturation in the insular and central cerebral regions of
normal cerebral fetuses from 13 to 28 GWs. This classifi-
cation organizes the sulcal and gyral formation of these
regions with respect to GW. It is the first gestational age-
dependent classification based on macroscopical data taken
from the human fetal brain. Stages are based on first ma-
croscopical sightings of sulci and gyri in insular and central
cerebral regions of the lateral cortex. This classification
reports the grade and the mechanism of insular, operculum
and central cerebral development.
Previous suggestions have been made to classify human
fetal cerebral maturation based on MR imaging in the
second and third trimesters ‘‘trimenon’’ of pregnancy.
Fig. 7 Photograph of the lateral view of the fetal cerebral hemisphere
at 22 GWs. a Complete but without pia mater and b without the
opercula, showing that both CIS and CCS (arrows) are better
identified in the inferior part
344 Brain Struct Funct (2007) 212:335–346
123

McArdle et al. (1987) proposed a classification of five
stages. This classification focuses on frontal and occipital
sulcation and gyration, observing the folds of the insular
cortex at the stage 4 only (37–39 GWs). The development
of the cerebral cortex was later classified into eight stages
using MR images in the Ac-Pc plane, with special attention
to the frontal and temporal lobes (Abe et al. 2003).
Comparing previous classifications using MR images
with the classification proposed in this study shows that
differences found could explain the discrepancies seen
between this and previous studies. Firstly, neither of the
MRI classifications observed complete development of the
insular cortex, and sulci appearance was described 4–8
weeks later than first seen in this study. The reason could
be that the MRI studies used thicker slices (6–8 mm slice
thickness). Secondly, the MRI studies are not tangential to
the convexity of the surface of the lateral cerebral cortex
and it is difficult to obtain true axial, sagittal, and coronal
planes, so they are not able to detect small ranges (the
linear furrows) of cortical surface modification.
Conclusion
To our knowledge this is the first study focusing on the
morphological and chronological development of the
human insular cortex and adjacent structures. We have
described an earlier appearance of insular and periinsular
sulci than previously described by neuroimaging studies.
This study suggests that the insular lobe is the first
cerebral lobe to be identified by macroscopic techniques
with earlier right side development. Periinsular sulci, CCS
and CIS are the first sulci to develop in the lateral surface
of the human fetal cerebral hemisphere. We have proposed
a five stage classification describing the precise chronology
of fetal insular cortex gyration in relation to fetal gesta-
tional age.
Acknowledgment We thank Ms Emily Witty and Mr Michel
Magnin for editorial review of the text.
References
Abe S, Takagi K, Yamamoto T, Okuhata Y, Kato T (2003)
Assessment of cortical gyrus and sulcus formation using MR
images in normal fetuses. Prenat Diagn 23:225–231
Bamiou DE, Musiek FE, Luxon LM (2003) The insula (Island of Reil)
and its role in auditory processing literature review. Brain Res Rev
Brooks JCW, Zambreanu L, Godinez A, Craig AD, Tracey I (2005)
Somatotopic organization of the human insula to painful heat
studied with high resolution functional imaging. Neuroimage
27:201–209
Chi JG, Dooling EC, Gilles FH (1977) Gyral development of the
human brain. Ann Neurol 1:86–93
Clark TE (1896) The comparative anatomy of the insula. J Comp
Neurol 6:59–100
Cohen-Sacher B, Lerman-Sagie T, Lev D, Malinger G (2006)
Sonographic developmental milestones of the fetal cerebral
cortex: a longitudinal study. Ultrasound Obstet Gynecol 27:494–
502
Cunningham DJ (1891) The development of the gyri and sulci on the
surface of the island of Reil of the brain. J Anat Physiol 25:338–
348
Dorovini-Zis K, Dolman CL (1977) Gestational development of
brain. Arch Pathol Lab Med 101:192–195
Feess-Higgins A, Larroche J-C (1987) Le developpement du cerveau
foetal humain. Atlas Anatomique. Masson, Paris (In French)
Friederici AD, Bahlmann J, Heim S, Schubotz RI, Anwander A
(2006) The brain differentiates human and non-human gram-
mars: functional localization and structural connectivity. Proc
Natl Acad Sci USA 14 103(7):2458–2463
Garel C, Chantrel E, Brisse H, Elmaleh M, Luton D, Oury JF, Sebag
G, Hassan M (2001) Fetal cerebral cortex: normal gestational
landmarks identified using prenatal MR imaging. AJNR Am J
Neuroradiol 22:184–189
Garel C, Chantrel E, Elmaleh M, Brisse H, Sebag G (2003) Fetal
MRI: normal gestational landmarks for cerebral biometry,
gyration and myelination. Childs Nerv Syst 19:422–425
Govaert P, Swarte R, De Vos A, Lequin M (2004) Sonographic
appearance of the normal and abnormal insula of Reil. Dev Med
Child Neurol 46(9):610–616
Guldberg A (1887) —Zur morphologie der insula Reillis. Anat Anz
2:659–665
Isnard J, Guenot M, Sindou M, Mauguiere F (2004) Clinical
manifestations of insular lobe seizures: a stereo-electroenceph-
alographic study. Epilepsia 45(9):1079–1090
Kodam S (1926) Uber die sogenannten Basalganglien, Morphoge-
netische und pathologisch-anatomische Untersuchunger.
Schweiz Arch Neurol Psychiatr 18:179–246
Lewis JM, Beauchamp MS, De Yoe EA (2000) A comparison of
visual and auditory motion processing in human cerebral cortex.
Cereb Cortex 10(9):873–888
Lockard I (1948) Certain developmental relations and fiber connec-
tions of the triangular gyrus in primates. J Comp Neurol 89:349–
386
McArdle CB, Joan Richardson C, Nicholas DA, Mirfakhraee M,
Keith Hayden C, Amparo EG (1987) Developmental features of
the neonatal brain: MR imaging. Part I. gray white matter
differentiation and myelination. Radiology 162:223–229
McCarthy G, Blamier AM, Rothman DL, Gruelter R, Shulman RG
(1993) Echo-planar magnetic resonance imaging studies of
frontal cortex activation during word generation in humans. Proc
Natl Acad Sci USA 90:4952–4956
Mesulam MM, Mufson EJ (1985) The Insula of Reil in man and
monkey. In: Jones EG, Peters AA (eds) Cerebral cortex. Plenum
Press, New York, pp 179–226
Mesulam MM, Mufson EJ (1982) Insula of the old world monkey. I.
Architectonics in the insulo-orbito-temporal component of the
paralimbic brain. J Comp Neurol 212:1–22
Monteagudo A, Timor-Tritsch IE (1997) Development of fetal gyri,
and fissures: a transvaginal sonographic study. Ultrasound Obstet
Gynecol 9:222–228
Ostrowsky K, Isnard J, Ryvlin P, Guenot M, Fischer C, Mauguiere F
(2000) Functional mapping of the insular cortex: clinical
implication in temporal lobe epilepsy. Epilepsia 41(6):681–686
Peyron R, Schneider F, Faillenot I, Convers P, Barral FG, Garcia-
Larrea L, Laurent B (2004) An fMRI study of cortical
representation of mechanical allodynia in patients with neuro-
pathic pain. Neurology 63(10):1838–1846
Brain Struct Funct (2007) 212:335–346 345
123

Reil JC (1809) Unterfuchungen uber den Bau des grofsen Gehirns im
Menfchen: Vierte Fortsetzung VIII. Arch Physiol Halle 9:136–146
Retzius G (1896) Das menschenhirn studien in der makroskopischen
morphologie. Morstedt, Stockholm
Ruiz A, Sembely-Taveau C, Paillet C, Sirinelli D (2006) Sonographic
cerebral sulcal pattern in normal fetuses. Radiologie 87:49–55
Schreckenberger M, Siessmeier T, Viertmann A, Landvogt C,
Buchholz HG, Rolke R, Treede RD, Bartenstein P, Birklein F
(2005) The unpleasantness of tonic pain is encoded by the
insular cortex. Neurology 64(7):1175–1183
Streeter GL (1912) The development of the nervous system. In:
Keibel F, Mall FP (eds) Manual of human embryology, vol II,
chapter XIV. Lippincott, Philadelphia
Toi A, Lister WS, Fong KW (2004) How early are fetal cerebral sulci
visible at prenatal ultrasound and what is the normal pattern of
early fetal sulcale development? Ultrasound Obstet Gynecol
24:706–715
Toro R, Burnod Y (2003) Geometric atlas: modeling the cortex as an
organized surface. Neuroimage 20:1468–1484
346 Brain Struct Funct (2007) 212:335–346
123





























