Embed Size (px)
Citation preview

96:2877-2888, 2006. First published Aug 9, 2006; doi:10.1152/jn.00664.2006 J NeurophysiolAceves Dagoberto Tapia, Verónica Anaya, Elvira Galarraga, José Bargas and Jorge Adán Hernández, Osvaldo Ibáñez-Sandoval, Arturo Sierra, René Valdiosera,
Dopamine Receptors4 and D2/3Globus Pallidus by DControl of the Subthalamic Innervation of the Rat
You might find this additional information useful...
114 articles, 47 of which you can access free at: This article cites http://jn.physiology.org/cgi/content/full/96/6/2877#BIBL
including high-resolution figures, can be found at: Updated information and services http://jn.physiology.org/cgi/content/full/96/6/2877
can be found at: Journal of Neurophysiologyabout Additional material and information http://www.the-aps.org/publications/jn
This information is current as of November 22, 2006 .
http://www.the-aps.org/.American Physiological Society. ISSN: 0022-3077, ESSN: 1522-1598. Visit our website at (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by the
publishes original articles on the function of the nervous system. It is published 12 times a yearJournal of Neurophysiology
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from

Control of the Subthalamic Innervation of the Rat Globus Pallidus by D2/3
and D4 Dopamine Receptors
Adan Hernandez,1 Osvaldo Ibanez-Sandoval,2 Arturo Sierra,1 Rene Valdiosera,1 Dagoberto Tapia,2
Veronica Anaya,3 Elvira Galarraga,2 Jose Bargas,2 and Jorge Aceves1
1Departamento de Fisiologıa, Biofısica y Neurociencias, Centro de Investigacion y Estudios Avanzados; 2Departamento de Biofısica,Instituto de Fisiologıa Celular; and 3Neurociencias, Facultad de Estudios Superiores-Iztacala, Universidad Nacional Autonoma deMexico, Federal District, Mexico City, Mexico
Submitted 26 June 2006; accepted in final form 3 August 2006
Hernandez, Adan, Osvaldo Ibanez-Sandoval, Arturo Sierra, ReneValdiosera, Dagoberto Tapia, Veronica Anaya, Elvira Galarraga,Jose Bargas, and Jorge Aceves. Control of the subthalamic inner-vation of the rat globus pallidus by D2/3 and D4 dopamine receptors.J Neurophysiol 96: 2877–2888, 2006. First published August 9, 2006;doi:10.1152/jn.00664.2006. The effects of activating dopaminergicD2/3 and D4 receptors during activation of the subthalamic projectionto the globus pallidus (GP) were explored in rat brain slices using thewhole cell patch-clamp technique. Byocitin labeling and both ortho-dromic and antidromic activation demonstrated the integrity of somesubthalamopallidal connections in in vitro parasagittal brain slices.Excitatory postsynaptic currents (EPSCs) that could be blocked byCNQX and AP5 were evoked onto pallidal neurons by local fieldstimulation of the subthalamopallidal pathway in the presence ofbicuculline. Bath application of dopamine and quinpirole, a dopami-nergic D2-class receptor agonist, reduced evoked EPSCs by about35%. This effect was only partially blocked by sulpiride, a D2/3
receptor antagonist. The sulpiride-sensitive reduction of the sub-thalamopallidal EPSC was associated with an increase in the paired-pulse ratio (PPR) and a reduction in the frequency but not the meanamplitude of spontaneous EPSCs (sEPSCs), indicative of a presynap-tic site of action, which was confirmed by variance–mean analysis.The sulpiride-resistant EPSC reduction was mimicked by PD 168,077and blocked by L-745,870, selective D4 receptor agonist and antag-onist, respectively, suggesting the involvement of D4 receptors. Thereduction of EPSCs produced by PD 168,077 was not accompanied bychanges in PPR or the frequency of sEPSCs; however, it was accom-panied by a reduction in mean sEPSC amplitude, indicative of apostsynaptic site of action. These results show that dopamine modu-lates subthalamopallidal excitation by presynaptic D2/3 and postsyn-aptic D4 receptors. The importance of this modulation is discussed.
I N T R O D U C T I O N
The globus pallidus (GP) plays a critical role in the controlof movement exerted by the basal ganglia (DeLong et al. 1985;Mitchell et al. 1987; Turner et al. 1998) through its projectionsto the subthalamic nucleus (STN) and to other basal gangliastructures that are inhibited by GABAergic synaptic inputs(Beckstead 1983; Gandia et al. 1993; Kita and Kita 2001; Kitaand Kitai 1991, 1994; Parent et al. 2000; Sato et al. 2000a;Shinonaga et al. 1992; Windels et al. 2005).
In turn, GP neurons receive and integrate several entries.Their main inhibitory afferents come from the striatum (Cooperand Stanford 2001; Engler et al. 2006; Fonnum et al. 1978;
Gimenez-Amaya and Graybiel 1990; Hazrati and Parent 1992;Kawaguchi et al. 1990; Pan and Walters 1988; Parent et al.1984; Shin et al. 2003; Tremblay and Filion 1989) and fromaxon collaterals that interconnect GP neurons themselves (Kitaet al. 2004; Parent et al. 2000; Shin et al. 2003), although theirmain excitatory inputs come from the STN (Hazrati and Parent1992; Kita and Kitai 1991; Nambu et al. 2000; Sato et al.2000b; Smith et al. 1990) and from thalamic intralaminarnuclei (Mouroux et al. 1997; Yasukawa et al. 2004). Connec-tions between STN and GP form the principal site of recipro-cally innervated excitatory and inhibitory neurons in the basalganglia (Bevan et al. 2002b). When inhibitory and excitatoryneuronal populations are reciprocally innervated, they couldform the basis of a central pattern generator (Grillner 2003;Terman et al. 2002; Yuste et al. 2005) that may transformexcitatory drives into spatiotemporal patterns of oscillatoryactivity for the generation of motor programs (Bevan et al.2002b; Plenz and Kitai 1999).
The above-mentioned afferents converging onto pallidalneurons follow a structured topographical arrangement: the GPnetwork (Hazrati and Parent 1992; Kaneda et al. 2002; Shink etal. 1996; Yasukawa et al. 2004). These inputs are integrated bypallidal neurons with the important contribution of a distinctdopaminergic innervation from neurons of the subtantia nigrapars compacta (Debeir et al. 2005; Gauthier et al. 1999;Lindvall and Bjorklund 1979; Prensa and Parent 2001; Ruskinand Marshall 1997). The way in which dopaminergic innerva-tion modulates the GP network is a matter of study (e.g.,Costall and Naylor 1972; Galvan et al. 2001; Hauber and Lutz1999; Kreiss et al. 1997; Nakanishi et al. 1985; Querejeta et al.2001). For example, a presynaptic modulation of the striato-pallidal projection through D2-class receptors (Cooper andStanford 2001; Floran et al. 1997) was previously described. Inaddition, a postsynaptic modulation of inhibitory inputsthrough D4-receptor activation (Ariano et al. 1997; Mauger etal. 1998; Mrzljak et al. 1996; Shin et al. 2003) was alsoproposed.
Because it was recently shown that subthalamic projectionsto subtantia nigra pars reticulata are presynaptically modulatedby dopamine (Ibanez-Sandoval et al. 2006), this work investi-gated the possibility that the subthalamopallidal connectionswere also modulated by dopamine. Correspondingly, it is
Address for reprint requests and other correspondence: J. Bargas, Biofısica,Instituto de Fisiologıa Celular, UNAM, PO Box 70-253, Mexico City, DF04510 Mexico (E-mail: [email protected]).
The costs of publication of this article were defrayed in part by thepayment of page charges. The article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
J Neurophysiol 96: 2877–2888, 2006.First published August 9, 2006; doi:10.1152/jn.00664.2006.
28770022-3077/06 $8.00 Copyright © 2006 The American Physiological Societywww.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from

shown here that D2/3 and D4 receptor activation modulates thesubthalamopallidal input onto GP neurons. A preliminary re-port of these results was previously presented in abstract form(Hernandez et al. 2004).
M E T H O D S
Preparation of slices
All procedures were carried out in accordance with the NationalInstitutes of Health Guide for Care and Use of Laboratory Animalsand were approved by the Institutional Animal Care Committees ofthe CINVESTAV and UNAM. The experiments were performed onbrain slices obtained from Wistar rats [postnatal day (PD) 14 to PD21]. The rats were anesthetized and decapitated. The brain was rapidlyimmersed for 1 min in cold oxygenated saline (about 4°C; 95%O2-5% CO2) of the following composition (in mM): 124 NaCl, 2.5KCl, 1.3 MgCl2, 1.2 NaH2PO4, 2.4 CaCl2, and 10 glucose. The samesaline but with choline chloride (124 mM) instead of NaCl was usedduring the slicing procedure. Parasagittal slices (300 �m) containingboth GP and STN (see Fig. 1) were cut on a vibroslicer (Lancer,Technical Products International, St. Louis, MO) and transferred tothe saline with NaCl (above). The slices were left for equilibration forat least 1 h in oxygenated saline at room temperature (about 25°C).After equilibration, a single slice was transferred to a recordingchamber placed on the stage of an upright microscope and wascontinuously superfused (2–3 ml min�1) with oxygenated saline atroom temperature.
Whole cell recordings
Recordings were made at room temperature (about 25°C) fromneurons located inside the GP boundaries as seen in the parasagittal
slice (Fig. 1). Neurons were visualized using infrared differentialinterference videomicrosocopy with a �40 water-immersion objec-tive (Hamamatsu C2400-50, Hamamatsu Photonics Systems USA andAxioscop, Carl Zeiss, Oberkochen, Germany.). Micropipettes forwhole cell recordings were pulled (Sutter Instrument, Novato, CA)from borosilicate glass tubes (1.5 mm outer diameter, WPI, Sarasota,FL) for a final resistance of 2–5 M� when filled with internal salineof the following composition (in mM): 120 KSO3CH3, 16 KCl, 2MgCl2, 10 HEPES, 1.1 K2-EGTA, 1.1 ATP-Mg, 1.1 GTP-Na, and 5mM QX-314 (pH 7.3 adjusted with KOH and 287–290 mOsm L�1).QX-314 was used to avoid contamination of synaptic responses byunclamped action currents. Voltage-clamp recordings were made withan Axopatch 200A amplifier (Axon Instruments, Foster City, CA).Liquid junction potentials (�5 mV) were not corrected.
Recordings were acquired at 10 kHz using a Digidata 1200 inter-face (Axon), Bessel filtered at 5 kHz, and analyzed with pCLAMP (v7.0) (Axon). Access resistance (7–20 M�) was monitored continu-ously and experiments were abandoned if changes �20% occurred.
Histochemistry
As described elsewhere (see Ibanez-Sandoval et al. 2006; Vergaraet al. 2003), a sharp micropipette (tip �0.5 �m) filled with externalsaline and 5% biocytin (Sigma-Aldrich, St. Louis, MO) was placedinside the STN borders (near the middle) and left there for 1 h whilethe slice was constantly superfused. Thereafter the pipette was retiredand the slice was superfused with oxygenated saline for another 6 h.The slice was then fixed overnight in 0.1 M phosphate-buffered saline(PBS; pH � 7.4; 4°C) with 4% paraformaldehyde and 1% picric acid.Afterward, slices were infiltrated with 30% sucrose and cut on avibratome (Ted Pella, Reading, CA) into 60-�m sections. Afterwashing with Tris-buffered saline (TBS) containing Triton X-100 andavidin–biotin–peroxidase complex (1:100; Vector Laboratories, Bur-
FIG. 1. Connectivity between globus pal-lidus (GP) and subthalamic nucleus (STN). A:field stimulation inside the STN (see inset)evoked an antidromic action potential in a GPneuron (top). Arrowhead signals stimulus ar-tifact. An action potential was evoked in thesame GP neuron with a depolarizing currentstep (bottom). Arrowhead signals stimulus ar-tifact of field stimulus in STN after this re-sponse; collision blocked the antidromicspike. Note absence of synaptic response afterthe artifact. B: field stimulation inside the GP(see inset) evoked an antidromic action po-tential in a STN neuron (top). Arrowheadsignals stimulus artifact. An action potentialwas evoked in the same STN neuron with adepolarizing current step (bottom). Arrow-head signals stimulus artifact of field stimulusin the GP. Collision blocked the antidromicspike. C: site of biocytin instillment withinthe confines of the STN. Note several filledSTN neurons. D: numerous fibers leaving theSTN and reaching the GP were observed inthe parasagittal slice. E: an axon (white arrowheads) emerging from a labeled STN neuron(black arrow) is shown entering the internalcapsule in route to the GP. F: some GPneurons were also filled retrogradely, withexample shown. This neuron was located atthe site marked by the rectangle in D. NSt,neostriatum; ic, internal capsule. S indicatesthe stimulation electrode.
2878 HERNANDEZ ET AL.
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from

lingame, CA) for 4 h at room temperature, the slices were reacted with3,3�-diaminobenzidine tetrahydrochloride (DAB; 0.05%) and H2O2
(0.003%) in TBS and mounted on slides to visualize the boundhorseradish peroxidase (HRP). This enabled resolution, through trans-illumination microscopy, of labeled neurons inside the STN (Fig. 1C)(Leica Imaging Systems, Cambridge, UK) and, most important, ofabundant subthalamopallidal fibers projecting to their target (Fig. 1, Dand E) (Hazrati and Parent 1992; Sato et al. 2000b; Smith et al. 1990).
Evoked synaptic currents
Synaptic currents were evoked in GP neurons by field stimulationwith concentric bipolar tungsten electrodes (50 �m at the tip, 1 k�DC resistance; FHC, Bowdoinham, ME). The stimulating electrodewas placed either on the STN itself or on the internal capsuleapproaching the caudoventral part of the GP (Ogura and Kita 2000),where subthalamopallidal projections were seen to run in histochem-istry experiments (see Fig. 1D). The likelihood of attaining an excit-atory connection was one out of 10 cases when field stimulation wasin the STN itself. The probability increased to at least eight out of 10cases when field stimulation was in the internal capsule, but decreasedto zero when the stimulus was located a few microns outside this path(Fig. 1, C and D) (Hanson and Jaeger 2002). The frequency ofstimulation was 0.1 Hz and the strength of the pulses was adjusted toobtain roughly 70% of the maximal excitatory postsynaptic current(EPSC) amplitude. The location of dopamine receptors involved in themodulation of the subthalamopallidal transmission was assessed bythree different methods: paired-pulse ratio (PPR), frequency andamplitude of spontaneous EPSCs (sEPSCs), and variance–mean anal-ysis (see following text). Glutamatergic EPSCs were isolated byrecording the currents in the presence of 30 �M bicuculline to avoidcontamination with GABAergic responses (Kita and Kitai 1991; Kitaet al. 2005; Nambu et al. 2000). Unless stated otherwise, a point in thetime-course graphs is the average of six records, at 0.1 Hz (above),which is one trace per 10 s and one point every minute. However,illustrated records of paired-pulse responses are the average of 25consecutive events during steady state. sEPSCs were also recorded inthe presence of bicuculline (30 �M) and tetrodotoxin (TTX, 1 �M).Traces of sEPSCs were filtered at 2 kHz and digitized at 10 kHz. Timeintervals of recorded events during 1.5 min were analyzed (Minianaly-sis, Jaejin Software, Leonia, NJ) in each experimental condition. Theglutamatergic nature of sEPSCs was assessed by their completeelimination by CNQX (30 �M). The N-methyl-D-aspartate (NMDA)–receptor antagonist AP5 (50 �M) was also added in some experimentsbut no qualitative difference was found at this holding potential (�80mV) with experiments that used only CNQX (cf. Ibanez-Sandoval etal. 2006).
When the paired-pulse protocol was performed with field stimula-tion, interpulse intervals of about 50 ms were used. Changes in thepaired-pulse ratio (PPR) of EPSC responses (PPR � second EPSC/first EPSC) are known to be linearly proportional to the probability ofrelease and the paired-pulse protocol was demonstrated to detectpresynaptic actions of transmitters (e.g., Baldelli et al. 2005; Bolsha-kov and Siegelbaum 1995; Dunwiddie and Hass 1985; Guzman et al.2003; Ibanez-Sandoval et al. 2006; Kamiya and Zucker 1994; Salgadoet al. 2005; Zucker 1999). Nonstationary fluctuations analysis wasperformed from 60 consecutive 50-Hz trains evoked at 0.1 Hz toperform variance–mean analyses in which mean amplitudes of evokedEPSCs (corrected for basal lines from previous EPSCs) were plottedagainst their peak variance: variance–mean (V–M) plots (Clementsand Silver 2000; Ibanez-Sandoval et al. 2006; Koos et al. 2004;Salgado et al. 2005). Then, a parabola of the form
y � Ax � Bx2 (1)
was fitted with a Marquardt algorithm (Fig. 9). where y representsevoked EPSC peak variance (ordinates), x represents EPSC mean
peak amplitude (abscissae), and A and B are free parameters. Basi-cally, parameter A indicates the initial slope of the parabola andparameter B depends on the width of the parabola. From this fit, aweighted average of the quantal amplitude (Qw) was obtained
Qw � A/�1 � CV2 (2)
where CV is the coefficient of variation of EPSC amplitudes. Inaddition, the approximate number of release sites (N) and the averageprobability of release (Pr) across release sites (assuming a binomialdistribution) can be approximated by
N � 1/B (3)
Pr � x�B/A�1 � CV2 (4)
Statistics
All data are given as means SE unless stated otherwise. Signif-icance of the effect of drugs as well as the significance of thedifferences in paired-pulse ratios was tested with nonparametricstatistics: the Wilcoxon t-test or Mann–Whitney’s U test, dependingon paired or unpaired samples. When the same sample had more thanone treatment Friedman’s statistics with post hoc Student–Newman–Keuls test was used. Statistical differences of fitted functions wereassessed by comparing the obtained parameters and their estimationerrors with Student’s t-test.
Effects on sEPSCs were estimated by Kolmogorov–Smirnov two-sample test from cumulative distribution amplitudes or cumulativedistributions of interevent intervals (Minianalysis).
Drugs
Drugs were stored in the freezer as dry aliquots and stock solutionswere prepared just before each experiment and added to the perfusionsolution in the final concentration. Bicuculline methiodide, 3,4-dihy-droxyphenethylamine (dopamine), trans-(�)-4aR-4,4a,5,6,7,8,8a,9-octahydro-5-propyl-1H-pyrazolo(3,4-g)quinoline (quinpirole), S-(�)-5-amino-sulfonyl-N-[(1-ethyl-2-pyrrolidinyl)-methyl]-2-methoxyben-zamide (sulpiride), 6-cyano-7-nitroquinoxalline-2,3-dione (CNQX),D-(�)-2-amino-5-phosphonopentanoic acid (AP5), N-[[4-(2-cyanophenyl)-1-piperazinyl]methyl]-3-methyl-benzamide maleate(PD 168,077), (3-[(4-[4-chlorophenyl]piperazin-1-yl)methyl]-1H-pyrrolo[2,3-b]pyridine) hydrochloride (L-745,870), lidocaine N-ethylbromide (QX-314), and tetrodotoxin (TTX) were obtained fromSigma (St. Louis, MO). Drugs were diluted into the bath solution.
R E S U L T S
Anatomical connectivity of the GP and STN neurons
All recordings were made from neurons situated inside theGP borders, during field stimulation of the subthalamopallidalpath (see METHODS). It is known that there are different popu-lations of neurons in the GP (Cooper and Stanford 2000; Kitaand Kitai 1994; Nambu and Llinas 1997). They can expressGAD, calbindin, parvalbumin, or enkephalin (Hoover andMarshall 1999; Kita and Kita 2001; Rajakumar et al. 1994;Voorn et al. 1999). Field stimulation inside the STN borders orin the internal capsule (see METHODS) could evoke antidromicaction potentials in some GP neurons, sometimes withoutevoking an excitatory postsynaptic potential (EPSP) with anorthodromic action potential (Fig. 1A, top; stimulus artifact issignaled with an arrowhead). This antidromic action potentialcould collide with an action potential evoked with a depolar-izing current injection [Fig. 1A, bottom; artifact of antidromicstimulation (arrowhead) can be seen inside the refractory
2879DOPAMINE CONTROLS THE SUBTHALAMIC INPUT TO THE PALLIDUM
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from

period of the direct spike; note the absence of synaptic re-sponse or autorregenerative spike] (Kita and Kitai 1991). Onthe other hand, field stimulation inside the GP borders couldevoke antidromic action potentials in some STN neurons,sometimes without evoking an inhibitory postsynaptic poten-tial (IPSP) (Fig. 1B, top; stimulus artifact is signaled with anarrowhead). This antidromic action potential could collide withan action potential evoked with a depolarizing current injectedon a STN neuron (Fig. 1B, bottom; arrowhead signals theartifact of antidromic stimulation). Staining after anterogradetransport of biocytin injected inside the STN borders (Fig. 1C;see labeled STN neurons around the site of injection) (seeIbanez-Sandoval et al. 2006) can disclose subthalamopallidalfibers in a parasagittal brain slice that includes both STN andGP (Fig. 1D). Axons emerging from STN neurons had avaricose appearance and could be followed entering the inter-nal capsule toward the GP with other labeled fibers (Fig. 1E;white arrows). In some cases, retrogradely labeled GP neuronscould also be observed (Fig. 1F), with morphology similar tothat described previously (Kita and Kita 2001; Kita and Kitai1994; Ogura and Kita 2000), i.e.: variable (oval or triangular)shape with three to five primary dendrites. Thus abundantanterograde labeling and some retrograde labeling, togetherwith antidromic activation in either way, suggested that somereciprocal connections between the GP and STN are preservedin a parasagittal brain slice of the present characteristics (seeMETHODS) (Ogura and Kita 2000). Taken together, these results(n � 6 slices) suggested that field stimulation in the presentconditions is mostly the product of the orthodromic activationof the subthalamopallidal pathway in vitro (see also Hansonand Jaeger 2002).
Glutamatergic synaptic responses recorded in GP neuronsafter stimulation of the subthalamopallidal pathway
The electrophysiological properties of GP neurons werethoroughly characterized in previous studies (e.g., Chan et al.2004; Cooper and Stanford 2000; Kita and Kitai 1991; Nambuand Llinas 1994; Overton and Greenfield 1995). During thepresent work, neurons of previously described classes wererecorded. For example, voltage records of a representativeneuron (without QX-314) that responded to subthalamopallidalstimulation are depicted in Fig. 2A. As in the case of thisneuron (Fig. 2A), many neurons recorded inside the GP bordersexhibited a characteristic slowly developing voltage- and time-dependent rectification when injected with hyperpolarizingcurrent pulses fired spontaneously, could fire at high frequen-cies, and showed a rebound firing after hyperpolarizing pulses(Chan et al. 2004; Cooper and Stanford 2000; Nambu andLlinas 1994). However, QX-314 was used in most cases toavoid contamination of the synaptic response with orthodromicunclamped active currents (see METHODS) (Kita and Kitai 1991;Kita et al. 2005; Nambu et al. 2000). Therefore in most cases,we did not characterize, electrophysiologically, the type ofpostsynaptic neuron recorded. Second, the criterion to acceptan experiment was the synaptic response to subthalamopallidalpathway stimulation (see above). For these reasons, our samplecould be biased toward some neuronal classes, i.e., those thatpreferentially receive STN inputs (Hoover and Marshall 1999;Kita and Kita 2001; Rajakumar 1994). The dopaminergic
modulation of the synaptic responses described below did notappear to differ from one experiment to the other once theconnection was stable.
Synaptic currents onto another neuron (in the presence ofQX-314; see METHODS), evoked by stimulating the subthalamo-pallidal pathway, are illustrated in the inset of Fig. 2B. Thesecurrents were recorded at different holding potentials in thepresence of bicuculline (30 �M) and AP5 (50 �M). Thecurrent–voltage relationship (I–V plot) for the peak EPSCamplitude had a linear relationship with a reversal potentialaround 0 mV (Fig. 2B). This I–V plot is typical of synapticcurrents flowing through �-amino-3-hydroxy-5-methyl-4-isox-azolepropionic acid (AMPA)/kainate receptor channels (e.g.,Jonas et al. 1993). In fact, most EPSC was carried by AMPA/kainate receptors in these conditions as confirmed by CNQX(10 �M) blockage (Fig. 2C), which practically abolished mostof the synaptic current when evoked at a holding potential of�80 mV. Subsequent addition of AP5 (50 �M), a selectiveblocker of NMDA receptor channels, eliminated a minimalresidual current (Fig. 2C, circles) (cf. Ibanez-Sandoval et al.2006). The actions of both blockers were reversible (Fig. 2C).When no blockers were added, currents were seen to remainstable for �30 min with these stimulation parameters (Fig. 2C,triangles). In the present conditions, an EPSC of these charac-teristics is expected, given the previous information about thesubthalamopallidal connection (see Kita and Kitai 1991; Kitaet al. 2005; Nambu et al. 2000).
FIG. 2. Subthalamopallidal excitatory postsynaptic current (EPSC). A: volt-age responses of one class of GP neuron to a series of current pulses. Neuronfires with little or no accommodation, exhibits a voltage-sag with hyperpolar-izing current injection, and shows a rebound firing on cessation of hyperpo-larization. B: stimulation of the subthalamopallidal pathway evokes synapticcurrents (inset) on a GP neuron (QX-314 avoids uncampled firing). Peakcurrent amplitude of these currents changed with holding potential. A current–voltage relationship (I–V plot) of these currents shows a reversal potential atabout 0 mV. D-(�)-2-amino-5-phosphonopentanoic acid (AP5, 50 �M) waspresent throughout the experiment. C: time course of synaptic current evokedat a holding potential of �80 mV. 6-Cyano-7-nitroquinoxaline-2,3-dione(CNQX, 10 �M), a selective antagonist for �-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA)/kainate receptors virtually abolished thiscurrent. Addition of AP5 (50 �M) blocked a minimal residual current. D:representative traces taken at indicated numbers during the time course of theexperiment. Bicuculline (30 �M) was present throughout.
2880 HERNANDEZ ET AL.
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from
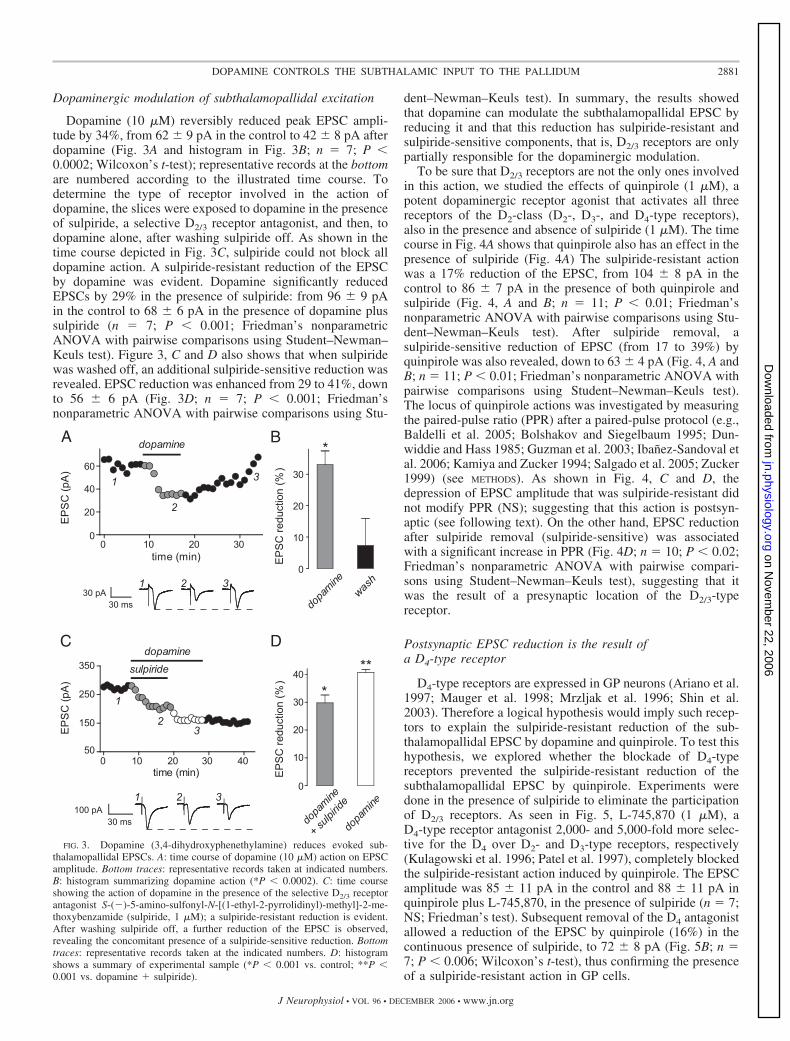
Dopaminergic modulation of subthalamopallidal excitation
Dopamine (10 �M) reversibly reduced peak EPSC ampli-tude by 34%, from 62 9 pA in the control to 42 8 pA afterdopamine (Fig. 3A and histogram in Fig. 3B; n � 7; P �0.0002; Wilcoxon’s t-test); representative records at the bottomare numbered according to the illustrated time course. Todetermine the type of receptor involved in the action ofdopamine, the slices were exposed to dopamine in the presenceof sulpiride, a selective D2/3 receptor antagonist, and then, todopamine alone, after washing sulpiride off. As shown in thetime course depicted in Fig. 3C, sulpiride could not block alldopamine action. A sulpiride-resistant reduction of the EPSCby dopamine was evident. Dopamine significantly reducedEPSCs by 29% in the presence of sulpiride: from 96 9 pAin the control to 68 6 pA in the presence of dopamine plussulpiride (n � 7; P � 0.001; Friedman’s nonparametricANOVA with pairwise comparisons using Student–Newman–Keuls test). Figure 3, C and D also shows that when sulpiridewas washed off, an additional sulpiride-sensitive reduction wasrevealed. EPSC reduction was enhanced from 29 to 41%, downto 56 6 pA (Fig. 3D; n � 7; P � 0.001; Friedman’snonparametric ANOVA with pairwise comparisons using Stu-
dent–Newman–Keuls test). In summary, the results showedthat dopamine can modulate the subthalamopallidal EPSC byreducing it and that this reduction has sulpiride-resistant andsulpiride-sensitive components, that is, D2/3 receptors are onlypartially responsible for the dopaminergic modulation.
To be sure that D2/3 receptors are not the only ones involvedin this action, we studied the effects of quinpirole (1 �M), apotent dopaminergic receptor agonist that activates all threereceptors of the D2-class (D2-, D3-, and D4-type receptors),also in the presence and absence of sulpiride (1 �M). The timecourse in Fig. 4A shows that quinpirole also has an effect in thepresence of sulpiride (Fig. 4A) The sulpiride-resistant actionwas a 17% reduction of the EPSC, from 104 8 pA in thecontrol to 86 7 pA in the presence of both quinpirole andsulpiride (Fig. 4, A and B; n � 11; P � 0.01; Friedman’snonparametric ANOVA with pairwise comparisons using Stu-dent–Newman–Keuls test). After sulpiride removal, asulpiride-sensitive reduction of EPSC (from 17 to 39%) byquinpirole was also revealed, down to 63 4 pA (Fig. 4, A andB; n � 11; P � 0.01; Friedman’s nonparametric ANOVA withpairwise comparisons using Student–Newman–Keuls test).The locus of quinpirole actions was investigated by measuringthe paired-pulse ratio (PPR) after a paired-pulse protocol (e.g.,Baldelli et al. 2005; Bolshakov and Siegelbaum 1995; Dun-widdie and Hass 1985; Guzman et al. 2003; Ibanez-Sandoval etal. 2006; Kamiya and Zucker 1994; Salgado et al. 2005; Zucker1999) (see METHODS). As shown in Fig. 4, C and D, thedepression of EPSC amplitude that was sulpiride-resistant didnot modify PPR (NS); suggesting that this action is postsyn-aptic (see following text). On the other hand, EPSC reductionafter sulpiride removal (sulpiride-sensitive) was associatedwith a significant increase in PPR (Fig. 4D; n � 10; P � 0.02;Friedman’s nonparametric ANOVA with pairwise compari-sons using Student–Newman–Keuls test), suggesting that itwas the result of a presynaptic location of the D2/3-typereceptor.
Postsynaptic EPSC reduction is the result ofa D4-type receptor
D4-type receptors are expressed in GP neurons (Ariano et al.1997; Mauger et al. 1998; Mrzljak et al. 1996; Shin et al.2003). Therefore a logical hypothesis would imply such recep-tors to explain the sulpiride-resistant reduction of the sub-thalamopallidal EPSC by dopamine and quinpirole. To test thishypothesis, we explored whether the blockade of D4-typereceptors prevented the sulpiride-resistant reduction of thesubthalamopallidal EPSC by quinpirole. Experiments weredone in the presence of sulpiride to eliminate the participationof D2/3 receptors. As seen in Fig. 5, L-745,870 (1 �M), aD4-type receptor antagonist 2,000- and 5,000-fold more selec-tive for the D4 over D2- and D3-type receptors, respectively(Kulagowski et al. 1996; Patel et al. 1997), completely blockedthe sulpiride-resistant action induced by quinpirole. The EPSCamplitude was 85 11 pA in the control and 88 11 pA inquinpirole plus L-745,870, in the presence of sulpiride (n � 7;NS; Friedman’s test). Subsequent removal of the D4 antagonistallowed a reduction of the EPSC by quinpirole (16%) in thecontinuous presence of sulpiride, to 72 8 pA (Fig. 5B; n �7; P � 0.006; Wilcoxon’s t-test), thus confirming the presenceof a sulpiride-resistant action in GP cells.
FIG. 3. Dopamine (3,4-dihydroxyphenethylamine) reduces evoked sub-thalamopallidal EPSCs. A: time course of dopamine (10 �M) action on EPSCamplitude. Bottom traces: representative records taken at indicated numbers.B: histogram summarizing dopamine action (*P � 0.0002). C: time courseshowing the action of dopamine in the presence of the selective D2/3 receptorantagonist S-(�)-5-amino-sulfonyl-N-[(1-ethyl-2-pyrrolidinyl)-methyl]-2-me-thoxybenzamide (sulpiride, 1 �M); a sulpiride-resistant reduction is evident.After washing sulpiride off, a further reduction of the EPSC is observed,revealing the concomitant presence of a sulpiride-sensitive reduction. Bottomtraces: representative records taken at the indicated numbers. D: histogramshows a summary of experimental sample (*P � 0.001 vs. control; **P �0.001 vs. dopamine � sulpiride).
2881DOPAMINE CONTROLS THE SUBTHALAMIC INPUT TO THE PALLIDUM
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from
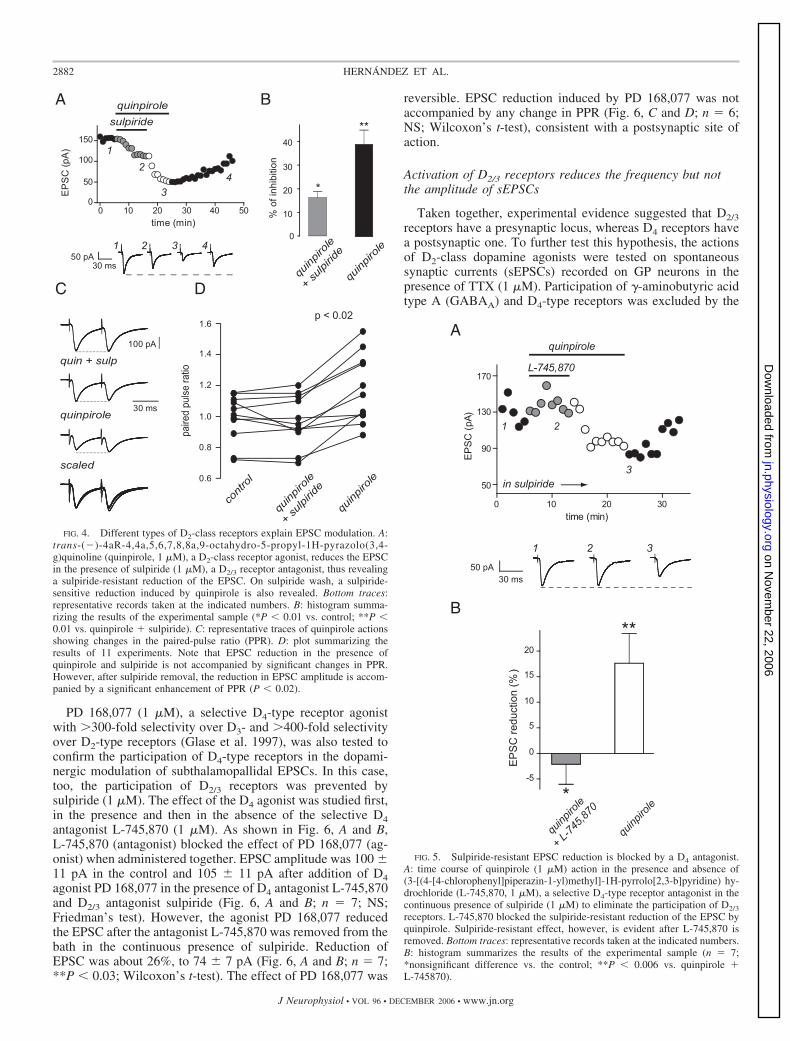
PD 168,077 (1 �M), a selective D4-type receptor agonistwith �300-fold selectivity over D3- and �400-fold selectivityover D2-type receptors (Glase et al. 1997), was also tested toconfirm the participation of D4-type receptors in the dopami-nergic modulation of subthalamopallidal EPSCs. In this case,too, the participation of D2/3 receptors was prevented bysulpiride (1 �M). The effect of the D4 agonist was studied first,in the presence and then in the absence of the selective D4antagonist L-745,870 (1 �M). As shown in Fig. 6, A and B,L-745,870 (antagonist) blocked the effect of PD 168,077 (ag-onist) when administered together. EPSC amplitude was 100 11 pA in the control and 105 11 pA after addition of D4agonist PD 168,077 in the presence of D4 antagonist L-745,870and D2/3 antagonist sulpiride (Fig. 6, A and B; n � 7; NS;Friedman’s test). However, the agonist PD 168,077 reducedthe EPSC after the antagonist L-745,870 was removed from thebath in the continuous presence of sulpiride. Reduction ofEPSC was about 26%, to 74 7 pA (Fig. 6, A and B; n � 7;**P � 0.03; Wilcoxon’s t-test). The effect of PD 168,077 was
reversible. EPSC reduction induced by PD 168,077 was notaccompanied by any change in PPR (Fig. 6, C and D; n � 6;NS; Wilcoxon’s t-test), consistent with a postsynaptic site ofaction.
Activation of D2/3 receptors reduces the frequency but notthe amplitude of sEPSCs
Taken together, experimental evidence suggested that D2/3receptors have a presynaptic locus, whereas D4 receptors havea postsynaptic one. To further test this hypothesis, the actionsof D2-class dopamine agonists were tested on spontaneoussynaptic currents (sEPSCs) recorded on GP neurons in thepresence of TTX (1 �M). Participation of �-aminobutyric acidtype A (GABAA) and D4-type receptors was excluded by the
FIG. 4. Different types of D2-class receptors explain EPSC modulation. A:trans-(�)-4aR-4,4a,5,6,7,8,8a,9-octahydro-5-propyl-1H-pyrazolo(3,4-g)quinoline (quinpirole, 1 �M), a D2-class receptor agonist, reduces the EPSCin the presence of sulpiride (1 �M), a D2/3 receptor antagonist, thus revealinga sulpiride-resistant reduction of the EPSC. On sulpiride wash, a sulpiride-sensitive reduction induced by quinpirole is also revealed. Bottom traces:representative records taken at the indicated numbers. B: histogram summa-rizing the results of the experimental sample (*P � 0.01 vs. control; **P �0.01 vs. quinpirole � sulpiride). C: representative traces of quinpirole actionsshowing changes in the paired-pulse ratio (PPR). D: plot summarizing theresults of 11 experiments. Note that EPSC reduction in the presence ofquinpirole and sulpiride is not accompanied by significant changes in PPR.However, after sulpiride removal, the reduction in EPSC amplitude is accom-panied by a significant enhancement of PPR (P � 0.02).
FIG. 5. Sulpiride-resistant EPSC reduction is blocked by a D4 antagonist.A: time course of quinpirole (1 �M) action in the presence and absence of(3-[(4-[4-chlorophenyl]piperazin-1-yl)methyl]-1H-pyrrolo[2,3-b]pyridine) hy-drochloride (L-745,870, 1 �M), a selective D4-type receptor antagonist in thecontinuous presence of sulpiride (1 �M) to eliminate the participation of D2/3
receptors. L-745,870 blocked the sulpiride-resistant reduction of the EPSC byquinpirole. Sulpiride-resistant effect, however, is evident after L-745,870 isremoved. Bottom traces: representative records taken at the indicated numbers.B: histogram summarizes the results of the experimental sample (n � 7;*nonsignificant difference vs. the control; **P � 0.006 vs. quinpirole �L-745870).
2882 HERNANDEZ ET AL.
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from
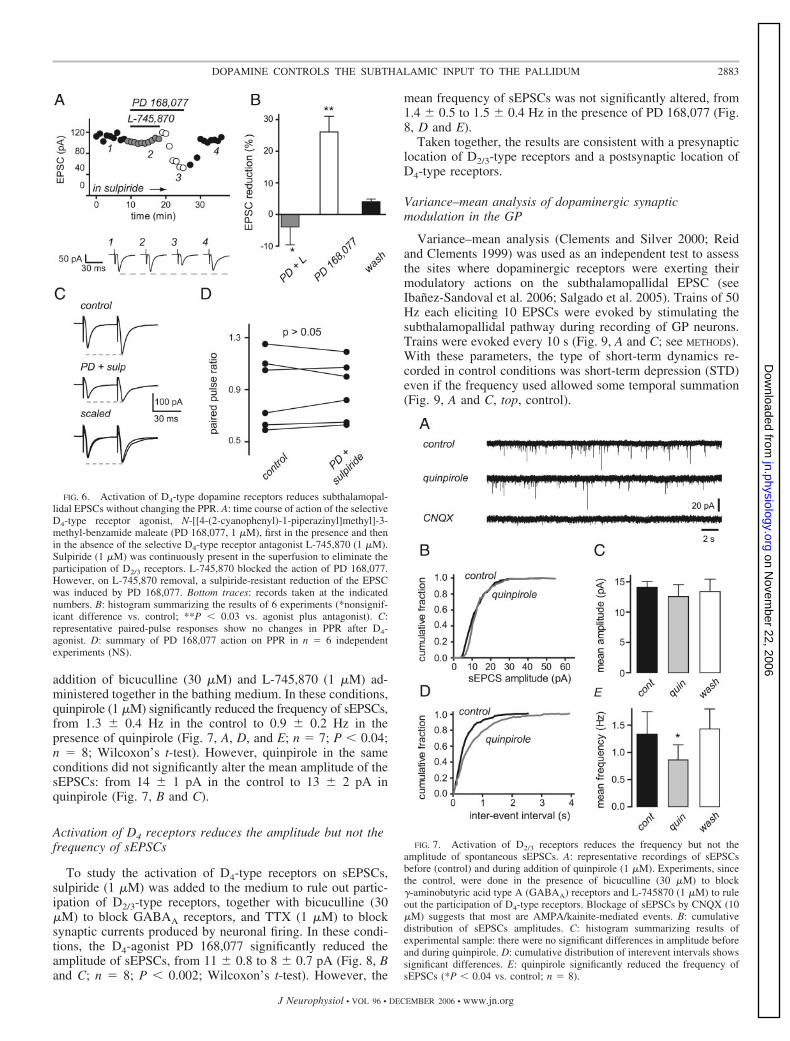
addition of bicuculline (30 �M) and L-745,870 (1 �M) ad-ministered together in the bathing medium. In these conditions,quinpirole (1 �M) significantly reduced the frequency of sEPSCs,from 1.3 0.4 Hz in the control to 0.9 0.2 Hz in thepresence of quinpirole (Fig. 7, A, D, and E; n � 7; P � 0.04;n � 8; Wilcoxon’s t-test). However, quinpirole in the sameconditions did not significantly alter the mean amplitude of thesEPSCs: from 14 1 pA in the control to 13 2 pA inquinpirole (Fig. 7, B and C).
Activation of D4 receptors reduces the amplitude but not thefrequency of sEPSCs
To study the activation of D4-type receptors on sEPSCs,sulpiride (1 �M) was added to the medium to rule out partic-ipation of D2/3-type receptors, together with bicuculline (30�M) to block GABAA receptors, and TTX (1 �M) to blocksynaptic currents produced by neuronal firing. In these condi-tions, the D4-agonist PD 168,077 significantly reduced theamplitude of sEPSCs, from 11 0.8 to 8 0.7 pA (Fig. 8, Band C; n � 8; P � 0.002; Wilcoxon’s t-test). However, the
mean frequency of sEPSCs was not significantly altered, from1.4 0.5 to 1.5 0.4 Hz in the presence of PD 168,077 (Fig.8, D and E).
Taken together, the results are consistent with a presynapticlocation of D2/3-type receptors and a postsynaptic location ofD4-type receptors.
Variance–mean analysis of dopaminergic synapticmodulation in the GP
Variance–mean analysis (Clements and Silver 2000; Reidand Clements 1999) was used as an independent test to assessthe sites where dopaminergic receptors were exerting theirmodulatory actions on the subthalamopallidal EPSC (seeIbanez-Sandoval et al. 2006; Salgado et al. 2005). Trains of 50Hz each eliciting 10 EPSCs were evoked by stimulating thesubthalamopallidal pathway during recording of GP neurons.Trains were evoked every 10 s (Fig. 9, A and C; see METHODS).With these parameters, the type of short-term dynamics re-corded in control conditions was short-term depression (STD)even if the frequency used allowed some temporal summation(Fig. 9, A and C, top, control).
FIG. 7. Activation of D2/3 receptors reduces the frequency but not theamplitude of spontaneous sEPSCs. A: representative recordings of sEPSCsbefore (control) and during addition of quinpirole (1 �M). Experiments, sincethe control, were done in the presence of bicuculline (30 �M) to block�-aminobutyric acid type A (GABAA) receptors and L-745870 (1 �M) to ruleout the participation of D4-type receptors. Blockage of sEPSCs by CNQX (10�M) suggests that most are AMPA/kainite-mediated events. B: cumulativedistribution of sEPSCs amplitudes. C: histogram summarizing results ofexperimental sample: there were no significant differences in amplitude beforeand during quinpirole. D: cumulative distribution of interevent intervals showssignificant differences. E: quinpirole significantly reduced the frequency ofsEPSCs (*P � 0.04 vs. control; n � 8).
FIG. 6. Activation of D4-type dopamine receptors reduces subthalamopal-lidal EPSCs without changing the PPR. A: time course of action of the selectiveD4-type receptor agonist, N-[[4-(2-cyanophenyl)-1-piperazinyl]methyl]-3-methyl-benzamide maleate (PD 168,077, 1 �M), first in the presence and thenin the absence of the selective D4-type receptor antagonist L-745,870 (1 �M).Sulpiride (1 �M) was continuously present in the superfusion to eliminate theparticipation of D2/3 receptors. L-745,870 blocked the action of PD 168,077.However, on L-745,870 removal, a sulpiride-resistant reduction of the EPSCwas induced by PD 168,077. Bottom traces: records taken at the indicatednumbers. B: histogram summarizing the results of 6 experiments (*nonsignif-icant difference vs. control; **P � 0.03 vs. agonist plus antagonist). C:representative paired-pulse responses show no changes in PPR after D4-agonist. D: summary of PD 168,077 action on PPR in n � 6 independentexperiments (NS).
2883DOPAMINE CONTROLS THE SUBTHALAMIC INPUT TO THE PALLIDUM
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from
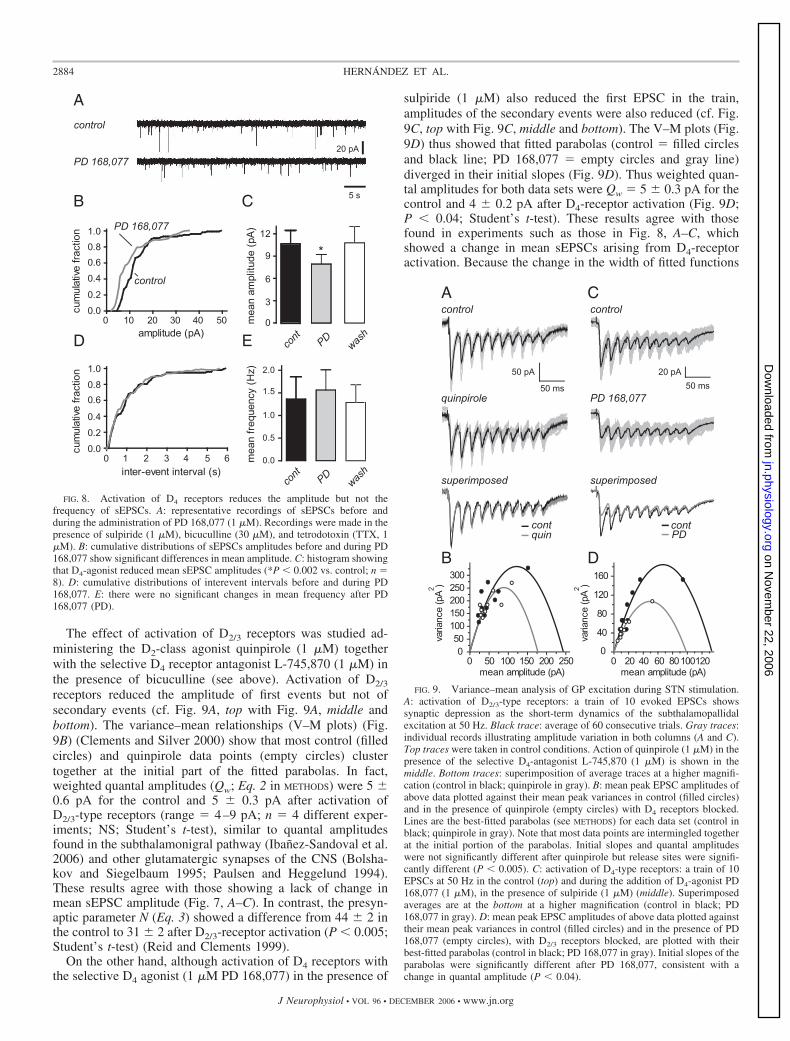
The effect of activation of D2/3 receptors was studied ad-ministering the D2-class agonist quinpirole (1 �M) togetherwith the selective D4 receptor antagonist L-745,870 (1 �M) inthe presence of bicuculline (see above). Activation of D2/3
receptors reduced the amplitude of first events but not ofsecondary events (cf. Fig. 9A, top with Fig. 9A, middle andbottom). The variance–mean relationships (V–M plots) (Fig.9B) (Clements and Silver 2000) show that most control (filledcircles) and quinpirole data points (empty circles) clustertogether at the initial part of the fitted parabolas. In fact,weighted quantal amplitudes (Qw; Eq. 2 in METHODS) were 5 0.6 pA for the control and 5 0.3 pA after activation ofD2/3-type receptors (range � 4–9 pA; n � 4 different exper-iments; NS; Student’s t-test), similar to quantal amplitudesfound in the subthalamonigral pathway (Ibanez-Sandoval et al.2006) and other glutamatergic synapses of the CNS (Bolsha-kov and Siegelbaum 1995; Paulsen and Heggelund 1994).These results agree with those showing a lack of change inmean sEPSC amplitude (Fig. 7, A–C). In contrast, the presyn-aptic parameter N (Eq. 3) showed a difference from 44 2 inthe control to 31 2 after D2/3-receptor activation (P � 0.005;Student’s t-test) (Reid and Clements 1999).
On the other hand, although activation of D4 receptors withthe selective D4 agonist (1 �M PD 168,077) in the presence of
sulpiride (1 �M) also reduced the first EPSC in the train,amplitudes of the secondary events were also reduced (cf. Fig.9C, top with Fig. 9C, middle and bottom). The V–M plots (Fig.9D) thus showed that fitted parabolas (control � filled circlesand black line; PD 168,077 � empty circles and gray line)diverged in their initial slopes (Fig. 9D). Thus weighted quan-tal amplitudes for both data sets were Qw � 5 0.3 pA for thecontrol and 4 0.2 pA after D4-receptor activation (Fig. 9D;P � 0.04; Student’s t-test). These results agree with thosefound in experiments such as those in Fig. 8, A–C, whichshowed a change in mean sEPSCs arising from D4-receptoractivation. Because the change in the width of fitted functions
FIG. 9. Variance–mean analysis of GP excitation during STN stimulation.A: activation of D2/3-type receptors: a train of 10 evoked EPSCs showssynaptic depression as the short-term dynamics of the subthalamopallidalexcitation at 50 Hz. Black trace: average of 60 consecutive trials. Gray traces:individual records illustrating amplitude variation in both columns (A and C).Top traces were taken in control conditions. Action of quinpirole (1 �M) in thepresence of the selective D4-antagonist L-745,870 (1 �M) is shown in themiddle. Bottom traces: superimposition of average traces at a higher magnifi-cation (control in black; quinpirole in gray). B: mean peak EPSC amplitudes ofabove data plotted against their mean peak variances in control (filled circles)and in the presence of quinpirole (empty circles) with D4 receptors blocked.Lines are the best-fitted parabolas (see METHODS) for each data set (control inblack; quinpirole in gray). Note that most data points are intermingled togetherat the initial portion of the parabolas. Initial slopes and quantal amplitudeswere not significantly different after quinpirole but release sites were signifi-cantly different (P � 0.005). C: activation of D4-type receptors: a train of 10EPSCs at 50 Hz in the control (top) and during the addition of D4-agonist PD168,077 (1 �M), in the presence of sulpiride (1 �M) (middle). Superimposedaverages are at the bottom at a higher magnification (control in black; PD168,077 in gray). D: mean peak EPSC amplitudes of above data plotted againsttheir mean peak variances in control (filled circles) and in the presence of PD168,077 (empty circles), with D2/3 receptors blocked, are plotted with theirbest-fitted parabolas (control in black; PD 168,077 in gray). Initial slopes of theparabolas were significantly different after PD 168,077, consistent with achange in quantal amplitude (P � 0.04).
FIG. 8. Activation of D4 receptors reduces the amplitude but not thefrequency of sEPSCs. A: representative recordings of sEPSCs before andduring the administration of PD 168,077 (1 �M). Recordings were made in thepresence of sulpiride (1 �M), bicuculline (30 �M), and tetrodotoxin (TTX, 1�M). B: cumulative distributions of sEPSCs amplitudes before and during PD168,077 show significant differences in mean amplitude. C: histogram showingthat D4-agonist reduced mean sEPSC amplitudes (*P � 0.002 vs. control; n �8). D: cumulative distributions of interevent intervals before and during PD168,077. E: there were no significant changes in mean frequency after PD168,077 (PD).
2884 HERNANDEZ ET AL.
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from

was accompanied with a change in initial slope, calculations ofthe presynaptic parameter N did not show any difference: 24 2 in the control versus 23 2 in PD 168,077 (Clements andSilver 2000). Further quantal analysis may be performed in thefuture to assess agonist effects on release probabilities becauseaverage probabilities (Eq. 4) did not show any difference withthis analysis (e.g., Salgado et al. 2005). For the goals of thepresent work, however, variance–mean analysis was consistentwith the view that dopaminergic modulation of subthalamo-pallidal transmission occurs at both presynaptic (D2/3 recep-tors) and postsynaptic sites (D4 receptors).
D I S C U S S I O N
Reciprocal connections between STN and GP are a source ofrhythmicity and physiological or pathological oscillations inneuronal firing (Bevan et al. 2002b; Plenz and Kitai 1999;Terman et al. 2002). In this work, biocytin anterograde andretrograde labeling, as well as antidromic activation, demon-strated that some reciprocal connections between GP and STN(Gandia et al. 1993; Hazrati and Parent 1992; Kita and Kita1991, 2001; Kita et al. 2004; Nambu et al. 2000; Parent et al.2000; Sato et al. 2000b; Shink et al. 1996; Shinonaga et al.1992; Smith et al. 1990) can functionally be preserved in invitro slice preparations (Beurrier et al. 2006; Hanson andJaeger 2002: Ogura and Kita 2000). Furthermore, moderate-strength orthodromic stimulation of the subthalamopallidalpathway in the presence of bicuculline to block inhibitoryinputs and intracellular QX-314 to block unclamped actioncurrents in the recorded cell can isolate single excitatorypostsynaptic currents (EPSCs) that, at �80 mV holding poten-tial, were mostly the product of AMPA/kainate receptor acti-vation (Ogura and Kita 2000). The main finding of this workwas that dopamine in the globus pallidus, presynaptically andpostsynaptically, modulates this excitatory subthalamopallidalinput. Both D2/3- and D4-type receptors participate, thus de-creasing the magnitude of the synaptic response. D2/3-typereceptors appear to be located presynaptically on subthalamo-pallidal afferents, whereas D4-type receptors appear to belocated postsynaptically on GP neurons. Physiologically, bothreceptors appear to act simultaneously to control the excitatorytone and the firing of GP neurons. Because STN also projectsto the subtantia nigra pars compacta (Iribe et al. 1999; Ka-nazawa et al. 1976; Smith and Grace 1992), the results thensuggest that subthalamic efferents to the GP may autocontroltheir output through the simultaneous activation of dopaminer-gic modulation.
Presynaptic control of subthalamic inputs by D2/3 receptors
Subthalamic neurons express both D1-class and D2-classreceptors (Baufreton et al. 2003; Ciliax et al. 2000; Dawson etal. 1986; Flores et al. 1999; Hurd et al. 2001; Johnson et al.1994; Svenningsson and Le Moine 2002), which mediatedopamine actions that modulate the firing of STN neurons(Baufreton et al. 2003; Kreiss et al. 1996, 1997; Mintz et al.1986; Ni et al. 2001; Svenningsson and Le Moine 2002; Zhu etal. 2002). Both receptor classes may be transported to thesynaptic terminals of axons arising from STN projection neu-rons, for example, to presynaptically modulate subthalamoni-gral inputs to the reticulata (Ibanez-Sandoval et al. 2006). This
suggests that they could also be present in the terminals of thesubthalamopallidal efferents.
In the present work we chose to study the subthalamopallidalmodulation by D2-class receptors only. This decision wasinfluenced by the fact that GP neurons express D4-type recep-tors (Ariano et al. 1997; Floran et al. 2004; Mauger et al. 1998;Mrzljak et al. 1996) and it was previously shown that D4receptors located on GP neurons exert an inhibitory control onGABAergic inputs (Shin et al. 2003). Moreover, the GP alsoreceives glutamatergic excitatory inputs from the thalamus(Mouroux et al. 1997; Yasukawa et al. 2004). Although tha-lamic afferents do not express D2/3 receptors, they may expressD5-type receptors (Meador-Woodruff et al. 1992), which mayalso be the most important type of D1-class receptor present inSTN neurons (Baufreton et al. 2003; Ciliax et al. 2000; Sven-ningsson and Le Moine 2002). Therefore modulation by D1- orD2-class receptors needs to be carefully separated to under-stand their actions in the GP.
This work reports that both dopamine and quinpirole, theD2-class nonselective receptor agonist, reduced the sub-thalamopallidal-evoked EPSCs. In both cases this reductionwas only partially blocked by the D2/3 receptor antagonistsulpiride, thus revealing a potentially important sulpiride-re-sistant action of dopamine D2-class agonists. Even at highconcentrations (e.g., 10 �M) sulpiride does not block D4receptors (Price and Pittman 2001; Shin et al. 2003; Wang et al.2002) and, because GP neurons express D4-type receptors, itwas logical to hypothesize that the sulpiride-resistant actionwas mediated by D4 receptors (see following text).
The sulpiride-sensitive action could then be mediated byD2/3 receptors located in the subthalamopallidal pathway (Cooperand Stanford 2001; Ibanez-Sandoval et al. 2006). The presentwork supported this hypothesis with three kinds of experimen-tal evidence. First, the sulpiride-sensitive reduction of synapticcurrents was associated with a change in the paired-pulse ratio,consistent with a presynaptic site of action (e.g., Baldelli et al.2005; Dunwiddie and Haas 1985; Guzman et al. 2003; Ibanez-Sandoval et al. 2006; Kamiya and Zucker 1994; Price andPittman 2001; Salgado et al. 2005; Zucker 1999). Second,selective activation of D2/3 receptors changed the frequencybut not the amplitude of sEPSCs (Cooper and Stanford 2001;Koga and Momiyama 2000; see also Mann-Metzer and Yarom2002; Sahara and Takahashi 2001). Finally, variance–meananalysis (Clements and Silver 2000; Ibanez-Sandoval et al.2006; Koos et al. 2004; Reid and Clements 1999; Salgado et al.2005) suggested that selective activation of D2/3 receptors didnot change the quantal amplitude. Instead, the parameter thatwas different in the fitted function (N) was consistent with apresynaptic change (Eq. 3; see METHODS).
Postsynaptic control of subthalamic inputs by D4 receptors
The hypothesis that the sulpiride-resistant action was medi-ated by D4 receptors located postsynaptically on GP neuronswas supported by three types of experiments. First, selectiveactivation of D4 receptors reduced subthalamopallidal EPSCswithout changing the paired-pulse ratio. Second, D4 receptoractivation reduced the mean amplitude of sEPSCs withoutaffecting the frequency. Finally, variance–mean analysis re-vealed a significant reduction in quantal amplitude withoutaffecting any presynaptic parameter.
2885DOPAMINE CONTROLS THE SUBTHALAMIC INPUT TO THE PALLIDUM
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from

To conclude, both D2/3 and D4 type receptors modulate thesubthalamopallidal response. Nevertheless, D2/3-type receptorsexert their modulation acting presynaptically, whereas D4-typereceptors modulate the inputs from the postsynaptic side.
Possible physiological consequences
Dopamine actions in the GP are necessary to achieve motorcontrol (Pan and Walters 1988; Raz et al. 2000). Bilateralinfusions of D1- or D2-class receptor antagonists into the GPinduce akinesia and catalepsia (Costall and Naylor 1972;Hauber and Lutz 1999). However, the infusion of dopamine inthe same nucleus causes amelioration of motor deficits inhemiparkinsonian rats (Galvan et al. 2001). It is then importantto know the actions of dopamine in the GP.
The GP and the STN assemble a reciprocally innervatednetwork of excitatory and inhibitory neurons (Bevan et al.2002b; Gandia et al. 1993; Hazrati and Parent 1992; Kita andKita 1991, 2001; Kita et al. 2004; Nambu et al. 2000; Parent etal. 2000; Sato et al. 2000a,b; Shink et al. 1996; Shinonaga et al.1992; Smith et al. 1990). Because of the intrinsic properties ofGP and STN neurons (Beurrier et al. 1999, 2000; Bevan andWilson 1999; Bevan et al. 2002a; Chan et al. 2004; Cooper andStanford 2000; Hallworth et al. 2003; Kita and Kitai 1991; Kitaet al. 2004; Nambu and Llinas 1994; Overton and Greenfield1995), this network forms a robust central pattern generator(CPG) in the basal ganglia (Grillner 2003; Terman et al. 2002;Yuste et al. 2005). Diverse spatiotemporal patterns of rhythmicand synchronous oscillatory activity can arise when the levelsof inhibitory or excitatory inputs impinging onto this networkare varied (Bevan and Wilson 1999; Bevan et al. 2002b; Kitaet al. 2004; Overton and Greenfield 1995; Plenz and Kitai1999; Terman et al. 2002; Urbain et al. 2000). The present andprevious (Cooper and Stanford 2001; Floran et al. 1997)findings showed that the levels of inhibitory and excitatoryinputs impinging onto GP neurons depend on dopaminergicmodulation. Therefore it is logical to infer that the variety ofspatiotemporal patterns of oscillatory activity that this networkexhibits to produce motor organization may be a product ofdopaminergic modulation. Perhaps, a detailed somatotopicorganization of dopaminergic neurons in the compacta deter-mines, according to the afferents active during a motor task,which inputs increase and which decrease their strength. Ifdopamine modulation is lost, however, a dramatic reduction inthe possible patterns is obtained. The system then becomeslocked in a few extreme and robust set of variations (Bergmanet al. 1994; Brown et al. 2001; Levy et al. 2002; Magill et al.2001; Magnin et al. 2000; Nini et al. 1995; Raz et al. 2000).
A C K N O W L E D G M E N T S
The technical assistance of H. Zavala and A. Laville is greatly appreciated.
G R A N T S
This work was supported by Consejo Nacional de Ciencia y Tecnologıa(Mexico) Grants G34706 to J. Aceves and 42636 to E. Galarraga, by grantsfrom Direccion General de Asuntos del Personal Academico, UniversidadNacional Autonoma de Mexico Grants IN201603 to J. Bargas and IN200803to E. Galarraga, and by Program Project IMPULSA 03 grants to E. Galarragaand J. Bargas.
R E F E R E N C E S
Ariano MA, Wang J, Noblett KL, Larson ER, and Sibley DR. Cellulardistribution of the rat D4 dopamine receptor protein in the CNS usinganti-receptor antisera. Brain Res 752: 26–34, 1997.
Baldelli P, Hernandez-Guijo JM, Carabelli V, and Carbone E. Brain-derived neurotrophic factor enhances GABA release probability and non-uniform distribution of N- and P/Q-type channels on release sites ofhippocampal inhibitory synapses. J Neurosci 25: 3358–3368, 2005.
Baufreton J, Garret M, Rivera A, de la Calle A, Gonon F, Dufy B, BioulacB, and Taupignon A. D5 (not D1) dopamine receptors potentiate burst-firing in neurons of the subthalamic nucleus by modulating an L-typecalcium conductance. J Neurosci 23: 816–825, 2003.
Beckstead RM. A pallidostriatal projection in the cat and monkey. Brain ResBull 11: 629–632, 1983.
Bergman H, Wichmann T, Karmon B, and DeLong MR. The primatesubthalamic nucleus. II. Neuronal activity in the MPTP model of parkin-sonism. J Neurophysiol 72: 5507–5520, 1994.
Beurrier C, Ben-Ari Y, and Hammond C. Preservation of the direct andindirect pathways in an in vitro preparation of the mouse basal ganglia.Neuroscience 140: 77–86, 2006.
Beurrier C, Bioulac B, and Hammond C. Slowly inactivating sodium current(INaP) underlies single-spike activity in rat subthalamic neurons. J Neuro-physiol 83: 1951–1957, 2000.
Beurrier C, Congar P, Bioulac B, and Hammond C. Subthalamic nucleusneurons switch from single-spike activity to burst-firing mode. J Neurosci19: 599–609, 1999.
Bevan MD, Magill PJ, Hallworth NE, Bolam JP, and Wilson CJ. Regula-tion of the timing and pattern of action potential generation in rat subtha-lamic neurons in vitro by GABA-A IPSPs. J Neurophysiol 87: 1348–1362,2002a.
Bevan MD, Magill PJ, Terman D, Bolam JP, and Wilson CJ. Move to therhythm: oscillations in the subthalamic nucleus-external globus pallidusnetwork. Trends Neurosci 25: 525–531, 2002b.
Bevan MD and Wilson CJ.Mechanisms underlying spontaneous oscillationand rhythmic firing in rat subthalamic neurons. J Neurosci 19: 7617–7628,1999.
Bolshakov VY and Siegelbaum SA. Regulation of hippocampal transmitterrelease during development and long-term potentiation. Science 269: 1730–1734, 1995.
Brown P, Oliviero A, Mazzone P, Insola A, Tonali P, and Di Lazzaro V.Dopamine dependency of oscillations between subthalamic nucleus andpallidum in Parkinson’s disease. J Neurosci 21: 1033–1038, 2001.
Chan CS, Shigemoto R, Mercer JN, and Surmeier DJ. HCN2 and HCN1channels govern the regularity of autonomous pacemaking and synapticresetting in globus pallidus neurons. J Neurosci 24: 9921–9932, 2004.
Ciliax BJ, Nash N, Heilman C, Sunahara R, Hartney A, Tiberi M, Rye DB,Caron MG, Niznik HB, and Levey AL. Dopamine D5 receptor immuno-localization in rat and monkey brain. Synapse 37: 125–145, 2000.
Clements JD and Silver RA. Unveiling synaptic plasticity: a new graphicaland analytical approach. Trends Neurosci 23: 105–113, 2000.
Cooper AJ and Stanford IM. Electrophysiological and morphological char-acteristics of three subtypes of rat globus pallidus neurone in vitro. J Physiol527: 291–304, 2000.
Cooper AJ and Stanford IM. Dopamine D2 receptor mediated presynapticinhibition of striatopallidal GABAA IPSCs in vitro. Neuropharmacology 41:62–71, 2001.
Costall B, Naylor RJ, and Olley JF. On the involvement of the caudate-putamen, globus pallidus and substantia nigra with neuroleptic and cholin-ergic modification of locomotor activity. Neuropharmacology 11: 317–330,1972.
Dawson TM, Gehlert DR, McCabe RT, Barnett A, and Wamsley JK. D1
dopamine receptors in the rat brain: a quantitative autoradiographic analysis.J Neurosci 6: 2352–2365, 1986.
Debeir T, Ginestet L, Francois C, Laurens S, Martel JC, Chopin P,Marien M, Colpaert F, and Raiman-Vozari R. Effect of intrastriatal6-OHDA lesion on dopaminergic innervation of the rat cortex and globuspallidus. Exp Neurol 193: 444–454, 2005.
DeLong MR, Crutcher MD, and Georgopoulos AP. Primate globus pallidusand subthalamic nucleus: functional organization. J Neurophysiol 53: 530–543, 1995.
Dunwiddie TV and Haas HL. Adenosine increases synaptic facilitation in thein vitro rat hippocampus: evidence for a presynaptic site of action. J Physiol369: 365–377, 1985.
2886 HERNANDEZ ET AL.
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from

Engler B, Freiman I, Urbanski M, and Szabo B. Effects of exogenous andendogenous cannabinoids on GABAergic neurotransmission between thecaudate-putamen and the globus pallidus in the mouse. J Pharmacol ExpTher 316: 608–617, 2006.
Floran B, Floran L, Erlij D, and Aceves J. Dopamine D4 receptors inhibitdepolarization-induced [3H]GABA release in the rat subthalamic nucleus.Eur J Pharmacol 498: 97–102, 2004.
Floran B, Floran L, Sierra A, and Aceves J. D2 receptor mediated inhibitionof GABA release by endogenous dopamine in the rat globus pallidus.Neurosci Lett 237: 1–4, 1997.
Flores G, Liang JJ, Sierra A, Martinez-Fong D, Quirion R, Aceves J, andSrivastava LK. Expression of dopamine receptors in the subthalamicnucleus of the rat: characterization using reverse transcriptase-polymerasechain reaction and autoradiography. Neuroscience 91: 549–556, 1999.
Fonnum F, Gottesfeld Z, and Grofova I. Distribution of glutamate decar-boxylase, choline acetyl-transferase and aromatic amino acid decarboxylasein the basal ganglia of normal and operated rats. Evidence for striatopallidal,striatoentopeduncular and striatonigral GABAergic fibres. Brain Res 143:125–138, 1978.
Galvan A, Floran B, Erlij D, and Aceves J. Intrapallidal dopamine restoresmotor deficits induced by 6-hydroxydopamine in the rat. J Neural Transm108: 153–166, 2001.
Gandia JA, De Las Heras S, Garcia M, and Gimenez-Amaya JM. Afferentprojections to the reticular thalamic nucleus from the globus pallidus and thesubstantia nigra in the rat. Brain Res Bull 32: 351–358, 1993.
Gauthier J, Parent M, Levesque M, and Parent A. The axonal arborizationof single nigrostriatal neurons in rats. Brain Res 834: 228–232, 1999.
Gimenez-Amaya JM, and Graybiel AM. Compartmental origins of thestriatopallidal projection in the primate. Neuroscience 34: 111–126, 1990.
Glase SA, Akunne HC, Georgic LM, Heffner TG, MacKenzie RG, ManleyPJ, Pugsley TA, and Wise LD. Substituted [(4-phenylpiperazinyl)-meth-yl]benzamide: selective dopamine D4 agonists. J Med Chem 40: 1771–1772,1997.
Grillner S. The motor infrastructure: from ion channels to neuronal networks.Nat Rev Neurosci 4: 573–586, 2003.
Guzman JN, Hernandez A, Galarraga E, Laville A, Vergara R, Aceves J,and Bargas J. Dopaminergic modulation of axon collaterals interconnectingspiny neurons of the rat sriatum. J Neurosci 23: 8931–8940, 2003.
Hallworth NE, Wilson CJ, and Bevan MD. Apamin-sensitive small conduc-tance calcium-activated potassium channels, through their selective cou-pling to voltage-gated calcium channels, are critical determinants of theprecision, pace, and pattern of action potential generation in rat subthalamicnucleus neurons in vitro. J Neurosci 23: 7525–7542, 2003.
Hanson JE and Jaeger D. Short-term plasticity shapes the response tosimulated normal and parkinsonian input patterns in the globus pallidus.J Neurosci 22: 5164–5172, 2002.
Hauber W and Lutz S. Dopamine D1 or D2 receptor blockade in the globuspallidus produces akinesia in the rat. Behav Brain Res 106: 143–150, 1999.
Hazrati LN and Parent A. Convergence of subthalamic and striatal efferentsat pallidal level in primates: an anterograde double-labeling study withbiocytin and PHA-L. Brain Res 569: 336–340, 1992.
Hernandez A, Valdiosera R, Floran B, Erlij D, and Aceves J. Pre- andpostsynaptic modulation of the glutamatergic transmission by dopamine inthe rat globus pallidus. Soc Neurosci Abstr 46: 16, 2004.
Hoover BR and Marshall JF. Population characteristics of preproenkephalinmRNA-containing neurons in the globus pallidus of the rat. Neurosci Lett265: 199–202, 1999.
Hurd YL, Suzuki M, and Sedvall GC. D1 and D2 dopamine receptor mRNAexpression in whole hemisphere sections of the human brain. J ChemNeuroanat 22: 127–137, 2001.
Ibanez-Sandoval O, Hernandez A, Floran B, Galarraga E, Tapia D,Valdiosera R, Erlij D, Aceves J, and Bargas J. Control of the subthalamicinnervation of substantia nigra pars reticulata by D1 and D2 dopaminereceptors. J Neurophysiol 95: 1800–1811, 2006.
Iribe Y, Moore K, Pang KC, and Tepper JM. Subthalamic stimulation-induced synaptic responses in substantia nigra pars compacta dopaminergicneurons in vitro. J Neurophysiol 82: 925–933, 1999.
Johnson AE, Coirini H, Kallstrom L, and Wiesel FA. Characterization ofdopamine receptor binding sites in the subthalamic nucleus. Neuroreport 5:1836–1838, 1994.
Jonas P, Major G, and Sakmann B. Quantal components of unitary EPSCsat the mossy fibre synapse on CA3 pyramidal cells of rat hippocampus.J Physiol 472: 615–663, 1993.
Kamiya H and Zucker RS. Residual Ca2� and short-term synaptic plasticity.Nature 371: 603–606, 1994.
Kanazawa I, Marshall GR, and Kelly JS. Afferents to the rat substantia nigrastudied with horseradish peroxidase, with special reference to fibers from thesubthalamic nucleus. Brain Res 115: 485–491, 1976.
Kaneda K, Nambu A, Tokuno H, and Takada M. Differential processingpatterns of motor information via striatopallidal and striatonigral projec-tions. J Neurophysiol 88: 1420–1432, 2002.
Kawaguchi Y, Wilson CJ, and Emson PC. Projection subtypes of ratneostriatal matrix cells revealed by intracellular injection of biocytin. J Neu-rosci 10: 3421–3438, 1990.
Kita H and Kita T. Number, origins, and chemical types of rat pallidostriatalprojection neurons. J Comp Neurol 437: 438–448, 2001.
Kita H and Kitai ST. Intracellular study of rat globus pallidus neurons:membrane properties and responses to neostriatal, subthalamic and nigralstimulation. Brain Res 564: 296–305, 1991.
Kita H and Kitai ST. The morphology of globus pallidus projection neuronsin the rat: an intracellular staining study. Brain Res 636: 308–319, 1994.
Kita H, Nambu A, Kaneda K, Tachibana Y, and Takada M. Role ofionotropic glutamatergic and GABAergic inputs on the firing activity ofneurons in the external pallidum in awake monkeys. J Neurophysiol 92:3069–3084, 2004.
Kita H, Tachibana Y, Nambu A, and Chiken S. Balance of monosynapticexcitatory and disynaptic inhibitory responses of the globus pallidus inducedafter stimulation of the subthalamic nucleus in the monkey. J Neurosci 25:8611–8619, 2005.
Koga E and Momiyama T. Presynaptic dopamine D2-like receptors inhibitexcitatory transmission onto rat ventral tegmental dopaminergic neurons.J Physiol 523: 163–173, 2000.
Koos T, Tepper JM, and Wilson CJ. Comparison of IPSCs evoked by spinyand fast-spiking neurons in the neostriatum. J Neurosci 24: 7916–7922,2004.
Kreiss DS, Anderson LA, and Walters JR. Apomorphine and dopamine D1
receptor agonists increase the firing rates of subthalamic nucleus neurons.Neuroscience 72: 863–876, 1996.
Kreiss DS, Mastropietro CW, Rawji SS, and Walters JR. The response ofsubthalamic nucleus neurons to dopamine receptor stimulation in a rodentmodel of Parkinson’s disease. J Neurosci 17: 6807–6819, 1997.
Kulagowski JJ, Broughton HB, Curtis NR, Mawer IM, Ridgill MP, BakerR, Emms F, Freedman SB, Marwood R, Patel S, Ragan CI, and LeesonPD. 3-((4-(4-Chlorophenyl)piperazin-1-yl)-methyl)-1H-pyrrolo-2,3-b-pyri-dine: an antagonist with high affinity and selectivity for the human dopa-mine D4 receptor. J Med Chem 39: 1941–1942, 1996.
Levy R, Ashby P, Hutchison WD, Lang AE, Lozano AM, and DostrovskyJO. Dependence of subthalamic nucleus oscillations on movement anddopamine in Parkinson’s disease. Brain 125: 1196–1209, 2002.
Lindvall O and Bjorklund A. Dopaminergic innervation of the globuspallidus by collaterals from the nigrostriatal pathway. Brain Res 172:169–173, 1979.
Magill PJ, Bolam JP, and Bevan MD. Dopamine regulates the impact of thecerebral cortex on the subthalamic nucleus-globus pallidus network. Neu-roscience 106: 313–330, 2001.
Magnin M, Morel A, and Jeanmonod D. Single-unit analysis of the palli-dum, thalamus and subthalamic nucleus in parkinsonian patients. Neuro-science 96: 549–564, 2000.
Mann-Metzer P and Yarom Y. Pre- and postsynaptic inhibition mediated byGABAB receptors in cerebellar inhibitory interneurons. J Neurophysiol 87:183–190, 2002.
Mauger C, Silvan B, Brockhaus M, Fuchs S, Civelli O, and Monsma F Jr.Development and characterization of antibodies directed against the mouseD4 dopamine receptor. Eur J Neurosci 10: 529–537, 1998.
Meador-Woodruff JH, Mansour A, Grandy DK, Damask SP, Civelli O,and Watson SJ Jr. Distribution of D5 dopamine receptor mRNA in ratbrain. Neurosci Lett 145: 209–212, 1992.
Mintz I, Hammond C, and Feger J. Excitatory effect of iontophoreticallyapplied dopamine on identified neurons of the rat subthalamic nucleus.Brain Res 375: 172–175, 1986.
Mitchell SJ, Richardson RT, Baker FH, and DeLong MR. The primateglobus pallidus: neuronal activity related to direction of movement. ExpBrain Res 68: 491–505, 1987.
Mouroux M, Hassani OK, and Feger J. Electrophysiological and Fosimmunohistochemical evidence for the excitatory nature of the parafascicu-lar projection to the globus pallidus. Neuroscience 81: 387–397, 1997.
2887DOPAMINE CONTROLS THE SUBTHALAMIC INPUT TO THE PALLIDUM
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from

Mrzljak L, Bergson C, Pappy M, Huff R, and n-Rakic PS. Localization ofdopamine D4 receptors in GABAergic neurons of the primate brain. Nature381: 245–248, 1996.
Nakanishi H, Hori N, and Kastuda N. Neostriatal evoked inhibition andeffects of dopamine on globus pallidal neurons in rat slice preparations.Brain Res 358: 282–286, 1985.
Nambu A and Llinas R. Electrophysiology of globus pallidus neurons invitro. J Neurophysiol 72: 1127–1139, 1994.
Nambu A and Llinas R. Morphology of globus pallidus neurons: its corre-lation with electrophysiology in guinea pig brain slices. J Comp Neurol 377:85–94, 1997.
Nambu A, Tokuno H, Hamada I, Kita H, Imanishi M, Akazawa T, IkeuchiY, and Hasegawa N. Excitatory cortical inputs to pallidal neurons via thesubthalamic nucleus in the monkey. J Neurophysiol 84: 289–300, 2000.
Ni Z, Gao D, Bouali-Benazzouz R, Benabid AL, and Benazzouz A. Effectof microiontophoretic application of dopamine on subthalamic nucleusneuronal activity in normal rats and in rats with unilateral lesion of thenigrostriatal pathway. Eur J Neurosci 14: 373–381, 2001.
Nini A, Feingold A, Slovin H, and Bergman H. Neurons in the globuspallidus do not show correlated activity in the normal monkey, but phase-locked oscillations appear in the MPTP model of parkinsonism. J Neuro-physiol 74: 1800–1805, 1995.
Ogura M and Kita H. Dynorphin exerts both postsynaptic and presynapticeffects in the globus pallidus of the rat. J Neurophysiol 83: 3366–3376,2000.
Overton PG and Greenfield SA. Determinants of neuronal firing pattern inthe guinea-pig subthalamic nucleus: an in vivo and in vitro comparison.J Neural Transm 10: 41–54, 1995.
Pan HS and Walters JR. Unilateral lesion of the nigrostriatal pathwaydecreases the firing rate and alters the firing pattern of globus pallidusneurons in the rat. Synapse 2: 650–656, 1988.
Parent A, Bouchard C, and Smith Y. The striatopallidal and striatonigralprojections: two distinct fiber systems in primate. Brain Res 303: 385–390,1984.
Parent A, Sato F, Wu Y, Gauthier J, Levesque M, and Parent M.Organization of the basal ganglia: the importance of axonal collateralization.Trends Neurosci 23: S20–S27, 2000.
Patel S, Freedman S, Chapman KL, Emms F, Fletcher AE, Knowles M,Marwood R, Mcallister G, Myers J, Curtis N, Kulagowski JJ, LeesonPD, Ridgill M, Graham M, Matheson S, Rathbone D, Watt AP, BristowLJ, Rupniak NM, Baskin E, Lynch JJ, and Ragan CI. Biological profileof L-745,870, a selective antagonist with high affinity for the dopamine D4
receptor. J Pharmacol Exp Ther 283: 636–647, 1997.Paulsen O and Heggelund P. The quantal size at retinogeniculate synapses
determined from spontaneous and evoked EPSCs in guinea-pig thalamicslices. J Physiol 480: 505–511, 1994.
Plenz D and Kitai ST. A basal ganglia pacemaker formed by the subthalamicnucleus and external globus pallidus. Nature 400: 677–682, 1999.
Prensa L and Parent A. The nigrostriatal pathway in the rat: a single-axonstudy of the relationship between dorsal and ventral tier nigral neurons andthe striosome/matrix striatal compartments. J Neurosci 21: 7247–7260,2001.
Price CJ and Pittman QJ. Dopamine D4 receptor activation inhibits presyn-aptically glutamatergic neurotransmission in the rat supraoptic nucleus.J Neurophysiol 86: 1149–1155, 2001.
Querejeta E, Delgado A, Valdiosera R, Erlij D, and Aceves J. IntrapallidalD2 dopamine receptors control globus pallidus neuron activity in the rat.Neurosci Lett 300: 79–82, 2001.
Rajakumar N, Rushlow W, Naus CC, Elisevich K, and Flumerfelt BA.Neurochemical compartmentalization of the globus pallidus in the rat: animmunocytochemical study of calcium-binding proteins. J Comp Neurol346: 337–348, 1994.
Raz A, Vaadia E, and Bergman H. Firing patterns and correlations ofspontaneous discharge of pallidal neurons in the normal and the tremulous1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine vervet model of parkinson-ism. J Neurosci 20: 8559–8571, 2000.
Reid CA and Clements JD. Postsynaptic expression of long-term potentiationin the rat dentate gyrus demonstrated by variance-mean analysis. J Physiol518: 121–130, 1999.
Ruskin DN and Marshall JF. Differing influences of dopamine agonists andantagonists on Fos expression in identified populations of globus pallidusneurons. Neuroscience 81: 79–92, 1997.
Sahara Y and Takahashi T. Quantal components of the excitatory postsyn-aptic currents at a rat central auditory synapse. J Physiol 536: 189–197,2001.
Salgado H, Tecuapetla F, Perez-Rosello T, Perez-Burgos A, Perez-GarciE, Galarraga E, and Bargas J. A reconfiguration of CaV2 Ca2� channelscurrent and its dopaminergic D2 modulation in developing neostriatalneurons. J Neurophysiol 94: 3771–3787, 2005.
Sato F, Lavallee P, Levesque M, and Parent A. Single-axon tracing study ofneurons of the external segment of the globus pallidus in primate. J CompNeurol 417: 17–31, 2000a.
Sato F, Parent M, Levesque M, and Parent A. Axonal branching pattern ofneurons of the subthalamic nucleus in primates. J Comp Neurol 424:142–152, 2000b.
Shin RM, Masuda M, Miura M, Sano H, Shirasawa T, Song WJ, Koba-yashi K, and Aosaki T. Dopamine D4 receptor-induced postsynapticinhibition of GABAergic currents in mouse globus pallidus neurones.J Neurosci 23: 11662–11672, 2003.
Shink E, Bevan MD, Bolam JP, and Smith Y. The subthalamic nucleus andthe external pallidum: two tightly interconnected structures that control theoutput of the basal ganglia in the monkey. Neuroscience 73: 335–357, 1996.
Shinonaga Y, Takada M, Ogawa-Meguro R, Ikai Y, and Mizuno N. Directprojections from the globus pallidus to the midbrain and pons in the cat.Neurosci Lett 135: 179–183, 1992.
Smith ID and Grace AA. Role of the subthalamic nucleus in the regulation ofnigral dopamine neuron activity. Synapse 12: 287–303, 1992.
Smith Y, Hazrati LN, and Parent A. Efferent projections of the subthalamicnucleus in the squirrel monkey as studied by the PHA-L anterograde tracingmethod. J Comp Neurol 294: 306–323, 1990.
Svenningsson P and Le Moine C. Dopamine D1/5 receptor stimulationinduces c-fos expression in the subthalamic nucleus: possible involvementof local D5 receptors. Eur J Neurosci 15: 133–142, 2002.
Terman D, Rubin JE, Yew AC, and Wilson CJ. Activity patterns in a modelfor the subthalamopallidal network of the basal ganglia. J Neurosci 22:2963–2976, 2002.
Tremblay L and Filion M. Responses of pallidal neurons to striatal stimula-tion in intact waking monkeys. Brain Res 498: 1–16, 1989.
Turner RS, Grafton ST, Votaw JR, Delong MR, and Hoffman JM. Motorsubcircuits mediating the control of movement velocity: a PET study.J Neurophysiol 80: 2162–2176, 1998.
Urbain N, Gervasoni D, Souliere F, Lobo L, Rentero N, Windels F, AstierB, Savasta M, Fort P, Renaud B, Luppi PH, and Chouvet G. Unrelatedcourse of subthalamic nucleus and globus pallidus neuronal activities acrossvigilance states in the rat. Eur J Neurosci 12: 3361–3374, 2000.
Vergara R, Rick C, Hernandez-Lopez S, Laville JA, Guzman JN, Galar-raga E, Surmeier DJ, and Bargas J. Spontaneous voltage oscillations instriatal projection neurons in a rat corticostriatal slice. J Physiol 553:169–182, 2003.
Voorn P, van de Witte S, Tjon G, and Jonker AJ. Expression of enkephalinin pallido-striatal neurons. Ann NY Acad Sci 877: 671–675, 1999.
Wang X, Zhong P, and Yan Z. Dopamine D4 receptors modulate GABAergicsignaling in pyramidal neurons of prefrontal cortex. J Neurosci 22: 9185–9193, 2002.
Windels F, Carcenac C, Poupard A, and Savasta M. Pallidal origin ofGABA release within the substantia nigra pars reticulata during high-frequency stimulation of the subthalamic nucleus. J Neurosci 25: 5079–5086, 2005.
Yasukawa T, Kita T, Xue Y, and Kita H. Rat intralaminar thalamic nucleiprojections to the globus pallidus: a biotinylated dextran amine anterogradetracing study. J Comp Neurol 471: 153–167, 2004.
Yuste R, MacLean JN, Smith J, and Lansner A. The cortex as a centralpattern generator. Nat Rev Neurosci 6: 477–483, 2005.
Zhu Z, Bartol M, Shen K, and Johnson SW. Excitatory effects of dopamineon subthalamic nucleus neurons: in vitro study of rats pretreated with6-hydroxydopamine and levodopa. Brain Res 945: 31–40, 2002.
Zucker RS. Calcium- and activity-dependent synaptic plasticity. Curr OpinNeurobiol 9: 305–313, 1999.
2888 HERNANDEZ ET AL.
J Neurophysiol • VOL 96 • DECEMBER 2006 • www.jn.org
on Novem
ber 22, 2006 jn.physiology.org
Dow
nloaded from















![D4 dVed fa AVXRdfd ac`SV aR_V] - Daily Pioneer](https://img.dokumen.tips/doc/110x75/631e48c225add517740af8e1/d4-dved-fa-avxrdfd-acsv-arv-daily-pioneer.jpg)












