Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Vaccine 26 (2008) 2935–2941
Contents lists available at ScienceDirect
Vaccine
journa l homepage: www.e lsev ier .com/ locate /vacc ine
Construction and evaluation of the eukaryotic expression plasmid encoding twocopies of somatostatin genes fused with hepatitis B surface antigen gene S
Aixin Lianga, Shaoxian Caob, Li Hana, Yanfeng Yaoa, M. Moaeen-ud-Dina, Liguo Yanga,∗
a Key Lab of Agricultural Animal Genetics, Breeding and Reproduction of Ministry of Education, Huazhong Agricultural University, Wuhan 430070, PR Chinab Institute of Animal Science, Jiangsu Academy of Agricultural Science, Nanjing 210014, PR China
a r t i c l e i n f o
Article history:Received 14 August 2007Received in revised form 18 March 2008Accepted 19 March 2008Available online 8 April 2008
Keywords:SomatostatinHepatitis B surface antigenDNA vaccine
a b s t r a c t
The aim of current study was to evaluate the prospects of somatostatin DNA vaccine. Two copies ofsomatostatin (SS) genes were fused with the hepatitis B surface antigen (HBsAg) S gene using geneticengineering methods, the identified recombinant plasmid designated as pcS/2SS was transfected intoHeLa cells to detect expression and antigenicity of target fusion protein, and its immunoreaction as wellas safety was evaluated with animal experiments. The expressed target protein had a specific reactionwith somatostatin antibody and showed a single strip result. A single injection of this vector stimulatedlong-term antigen-specific antibody responses in rats, and peak antibody levels occurred at the 2nd weekof the initial injection. Additionally, the 50 �g immunized group resulted in a 13.5% increase in growth rateas compared with control group (111.7 g vs. 98.4 g). The genomic DNA was assayed for integrated plasmidusing a sensitive PCR method, and the risk of mutation due to integration of pcS/2SS plasmid followingintramuscular injection in mice was negligible. The successful construction of pcS/2SS DNA vaccine withgood immunogenicity and safety has prospects to promote growth of animals.
© 2008 Elsevier Ltd. All rights reserved.
Introduction
Growth is a very complex process which is governed bygenotype, environment, nutrition and the endocrine system. Theprincipal hormone involved in stimulating growth is pituitarygrowth hormone (GH) and the secretion of GH is regulated througha complex neuroendocrine control system, especially by the func-tional interplay of two hypothalamic hypophysiotropic hormones,growth hormone-releasing hormone (GHRH) and somatostatin(SS), exerting stimulatory and inhibitory influences, respectively[1–4]. Somatostatin, which is not species-specific and which occursin a 14- and 28-amino acid form, inhibits the secretion of mosthormones [5]. Immuno-neutralization of somatostatin has shownsome promise as a technique for the enhancement of growth. Therehas been a lot of literature that immunization against synthetic SSconjugated to a carrier protein could promote the growth rates ofimmunized animals, such as lambs, steers and pigs [6–9]. Recom-binant vaccinia virus expressing SS has also been described topromote the growth performance of immunized animals [10–13].Unfortunately, the purification steps of these vaccines were com-plicated and tedious, thereby increasing cost and labor extensively.Additionally, the vaccinia virus could not be used for booster immu-
∗ Corresponding author. Tel.: +86 27 8728 1813; fax: +86 27 8728 1813.E-mail address: [email protected] (L. Yang).
nization and had the risk of virulence recovery. Therefore, it is nec-essary to develop a novel strategy of somatostatin immunization.
DNA vaccines have become a reliable and major means to elicitimmune response in the past decade, and immune responses areinduced subsequent to the in vivo expression of antigen fromdirectly introduced plasmid DNA [14–17]. The potential advantagesof DNA vaccines include the induction of cellular and humoralimmune responses, flexible genetic design, lack of infection risk,stability of reagents, and the relatively low cost of production ina microbial host. So far, it has been used successfully in raisingimmune responses against various infectious agents, i.e. HIV-1,influenza, parasites, Mycoplasma pulmonis, Mycobacterium tuber-culosis and tumor cells [18,19].
It was concluded in light of above-mentioned studies thatsomatostatin DNA vaccine potency should be tested through choos-ing the hepatitis B surface antigen (HBsAg) particle as a vehiclefor presentation of SS antigenic determinants. The surface anti-gen of HBV (HBsAg) is highly immunogenic both as recombinantparticles as well as DNA vaccines. Previous studies have shownHBsAg is an effective tool for the presentation of foreign epitopes,i.e. HIV-V3 [13,20,21]. This prompted us to investigate whetherthe potent immunogenicity of the HBsAg particles could be usedto enhance the immune response against somatostatin vaccine.Thus, we constructed the fusion plasmid encoding somatostatingene fused with hepatitis B surface antigen gene S, and studiedthe antibody response as well as the growth rate of rats. Moreover,
0264-410X/$ – see front matter © 2008 Elsevier Ltd. All rights reserved.doi:10.1016/j.vaccine.2008.03.036

Author's personal copy
2936 A. Liang et al. / Vaccine 26 (2008) 2935–2941
the potential of recombinant plasmid to integrate into the genomicDNA was also investigated.
Materials and methods
Vectors
SS gene was synthesized chemically by Hao Jia TechnologyCompany (Shanghai, China), pcMV-S vector encoding hepatitis Bvirus surface protein S was conserved by our laboratory, prokary-otic expression vector of pUC18 and eukaryotic expression vectorpcDNA3.1(−) containing the cytomegalovirus (CMV) promoterwere purchased from Invitrogen Company (Shanghai, China).
Recombinant DNA techniques
DNA preparation, genetic manipulations, PCR, and transforma-tion of bacteria were carried out according to standard protocols[22].
Construction of fusion expression plasmid harboring SS and Sgenes
S gene was amplified by PCR from pcMV-S using a pair of primers(Table 1), which contained a Kozak sequence [23,24] incorporatingthe start codon (bold) and EcoRI site (underline) as well as KpnI site(underline). The PCR products were analyzed by agarose gel elec-trophoresis. The fragment of the expected (696 bp) size was thendigested using EcoRI and KpnI restriction enzymes and inserted intopUC18 plasmid. Hereafter, the synthetic SS gene was inserted intothe 3′ end of S gene to form the fusion gene S/SS [12]. The 5′ endof SS included KpnI site, the 3′ end was stop codon and includedHindIII site, the sequence was shown as follows:
After that, the fusion gene was subcloned into pcDNA3.1(−) cre-ating pcS/SS plasmid. In order to construct two copies of SS plasmidnamed pcS/2SS, PSSB primers (Table 1) were designed that changedthe restriction enzyme site and deleted the stop codon, which con-tained two BamHI sites (underline), and then amplified SS genefrom pcS/SS. The PCR products of the above reaction were digestedby BamHI and were incorporated between the 112 and 113 aminoacids [13] in the coding region of S gene to construct pcS/2SS. Theinsertion site, direction and sequence of the genes were identifiedto be correct by restriction endonuclease digestion and sequencing.
Expression of recombinant plasmid pcS/2SS in HeLa cells
HeLa cells were cultured in Dulbecco’s Modified Eagles Medium(DMEM, Invitrogen, Shanghai, China) with 10% fetal bovine serum(FBS, Invitrogen, Shanghai, China). Transfection experiments wereperformed on 70–80% confluent cells using LipofectamineTM 2000Kit (Invitrogen, Shanghai, China) according to manufacturer’sinstructions. After incubation for 72 h at 37 ◦C in 5% CO2 incuba-tor, the growth media were replaced and G418 (400 �g/ml) wasadded, then incubation continued for 2 weeks until ready to assay.The samples were boiled for 10 min and then separated by discon-tinuous sodium dodecylsulfate polyacrylamide gel electrophoresis(SDS-PAGE). Protein bands were visualized by Coomassie Bril-liant Blue staining. Following SDS-PAGE separation, proteins weretransferred to a nitrocellulose membrane. After blocking with 5%(w/v) dry skimmed milk in TBS for 1 h at 37 ◦C, membrane stripswere incubated with a 1:200 dilution of rabbit anti-SS (Boster,Wuhan, China) for 3 h at 37 ◦C. Following extensive washing,membrane strips were incubated with a 1:200 dilution of HRP(horseradish peroxidase)-conjugated goat anti-rabbit IgG (Boster,Wuhan, China) for 1 h at 37 ◦C, then extensive washing again. Detec-tion was performed by diaminobenzidine (DAB, Sigma–Aldrich).The immune enzymic reaction was stopped by rinsing in distilledwater.
DNA vaccination
RatsForty female SD rats of 6 weeks of age were purchased from ani-
mal field of Jiangning QingLong Hill, and divided into four groupsrandomly. One-week acclimatization period was given beforeimmunization. An injection with 100 �l of procaine hydrochlo-ride (0.5%) was performed at 24 h before the immunization
as a pretreatment, then rats were immunized intramuscularly (i.m)in tibialis anterior with different doses (10 �g/100 �l, 50 �g/100 �l,100 �g/100 �l) of pcS/2SS plasmid as tested group and the samevolume of physiological saline as a negative control. A booster wasperformed once with the same dose of plasmid as the primaryimmunization 4 weeks later. The blood samples were harvestedby retroorbital puncture with heparinized tubes at weeks 0, 2, 4, 6,9 and 12. Following centrifugation at 4000 × g for 5 min, the plasmawas separated from the whole blood and stored at −20 ◦C for use.
Table 1The primer sets for PCR in this study
Primer names Sequences of primer Anneal temperature (◦C) Fragment length (bp)
Ps F: TAATATTGAATTCCCTGCGCTGAACATGG 60 696R: GCGAAGGTACCAACGTATACCCAAAGACAA
PSSB F: TAGGGATCCGCTGGCTGCA AGAAT 52.1 42R: CCGGGATCCACAGGATGTGAAAGTC
PGAPDH F: AGCCTCGTCCCGTAGACAA AATGGT 60 1002R: GTGGGTGGTCCAGGGTTTCTTACTC
PS/2SS F: GAATTCCCTGCGCTGAACATGGA 65 792R: GAGTTTTTGTTCTACGTAA
Notes: The restriction endonucleases of EcoRI and KpnI were designed in the Ps primer marked with underline, and the Kozak sequence was shown in bold. In the primerPSSB, the BamHI sites were also underlined. F: forward; R: reverse.
Author's personal copy
A. Liang et al. / Vaccine 26 (2008) 2935–2941 2937
MiceFifteen Balb/C female mice (SPF grade, 4 weeks old, purchased
from Medical Laboratory Animal Center of Hubei Province) weredivided into two groups. All animals were injected with 100 �l ofprocaine hydrochloride (0.5%) as the pretreatment at 24 h beforethe immunization, and then 10 mice were injected intramuscu-larly in quadriceps with 50 �g of pcS/2SS plasmid, and five micewere immunized with physiological saline as a control. To investi-gate the integration into host cellular DNA following intramuscularinjection in mice, multiple tissues such as heart, liver, spleen, lung,kidney, stomach, muscle were removed from each animal after 3months of immunization. Quadriceps muscle was removed lastto avoid contamination of other tissues. Tissues were rinsed inphosphate-buffered saline (PBS), and stored at −70 ◦C until DNAwas isolated.
Detection of antibodies against somatostatin
Specific antibodies were detected with an indirect ELISAmethod, using somatostatin-14 (S9129, Sigma–Aldrich) as standardantigen. Briefly, 96-well immunoplates (Greiner bio-one, Germany)were coated with 100 ng of somatostatin diluted in bicarbonatebuffer (pH 9.6) overnight at 4 ◦C. Following blocking with 5% (w/v)dry skimmed milk in PBS (pH 7.2) for 1 h at 37 ◦C, 1:50 dilutionof plasma samples in PBST (0.05% Tween-20 in PBS) were addedto the wells and the plates were incubated for 1 h at 37 ◦C. Eachsample was tested in triplicate, meanwhile positive and negativecontrols were also used. Bound antibody was detected by the addi-tion of HRP-labeled goat anti-rat IgG antibodies (Boster, Wuhan,China) diluted 1/2500 in PBST and incubated for 1 h at 37 ◦C.To develop the ELISA result, 10 mg of tetramethylbenzidine (TMBtablets, Sigma–Aldrich) dissolved in absolute ethanol was mixedwith phosphate–citrate buffer (pH 5.0) using H2O2 as the substrate,then incubated for 10 min at 37 ◦C. Reactions were terminated with2 M H2SO4 and the resulting optical density (OD) was measured at450 nm in a plate reader (Thermo Electron Corporation).
Analysis of genomic DNA
The genomic DNA was extracted according to a protocolreported by literature [22]. The purified DNA preparations wereshown to have A260/A280 ratios of ≥1.8 (data not shown), be of highmolecular weight, and be devoid of detectable RNA. Then sampleswere analyzed by sensitive PCR method. Generally, genomic DNAsamples were assayed for house gene, i.e. GAPDH to detect the qual-ity of isolation DNA, then assayed for the target gene (S/2SS). Theprimers of PGAPDH and PS/2SS were shown in the Table 1. The resultswere analyzed by agarose gel electrophoresis.
Statistics
Immunized rats were weighted at 0, 1, 2, 4 and 9 weeks of age.Data were analyzed using the Microsoft Excel statistics package.The comparison between two means was statistically analyzed byStudent’s t-test with Stat view.
Results
Identification of recombinant pcS/2SS plasmid
To construct the S/2SS expression vector, the entire S gene wasamplified by PCR from pcMV-S vector, the cloned fragment (696 bp)was verified by agarose gel and sequencing (Fig. 1). By double diges-tion with EcoRI and KpnI, S gene was subcloned into the pUC vector,designated as pUS (Fig. 2). Accordingly, SS gene was synthesized and
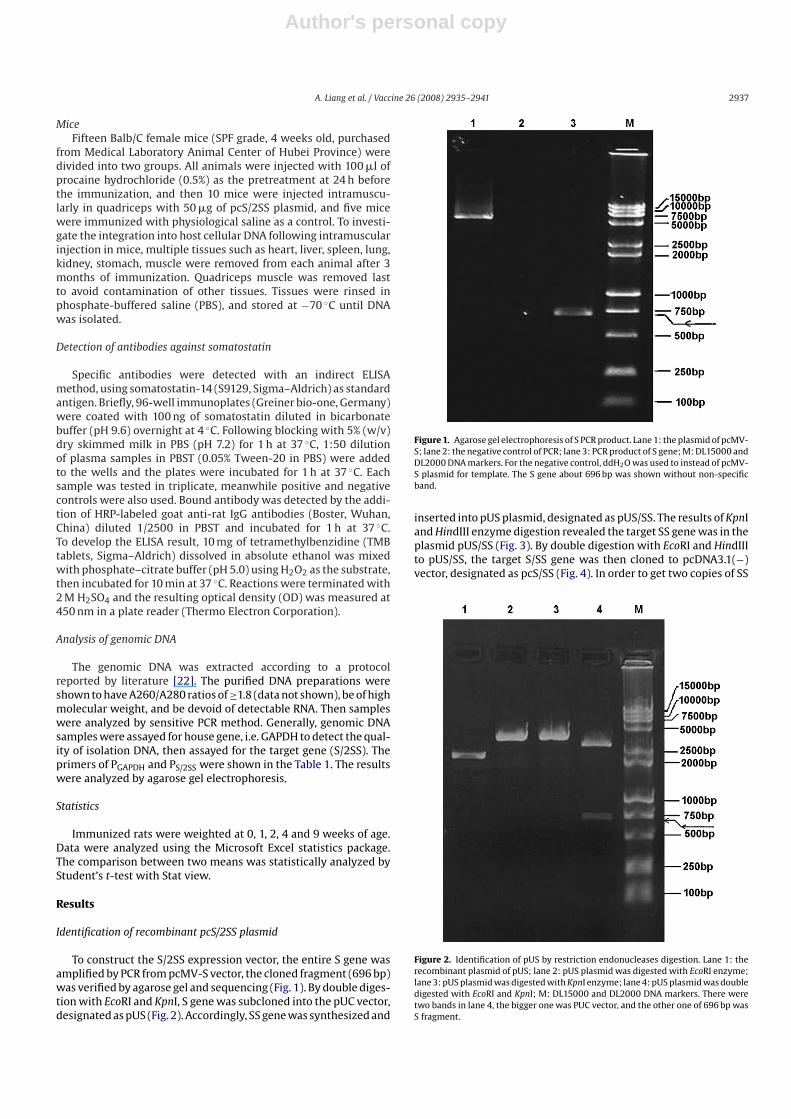
Figure 1. Agarose gel electrophoresis of S PCR product. Lane 1: the plasmid of pcMV-S; lane 2: the negative control of PCR; lane 3: PCR product of S gene; M: DL15000 andDL2000 DNA markers. For the negative control, ddH2O was used to instead of pcMV-S plasmid for template. The S gene about 696 bp was shown without non-specificband.
inserted into pUS plasmid, designated as pUS/SS. The results of KpnIand HindIII enzyme digestion revealed the target SS gene was in theplasmid pUS/SS (Fig. 3). By double digestion with EcoRI and HindIIIto pUS/SS, the target S/SS gene was then cloned to pcDNA3.1(−)vector, designated as pcS/SS (Fig. 4). In order to get two copies of SS
Figure 2. Identification of pUS by restriction endonucleases digestion. Lane 1: therecombinant plasmid of pUS; lane 2: pUS plasmid was digested with EcoRI enzyme;lane 3: pUS plasmid was digested with KpnI enzyme; lane 4: pUS plasmid was doubledigested with EcoRI and KpnI; M: DL15000 and DL2000 DNA markers. There weretwo bands in lane 4, the bigger one was PUC vector, and the other one of 696 bp wasS fragment.
Author's personal copy
2938 A. Liang et al. / Vaccine 26 (2008) 2935–2941
Figure 3. Identification of pUS/SS by restriction endonucleases digestion. Lane 1:the recombinant plasmid of pUS/SS; lane 2: pUS/SS plasmid was digested with KpnIenzyme; lane 3: pUS/SS plasmid was digested with HindIII enzyme; lane 4: pUS/SSwas digested with KpnI and HindIII; M: DL15000 and DL2000 DNA markers. Thechemically synthesized SS gene was inserted into the 3′ end of S gene in PUS to formthe fusion gene S/SS. From results of single digestion (lanes 2 and 3) and doubledigestion (lane 4), we found that the SS gene was cloned into pUS vector.
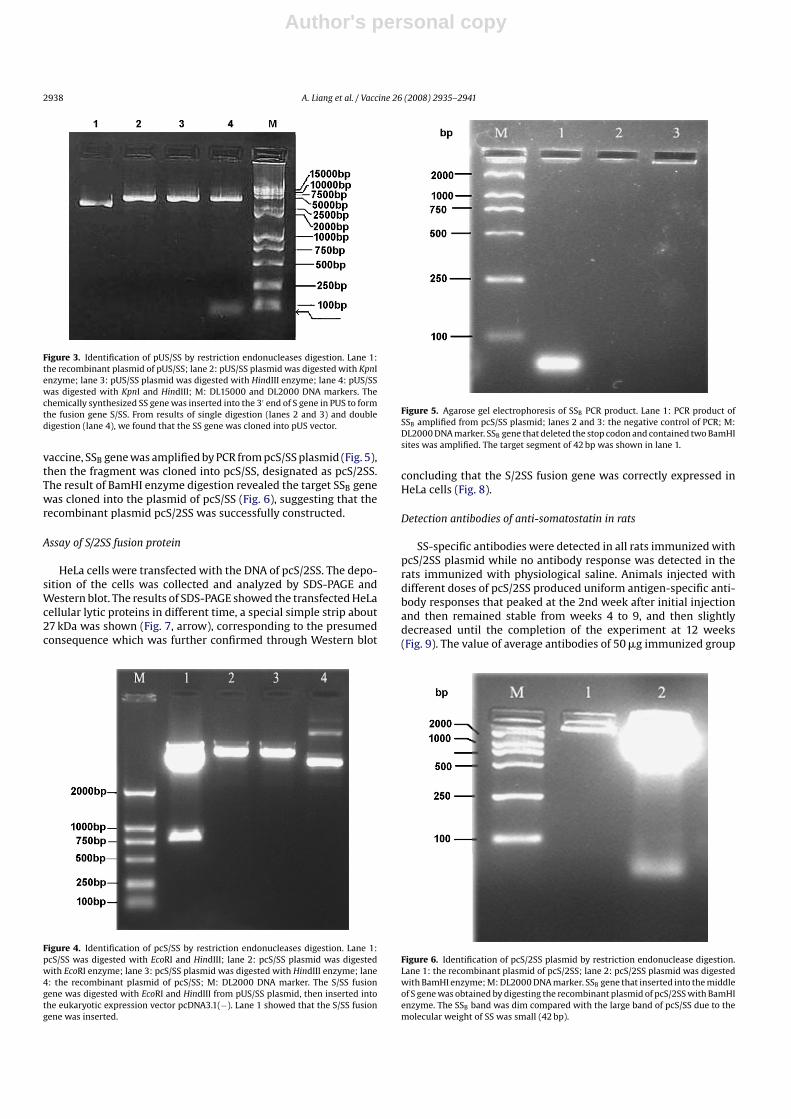
vaccine, SSB gene was amplified by PCR from pcS/SS plasmid (Fig. 5),then the fragment was cloned into pcS/SS, designated as pcS/2SS.The result of BamHI enzyme digestion revealed the target SSB genewas cloned into the plasmid of pcS/SS (Fig. 6), suggesting that therecombinant plasmid pcS/2SS was successfully constructed.
Assay of S/2SS fusion protein
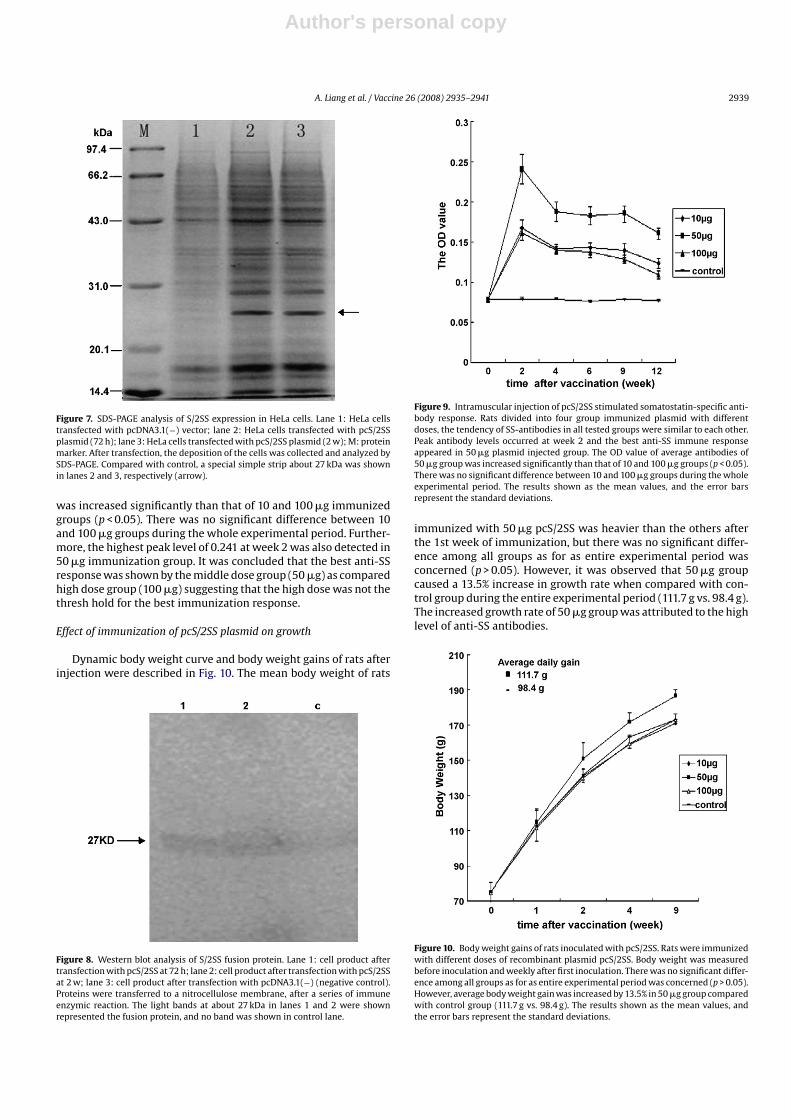
HeLa cells were transfected with the DNA of pcS/2SS. The depo-sition of the cells was collected and analyzed by SDS-PAGE andWestern blot. The results of SDS-PAGE showed the transfected HeLacellular lytic proteins in different time, a special simple strip about27 kDa was shown (Fig. 7, arrow), corresponding to the presumedconsequence which was further confirmed through Western blot
Figure 4. Identification of pcS/SS by restriction endonucleases digestion. Lane 1:pcS/SS was digested with EcoRI and HindIII; lane 2: pcS/SS plasmid was digestedwith EcoRI enzyme; lane 3: pcS/SS plasmid was digested with HindIII enzyme; lane4: the recombinant plasmid of pcS/SS; M: DL2000 DNA marker. The S/SS fusiongene was digested with EcoRI and HindIII from pUS/SS plasmid, then inserted intothe eukaryotic expression vector pcDNA3.1(−). Lane 1 showed that the S/SS fusiongene was inserted.
Figure 5. Agarose gel electrophoresis of SSB PCR product. Lane 1: PCR product ofSSB amplified from pcS/SS plasmid; lanes 2 and 3: the negative control of PCR; M:DL2000 DNA marker. SSB gene that deleted the stop codon and contained two BamHIsites was amplified. The target segment of 42 bp was shown in lane 1.
concluding that the S/2SS fusion gene was correctly expressed inHeLa cells (Fig. 8).
Detection antibodies of anti-somatostatin in rats
SS-specific antibodies were detected in all rats immunized withpcS/2SS plasmid while no antibody response was detected in therats immunized with physiological saline. Animals injected withdifferent doses of pcS/2SS produced uniform antigen-specific anti-body responses that peaked at the 2nd week after initial injectionand then remained stable from weeks 4 to 9, and then slightlydecreased until the completion of the experiment at 12 weeks(Fig. 9). The value of average antibodies of 50 �g immunized group
Figure 6. Identification of pcS/2SS plasmid by restriction endonuclease digestion.Lane 1: the recombinant plasmid of pcS/2SS; lane 2: pcS/2SS plasmid was digestedwith BamHI enzyme; M: DL2000 DNA marker. SSB gene that inserted into the middleof S gene was obtained by digesting the recombinant plasmid of pcS/2SS with BamHIenzyme. The SSB band was dim compared with the large band of pcS/SS due to themolecular weight of SS was small (42 bp).
Author's personal copy
A. Liang et al. / Vaccine 26 (2008) 2935–2941 2939
Figure 7. SDS-PAGE analysis of S/2SS expression in HeLa cells. Lane 1: HeLa cellstransfected with pcDNA3.1(−) vector; lane 2: HeLa cells transfected with pcS/2SSplasmid (72 h); lane 3: HeLa cells transfected with pcS/2SS plasmid (2 w); M: proteinmarker. After transfection, the deposition of the cells was collected and analyzed bySDS-PAGE. Compared with control, a special simple strip about 27 kDa was shownin lanes 2 and 3, respectively (arrow).
was increased significantly than that of 10 and 100 �g immunizedgroups (p < 0.05). There was no significant difference between 10and 100 �g groups during the whole experimental period. Further-more, the highest peak level of 0.241 at week 2 was also detected in50 �g immunization group. It was concluded that the best anti-SSresponse was shown by the middle dose group (50 �g) as comparedhigh dose group (100 �g) suggesting that the high dose was not thethresh hold for the best immunization response.
Effect of immunization of pcS/2SS plasmid on growth
Dynamic body weight curve and body weight gains of rats afterinjection were described in Fig. 10. The mean body weight of rats
Figure 8. Western blot analysis of S/2SS fusion protein. Lane 1: cell product aftertransfection with pcS/2SS at 72 h; lane 2: cell product after transfection with pcS/2SSat 2 w; lane 3: cell product after transfection with pcDNA3.1(−) (negative control).Proteins were transferred to a nitrocellulose membrane, after a series of immuneenzymic reaction. The light bands at about 27 kDa in lanes 1 and 2 were shownrepresented the fusion protein, and no band was shown in control lane.
Figure 9. Intramuscular injection of pcS/2SS stimulated somatostatin-specific anti-body response. Rats divided into four group immunized plasmid with differentdoses, the tendency of SS-antibodies in all tested groups were similar to each other.Peak antibody levels occurred at week 2 and the best anti-SS immune responseappeared in 50 �g plasmid injected group. The OD value of average antibodies of50 �g group was increased significantly than that of 10 and 100 �g groups (p < 0.05).There was no significant difference between 10 and 100 �g groups during the wholeexperimental period. The results shown as the mean values, and the error barsrepresent the standard deviations.
immunized with 50 �g pcS/2SS was heavier than the others afterthe 1st week of immunization, but there was no significant differ-ence among all groups as for as entire experimental period wasconcerned (p > 0.05). However, it was observed that 50 �g groupcaused a 13.5% increase in growth rate when compared with con-trol group during the entire experimental period (111.7 g vs. 98.4 g).The increased growth rate of 50 �g group was attributed to the highlevel of anti-SS antibodies.
Figure 10. Body weight gains of rats inoculated with pcS/2SS. Rats were immunizedwith different doses of recombinant plasmid pcS/2SS. Body weight was measuredbefore inoculation and weekly after first inoculation. There was no significant differ-ence among all groups as for as entire experimental period was concerned (p > 0.05).However, average body weight gain was increased by 13.5% in 50 �g group comparedwith control group (111.7 g vs. 98.4 g). The results shown as the mean values, andthe error bars represent the standard deviations.
Author's personal copy
2940 A. Liang et al. / Vaccine 26 (2008) 2935–2941
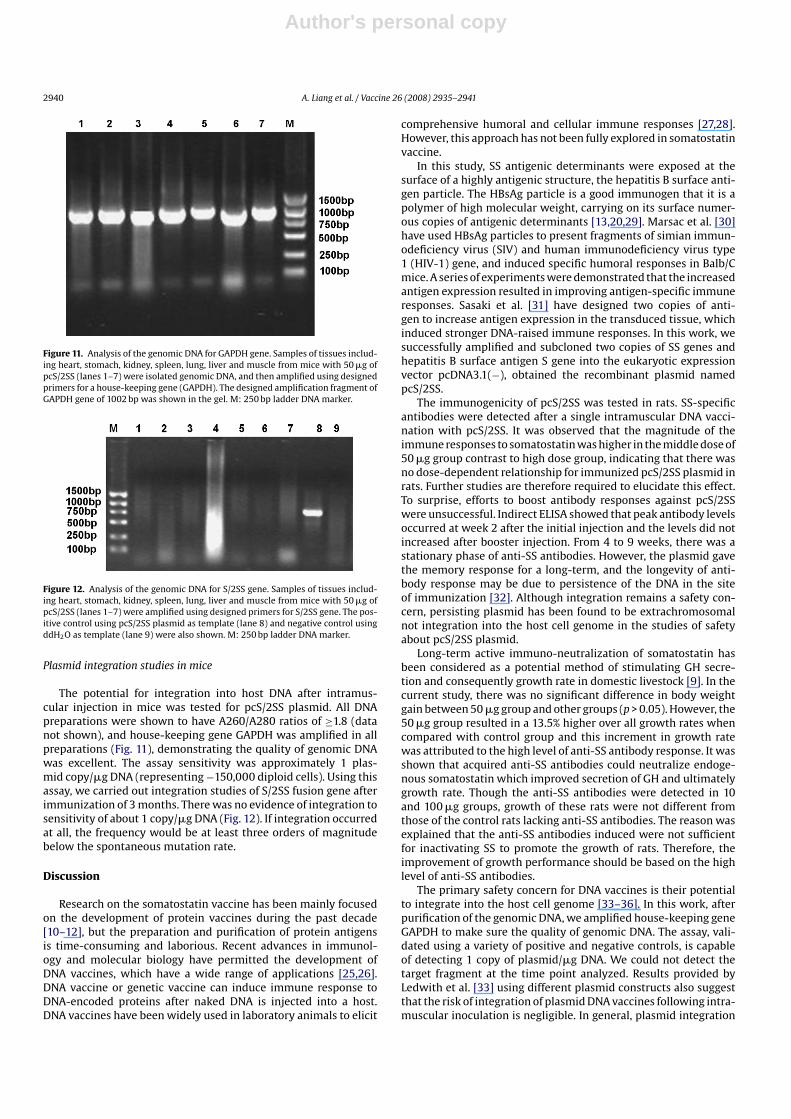
Figure 11. Analysis of the genomic DNA for GAPDH gene. Samples of tissues includ-ing heart, stomach, kidney, spleen, lung, liver and muscle from mice with 50 �g ofpcS/2SS (lanes 1–7) were isolated genomic DNA, and then amplified using designedprimers for a house-keeping gene (GAPDH). The designed amplification fragment ofGAPDH gene of 1002 bp was shown in the gel. M: 250 bp ladder DNA marker.
Figure 12. Analysis of the genomic DNA for S/2SS gene. Samples of tissues includ-ing heart, stomach, kidney, spleen, lung, liver and muscle from mice with 50 �g ofpcS/2SS (lanes 1–7) were amplified using designed primers for S/2SS gene. The pos-itive control using pcS/2SS plasmid as template (lane 8) and negative control usingddH2O as template (lane 9) were also shown. M: 250 bp ladder DNA marker.
Plasmid integration studies in mice
The potential for integration into host DNA after intramus-cular injection in mice was tested for pcS/2SS plasmid. All DNApreparations were shown to have A260/A280 ratios of ≥1.8 (datanot shown), and house-keeping gene GAPDH was amplified in allpreparations (Fig. 11), demonstrating the quality of genomic DNAwas excellent. The assay sensitivity was approximately 1 plas-mid copy/�g DNA (representing −150,000 diploid cells). Using thisassay, we carried out integration studies of S/2SS fusion gene afterimmunization of 3 months. There was no evidence of integration tosensitivity of about 1 copy/�g DNA (Fig. 12). If integration occurredat all, the frequency would be at least three orders of magnitudebelow the spontaneous mutation rate.
Discussion
Research on the somatostatin vaccine has been mainly focusedon the development of protein vaccines during the past decade[10–12], but the preparation and purification of protein antigensis time-consuming and laborious. Recent advances in immunol-ogy and molecular biology have permitted the development ofDNA vaccines, which have a wide range of applications [25,26].DNA vaccine or genetic vaccine can induce immune response toDNA-encoded proteins after naked DNA is injected into a host.DNA vaccines have been widely used in laboratory animals to elicit
comprehensive humoral and cellular immune responses [27,28].However, this approach has not been fully explored in somatostatinvaccine.
In this study, SS antigenic determinants were exposed at thesurface of a highly antigenic structure, the hepatitis B surface anti-gen particle. The HBsAg particle is a good immunogen that it is apolymer of high molecular weight, carrying on its surface numer-ous copies of antigenic determinants [13,20,29]. Marsac et al. [30]have used HBsAg particles to present fragments of simian immun-odeficiency virus (SIV) and human immunodeficiency virus type1 (HIV-1) gene, and induced specific humoral responses in Balb/Cmice. A series of experiments were demonstrated that the increasedantigen expression resulted in improving antigen-specific immuneresponses. Sasaki et al. [31] have designed two copies of anti-gen to increase antigen expression in the transduced tissue, whichinduced stronger DNA-raised immune responses. In this work, wesuccessfully amplified and subcloned two copies of SS genes andhepatitis B surface antigen S gene into the eukaryotic expressionvector pcDNA3.1(−), obtained the recombinant plasmid namedpcS/2SS.
The immunogenicity of pcS/2SS was tested in rats. SS-specificantibodies were detected after a single intramuscular DNA vacci-nation with pcS/2SS. It was observed that the magnitude of theimmune responses to somatostatin was higher in the middle dose of50 �g group contrast to high dose group, indicating that there wasno dose-dependent relationship for immunized pcS/2SS plasmid inrats. Further studies are therefore required to elucidate this effect.To surprise, efforts to boost antibody responses against pcS/2SSwere unsuccessful. Indirect ELISA showed that peak antibody levelsoccurred at week 2 after the initial injection and the levels did notincreased after booster injection. From 4 to 9 weeks, there was astationary phase of anti-SS antibodies. However, the plasmid gavethe memory response for a long-term, and the longevity of anti-body response may be due to persistence of the DNA in the siteof immunization [32]. Although integration remains a safety con-cern, persisting plasmid has been found to be extrachromosomalnot integration into the host cell genome in the studies of safetyabout pcS/2SS plasmid.
Long-term active immuno-neutralization of somatostatin hasbeen considered as a potential method of stimulating GH secre-tion and consequently growth rate in domestic livestock [9]. In thecurrent study, there was no significant difference in body weightgain between 50 �g group and other groups (p > 0.05). However, the50 �g group resulted in a 13.5% higher over all growth rates whencompared with control group and this increment in growth ratewas attributed to the high level of anti-SS antibody response. It wasshown that acquired anti-SS antibodies could neutralize endoge-nous somatostatin which improved secretion of GH and ultimatelygrowth rate. Though the anti-SS antibodies were detected in 10and 100 �g groups, growth of these rats were not different fromthose of the control rats lacking anti-SS antibodies. The reason wasexplained that the anti-SS antibodies induced were not sufficientfor inactivating SS to promote the growth of rats. Therefore, theimprovement of growth performance should be based on the highlevel of anti-SS antibodies.
The primary safety concern for DNA vaccines is their potentialto integrate into the host cell genome [33–36]. In this work, afterpurification of the genomic DNA, we amplified house-keeping geneGAPDH to make sure the quality of genomic DNA. The assay, vali-dated using a variety of positive and negative controls, is capableof detecting 1 copy of plasmid/�g DNA. We could not detect thetarget fragment at the time point analyzed. Results provided byLedwith et al. [33] using different plasmid constructs also suggestthat the risk of integration of plasmid DNA vaccines following intra-muscular inoculation is negligible. In general, plasmid integration

Author's personal copy
A. Liang et al. / Vaccine 26 (2008) 2935–2941 2941
into genomic DNA occurs by tandem repeats [37]. The frequencyof integration into the cellular genome could be affected by sev-eral factors, such as the plasmid sequence, the presence of chi-likeelements [37], Alu segments [38] and minisatellite regions [39].However, the integration of bacterial plasmid DNA is not quite sim-plistic. The mammalian genome appears to possess a mechanism toprotect its integrity [40]. If integration had occurred, the frequencywas ≤1–8 integrations per 150,000 diploid cells, which would be atleast three orders of magnitude below the spontaneous mutationrate. So, the risk of mutation plasmid due to integration of pcS/2SSplasmid in mice could be nil in the current study.
In summary, we have constructed a potent vaccine to promotegrowth of animals. The recombinant plasmid pcS/2SS could expresstarget protein with good immunogenicity and safety in eukaryotichosts. Moreover, an injection of the middle dose plasmid couldenhance the growth of rats. Further studies will be required toexplore its mechanism especially in dose aspect and apply thistechnology to large animals.
Acknowledgement
This study was carried out with the financial support by theNational Natural Science Foundation of China (no.: 30771549).
References
[1] Mullis PE. Genetic control of growth. Eur J Endocrinol 2005;152(1):11–31.[2] Tannenbaum GS, Epelbaum J, Bowers CY. Interrelationship between the
novel peptide ghrelin and somatostatin/growth hormone-releasing hor-mone in regulation of pulsatile growth hormone secretion. Endocrinology2003;144(3):967–74.
[3] Rettori V, Milenkovic L, Aquila MC, McCann SM. Physiologically significanteffect of neuropeptide Y to suppress growth hormone release by stimulatingsomatostatin discharge. Endocrinology 1990;126(5):2296–301.
[4] Bluet-Pajot MT, Bertherat J, Epelbaum J, Kordon C. Neural and pituitarymechanisms involved in growth hormone regulation. J Pediatr Endocrinol1993;6(3/4):357–69.
[5] Katakami H, Arimura A, Frohman LA. Growth hormone (GH)-releasing factorstimulates hypothalamic somatostatin release: an inhibitory feedback effect onGH secretion. Endocrinology 1986;118(5):1872–7.
[6] Dawson JM, Soar JB, Buttery PJ, Craigon J, Gill M, Beever DE. The effect ofimmunization against somatostatin and �-agonist administration alone andin combination on growth carcass composition in young steers. Anim Sci1997;64:37–51.
[7] Elsaesser F, Drath S. The potential of immuno-neutralization of somatostatinfor improving pig performance. Livest Prod Sci 1995;42:255–63.
[8] Mears GJ. Immunization of lambs against somatostatin improve growth rate.Can J Anim Sci 1990;70:1091–7.
[9] Magnan E, Mazzocchi L, Cataldi M, Guillaume V, Dutour A, Dadoun F, et al. Effectof actively immunizing sheep against growth hormone-releasing hormone orsomatostatin on spontaneous pulsatile and neostigmine-induced growth hor-mone secretion. J Endocrinol 1995;144(1):83–90.
[10] Wang NX, Chen WH, Du NX. Effect of somatostatin genetic engineering vaccineon growth and immuno-function of rat. Chin J Vet Sci 2002;22:427–9.
[11] Du NX, Yang H, Ji CY, Gong WB, Zhu HF. Reports on field trial of somatostatingenetic engineering vaccine. Anim Husband Vet Med 2001;33(2):23–4.
[12] Xu WZ, Du NX, Li GD, Wang Y, Li ZP. A new genetically engineered vaccine foranimal growth promotion. Sci China B 1994;37(10):1234–41.
[13] Xu WZ, Du NX, Li GD. A antigen vector of polyvalent vaccine in gene engineeringHBsAg polymer particular antigen. Chin J Virol 1991;7:383–6.
[14] Leitner WW, Ying H, Restifo NP. DNA and RNA-based vaccines: principles,progress and prospects. Vaccine 2000;18(9–10):765–77.
[15] Ada G, Ramshaw I. DNA vaccination. Expert Opin Emerg Drugs2003;8(1):27–35.
[16] Apostolopoulos V, Plebanski M. The evolution of DNA vaccines. Curr Opin MolTher 2000;2(4):441–7.
[17] Donnelly JJ, Wahren B, Liu MA. DNA vaccines: progress and challenges. JImmunol 2005;175(2):633–9.
[18] Ihata A, Watabe S, Okuda K. DNA vaccination. Nippon Rinsho2006;64(10):1865–9.
[19] Liu MA, Wahren B, Karlsson Hedestam GB. DNA vaccines: recent developmentsand future possibilities. Hum Gene Ther 2006;17(11):1051–61.
[20] Fomsgaard A, Nielsen HV, Bryder K, Nielsen C, Machuca R, Bruun L, et al.Improved humoral and cellular immune responses against the gp120 V3 loopof HIV-1 following genetic immunization with a chimeric DNA vaccine encod-ing the V3 inserted into the hepatitis B surface antigen. Scand J Immunol1998;47(4):289–95.
[21] Le Borgne S, Mancini M, Le Grand R, Schleef M, Dormont D, Tiollais P, et al. Invivo induction of specific cytotoxic T lymphocytes in mice and rhesus macaquesimmunized with DNA vector encoding an HIV epitope fused with hepatitis Bsurface antigen. Virology 1998;240(2):304–15.
[22] Sambrook J, Fritsch EF, Maniatis T. Molecular cloning: a laboratory manual. 3rded. New York: Cold Spring Harbor Laboratory Press; 2001.
[23] Kozak M. An analysis of 5′-noncoding sequences from 699 vertebrate messen-ger RNAs. Nucleic Acids Res 1987;15(20):8125–48 [Review].
[24] Iida Y, Masuda T. Strength of translation initiation signal sequence of mRNAas studied by quantification method: effect of nucleotide substitutionsupon translation efficiency in rat preproinsulin mRNA. Nucleic Acids Res1996;24(17):3313–6.
[25] Ulmer JB, Wahren B, Liu MA. DNA vaccines: recent technological and clinicaladvances. Discov Med 2006;6(33):109–12.
[26] Garmory HS, Brown KA, Titball RW. DNA vaccines: improving expression ofantigens. Genet Vaccines Ther 2003;1(1):2.
[27] Ellis RW. Technologies for the design, discovery, formulation and administra-tion of vaccines. Vaccine 2001;19(17–19):2681–7.
[28] Tang DC, DeVit M, Johnston SA. Genetic immunization is a simple method foreliciting an immune response. Nature 1992;356(6365):152–4.
[29] Woo WP, Doan T, Herd KA, Netter HJ, Tindle RW. Hepatitis B surface antigen vec-tor delivers protective cytotoxic T-lymphocyte responses to disease-relevantforeign epitopes. Virology 2006;80(8):3975–84.
[30] Marsac D, Puaux AL, Riviere Y, Michel ML. In vivo induction of cel-lular and humoral immune responses by hybrid DNA vectors encodingsimian/human immunodeficiency virus/hepatitis B surface antigen virus par-ticles in BALB/c and HLA-A2-transgenic mice. Immunobiology 2005;210(5):305–19.
[31] Sasaki S, Takeshita F, Oikawa T, Kojima Y, Xin KQ, Okuda K, et al. Improvementof DNA vaccine immunogenicity by a dual antigen expression system. BiochemBiophys Res Commun 2004;315(1):38–43.
[32] Van Harmelen JH, Shephard E, Thomas R, Hanke T, Williamson AL, Williamson C.Construction and characterisation of a candidate HIV-1 subtype C DNA vaccinefor South Africa. Vaccine 2003;21(27–30):4380–9.
[33] Ledwith BJ, Manam S, Troilo PJ, Barnum AB, Pauley CJ, Griffiths II TG, et al. Plas-mid DNA Vaccines: investigation of integration into host cellular DNA followingintramuscular injection in mice. Intervirology 2000;43(4–6):258–72.
[34] Medjitna TD, Stadler C, Bruckner L, Griot C, Ottiger HP. DNA vaccines: safetyaspect assessment and regulation. Dev Biol (Basel) 2006;126:261–70.
[35] Ledwith BJ, Manam S, Troilo PJ, Barnum AB, Pauley CJ, Griffiths II TG, et al.Plasmid DNA vaccines: assay for integration into host genomic DNA. Dev Biol(Basel) 2000;104:33–43.
[36] Wang Z, Troilo PJ, Wang X, Griffiths TG, Pacchione SJ, Barnum AB, et al. Detectionof integration of plasmid DNA into host genomic DNA following intramuscularinjection and electroporation. Gene Ther 2004;11(8):711–21.
[37] Manam S, Ledwith BJ, Barmum AB, Troilo PJ, Pauley CJ, Harper LB, et al.Plasmid DNA vaccines: tissue distribution and effects of DNA sequence,adjuvants and delivery method on integration into host DNA. Intervirology2000;43(4–6):273–81.
[38] Rudiger NS, Gregersen N, Kielland-Brandt MC. One short well conserved regionof Alu-sequences is involved in human gene rearrangements and has homologywith prokaryotic chi. Nucleic Acids Res 1995;23(2):256–60.
[39] Krowczynska AM, Rudders RA, Krontiris TG. The human minisatelliteconsensus at breakpoints of oncogene translocations. Nucleic Acids Res1990;18(5):1121–7.
[40] Scrable H, Stambrook PJ. A genetic program for deletion of foreign DNA fromthe mammalian genome. Mutat Res 1999;429(2):225–37.





























