Embed Size (px)
Citation preview

Conserved shell disguises diversity in Mesodontrachia landsnails from the Australian Monsoon Tropics (Gastropoda:Camaenidae)FRANCESCO CRISCIONE & FRANK KÖHLER
Submitted: 6 February 2013Accepted: 12 March 2013doi:10.1111/zsc.12011
Criscione, F., K€ohler F. Conserved shell disguises diversity in Mesodontrachia land snailsfrom the Australian Monsoon Tropics (Gastropoda: Camaenidae). —Zoologica Scripta, 42,389–405.We comprehensively revise the taxonomy of the camaenid genus Mesodontrachia Solem,1985, which is endemic to the Victoria River District and East Kimberley in the north-wes-tern Australian Monsoon Tropics based on comparative analyses of key morphological fea-tures and mitochondrial DNA sequences. We examined newly collected samples fromseveral collection sites spread over nearly 20 000 km2 of mostly inaccessible land, whichrepresented all three currently known and three previously undescribed species. All specieswere initially identified as members of Mesodontrachia based on their similar, putativelytypical shell. However, Mesodontrachia as so delimited was polyphyletic in a mitochondrialphylogeny with respect to several other camaenid genera from NW Australia. Contrary tothe shell, we found considerable variation in the penial anatomy that was consistent withthe mtDNA differentiation. To retain monophyletic taxa, we propose a revised taxonomywhereby Mesodontrachia is maintained as a monotypic taxon. In addition, four genera (Nodu-labium, Ototrachia, Pseudomesodontrachia and Vincentrachia) and three species (P. gregoriana,O. compressa and N. solidum) are newly described. The shell of all these taxa is highly con-served and of little taxonomic utility. Shell similarity is attributed to a similar life style in asimilar and harsh environment.Corresponding author: Frank K€ohler, Australian Museum, 6 College St., Sydney, NSW 2010,Australia. E-mail: [email protected] Criscione, Australian Museum, 6 College Street, Sydney, NSW, 2010, Australia.Frank Köhler, Australian Museum, 6 College Street, Sydney, NSW, 2010, Australia.
IntroductionThe Camaenidae arguably are the most diverse and proba-bly also the best known land snail family in Australia. Cam-aenids have extensively radiated throughout most of thecontinent occupying a wide range of habitats (Solem 1998).Their species richness is generally larger in humid andtopologically more complex regions, such as the WesternAustralian Kimberley, the rocky ranges in central Australiaand in particular in the Australian Mesic Zone along theeastern coast (Hugall & Stanisic 2011). By contrast, morearid and topologically less complex regions, such as most ofthe Australian Monsoon Tropics (AMT), support lowernumbers of species with generally wider distributions ofsingle species (Cameron 1992; Slatyer et al. 2007).The Camaenidae of the western part of the AMT (com-
prising the Western Australian Kimberley region and
adjacent areas in the Northern Territory) have comprehen-sively been revised by Alan Solem, who published a seriesof morphology-based taxonomic revisions (e.g. Solem 1979,1981a,b, 1985, 1997). These treatments achieved the mostcomplete documentation of the camaenid fauna in NWAustralia so far, providing the systematic framework forongoing studies.However, vast areas in the northern part of the North-
ern Territory have remained poorly surveyed mostly dueto their inaccessibility. Studies based on historical museummaterial have recently indicated that yet unsurveyedregions in the AMT may harbour phylogenetically distinctand endemic groups of camaenids (K€ohler 2012; K€ohler &Criscione 2013).To improve our understanding of the land snail diversity
in the AMT, between 2009 and 2010, a helicopter-based
ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405 389
Zoologica Scripta

survey has been conducted in the Victoria River District(VRD) and adjacent East Kimberley (EK). Based on mate-rial collected during this survey, recently we revised thetaxonomy of the camaenid genus Exiligada Iredale, 1939 bycombining analyses of molecular and anatomical data (Cri-scione et al. 2012). In this study, we found an unexpectedhigh number of previously unknown species boosting thenumber of described Exiligada species from one to fifteen.Most of these species were found in previously poorly sam-pled areas and were collected for the first time ever. How-ever, we also found several cryptic species that possessedvery similar shells. These species were not recognized asdistinct in previous, predominantly shell-based treatments,but had been subsumed under the name Exiligada negriensis(see Iredale 1939; Solem 1985). The Exiligada studyrevealed that camaenids from similar habitats may exhibitalmost identical shells and that species can properly bedelimited only through comparative examination of severalmorphological and molecular markers (Criscione et al.2012).Here, we continue our revision of the Camaenidae in
the VRD and EK by examining newly collected samplesattributed to Mesodontrachia Solem, 1985. Three specieshave previously been recognized within this genus mainlybased on their overall similar but in comparison with othercamaenid genera rather distinct shell (Solem 1985, 1988).Compared with other camaenids from the same region,shells of Mesodontrachia differ particularly in their largersize, broadly conical shape, smooth sculpture and browncolour. Having rather small known ranges, these speciesare considered to be narrowly endemic. In the VRD, twospecies are known: Mesodontrachia fitzroyana Solem, 1985 isfound on limestone outcrops between the Timber andSkull Creeks, and M. desmonda Solem, 1985 occurs onsandstone cliffs in the Saddle Creek drainage (Solem 1985).The third species M. cockburnensis inhabits mixed rock talusin the Cockburn Ranges of the EK (Solem 1988). Based onmaterial of all three known and further undescribed spe-cies, this study aimed at resolving the phylogenetic rela-tionships of Mesodontrachia and to complete thedocumentation of patterns of camaenid diversity in theVRD.
Materials and methodsMaterialThis study is based on newly collected, ethanol-preservedsamples and dry shells deposited in the malacological col-lections of the Australian Museum (AM), the Museum andArt Galleries of the Northern Territory (NTM) and theWestern Australian Museum (WAM) (Table S1). Collec-tion sites spanned an area of about 20 000 km2 throughoutthe Victoria River District in the Northern Territory and
the Eastern Kimberley in Western Australia. Samples fromthe same collection site are referred to as lots.
Molecular studiesDNA was extracted from small pieces of foot muscle fromup to four specimens per lot by use of a QIAGEN DNAextraction kit for animal tissue following the standard pro-cedure of the manual. Fragments of the mitochondrial 16SrRNA (16S) and cytochrome c oxidase subunit 1 (COI)genes were amplified by PCR using the primer pairs16Scs1 (Chiba 1999) and 16Sbd1 (Sutcharit et al. 2007),and L1490 and H2198 (Folmer et al. 1994), respectively.PCRs were performed using 0.5–1.0 Units of Red HotTM
thermostable DNA polymerase in a 10-fold dilution of buf-fer (Advanced Biotechnologies, Columbia, MD, USA:20 mM (NH4)2SO4, 750 mM Tris–HCL pH 9.0, 0.1% (w/v) Tween), with a final concentration of 0.05 mM dNTPs,3.5–4.5 mM MgCl2 and 12.5–25 pmol of each primer in atotal reaction volume of 25 ll. The cycling profile was asfollows: (a) for the 16S gene: (94°C for 5 min, annealing at55°C for 90 s, 72°C for 1 min) for one cycle, (94°C for30 s, annealing at 55°C for 90 s, 72°C for 1 min) for 40cycles and (94°C for 30 s, annealing at 55°C for 60 s, 72°Cfor 7 min); (b) for the COI gene: (94°C for 7 min, anneal-ing at 50°C for 1 min, 72°C for 2 min) for one cycle,(94°C for 1 min, annealing at 50°C for 1 min, 72°C for1 min) for 35 cycles and (94°C for 1 min, annealing at50°C for 1 min, 72°C for 7 min). Both strands of PCRfragments were purified and cycle sequenced by use of thePCR primers. Electropherograms were corrected for mis-reads, and forward and reverse strands were merged intoone sequence file using CodonCode Aligner v.3.6.1(CodonCode Corporation, Dedham, MA, USA). Sequenceshave been deposited in GenBank (Table S1).Sequence alignments were generated using MUSCLE as
implemented in MEGA5 (Tamura et al. 2011). Sequencesaturation was assessed for each mtDNA fragment usingtests implemented in DAMBE (Xia & Xie 2001; Xia et al.2003). Uncorrected pairwise genetic distances were calcu-lated using MEGA5 under the option ‘pairwise deletion ofgaps’. For phylogenetic analyses, 16S and COI sequenceswere concatenated into one partitioned data set. Prior tothe model-based phylogenetic analyses, the best-fit modelof nucleotide substitution was identified for each gene par-tition separately using the model proposal function ofMEGA5. Partitioned models were applied in the BI analy-ses with parameters estimated from the data set. ML analy-ses were performed by employing the more complex modelof sequence evolution to the entire data set as MEGA5 didnot allow for data partitioning. One-thousand ML boot-strap replicates were performed to assess the topology sup-port of the ML tree. Bayesian posterior probabilities of
390 ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405
Revision of the Australian camaenid Mesodontrachia � F. Criscione & F. K€ohler

phylogenetic trees were estimated by running a 107 genera-tions Metropolis-coupled Markov chain Monte Carlo (2runs with each 4 chains, of which one was heated) asimplemented by MrBayes v. 3.1.2 (Ronquist & Huelsen-beck 2003). A data partition was applied that allowedparameters to be estimated separately for each gene frag-ment and for each codon position of the COI gene. Sam-pling rate of the trees was 1000 generations. Generationssampled before the chain reached stationary were discardedas burn-in. Stationarity was reached when the average stan-dard deviation of split frequencies shown in MrBayes was<0.01 and the log likelihood of sampled trees reached a sta-tionary distribution (Ronquist & Huelsenbeck 2003).
Morphological studiesMorphological characters of adult shells (dimension, col-ouration and sculpture) were assessed from a representativenumber of randomly chosen adult specimens from each lot(more than 20, when possible). If the number of availablespecimens from one lot was too small to permit meaningfulstatistical comparisons, we included specimens from addi-tional collection sites in the immediate vicinity (i.e. in <10-km distance), if these had an identical shell sculpture.Adults were recognized by a complete apertural lip. Shellswere measured with callipers precise to 0.1 mm. Charactersmeasured were height (H = maximum dimension parallelto axis of coiling, including lip) and diameter (D = maxi-mum dimension perpendicular to H, including lip). Thenumber of whorls (W), including protoconch, was countedprecise to 0.1 as shown in K€ohler (2011a: fig. 2). Values ofD and H were plotted in SYSTAT v.15 (Systat SoftwareInc., Chicago, IL, USA) using the mtDNA tree topologyas a constraint. The 95% confidence ellipse option wasused to visualize the amount of overlap in the morphospaceoccupied by specimens of each clade. When there was littleoverlap, shells of clades were considered to be morphologi-cally distinct. Values of W were represented by means offrequency histograms to check for discontinuous distribu-tions among clades.Representatives of each mtDNA clade were dissected to
study the genital anatomy by use of a Leica M8 stereomicroscope with drawing mirror. Genital anatomy was stud-ied in two (in small lots) to five specimens per sample toconfirm that morphological features were consistent in spec-imens from the same clade. Anatomical features are reportedin the taxonomic descriptions at the end of the manuscript;the terminology used is the same as in K€ohler (2011a).
Abbreviations used16S, 16S rRNA gene; AM, Australian Museum, Sydney;AMT, Australian Monsoon Tropics; BI, Bayesian Infer-ence; COI, cytochrome c oxidase subunit 1 gene; D, shell
diameter; EK, Eastern Kimberley; ep, epiphallus; epd, epi-phallus-penis connecting duct; dry, number of dry shells;H, shell height; lp, longitudinal pilasters; ML, MaximumLikelihood; NP, National Park; NT, Northern Territory;NTM, Museum and Art Galleries of the Northern Terri-tory, Darwin; p, penis; rm, retractor muscle; sh, penialsheath; tp, tuberculated pilasters; va, vagina; vd, vas defer-ens; VRD, Victoria River District, NT; W, number ofwhorls of shell; WA, Western Australia; WAM, WesternAustralian Museum, Perth; wet, number of ethanol-pre-served specimens.
ResultsMolecular analysesBased on an explorative phylogenetic analysis of a compre-hensive 16S data set of the Australian Camaenidae, weselected a subset of more closely related camaenids, includ-ing a suitable outgroup, for more thorough analyses. Thisfinal data set contained concatenated sequences from 81camaenid specimens from the NT and WA, which repre-sented species of Mesondontrachia s. l. as well as eight othergenera (Table S1). The in-group comprised sequences of51 specimens of Mesodontrachia s. l., 39 of which were rep-resented by sequences from both genes, two by COIsequences only and ten by 16S sequences only. The miss-ing sequence fragments were coded as unknown. In addi-tion, species of the genera Cristilabrum, Exiligada,Mouldingia, Ningbingia, Ordtrachia, Prototrachia, Turgenitub-ulus and Westraltrachia from the EK and VRD wereincluded to test the monophyly of Mesodontrachia sensulato. Sequences of the Kimberley endemic Kimboraga wereused as outgroup to root the trees. The final concatenateddata set of aligned sequences of COI and 16S had a totallength of 1457 bp (16S: 802 bp, COI: 655 bp). Tests indi-cated no or little saturation in both mitochondrial frag-ments (Iss < Iss.c with P < 0.01). The hierarchicallikelihood test revealed the TN93 model (Tamura & Nei1993) with gamma distribution and proportions of invari-able sites (TN93 + Γ+I) as the best-fit model of sequenceevolution for 16S and the HKY model (Hasegawa et al.1985) with gamma distribution and proportions of invari-able sites (HKY+Γ+I) for COI.The BI and ML analyses produced trees with nearly
identical topologies (Figs 1 and 2). We generated a strictconsensus cladogram showing only the branches that wereconsistently resolved in both trees (Fig. 3). For clarity, thetrees depicted in Figs 1–3 were labelled with the names oftaxa recognized or described below. Accordingly, Mesodon-trachia s. l. was not a monophyletic group but formed apolyphyletic assemblage of six distinct, highly supportedclades that were separated from each other by furtherinterspersed clades occupied by species from different
ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405 391
F. Criscione & F. K€ohler � Revision of the Australian camaenid Mesodontrachia

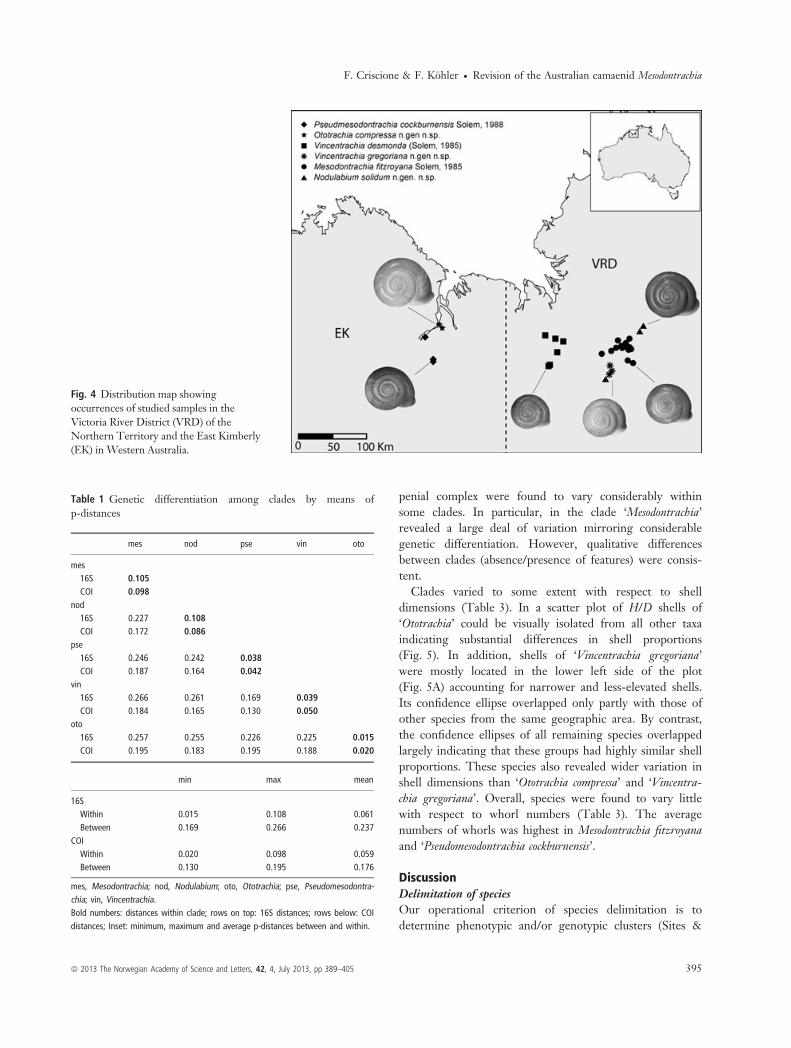
genera. The basal relationships between these cladesremained poorly supported or unresolved. Most clades haveallopatric distributions (Fig. 4). The clade ‘Mesodontrachiasensu stricto’ (BPP = 100%; BTSP = 99%) contained themajority of in-group sequences, which originated from theTimber Creek area in the VRD. While this clade revealedconsiderable genetic structuring, specimens from a singlepopulation (AM C.462802a-d) were distributed acrossalmost the entire clade. The clade ‘Nodulabium’
(BPP = 100%; BTSP = 99%) contained sequences fromlots with a disjunct distribution in the Western VRD. The
clade ‘Vincentrachia’ (BPP = 100%; BTSP = 99%) includedtwo clusters, one with sequences of specimens from a smallarea within the Gregory National Park and one with speci-mens identified as ‘Mesodontrachia desmonda Solem, 1985’.The clade ‘Ototrachia’ (BPP = 100%; BTSP = 99%) con-tained sequences of specimens from the western part of theEast Kimberley. All previous clades formed an essentiallyunresolved radiation together with clades formed by speciesof Mouldingia, Westraltrachia, Exiligada, Ordtrachia, Cristila-brum + Turgenitubulus, Ningbingia and Prototrachia, respec-tively. The clade ‘Pseudomesodontrachia’ contained a
Fig. 1 Bayesian phylogram based on analyses of the concatenated COI and 16S sequences. Numbers above branches indicate nodal support(%) by Bayesian posterior clade probabilities (BPP). Only BPP values � 90% are indicated. Support values of 100% are represented byasterisks.
392 ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405
Revision of the Australian camaenid Mesodontrachia � F. Criscione & F. K€ohler

specimen of ‘Mesodontrachia cockburnensis Solem, 1988’ andwas found to form the sister group of Prototrachia.The clades circumscribed above differed from each other
by mean uncorrected pairwise distances of about 17–27%(average = 24%) in 16S and 13–19% (average = 18%) inCOI (Table 1). There has been no overlap with meanuncorrected pairwise distances within clades rangingbetween 1% and 11% (average 6%) in 16S and between2% and 10% (average 6%) in COI.
Morphological analysesMorphological examinations and analyses of shell dimen-sions aimed to document and evaluate the amount ofmorphological differentiation within and between clades
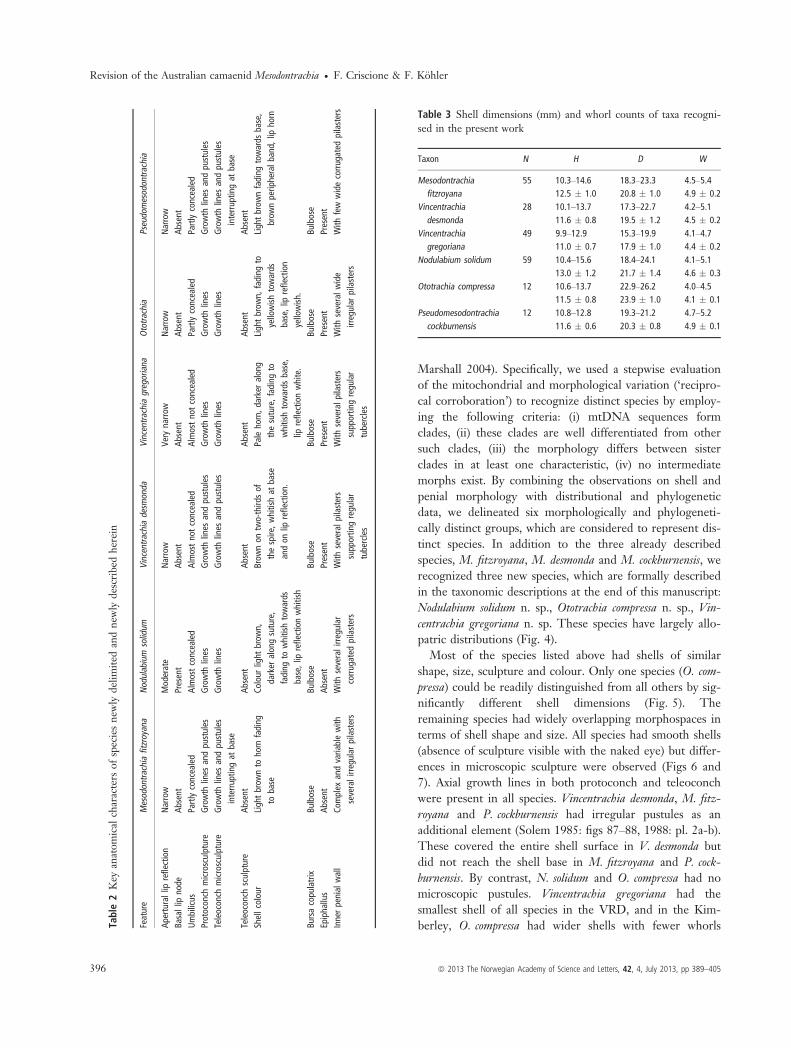
and species. For clarity, the following anatomical compari-sons are presented with reference to taxon names formallyintroduced further below. The underlying taxon delimita-tions resulted from combined analyses of molecular andanatomical differentiation but were not foregone conclu-sions preceding these analyses.We found that each clade was characterized by peculiar
features of the penial anatomy. Particularly, informativecharacters were thickness of penial sheath, length of penis,presence and development of epiphallus and sculpture ofinner penial wall. Characteristic features for each clade aregiven in Table 2; for further details refer to the anatomicaldescriptions at the end of this manuscript. The develop-ment of penial wall pilasters and the absolute size of the
Fig. 2 Maximum Likelihood phylogram based on analyses of the concatenated COI and 16S sequences. Numbers above branches indicatenodal support (%) by ML bootstrapping. Only BTSP values � 70% are indicated. Support values of 100% are represented by asterisks.
ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405 393
F. Criscione & F. K€ohler � Revision of the Australian camaenid Mesodontrachia
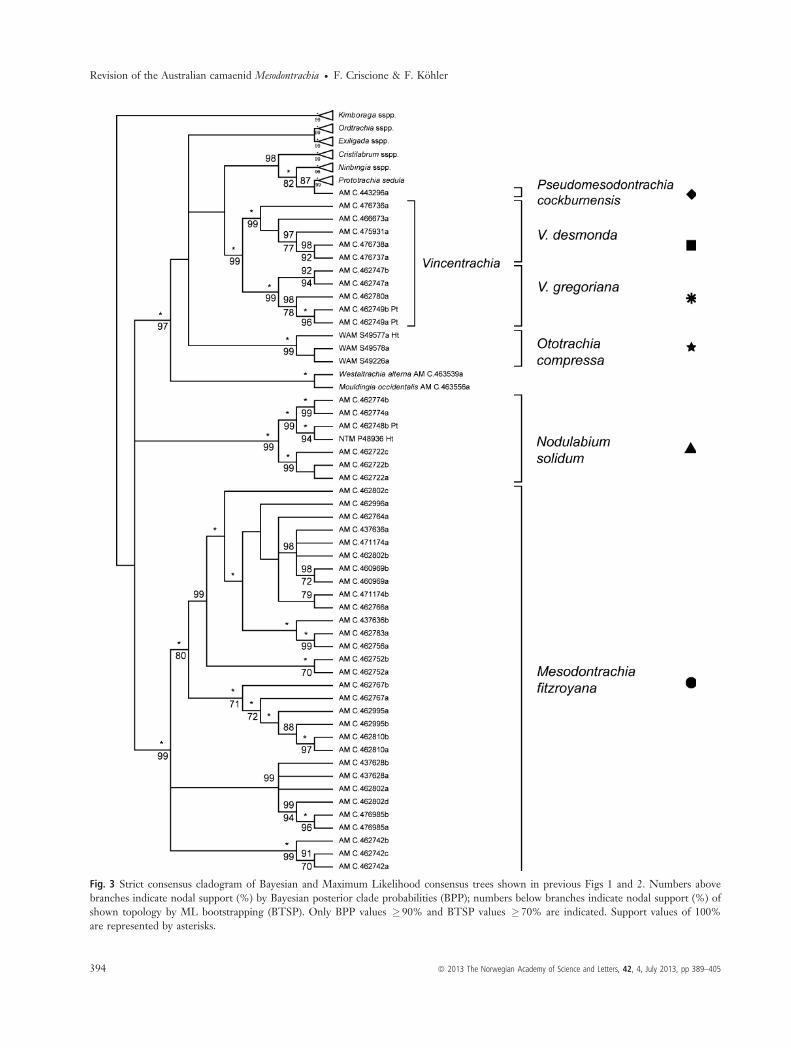
Fig. 3 Strict consensus cladogram of Bayesian and Maximum Likelihood consensus trees shown in previous Figs 1 and 2. Numbers abovebranches indicate nodal support (%) by Bayesian posterior clade probabilities (BPP); numbers below branches indicate nodal support (%) ofshown topology by ML bootstrapping (BTSP). Only BPP values � 90% and BTSP values � 70% are indicated. Support values of 100%are represented by asterisks.
394 ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405
Revision of the Australian camaenid Mesodontrachia � F. Criscione & F. K€ohler
penial complex were found to vary considerably withinsome clades. In particular, in the clade ‘Mesodontrachia’revealed a large deal of variation mirroring considerablegenetic differentiation. However, qualitative differencesbetween clades (absence/presence of features) were consis-tent.Clades varied to some extent with respect to shell
dimensions (Table 3). In a scatter plot of H/D shells of‘Ototrachia’ could be visually isolated from all other taxaindicating substantial differences in shell proportions(Fig. 5). In addition, shells of ‘Vincentrachia gregoriana’were mostly located in the lower left side of the plot(Fig. 5A) accounting for narrower and less-elevated shells.Its confidence ellipse overlapped only partly with those ofother species from the same geographic area. By contrast,the confidence ellipses of all remaining species overlappedlargely indicating that these groups had highly similar shellproportions. These species also revealed wider variation inshell dimensions than ‘Ototrachia compressa’ and ‘Vincentra-chia gregoriana’. Overall, species were found to vary littlewith respect to whorl numbers (Table 3). The averagenumbers of whorls was highest in Mesodontrachia fitzroyanaand ‘Pseudomesodontrachia cockburnensis’.
DiscussionDelimitation of speciesOur operational criterion of species delimitation is todetermine phenotypic and/or genotypic clusters (Sites &
Fig. 4 Distribution map showingoccurrences of studied samples in theVictoria River District (VRD) of theNorthern Territory and the East Kimberly(EK) in Western Australia.
Table 1 Genetic differentiation among clades by means ofp-distances
mes nod pse vin oto
mes16S 0.105COI 0.098
nod16S 0.227 0.108COI 0.172 0.086
pse16S 0.246 0.242 0.038COI 0.187 0.164 0.042
vin16S 0.266 0.261 0.169 0.039COI 0.184 0.165 0.130 0.050
oto16S 0.257 0.255 0.226 0.225 0.015COI 0.195 0.183 0.195 0.188 0.020
min max mean
16SWithin 0.015 0.108 0.061Between 0.169 0.266 0.237
COIWithin 0.020 0.098 0.059Between 0.130 0.195 0.176
mes, Mesodontrachia; nod, Nodulabium; oto, Ototrachia; pse, Pseudomesodontra-chia; vin, Vincentrachia.Bold numbers: distances within clade; rows on top: 16S distances; rows below: COIdistances; Inset: minimum, maximum and average p-distances between and within.
ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405 395
F. Criscione & F. K€ohler � Revision of the Australian camaenid Mesodontrachia
Marshall 2004). Specifically, we used a stepwise evaluationof the mitochondrial and morphological variation (‘recipro-cal corroboration’) to recognize distinct species by employ-ing the following criteria: (i) mtDNA sequences formclades, (ii) these clades are well differentiated from othersuch clades, (iii) the morphology differs between sisterclades in at least one characteristic, (iv) no intermediatemorphs exist. By combining the observations on shell andpenial morphology with distributional and phylogeneticdata, we delineated six morphologically and phylogeneti-cally distinct groups, which are considered to represent dis-tinct species. In addition to the three already describedspecies, M. fitzroyana, M. desmonda and M. cockburnensis, werecognized three new species, which are formally describedin the taxonomic descriptions at the end of this manuscript:Nodulabium solidum n. sp., Ototrachia compressa n. sp., Vin-centrachia gregoriana n. sp. These species have largely allo-patric distributions (Fig. 4).Most of the species listed above had shells of similar
shape, size, sculpture and colour. Only one species (O. com-pressa) could be readily distinguished from all others by sig-nificantly different shell dimensions (Fig. 5). Theremaining species had widely overlapping morphospaces interms of shell shape and size. All species had smooth shells(absence of sculpture visible with the naked eye) but differ-ences in microscopic sculpture were observed (Figs 6 and7). Axial growth lines in both protoconch and teleoconchwere present in all species. Vincentrachia desmonda, M. fitz-royana and P. cockburnensis had irregular pustules as anadditional element (Solem 1985: figs 87–88, 1988: pl. 2a-b).These covered the entire shell surface in V. desmonda butdid not reach the shell base in M. fitzroyana and P. cock-burnensis. By contrast, N. solidum and O. compressa had nomicroscopic pustules. Vincentrachia gregoriana had thesmallest shell of all species in the VRD, and in the Kim-berley, O. compressa had wider shells with fewer whorlsTa
ble2Key
anatom
ical
characters
ofspeciesne
wly
delim
itedandne
wly
describedhe
rein
Feature
Mesodontra
chia
fitzroyana
Nodulabium
solidum
Vincentra
chia
desm
onda
Vincentra
chia
gregoriana
Ototra
chia
Pseudomesodontra
chia
Aperturallip
reflection
Narrow
Moderate
Narrow
Very
narro
wNarrow
Narrow
Basallip
node
Absent
Present
Absent
Absent
Absent
Absent
Umbilicus
Partlyconcealed
Almostconcealed
Almostnotconcealed
Almostnotconcealed
Partlyconcealed
Partlyconcealed
Protoconch
microsculpture
Growth
lines
andpustules
Growth
lines
Growth
lines
andpustules
Growth
lines
Growth
lines
Growth
lines
andpustules
Teleoconch
microsculpture
Growth
lines
andpustules
interru
ptingat
base
Growth
lines
Growth
lines
andpustules
Growth
lines
Growth
lines
Growth
lines
andpustules
interru
ptingat
base
Teleoconch
sculpture
Absent
Absent
Absent
Absent
Absent
Absent
Shellcolour
Lightbrow
nto
horn
fading
tobase
Colour
light
brow
n,darker
alongsuture,
fading
towhitishtowards
base,lip
reflectionwhitish
Brow
non
two-thirdsof
thespire,w
hitishat
base
andon
lipreflection.
Palehorn,d
arkeralong
thesuture,fadingto
whitishtowards
base,
lipreflectionwhite.
Lightbrow
n,fading
toyellowish
towards
base,lip
reflection
yellowish
.
Lightbrow
nfading
towards
base,
brow
nperipheralb
and,
liphorn
Bursacopulatrix
Bulbose
Bulbose
Bulbose
Bulbose
Bulbose
Bulbose
Epiphallus
Absent
Absent
Present
Present
Present
Present
Innerpenial
wall
Complex
andvariablewith
severalirre
gularpilasters
With
severalirre
gular
corru
gatedpilasters
With
severalp
ilasters
supportingregular
tubercles
With
severalp
ilasters
supportingregular
tubercles
With
severalw
ide
irregular
pilasters
With
few
widecorru
gatedpilasters Table 3 Shell dimensions (mm) and whorl counts of taxa recogni-
sed in the present work
Taxon N H D W
Mesodontrachiafitzroyana
55 10.3–14.6 18.3–23.3 4.5–5.412.5 � 1.0 20.8 � 1.0 4.9 � 0.2
Vincentrachiadesmonda
28 10.1–13.7 17.3–22.7 4.2–5.111.6 � 0.8 19.5 � 1.2 4.5 � 0.2
Vincentrachiagregoriana
49 9.9–12.9 15.3–19.9 4.1–4.711.0 � 0.7 17.9 � 1.0 4.4 � 0.2
Nodulabium solidum 59 10.4–15.6 18.4–24.1 4.1–5.113.0 � 1.2 21.7 � 1.4 4.6 � 0.3
Ototrachia compressa 12 10.6–13.7 22.9–26.2 4.0–4.511.5 � 0.8 23.9 � 1.0 4.1 � 0.1
Pseudomesodontrachiacockburnensis
12 10.8–12.8 19.3–21.2 4.7–5.211.6 � 0.6 20.3 � 0.8 4.9 � 0.1
396 ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405
Revision of the Australian camaenid Mesodontrachia � F. Criscione & F. K€ohler

than P. cockburnensis (Figs 5B and S1B). Shell colour wasgenerally similar, ranging from pale horn to light brown(Fig. 6). The colour was always more intense on the upperpart of whorls and faded towards the shell base. An indis-tinct brown peripheral band was found in P. cockburnensisonly (Fig. 6F).Penial anatomy has been shown to be an informative
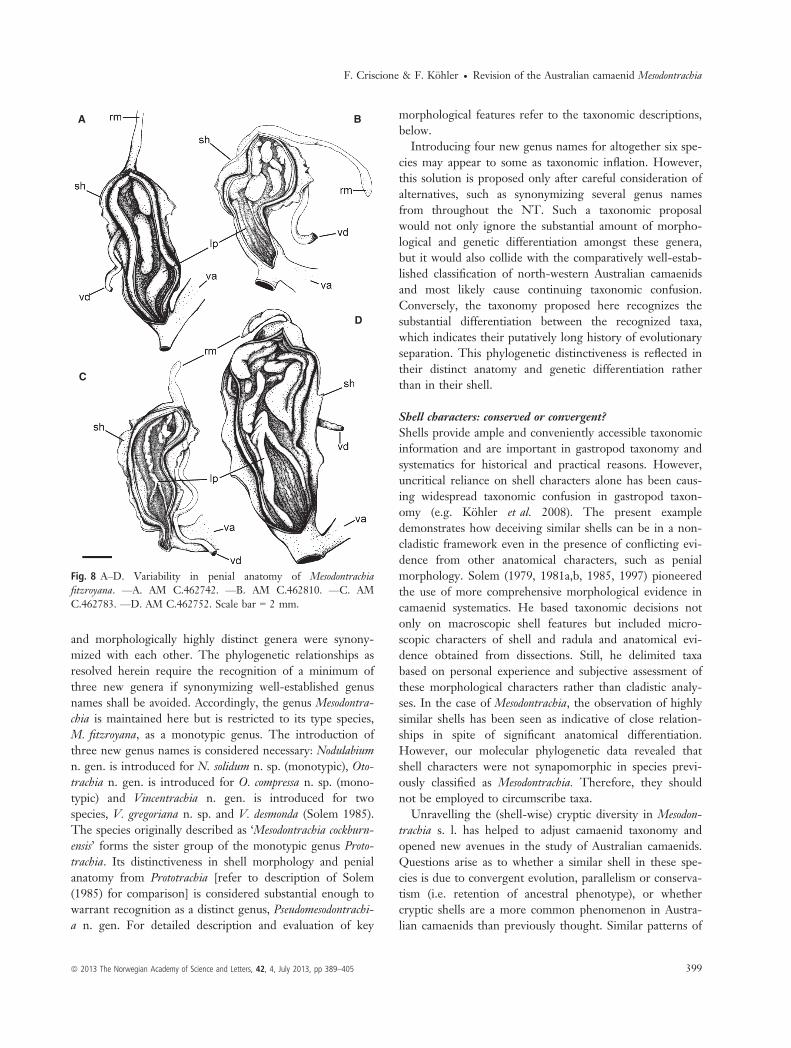
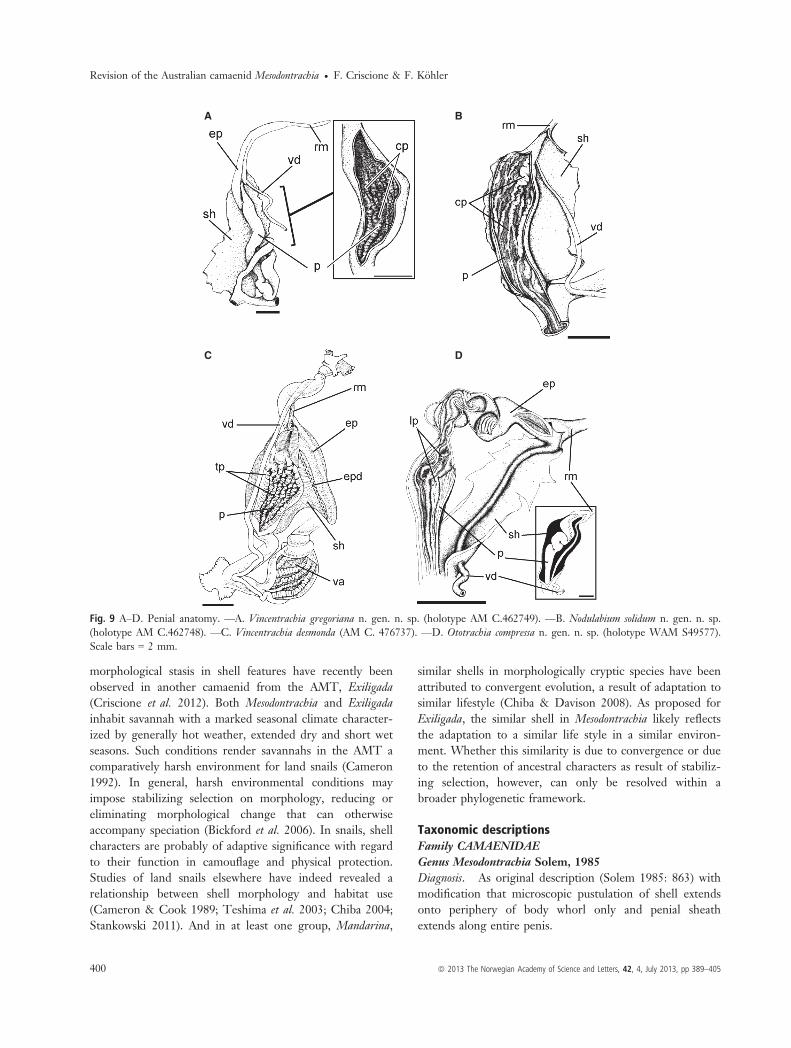
source of taxonomic information in Australian camaenidseven when shells provide little taxonomic information(e.g. K€ohler 2011a,b; Criscione & K€ohler 2013). In con-cordance with this general finding, species studied hererevealed substantial differences in their penial anatomy(Figs 8 and 9). An epiphallus was absent in two species(M. fitzroyana [Fig. 8], N. solidum [Fig. 9D]) but presentin all others. Where present, the epiphallus differed indevelopment, length, interior wall structure and withrespect to its connection to the penis with each speciesrevealing peculiar features (Fig. 9; Solem 1988: fig. 34).
The penis was coiled within the penial sheath in bothspecies of Vincentrachia, O. compressa and P. cockburnensis,while all other species had an uncoiled penis. In addition,the species had vastly different sculptures of the innerpenial wall (for detailed comparisons refer to taxo-nomic descriptions, below) apart from V. desmonda andV. gregoriana, which had a similar penial wall sculpture(Fig. 9A,C).The amounts of genetic differentiation of all six species
recognized here (Table 2) were equal to or even largerthan in most other camaenids from north-western Austra-lia. Previous studies of north-western Australian camaenidsfound that intraspecific distances did generally not exceed3% in Exiligada (Criscione et al. 2012), 4% in Rhagada(Johnson et al. 2012) and 6% in Amplirhagada (K€ohler &Johnson 2012) with no or little overlap with interspecificdistances. These values are also comparable with <4%intraspecific divergence in a wide range of stylommatophorans
A B
Fig. 5 A–B. H/D scatter plots with 95%confidence ellipses. —A. Mesodontrachiafitzroyana, Nodulabium solidum gen. n. sp.n. and Vincentrachia gregoriana gen. n., sp.n. —B. Ototrachia compressa gen. n., sp. n.,Vincentrachia desmonda andPseudomesodontrachia cockburnensis.
A
B
C
D
E
F
Fig. 6 A–F. Shells of species studied herein. —A. Mesodontrachia fitzroyana (AM C.462764). —B. Vincentrachia gregoriana n. gen. n. sp.(paratype AM C.462749). —C. Nodulabium solidum n. gen. n. sp. (paratype AM C.462774). —D. Ototrachia compressa n. gen. n. sp. (holotypeWAM S49577). —E. Vincentrachia desmonda (AM C. 476737). —F. Pseudomesodontrachia cockburnensis (AM C. 443296). Scale bar = 1 cm.
ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405 397
F. Criscione & F. K€ohler � Revision of the Australian camaenid Mesodontrachia

(Davison et al. 2009). Two species delimited here, Mes-odontrachia fitzroyana and Nodulabium solidum n. sp.,revealed considerable intraspecific genetic structuring andextreme intraspecific divergence with p-distances amount-ing up to 10%. However, for two reasons we do not con-sider this considerable genetic variability as indicative forthe existence of further cryptic species. Firstly, the mor-phological variation within clades was found to be insignif-icant in both shell and anatomy. Where present (Fig. 8),anatomical differences were small and the variation wasnot consistent with genetic structuring (i.e. large geneticdistances were not correlated with large anatomical differ-ences). Secondly, one population of M. fitzroyana con-tained haplotypes from four different lineages. The morecomprehensive sampling of haplotypes in M. fitzroyanasuggests that populations may reveal considerable geneticstructuring despite ongoing gene flow. Resolving the phy-logeographic patterns within species completely may,therefore, require a more extensive sampling than could beachieved here in most cases. Consequently, we preferred aconservative approach in delimiting species that requiredthe presence of concurrent patterns of genetic and mor-phological differentiation.
Genus-level taxonomyThree species have previously been assigned to the genusMesodontrachia in morphology-based taxonomic accounts(Solem 1985, 1988). This taxonomic treatment reflectedthe conserved shell morphology but ignored the substantialdifferentiation in the penial anatomy. However, no attempthas previously been made to test the genus-level taxonomywithin a cladistic or phylogenetic framework. Contrary tothe earlier taxonomic treatment, the present work con-firmed that a rather homogenous shell in altogether sixspecies is of little taxonomic utility. Other than in theirshell, these species varied considerably in their penial anat-omy. The variation in penial anatomy and the considerablegenetic differentiation are indicative of shell-wise cryptictaxonomic diversity in Mesodontrachia sensu lato. However,the observed anatomical differences per se do not provideunambiguous means for the delimitation of genera as acomparable range of anatomical variability in penial fea-tures has been documented in other species-rich camaenidgenera, such as Amplirhagada (K€ohler 2010, 2011c) andExiligada (Criscione et al. 2012), for instance.To maintain monophyletic taxa, a split-up of Mesodontrachi-
a s. l. is inevitable unless a large number of well established
A
C
E
B
D
F
Fig. 7 SEM micrographs of shells showingprotoconch and teleoconch microsculpture.—A–B. Vincentrachia gregoriana n. gen. n.sp. (paratype AM C.462747). —C–D.Nodulabium solidum n. gen. n. sp. (paratypeAM C.462774). —E–F. Ototrachia compressan. gen. n. sp. (paratype WAM S49101).Scale bars: A, C, E = 1 mm; B, D,F = 200/m.
398 ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405
Revision of the Australian camaenid Mesodontrachia � F. Criscione & F. K€ohler
and morphologically highly distinct genera were synony-mized with each other. The phylogenetic relationships asresolved herein require the recognition of a minimum ofthree new genera if synonymizing well-established genusnames shall be avoided. Accordingly, the genus Mesodontra-chia is maintained here but is restricted to its type species,M. fitzroyana, as a monotypic genus. The introduction ofthree new genus names is considered necessary: Nodulabiumn. gen. is introduced for N. solidum n. sp. (monotypic), Oto-trachia n. gen. is introduced for O. compressa n. sp. (mono-typic) and Vincentrachia n. gen. is introduced for twospecies, V. gregoriana n. sp. and V. desmonda (Solem 1985).The species originally described as ‘Mesodontrachia cockburn-ensis’ forms the sister group of the monotypic genus Proto-trachia. Its distinctiveness in shell morphology and penialanatomy from Prototrachia [refer to description of Solem(1985) for comparison] is considered substantial enough towarrant recognition as a distinct genus, Pseudomesodontrachi-a n. gen. For detailed description and evaluation of key
morphological features refer to the taxonomic descriptions,below.Introducing four new genus names for altogether six spe-
cies may appear to some as taxonomic inflation. However,this solution is proposed only after careful consideration ofalternatives, such as synonymizing several genus namesfrom throughout the NT. Such a taxonomic proposalwould not only ignore the substantial amount of morpho-logical and genetic differentiation amongst these genera,but it would also collide with the comparatively well-estab-lished classification of north-western Australian camaenidsand most likely cause continuing taxonomic confusion.Conversely, the taxonomy proposed here recognizes thesubstantial differentiation between the recognized taxa,which indicates their putatively long history of evolutionaryseparation. This phylogenetic distinctiveness is reflected intheir distinct anatomy and genetic differentiation ratherthan in their shell.
Shell characters: conserved or convergent?Shells provide ample and conveniently accessible taxonomicinformation and are important in gastropod taxonomy andsystematics for historical and practical reasons. However,uncritical reliance on shell characters alone has been caus-ing widespread taxonomic confusion in gastropod taxon-omy (e.g. K€ohler et al. 2008). The present exampledemonstrates how deceiving similar shells can be in a non-cladistic framework even in the presence of conflicting evi-dence from other anatomical characters, such as penialmorphology. Solem (1979, 1981a,b, 1985, 1997) pioneeredthe use of more comprehensive morphological evidence incamaenid systematics. He based taxonomic decisions notonly on macroscopic shell features but included micro-scopic characters of shell and radula and anatomical evi-dence obtained from dissections. Still, he delimited taxabased on personal experience and subjective assessment ofthese morphological characters rather than cladistic analy-ses. In the case of Mesodontrachia, the observation of highlysimilar shells has been seen as indicative of close relation-ships in spite of significant anatomical differentiation.However, our molecular phylogenetic data revealed thatshell characters were not synapomorphic in species previ-ously classified as Mesodontrachia. Therefore, they shouldnot be employed to circumscribe taxa.Unravelling the (shell-wise) cryptic diversity in Mesodon-
trachia s. l. has helped to adjust camaenid taxonomy andopened new avenues in the study of Australian camaenids.Questions arise as to whether a similar shell in these spe-cies is due to convergent evolution, parallelism or conserva-tism (i.e. retention of ancestral phenotype), or whethercryptic shells are a more common phenomenon in Austra-lian camaenids than previously thought. Similar patterns of
A B
C
D
Fig. 8 A–D. Variability in penial anatomy of Mesodontrachiafitzroyana. —A. AM C.462742. —B. AM C.462810. —C. AMC.462783. —D. AM C.462752. Scale bar = 2 mm.
ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405 399
F. Criscione & F. K€ohler � Revision of the Australian camaenid Mesodontrachia
morphological stasis in shell features have recently beenobserved in another camaenid from the AMT, Exiligada(Criscione et al. 2012). Both Mesodontrachia and Exiligadainhabit savannah with a marked seasonal climate character-ized by generally hot weather, extended dry and short wetseasons. Such conditions render savannahs in the AMT acomparatively harsh environment for land snails (Cameron1992). In general, harsh environmental conditions mayimpose stabilizing selection on morphology, reducing oreliminating morphological change that can otherwiseaccompany speciation (Bickford et al. 2006). In snails, shellcharacters are probably of adaptive significance with regardto their function in camouflage and physical protection.Studies of land snails elsewhere have indeed revealed arelationship between shell morphology and habitat use(Cameron & Cook 1989; Teshima et al. 2003; Chiba 2004;Stankowski 2011). And in at least one group, Mandarina,
similar shells in morphologically cryptic species have beenattributed to convergent evolution, a result of adaptation tosimilar lifestyle (Chiba & Davison 2008). As proposed forExiligada, the similar shell in Mesodontrachia likely reflectsthe adaptation to a similar life style in a similar environ-ment. Whether this similarity is due to convergence or dueto the retention of ancestral characters as result of stabiliz-ing selection, however, can only be resolved within abroader phylogenetic framework.
Taxonomic descriptionsFamily CAMAENIDAEGenus Mesodontrachia Solem, 1985Diagnosis. As original description (Solem 1985: 863) withmodification that microscopic pustulation of shell extendsonto periphery of body whorl only and penial sheathextends along entire penis.
A B
C D
Fig. 9 A–D. Penial anatomy. —A. Vincentrachia gregoriana n. gen. n. sp. (holotype AM C.462749). —B. Nodulabium solidum n. gen. n. sp.(holotype AM C.462748). —C. Vincentrachia desmonda (AM C. 476737). —D. Ototrachia compressa n. gen. n. sp. (holotype WAM S49577).Scale bars = 2 mm.
400 ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405
Revision of the Australian camaenid Mesodontrachia � F. Criscione & F. K€ohler

Taxonomic remarks. Originally described to contain twospecies, Mesodontrachia fitzroyana (type species) and M. des-monda; a third species, M. cockburnensis, was subsequentlydescribed (Solem 1988). These three species do not form amonophyletic group. Therefore, M. desmonda and M. cock-burnensis are here removed from the genus, which becomesmonotypic. The updated generic diagnosis reflects thesetaxonomic changes.
Comparative remarks. Shells of M. fitzroyana differ fromVincentrachia by wider columellar reflection, from Turgeni-tubulus, Cristilabrum and Exiligada by less-developed lipreflection, from Nodulabium by lack of basal nodule, fromOtotrachia and Pseudomesodontrachia by more elevated spire.Microsculpture similar to Vincentrachia, Pseudomesodontrachi-a and some Exiligada species. Teleoconch sculpture differsfrom Prototrachia, Ordtrachia by lack of prominent axialribs. Colour pattern very similar to Nodulabium. Generatreated herein differ by presence of epiphallus (Vincentra-chia, Ototrachia), coiled penis (Vincentrachia) and distinct pe-nial wall sculpture (Vincentrachia, Ototrachia). Developmentof penial pilasters variable (Fig. 8). Genital anatomy mostsimilar to Nodulabium but inner penial wall with finer andmore numerous longitudinal pilasters.
Pseudomesodontrachia n. gen. (Fig. 6F; Solem, 1988: pl.2a,b, figs 29–34)Type species. Mesodontrachia cockburnensis Solem, 1988Material examined. See Table S1.
Etymology. For being similar to Mesodontrachia, derivedfrom ‘pseud�es’ (Greek = false).
Description.Shell. Moderately large, subglobose, periphery rounded,
spire moderately elevated, moderately thin; umbilicus wideopen, partly concealed by columellar reflection; protoconchand teleoconch with moderately fine radial growth lines andfine pustulations (Fig. 6E). Last whorl wide in cross-section,periphery rounded. Apertural lip reflected, slightlyexpanded, parietal wall very thin. Shell colour brown onupper third of spire (some specimens with a darker subsutur-al area resembling a band), whitish at base and on lip reflec-tion, with a brownish-reddish band on whorl periphery.
Genital anatomy. Refer to description of M. cockburnen-sis by Solem (1988: 39) (Fig. 9C).
Comparative remarks. Shell differing from Nodulabiummainly by lack of basal nodule. Shell sculpture similar toMesodontrachia, Ningbingia, Turgenitubulus and some Cristi-labrum species differing from V. gregoriana, Ototrachia,
Nodulabium and other Cristilabrum species by presence ofpustules, from V. desmonda by pustules not continuing ontoshell base. Shell colour similar to some banded Exiligadaspecies but differing from other taxa in the area by distinc-tive sharp transition (brown-whitish) between top of shelland its base.
Vincentrachia n. gen. (Figs 6D,E, 7A,B and 8A,C)Type species. Mesondontrachia desmonda Solem, 1985Material examined. See Table S1.
Etymology. In honour of Vince Kessner in recognition ofhis lasting achievements as ardent snail collector and fieldworker, derived from his first name and the word ‘trache�ıa’(Greek = trachea) that indicates the similarity in shell mor-phology with other genera with the same suffix.
Description.Shell. Relatively small to average, subglobose, periphery
rounded, spire moderately elevated to elevated, thin tomoderately thin; umbilicus wide open, partially concealedby columellar reflection; protoconch and teleoconch withfine to moderately fine radial growth lines and with or with-out fine pustulations (Figs 6E,D and 7A,B). Subsuturaltubercles may be present (V. gregoriana). Last whorl moder-ately wide in cross-section, periphery slightly shouldered.Apertural lip reflected, slightly to moderately expanded,parietal wall very thin. Shell colour horn or brown on uppertwo-thirds of spire, lighter at base and on lip reflection.
Genital anatomy. Penial sheath well-developed, extend-ing entire length (Figs. 9A,C). Epiphallus forming a tubesurrounded by thick layers of muscle, well separated frompenis and connected to penis through narrow zigzag duct; aslong as penis. Penial retractor muscle inserting on vas-epi-phallus junction, with fibres continuing along epiphallusattaching to penial apex. Vas deferens entering sheath api-cally. Penis coiled or uncoiled inside sheath, with thick wall,inner penial wall structure consisting of several wide longitu-dinal pilasters supporting regular tubercles. Pilaster runningalong entire wall length. Vagina as long as penis, two timeslonger than free oviduct. Bursa copulatrix simple, as long asfree oviduct, reaching posterior end of spermoviduct.
Vincentrachia desmonda (Solem, 1985)Mesodontrachia desmonda – Solem 1985: 867–870; pl. 87a–f;figs 218a–c, 219a–e.Material examined. See Table S1
Diagnosis.Shell. Relatively small to average, subglobose, periphery
rounded, spire moderately elevated, moderately thin;
ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405 401
F. Criscione & F. K€ohler � Revision of the Australian camaenid Mesodontrachia

umbilicus wide open, partially concealed by columellarreflection; protoconch and teleoconch with moderately fineradial growth lines and fine pustulations (Fig. 6E). Lastwhorl moderately wide in cross-section, periphery slightlyshouldered. Apertural lip reflected, slightly expanded, pari-etal wall very thin. Shell colour brown on upper two-thirdsof spire, whitish at base and on lip reflection.
Genital anatomy. Penial sheath well-developed, extend-ing entire length (Fig. 9C). Epiphallus forming a tube sur-rounded by thick layers of muscle, well separated frompenis and connected to penis through narrow zigzag duct;as long as penis. Penial retractor muscle inserting on vas-epiphallus junction, with fibres continuing along epiphallusattaching to penial apex. Vas deferens entering sheath api-cally. Penis uncoiled, with thick wall, inner penial wallstructure consisting of several wide longitudinal pilasterssupporting regular tubercles. Pilaster running along entirewall length. Vagina as long as penis, two times longer thanfree oviduct. Bursa copulatrix simple, as long as free ovi-duct, reaching posterior end of spermoviduct.
Comparative remarks. Shell size and shape similar to Pseu-domesodontrachia and some Exiligada species, differing fromthose taxa by always reduced columellar and lip reflection.Shell differing from V. gregoriana by more developed lipreflection and from Nodulabium mainly by lack of basalnodule. Protoconch and teleoconch microsculpture differ-ing from Pseudomesodontrachia, Ototrachia, Prototrachia, someExiligada and Ordtrachia species; similar to remaining taxain the area and differing by those by pustules reachingbase; teleoconch sculpture similar to most taxa in samegeographic area and differing from Prototrachia, Ordtrachia,Turgenitubulus and Cristilabrum by lack of axial ribs. Colourdarker than other taxa in the area; colour pattern distinc-tive by showing sharp transition (brown-whitish) betweenrest of shell and its base. Genitalia differing from Mesodon-trachia, Nodulabium, Ototrachia, Ningbingia, Turgenitubulusand Cristilabrum by presence of epiphallus. An epiphallus-penis connection was not visible in the holotype (dashedline in Fig. 9C), probably an old specimen, exclusively act-ing as a female, with a non-functional male reproductivesystem. However, a thin duct, connecting epiphallus andpenis, was consistently observed in dissections of youngerspecimens (not figured). Details of the reproductive anat-omy of this species were previously based on dissection ofa subadult specimen (Solem 1985; fig. 219), not displayinga mature reproductive system.
Vincentrachia gregoriana n. spHolotype. AUSTRALIA, NT, VRD, Gregory NP,
7.7 km WNW of Bullita O/S, 16° 05′ 43″ S, 130° 21′ 42″
E, limestone karst, pockets of vine thicket, in talus (V.Kessner, S. Cordell, 13 Jul 2008), dissected specimen(NTM P48936).
Paratypes. Same as holotype, 34 dry, 6 wet (AMC.462749).
Additional material. See Table S1.
Etymology. For Gregory National Park, derived from‘gregorianus’ (Latin), adjective of feminine gender.
Description.Shell. Relatively small to average, subglobose, periphery
rounded, spire elevated, thin; umbilicus moderately wide,partially concealed by columellar reflection; protoconchwith fine radial growth lines, teleoconch with fine radialgrowth lines and subsutural tubercles (Figs 6D and 7A,B).Last whorl moderately wide in cross-section, peripheryshouldered. Apertural lip reflected, slightly expanded, pari-etal wall very thin. Shell colour pale horn, darker along thesuture, fading to whitish towards base, lip reflection white.
Genital anatomy. Like type species but penis about twotimes longer, coiled within sheath (Fig. 9A).
Comparative remarks. Smallest shell among species studiedherein, shell shape similar to V. desmonda, Mesodontrachia,and Exiligada, differing from those by narrower lip reflec-tion. Shell differing from Nodulabium by lack of basal nod-ule. Protoconch and teleoconch microsculpture similar toOtotrachia and some Exiligada species; teleoconch sculpturesimilar to most taxa in same geographic area and differingfrom Prototrachia, Ordtrachia, Turgenitubulus and Cristila-brum by lack of axial ribs. Colour pattern similar to Nodula-bium from which it can be differentiated by lighterbackground colour. Genitalia differing from Mesodontrachia,Nodulabium, Ototrachia, Ningbingia, Turgenitubulus and Cris-tilabrum by presence of epiphallus.
Nodulabium gen. n. (Figs 6B, 7C,D and 9B)Type species. Nodulabium solidum n. sp.
Etymology. For the basal nodule of the shell lip of thetype species, derived from ‘nodulus’ (Latin = nodule) and‘labium’ (Latin = lip).
Description. As for type species (see below).
Nodulabium solidum n. spHolotype. AUSTRALIA, NT, VRD, Gregory NP,
13 km WSW of Bullita O/S, 16° 08′ 49″ S, 130° 18′ 45″ E,
402 ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405
Revision of the Australian camaenid Mesodontrachia � F. Criscione & F. K€ohler

limestone karst, vine thicket patches, in talus (V. Kessner,14 Jul 2008), dissected specimen (NTM P48937).
Paratypes. Same as holotype, 18 dry, 1 wet (AMC.462748).
Additional material. See Table S1.
Etymology. In reference to the particularly thick shell ofthe type species which gives it a solid appearance, derivedfrom ‘solidum’ (Latin = solid), adjective in the neutral gen-der.
Description.Shell. Moderately large, subglobose, periphery
rounded, spire moderately elevated, thick; umbilicus almostcompletely concealed by columellar reflection; protoconchwith moderately fine radial growth lines; teleoconch withmoderately fine radial growth lines and fine micro-pustula-tions (Figs 6B and 7C,D). Last whorl wide in cross-section,periphery uniformly rounded. Apertural lip reflected, mod-erately expanded, parietal wall thick; with a weak basalnodule. Shell colour light brown, darker along suture, fad-ing to whitish towards base, lip reflection whitish.
Genital anatomy. Penial sheath well-developed, extend-ing entire length. Epiphallus absent (Fig. 9B). Penial retrac-tor muscle inserting directly onto apical end of penis. Vasdeferens entering penial sheath subapically, continuing api-cally within sheath until reflexing into penis. Penis uncoiled,with moderately thick wall; inner penial wall structure con-sisting of several corrugated, wide longitudinal pilasters.Pilaster running along entire wall length. Vagina lengthequivalent of two-thirds of penis and two times longer thanfree oviduct; bursa copulatrix simple, usually as long as freeoviduct, reaching anterior end of spermoviduct.
Comparative remarks. Thickest shell among taxa studiedherein and (along with Mesodontrachia) highest spired, dif-fering from them by presence of basal nodule and by widercolumellar and lip reflections, the former less pronouncedthan in Turgenitubulus and Cristilabrum, the latter narrowerthan in Prototrachia, Ningbingia, Turgenitubulus and Cristila-brum.Protoconch microsculpture differing from Pseudomesodon-
trachia, Ototrachia, Prototrachia and some Exiligada and Ord-trachia species by presence of pustules; teleoconchsculpture similar to most taxa in same geographic area anddiffering from Prototrachia, Ordtrachia, Turgenitubulus andCristilabrum by lack of axial ribs. Colour pattern similar toPseudomesodontrachia from which it can be differentiated bydarker background colour. Genitalia differing from
Vincentrachia, Ototrachia, Pseudomesodontrachia, Exiligada,Prototrachia, Ordtrachia by absence of epiphallus.
Ototrachia n. gen. (Figs 6C, 7E,F and 9D)Type species. Ototrachia compressa n. sp.
Etymology. For the ear-shaped shell of the type species,derived from ‘ot�os’ (Greek = ear) and ‘trache�ıa’(Greek = trachea; to indicate the similarity in shell mor-phology with other genera with the same suffix).
Description. As for type species (see below).
Ototrachia compressa n. spHolotype. AUSTRALIA, WA, EK, Wyndham, Two
Mile Valley, 15° 28′ 01″ S, 128° 08′ 10″ E, vine thicket onsteep slopes, scree, under pile of rocks (V. Kessner, 8 Jan2009), dissected specimen (WAM S49577).
Paratypes. Same data as holotype, 3 dry (WAMS49686).
Additional material. See Table S1.
Etymology. For the weakly elevated shell spire, derivedfrom ‘compressus’ (Latin = compressed), adjective in femi-nine gender.
Description.Shell. Large, subglobose, periphery rounded, spire
weakly elevated, thick; umbilicus wide only slightly con-cealed by columellar reflection; protoconch and teleoconchwith moderately fine radial growth lines (Figs 6C and 7E,F). Last whorl considerably wide in cross-section, peripheryslightly shouldered. Apertural lip reflected, moderatelyexpanded, parietal wall thin. Shell colour light brown, fad-ing to yellowish towards base, lip reflection yellowish.
Genital anatomy. Penial sheath well-developed, extend-ing entire length (Fig. 9D). Epiphallus consisting of a nar-row zigzag duct surrounded by thick walls, connected topenis with a very narrow duct; almost three times shorterthan penis. Penial retractor muscle inserting directly at thevas deferens-epiphallus junction. Penis coiled, with moder-ately thick wall; inner penial wall structure consisting ofseveral wide irregular longitudinal pilasters. Pilasters run-ning along entire wall length. Vagina half of penis and twotimes longer than free oviduct; bursa copulatrix relativelysimple, as long as free oviduct, reaching anterior end ofspermoviduct, with well-differentiated end. Albumen glandrelatively small. Talon embedded in albumen gland withinproximal portion. Hermaphroditic duct tightly undulating.
ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405 403
F. Criscione & F. K€ohler � Revision of the Australian camaenid Mesodontrachia

Comparative remarks. Largest shell of taxa studied here;shape distinctive by large body whorl in cross- section; sim-ilar to some Exiligada species, but differing by more devel-oped columellar and lip reflection, from the latter ones byshell colour. Shell differing from Nodulabium mainly bylack of basal nodule. Protoconch and teleoconch micro-sculpture similar to Pseudomesodontrachia and some Exiligadaspecies; teleoconch sculpture similar to most taxa in samegeographic area and differing from Prototrachia, Ordtrachia,Turgenitubulus and Cristilabrum by lack of axial ribs. Coloursimilar to Mesodontrachia but differing by lip reflectionbeing yellowish. Genitalia differing from Mesodontrachia,Nodulabium, Ototrachia, Ningbingia, Turgenitubulus and Cris-tilabrum by presence of epiphallus.
AcknowledgementsThis work has been made possible through financial sup-port from the Australian Government (ABRS grantsRF210-05 and RF211-10 to FK). Vince Kessner (AdelaideRiver) and Michael Braby (Darwin) prepared and con-ducted the field work, thereby providing the foundationsfor this work. Their efforts are most thankfully acknowl-edged. Special thanks are further due to Morgan Laudine,Martin P€uschel and Michael Shea (Sydney) for performingdissections and producing illustrations, and to Sue Lindsay(Sydney) for conducting SEM work. Thanks are due totwo reviewers; their constructive comments helped toimprove the manuscript.
ReferencesBickford, D. P., Lohman, D. J., Sodhi, N. S., Ng, P. K. L., Meier,R., Winker, K., Ingram, K. K. & Das, I. (2006). Cryptic speciesas a window on diversity and conservation. Trends in Ecology andEvolution, 22, 148–155.
Cameron, R. A. D. (1992). Land snail faunas of the Napier andOscar Ranges, Western Australia; diversity, distribution and spe-ciation. Biological Journal of the Linnean Society, 45, 271–286.
Cameron, R. A. D. & Cook, L. M. (1989). Shell size and shape inMadeiran land snails: do niches remain unfilled? Biological Jour-nal of the Linnean Society, 36, 79–96.
Chiba, S. (1999). Accelerated evolution of land snails Mandarina inthe oceanic Bonin Islands: evidence from mitochondrial DNAsequences. Evolution, 53, 460–471.
Chiba, S. (2004). Ecological and morphological patterns in com-munities of land snails of the genus Mandarina from the BoninIslands. Journal of Evolutionary Biology, 17, 131–143.
Chiba, S. & Davison, A. (2008). Anatomical and molecular studiesreveal several cryptic species of the endemic genus Mandarina(Pulmonata: Helicoidea) in the Ogasawara Islands. Journal ofMolluscan Studies, 74, 373–382.
Criscione, F. & K€ohler, F. (2013). Six new species of Australocosmi-ca K€ohler, 2011 from the Kimberley islands, Western Australia(Mollusca: Pulmonata: Camaenidae). Zootaxa, 3608, 101–115.
Criscione, F., Law, M. L. & K€ohler, F. (2012). Land snail diversityin the monsoon tropics of Northern Australia: revision of the
genus Exiligada Iredale, 1939 (Mollusca: Pulmonata: Camaeni-dae), with description of 13 new species. Zoological Journal of theLinnean Society, 166, 689–722.
Davison, A., Blackie, R. L. E. & Scothern, G. P. (2009). DNA bar-coding of stylommatophoran land snails: a test of existingsequences. Molecular Ecology Resources, 9, 1092–1101.
Folmer, O., Black, M., Hoeh, W., Lutz, R. & Vrijenhoek, R.(1994). DNA primers for amplification of mitochondrial cyto-chrome c oxidase subunit I from diverse metazoan invertebrates.Molecular Marine Biology and Biotechnology, 3, 294–299.
Hasegawa, M., Kishino, H. & Yano, T. (1985). Dating of thehuman-ape splitting by a molecular clock of mitochondrialDNA. Journal of Molecular Evolution, 22, 160–174.
Hugall, A. F. & Stanisic, J. (2011). Beyond the prolegomenon: amolecular phylogeny of the Australian camaenid land snail radia-tion. Zoological Journal of the Linnean Society, 161, 531–572.
Iredale, T. (1939). A review of the land Mollusca of Western Aus-tralia. Records of the Western Australian Museum and Art Gallery,2, 1–88.
Johnson, M. S., Hamilton, Z. R., Teale, R. O. Y. & Kendrick, P.G. (2012). Endemic evolutionary radiation of Rhagada land snails(Pulmonata: Camaenidae) in a continental archipelago in north-ern Western Australia. Biological Journal of the Linnean Society,106, 316–327.
K€ohler, F. (2010). Uncovering local endemism in the Kimberley,Western Australia: description of new species of the genus Am-plirhagada Iredale, 1933 (Pulmonata, Camaenidae). Records of theAustralian Museum, 62, 217–284.
K€ohler, F. (2011a). The camaenid species of the KimberleyIslands, Western Australia (Stylommatophora: Helicoidea). Mal-acologia, 54, 203–406.
K€ohler, F. (2011b). Australocosmica, a new genus of land snails fromthe Kimberley, Western Australia (Eupulmonata, Camaenidae).Malacologia, 53, 199–216.
K€ohler, F. (2011c). Descriptions of new species of the diverseand endemic land snail Amplirhagada Iredale, 1933 from rain-forest patches across the Kimberley, Western Australia (Pul-monata, Camaenidae). Records of the Australian Museum, 63,167–202.
K€ohler, F. (2012). Taxonomic revision of two endemic land snailgenera from the Top End of Northern Australia with remarkson two problematic species named by de F�erussac and Le Guil-lou (Eupulmonata, Camaenidae). Zoosystematics and Evolution, 88,53–62.
K€ohler, F. & Criscione, F. (2013). Arnhemtrachia ramingining – anew genus and species of land snail from Arnhem Land,Australia (Stylommatophora: Camaenidae). Molluscan Research,in press.
K€ohler, F. & Johnson, M. S. (2012). Species limits in molecularphylogenies: a cautionary tale from Australian land snails (Cama-enidae: Amplirhagada). Zoological Journal of the Linnean Society,165, 337–362.
K€ohler, F., Brinkmann, N. & Glaubrecht, M. (2008). Convergencecauses confusion: on the systematics of the freshwater gastropodSulcospira pisum (Brot, 1868) (Cerithioidea, Pachychilidae). Mal-acologia, 50, 331–339.
Ronquist, F. & Huelsenbeck, J. P. (2003). MRBAYES 3: Bayesianphylogenetic inference under mixed models. Bioinformatics, 19,1572–1574.
404 ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405
Revision of the Australian camaenid Mesodontrachia � F. Criscione & F. K€ohler

Sites, J. W. & Marshall, J. C. (2004). Operational criteria fordelimiting species. Annual Review of Ecology Evolution and System-atics, 35, 199–227.
Slatyer, C., Ponder, W. F., Rosauer, D. & Davis, L. (2007).Between a rock and a dry place – land snails in arid Australia. InC. Dickman, D. Lunney & S. Burgin (Eds) Animals of arid Aus-tralia: Out on their Own? (pp. 30–41). Mosman, NSW: RoyalZoological Society of New South Wales.
Solem, A. (1979). Camaenid land snails from Western and centralAustralia (Mollusca: Pulmonata: Camaenidae). I. Taxa withtrans-Australian distribution. Records of the Western AustralianMuseum, 10(Suppl), 5–142.
Solem, A. (1981a). Camaenid land snails from Western and centralAustralia (Mollusca: Pulmonata: Camaenidae). II. Taxa from theKimberley, Amplirhagada Iredale 1933. Records of the WesternAustralian Museum, 11(Suppl), 147–320.
Solem, A. (1981b). Camaenid land snails from Western and centralAustralia (Mollusca: Pulmonata: Camaenidae). III. Taxa from theNingbing Ranges and nearby areas. Records of the Western Austra-lian Museum, 11(Suppl), 321–425.
Solem, A. (1985). Camaenid land snails from Western and centralAustralia (Mollusca: Pulmonata: Camaenidae). V. RemainingKimberley genera and addenda to the Kimberley. Records of theWestern Australian Museum, 20(Suppl), 707–981.
Solem, A. (1988). New camaenid land snails from the northeastKimberley, Western Australia. Journal of the Malacological Societyof Australia, 9, 27–58.
Solem, A. (1997). Camaenid land snails from Western and centralAustralia (Mollusca: Pulmonata: Camaenidae). VII. Taxa fromDampierland trough the Nullabor. Records of the Western Austra-lian Museum, 50(Suppl), 1461–1906.
Solem, A. (1998). Family Camaenidae. In P. L. Beesley, G. J. B.Ross & A. Wells (Eds) Mollusca: The Southern Synthesis (pp.1112–1113). Melbourne: CSIRO Publishing.
Stankowski, S. (2011). Extreme, continuous variation in an islandsnail: Local diversification and association of shell form with the
current environment. Biological Journal of the Linnean Society,104, 756–769.
Sutcharit, C., Naggs, F. & Panha, S. (2007). Systematic review ofthe land snail genus Neocepolis Pilsbry, 1891 (Pulmonata: Camae-nidae) from north Vietnam. Journal of Natural History, 41, 619–631.
Tamura, K. & Nei, M. (1993). Estimation of the number of nucle-otide substitutions in the control region of mitochondrial DNAin humans and chimpanzees. Molecular Biology and Evolution, 10,512–526.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M. &Kumar, S. (2011). MEGA5: molecular evolutionary geneticsanalysis using maximum likelihood, evolutionary distance, andmaximum parsimony methods. Molecular Biology and Evolution,28, 2731–2739.
Teshima, H., Davison, A., Kuwahara, Y., Yokoyama, J., Chiba, S.,Fukuda, T., et al. (2003). The evolution of extreme shell shapevariation in the land snail Ainohelix editha: a phylogeny andhybrid zone analysis. Molecular Ecology, 12, 1869–1878.
Xia, X. & Xie, Z. (2001). DAMBE: data analysis in molecular biol-ogy and evolution. Journal of Heredity, 92, 371–373.
Xia, X., Xie, Z., Salemi, M., Chen, L. & Wang, Y. (2003). Anindex of substitution saturation and its application. MolecularPhylogenetics and Evolution, 26, 1–7.
Supporting InformationAdditional Supporting Information may be found in theonline version of this article:Table S1. Museum registration numbers, locality data andGenBank accession numbers of studied samples.Fig. S1. H/D scatter plots with 95% confidence ellipses ofall shells measured herein.Fig. S2. A–B. Frequency bar charts of whorl numbers (W).
ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, pp 389–405 405
F. Criscione & F. K€ohler � Revision of the Australian camaenid Mesodontrachia

Corrigendumdoi:10.1111/zsc.12021
The article by Criscione and K€ohler (2013) contained errors in the legend of Fig. 6. We reprint the correct figure legendbelow. The text references to Fig. 6 are generally correct apart from references to ‘Fig. 6E’ in the paragraph on Pseudomes-odontrachia, which should read ‘Fig. 6F’ instead.
Fig. 6 A–F. Shells of species studied herein. —A. Mesodontrachia fitzroyana (AM C.462764). —B. Nodulabium solidum n.gen. n. sp. (paratype AM C.462774). —C. Ototrachia compressa n. gen. n. sp. (holotype WAM S49577). —D. Vincentrachiagregoriana n. gen. n. sp. (paratype AM C.462749). —E. Vincentrachia desmonda (AM C.476737). —F. Pseudomesodontrachiacockburnensis (AM C.443296). Scale bar = 1 cm.
ReferenceCriscione, F. & K€ohler, F. (2013) Conserved shell disguises diversity in Mesodontrachia land snails from the Australian Monsoon Tropics(Gastropoda: Camaenidae). Zoologica Scripta, 42, 389–405.
440 ª 2013 The Norwegian Academy of Science and Letters, 42, 4, July 2013, p 440
Zoologica Scripta





























