Embed Size (px)
Citation preview

15
ISSN 0031-0301, Paleontological Journal, 2007, Vol. 41, No. 1, pp. 15–27. © Pleiades Publishing, Ltd., 2007.Original Russian Text © O.V. Amitrov, E.A. Zhegallo, 2007, published in Paleontologicheskii Zhurnal, 2007, No. 1, pp. 15–26.
INTRODUCTION
The gastropods studied represent a group widelydistributed in the Paleogene (especially in the Oli-gocene) of western Europe (mainly the North SeaBasin). It is also present in the south of the formerUSSR, including the Transcaspian Region, where ninedifferent varieties (at least eight species) are recognizedfrom the Upper Eocene to the Oligocene–Mioceneboundary beds.
In the papers of the 19th–early 20th century, andeven in later publications,
Scalaspira
species were usu-ally described under generic name
Fusus
. Wenz (1938–1944) believed that the valid name for this genus was
Fusinus
Rafinesque, 1810 (=
Fusus
Bruguière, 1789,non Halbling, 1779). Kautsky (1925) assigned a groupof species from the Oligocene and Miocene of Europeto the genus
Aquilofusus
. Tembrock (1964, 1965, 1968)proposed a detailed revision of all these gastropods.She concluded that the genus
Aquilofusus
should beunited with several other gastropod genera including
Scalaspira
Conrad, 1862 and
Mohnia
Friele, 1879. Pre-viously, these genera were assigned to different fami-lies. According to Wenz, the genus
Aquilofusus
belongsto Fasciolariidae, subfamily Fusinidae, while
Sca-laspira
belongs to Muricidae (at that time only the typespecies of this genus was recorded from the Miocene ofNorth America), whereas
Mohnia
belonged to Buc-cinidae (it was thought that this genus was representedby several extant species from northern seas). Tem-brock suggested that the oldest available name
Sca-laspira
should be used for this united genus andassigned this genus to Buccinidae based on its proto-conch and on the structure of the radula of extant spe-cies previously assigned to
Mohnia
. Most workers(including this study) accepted the view of Tembrock,
although the spindle-shaped shell with a long siphonalcanal makes the species from the Cenozoic of Eurasiaconsiderably more similar to the typical
Fusus
than to
Buccinum
. It should be said that in one of the latestpapers on these gastropods (Gürs and Schnetler, 2004)the genus
Aquilofusus
is not synonymized with
Sca-laspira
(
Mohnia
is not mentioned), but assigned toBuccinidae.
Several issues arose while studying the Transcas-pian
Scalaspira
. Firstly, a wide range of opinions onspecies identifications has been published in westernEuropean works, which were also extensively used byRussian workers. Differences in identification were sig-nificantly reduced by the 1960s after Tembrock’s revi-sion, but no complete consensus has been achieved upto now. Secondly, the precision and quality of identifi-cations of the Transcaspian material are reducedbecause of its state of preservation. In some species noprotoconchs are known and fine details of ornamenta-tion are not preserved (especially in early whorls), i.e.,characters which Tembrock and some other workersconsider of having high taxonomic value. Thirdly, up tothe end of the 1950s the stratigraphic distribution of ourspecies was difficult to estimate because of uncertain-ties in the geology of the Transcaspian Region. Amitrov(2005) published a paper on the Eocene–Oligocenegastropod assemblages of western Kazakhstan, inwhich he also discussed the stratigraphy of these beds.Having mentioned the above paper, we still want to reit-erate several of the most important issues (see alsoTable 1). Up to the end of the 1950s it was not acknowl-edged that in the northern Ustyurt Plateau and the AralSea Region, the Chegan Formation (a long knownseries with a rich fauna presently dated as UpperEocene and possibly upper part of the Middle Eocene)is overlain by at least two series with normal marine
Scalaspira
(Gastropoda) from the Paleogene of the Transcaspian Region
O. V. Amitrov and E. A. Zhegallo
Paleontological Institute, Russian Academy of Sciences, Profsoyuznaya ul. 123, Moscow, 117997 Russiae-mail: [email protected]; [email protected]
Received November 28, 2005
Abstract
—The composition and distribution of the species of the genus
Scalaspira
Conrad, 1862 in theEocene–Oligocene of western Kazakhstan are revised. Following Tembrock we assign this genus to Buccinidaeand synonymize
Aquilofusus
Kautsky, 1925 with it. The stratigraphic interval studied contains at least eight spe-cies. Three species are new:
S. alexeevi
(Middle? and Upper Eocene, Chegan Formation),
S. korobkovi
, and
S. kumsuatensis
(Upper Oligocene, Karatomak Beds).
DOI:
10.1134/S0031030107010029
Key words
:
Scalaspira
, Buccinidae, Upper Eocene, Oligocene, western Kazakhstan, Mangyshlak, Ustyurt.
PALEONTOLOGICAL JOURNAL
Vol. 41
No. 1
2007
SCALASPIRA
(GASTROPODA) FROM THE PALEOGENE OF THE TRANSCASPIAN REGION 17
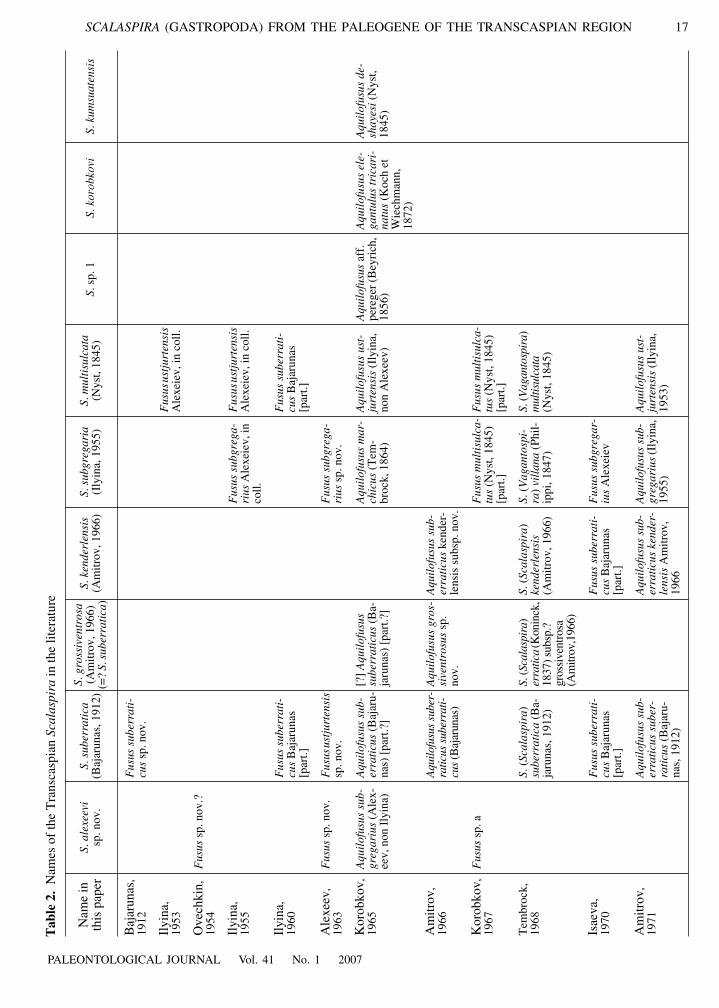
Tab
le 2
.
Nam
es o
f th
e T
rans
casp
ian
Scal
aspi
ra
in th
e lit
erat
ure
Nam
e in
th
is p
aper
S
.
alex
eevi
sp. n
ov.
S
.
sube
rrat
ica
(Baj
arun
as, 1
912)
S
.
gros
sive
ntro
sa
(A
mitr
ov, 1
966)
(=
?
S
.
sube
rrat
ica
)
S
.
kend
erle
nsis
(A
mitr
ov, 1
966)
S
.
subg
rega
ria
(I
lyin
a, 1
955)
S
.
mul
tisu
lcat
a
(N
yst,
1845
)
S
. sp.
1
S
.
koro
bkov
i
S
.
kum
suat
ensi
s
Baj
arun
as
, 19
12
Fus
us
sube
rrat
i-cu
s
sp.
nov
.
Ilyi
na
,19
53
Fus
us
ustj
urte
nsis
A
lexe
iev,
in c
oll.
Ove
chki
n
, 19
54
Fus
us
sp.
nov
.?
Ilyi
na
,19
55
Fus
us
subg
rega
-ri
us
Ale
xeie
v, in
co
ll.
Fus
us
ustj
urte
nsis
A
lexe
iev,
in c
oll.
Ilyi
na
,19
60
Fus
us
sube
rrat
i-cu
s
Baj
arun
as
[par
t.]
Fus
us
sube
rrat
i-cu
s
Baj
arun
as
[par
t.]
Ale
xeev
, 19
63
Fus
us
sp.
nov
.
Fus
us
ustj
urte
nsis
sp
. nov
.
Fus
us
subg
rega
-ri
us
sp.
nov
.
Kor
obko
v
, 19
65
Aqu
ilof
usus
sub-
greg
ariu
s
(A
lex-
eev,
non
Ily
ina)
Aqu
ilof
usus
sub-
erra
ticu
s
(B
ajar
u-na
s) [
part
.?]
[?]
Aqu
ilof
usus
su
berr
atic
us (
Ba-
jaru
nas)
[pa
rt.?
]
Aqu
ilof
usus
mar
-ch
icus
(T
em-
broc
k, 1
864)
Aqu
ilof
usus
ust
-ju
rten
sis
(Ily
ina,
no
n A
lexe
ev)
Aqu
ilof
usus
aff
. pe
rege
r (B
eyri
ch,
1856
)
Aqu
ilof
usus
ele
-ga
ntul
us tr
icar
i-na
tus
(Koc
h et
W
iech
man
n,
1872
)
Aqu
ilof
usus
de-
shay
esi (
Nys
t, 18
45)
Am
itrov
, 19
66A
quilo
fusu
s su
ber-
ratic
us s
uber
rati-
cus
(Baj
arun
as)
Aqu
ilof
usus
gro
s-si
vent
rosu
s sp
. no
v.
Aqu
ilof
usus
sub
-er
rati
cus
kend
er-
lens
is s
ubsp
. nov
.
Kor
obko
v,
1967
Fus
us s
p. a
Fus
us m
ulti
sulc
a-tu
s (N
yst,
1845
) [p
art.]
Fus
us m
ulti
sulc
a-tu
s (N
yst,
1845
) [p
art.]
Tem
broc
k,
1968
S. (
Scal
aspi
ra)
sube
rrat
ica
(Ba-
jaru
nas,
191
2)
S. (S
cala
spir
a)er
ratic
a (K
onin
ck,
1837
) sub
sp.?
gr
ossi
vent
rosa
(A
mitr
ov,1
966)
S. (
Scal
aspi
ra)
kend
erle
nsis
(A
mitr
ov, 1
966)
S. (
Vag
anto
spi-
ra)
vill
ana
(Phi
l-ip
pi, 1
847)
S. (V
agan
tosp
ira)
m
ultis
ulca
ta
(Nys
t, 18
45)
Isae
va,
1970
Fus
us s
uber
rati
-cu
s B
ajar
unas
[p
art.]
Fus
us s
uber
rati
-cu
s B
ajar
unas
[p
art.]
Fus
us s
ubgr
egar
-iu
s A
lexe
iev
Am
itrov
, 19
71A
quil
ofus
us s
ub-
erra
ticu
s su
ber-
rati
cus
(Baj
aru-
nas,
191
2)
Aqu
ilof
usus
sub
-er
rati
cus
kend
er-
lens
is A
mitr
ov,
1966
Aqu
ilof
usus
sub
-gr
egar
ius
(Ily
ina,
19
55)
Aqu
ilof
usus
ust
-ju
rten
sis
(Ily
ina,
19
53)

18
PALEONTOLOGICAL JOURNAL Vol. 41 No. 1 2007
AMITROV, ZHEGALLO
overlying beds, including those with a specific Kenderlifaunal assemblage, are of Rupelian age, being equiva-lents of the Astsheairyk Formation, and that the marineRupelian beds in Mangyshlak (like in the northernUstyurt) are overlain by the Solionovskii Horizon, andabove that by the marine Karagie Formation (Chattian,equivalent of the Karatomak Beds). The Karagie For-mation contains the faunal locality near the KolmyshRavine that was previously thought to belong to theUzunbas Formation. Finally, the fourth issue compli-cating the situation (referred to by Amitrov, 2005, seealso Table 2) is that the large monograph on Cheganmollusks written by A.K. Alexeev before World War IIwas only published in 1963. Some authors, using Alex-eev’s manuscript redescribed species established byAlexeev, but provided their own descriptions and fig-ured their own material. This also applies to Scalaspiraspecies described as Fusus ustjurtensis Alexeiev (Ily-ina, 1953) and F. subgregarius Alexeiev (Ilyina, 1955).In addition, at least one species was misidentified byA.P. Ilyina. The species Fusus ustjurtensis identified byIlyina is found in the Chattian (Karatomak) Beds,whereas Alexeev’s species is Rupelian (described fromtwo shells from the Astsheairyk Formation); this is asynonym of F. suberraticus Bajarunas, 1912. Such sit-uations emphasize the importance of the Code of Zoo-logical Nomenclature. In this case the authorship ofthese species should be attributed to Ilyina, irrespectiveof whether her understanding of the species is same asAlexeev’s. Another problem is whether the Transcas-pian species are synonyms or subspecies of well-knownEuropean species. It is clear from Table 2 that we fol-low Tembrock and synonymize Scalaspira ustjurtensiswith S. multisulcata (Nyst, 1845). This point is dis-cussed below.
MORPHOLOGICAL CHARACTERS OF SCALASPIRA AND THEIR TAXONOMIC
APPLICATION
Tembrock attached particular importance to themorphology of the apical part of the shell. Based on thisshe recognized the subgenus Vagantospira Tembrock,1968, with the type species S. multisulcata (to this sub-genus she also assigned S. villana and S. houchei, whileother species she assigned to the nominal subgenus). InScalaspira s. s. smooth whorls of the protoconch growin almost the same plane, and the apex appears blunt,whereas in Vagantospira, the protoconch is conical.Another difference is that in some species of the nomi-nal subgenus, axial ridges appear at the end of theembryonic part of the shell, along with the first spiralribs, to form together a reticulate pattern. InVagantospira this is not observed. Tembrock warnedthat species very similar in ornamentation of the lastwhorls may in fact belong to different subgenera. In theTranscaspian material, the early whorls are rarely pre-served, but it is clear that S. suberratica and similar spe-cies belong to the nominal subgenus, whereas S. sub-
gregaria and S. “ustjurtensis” certainly belong toVagantospira. The value of these differences is anothermatter. Tembrock herself noted that there are speciesthat appear intermediate between the two subgenera.We believe that the “axial ridges” are only morestrongly developed regular growth lines, which can alsobe observed in Vagantospira.
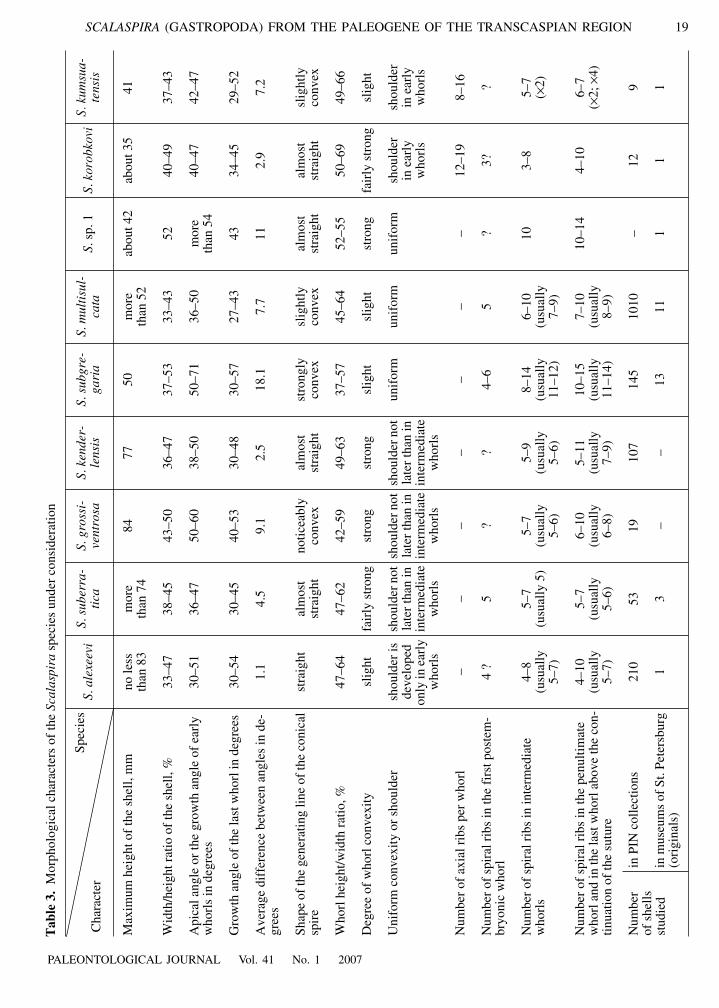
The species differ (even if slightly) in the size oftheir shells. Table 3 shows that among the Transcaspiantaxa four species from the S. suberratica group have amaximum shell height of 74–83 mm; S. subgregaria,and S. multisulcata, about 50 mm; and the other spe-cies, no more than 42 mm. However, it should be saidthat large specimens of each species are rarely foundand may be broken; therefore, there is a possibility thatfuture collecting will result in the discovery of largershells (this especially concerns species represented byfew specimens). It should also be remembered that themaximum (and average) sizes of shells of the same spe-cies may be different in specimens from different beds.For instance, the Uzunbas fauna is different from theKulandy and Astsheairyk faunas in the smaller size ofthe specimens, which is also true for Scalaspira(Amitrov, 1966, 1971).
The general shape of the shell is described by quan-titative characters such as the apical angle (if the apexis broken, the angle of the earliest preserved whorl hasto be measured), the growth angle of the last whorl, andthe width-to-height (W/H) ratios of the shell, aperture,a whorl, etc. Differences in these characters betweenspecies are not great, but for every two pairs of speciesthey can be very distinct and can be used for identifica-tion. In gastropods, the shell does not usually growcompletely isometrically. The spire is usually conicalwith a slightly convex generating line, the growth angleof the last whorl is smaller than the apical angle, and theW/H ratio in large shells is smaller than in small shells.This also applies to Scalaspira, but in our species,allometry is not very distinct. It is best observed inS. subgregaria and S. grossiventrosa, whereas in S.alexeevi the difference between the growth angles ofearly and late whorls is on average only one degree, thegenerating line may be considered straight, evenslightly concave in some specimens.
Species are also distinguished by the degree of con-vexity or angularity of whorls and the position of themaximum convexity or curvature.
Presence or absence of axial ornamentation is animportant character. However, as noted by Tembrock,there are Scalaspira species in which some specimenspossess axial ribs, whereas others do not. Sometimes itis difficult to distinguish elements of axial ornamenta-tion from growth lines. However, Transcaspian speciesdo not impose such difficulties. Seven of them do nothave axial ornamentation (if ridges in the earliestwhorls are not counted), whereas two species haveclear axial ribs.
Morphology of spiral ornamentation is very impor-tant for identification. Usually, the ornamentation of theearly whorls is the least changeable and therefore most
PALEONTOLOGICAL JOURNAL Vol. 41 No. 1 2007
SCALASPIRA (GASTROPODA) FROM THE PALEOGENE OF THE TRANSCASPIAN REGION 19
Tab
le 3
. M
orph
olog
ical
cha
ract
ers
of th
e Sc
alas
pira
spe
cies
und
er c
onsi
dera
tion
Spec
ies
Cha
ract
erS.
ale
xeev
iS.
sub
erra
-ti
caS.
gro
ssi-
vent
rosa
S. k
ende
r-le
nsis
S. s
ubgr
e-ga
ria
S. m
ulti
sul-
cata
S. s
p. 1
S. k
orob
kovi
S. k
umsu
a-te
nsis
Max
imum
hei
ght o
f th
e sh
ell,
mm
no le
ssth
an 8
3m
ore
than
74
8477
50m
ore
than
52
abou
t 42
abou
t 35
41
Wid
th/h
eigh
t rat
io o
f th
e sh
ell,
%33
–47
38–4
543
–50
36–4
737
–53
33–4
352
40–4
937
–43
Api
cal a
ngle
or
the
grow
th a
ngle
of
earl
y w
horl
s in
deg
rees
30–5
136
–47
50–6
038
–50
50–7
136
–50
mor
eth
an 5
440
–47
42–4
7
Gro
wth
ang
le o
f th
e la
st w
horl
in d
egre
es30
–54
30–4
540
–53
30–4
830
–57
27–4
343
34–4
529
–52
Ave
rage
dif
fere
nce
betw
een
angl
es in
de-
gree
s1.
14.
59.
12.
518
.17.
711
2.9
7.2
Shap
e of
the
gene
ratin
g lin
e of
the
coni
cal
spir
est
raig
htal
mos
t st
raig
htno
ticea
bly
conv
exal
mos
t st
raig
htst
rong
lyco
nvex
slig
htly
conv
exal
mos
t st
raig
htal
mos
t st
raig
htsl
ight
lyco
nvex
Who
rl h
eigh
t/wid
th r
atio
, %47
–64
47–6
242
–59
49–6
337
–57
45–6
452
–55
50–6
949
–66
Deg
ree
of w
horl
con
vexi
tysl
ight
fair
ly s
tron
gst
rong
stro
ngsl
ight
slig
htst
rong
fair
ly s
tron
gsl
ight
Uni
form
con
vexi
ty o
r sh
ould
ersh
ould
er is
de
velo
ped
only
in e
arly
w
horl
s
shou
lder
not
la
ter
than
in
inte
rmed
iate
w
horl
s
shou
lder
not
la
ter
than
in
inte
rmed
iate
w
horl
s
shou
lder
not
la
ter
than
in
inte
rmed
iate
w
horl
s
unif
orm
unif
orm
unif
orm
shou
lder
in e
arly
w
horl
s
shou
lder
in e
arly
w
horl
s
Num
ber
of a
xial
rib
s pe
r w
horl
––
––
––
–12
–19
8–16
Num
ber
of s
pira
l rib
s in
the
firs
t pos
tem
-br
yoni
c w
horl
4 ?
5?
?4–
65
?3?
?
Num
ber
of s
pira
l rib
s in
inte
rmed
iate
w
horl
s4–
8(u
sual
ly5–
7)
5–7
(usu
ally
5)
5–7
(usu
ally
5–6)
5–9
(usu
ally
5–6)
8–14
(usu
ally
11–1
2)
6–10
(usu
ally
7–9)
103–
85–
7(×
2)
Num
ber
of s
pira
l rib
s in
the
penu
ltim
ate
who
rl a
nd in
the
last
who
rl a
bove
the
con-
tinua
tion
of th
e su
ture
4–10
(usu
ally
5–7)
5–7
(usu
ally
5–6)
6–10
(usu
ally
6–8)
5–11
(usu
ally
7–9)
10–1
5(u
sual
ly11
–14)
7–10
(usu
ally
8–9)
10–1
44–
106–
7(×
2; ×
4)
Num
ber
of s
hells
stud
ied
in P
IN c
olle
ctio
ns21
053
1910
714
510
10–
129
in m
useu
ms
of S
t. Pe
ters
burg
(o
rigi
nals
)1
3–
–13
111
11

20
PALEONTOLOGICAL JOURNAL Vol. 41 No. 1 2007
AMITROV, ZHEGALLO
indicative. In many gastropod groups the onset of orna-mentation is clearly fixed after the smooth whorls of theprotoconch. Sometimes, the early ornamentation is dif-ferent from that in most of the teleoconch, and the bor-der between the “intermediate” and “main” zones maybe very distinct. However, in Scalaspira these bordersare not easily found. The initial portions of spiral ribsare very weak, and their place of appearance may beartificially “shifted” when the shell surface is evenslightly tattered or because of the quality of the micro-scope. In some species the “main whorls” may be dis-tinguished from the “intermediate” by the change in thecharacter of ribs from thin and widely spaced to ribbon-shaped, separated by narrow grooves. However, thistransition is gradual, occupying some part of the whorl,whereas in S. suberratica (unlike the closely similar S.alexeevi) the primary ribs from the very beginning areribbon-shaped and the “intermediate” whorls can onlybe distinguished by the presence of axial ridges.
Tembrock indicated the number of “primary spirals”for each Scalaspira species. In some species this num-ber is variable. In addition, the spiral ribs do not appearsimultaneously, and there is an element of subjectivismin the recognition of “primary” ribs. Tembrock did notinclude poorly visible ribs near the sutures in the num-ber of “primary ribs.” However, in different specimensof the same species and in different parts of the sameshell, such a rib may run alternately along the sutureand at a distance from it.
In later whorls growth changes are observed (as theshell grows, the number of ribs always increases), whilevariability increases (number of spiral ribs and the ribwidth-to-intercostal space ratio vary). The speciesS. subgregaria and S. multisulcata are not distin-guished based on the last criteria.
In Table 3 and in the identification key, where theentire range of variability has to be taken into account,the degree of overlap of different species appearsgreater than during the visual examination of the mate-rial. For S. subgregaria and S. multisulcata this con-cerns differences not only in the number of ribs, butalso in the W/H ratio of whorls. In Table 3 the overlapis so large that mentioning this character in the key isuseless, but in fact in most shells of S. subgregariawhorl height is less than half of its width, whereas inS. multisulcata it is greater than half of its width; thelast whorls of the largest specimens show no overlap inthis character.
Table 3 does not contain data on the ornamentationof the base of the shell. In the lower part of the siphonalcanal, the ribs become thin, closely spaced, and oftenfused with each other; and hence, difficult to calculate.In the upper part of the base (below continuation of thesuture), the ribs have approximately the same size,shape, frequency, as in the zones above the suture in thelast whorls. The presence or absence of spiral folds onthe inner surface of outer lip is discussed below. Foldsare present in one of 12 shells of Scalaspira korobkovi,two of nine shells of S. kumsuatensis, and on four of53 shells of S. suberratica. In many Chegan S. alexeevi,
the apertures are incomplete or filled with matrix; how-ever, in one shell, folds are visible, and in several othersthey are certainly absent. In S. multisulcata folds werenot observed in any of hundreds of shells from out-crops, but were observed in two shells from Mangy-shlak bore cores. In the other Transcaspian species,folds were not observed in any specimens. While itcould be suggested that in S. kenderlensis and S. gros-siventrosa they may not have been preserved, or wereunnoticed, in S. subgregaria many shells are reason-ably well preserved, and the absence of the folds onthem is certain.
Usually, folds do not extend far inside the apertureand cover only some areas. It is possible that this isrelated to unfavorable periods in the life of the mollusk.S. suberratica, in which folds occur in zones with vari-cose extensions of the shell wall, provides most supportfor this hypothesis. Unlike many other gastropodgroups, in Scalaspira varices are formed only episodi-cally, are present in a few specimens, and are mostlikely traces of illness.
Tembrock (1968) sometimes mentioned the pres-ence of folds in her species descriptions. However, shedid not attribute much importance to this character.Gründel (1991) agreed with this interpretation, but hementions presence or absence of folds among the char-acters distinguishing subspecies of S. multisulcata (seebelow).
It should be added that shells lacking true folds,including those species in which folds are neverpresent, sometimes also show indistinct spiral eleva-tions and depressions on the inner surface, which usu-ally but not always reflect ribbing on the outer surface.Fractures formed in a decaying shell have the samedirection. Apparently, this is connected to the shellmicrostructure and the way the shell grew.
Another character, pointed out by Gründel is thepattern of the growth line. In some specimens they areslightly incised or wavy (each incision corresponds to aspiral rib). It appears that the degree of incision varieseven within shells of the same species from the samesample. It partly depends on preservation (the incisionsare best visible when the outer layer is slightlyleached). However, in the Transcaspian S. multisulcatathe growth lines are generally considerably more undu-late than in S. subgregaria or in any of specimens ofS. multisulcata that we received from Belgium. Weagree with Gründel that this character can be used incombination with others to distinguish between subspe-cies and species.
IDENTIFICATION KEY TO TRANSCASPIAN SCALASPIRA
1.A. Spiral and axial ornamentations are present……2B. Only spiral ornamentation is present……………32(1A). A. Whorls are relatively strongly convex;
three thin spiral primary ribs are prominent in earlywhorls; axial ribs are approximately the same shape andwidth………………………………Scalaspira korobkovi

PALEONTOLOGICAL JOURNAL Vol. 41 No. 1 2007
SCALASPIRA (GASTROPODA) FROM THE PALEOGENE OF THE TRANSCASPIAN REGION 21
B. Whorls are weakly and unevenly convex; spiralribs are numerous and small, axial ribs are ridgelike andlarge……………………………………S. kumsuatensis
3(1B). A. Whorls (at least early) are bent above themiddle. The bent area and the area below possess largewide ribs, usually five. Weak ribs may be developedabove the shoulder……………………………………4
B. Whorls are evenly convex, evenly covered by spi-ral ribs, usually narrow; at least six or seven ribs arepresent…………………………………………………7
4(3A).A. Whorls are weakly convex, the shoulder isdeveloped only in early whorls………………S. alexeevi
B. Whorls are strongly convex, the shoulder ispresent at least until intermediate whorls………………5
5(4B).A. The ribs in the last whorl are clear, evenlycovering the entire shell surface, including thebase………………………………………S. kenderlensis
B. In the upper part of the last whorl, ribs are usuallysmoothened, while they are prominent and large at thebase……………………………………………………6
6(5B).A. The spire is highly conical with an almoststraight generating line: the apical angle is 36°–47°, thegrowth angle of the last whorl is on average smalleronly by 4.5°; W/H ratio of the shell is 38–45%………………………………………………………S. suberratica
B. The spire is shaped like a low cone with a notice-ably convex generating line: the apical angle is 50°–60°,the growth angle of the last whorl is on average by 9°smaller, W/H ratio of the shell is 43–50%………………………………………………………S. grossiventrosa
7(3B).A. Whorls are strongly convex; ornamenta-tion is weak……………………………Scalaspira sp. 1
B. Whorls are weakly convex, ornamentation strong………………………………………………………8
8(7B).A. The spire is conical with weakly convexgenerating line: the apical angle is 36°–50°, the growthangle of the last whorl is on average by 7.7° smaller; sixto ten (usually seven to nine) ribs in intermediate andlater whorls………………………………S. multisulcata
B. The spire is conical with strongly curved gener-ating line: the apical angle is 50°–71°, the growth angleof the last whorl is on average by 18° smaller; 8–15(usually 11–14) ribs are present in the intermediate andlater whorls……………………………S. subgregaria
COMPARISON OF TRANSCASPIAN AND EUROPEAN SPECIES
Table 3 and identification key show the differencesbetween the Transcaspian species. For three new spe-cies, comparisons follow their descriptions. Here, theremaining six taxa are compared with the westernEuropean species.
Shells of five of our species were sent to Tembrockand she published her opinion (Tembrock, 1968). Shefound that Fusus subgregarius Ilyina is a synonym ofScalaspira villana (Philippi). Indeed, the European
specimens that Tembrock identified as S. villana is verysimilar to S. subgregaria. However, Gründel (1991),who revised this group, found it impossible to deter-mine which species was meant from Philippi’s (1847)original description, while the holotype is lost. Appar-ently this is correct, but Gründel (1991) proposed a newname S. multisulcata magdeburgensis Gründel, 1991for the specimens that Tembrock identified as S. vil-lana. If Tembrock’s specimens are identical to the Tran-scaspian species (and it seems so), then they should benamed S. subgregaria (Ilyina, 1955), whereas the nameS. subgregaria remains valid for the Transcaspian spe-cies even if it is different from the European taxon.
As can be seen from Table 3, Tembrock synony-mized Fusus ustjurtensis Ilyina, 1953 with Scalaspiramultisulcata (Nyst, 1845), a view we support. This spe-cies is not very close to S. subgregaria (=? magdebur-gensis), and we, unlike Gründel, believe them to repre-sent different species. However, according to Gründelthe species S. multisulcata includes the subspeciessequens (Warneck, 1926) (= houchei Glibert, 1957) andmarchica Tembrock, 1964. Apparently these are indeedsubspecies of S. multisulcata, with very subtle differ-ences from the type subspecies and each other. Theshells of the type subspecies are 40 mm high, withgrowth lines not undulate, and almost always with foldsin the aperture. The shells of S. multisulcata marchicaare up to 35 mm high, with undulate growth lines and,usually, no folds in the aperture. This subspecies hasslightly more convex whorls and a more strongly deep-ened suture. In S. multisulcata sequens shells are larger(up to 50 mm), also with undulate growth lines andwithout folds in the aperture; whorls in this subspeciesare the most strongly flattened, and ornamentation isweak.
To which subspecies do the Transcaspian specimensbelong? The absence of folds in the aperture and wavygrowth lines distinguish them from the type subspecies.Tembrock, who did not consider these charactersimportant, identified the Transcaspian specimens as thetype subspecies, but it appears that, from their whorlshape and ornamentation, they are close to marchica,whereas they are very much different from the subspe-cies sequens in all characters except size. According toGründel, the three subspecies have different strati-graphic ranges: multisulcata s. s. comes from theLower and Middle Rupelian, marchica comes from theUpper Rupelian, and sequens comes from the Chattian.In the Transcaspian Region S. “ustjurtensis” are foundalmost exclusively in the Chattian (isolated and there-fore doubtful specimens were found in the Rupelian). Itremains unclear whether the Transcaspian specimensbelong to one of the European subspecies or representa separate subspecies of S. multisulcata ustjurtensis.
Tembrock expressed opinions different from ourown on the specimens that Amitrov (1966) identified asAquilofusus suberratica suberratica (Bajarunas),A. suberratica kenderlensis Amitrov and A. grossivent-
16
PALEONTOLOGICAL JOURNAL
Vol. 41
No. 1
2007
AMITROV, ZHEGALLO
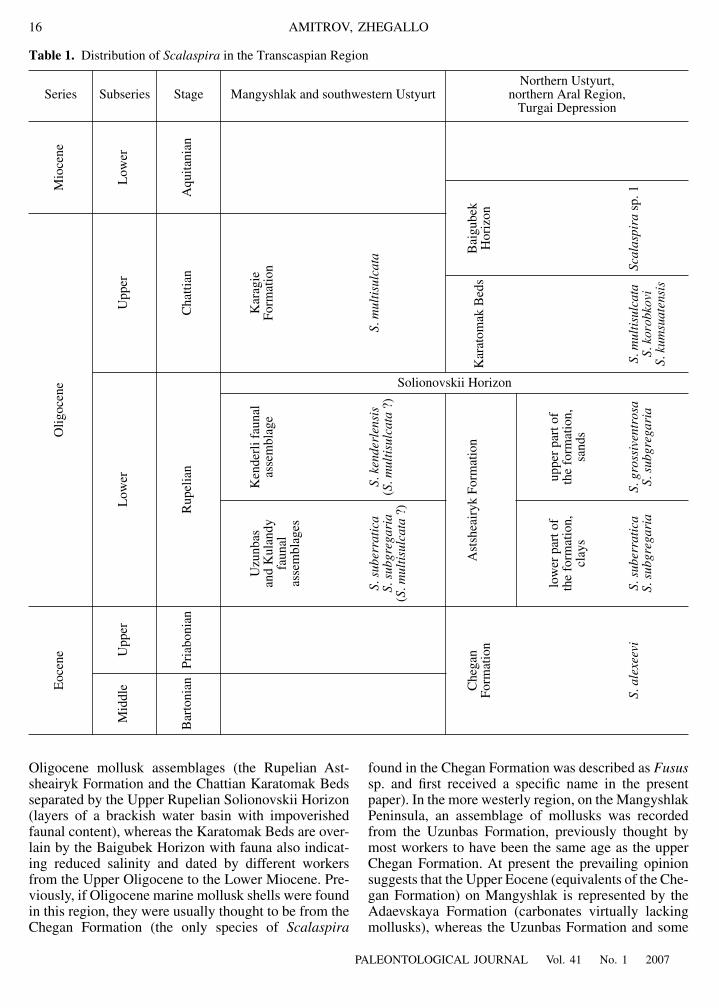
Oligocene mollusk assemblages (the Rupelian Ast-sheairyk Formation and the Chattian Karatomak Bedsseparated by the Upper Rupelian Solionovskii Horizon(layers of a brackish water basin with impoverishedfaunal content), whereas the Karatomak Beds are over-lain by the Baigubek Horizon with fauna also indicat-ing reduced salinity and dated by different workersfrom the Upper Oligocene to the Lower Miocene. Pre-viously, if Oligocene marine mollusk shells were foundin this region, they were usually thought to be from theChegan Formation (the only species of
Scalaspira
found in the Chegan Formation was described as
Fusus
sp. and first received a specific name in the presentpaper). In the more westerly region, on the MangyshlakPeninsula, an assemblage of mollusks was recordedfrom the Uzunbas Formation, previously thought bymost workers to have been the same age as the upperChegan Formation. At present the prevailing opinionsuggests that the Upper Eocene (equivalents of the Che-gan Formation) on Mangyshlak is represented by theAdaevskaya Formation (carbonates virtually lackingmollusks), whereas the Uzunbas Formation and some
Table 1.
Distribution of
Scalaspira
in the Transcaspian Region
Series Subseries Stage Mangyshlak and southwestern UstyurtNorthern Ustyurt,
northern Aral Region,Turgai Depression
Mio
cene
Low
er
Aqu
itani
an
Bai
gube
kH
oriz
on
Scal
aspi
ra
sp.
1
Olig
ocen
e
Upp
er
Cha
ttian
Kar
agie
Form
atio
n
S
.
mul
tisu
lcat
a
Kar
atom
ak B
eds
S
.
mul
tisu
lcat
aS
.
koro
bkov
i
S
.
kum
suat
ensi
s
Low
er
Rup
elia
n
Solionovskii Horizon
Ken
derl
i fau
nal
asse
mbl
age
S
.
kend
erle
nsis
(
S
.
mul
tisu
lcat
a
?)
Ast
shea
iryk
For
mat
ion
uppe
r pa
rt o
fth
e fo
rmat
ion,
sand
s
S
.
gros
sive
ntro
saS
.
subg
rega
ria
Uzu
nbas
and
Kul
andy
faun
alas
sem
blag
es
S
.
sube
rrat
ica
S
.
subg
rega
ria
(
S
.
mul
tisu
lcat
a
?)
low
er p
art o
fth
e fo
rmat
ion,
clay
s
S
.
sube
rrat
ica
S
.
subg
rega
ria
Eoc
ene U
pper
Pria
boni
an
Che
gan
Form
atio
n
S
.
alex
eevi
Mid
dle
Bar
toni
an


PALEONTOLOGICAL JOURNAL Vol. 41 No. 1 2007
SCALASPIRA (GASTROPODA) FROM THE PALEOGENE OF THE TRANSCASPIAN REGION 23
rosa Amitrov. Perhaps, this is partly related to the factthat the Transcaspian material which she saw was notwell preserved. We still do not have shells with proto-conchs for Scalaspira kenderlensis and S. grossivent-rosa, whereas in S. suberratica protoconchs are pre-served in only a few specimens (types, which we couldnot send abroad). A photograph of one of these pub-lished by Tembrock as a drawing (Tembrock, 1968,pl. 9, fig. 5) was also not of sufficient quality. Tembrocksaw on the shell only two “primary spirals” and thoughtthat this species is similar to S. erratica (Koninck). Infact S. suberratica has five primary “spirals” and fiveribs remain in the later whorls, whereas in S. erraticatwo large ribs are present even in later whorls.
Tembrock considered the species S. kenderlensisdistant from S. suberratica and believed it to be close tothe British species S. regularis (Sowerby). These spe-cies are similar in general shell shape and the strongconvexity of whorls, but their ornamentation is sharplydifferent (shells of S. regularis possess distinct axialribs). We noted (Amitrov, 2005) that the Kenderli Fau-nal Assemblage, despite its peculiarity, is still similar tothe synchronous and slightly earlier assemblages ofneighboring regions; the origin of the Kenderli Sca-laspira from the Uzunbas S. suberratica seems quiteprobable, whereas the immigration of the S. regularisgroup to the Kenderli basin would have appeared odd.Even morphologically, the Kenderli specimens are sosimilar to the Uzunbas–Kulandy species that at firstthey were described as a subspecies. Later, taking intoaccount Tembrock’s opinion we began consideringthem as a separate species.
Scalaspira grossiventrosa, in contrast, was from thebeginning described as a separate species because,based on the material available, it was more clearly sep-arated from S. suberratica. However, it seems possiblenow that this is not even a subspecies, but in Tem-brock’s terminology, a facial variety. In the Ustyurt,S. suberratica is found in the clayey facies in the lowerpart of the Astsheairyk Formation, whereas the morethick walled, low S. grossiventrosa (the name means“with a large belly”) come from the sandy facies of theupper part of the formation. The majority of our speci-
mens were collected on Mt. Karashoky, but the shellsfigured in the dissertation of A.I. Korobkov as Aquilo-fusus suberraticus also come from this locality (possi-bly from more fine-grained rocks). Judging from theapical angle (42° and 47°) these specimens should morelikely to be placed near S. suberratica than near S. gros-siventrosa. It is possible that the discovery of bettermaterial will allow a clearer separation of these formsor a justified synonymy. Tembrock did not find reliablecharacters in S. grossiventrosa distinguishing it fromS. erratica, but it seems that the Upper Astsheairykspecimens are even less similar to S. erratica than thetypical S. suberratica. The species from the BaigubekDeposits that we identify as Scalaspira sp. 1 was deter-mined by A.I. Korobkov as Aquilofusus aff. peregerBeyrich, but it seems that this species is not similar theNorth European Neogene species S. pereger, which hasaxial ornamentation (although weak) and relativelystrong, irregular spiral ornamentation.
SYSTEMATIC PALEONTOLOGYFamily Buccinidae Rafinesque, 1815
Genus Scalaspira Conrad, 1862Scalaspira alexeevi Amitrov et Zhegallo, sp. nov.
Plate 3, figs. 1–4
Fusus sp. nov. ?: Ovechkin, 1954, p. 80, pl. 11, figs. 11 and 12.Fusus sp. nov.: Alexeev, 1963, p. 111, pl. 21, figs. 1 and 2.Fusus sp. a: Korobkov, 1967, p. 68, pl. 1, fig. 25.E t y m o l o g y. In memory of A.K. Alexeev, spe-
cialist on the Paleogene mollusks of the TranscaspianRegion.
H o l o t y p e. PIN, no. 1470/41255; Kazakhstan,Turgai Depression, Atambas-Chink; Upper Eocene,Chegan Formation.
D e s c r i p t i o n. The shell is spindle-shaped, up to83 mm high (possibly even higher). W : H = 33–47%,AH : H = 52–61%. The spire is conical, with a generat-ing line varying from weakly convex to weakly concave(on average straight). The growth angle of early whorlsis 30°–51°, the growth angle of the last whorl is on aver-age smaller than one degree, but can be greater (up to54°). The whorl growth is regular. WH : WW = 47–64%.
E x p l a n a t i o n o f P l a t e 3All specimens are housed in the Depository of the Paleontological Institute of the Russian Academy of Sciences.Figs. 1–4. Scalaspira alexeevi sp. nov. Upper Eocene, Chegan Formation: (1) holotype PIN, no. 1470/41255, from the side oppositeto the aperture, ×1; Turgai Depression, Atambas-Chink; (2) specimen PIN, no. 1470/25011, apertural view, ×1.5, northern AralRegion, Mt. Termenbes; (3) specimen PIN, no. 1470/2657, early whorls, (3a) ×12, (3b) ×23, the same locality; (4) specimen PIN,no. 1470/39216, from the side opposite to the aperture, ×4, Turgai Depression, Chelkar-Nura Chink.Figs. 5 and 6. Scalaspira suberratica (Bajarunas, 1912). Lower Oligocene: (5) specimen PIN, no. 1470/48025, from the side oppo-site to the aperture, ×1.5, ? Mangyshlak, Uzunbas Formation; (6) specimen PIN, no. 1470/1680, early whorls, (6a) ×9.5, (6b) ×23,Mangyshlak, Uzunbas ravine, Uzunbas Formation.Figs. 7 and 8. Scalaspira grossiventrosa (Amitrov, 1966). Lower Oligocene, northern Ustyurt, upper part of the Astsheairyk For-mation: (7) holotype PIN, no. 1470/929, apertural view, ×1, Astsheairyk ravine (= Amitrov, 1966, pl. 2, fig. 11); (8) specimen PIN,no. 1470/1686, apertural view, ×1, Mt. Karashoky (= Amitrov, 1966, pl. 2, fig. 10).Figs. 9 and 10. Scalaspira kenderlensis (Amitrov, 1966). Lower Oligocene, Mangyshlak, Kenderli solonchak playa: (9) holotypePIN, no. 1470/1684, (9a) from the side of the outer lip, ×1, (9b) apertural view, ×1 (= Amitrov, 1966, pl. 2, fig. 8 = Amitrov, 1971,pl. 2, fig. 5); (10) specimen PIN, no. 1470/1683, from the side opposite to the aperture, ×1 (= Amitrov, 1966, pl. 2, fig. 9).


PALEONTOLOGICAL JOURNAL Vol. 41 No. 1 2007
SCALASPIRA (GASTROPODA) FROM THE PALEOGENE OF THE TRANSCASPIAN REGION 25
The whorls are weakly convex, in early whorls a moreor less noticeable shoulder occurs above the middle ofthe whorl, although the convexity remains somewhatirregular. The shell possesses only spiral ribs. Judgingfrom the specimen in which the early whorls are betterpreserved (Pl. 3, fig. 3), three identical ribs appear first,and soon the fourth similar rib appears from under thesuture. The upper rib runs above the mid-whorl and cor-responds to the shoulder. Later, between it and the sec-ond rib, two weaker ribs appear. In the subsequentwhorl they are the same size as the four primary ribs.One rib may appear before the shoulder. All ribs arethin at first, separated by considerably wider spaces, butthey gradually become wide, ribbonlike, whereas thespaces in the last whorls are usually slightly narrowerthan the ribs. In the whorls of the spire and in the lastwhorl above the continuation of the suture, the numberof ribs is usually five to seven, sometimes four, but mayoccasionally increase to ten due to the intercalation ofsecondary ribs. On the base of the shell, the ribs areapproximately of the same shape and size as in theupper part of the last whorl. Some large shells, espe-cially in their last whorls, are almost smooth, but evenin these ribbing is present on the base. In some speci-mens, thin folds (about 12) are present on the inner sur-face of the outer lip of the aperture.
M e a s u r e m e n t s o f t h e h o l o t y p e. H =64 mm, W = about 27 mm, AH = about 38 mm, AW =13 mm, apical angle is 39°, growth angle of the whorlis 47°, the number of spiral ribs in the early whorls is six,in the penultimate whorl there are eight ribs per whorl.
C o m p a r i s o n. Of all know Scalaspira species,the Chegan species most closely resembles the Rupe-lian S. suberratica (which is apparently its directdescendant). They share such characters as an irregularconvexity of the whorl, the presence of a shoulder inearly whorls, and a low number of ribs. However,already in the early whorls, the shoulder in S. alexeeviis weaker, whereas the convexity in the last whorls ismore regular. Judging from the few specimens withpreserved first postembryonic whorls, the ribs in S. sub-erratica from the very beginning are wide and ribbon-like, whereas in S. alexeevi, the ribs remain thin for
some time. The new species differs even more stronglyfrom S. grossiventrosa and S. kenderlensis. In S. gros-siventrosa the shell is relatively low, with a noticeablyconvex generating line, whereas in S. kenderlensis thelast whorls are covered by ribs that show no trendtoward smoothening. In S. erratica in early and usuallyin last whorls two strong ribs are sharply predominant.Even if the Transcaspian species are related to S. errat-ica, as was believed by Tembrock, S. alexeevi is lesssimilar to it than S. suberratica and S. grossiventrosa.This species is distinguished from S. subgregaria andS. multisulcata by the presence of a shoulder in theearly whorls and in having fewer ribs. The Chegan spe-cies is more similar to S. multisulcata from the Chattianof the Transcaspian Region than to the Rupelian S. sub-gregaria, from which it is also distinguished by theshape of the spire (in S. subgregaria the generating lineof the cone is strongly convex) and in the number of ribs.
O c c u r r e n c e. The upper Middle? and UpperEocene, Chegan Formation; Kazakhstan.
M a t e r i a l. Two hundred and ten specimens; ofwhich 203 are from the Turgai Depression (Chelkar-Nura Chink, Atambas-Chink, Mt. Atambas); others arefrom the northern Aral Region: Mt. Termenbes (fivespecimens), Altan-Chokusu Uplift (one specimen),Tuguzken Depression (one specimen). Almost all shellsare to some extent damaged and worn, none has a pro-toconch.
Scalaspira korobkovi Amitrov et Zhegallo, sp. nov.
Plate 4, figs. 10–12
E t y m o l o g y. In honor of A.I. Korobkov, special-ist in Cenozoic mollusks.
H o l o t y p e. PIN, no. 1470/29335; Kazakhstan,coast of the Aral Sea, Kumsuat Bay; Upper Oligocene,Karatomak Beds.
D e s c r i p t i o n. The shell is spindle-shaped, up to35 mm high. W : H = 40–49%, AH : H = 54–62%. Thespire is shaped like a cone with an almost straight gen-erating line: the growth angle of the early whorls is40°–47° (in one specimen 54°), the growth angle of the
E x p l a n a t i o n o f P l a t e 4Figs. 1–5. Scalaspira subgregaria (Ilyina, 1953). Lower Oligocene; (1) specimen PIN, no. 1470/2702, from the side opposite to theaperture, ×2, Mangyshlak, Kulandy Cliffs (= Amitrov, 1971, pl. 2, fig. 2); (2) specimen PIN, no. 1470/47626, from the side oppositeto the aperture, ×2, Mangyshlak, Kulandy Cliffs; (3) specimen PIN, no. 1470/784, from the side opposite to the aperture, ×2, north-ern Ustyurt, Astsheairyk Ravine, Astsheairyk Formation; (4) specimen PIN, no. 1470/1005 (juvenile), (4a) from the side oppositeto the aperture, ×13, (4b) early whorls, ×17.5, southern Aral Sea Region, from a borehole of the Karakol’ Branch of SGPK;(5) specimen PIN, no. 1470/930, apertural view, ×1, northern Ustyurt, Mt. Tamdy, Astsheairyk Formation.Figs. 6–9. Scalaspira multisulcata (Nyst, 1845). Upper Oligocene: (6) specimen PIN, no. 1470/28562, from the side opposite to theaperture, ×1.5, northern Aral Sea Region, Kumsuat Bay, Karatomak Beds; (7) specimen PIN, no. 1470/28564, apertural view, ×1.5,the same locality; (8) specimen PIN, no. 1470/28580, apertural view, ×2, the same locality; (9) specimen PIN, no. 1470/973, earlywhorls, ×12, southern Aral Sea Region, borehole of the Tashauz Branch of SGPK.Figs. 10–12. Scalaspira korobkovi sp. nov. Upper Oligocene, northern Aral Sea Region, Kumsuat Bay, Karatomak Beds: (10) holo-type PIN, no.1470/29335, from the side opposite the aperture, ×2; (11) specimen PIN, no. 1470/29333, apertural view, ×2;(12) specimen PIN, no. 1470/29342, from the side opposite to the aperture, ×3.Figs. 13 and 14. Scalaspira kumsuatensis sp. nov. Upper Oligocene, northern Aral Sea Region, Kumsuat Bay, Karatomak Beds:(13) specimen PIN, no. 1470/29350, from the side opposite to the aperture, ×2; (14) holotype PIN, no. 1470/29349, apertural view, ×2.

26
PALEONTOLOGICAL JOURNAL Vol. 41 No. 1 2007
AMITROV, ZHEGALLO
last whorl is 34°–45°; the difference between theseangles is on average 3°. WH : WW = 50–69%; this ratioslightly decreases from earlier to later whorls. Whorlsare relatively strongly convex. The early whorls quiteclearly show a shoulder above the middle of the whorlthat becomes smoothened in the last whorls in largespecimens, but the convexity remains irregular.
The ornamentation is represented by spirals andaxial ribs. The early whorls possess three spiral ribs(the upper rib is on the shoulder). The ribs are large, allthe same size, and are narrower than the intercostalspaces. Later, secondary ribs appear in the spaces andabove the shoulder. The secondary ribs quickly reachthe size of the three primary ribs. In larger shells, thetotal number of ribs in the penultimate whorl and in thelast whorl above the continuation of the suture reaches10. Similar ribs cover the base, where there are 12 ofthem. The axial ribs are usually more distinct in theearly whorls, where in their size and shape they are sim-ilar to spiral ribs (also narrow, with wider spaces) andform a regular lattice crossing the spiral ribs. In the lastwhorls, the axial ribs may become smoothened, loseregularity, and become very similar to growth lines.The number of the axial ribs is from 12 to 19 per whorl.On average it becomes slightly larger from the early tothe last whorls. In one of the specimens in our collec-tion the inner surface of the outer lip possesses 12 thinfolds of varying size.
M e a s u r e m e n t s o f t h e h o l o t y p e. Theholotype is at least 34 mm high and 13.6 mm wide;WH = about 21 mm; WW = 7.5 mm; growth angle ofthe early whorls is 47°, of the last whorl 42°, the num-ber of spiral ribs in early whorls is 4–5, 8 ribs in thepenultimate whorl above the suture, 12 ribs on the base,13 axial ribs in early whorls, and up to 19 ribs per whorlin later whorls.
C o m p a r i s o n. This species is distinguished fromother Transcaspian species of Scalaspira, except forS. kumsuatensis, by the presence of axial ribs. It differsfrom S. kumsuatensis also in the shape of the whorl (thepresence of the shoulder in early whorls and in the morestrongly developed and more irregular convexity in thelater whorls) and in the ornamentation, both spiral (inthe early whorls three large ribs instead of numerousweak ribs) and axial (the ribs are thin rather than wideand rounded ridgelike). Korobkov (1965) identified thisform as Aquilofusus elegantulus tricarinatus. However,the species Scalaspira elegantula (Philippi, 1843) fromthe Chattian of northern Europe retains a sharp shoul-der up to the later whorls and has only two spiral ribs oreven only one (the second may run below the suture andbecome visible only in the last whorl). In S. tricarinata(Koch et Wiechmann, 1872) (according to Tembrock,an independent species that also comes from the Chat-tian of northern Europe) there are three spiral ribs, butthis number remains the same in the later whorls; andneither the shoulder nor the axial ribs become smooth-ened. In its shape and ornamentation of the later whorls,
the new species resembles some Neogene species,especially S. semiglabra (Beyrich, 1856), but the latterspecies and other similar species have two strong spiralribs, whereas the third (upper), when present, is muchweaker.
M a t e r i a l. Twelve shells from the Karatomak areain the Kumsuat Bay. All shells are to some extent dam-aged, and none has a protoconch.
Scalaspira kumsuatensis Amitrov et Zhegallo, sp. nov.
Plate 4, figs. 13 and 14
E t y m o l o g y. From the Kumsuat Bay in theAral Sea.
H o l o t y p e. PIN, no. 1470/29349; Kazakhstan,coast of the Aral Sea, Kumsuat Bay; Upper Oligocene,Karatomak Beds.
D e s c r i p t i o n. The shell is spindle shaped, up to41 mm high, W : H = 37–43%, AH : H = 51–60%, thespire is in the shape of a cone with a weakly convexgenerating line: the growth angle in the early whorls is42°–47°, that of the last whorl is on average by 7°smaller. WH : WW = 49–66%, is almost independent ofthe size of the whorl. Whorls are weakly, evenly convex.
The shell possesses spiral and axial ribs. The spiralribs are weak and densely spaced. There are five toseven primary ribs in the intermediate and later whorls,but even weaker secondary ribs and, in larger speci-mens, tertiary ribs intercalate. Hence, the total numberof ribs may increase fourfold. The axial ribs are wide,ridgelike, usually 8 to 11 per whorl, in one shell even16. In some shells, the inner surface of the outer lip hasabout eight folds.
M e a s u r e m e n t s o f t h e h o l o t y p e. H =24.8 mm, W = 9.8 mm, AH = 13.6 mm, AW = 4.2 mm,the growth angle in the early whorls is 47°, that of thelast whorl is 33°, the number of spiral ribs is 5 × 2 in theearly whorls and 6 × 4 in the later whorls, and the num-ber of axial ribs is 10 per whorl.
C o m p a r i s o n. The comparison with S. korobkoviis above. The new species is distinguished from otherTranscaspian species by the presence of axial ribs. It isdistinguished from the northern European speciesS. deshayesi (Nyst, 1845), which was identified byKorobkov as the Transcaspian species, by the cleardivision of spiral ribs into the primary and secondaryribs. It differs from S. waeli (Nyst, 1852) in the samecharacter, and in the higher, less convex whorls andmore densely spaced axial ribs.
R e m a r k s. R. Janssen and J. Gründel (Germany)examined our material and agreed that this species isdifferent from the northern European species, and sug-gested that it possibly belongs to Streptochetus ratherthan to Scalaspira. Similar species of both genera aredistinguished based on protoconchs, which are not pre-served in our specimens.
O c c u r r e n c e. Upper Oligocene, KaratomakBeds; Kazakhstan.

PALEONTOLOGICAL JOURNAL Vol. 41 No. 1 2007
SCALASPIRA (GASTROPODA) FROM THE PALEOGENE OF THE TRANSCASPIAN REGION 27
M a t e r i a l. Nine specimens, of which seven comefrom the Karatomak area, Kumsuat Bay, northern AralSea Region, and two specimens come from theAshchik-Taipak area, northern Ustyurt.
ACKNOWLEDGMENTS
The manuscript of this paper and the materialdescribed was examined and commented on by R. Jan-ssen and J. Gründel (Germany) and by L.B. Iljina,L.A. Nevesskaja, S.V. Popov, and A.A. Shevyrev, ourcolleagues from the Laboratory of Mollusks (Paleonto-logical Institute, Russian Academy of Sciences).A.V. Guzhov helped in the preparation of the manu-script for publication. Some specimens were photo-graphed by V.T. Antonova. We are very grateful to allthese people.
The study was supported by the Russian Foundationfor Basic Research, project no. 04-04-48703.
REFERENCES
1. A. K. Alexeev, Paleogene Molluscan Fauna of theNorthern Urals Region (Akad. Nauk ArmSSR, Yerevan,1963) [in Russian].
2. O. V. Amitrov, “On the Distribution of Some AquilofususSpecies in the Paleogene of the Transcaspian Region,”Paleontol. Zh., No. 1, 28–37 (1966).
3. O. V. Amitrov, “Oligocene Gastropod Assemblages inthe Mangyshlak Peninsula and Southwestern UstyurtPlateau,” in Stratigraphy and Paleogeography of theCenozoic Oil- and Gas-Bearing Regions of the SouthernSoviet Union (Nedra, Moscow, 1971), pp. 65–81 [inRussian].
4. O. V. Amitrov, “Marine Gastropod Assemblages of theEocene and Oligocene of Western Kazakhstan,” Byull.Mosk. O–va Ispyt. Prir., Otd. Geol. 80 (1), 37–55 (2005).
5. M. V. Bajarunas, “Lower Oligocene Deposits of theMangyshlak Peninsula,” Zap. S.-Peterb. Mineral. O–va,Ser. 2 49, 19–69 (1912).
6. J. Gründel, “Die Untergattung Scalaspira (Vagantospira)Tembrock, 1968 (Gastropoda) im Oligozän des Nord-see-Beckens,” Z. Angew. Geol. 37 (3), 101–106 (1991).
7. K. Gürs and K. I. Schnetler, “Aquilofusus klugorum n. sp.(Gastropoda, Buccinidae) from Late Miocene of theNorth Sea Basin and Stratigraphical Implications of the
Genus Aquilofusus Kautsky 1925,” Meyniana 56, 5–20(2004).
8. A. P. Ilyina, “Mollusks of the Chegan Formation in theNorthern Scarps of the Ustyurt Plateau,” Paleontol. Sb.,No. 1, 93–134 (1953).
9. A. P. Ilyina, Paleogene Mollusks of the Northern UstyurtPlateau (Chegan and Ashcheairyk Formations) (Gostop-tekhizdat, Leningrad, 1955) [in Russian].
10. A. P. Ilyina, “Lower Oligocene Mollusks of the Mangy-shlak Peninsula,” Paleontol. Sb., No. 2, 265–298 (1960).
11. A. G. Isaeva, “Macropaleontological Characterization ofthe Spiroplectammina carinata–Cyclammina constricti-margo Zone of the Oligocene in the Western Scarp of theUstyurt Plateau,” in Stratigraphy and Paleontology ofthe Mesozoic and Paleogene Deposits of Central Asia(Nedra, Leningrad, 1970), pp. 175–179 [in Russian].
12. F. Kautsky, “Das Miocän von Hemoor und Basbeck-Osten,” Abh. Preuss. Geol. Landesanst., N.F., Issue 97,1–255 (1925).
13. A. I. Korobkov, “Mollusk-Based Stratigraphy of the Oli-gocene Deposits of the Northern Ustyurt Plateau and theNorthern Aral Region,” Extended Abstract of Candi-date’s Dissertation in Geology and Mineralogy (Lenin-grad, 1965).
14. A. I. Korobkov, “On the Age and Relationships betweenthe Ashcheairyk and Chegan Formations of the NorthernUstyurt Plateau and the Uzunbas Formation of theMangyshlak Peninsula,” in Geology of the TurgayDepression and the Aral Sea Region (Nedra, Leningrad,1967), pp. 62–73 [in Russian].
15. N. K. Ovechkin, Deposits of the Middle Paleogene of theTurgay Valley and the Northern Ural Mountains Region(Gosgeoltekhizdat, Moscow, 1954) [in Russian].
16. R. A. Philippi, “Verzeichniss der in der Gegend vonMagdeburg aufgefundenen Tertiärversteinerungen,” Palae-ontographica 1 (2), 45–90 (1847).
17. M. L. Tembrock, “Taxonomie des Formenkreises ‘Fususmultisulcatus’ Nyst, 1843 (Gastropoda),” Ber. Geol.Ges. DDR 9 (3), 308–310, 407–412 (1964).
18. M. L. Tembrock, “Zum Artproblem bei Gastropoden,erläutert an den Scalaspira (al. ‘Aquilofusus’)-Arten desnorddeutschen Oligozäns und Miozäns,” Ber. Geol. Ges.DDR 10 (4), 429–438 (1965).
19. M. L. Tembrock, “Taxonomisch-stratigraphische Studiezur Scalaspira-Gruppe (Gastropoda, Tertiär),” Paläon-tol. Abh. Abt. A 3 (2), 193–366 (1968).
20. W. Wenz, Gastropoda, Vol. 6: Th. 1. Allgemeiner Teilund Prosobranchia (Berlin-Zehlendorf, 1938).




![[Terrestrial mollusks (Mollusca: Gastropoda) of Costa Rica: classification, distribution and conservation]](https://img.dokumen.tips/doc/110x75/6359f385a93b2583d603882f/terrestrial-mollusks-mollusca-gastropoda-of-costa-rica-classification-distribution.jpg)
























