Embed Size (px)
Citation preview

GENOMICS 11,24-32 (1991)
Characterization of a Set of Variable Number of Tandem Repeat Markers Conserved in Bovidae
MICHEL GEORGES, ASOKA GUNAWARDANA, * DAVID W. THREADGILL, t MARK LATHROP, * INGRID OLSAKER,’ ANURADHA MISHRA, LESLIE L. SARGEANT, ANDREINA SCHOEBERLEIN,* MIKE R. STEELE, CHRISTIE TERRY,
DEBORAH S. THREADGILL, XUYUN ZHAO, TOM HOLM, RUDY FRIES,* AND JAMES E. WorwicKt
Genmark Inc., 47 7 Wakara Way, Salt Lake City, Utah 84108; l Gruppe Tierzucht, lnstitut fur Nutztierwissenschaften, ETH-Zentrum, Zurich, Switzerland; tDepartment of Veterinary Pathology, Texas A&M University, College Station, Texas 77843;
and Kentre d’Etude du Polymorphisme Humain (CEPH), Hepita/ St. Louis, Paris, France
Received February 2 1, 1991; revised May 2, 1991
Screening purpose-built libraries with minisatellite probes, we have isolated 36 bovine variable number of tan- dem repeat markers (VNTRs) characterized by a mean he- terozygosity of 59.3 within the American Holstein breed. Matching probabilities and exclusion powers were esti- mated by Monte-Carlo simulation, showing that the top 5 to 10 markers could be used as a very efficient DNA-based system for individual identillcation and paternity diagno- sis. The isolated VNTR systems should contribute signifi- cantly to the establishment of a bovine primary DNA marker map. Linkage analysis, use of somatic cell hybrids, and in situ hybridization demonstrate that these bovine VNTRs are scattered throughout the bovine genome, with- out evidence for proterminal confinement as in the human, and that at least some of them are organized as clusters. Moreover, Southern blot analysis and in situ hybridization demonstrate conservation of sequence and map location of minisatellites within Bovidae. o 1991 Academic PWSS, lnc.
INTRODUCTION
Human minisatellite sequences, exhibiting very high levels of genetic polymorphism due to variation in the number of tandem repetitions, have proven an invaluable source of genetic markers commonly termed “VNTRs” (Jeffreys et al., 1985; Nakamura et al., 1987a). VNTRs have been instrumental in the ge- netic mapping of several disease-causing genes, as tools for individual identification and paternity diag- nosis, and to address a variety of biological issues, including imprinting, loss of heterozygosity in malig- nancies, etc.
’ Present address: Department of Animal Genetics, Norwegian College of Veterinary Medicine, Oslo.
o&38-7543/91 $3.00 Copyright 0 1991 by Academic Press, Inc. All rights of reproduction in any form reserved.
In animal genetics, highly polymorphic markers such as VNTRs can similarly be used for individual identification and paternity diagnosis-relying today on less informative biochemical polymorphisms and blood group systems-and for the mapping of so- called economic trait loci (ETL) or genes involved in the determination of production traits. Classically, artificial selection has relied on the biometrical evalu- ation of breeding values from individual performance records and from performances of relatives (Falconer, 1989). One of the powers of the biometrical approach is that it obviates the need for any detailed molecular knowledge of the underlying genes or ETL. However, it is believed that the genetic mapping of ETL could be used to increase genetic response by affecting the accuracy and speed of selection, through a procedure called marker-assisted selection (MAS) (Soiler and Beckman, 1982; Smith and Simpson, 1986). More- over, defined alleles could be moved efficiently be- tween genetic backgrounds by velogenetics or the combined use of marker-assisted introgression and germline manipulations aimed to reduce the genera- tion interval (Georges and Massey, 1991; Georges, 1991). We report the cloning and characterization of 36 bovine variable number of tandem repeat (VNTR) markers, characterized by a high degree of polymor- phism within commercial herds and shown to be con- served within Bovidae.
MATERIALS AND METHODS
Cloning of Bovine VNTRs and IIetection of Polymorphism
Genomic DNA (500 pg) from 20 unrelated cows was digested to completion with Mb01 or HaeIII. After two rounds of size fractionation by agarose gel electropho- resis, electroelution, and addition of EcoRI linkers
24
VNTR MARKERS CONSERVED IN BOVIDAE 25
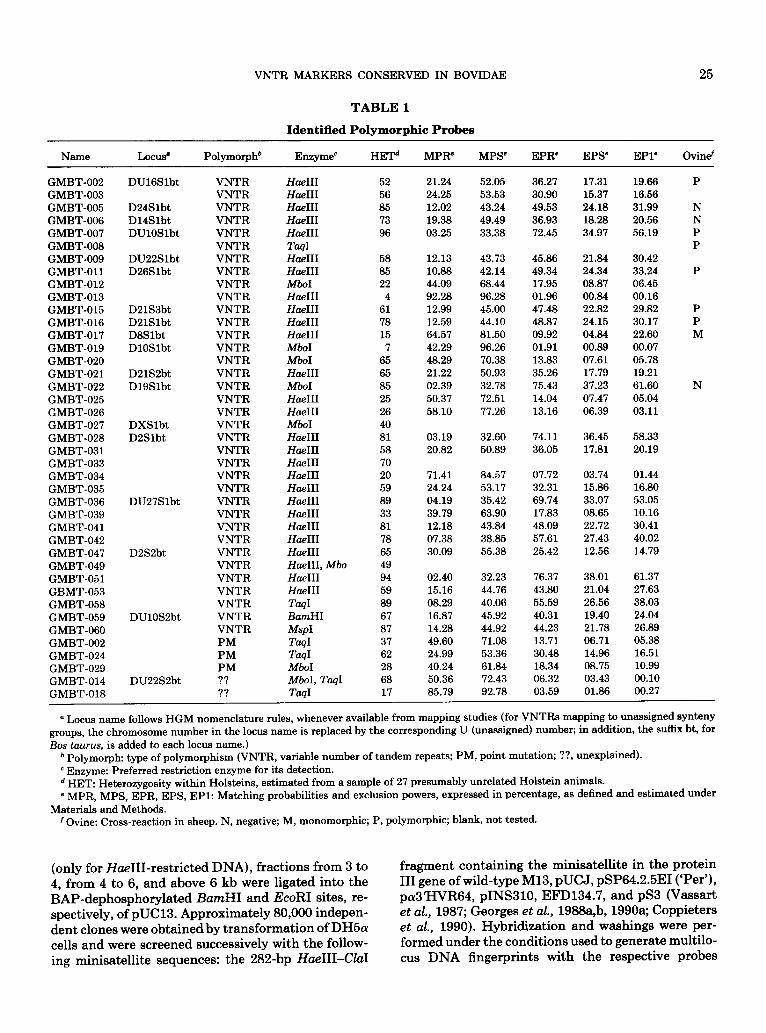
TABLE 1
Identified Polymorphic Probes
Name Locus” Polymorph* Enzyme’ HET” MPR’ MPS’ EPR’ EPS EPl’ Ovinef
GMBT-002 GMBT-003 GMBT-005 GMBT-006 GMBT-007 GMBT-008 GMBT-009 GMBT-011 GMBT-012 GMBT-013 GMBT-015 GMBT-016 GMBT-017 GMBT-019 GMBT-020 GMBT-021 GMBT-022 GMBT-025 GMBT-026 GMBT-027 GMBT-028 GMBT-031 GMBT-033 GMBT-034 GMBT-035 GMBT-036 GMBT-039 GMBT-041 GMBT-042 GMBT-047 GMBT-049 GMBT-051 GBMT-053 GMBT-058 GMBT-059 GMBT-060 GMBT-002 GMBT-024 GMBT-029 GMBT-014 GMBT-018
DUlGSlbt
D24Slbt DllSlbt DUlOSlbt
DU22Slbt D26Slbt
D21S3bt DZlSlbt D8Slbt DlOSlbt
D21S2bt DlSSlbt
DXSlbt D2Slbt
DU27Slbt
D2S2bt
DUlOS2bt
DU22S2bt
VNTR Hue111 VNTR Hoe111 VNTR Hoe111 VNTR Hoe111 VNTR Hoe111 VNTR TaqI VNTR Hoe111 VNTR Hoe111 VNTR Mb01 VNTR Hoe111 VNTR Hoe111 VNTR Hue111 VNTR Hoe111 VNTR Mb01 VNTR Mb01 VNTR Hoc111 VNTR Mb01 VNTR Hoc111 VNTR Hue111 VNTR Mb01 VNTR Hoe111 VNTR Hue111 VNTR Hue111 VNTR Hue111 VNTR Hue111 VNTR Hue111 VNTR Hue111 VNTR Hue111 VNTR Hoe111 VNTR HaeIII VNTR H&II, Mbo VNTR Hoc111 VNTR Hue111 VNTR TaqI VNTR BamHI VNTR MspI PM Td PM Td PM Mb01 ?? MboI, TaqI ?? TasI
52 21.24 52.05 36.27 17.31 56 24.25 53.53 30.90 15.37 85 12.02 43.24 49.53 24.18 73 19.38 49.49 36.93 18.28 96 03.25 33.38 72.45 34.97
58 85 22
4 61 78 15
7 65 65 85 25 26 40 81 58 70 20 59 89 33 81 78 65 49 94 59 89 67 87 37 62 28 68 17
12.13 43.73 45.86 21.84 10.88 42.14 49.34 24.34 44.09 68.44 17.95 08.87 92.28 96.28 01.96 00.84 12.99 45.00 47.48 22.82 12.59 44.10 48.87 24.15 64.57 81.50 09.92 04.84 42.29 96.26 01.91 00.89 48.29 70.38 13.83 07.61 21.22 50.93 35.26 17.79 02.39 32.78 75.43 37.23 50.37 72.51 14.04 07.47 58.10 77.26 13.16 06.39
03.19 32.60 74.11 36.45 58.33 20.82 50.89 36.05 17.81 20.19
71.41 84.57 07.72 03.74 01.44 24.24 53.17 32.31 15.86 16.80 04.19 35.42 69.74 33.07 53.05 39.79 63.90 17.83 08.65 10.16 12.18 43.84 48.09 22.72 30.41 07.38 38.85 57.61 27.43 40.02 30.09 55.38 25.42 12.56 14.79
02.40 32.23 76.37 38.01 61.37 15.16 44.76 43.80 21.04 27.63 08.29 40.06 55.59 26.56 38.03 16.87 45.92 40.31 19.40 24.04 14.28 44.92 44.23 21.78 26.89
49.60 71.08 13.71 06.71 05.38 24.99 53.36 30.48 14.96 16.51 40.24 61.84 18.34 08.75 10.99 50.36 72.43 06.32 03.43 00.10 85.79 92.78 03.59 01.86 00.27
19.66 P 16.56 31.99 N 20.56 N 56.19 P
P 30.42 33.24 P 06.45 00.16 29.82 P 30.17 P 22.60 M 00.07 05.78 19.21 61.60 N 05.04 03.11
a Locus name follows HGM nomenclature rules, whenever available from mapping studies (for VNTRs mapping to unassigned synteny groups, the chromosome number in the locus name is replaced by the corresponding U (unassigned) number; in addition, the suffix bt, for Bos taurw, is added to each locus name.)
* Polymorph: type of polymorphism (VNTR, variable number of tandem repeats; PM, point mutation; ??, unexplained). ’ Enzyme: Preferred restriction enzyme for its detection. d HET: Heterozygosity within Holsteins, estimated from a sample of 27 presumably unrelated Holstein animals. ’ MPR, MPS, EPR, EPS, EPl: Matching probabilities and exclusion powers, expressed in percentage, as defined and estimated under
Materials and Methods. f Ovine: Cross-reaction in sheep. N, negative; M, monomorphic; P, polymorphic; blank, not tested.
(only for HueIII-restricted DNA), fractions from 3 to fragment containing the minisatellite in the protein 4, from 4 to 6, and above 6 kb were ligated into the III gene of wild-type M13, pUCJ, pSP64.2.5EI (‘Per’), BAP-dephosphorylated BumHI and EcoRI sites, re- pa3HVR64, pINS310, EFD134.7, and pS3 (Vassart spectively, of pUC13. Approximately 80,000 indepen- et al., 1987; Georges et CLZ., 1988a,b, 1990a; Coppieters dent clones were obtained by transformation of DH5a et al., 1990). Hybridization and washings were per- cells and were screened successively with the follow- formed under the conditions used to generate multilo- ing minisatellite sequences: the 282-bp H&II-&I cus DNA fingerprints with the respective probes
26 GEORGES ET AL.
b 0.24
0.22 T
GMBT - 028 Hae III
a 0.2 Mean heterozygosiiy: 59.3%
5 0.18
95 0.16
2 0.14
ii 0.12
6 0.1
: s o.c@
g 0.06
IL 0134
0.02
0
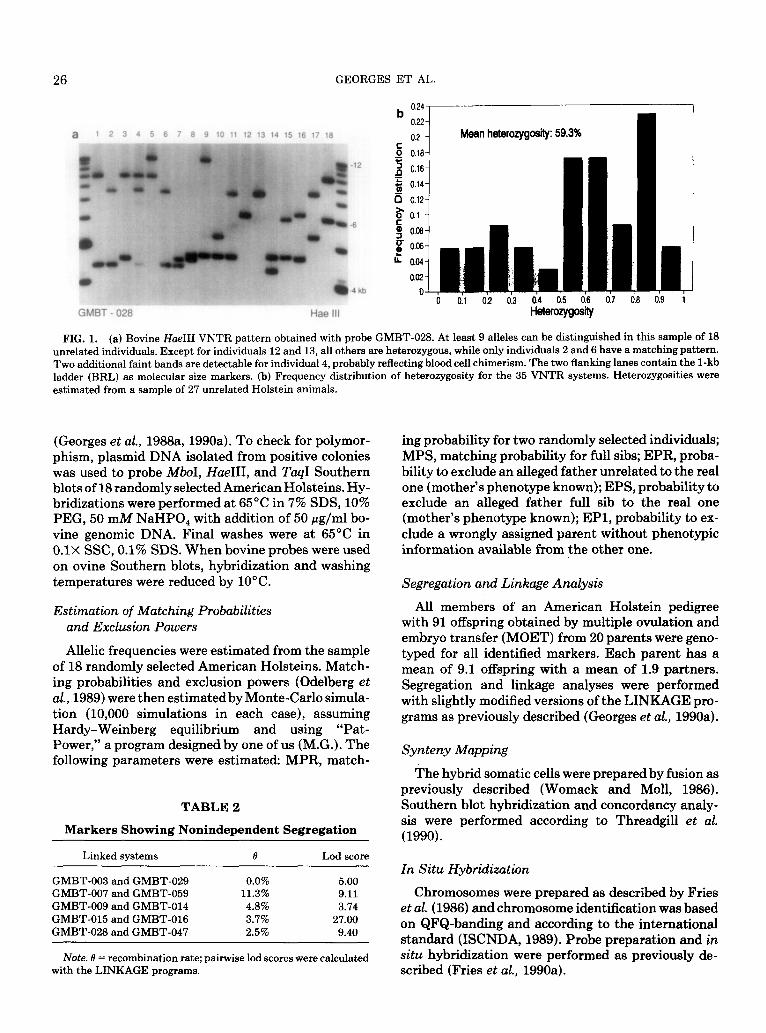
FIG. 1. (a) Bovine Hoe111 VNTR pattern obtained with probe GMBT-028. At least 9 alleles can be distinguished in this sample of 18 unrelated individuals. Except for individuals 12 and 13, all others are heterozygous, while only individuals 2 and 6 have a matching pattern. Two additional faint bands are detectable for individual 4, probably reflecting blood cell chimerism. The two flanking lanes contain the 1-kb ladder (BRL) as molecular size markers. (b) Frequency distribution of heterozygosity for the 35 VNTR systems. Heterozygosities were estimated from a sample of 27 unrelated Holstein animals.
(Georges et al., 1988a, 1990a). To check for polymor- phism, plasmid DNA isolated from positive colonies was used to probe MboI, HaeIII, and TaqI Southern blots of 18 randomly selected American Holsteins. Hy- bridizations were performed at 65°C in 7% SDS, 10% PEG, 50 mM NaHPO, with addition of 50 pg/ml bo- vine genomic DNA. Final washes were at 65°C in 0.1X SSC, 0.1% SDS. When bovine probes were used on ovine Southern blots, hybridization and washing temperatures were reduced by 10°C.
Estimation of Matching Probabilities and Exclusion Powers
Allelic frequencies were estimated from the sample of 18 randomly selected American Holsteins. Match- ing probabilities and exclusion powers (Odelberg et al, 1989) were then estimated by Monte-Carlo simula- tion (10,000 simulations in each case), assuming Hardy-Weinberg equilibrium and using “Pat- Power,” a program designed by one of us (M.G.). The following parameters were estimated: MPR, match-
TABLE 2
Markers Showing Nonindependent Segregation
Linked systems e Lod score
GMBT-003 and GMBT-029 GMBT-067 and GMBT-059 GMBT-009 and GMBT-014 GMBT-015 and GMBT-016 GMBT-028 and GMBT-047
0.0% 5.00 11.3% 9.11
4.8% 3.74 3.7% 27.06 2.5% 9.40
Note. 0 = recombination rate; pairwise lod scores were calculated with the LINKAGE programs.
ing probability for two randomly selected individuals; MPS, matching probability for full sibs; EPR, proba- bility to exclude an alleged father unrelated to the real one (mother’s phenotype known); EPS, probability to exclude an alleged father full sib to the real one (mother’s phenotype known); EPl, probability to ex- clude a wrongly assigned parent without phenotypic information available from the other one.
Segregation and Linkage Analysis
All members of an American Holstein pedigree with 91 offspring obtained by multiple ovulation and embryo transfer (MOET) from 20 parents were geno- typed for all identified markers. Each parent has a mean of 9.1 offspring with a mean of 1.9 partners. Segregation and linkage analyses were performed with slightly modified versions of the LINKAGE pro- grams as previously described (Georges et al., 1990a).
Synteny Mapping
The hybrid somatic cells were prepared by fusion as previously described (Womack and Moll, 1986). Southern blot hybridization and concordancy analy- sis were performed according to Threadgill et al. (1990).
In Situ Hybridization
Chromosomes were prepared as described by Fries et al. (1986) and chromosome identification was based on QFQ-banding and according to the international standard (ISCNDA, 1989). Probe preparation and in situ hybridization were performed as previously de- scribed (Fries et al., 1990a).

27 VNTR MARKERS CONSERVED IN BOVIDAE
TABLE 3
Percentage Concordancy Analysis for Synteny Mapping of 16 Bovine VNTRs, Using Somatic Cell Hybrids
Syntenic grp: 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 X Chromosome: 5 21 10 18 6 8 15 23 19 14 26
GMBT-002 65 47 55 55 45 50 45 25 69 40 75 60 70 37 45 100 70 75 55 70 40 60 55 40 50 45 70 55 45 60 GMBT-006 45 42 35 65 55 70 45 65 56 80 45 40 60 58 75 40 60 45 55 60 70 60 75 100 50 55 40 55 85 30 GMBT-007 55 53 45 75 35 70 45 75 56 100 35 50 60 63 65 40 60 45 55 70 90 70 75 80 50 55 50 75 75 10 GMBT-009 45 32 55 45 55 80 65 55 63 70 55 50 50 63 65 60 50 65 85 70 60 100 55 60 33 85 60 55 65 30 GMBT-011 30 37 50 40 50 65 60 50 63 55 50 35 35 68 50 45 35 60 70 55 55 85 40 55 50 100 45 40 60 45 GMBT-015 65 63 65 96 25 60 25 65 75 70 55 60 80 47 55 60 80 75 45 80 60 50 65 70 50 45 60 55 45 40 GMBT-016 78 61 65 96 61 87 83 74 74 70 57 74 70 83 79 56 83 88 70 83 80 80 24 72 55 80 0 54 0 30 GMBT-017 70 58 70 80 40 65 40 60 63 65 60 65 80 42 60 65 96 70 50 75 55 45 70 65 33 30 65 60 60 35 GMBT-019 40 32 50 20 100 55 80 40 31 35 70 45 45 63 60 45 45 50 70 25 35 55 50 55 50 60 45 30 60 65 GMBT-021 70 68 60 100 20 55 30 60 81 75 50 65 75 53 50 55 75 70 40 75 65 45 60 65 50 40 55 60 60 35 GMBT-022 50 47 30 60 30 55 40 60 50 85 30 35 45 58 50 35 45 30 40 55 95 55 60 75 67 60 35 60 70 25 GMBT-027 55 58 55 35 65 20 45 25 44 10 75 60 40 47 25 60 40 65 35 30 20 30 35 30 83 45 50 25 35 100 GMBT-036 65 58 75 55 45 50 45 45 56 50 75 70 60 42 45 70 60 65 55 70 40 60 55 40 50 45 100 65 45 50
Note. Reference markers for the respective synteny groups are Ul, GNBl; U2, MEl; U3, NKNB; U4, MPI; U5, FOS; U6, AMYl; U7, LDHA; U8, GNBZ; U9, GPI; UlO, SODl; Ull, VIM; U12, GPXl; U13, MET; U14, GSR; U15, CASK; U16, ABL: U17, CRYG; U18, GGTB2; U19, CAT; U20, GLOl; U21, GH; U22, AMH; U23, ALDHL; U24, TG; U25, CLTLA; U26, OAT, U27, DU27Slb; U28, MBP; U29, RBP3; and X, DMD. Synteny groups with highest concordancy scores, to which corresponding VNTRs were assigned, are shown in boldface.
RESULTS
A Set of Bovine VNTR Markers
Using the strategy described above, we have iso- lated a total of 36 bovine VNTRs, listed in Table 1. Polymorphic patterns were attributed to minisatellite sequences when characterized by more than two al- leles distinguishable with more than one restriction enzyme. Seven additional, non-VNTR-type polymor- phisms were detected during this experiment and are reported as well. Figure la shows a typical VNTR pattern obtained with probe GMBT-028. Within the American Holstein breed, the mean heterozygosity over all VNTR systems was 59.3%; the heterozygosity distribution is shown in Fig. lb. When probe GMBT- 016 was used with Mb01 insteadof HaeIII, and suppos- edly because of the presence of minisatellite variant repeats (MVR) (Jeffreys et al., 1990) harboring Mb01 sites, an extremely variable, locus-specific “midisa- tellite” pattern (Nakamura et al., 1987b) was gener- ated (data not shown). Used with MboI, this probe is particularly powerful for individual identification.
We found one clear instance of maternal neomuta- tion with probe GMBT-022. With this exception, all probes showed proper Mendelian segregation.
Table 1 reports estimated matching probabilities and exclusion powers as well. Systems GMBT-009, GMBT-011, and GMBT-022 were treated as “open” systems, meaning that-because of their small size- some alleles were not detectable under our conditions. To avoid ambiguities in identification and paternity diagnosis, these unidentified alleles were pooled in a
single “recessive” class. For individuals showing a single band, no distinction was made between homo- zygosity and heterozygosity based on band intensity. Discrepancies between probe ranking according to heterozygosity versus ranking according to matching probabilities and exclusion power most probably re- sult from the small sample size used to estimate both types of parameters. However, heterozygote advan- tage at some loci could be an alternative although un- likely explanation in view of the apparent neutral be- havior of human minisatellite sequences (Jeffreys et al., 1988).
Genomic Distribution
We performed pairwise linkage analysis between all markers. As it is known that at least some of these sequences are organized as minisatellite clusters (Royle et al., 1988, Georges et aZ., 1990a; Julier et al., 1990), we were expecting to find tightly linked sys- tems. We found evidence for five pairs of linked markers, four of which were characterized by a recom- bination rate inferior to 5% (Table 2). However, two of the systems involved in a tight linkage detect non- VNTR-type polymorphisms (GMBT-014, GMBT- 022). The corresponding probes were probably iso- lated because they contain a genuine although nonpo- lymorphic minisatellite, and were fortuitously detecting other types of polymorphism. Despite these five linked pairs, results of the linkage analysis pointed toward a scattering of these markers throughout the bovine genome.
Evidence for a broad genomic distribution of our
28 GEORGES ET AL.
a Number of grains
r--F-44
2
GMBT-028
Number of grains
-
14
GMBT-006
Number of grains 10 20 30 40 50 50
GMBT-016
Number of grains
-
24
GMBT-005
Number of grains
I+-++
10
GMBT-019
Number of grains
G-+===O
GMBT-022
Number of grains
r-f+++
21
GMBT-015
Number of grains
I-++-+
26
GMBT-0 11
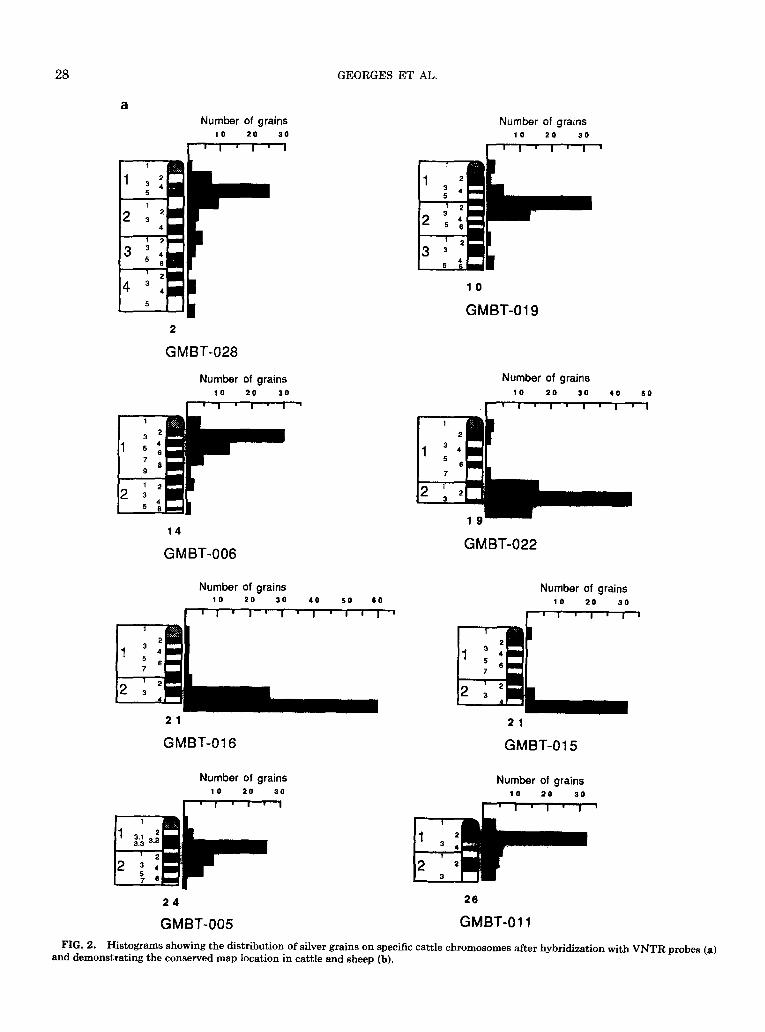
FIG. 2. Histograms showing the distribution of silver grains on specific cattle chromosomes after hybri&&,ion with VNTR probes (a) and demonstrating the conserved map location in cattle and sheep (b).
VNTR MARKERS CONSERVED IN BOVIDAE 29
b
Bos taurus Ovis aries
Number of grains Number of grains
10 7
GMBT-019
Number of grains Number of grains
19 11
GMBT-022
Number of grains Number of grains
21 18
GMBT-016
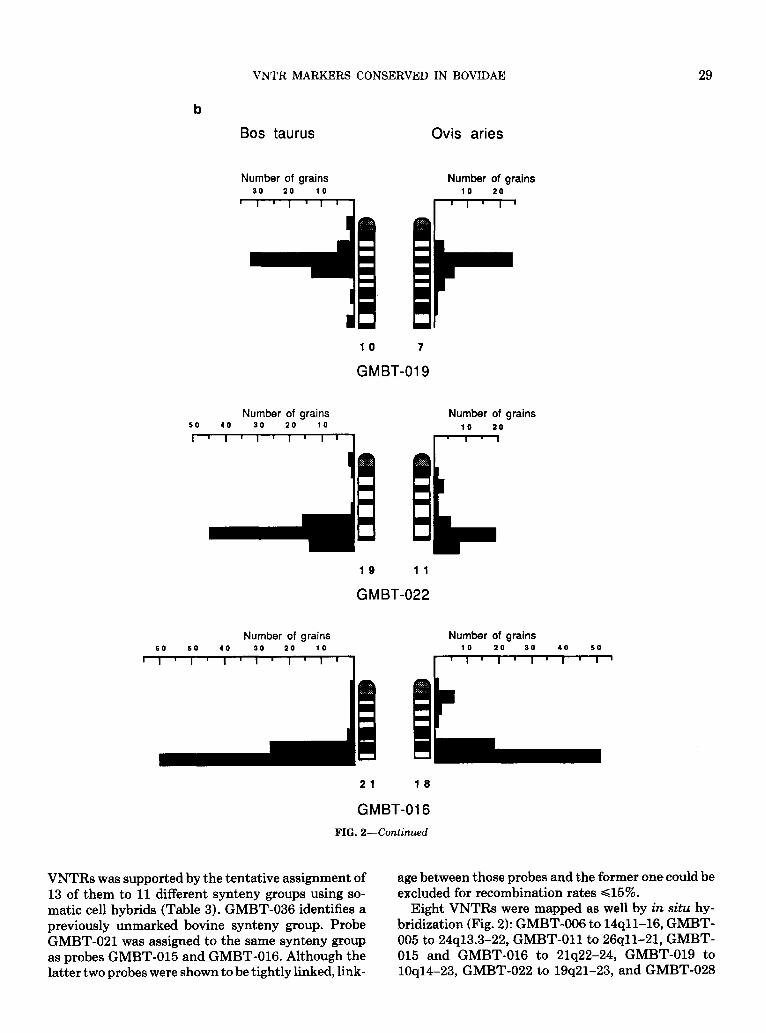
FIG. 2-Continued
VNTRs was supported by the tentative assignment of 13 of them to 11 different synteny groups using so- matic cell hybrids (Table 3). GMBT-036 identifies a previously unmarked bovine synteny group. Probe GMBT-021 was assigned to the same synteny group as probes GMBT-015 and GMBT-016. Although the latter two probes were shown to be tightly linked, link-
age between those probes and the former one cot&l be excluded for recombination rates ~15%.
Eight VNTRs were mapped as well by in situ hy- bridization (Fig. 2): GMBT-006 to 14qll-16, GMBT- 005 to 24q13.3-22, GMBT-011 to 26qll-21, GMBT- 015 and GMBT-016 to 21q22-24, GMBT-019 to lOq14-23, GMBT-022 to 19q21-23, and GMBT-028

30 GEORGES ET AL.
Cattle Sheep
1 2 3 4 5 6 7 8 9 10 11 12 12345
GMBT - 007 Mbol
FIG. 3. Sequence conservation of minisatellites within Bovi- dae. VNTR patterns obtained with bovine probe GMBT-007 in cattle (a) and sheep (b).
to 2q13-21. Again, good genomic coverage was evi- dent; six probes mapped to five different chromo- somes. Probes GMBT-015 and -016 both mapped to 21q23-24 as expected from the linkage study and the assignment on the hybrid panel. Surprisingly, five of the eight VNTRs clearly showed an interstitial map location. Only probes GMBT-015, -016, and -022 were located proterminally, the former two identify- ing the same minisatellite cluster. These results seem to contrast with those of Royle et al. (1988), which demonstrated preferential proterminal mapping of human VNTRs. Probe GMBT-011, previously lo- cated on U26, was mapped to chromosome 26, allow- ing us to tentatively assign synteny group U26 to chromosome 26.
Conservation of Sequence and Map Location within Bovidae
We hybridized 10 bovine VNTRs to sheep South- ern blots, under slightly reduced stringency condi-
tions. Seven of them were yielding locus-specific pat- terns, and six of these were showing a substantial de- gree of polymorphism in a sample of 5 unrelated sheep (Table 1). Figure 3 illustrates the patterns obtained with probe GMBT-007.
Probes GMBT-016, -019, and -022, mapping in the bovine to 21q23-qter, lOq15-q24, and 19q21-qter, re- spectively, were mapped by in situ in sheep as well (Figs. 2b and Table 4). The three probes produced signals on chromosomes 18, 7, and 11, recognized as evolutionary homologs of bovine chromosomes 21,10, and 19 (ISCNDA, 1989). Moreover, the signals were found over the exact positions as expected in the case of conservation of chromosomal location in cattle and sheep.
DISCUSSION
To isolate bovine VNTRs, we have used a strategy similar to that of Wong et al. (1986, 1987), based on the screening of size-selected restriction fragments obtained by complete digestion with the four-cutters Mb01 and HaeIII. Advantages of this strategy are as follows: (i) The complexity of this size range is sub- stantially reduced. Following Bishop et al. (1983) and assuming an exponential distribution of restriction fragment lengths, the fragments >2 kb represent about 10e3 of the total number of Mb01 or Hue111 frag- ments, corresponding to approximately lo4 frag- ments. This allows us to work readily with plasmid vectors. (ii) The subsequent search for and use of the polymorphism are performed with the same enzyme used to generate the libraries, obviating the need to screen several restriction enzymes, hence reducing costs. (iii) Relying on frequent four-cutters, the
TABLE 4
Analysis of Autoradiographic Silver Grain Distribution after Hybridization of the VNTR Probes with Cattle and Sheep Chromosomes
Probe
GMBT005 GMBTOOG GMBTOll GMBT015 GMBTO16 GMBTOl9 GMBT022 GMBT028
GMBTO16 GMBTO19 GMBT022
a Sheep.
Locus
D24Slbt D14Slbt D26Slbt D21S3bt DflSlbt DlOSlbt DlSSlbt D2Slbt
D2lSloa DlOSloa DlSSloa
No. metaphases scored
42 78 99 68
107 50 29
50
94 50 62
Total No. of grains
231
334 284 196 537 215 234
294
298 195 145
No. grains No. grains Peak of on specific at specific grain
chromosome region accumulation
46 (on 24) 44 (24q12-q24) 24q13.3/21 56 (on 14) 47(14qll-q16) 14q12113 55 (on 26) 42 (26qll-q21) 26q12/13 34 (on 21) 33 (21q22-qter) 21q24
135 (on 21) 130 (21q21-qter) 21q24 56 (on 10) 50 (lOq13-q24) lOq15-22 79 (on 19) 75 (19ql7-qter) 19q22/23 58 (on 2) 41 (2q12-q21) 2q13-15
77 (on 18)” 69 (lSq21-qter) 18q24 37 (on 7)” 30 (7q13-q24) 7q15-22 38 (on 11)” 32 (llql7-qter) llq22/23

VNTR MARKERS CONSERVED IN BOVIDAE 31
cloned minisatellites contain very little flanking se- quences and only very few of them carry highly re- peated sequences which would interfere during hy- bridization. (iv) Theoretically, the larger minisatel- lites targeted by this approach are more likely to be involved in mutational events and could therefore be the more polymorphic ones.
A disadvantage of this approach is the unequal rep- resentation of minisatellite loci in our library. The libraries were generated with a mixture of DNA from 20 unrelated individuals, to increase the number of clonable microsatellites. As a consequence, loci for which most alleles are within the selected size range will be overrepresented, compared to loci for which the majority of alleles in the population are below this range.
This collection of bovine VNTRs could be used for DNA-based individual identification and paternity diagnosis. Combining our top five probes, matching probabilities and exclusion powers at least as good as those obtained with classical systems are obtained: MPR, 8 X lo-*; MPS, 4 x 10-3; EPR, 0.999; EPS, 0.893; EPl, 0.987. Adding more probes will of course only increase the power of the system. In fact, these probes have been used efficiently to solve paternity problems beyond the power of blood group systems. DNA typing is not limited to blood samples as present systems are, which expands its spectrum of applica- tions and power. As an example, DNA typing has been used to deal efficiently with fetal blood cell chi- merism (Farber et al, 1989), frequently encountered in cattle. Compared with multilocus DNA finger- prints, locus-specific VNTRs are much easier to in- terpret and are more reproducible. Following properly established standardization procedures, a “common language” could be established, allowing exchange of information between laboratories. It is noteworthy that heterozygosity and allelic frequencies for some probes seem to vary substantially between breeds. As an example, probe GMBT-012 is characterized by a heterozygosity of 22% in Holsteins, but higher than 50% in both Herefords and Brown Swiss. Hence, proper use of these probes may initially require accu- rate estimation of genetic variation for different breeds.
Assuming a coverage of 20 CM per marker in link- age studies, the set of markers described in this paper would allow the scanning of approximately 7 Mor- gans. Accepting a total map length for the bovine ge- nome of 25 Morgans (Steele and Georges, 1991), this represents close to 33%. We have complemented the set of bovine VNTR described in this paper with over 80 multisite haplotypes, generated with cosmid probes, and more than 100 microsatellite systems (Georges et al., 1990a; Fries et aZ., 199Ob; Steele and Georges, 1991). Therefore, the majority of the bovine
genome is now amenable to linkage scanning. Since several of these markers are already “anchored” to specific chromosomes or synteny groups, a primary bovine DNA marker map should soon be available. Moreover, the remarkable conservation of mini- and microsatellites within Bovidae will substantially ac- celerate the construction of genetic maps in sheep and goats and offer the possibility to address interesting evolutionary issues.
ACKNOWLEDGMENTS
We are indebted to Dr. Joe Massey, Professor Ray White, Pro- fessor Ray Gesteland, Professor Jean-Marc Lalouel, Professor Gerald Stranzinger, and Mr. Jim Fordyce for their continuous in- terest and advice. We thank Dr. Stan Henderson andDr. Pat Mad- dox for giving us access to the bovine pedigrees and Dr. Jay Hetzel for providing us with sheep DNAs. We acknowledge Randy Weatherly for his advice with the “Pat-Power” program and Sa- bina Solinas for the initial help with in situ hybridization.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
REFERENCES
BISHOP, D. T., WILLIAMSON, J. A., AM) SKOLNICK, M. H. (1983). A model for restriction fragment length distributions. Am. J. Hum. Genet 35: ‘795-815.
COPPIETEF~S, W., VAN DE WEGHE, A., DEPICKER, A., Bou- QUET, Y., AND VAN ZEVEREN, A. (1990). A hypervariable pig DNA fragment. Anim. Genet. 21: 29-30.
FALCONER, D. S. (1983). “Introduction to Quantitative Genet- ics,” 3rd ed., Longman Scientific & Technical, New York.
FARBER, C. M., GEORGES, M., DEBOCK, G., VERHEST, A., SI- MON, P., AND VERSCHRAECEN-SPAE, M. (1989). Demonstra- tion of spontaneous XX/XY chimerism by DNA fingerprint- ing. Hum. Genet. 182: l-3.
FRIES, R., HEDIGER, R., AND STRANZINGER, G. (1986). Tenta- tive chromosomal localization of the bovine major histocom- patibility complex by in situ hybridization. Anim. Genet. 17: 281-294.
FRIES, R., SOLINAS, S., AND GUNAWARDANA, A. (1990a). Chro- mosomal assignment of marker loci to monitor spacing. in “Gene Mapping: Strategies, Techniques and Applications” (L. B. &hook, H. A. Lewin, and D. G. McLaren, Eds.), Dekker, New York/Basel.
FRIES, R., EGGEN, A., AND STFUNZINGER, G. (199Ob). The bovine genome contains polymorphic microsatellites. Geno- mics 8: 403-406.
GEORGES, M., LEQUARR& A. S., CASTELLI, M., HANSET, R., AND VASSART, G. (1988a). DNA fingerprinting in domestic an- imals using four different minisatellite probes. Cytogenet. Cell Genet. 47: 127-131.
GEORGES, M., COCHAUX, P., LF.QUAR&, A. S., YOUNG, M. W., AND VASSART, G. (1988b). DNA fingerprinting in man using a mouse probe related to part of the Drosophila “Per” gene. Nucleic Acids Res. 16: 7193.
GEORGES, M., LATHROP, M., HILBERT, P., MARCO=, A., SCHWERS, A., SWILLENS, S., VASSART, G., AND HANSET, R. (1990a). On the use of DNA fingerprints for linkage studies in cattle. Gerwmics 6: 461-474.
GEORGES, M., MISHRA, A., SARGEANT, L., STEELE, M., AND ZHAO, X. (1990b). Progress towards a primary DNA marker map in cattle. In “Proceedings, 4th World Congress on Genet-

GEORGES ET AL. 32
12.
13.
14.
15.
16.
17.
18.
19.
20.
its Applied to Livestock Production, Edinburgh, July 1990,” Vol. XIII, pp. 107-111.
GEORGES, M. (1991). Perspectives for marker assisted selec- tion and velogenetics in animal breeding. In “Applications of Research in Mammalian Development: Current Communica- tions in Cell and Molecular Biology 4” (Pedersen, McLaren, and First, Eds.), Cold Spring Harbor Laboratory Press. Cold Spring Harbor, NY.
GEORGES, M., AND MASSEY, J. M. (1991). Velogenetics or the synergistic use of marker assisted selection and germ-line ma- nipulation. Theriogenology 35: 151-156.
ISCNDA (1989). International System for Cytogenetic No- menclature of Domestic Animals Cytogenet. Cell Genet 53: 65-79.
JEFFREYS, A. J., WILSON, V., AND THEIN, S. L. (1985). Hyper- variable “minisatellites” regions in human DNA. Nature 314: 67-73.
JEFFREYS, A. J., ROYLE, N. J., WILSON, V., AND WONG, Z. (1988). Spontaneous mutation rates to new length alleles at tandem-repetitive hypervariable loci in human DNA. Nature 332: 278280.
JEFFREYS, A. J., NEUMANN, R., AND WILSON, V. (1990). Re- peat unit sequence variation in minisatellites: A novel source of DNA polymorphism for studying variation and mutation by single molecule analysis. Cell.
JULIER, C., DE GOUYON, B., GEORGES, M., GUENET, J. L., AVNER, P., AND LATHROP, G. M. (1990). Minisatellite linkage maps in the mouse by cross-hybridization with human VNTR probes. PFOC. Natl. Acad. Sci. USA 87: 4585-4589.
NAKAMURA, Y., LEPPERT, M., O’CONNELL, P., WOLFF, R., HOLM, T., CULVER, M., MARTIN, C., FUJIMOTO, E., HOFF, M., KUMLIN, E., AND WHITE, R. (1987a). Variable number of tan- dem repeat (VNTR) markers for human gene mapping. Science 235: 1616-1622.
NAKAMURA, Y., JULIER, C., WOLFF, R., HOLM, T., O’CON- NELL, P., LEPPERT, M., AND WHITE, R. (198713). Characteriza- tion of a human ‘midisatellite’ sequence. Nucleic Acids Res. 15: 2537-2547.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
ODELBERG, S. J., PLAETKE, R., ELDRIDGE, J. R., BALLARD, L., O’CONNELL, P., NAKAMURA, Y., LEPPERT, M., LALOUEL, J.-M., AND WHITE, R. (1989). Characterization of eight VNTR loci by agarose gel electrophoresis. Genomics 5: 915- 924.
ROYLE, N. J., CLARKSON, R. E., WONG, Z., AND JEFFREYS, A. J. (1988). Clustering of hypervariable minisatellites in the proterminal regions of human autosomes. Genomics 3: 352- 360.
SMITH, C., AND SIMPSON, S. P. (1986). The use of genetic polymorphism in livestock improvement. J. Anim. Breedg. Genet. 103: 205-217.
SOLLER, M., AND BECKMAN, J. S. (1982). Restriction frag- ment length polymorphisms and genetic improvement. In “Proceedings, 2nd World Congress on Genetics Applied to Livestock Production, Madrid,” Vol. 6, pp. 396-404.
STEELE, M. R., AND GEORGES, M. (1991). Generation of bo- vine multisite haplotypes using random cosmid clones. Geno- mics 10: 889-904.
THREADGILL, D. W., ADKINSON, L. R., AND WOMACK, J. E. (1990). Syntenic conservation between humans and cattle. I. Human chromosome 9. Genomics 8: 22-28.
VASSART, G., GEORGES, M., MONSIEUR, R., BROCAS, H., LE- QUARRE, A. S., AND CHRISTOPHE, D. (1987). A sequence in Ml3 phage detects hypervariable minisatellites in human and animal DNA. Science 235: 683-684.
WOMACK, J. E., AND MOLL, Y. D. (1986). Gene map of the cow: Conservation of linkage with mouse and man. J. Hered. 77: 2-7.
WONG, A., WILSON, V., PATEL, I., POVEY, S., AND JEFFREYS, A. J. (1987). Characterization of a panel of highly variable minisatellites cloned from human DNA. Ann. Hum. Gene& 51: 269-288.
WONG, Z., WILSON, V., JEFFREYS, A. J., AND THEIN, S. L. (1986). Cloning a selected fragment from a human DNA “fin- gerprint”: Isolation of an extremely polymorphic minisatel- lite. Nucleic Acids Res. 14: 4605-4616.





























