Embed Size (px)
Citation preview

ORIGINAL ARTICLE
Characterization and Distribution of Colonic Dendritic Cellsin Crohn’s Disease
Manuel A. Silva, MD,* Carolina B. López, PhD,† Francine Riverin,‡ Luc Oligny, MD,‡José Menezes, PhD,§ and Ernest G. Seidman, MD*
Abstract: Dendritic cells (DCs) are thought to play an importantrole in the pathogenesis of autoimmune inflammation, includingCrohn’s disease (CD). We investigated the distribution and stateof maturation of DCs in the colon in relation to the severity of inflamma-tion and therapy. Using archival specimens from colonic resections in 19pediatric patients with CD and 14 controls, we identified and character-ized the DCs within the lamina propria, submucosa, and muscularis com-partments using morphologic and quantitative immunohistochemicalmethods. The distribution of CD11c+CD83+CD68−DC-SIGN+ andimmature CD11c+CD83−CD68−DC-SIGN+ DCs within the differentcompartments varied according to the presence or absence of CD aswell as to the severity of inflammation and systemic corticoid treatment.Immature DCs were only found in non-inflamed control colonic tissue.Marked reductions (60% and 30%) in total CD11c+ and CD83+ DCnumbers were observed in CD tissue samples compared with controls(P < 0.05). CD samples from patients on corticosteroid therapy weresignificantly more depleted than in tissue from untreated patients orthose on other drugs. Colonic tissue with severe inflammation hadreduced numbers of CD11c+ and CD83+ DCs in the lamina propriaand submucosal compartments (80% and 76% for CD11c; 75% and76% for CD83, respectively, P < 0.05), with a concomitant increase(525% for CD11c and 700% for CD83 P < 0.05) of DCs in the muscu-laris compartment, compared to moderately inflamed and non-inflamedCD tissue. Our data suggest that an imbalance in intestinal DC subpopu-lations may play a role in the initiation and/or the maintenance of chronic
inflammation in CD. Corticosteroid therapy is associated with colonicDC depletion.
Key Words: Crohn’s disease, dendritic cells, immunopathogenesis,inflammation.
(Inflamm Bowel Dis 2004;10:504–512)
Despite recent advances in knowledge about mucosal im-munology, the etiology and pathogenesis of Crohn’s dis-
ease (CD) remain largely unknown. Increasing evidence sug-gests that the interplay between genetic, environmental, andimmune factors is crucial for the initiation of CD.1 Recently,dendritic cells (DCs) residing in the gut-associated lymphoidtissue (GALT) have become a focus of intense investigations.2
Thought to represent efficient antigen-presenting cells, DCsare likely to have a major role in the initiation, regulation, andmaintenance of the chronic inflammatory response observedin CD.
Studies have indicated that immature DCs acquire anti-gens from peripheral tissues and undergo maturation associ-ated with migration to secondary lymphoid tissues.3 MatureDCs present processed peptides to antigen-specific T cells, re-sulting in immune priming, whereas immature DCs induce Tcell anergy.3 During the maturation process, a variety of cyto-kines and cell surface markers such as co-stimulatory mol-ecules are induced and expressed, characterizing distinctstages of DC maturation. Recent studies suggest that semi-mature DCs induce immune tolerance.4 CD11c and DC-SIGNare markers of DCs, and mature and semi-mature DCs highlyexpress CD83.5–7 CD68 is established as an intracellularmarker for macrophages, allowing for their differentiationfrom DCs as antigen-presenting cells in IBD.8,9
Although the precise distribution of intestinal DCs is notwell delineated, murine studies suggest that they are predomi-nantly found in the Peyers patches of the lamina propria (LP).2
Studies in human intestinal tissue have revealed that smallpopulations of DCs are dispersed throughout the LP.10–20 Inview of the fact that CD is a transmural disease, it is importantto characterize the distribution of DCs throughout the compart-ments of the bowel wall compared with non-IBD tissue. In
Received for publication February 28, 2004; accepted May 6, 2004.From the *Mucosal Immunology Laboratory, Division of Gastroenterology,
Hepatology & Nutrition, Sainte-Justine Hospital Research Centre, Depart-ment of Paediatrics, University of Montreal, Montreal, Quebec, CanadaH3TICS; the †Department of Microbiology, Mount Sinai School of Medi-cine, New York, New York 10029; and the Sainte-Justine Hospital Re-search Centre, Departments of ‡Pathology and §Immunology, Universityof Montreal, Montreal, Quebec, Canada H3TICS.
Supported by a Crohn’s & Colitis Foundation of Canada (CCFC) operatinggrant (E.G.S., J.M.), by a Research Fellowship Award from the CanadianAssociation of Gastroenterology/Axcan Pharma (M.S.), by a Canada Re-search Chair in IBD from the CCFC and Canadian Institutes of HealthResearch (E.G.S.), and by a National Research Award of Excellence fromthe Fonds de la Recherche en Santé du Québec (E.G.S.).
Reprints: Ernest Seidman, MD, Chief, Division of Gastroenterology, Ste. JustineHospital, 3175 Cote Ste. Catherine Rd., Montreal, Quebec, H3T 1C5,Canada (e-mail: [email protected]).
Copyright © 2004 by Lippincott Williams & Wilkins
504 Inflamm Bowel Dis • Volume 10, Number 5, September 2004

addition, their characterization with respect to the severity ofinflammation would afford further insights into the role of DCsin the pathogenesis of CD.
PATIENTS AND METHODS
Patients and Tissue SpecimensParaffin-embedded colonic tissue was obtained from ar-
chival specimens from CD and controls. The diagnosis of CDwas made on the basis of established clinical, bacteriological,radiologic, endoscopic, and histopathological criteria.21 Indi-cations for bowel resection were the presence of intestinal per-foration, stenosis with clinical signs of obstruction, or therapy-resistant CD. Nineteen consecutive colonic resections (1999–2002) from pediatric CD patients were used (6 male, 13female; mean age 16 ± 2 years). All but 4 patients were receiv-ing various medications, including 5-aminosalicylic acid, cor-ticosteroids, and 6-mercaptopurine. In 8, corticosteroids wereused in combination with the other drugs, whereas 7 otherswere not on corticosteroids. A total of 25 paraffin-blocks wereobtained with available tissue. In 5 cases, more than 1 blockoriginated from the same patient. The histopathology wasevaluated by an experienced pathologist and scored on the ba-sis of previously described criteria.22 Tissue samples were thenclassified into 3 groups: non-inflamed CD (11 blocks: 9 ce-cum, 1 transverse colon, 1 sigmoid); mildly inflamed CD (7blocks: 6 cecum, 1 descending colon); and moderately in-flamed CD (7 blocks: 5 cecum, 2 sigmoid). Multiple blocksoriginating from the same patient were classified individually,according to the index of severity of inflammation22: one non-inflamed and one mildly inflamed block in 2 cases and non-inflamed and moderately inflamed specimens in 3 other cases.
As non-inflammatory controls, 14 paraffin-blocks wereobtained from archival surgical specimens (6 rectum, 5 sig-moid, 2 ascending colon, 1 descending colon) collected duringthe past 8 years from pediatric patients (8 male, 6 female; meanage, 4.3 ± 3 years). The indications for colonic resection wereHirschsprung’s disease (n = 12) or pelvic trauma (n = 2). In-formed consent for use of tissue was obtained from all the pa-tients, or if less than age 14, from their parents or legal guard-ians. The study protocol and consent forms were approved bythe Research Ethics Committee of the Sainte-Justine HospitalResearch Center.
Immunoperoxidase Staining for QuantitativeAnalysis of DCs
Three serial sections were sequentially stained with anti-CD11c, anti-CD83, and anti-CD68 antibodies using standardimmunoperoxidase techniques. The sections were deparaf-finized and rehydrated. Before carrying out the anti-CD11c oranti-CD83 immunoreactions, the sections were submerged in0.5% H2O2 to block endogenous peroxidase activity. Afterwashing in PBS, they were treated with a 0.05% trypsin solu-
tion in 0.05% calcium chloride (pH 7.8, 30 minutes) to unmaskthe antigenicity. They were then incubated (10 minutes) withuniversal blocking solution (Dako, Glostrup, Denmark) toblock any nonspecific antibody binding. After washing inPBS, they were incubated with anti-human CD11c (mouseIgG1; mAb B-ly6; 1:50; BD Biosciences-Pharmigen, SanDiego, CA) and anti-human CD83 (mouse IgG; mAbHB15A17.11; 1:50; Accurate Chemical & Scientific Corpora-tion, New York, NY) overnight at 4°C. Subsequently, the sec-tions were treated (90 minutes) with biotinylated anti-mouseIgG (Dako) and with peroxidase-labeled streptavidin(LSAB+plus System, Dako). Sections stained for anti-CD68were first blocked for endogenous peroxidase activity (10 min-utes) with 0.5% H2O2 followed by universal blocking solution(Dako) for another 10 minutes. Incubation (1 hour) with anti-human CD68 (mouse IgG mAb; 1/100; Dako) was then per-formed, followed by a peroxidase-labeled polymer anti-mouseIgG (EnVision System, Dako, Mississauga, ON, Canada). Per-oxidase activity was determined with diaminobenzidine solu-tion for 30 minutes. (Sigma D9015). Sections were counter-stained with Mayer’s haematoxylin and mounted in glycerolmedium. Human acute-inflamed tonsil tissue was used as apositive control for all the antibodies. As a negative control,the same tissue was stained with either an isotype-matched an-tibody of inappropriate specificity or by omitting the primaryantibody. Forty high-power fields were used to evaluate thelocation and morphology of stained cells. Only stained cellswith a monocyte morphology were counted. Cells with a lym-phocytic and polynuclear neutrophil morphology or lacking anucleus were not considered. Macrophages were characterizedby their typical round or oval nuclei and relative abundant cy-toplasm containing phagocytozed vesicles or fragments andCD68 positive staining.23 The number of CD83 and CD11cpositive cells in each compartment of the colonic wall was de-termined in 1 or 2 different randomly selected areas/slide, de-pending on whether the tissue sample was less than or longerthan 1 cm, respectively. A total of 33 different areas from the25 CD blocks and 20 areas from the 14 non-inflamed controlblocks were evaluated for each immunoperoxidase stainingtechnique. The same areas were selected in the consecutiveserial sections. Cell number was expressed in terms of area(per mm2) of LP, submucosa, and muscularis compartments.
Immunoperoxidase Double StainingThe last 2 serial sections were deparaffinized, rehy-
drated, blocked with 0.5% H2O2, and pretreated with 0.05%trypsin solution in 0.05% calcium chloride (pH 7.8) for 30minutes. After universal blocking as above for 10 minutes(Dako), an overnight incubation at 4°C was performed withone of the following pairs of antibodies: (1) CD11c and CD83double staining: anti-human CD11c (goat, IgG sc-6620, 1/50,Santa Cruz Biotechnology, Santa Cruz, CA) and anti-humanCD83 (1/50 dilution); (2) CD11c and DC-SIGN double stain-
Inflamm Bowel Dis • Volume 10, Number 5, September 2004 Characterization and Distribution of Colonic Dendritic Cells in CD
© 2004 Lippincott Williams & Wilkins 505
ing: mouse anti-human CD11c (1/10 dilution) and anti-humanDC-SIGN (goat, IgG sc-1138, 1/50, Santa Cruz Biotechnol-ogy).
Sections were then treated with biotinylated rabbit anti-goat IgG for 90 minutes (1/400) followed by streptavidin-alkaline phosphatase for 30 minutes (1/100, both from Dako).Staining was then performed with Fast Red System for 15 min-utes (Dako), giving a red precipitate. Subsequent to biotinblocking for 20 minutes, incubations with biotinylated goatanti-mouse IgG using 1/100 and 1/25 dilutions for the first andthe second double staining were carried out, followed by strep-tavidin-peroxidase incubation for 30 minutes (Dako). Peroxi-dase activity was determined with diaminobenzidine solutionfor 15 minutes, yielding a brown precipitate. Sections werecounterstained with Mayer hematoxylin as well. Human tonsiltissue was used as a positive inflammatory control, whereasthe same tissue immunoassayed with an isotype-matched an-tibody of inappropriate specificity served as a negative control.Also, additional serial sections were subjected to single stain-ing with each of the markers used in both double staining tech-niques.
Statistical AnalysisThe mean number of stained cells (±SD) per mm2 within
each of the compartments was calculated as the numerical sum
of each of the values obtained for the individual compartmentsand defined in terms of the index of severity of inflammation.22
Differences were evaluated by the Student t test or by ANOVAfor comparison of the mean number of cells within each com-partment, compared with their severity of inflammation. Logtransformation of the data was performed to obtain near-normal distributions prior to application of the ANOVA test.Significance was assessed at the 5% level (P value < 0.05). Allanalyses were carried out using SPSS version 10.0.5 (SPSSInc., Chicago, IL).24
RESULTS
Colonic DC Morphology and DistributionThe results for the serial sections subjected to single
staining for CD11c and CD83 are shown in Figures 1 and 2,respectively. An intense reaction for both markers was ob-served in cells located in all 3 colonic compartments, butmainly in the submucosa of control and non-inflamed CD tis-sue sections. Determination of CD11c and DC-SIGN revealeddouble-stained monocytes in all control and CD samples (Fig.3). Whereas staining for CD83 and CD11c showed double-positive monocytes in all CD samples, some CD11c+ mono-cytes were CD83− in the control tissue sections (Fig. 4). Stain-ing detected CD68+ macrophages in all areas selected for the
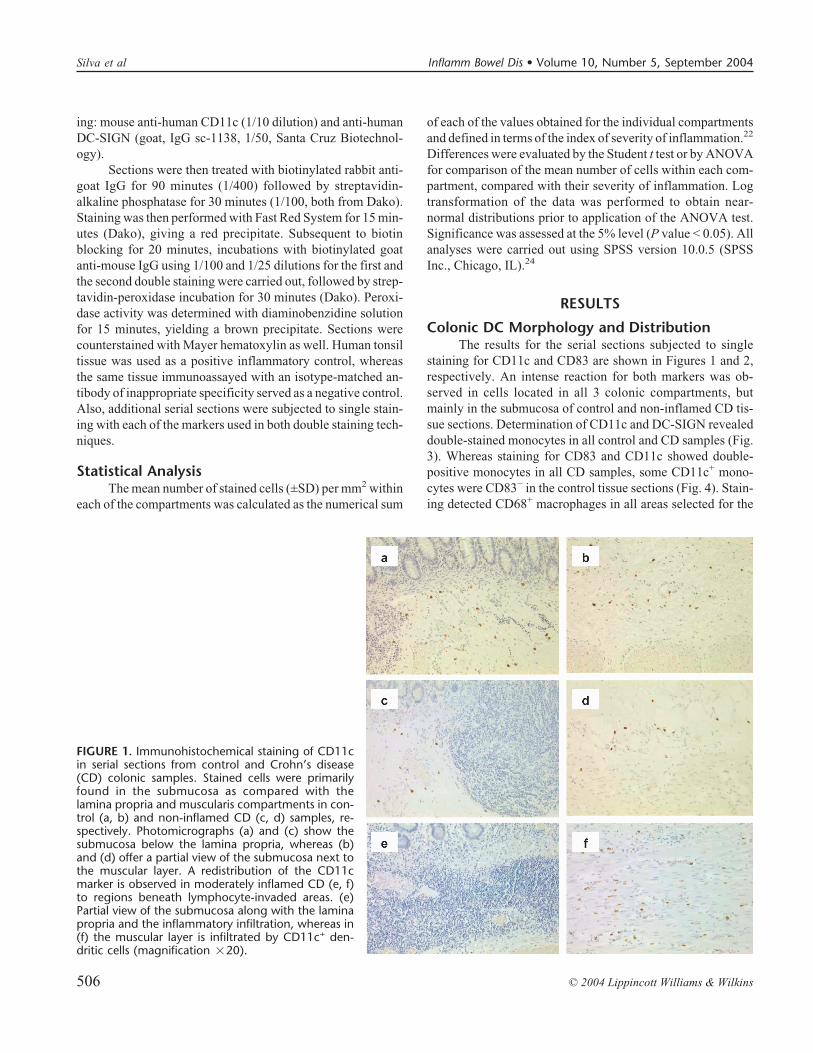
FIGURE 1. Immunohistochemical staining of CD11cin serial sections from control and Crohn’s disease(CD) colonic samples. Stained cells were primarilyfound in the submucosa as compared with thelamina propria and muscularis compartments in con-trol (a, b) and non-inflamed CD (c, d) samples, re-spectively. Photomicrographs (a) and (c) show thesubmucosa below the lamina propria, whereas (b)and (d) offer a partial view of the submucosa next tothe muscular layer. A redistribution of the CD11cmarker is observed in moderately inflamed CD (e, f)to regions beneath lymphocyte-invaded areas. (e)Partial view of the submucosa along with the laminapropria and the inflammatory infiltration, whereas in(f) the muscular layer is infiltrated by CD11c+ den-dritic cells (magnification �20).
Silva et al Inflamm Bowel Dis • Volume 10, Number 5, September 2004
506 © 2004 Lippincott Williams & Wilkins
study in both CD and control samples. CD68+ macrophageswere mainly found in the LP, immediately beneath the surfaceepithelium, and in lymphocyte-infiltrated areas of the submu-cosa in mildly and moderately inflamed CD samples (Fig. 5).
CD68+ macrophages were uniformly CD11c+ and DC-SIGN−
(data not shown).Control samples were characterized by the presence of both
CD11c+CD83+CD68−DC-SIGN+ and CD11c+CD83−CD68−DC-
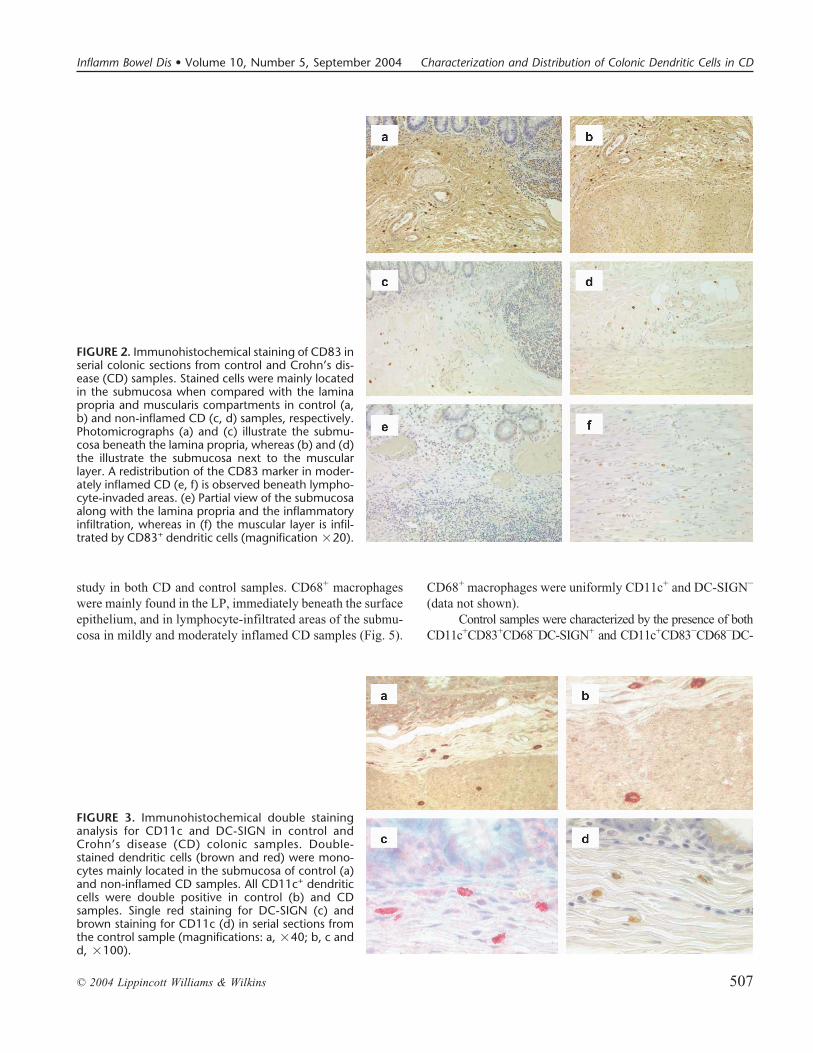
FIGURE 2. Immunohistochemical staining of CD83 inserial colonic sections from control and Crohn’s dis-ease (CD) samples. Stained cells were mainly locatedin the submucosa when compared with the laminapropria and muscularis compartments in control (a,b) and non-inflamed CD (c, d) samples, respectively.Photomicrographs (a) and (c) illustrate the submu-cosa beneath the lamina propria, whereas (b) and (d)the illustrate the submucosa next to the muscularlayer. A redistribution of the CD83 marker in moder-ately inflamed CD (e, f) is observed beneath lympho-cyte-invaded areas. (e) Partial view of the submucosaalong with the lamina propria and the inflammatoryinfiltration, whereas in (f) the muscular layer is infil-trated by CD83+ dendritic cells (magnification �20).
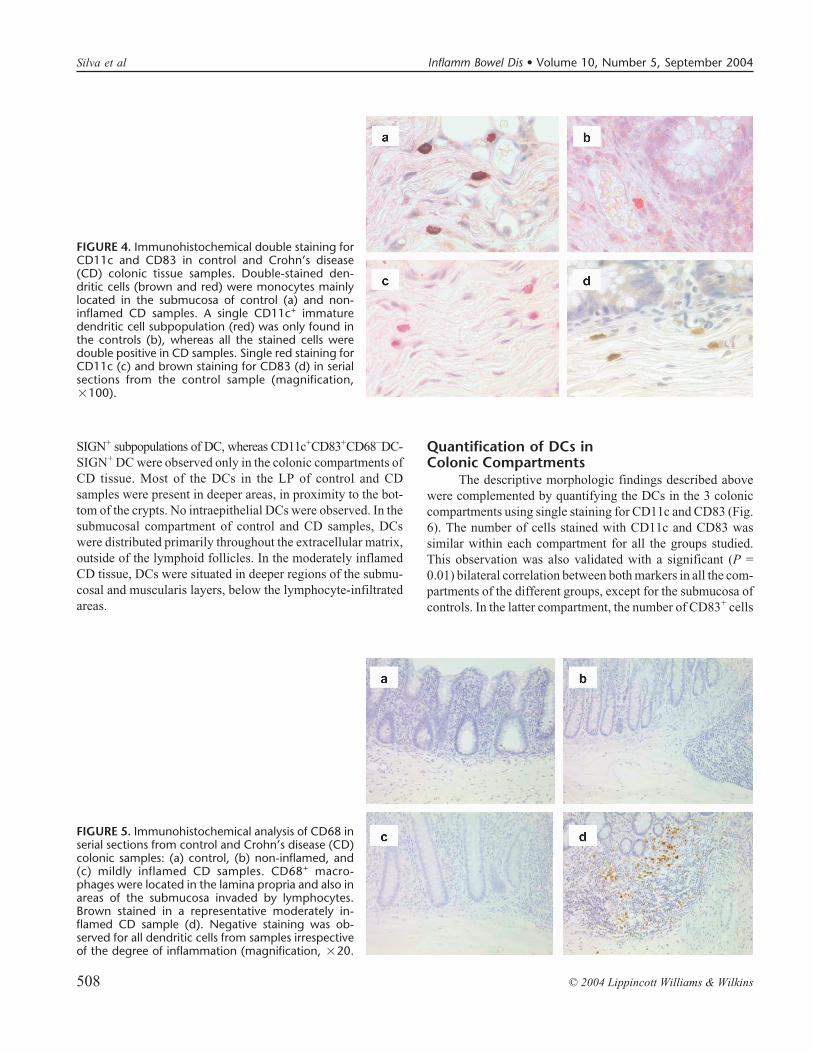
FIGURE 3. Immunohistochemical double staininganalysis for CD11c and DC-SIGN in control andCrohn’s disease (CD) colonic samples. Double-stained dendritic cells (brown and red) were mono-cytes mainly located in the submucosa of control (a)and non-inflamed CD samples. All CD11c+ dendriticcells were double positive in control (b) and CDsamples. Single red staining for DC-SIGN (c) andbrown staining for CD11c (d) in serial sections fromthe control sample (magnifications: a, �40; b, c andd, �100).
Inflamm Bowel Dis • Volume 10, Number 5, September 2004 Characterization and Distribution of Colonic Dendritic Cells in CD
© 2004 Lippincott Williams & Wilkins 507
SIGN+ subpopulations of DC, whereas CD11c+CD83+CD68−DC-SIGN+ DC were observed only in the colonic compartments ofCD tissue. Most of the DCs in the LP of control and CDsamples were present in deeper areas, in proximity to the bot-tom of the crypts. No intraepithelial DCs were observed. In thesubmucosal compartment of control and CD samples, DCswere distributed primarily throughout the extracellular matrix,outside of the lymphoid follicles. In the moderately inflamedCD tissue, DCs were situated in deeper regions of the submu-cosal and muscularis layers, below the lymphocyte-infiltratedareas.
Quantification of DCs inColonic Compartments
The descriptive morphologic findings described abovewere complemented by quantifying the DCs in the 3 coloniccompartments using single staining for CD11c and CD83 (Fig.6). The number of cells stained with CD11c and CD83 wassimilar within each compartment for all the groups studied.This observation was also validated with a significant (P =0.01) bilateral correlation between both markers in all the com-partments of the different groups, except for the submucosa ofcontrols. In the latter compartment, the number of CD83+ cells
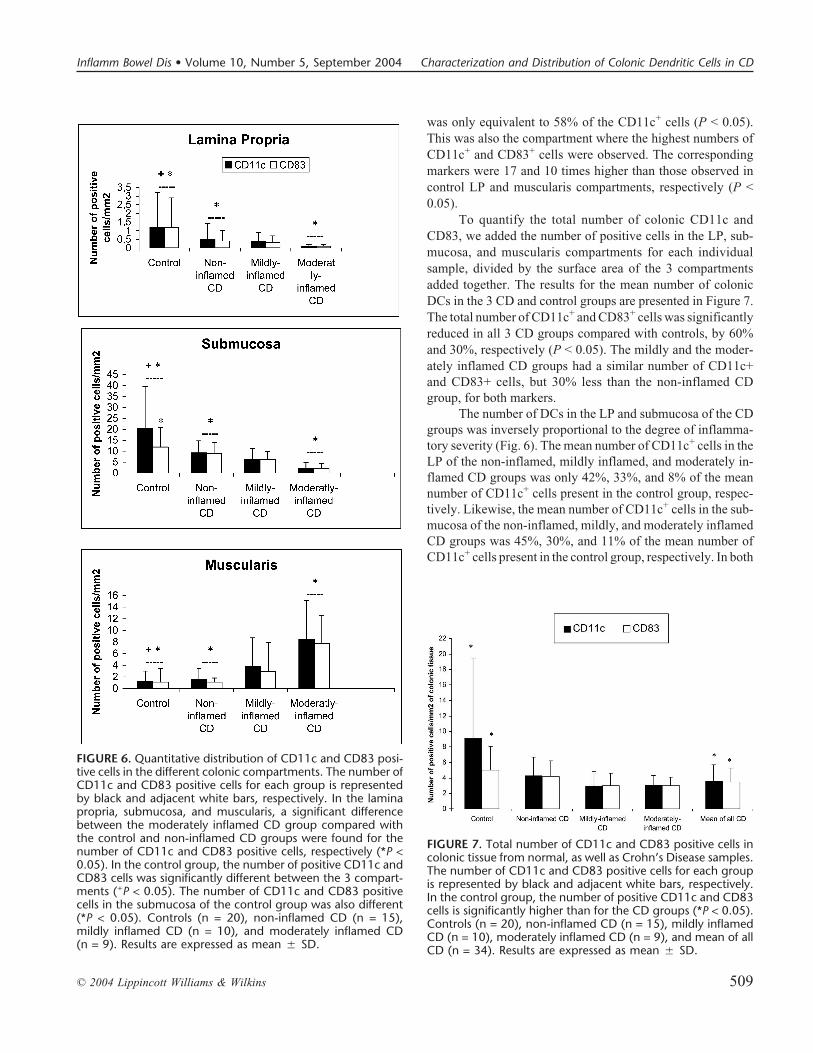
FIGURE 4. Immunohistochemical double staining forCD11c and CD83 in control and Crohn’s disease(CD) colonic tissue samples. Double-stained den-dritic cells (brown and red) were monocytes mainlylocated in the submucosa of control (a) and non-inflamed CD samples. A single CD11c+ immaturedendritic cell subpopulation (red) was only found inthe controls (b), whereas all the stained cells weredouble positive in CD samples. Single red staining forCD11c (c) and brown staining for CD83 (d) in serialsections from the control sample (magnification,�100).
FIGURE 5. Immunohistochemical analysis of CD68 inserial sections from control and Crohn’s disease (CD)colonic samples: (a) control, (b) non-inflamed, and(c) mildly inflamed CD samples. CD68+ macro-phages were located in the lamina propria and also inareas of the submucosa invaded by lymphocytes.Brown stained in a representative moderately in-flamed CD sample (d). Negative staining was ob-served for all dendritic cells from samples irrespectiveof the degree of inflammation (magnification, �20.
Silva et al Inflamm Bowel Dis • Volume 10, Number 5, September 2004
508 © 2004 Lippincott Williams & Wilkins
was only equivalent to 58% of the CD11c+ cells (P < 0.05).This was also the compartment where the highest numbers ofCD11c+ and CD83+ cells were observed. The correspondingmarkers were 17 and 10 times higher than those observed incontrol LP and muscularis compartments, respectively (P <0.05).
To quantify the total number of colonic CD11c andCD83, we added the number of positive cells in the LP, sub-mucosa, and muscularis compartments for each individualsample, divided by the surface area of the 3 compartmentsadded together. The results for the mean number of colonicDCs in the 3 CD and control groups are presented in Figure 7.The total number of CD11c+ and CD83+ cells was significantlyreduced in all 3 CD groups compared with controls, by 60%and 30%, respectively (P < 0.05). The mildly and the moder-ately inflamed CD groups had a similar number of CD11c+and CD83+ cells, but 30% less than the non-inflamed CDgroup, for both markers.
The number of DCs in the LP and submucosa of the CDgroups was inversely proportional to the degree of inflamma-tory severity (Fig. 6). The mean number of CD11c+ cells in theLP of the non-inflamed, mildly inflamed, and moderately in-flamed CD groups was only 42%, 33%, and 8% of the meannumber of CD11c+ cells present in the control group, respec-tively. Likewise, the mean number of CD11c+ cells in the sub-mucosa of the non-inflamed, mildly, and moderately inflamedCD groups was 45%, 30%, and 11% of the mean number ofCD11c+ cells present in the control group, respectively. In both
FIGURE 6. Quantitative distribution of CD11c and CD83 posi-tive cells in the different colonic compartments. The number ofCD11c and CD83 positive cells for each group is representedby black and adjacent white bars, respectively. In the laminapropria, submucosa, and muscularis, a significant differencebetween the moderately inflamed CD group compared withthe control and non-inflamed CD groups were found for thenumber of CD11c and CD83 positive cells, respectively (*P <0.05). In the control group, the number of positive CD11c andCD83 cells was significantly different between the 3 compart-ments (+P < 0.05). The number of CD11c and CD83 positivecells in the submucosa of the control group was also different(*P < 0.05). Controls (n = 20), non-inflamed CD (n = 15),mildly inflamed CD (n = 10), and moderately inflamed CD(n = 9). Results are expressed as mean � SD.
FIGURE 7. Total number of CD11c and CD83 positive cells incolonic tissue from normal, as well as Crohn’s Disease samples.The number of CD11c and CD83 positive cells for each groupis represented by black and adjacent white bars, respectively.In the control group, the number of positive CD11c and CD83cells is significantly higher than for the CD groups (*P < 0.05).Controls (n = 20), non-inflamed CD (n = 15), mildly inflamedCD (n = 10), moderately inflamed CD (n = 9), and mean of allCD (n = 34). Results are expressed as mean � SD.
Inflamm Bowel Dis • Volume 10, Number 5, September 2004 Characterization and Distribution of Colonic Dendritic Cells in CD
© 2004 Lippincott Williams & Wilkins 509

compartments, significant differences were found between themoderately inflamed CD group compared with the control andnon-inflamed CD groups (P < 0.05). The same profile of dif-ferences was observed for the distribution of CD83+ cells inthe LP and submucosa layers (Fig. 6). In the muscularis layer,the number of DCs increased gradually in parallel with the se-verity of inflammation; this pattern was the reverse of the onedescribed for the LP and submucosal compartments. The meannumber of CD11c+ cells in the non-inflamed, mildly inflamed,and moderately inflamed CD groups was 133%, 317%, and700% of the mean number of CD11c+ cells in the controlgroup, respectively. Significant differences between the mod-erately inflamed CD group compared with the control and non-inflamed CD groups were found (P < 0.05). The same patternof results was observed in the CD83+ cells (Fig. 6).
Influence of Treatment on Colonic DCsWe also examined the effect of drug treatments on the
CD colonic samples, classifying the results for the total num-ber of colonic CD11c+ and CD83+ DCs into 3 therapeutic cat-egories: tissue from CD patients without treatment (n = 6),tissue from CD patients with drugs other than corticosteroids(n = 11), and tissue from CD patients on corticosteroid therapy(n = 16). The total number of CD11c+ and CD83+ cells in thefirst (4.3 ± 2.8 and 4.9 ± 3.7) and second (4.7 ± 2.1 and 5.0 ±2.5) categories were similar, respectively. However, for thethird category of CD patients on corticosteroids, the numberof CD11c+ and CD83+ cells (2.8 ± 1.3 and 2.9 ± 1.3) wassignificantly decreased compared with the other categories(P < 0.002). The same analysis was carried out for the non-inflamed and moderately inflamed CD groups. However, in-sufficient numbers of untreated patients were available foranalysis. In the non-inflamed CD group, the total number ofCD11c+ and CD83+cells for the noncorticosteroid (6.5 ± 1.9and 6.2 ± 1.2; n = 6) and corticosteroid therapy groups (2.9 ±1.1 and 3.0 ± 0.7; n = 7) were significantly different (P < 0.05).For the moderately inflamed CD group, the total numberof CD11c+ and CD83+ cells for the noncorticosteroid (3.2 ±0.2 and 3.6 ± 0.9; n = 3) and corticosteroid treated cases (2.5 ±1.0 and 2.3 ± 0.9; n = 4) did not reach statistical significance.Comparing the non-inflamed and moderately inflamed CDgroups, a decrease in the total number of CD11c+ andCD83+cells was observed in both moderately inflamed therapygroups, especially in the noncorticoid therapy group (P <0.005).
DISCUSSIONDespite increasing evidence for the importance of DCs
in T cell tolerance and mucosal immune response,3 their role inthe pathogenesis of IBD remains poorly understood.25 In thisstudy we report, for the first time, alterations in the distributionof DCs throughout the colonic wall in tissue obtained fromresections in pediatric CD patients compared with control
specimens. Previous studies have focused on DCs in the LP ofsuperficial colonic mucosal biopsies obtained in adult pa-tients.10–20 Although the LP is a polarized compartment whereintestinal immune responses are believed to be initiated, littleattention has been focused on the submucosal compartment aspart of an integrated intestinal immune system. In this study,we observed that colonic DCs were primarily located in thesubmucosa of resected tissue obtained from control and non-inflamed CD groups. These findings suggest that DCs in thesubmucosal compartment likely play a key regulatory role inthe intestinal immune system, and that potential aberrant regu-latory function of submucosal DCs may be involved in thepathogenesis of CD.
Early reports on human intestinal DCs were limited tothose in the LP from IBD patients.10–11 The functional charac-teristics of intestinal DCs were subsequently reported in nor-mal and IBD colonic biopsies.12–13 DCs are generally founddeep in the LP compartment, beside the basement membraneof the crypts,18,19,23 as observed herein. They have been iso-lated from the colon of normal and CD adult patients and char-acterized as immature CD11c+ with low levels of CD83 andCD86 expression.14 These findings resemble those observed inour CD and control groups, where CD11c+CD83+DC werefound, but differ with respect to our CD11c+CD83−DC popu-lation found in controls. The isolation procedure used to ex-tract DCs could have influenced the expression of cell surfacemarkers and their maturation status. Also, the immunohisto-chemical methods we employed cannot differentiate betweenhigh or low levels of CD83 expression. Thus, we could notestablish if our CD83+DC were low CD83+ immature, highCD83+ mature, or semi-mature DCs. A subsequent report inadult patients described a mature CD83+DC-SIGN− DC popu-lation in normal colon and an immature CD83−DC-SIGN+ DCpopulation in colonic samples from patients with CD.26 How-ever, DCs were not identified on the basis of CD11c expres-sion, and both markers identified different cellular popula-tions, in contrast to their in vitro results.26 In the current study,CD11c and DC-SIGN were found to co-localize to the sameDC population, and the absence of CD83 defined an immatureDC subpopulation. It is noteworthy that our study is based on aselected subgroup of CD patients that required colonic resec-tion. We demonstrate for the first time in a pediatric CD studythat a CD11c+CD83+CD68−DC-SIGN+ subpopulation of DCsis present in all 3 compartments of the colonic wall. Thesefindings cannot easily be compared with other DC studies inCD because of the disparate ages of the patient groups and thedifferent experimental strategies used.
Chronic inflammation characterizes the immunologicand histopathological profile of the tissues involved in CD. Weevaluated the mucosa inflammatory status of our CD samplesby using a recently established semi-quantitative index thatconsiders the 6 principal histopathological features that char-acterize CD.22 We found that the total number of DCs was
Silva et al Inflamm Bowel Dis • Volume 10, Number 5, September 2004
510 © 2004 Lippincott Williams & Wilkins

significantly reduced in CD compared with control tissue. Thenumber of DCs was less in inflamed tissue compared with non-inflamed CD specimens (Fig. 7). Moreover, with increasingseverity of inflammation, DCs disappeared from the LP andthe submucosal compartments and tended to concentrate in themuscularis layer (Fig. 6). Significant quantitative differencesbetween the non-inflamed and the moderately inflamed CDgroups support our observations. In view of the fact that in-flammation is a continuous process, the location of the mildlyinflamed group between the two others would result in someoverlap, making it difficult to find any statistically significantdifference.
To evaluate the potential effects of drug therapy on DCdistribution in our CD samples, we analyzed the data accord-ing to 3 variables: no medical treatment, immunomodulatorytherapy without corticosteroids, and corticosteroid therapy. Nodifference was observed in DC distribution between thesamples from patients with noncorticosteroid and those with-out therapy. However, a statistically significant decrease in DCnumber was found in the samples from patients on corticoste-roids compared with the nonsteroid therapy group. Moreover,subgroup analysis revealed that the differences between bothgroups were statistically significant for the non-inflamed CDgroup. In the moderately inflamed CD group, the differencesdid not reach statistical significance. Also, for both groups oftherapy, the number of DCs was less in the moderately in-flamed group than in the non-inflamed one, reaching statisticalsignificance for the noncorticoid therapy group. Our data sug-gests that DC depletion induced by corticosteroid therapycould represent a potential pharmacological mechanism of im-munosuppression in CD.20 Overall, however, our data showthat in pediatric CD under the distribution of colonic DCs, aswell as their number, changes with different degrees of inflam-mation, irrespective of drug therapy.
In the current investigation, we observed both immatureCD11c+CD83−CD68−DC-SIGN+ and CD11c+CD83+CD68−DC-SIGN+ DC subpopulations in control tissue. In colonic tissuefrom patients with CD, the immature subpopulation of DCswas absent and the CD11c+CD83−CD68−DC-SIGN+ subpop-ulation partially reduced, even for the non-inflamed subgroup.This observation may be a consequence of the effect of thecorticosteroid therapy in CD. It is also possibly be due to animmunologic imbalance of the DC population in CD, becausethese findings were also valid for the samples without treat-ment and for the noncorticoid therapy group. In the normalintestine, all the subpopulations of DCs maintain a regulatedstate of homeostatic immune activation. When the intestinalsubpopulations of DCs are altered by depletion, immunologicimbalance likely ensues. This could render the intestine moresensitive to a deficient regulatory response and/or an excessiveeffector cell response when the immune system is challengedand activated. Overall, our data demonstrates that in CD, thedistribution of colonic DCs, as well as their number, changes
with different degrees of inflammation. We hypothesize thatthe depletion of DCs is responsible, in part, for the loss ofphysiological immune tolerance toward the host bacterial florathat has observed in CD.25–29 We did not find any difference interms of the distribution and marker expression of DCs in thedifferent parts of the colon. However, our groups were not per-fectly homogeneous in that the control tissue consisted mainlyof distal colonic samples, whereas the CD group was mainlybased on more proximal colonic samples. Also, our sampleswere not perfectly age-matched, due to difficulties in finding anon-inflamed control group.
In conclusion, we describe herein the distribution of DCsthroughout the compartments of the colon in pediatric CD pa-tients with varying degrees of inflammation severity, as com-pared with controls. We demonstrate that DCs are predominantlylocated in the submucosa in control tissue, where both immatureCD11c+CD83−CD68−DC-SIGN+ and CD11c+CD83+CD68−DC-SIGN+ DC phenotypes were detected. We observed variationsin the quantity and localization of DCs, correlating with theseverity of colonic inflammation in CD and also with drugtherapy. Our data imply that corticosteroid therapy leads todepletion of DCs in colonic CD samples. Our observationsalso suggest that the presence of different subpopulations ofcolonic DCs could be implicated in the homeostatic regulationof physiological intestinal immune activation to the host flora.Perturbation of the intestinal DC subpopulations in CD withthe loss of immature CD11c+CD83−CD68−DC-SIGN+DC anda reduction of CD11c+CD83+CD68−DC-SIGN+DC may causean immunologic imbalance that could be relevant to the patho-genesis of CD.
ACKNOWLEDGMENTSThe authors thank Roxanne Gendron for her assistance
with statistical analyses, Drs Devendra Amre and ErnestoGuiraldes for their critical review, and Sylvie Marineau forassistance in the preparation of this manuscript.
REFERENCES1. Bouma G, Strober W. The immunological and genetic basis of inflamma-
tory bowel disease. Nat Rev Immunol. 2003;3:521–533.2. Mowat A. Anatomical basis of tolerance and immunity to intestinal anti-
gens. Nat Rev Immunol. 2003;3:331–341.3. Morel PA, Feili-Hariri M, Coates PT, et al. Dendritic cells, T cell toler-
ance and therapy of adverse immune reactions. Clin Exp Immunol. 2003;133:1–10.
4. Lutz M, Schuler G. Immature, semi-mature and fully mature dendriticcells: which signals induce tolerance or immunity? Trends Immunol.2002;23:445–449.
5. Geijtenbeek T, Torensma R, van Vliet S, et al. Identification of DC-SIGN,a novel dendritic cell-specific ICAM-3 receptor that supports primary im-mune responses. Cell. 2000;100:575–585.
6. Steinman R, Pack M, Inaba K. Dendritic cells in the T-cell areas of lym-phoid organs. Immunol Rev. 1997;156:25–37.
7. Zhou L, Tedder T. Human blood dendritic cells selectively express CD83,a member of the immunoglobulin superfamily. J Immunol. 1995;154:3821–3835.
Inflamm Bowel Dis • Volume 10, Number 5, September 2004 Characterization and Distribution of Colonic Dendritic Cells in CD
© 2004 Lippincott Williams & Wilkins 511

8. Salto-Tellez M, Price AB. What is the significance of muciphages in co-lorectal biopsies? The significance of muciphages in otherwise normalcolorectal biopsies. Histopathology. 2000;36:556–562.
9. Falini B, Flenghi L, Pileri S, et al. PG-M1: a new monoclonal antibodydirected against a fixative-resistant epitope on the macrophage-restrictedform of the CD68 molecule. Am J Pathol. 1993;142:1359–1372.
10. Wilders M, Drexhage H, Kokjé M, et al. Veiled cells in chronic idiopathicinflammatory bowel disease. Clin Exp Immunol. 1984;55:377–387.
11. Wilders M, Drexhage H, Kokjé M, et al. Peripolesis followed by cytotox-icity in chronic idiopathic inflammatory bowel disease. Clin Exp Immu-nol. 1984;58:614–620.
12. Mahida Y, Wu K, Jewell D. Characterization of antigen-presenting activ-ity of intestinal mononuclear cells isolated from normal and inflammatorybowel disease colon and ileum. Immunology. 1988;65:543–549.
13. Pavli P, Hume DA, Van de Pol E, et al. Dendritic cells, the major antigen-presenting cells of the human colonic lamina propria. Immunology. 1993;78:132–141.
14. Bell S, Rigby R, English N, et al. Migration and maturation of humancolonic dendritic cells. J Immunol. 2001;166:4958–4967.
15. Ikeda Y, Akbar F, Matsui H, et al. Characterization of antigen-presentingdendritic cells in the peripheral blood and colonic mucosa of patients withulcerative colitis. Eur J Gastroenterol Hepatol. 2001;13:841–850.
16. Vuckovic S, Florin T, Khalil D, et al. CD40 and CD86 up-regulation withdivergent CMRF44 expression on blood dendritic cells in inflammatorybowel disease. Am J Gastroenterol. 2001;96:2946–2956.
17. Murakami H, Akbar F, Matsui H, et al. Macrophage migration inhibitoryfactor activates antigen-presenting dendritic cells and induces inflamma-tory cytokines in ulcerative colitis. Clin Exp Immunol. 2002;128:504–510.
18. Pavli P, Maxwell L, van de Pol E, et al. Distribution of human colonicdendritic cells and macrophages. Clin Exp Immunol. 1996;104:124–132.
19. Wilders M, Sminia T, Plesch B, et al. Large mononuclear Ia-positiveveiled cells in the Peyer’s patches. Immunology. 1983;48:461–467.
20. de Baey A, Mende I, Baretton G, et al. A subset of human dendritic cell inthe T cell area of mucosa-associated lymphoid tissue with a high potentialto produce TNF-�. J Immunol. 2003;170:5089–5094.
21. Seidman EG. Inflammatory bowel diseases. In: Roy C, Silverman A, Ala-gille D, eds. Pediatric Gastriontestinal Disease. 4th ed. St. Louis: Mosby;1996:435–483
22. Silva M, Menezes J, Wizman S, et al. Cytokine tissue levels as markers ofdisease activity in pediatric Crohn’s disease. Pediatr Res. 2003;54:456–461.
23. Nagashima R, Maeda K, Imai Y, et al. Lamina propria macrophages in thehuman gastrointestinal mucosa: their distribution, immunohistologicalphenotype, and function. J Histochem Cytochem. 1996;44:721–731.
24. Neter J, Kutner M, Nachtsheim C, Wasserman R. Edited by Irwin bookteam. Applied Linear Statistical Models. Chicago: McGraw-Hill; 1996
25. Stagg AJ, Hart AL, Knight SC, et al. The dendritic cell: its role in intes-tinal inflammation and relationship with gut bacteria. Gut. 2003;52:1522–1529.
26. Te Velde A, van Kooyk Y, Braat H, et al. Increased expression of DC-SIGN+IL-12+IL-18+ and CD83+IL-12-IL-18- dendritic cell populationsin the colonic mucosa of patients with Crohn’s disease. Eur J Immunol.2003;33:143–151.
27. Duchmann R, Kaiser I, Hermann E, et al. Tolerance exists towards resi-dent intestinal flora but is broken in active inflammatory bowel disease(IBD). Clin Exp Immunol. 1995;102:448–455.
28. Landers C, Cohavy O, Misra R, et al. Selected loss of tolerance evidencedby Crohn’s disease-associated immune responses to auto- and microbialantigens. Gastroenterology. 2002;123:689–699.
29. Sydora B, Tavernini M, Wessler A, et al. Lack of interleukin-10 leads tointestinal inflammation, independent of the time at which luminal micro-bial colonization occurs. Inflamm Bowel Dis. 2003;9:87–97.
Silva et al Inflamm Bowel Dis • Volume 10, Number 5, September 2004
512 © 2004 Lippincott Williams & Wilkins





























