Embed Size (px)
Citation preview

Research Collection
Doctoral Thesis
Cereal β-glucan processing for improved molecular interactions
Author(s): Marasca, Elena
Publication Date: 2019
Permanent Link: https://doi.org/10.3929/ethz-b-000382795
Rights / License: Creative Commons Attribution 4.0 International
This page was generated automatically upon download from the ETH Zurich Research Collection. For moreinformation please consult the Terms of use.
ETH Library

Diss. ETH N° 26279
Cereal β-glucan processing for improved
molecular interactions
A thesis submitted to attain the degree of
DOCTOR OF SCIENCES of ETH ZURICH
(Dr. sc. ETH Zurich)
presented by
Elena Marasca
MSc Chemistry, Università degli Studi di Torino
born on 08.06.1984
citizen of Italy
accepted on the recommendation of
Prof. Dr. Laura Nyström, examiner
Prof. Dr. Costas Biliaderis, co-examiner
Dr. Samy Boulos, co-examiner
2019

2

3
For Ginevra,
may you one day be proud of your Mum.

4

5
ABSTRACT
Cereal (1→3),(1→4)-β-D-glucan, a soluble dietary fiber found predominantly in oat and barley,
is a well-known contributor to several health promoting properties, such as the reduction of
serum cholesterol and the attenuation of postprandial glucose response. These effects have often
been associated to its high molecular weight (Mw) and its ability to form highly viscous
solutions in the gastrointestinal tract. It is well known that the chemical structure of β-glucan
varies among different cereal sources and that common food processing, such as milling, baking
and extrusion can affect its molecular structure and, in turn, its health and technological
properties. In particular processing can cause molecular degradation, which leads to a decrease
in Mw and, as a consequence, to loss of viscosity. Furthermore, oxidative degradation as a result
of hydroxyl radical attack has recently been established as a mechanism that modifies β-glucan,
especially in aqueous systems. Oxidation can cause chain cleavage in the polymer backbone or
oxidation of any of the hydroxyl groups of the glucose monomers, leading to the formation of
carbonyl or carboxyl groups, or even to a ring opening. However, there is still a lack of
information on how this variability in structure impacts on the biological value of β-glucan. The
aim of this thesis was the investigation of the changes caused to the β-glucan structure by
processing and the clarification of the relationship between processed β-glucan and its
physicochemical properties in food and in the gastrointestinal tract.
Recent literature has shown that oxidized and low Mw cereal β-glucan has better bile acid
binding capacity, i.e. stronger cholesterol reduction effect, compared to the native counterparts.
We investigated this by studying the kinetics of passage of a bile salt mix across a dialysis
membrane, in the presence or absence of native and modified (TEMPO or sodium periodate
oxidized or hydrolyzed) oat and barley β-glucan extracts, simulating the mechanism of
diffusion through the unstirred water layer of the small intestine. The results showed that native
oat and barley β-glucan extracts, which were also the most viscous ones, had the strongest bile
acid binding power, with oat β-glucan exerting a stronger effect than barley. Despite their lower
viscosity, TEMPO oxidized and acid hydrolyzed oat and barley β-glucan were also quite
effective in retaining the bile acids. However, in contrast with what is suggested in the literature,
oxidation and reduction of Mw did not increase the bile acid binding ability of the fibers.
Cereals can potentially be good sources of minerals such as iron. β-Glucan, however, is often
found in association with metal ions in cereals, and therefore its presence can reduce the
bioavailability of iron. In the past, a direct association between β-glucan and iron was generally

6
accepted. Recents reports in literature, however, have shown that the main culprit for the
reduction of iron bioavailavility is phytic acid, which is often found in association with dietary
fibers and which can form a strong complex with iron rendering it unavailable for absorption.
However, since processing can alter the molecular structure of β-glucan, the equilibrium
between this fiber, phytic acid and iron can be affected. In our study, we tried to clarify the
relationship between the structural properties of cereal β-glucan and its ability to bind iron,
focusing also on the role of phytic acid. Different treatments were used to modify the β-glucan
structure in a controlled way, namely TEMPO oxidation, sodium periodate (NaIO4) oxidation
and acid hydrolysis. The native and modified fibers where subsequently characterized and
subjected to an in vitro digestion in the presence of iron, with or without prior enzymatic
dephytinization. Through a reverse dialysis system, soluble iron (i.e. available for absorption)
was measured by atomic absorption spectrometry. The results obtained showed that no direct
binding between β-glucan and iron exists, confirming the major role of phytic acid as an
inhibitor of iron absorption in cereal foods.
The structure of cereal β-glucan can undergo several alterations during food processing, which
can reduce its Mw and viscosity. Also, hydroxyl-mediated oxidative degradation, linked to
Fenton chemistry, has been shown to take place already during storage of cereal β-glucan
solutions. In our study, we examined the effects of incubation at high temperature (125°C and
175°C) and extrusion cooking (120°C and 150°C, 25% moisture content) on the molecular
structure of β-glucan extracted from oat and barley flour. The treated extracts were
characterized by Mw and viscosity measurements, followed by investigation of oxidation
products by labelling of the carbonyl groups and analysis by UPLC-MS with fluorescent
detection. After the thermal treatments, a significant decrease in viscosity and Mw was recorded,
together with the formation of new reducing ends and cross-ring cleavage oxidation products,
with a more dramatic effect at higher temperature, confirming the hydroxyl radical induced
depolymerization of the fiber. For the extruded samples, no significant change in viscosity and
Mw took place in the case of oat β-glucan, while viscosity had a dramatic increase for barley,
in particular for the samples extruded at 120°C. No conclusive results in terms of the oxidation
products formed was reached for these extruded samples, therefore further investigation will be
required to clarify this process.
This study helped to shed some light on important aspects of the cereal β-glucan structure-
function relationship. In particular, it was shown that no direct binding between bile acids and
β-glucan exists and that the attenuation of serum cholesterol is due to the viscosity effects of

7
the β-glucan solutions. Also, the crucial role of phytic acid in the iron binding effect in the
presence of cereal β-glucan was confirmed, showing that no direct link between fiber and iron
exists. Finally, this study helped moving the first steps towards a more thorough
characterization of β-glucan in food, which could help in the future to choose the best
processing conditions to ensure maximum health effects.

8

9
ABSTRACT (Italian version) I (1→3),(1→4)-β-D-glucani di origine cerealicola sono fibre alimentari solubili, presenti
principalmente in orzo e avena, e sono noti per le loro proprietà benefiche per la salute, come
la riduzione del colesterolo nel siero e l’attenuazione della risposta glicemica postprandiale.
Questi effetti sono stati spesso associati al loro alto peso molecolare (Mw) e alla capacità di
queste fibre di formare soluzioni ad alta viscosità nel tratto gastrointestinale. È noto che la
struttura chimica dei β-glucani vari a seconda dei cereali e che le comuni tecniche di lavorazion
degli alimenti quali la cottura in forno, la macinatura e l’estrusione, possano portare a
un’alterazione della loro struttura molecolare e, di conseguenza, a una modifica delle loro
proprietà salutari e tecnologiche. In particolare, può causare degradazione molecolare, che porta
ad una diminuzione del Mw e, di conseguenza, a una perdita di viscosità. Inoltre, la degradazione
ossidativa dovuta all'attacco dei radicali idrossilici è stata dimostrata come il meccanismo
responsabile dell’alterazione dei β-glucani, in particolare in soluzione. L'ossidazione può
causare la scissione della catena polimerica o l'ossidazione di uno qualsiasi dei gruppi ossidrilici
dei monomeri di glucosio, portando alla formazione di gruppi carbonilici o carbossilici o anche
all'apertura dell’anello. Tuttavia, mancano ancora informazioni dettagliate su come questa
variabilità nella struttura influenzi il valore biologico dei β-glucani. Lo scopo di questa tesi è
stato quello di esaminare la struttura dei β-glucani per chiarire la relazione tra il processing e le
loro proprietà fisico-chimiche negli alimenti e nel tratto gastrointestinale.
La letteratura recente ha dimostrato che i β-glucani ossidati e a basso peso molecolare hanno
una migliore capacità legante degli acidi biliari, cioè un maggiore effetto di riduzione del
colesterolo, rispetto alle fibre non modificate. Abbiamo investigato questo aspetto stuiando la
cinetica del passaggio di una miscela di sali biliari attraverso una membrana semipermeabile,
in presenza o assenza di β-glucani estratti da avena e orzo, allo stato naturale o modificati
(ossidati tramite TEMPO o periodato di sodio oppure idrolizzati), simulando il meccanismo di
diffusione attraverso lo strato di acqua non agitato dell'intestino tenue. Secondo i nostri risultati,
i β-glucani non modificati, dotati anche della viscosità più alta, hannoilpoterelegantepiù
forte,inparticolarequelliestrattidall’avena.Nonostantelalorobassaviscositàridotta,
ancheiβ-glucaniossidaticonTEMPOeidrolizzatisonorisultatiefficacineltratteneregli
acidibiliari.Tuttavia,contrariamenteaquantosuggeritoinletteratura,l'ossidazioneela
riduzionediMwnonaumentanoilpoterediritenzionedegliacidibiliaridapartediqueste
fibre.

10
Icerealisonopotenzialmentebuonefontidimineralicomeilferro.Iβ-glucani,tuttavia,si
trovanospessoinassociazioneconionimetallicineicereali,equindilaloropresenzapuò
ridurre la biodisponibilità del ferro. In passato si pensava esistesse un’associazione
diretta traβ-glucani e ferro.Tuttavia, studi recentihannodimostrato che il principale
responsabiledellariduzionedellabiodisponibilitàdel ferroè l'acidofitico,chesitrova
spessoinassociazioneconlefibrealimentariechepuòformareuncomplessoconilferro,
rendendolo non disponibile per l'assorbimento. Poiché la lavorazione dei cereali può
alterarelastrutturamolecolaredeiβ-glucani,l'equilibriotraquestafibra,l'acidofiticoe
ilferropuòrisentirne.Nelnostrostudio,abbiamocercatodichiarirelarelazionetrale
proprietàstrutturalideiβ-glucanidioriginecerealicolae lorosuacapacitàdi legare il
ferro,concentrandocianchesulruolodell'acidofitico.Diversitrattamentisonostatiusati
permodificare la strutturadeiβ-glucani inmodocontrollato, vale adire l'ossidazione
tramiteTEMPO, l'ossidazioneconperiodatodi sodio (NaIO4)e l'idrolisiacida.Le fibre
originali e modificate sono state successivamente caratterizzate e sottoposte a una
digestione in vitro in presenza di ferro, con o senza preventiva digestione enzimatica
dell’acidofitico.Attraversounsistemadidialisiinversa,ilferrosolubile(cioèdisponibile
perl'assorbimento)èstatomisuratomediantespettrometriadiassorbimentoatomico.I
risultatiottenutihannomostratochenonesistealcunlegamedirettotraβ-glucanieferro,
confermando il ruolo principale dell'acido fitico come inibitore dell'assorbimento del
ferroneglialimentiabasedicereali.
La struttura dei β-glucani può subire diverse alterazioni durante la lavorazione dei
cereali, che possono portare alla riduzione del loro Mw e viscosità. Inoltre, è stato
dimostratoche ladegradazioneossidativamediatadal radicale idrossile, collegataalla
chimicadiFenton,siverificagiàdurantelostoccaggioβ-glucaniinsoluzione.Nelnostro
studioabbiamoesaminatoglieffettidell'incubazioneadaltatemperatura(125°Ce175°C)
e della cottura a estrusione (120°C e 150°C, con contenuto di umidità del 25%) sulla
strutturamolecolaredeiβ-glucaniestrattidallafarinadiorzoeavena. Gliestrattitrattati
sonostaticaratterizzatitramitelamisuradipesomolecolareviscosità,oltreallostudio
deiprodottidiossidazionemediantelabellingdeigruppicarboniliciesuccessivaanalisi
con UPLC-MS con detector a fluorescenza. I trattamenti ad alta temperatura hanno
portatoaunasignificativariduzionedellaviscositàedelpesomolecolare, insiemealla
formazionedinuovigruppiriducentieprodottidiossidazioneconaperturadell’anello,

11
con un effetto più significativo a temperatura più elevata, confermando la
depolimerizzazionedellafibraindottadairadicaliidrossilici.
Pericampioniestrusi,nessunavariazionesignificativadiviscositàeMwsièverificatanel
caso dell'avena,mentre la viscosità ha avuto un drammatico aumento per i β-glucani
estratti dall’orzo. Non sono stati raggiunti risultati conclusivi per quanto riguarda
prodotti di ossidazione formati per i campioni estrusi, pertanto saranno necessarie
ulteriori indaginiperchiarirequestoprocesso.Questostudiohacontribuitoachiarire
alcuniaspetti importantidella relazionestruttura-funzionedeiβ-glucani contenutinei
cereali. Inparticolare, è statodimostrato chenonesistealcun legamediretto traacidi
biliari e β-glucani e che l'attenuazionedel colesterolonel siero èdovuta agli effetti di
viscositàdellesoluzionidiβ-glucani.Inoltre,èstatoconfermatoilruolocrucialedell'acido
fitico nell'effetto legante del ferro in presenza di β-glucani di origine cerealicola,
dimostrandochenonesistealcunlegamedirettotrafibraeferro.Infine,questostudioha
contribuitoamuovereiprimipassiversounacaratterizzazionepiùapprofonditadeiβ-
glucanineglialimenti,chepotrebbeaiutareinfuturoasceglierelemiglioricondizionidi
lavorazionepergarantireimassimieffettisullasalute.

12

13
AKNOWLEDGEMENTS Every good journey comes to an end and being at the finishing line of this incredible one at
ETH Zurich, I feel like I have to express my deepest thanks to several people, who supported
me through its up and downs and allowed me to be here to write these words.
First and foremost, I would like to warmly thank my supervisor Prof. Dr. Laura Nyström for
giving me the opportunity to do my PhD in the Food Biochemistry group. When first applying
for a position in the group in 2013, I immediately had the feeling that something was special
about her and her group. Despite not being chosen in that instance, which turned out to be the
best thing for everyone in the end, the thought of the FBC group stuck with me and Laura and
I kept in contact, until I finally got in touch with her just when she needed me and I found out
I was right. This was a special group to be part of and I will always remember the nice scientific
exchange but also the game nights, the laughter and the happiness of coming to work in this
nice atmosphere every day. Thank you Laura, thank you for you scientific and personal support,
for allowing me to have time to learn how to conciliate having a baby with finishing a PhD and
for supporting me through it all. It was great to have you a supervisor, both on a professional
and personal level.
Special thanks to Dr. Samy Boulos, I really couldn’t have done it without you, thank you for
all your support and for showing me I could do it when I was not sure of anything anymore.
Thanks for your precious guidance with the experiments, with the students and with the writing
but thank you also for being there when I needed a shoulder to cry on or someone to share my
successes with, for the nice conversations in the office and for the many little moments we had
together. You helped me more than you will ever know. I will forever be grateful to have found
you on my path and it is a great honor for me to have you as my co-examiner at my defense.
I would like to thank also Prof. Costas Biliaderis, from the Aristotle University of Thessaloniki
for being my co-examiner and for travelling all the way from Greece for my defense. Your
precious work has been the fundamental block of most of the things I have done and learnt
during my PhD, therefore it is an honor and a pleasure to have you as part of my examining
committee.
Deep thanks also to my dear colleague and friend Nadja Steiger, I cannot express how happy I
was to share this journey (and the office!!) with you. You helped me through difficult moments
and celebrated with me the happy ones and I will always cherish the many memories we have
together. I will miss you deeply but I am sure we will always be in touch.

14
Thank you to Dan Zhu, Chunyue Wei, Dr. Melanie Erzinger and Cristina Lupo for all the nice
lunch and coffee breaks, for your scientific and organizational support and in general for being
so great and so fun to work with!!!
I also want to thank the students I supervised, Viktorija Krivova, Béatrice Schmid, Meret
Allemann, Sybille Weber, Lukas Jung and, in particular Olivia Wyss. Olivia, thank you so much
for persevering in a project that turned out to be so much more challenging than originally
anticipated, thank you for not giving up despite the many Chelex purifications and all the other
“little” problems that we faced!! It was truly a pleasure to work with you, thank you for being
such a nice person and for all your restaurant recommendations and the food talk!!
I also want to thank all the other members of the group, past and present, who helped providing
a nice atmosphere and a fruitful working environment! Thank you Daniela Kalbermatter and
Aida Huber for your support in ensuring that everything run smoothly.
Ringrazio i miei genitori Mario e Giancarla per avermi permesso di arrivare fino qui, siete
sempre stati nella mia mente e nel mio cuore, nonostante la lontananza, e spero possiate essere
fieri di me. Grazie per aver sempre supportato le mie scelte, con affetto e discrezione nonostante
la sofferenza di vedermi partire.
Ringrazio Carlo per il suo immenso supporto durante questo percorso, anche nelle cose più
piccole, per essere stato la mia isola di pace e serenità nei momenti difficili e per avermi aiutato
a conciliare il fatto di essere una neomamma con le sfide di un PhD. E per aver sopportato tutte
le mie paturnie nel corso degli anni!!! Non ce l’avrei mai fatta senza di te.
Un ringraziamento meritatissimo anche alla mia piccola Ginevra, che (nonostante le notti
insonni!!!) mi ha regalato un nuovo livello di felicità che prima non conoscevo. Spero un giorno
potrai essere orgogliosa della tua mamma.

15
LIST OF ABBREVIATIONS
2-AB 2- aminobenzamide (anthranilic amide)
ACN acetonitrile
Ar aromatic ring
Ara arabinose
BA bile acid
BBG barley β-glucan
BG mixed-linkage cereal (1→3, 1→4)-β-D-glucan
BPI base peak ion chromatogram
C=O carbonyl group
DP degree of polymerization
Ery erythrose
ESI electrospray ionization
FLD fluorescent detection
Glc glucose unit
OBG oat β-glucan
•OH hydroxyl radical

16

17
CONTENTS
ABSTRACT ............................................................................................................................... 5
ABSTRACT (Italian version) ................................................................................................... 9
AKNOWLEDGEMENTS ........................................................................................................ 13
LIST OF ABBREVIATIONS ................................................................................................. 15
INTRODUCTION ................................................................................................................... 19
PART 1: REVIEW OF THE LITERATURE ........................................................................ 23
1. Cereal β-glucans .............................................................................................................. 25
1.1 Occurrence and structural characteristics ........................................................... 25
1.2 Rheology ................................................................................................................... 27
1.3 Health aspects .......................................................................................................... 28 1.3.1 Blood glucose regulation ................................................................................... 29 1.3.2 Cholesterol lowering .......................................................................................... 30 1.3.3 Additional health implications ........................................................................... 35
2. β-glucan oxidation ........................................................................................................... 37
2.1 Unselective Fenton oxidation .................................................................................. 37
2.2 Selective TEMPO oxidation ................................................................................... 42
2.3 Selective sodium periodate oxidation .................................................................... 45
3. Effect of β-glucan processing on structure and health properties ................................ 46
3.1 Chemical and enzymatic modification of β-glucan .............................................. 48 3.1.1 Oxidation ............................................................................................................ 48 3.1.2 Acidic and enzymatic hydrolysis ....................................................................... 49
3.2 Thermal and mechanical processing ..................................................................... 51
3.3 Impact of processing on β-glucan health promoting properties ......................... 54
4. Aims of the study .............................................................................................................. 67
PART 2: RESEARCH PAPERS ............................................................................................. 69
I. Bile acid binding by native and modified oat and barley β-glucan ............................... 71
II. In vitro estimation of iron binding by modified cereal β-glucan extracts ..................... 99
III. Investigation of cereal β-glucan degradation following thermal treatments and extrusion ................................................................................................................................ 127
PART 3: CONCLUSIONS & OUTLOOK ............................................................................ 165

18

19
INTRODUCTION Cereal β-glucan, a soluble dietary fiber found predominantly in oat and barley, has drawn the
attention of the scientific community in the last decades due to its beneficial health properties,
such as cholesterol lowering and blood glucose regulation, confirmed by governmental health
and food agencies (FDA, 2009; Efsa, 2011). These favorable health effects have often been
linked to the ability of this polysaccharide to form highly viscous solutions in the
gastrointestinal tract, slowing down gastric emptying and absorption of glucose as well as
inhibiting the uptake of food lipids and bile acids, preventing illnesses such as coronary heart
disease and diabetes (Lan-Pidhainy et al., 2007; Wolever et al., 2010; Wood, 2010). Since the
viscosity of the solutions formed by β-glucan depends on its molecular weight (Mw) and
concentration, the hypothesis is that the higher the Mw, the better the health properties.
Even though natural occurring cereal β-glucan possess very high Mw, food processing, such as
milling, baking, extrusion, etc., can alter β-glucan’s structural characteristics, which can lead to
a modification of the health and technological properties of the fiber. In particular, molecular
degradation can occur, lowering the Mw and, as a consequence, viscosity. Oxidative degradation
as a result of hydroxyl radical attack has recently been established to have a significant impact
on the β-glucan molecular structure, especially in aqueous systems (Kivelä et al, 2009; Kivelä
et al., 2012). Oxidation can cause chain cleavage in the polymer backbone or oxidation of any
of the hydroxyl groups of the glucose monomers, leading to the formation of carbonyl or
carboxyl groups, or even to a ring opening. However, the data available suggest that oxidation
may, in fact, enhance both β-glucan’s health benefits and technological properties (de Moura et
al., 2011; Lee et al., 2011). For example, it has been shown that oxidation can increase water
extractability by solubilizing the non-water soluble β-glucan and that the inclusion of oxidised
β-glucan in a wheat bread dough improves the softness of bread (Park et al., 2009; Lee et al.,
2011). Also, the β-glucan with low or medium Mw had a better bile acid binding capacity, linked
to the cholesterol reduction mechanism, than the high Mw ones (Kahlon and Woodruff, 2003;
Sayar et al., 2011). The same applies for oxidized β-glucan, which was shown to have a better
bile acid binding capability than the native counterpart (Park et al., 2009).
Another aspect to consider is that cereals can be a good source of minerals such as iron and
zinc, but the presence of dietary fibre and phytic acid can lower their bioavailability. Metal
complexation by β-glucans and other polysaccharides is known (Platt and Clydesdale, 1984),
but information about the process-induced changes in metal binding capacity is not available.

20
The relationships between β-glucan structure and its resulting biological functions have yet to
be fully established and this lack of knowledge is a major limitation for effectively utilizing
β-glucan as functional food. This thesis aimed at investigating the process-induced changes of
oat and barley β-glucan’s structure and the impact of these modification on the health properties
of this fiber.
References
(FDA) F.a.D.A. 21 CFR 101.81 (2009) ‘Health claims: Soluble fiber from certain foods and risk of coronary heart disease (CHD)’, in Regulatory Information, pp. 143–148.
Efsa (2011) ‘Scientific Opinion on the substantiation of health claims related to beta-glucans from oats and barley and maintenance of normal blood LDL-cholesterol concentrations ( ID 1236 , 1299 ), increase in satiety leading to a reduction in energy intake ( ID 851’, EFSA Journal, 9(6), pp. 1–21. doi: 10.2903/j.efsa.2011.2207.
Kahlon, T. S. and Woodruff, C. L. (2003) ‘In vitro binding of bile acids by rice bran, oat bran, barley and β-glucan enriched barley’, Cereal Chemistry. doi: 10.1094/CCHEM.2003.80.3.260.
Kivelä, R., Gates, F. and Sontag-Strohm, T. (2009) ‘Degradation of cereal beta-glucan by ascorbic acid induced oxygen radicals’, Journal of Cereal Science, 49(1), pp. 1–3. doi: 10.1016/j.jcs.2008.09.003.
Kivelä, R., Henniges, U., Sontag-Strohm, T. and Potthast, A. (2012) ‘Oxidation of oat β-glucan in aqueous solutions during processing’, Carbohydrate Polymers, 87(1), pp. 589–597. doi: 10.1016/j.carbpol.2011.08.028.
Lan-Pidhainy, X., Brummer, Y., Tosh, S. M., Wolever, T. M. and Wood, P. J. (2007) ‘Reducing beta-glucan solubility in oat bran muffins by freeze-thaw treatment attenuates its hypoglycemic effect’, Cereal Chemistry, 84(5), pp. 512–517. doi: 10.1094/CCHEM-84-5-0512.
Lee, K. Y., Park, S. Y. and Lee, H. G. (2011) ‘Effect of oat β-glucan and its oxidised derivative on the quality characteristics of sponge cake’, International Journal of Food Science & Technology, 46(12), pp. 2663–2668. doi: 10.1111/j.1365-2621.2011.02798.x.
de Moura, F. A., Pereira, J. M., da Silva, D. O., Zavareze, E. da R., da Silveira Moreira, A., Helbig, E. and Dias, A. R. G. (2011) ‘Effects of oxidative treatment on the physicochemical, rheological and functional properties of oat β-glucan’, Food Chemistry, 128(4), pp. 982–987. doi: 10.1016/j.foodchem.2011.04.003.
Park, S. Y., Bae, I. Y., Lee, S. and Lee, H. G. (2009) ‘Physicochemical and Hypocholesterolemic Characterization of Oxidized Oat β-Glucan’, Journal of Agricultural and Food Chemistry, 57(2), pp. 439–443. doi: 10.1021/jf802811b.
Platt, S. R. and Clydesdale, F. M. (1984) ‘Binding of Iron by Cellulose, Lignin, Sodium Phytate and Beta-Glucan, Alone and in Combination, Under Simulated Gastrointestinal pH Conditions’, Journal of Food Science. Blackwell Publishing Ltd, 49(2), pp. 531–535. doi: 10.1111/j.1365-2621.1984.tb12460.x.
Sayar, S., Jannink, J.-L. and White, P. J. (2011) ‘Textural and Bile Acid-Binding Properties of Muffins Impacted by Oat β-Glucan with Different Molecular Weights’, Cereal Chemistry Journal, 88(6), pp. 564–569. doi: 10.1094/CCHEM-02-11-0014.
Wolever, T. M., Tosh, S. M., Gibbs, A. L., Brand-Miller, J., Duncan, A. M., Hart, V., Lamarche, B.,

21
Thomson, B. A., Duss, R. and Wood, P. J. (2010) ‘Physicochemical properties of oat β-glucan influence its ability to reduce serum LDL cholesterol in humans: a randomized clinical trial’, The American Journal of Clinical Nutrition, 92(4), pp. 723–732. doi: 10.3945/ajcn.2010.29174.
Wood, P. J. (2010) ‘REVIEW: Oat and rye -glucan: Properties and function.’, Cereal Chem., 87(4), pp. 315–330.

22

23
PART 1: REVIEW OF THE LITERATURE

24

25
1. Cereal β-glucans
1.1 Occurrence and structural characteristics
Cereal β-glucan is a major component of the cell walls of the starchy endosperm and aleurone
layer of grains such as barley, oat, rye and wheat (Lazaridou and Biliaderis, 2007). In oat, barley
and rye, β-glucan is predominantly located in the starchy endosperm, while in wheat the highest
concentration is found in the subaleurone layer (Biliaderis and Izydorczyk, 2006; Wood, 2010).
This different distribution of β-glucan in the grains of different cereals is an aspect that needs
to be kept in consideration when choosing extraction and isolation procedures aimed at
obtaining fractions enriched in β-glucan (Izydorczyk and Biliaderis, 2000). The β-glucan
contents vary significantly among different cereals, with the highest amounts in barley (2.5–
11.3%) and oat (2.2–7.8%) followed by rye (1.2–2.9%) and wheat (0.4–1.4%) This differences
depend mainly on genotypical factors, but also environmental aspects, such as the availability
of water during grain maturation, can affect β-glucan content (Biliaderis and Izydorczyk, 2006).
As for the structure, cereal β-glucan (mixed linkage (1→3, 1→4)-β-D-glucan) is a
homopolysaccharide of D-glucopyranose made of blocks of consecutive (1→4)-linked β-D-
glucose residues separated by (1→3)-linkages. The β-(1→4)-linkages are predominant (about
70%) and generally occur in groups of two or three, while the β-(1→3)-linkages occur
individually (Lazaridou and Biliaderis, 2007). The flexible (1→3)-linkages interrupt the rigid
cellulose-like sections formed by (1→4)-linked glucose units, increasing the solubility of this
polymer in water and its ability to form highly viscous solutions. The resultant structure is a
polysaccharide built mainly from β-(1→3)-linked cellotriosyl (degree of polymerization DP3)
and cellotetraosyl (degree of polymerization DP4) units, but there is evidence of a minor
amount of sequences (between 5 and 10%) with consecutive (1→4) linkages longer than the
tetraose type and up to 20 glucosyl residues (Wood et al., 1991; Izydorczyk et al., 1998;
Biliaderis and Izydorczyk, 2006). Of these, DP5, DP6 and DP9 were shown to be the most
abundant (Wood et al., 1994; Lazaridou et al., 2003).

26
Figure 1. Molecular structure of cereal β-glucan.
β-Glucan from different cereal sources present similar structural characteristics but they differ
in fine structure, i.e. the molar ratio between cellotriose and cellotetraose units (DP3/DP4),
which is seen as the fingerprint of the structure of this polysaccharide. This characteristic ratio
has been shown to affect several physico-chemical properties of β-glucan, such as water
solubility, viscosity and gel formation behaviour, which are crucial for the technological and
health implications of this fiber. In particular, lower DP3/DP4 ratios translate into higher
solubility and viscosity and into diminished gel-forming capacity (Tosh et al., 2004; Cui and
Wang, 2009; Wood, 2010; Ryu et al., 2012). In literature, several studies have investigated the
fine structure of β-glucan from several sources, finding the highest DP3/DP4 ratio in wheat (3.0
to 4.5), followed by barley (1.8 to 3.5), rye (1.9 to 3.0) and oat (1.5 to 2.3) (Wood et al., 1994;
Lazaridou et al., 2004; Biliaderis and Izydorczyk, 2006).
Together with fine structure, the molecular weight is another important characteristic regulating
the technological and physiological properties of β-glucan, in particular its ability to form
highly viscous solutions. Since polysaccharides are polydisperse (i.e. they do not possess a
particular degree of polymerization), the weight average molecular weight (Mw) is normally
used to describe them. In their review, Lazaridou et al. (2007) report values of 65-3100 kDa
for oat, 31-2700 kDa for barley, 21-1100 kDa for rye and 209-487 kDa for wheat. The
significant discrepancies in the Mw values for each cereal β-glucan might be due to several
factors, such as variety and environmental aspects, extraction methods (e.g. extraction medium,
pH, temperature, solid to liquid ratio, extraction times, etc.) but can also depend on the
analytical methods used for the determination (instruments, detectors, standards, etc.)
(Biliaderis and Izydorczyk, 2006). On top of this, during extraction, depending on the procedure
used, endogenous enzymes such as β-glucanase can cleave the β-glucan structure, shortening
the chains and causing a reduction in Mw (Burkus and Temelli, 1998). In order to inactivate

27
these enzymes and avoid β-glucan degradation, a treatment with hot aqueous ethanol (70 to
90%) is often used as the first step of the extraction. This also has the additional side benefit of
removing impurities such as lipids, free sugars, amino acids, small proteins, and some phenolics
(Lazaridou et al., 2004).
1.2 Rheology
The solution properties of these conformationally disordered polysaccharides, and in particular
their ability to generate highly viscous solutions, depend on the concentration and on the Mw of
the β-glucan chains. The rate and extent of dissolution, but also the concentration and Mw highly
influence their physiological effects (Judd and Ellis, 2005; Wang and Ellis, 2014).
Shear viscosity diagrams of β-glucan normally have three different linear regions for dilute,
semi-dilute and concentrated solutions, separated by points called critical concentrations c* and
c**. The first critical concentration, c*, separating the dilute from the semi dilute solutions
pinpoints the state in which the polymer molecules begin to interact with each other in solution.
The second critical concentration, c**, between the semi-dilute and concentrated region,
represents the beginning of entanglements. In the concentrated region, the viscosity increases
more rapidly with increasing concentration (Lazaridou et al., 2003; Skendi et al., 2003). In the
dilute region, β-glucan solutions are Newtonian while in the concentrated region they are non-
Newtonian and present shear thinning (pseudo plastic) behavior, which consists in a low shear
rate plateau region followed by a decrease of the apparent viscosities with increasing shear rate
(Wood, 2010). The shear thinning behaviour is due to a decline in the entanglements resulting
from the chains orientating themselves along the flow. Wood (2010) showed that a solution
with low concentration (0.18%) of high Mw oat β-glucan and a solution with high concentration
(2%) of low Mw β-glucan had Newtonian behavior. However, increasing the Mw and
concentration of β-glucan led to a shear thinning behavior with a low shear rate plateau.
Literature suggests that low Mw oat β-glucan (<100 kDa) does not behave as random coil in
solution which is a result of a change from flow to gel-like properties (Lazaridou et al., 2003;
Wood, 2010). β-Glucan has been shown to be able to form gels under certain conditions and its
gelation properties are extremely sensitive to structure. In particular, the higher the proportion
of β-(1→3)-linked cellotriosyl units, the faster the gelation rate (Böhm and Kulicke, 1999).
Therefore, among the cereal β-glucans, the gelation time decreases in the order oat, barley and
wheat, which reflects the DP3/DP4 ratio (Lazaridou et al., 2007; Wood, 2010). Two different
models have been proposed for the gelation of β-glucan solutions: the first one suggests that

28
cellulose-like sequences of more than three contiguous β-(1→4)-linked glucosyl units stick
together and lead to gelation (Fincher and Stone, 1986). The other model hypothesizes that the
association of consecutive cellotriose units (linked by β-(1→3)-bonds) may form extended
junction zones and lead to the development of a gel network structure (Böhm et al., 1999). In
general, the rate of gelation of β-glucan increases as the Mw decreases, which is due to the fact
that smaller β-glucan molecules have a better mobility and are therefore more likely to interact
with each other (Lazaridou et al., 2003; Skendi et al., 2003; Wood, 2010).
1.3 Health aspects
According to the American Association of Cereal Chemists, “dietary fiber is the edible parts of
plants or analogous carbohydrates that are resistant to digestion and absorption in the human
small intestine with complete or partial fermentation in the large intestine. Dietary fiber includes
polysaccharides, oligosaccharides, lignin, and associated plant substances. Dietary fibers
promote beneficial physiological effects including laxation, and/or blood cholesterol
attenuation, and/or blood glucose attenuation.” (AACCI, 2001).
Confirming this definition, cereal β-glucan, one of the most common partially soluble dietary
fibers, has been associated with several beneficial health effects, in particular the attenuation of
postprandial blood glucose and insulin levels and the reduction of serum cholesterol (Wood,
2007). On top of this, β-glucan has been shown to have a positive effect on satiety, potentially
helping to prevent obesity (Barone Lumaga et al., 2012; Pentikäinen et al., 2014), and some
immunomodulatory properties (Estrada et al., 1997). Also, it has been suggested that diets rich
in dietary fibers such as cereal β-glucan may have a protective effect against certain types of
cancers of the digestive system, in particular of colon cancer (Daou and Zhang, 2012;
Choromanska et al., 2015).
Literature is very rich of studies about β-glucan physiological effects, which can sometimes
present variable or conflicting results. This may be due to several factors, such as the
experimental design, the dose and form of β-glucan consumed (liquid, solid, isolate,
concentrate, gum, etc.) but also the health status of the individuals recruited for the trials. For
example, the ability of soluble fibers to attenuate postprandial glucose and insulin responses is
stronger in the case of hypercholesterolemic, older, obese and diabetic subjects and far less
significant for young and healthy individuals (Beer et al., 1995; Hallfrisch and Behall, 2000;
Davy et al., 2002). The same applies for the cholesterol lowering properties, which are more
pronounced in the case of subjects with initial high blood cholesterol levels (Bell et al., 1999).

29
As already mentioned, the ability of β-glucan to form highly viscous solutions in the intestinal
lumen is the main factor determining its beneficial health effects. Several factors affect this
ability, in particular the amount, solubility or extractability at physiological conditions, and the
Mw and structure of the fiber. Changes to any of these particular characteristics can profoundly
affect β-glucan’s health properties (Wood, 2007).
1.3.1 Blood glucose regulation
The postprandial blood glucose and insulin responses greatly vary depending on the food
consumed and are higher the higher the glycemic load (quantity of glucose ingested). An
attenuated response has been shown to be beneficial for both healthy and diabetic subjects
(Wolever et al., 1991; Jenkins et al., 2002; Radulian et al., 2009; Kwong et al., 2013). The
response to a particular food can be measured by the glycemic index (GI), which is calculated
as the area under the curve of the glucose response after the consumption of a carbohydrate
containing meal compared to the response generated by the intake of a specific glucose dose or
of a specific amount of white bread (Wolever et al., 1991). Several studies have shown that
foods rich in oat or barley β-glucan can attenuate both the GI and the GII (insulinemic
response), both in diabetic and healthy patients (Hallfrisch et al., 1995; Cavallero et al., 2002;
Kim et al., 2009). Viscous soluble dietary fibers such as β-glucan, in fact, can slow down the
digestion of macronutrients by delaying gastric emptying, reducing the transport and mixing of
digestive enzymes and increasing the barrier properties of the unstirred water layer (UWL)
separating the intestinal lumen from the enterocytes (Gunness et al., 2012). Cavallero et al.
(2002) conducted a study were healthy non diabetic patients were fed different types of bread:
a plain wheat bread and other three breads enriched with varying amounts of barley β-glucan
replacing portions of the wheat flour. The results showed that the β-glucan enriched bread
attenuated the glycaemic response compared to the plain wheat bread (by up to 28%),
confirming the effectiveness of cereal β-glucan in reducing blood glucose levels, even in foods
with a high glycemic index such as bread.
The attenuation of postprandial blood glucose and insulin observed after consumption of oat or
barley products is due to the formation of viscous intestinal contents, which depends on both
the concentration and Mw of the solubilized β-glucan. For example, in a study by Wood et al.
(1994), healthy individuals were fed model drinks containing 50 g glucose and 1.3 to 10.5 g of
oat gum, an extract composed mainly of β-glucan. A linear relationship between log (viscosity)
of the meal and postprandial glucose and insulin responses was found, showing that 79 to 96%

30
of the changes in plasma glucose and insulin levels are due to viscosity. Moreover, following
partial acid hydrolysis of the oat gum and consequent loss of viscosity, the ability of the gum
to attenuate postprandial glucose and insulin levels was reduced or even eliminated. Similar
effects have also been shown for solid foods enriched with oat or barley β-glucan (Tappy et al.,
1996; Östman et al., 2006), confirming the key role of viscosity and, as a consequence, of
β-glucan concentration and Mw.
1.3.2 Cholesterol lowering
The potential cholesterol lowering effect of oat was first brought to public attention with a study
by Anderson et al. (1984), which showed that oat-bran enriched diets decreased serum
cholesterol concentrations by 19% and calculated LDL cholesterol by 23% with no change in
HDL cholesterol in 20 hypercholesterolemic patients. Anderson and his colleagues concluded
that the soluble fiber content of the oat bran was probably responsible for the observed
reduction, which was confirmed with another study in 1990 (Anderson et al., 1990), thus
indicating that oat products and other soluble-fiber sources can have a significant impact on
lowering serum total cholesterol and other lipid risk factors for cardiovascular disease (CVD).
Since oat bran was shown to be very rich in β-glucan (Wood et al., 1989) and the ability of
soluble fiber to lower serum cholesterol was well known, it was first hypothesized and then
confirmed that β-glucan was the active component of oat responsible for this beneficial health
effect (Braaten et al., 1994; Wolever et al., 2010). Similar results were also published for barley
rich diets, once more confirming the active role of β-glucan in lowering serum cholesterol and
in the prevention of CVD (McIntosh et al., 1991; Bourdon et al., 1999; Smith et al., 2008).
In 1997, the U.S. Food and Drug Administration (FDA) authorized the health claim on the
association between soluble dietary fiber from whole oat and a reduced risk of coronary heart
disease (CHD), recognizing β-glucan as the main bioactive compound (FDA, 1997). In
particular, 3 g of β-glucan per day from 0.75 g/serving was recognized as the minimum dose to
exert the cholesterol lowering effect. As reviewed by Lazaridou and Biliaderis (2007), a
decrease of 1% in serum cholesterol corresponds to a reduction of 2% in the risk of mortality
by hearth disease, therefore even a small change is of the greatest importance, in particular for
hypercholesterolemic subjects. Since it has been shown that soluble fibers can decrease serum
cholesterol levels by 10 to 20%, the risk of coronary heart disease can be theoretically be
reduced by 20 to 40% (Anderson et al., 1990).

31
The exact mechanism by which soluble fibers such as β-glucan attenuate the serum cholesterol
is still unclear, even though several hypothesis have been proposed (Anderson et al., 1990; Bell
et al., 1999; Gunness and Gidley, 2010). The first theory suggests an interaction between
soluble fibers and bile acids, whose synthesis and metabolism are strictly related to the
cholesterol levels in the blood. In humans, primary bile acids such as cholic acid and
chenodeoxycholic acid are synthesized in the liver starting from cholesterol and are then
conjugated with glycine or taurine via N-acyl linkage before being stored in the gall bladder.
The conjugated bile acids are commonly named “bile salts” since at the pH found in the gall
bladder and small intestine they are mostly found in the ionized form, unlike the primary bile
acids, which are weaker acids and therefore usually undissociated (Gunness and Gidley, 2010).
In the gall bladder bile salts reach concentrations up to 300mM, significantly higher than their
critical micellar concentration (CMC, usually <10mM), which lead these amphipathic
molecules to aggregate and form mixed micelles with biliary cholesterol and
phosphatidylcholine, which are then excreted into the duodenum after lipid containing meals
(Hofmann, 1999). On top of their role in the solubilization of dietary and biliary cholesterol and
liposoluble vitamins, these micelles also act as the site of lipase digestion of lipids (Armand,
2007). After digestion, the mixed micelles containing cholesterol, vitamins and the lipolysis
products diffuse through the unstirred water layer (UWL) lining the brush border membrane of
the intestinal lumen, allowing uptake of cholesterol and other lipophilic compounds by the
enterocytes.
Figure 2. Schematic representation of the mechanism of sequestration of bile salts by soluble dietary fibers (SDF).
32
Once this task is completed, the micelles travel through the rest of the small and large intestine
and during this journey the bile salts are progressively reabsorbed into the enterohepatic
circulation, by active and passive transport mechanisms, via the hepatic portal vein (Gunness
et al., 2012). It has been shown that the presence of soluble fibers in the small intestine can
prevent at least part of the bile salts from being re-absorbed in the enterohepatic circulation,
leading instead to their excretion in the faeces (Figure 2). In order to replenish the bile salts
pool in the liver, cholesterol is catabolized by the hepatocyte, causing a reduction of cholesterol
levels in the blood (Marlett and Fischer, 2002; Ellegård and Andersson, 2007). The way bile
salts and soluble fiber interact is still not completely clear and in recent years three mechanisms
have been proposed:
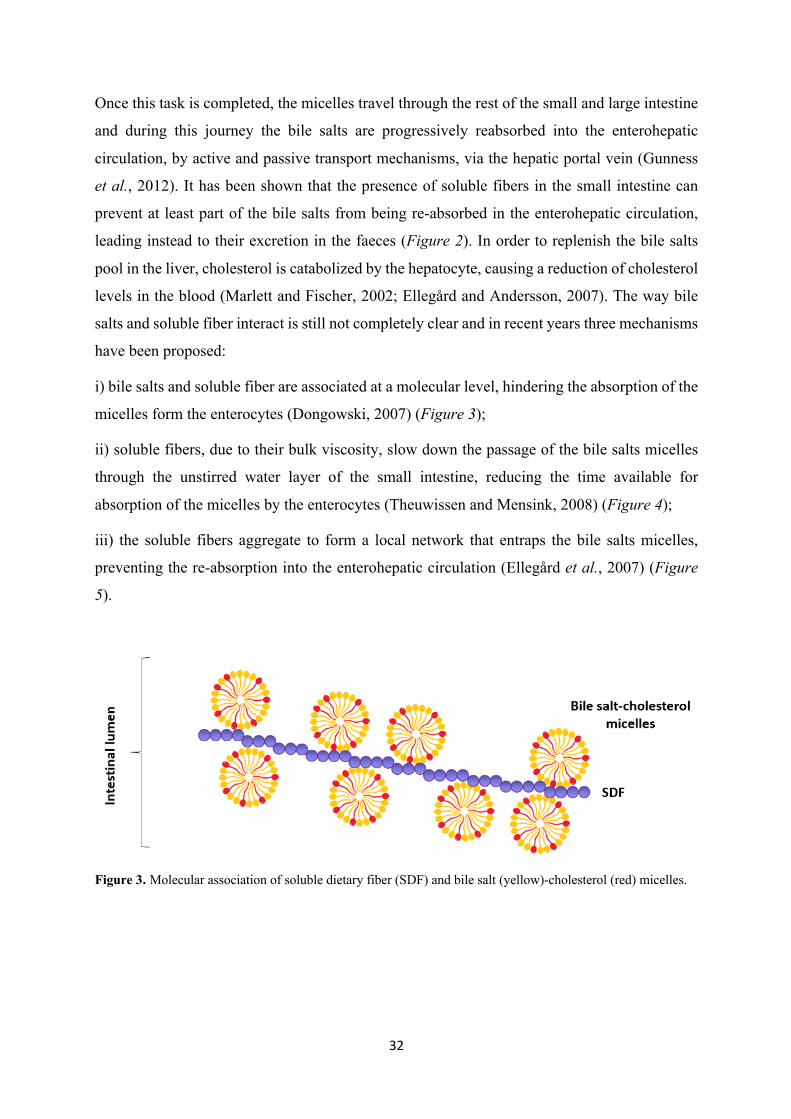
i) bile salts and soluble fiber are associated at a molecular level, hindering the absorption of the
micelles form the enterocytes (Dongowski, 2007) (Figure 3);
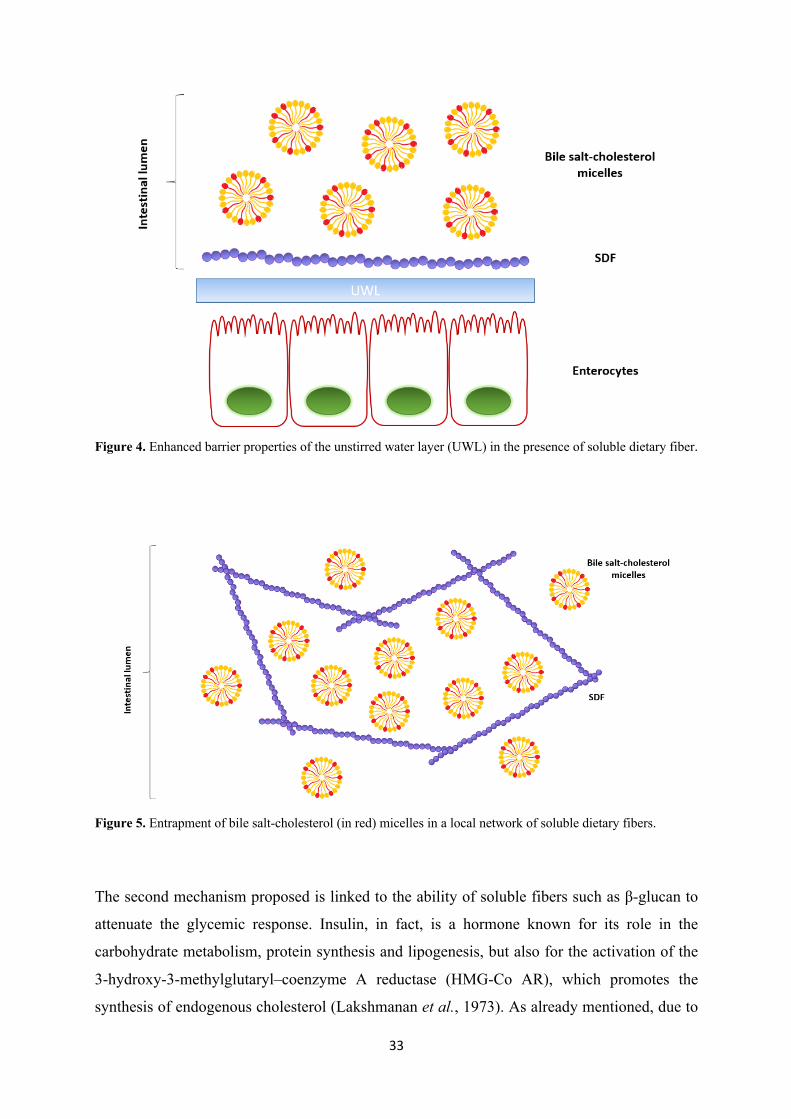
ii) soluble fibers, due to their bulk viscosity, slow down the passage of the bile salts micelles
through the unstirred water layer of the small intestine, reducing the time available for
absorption of the micelles by the enterocytes (Theuwissen and Mensink, 2008) (Figure 4);
iii) the soluble fibers aggregate to form a local network that entraps the bile salts micelles,
preventing the re-absorption into the enterohepatic circulation (Ellegård et al., 2007) (Figure
5).
Figure 3. Molecular association of soluble dietary fiber (SDF) and bile salt (yellow)-cholesterol (red) micelles.
33
Figure 4. Enhanced barrier properties of the unstirred water layer (UWL) in the presence of soluble dietary fiber.
Figure 5. Entrapment of bile salt-cholesterol (in red) micelles in a local network of soluble dietary fibers.
The second mechanism proposed is linked to the ability of soluble fibers such as β-glucan to
attenuate the glycemic response. Insulin, in fact, is a hormone known for its role in the
carbohydrate metabolism, protein synthesis and lipogenesis, but also for the activation of the
3-hydroxy-3-methylglutaryl–coenzyme A reductase (HMG-Co AR), which promotes the
synthesis of endogenous cholesterol (Lakshmanan et al., 1973). As already mentioned, due to

34
their ability to form a highly viscous intestinal contents, soluble fibers can slow down the
absorption of macronutrients such as glucose, with a consequent reduced postprandial glucose
level. This reduction is reflected in a decline in insulin level, which can cause an inhibition of
HMG-Co AR, leading to a decrease in cholesterol synthesis (Potter et al., 1981; Lazaridou and
Biliaderis, 2007). The mechanism by which insulin affects the synthesis of cholesterol,
however, is very complex and since this hormone is involved in many other diet-induced
responses, it is difficult to establish its exact contribution in the cholesterol synthesis and
metabolism. Also, since the formation of highly viscous intestinal contents by soluble dietary
fibers also causes a reduction in the re-absorption of bile salts, it is difficult to disentangle the
two mechanisms, which inevitably affect each other (Gunness and Gidley, 2010).
The third mechanism is associated with the anaerobic bacterial fermentation that soluble dietary
fibers undergo in the small intestine, which leads to the production of short chain fatty acids
(SCFA) such as acetate, propionate and butyrate. SCFA, in particular propionate, have been
shown to limit the action of HMG-Co A, that catalyzes the rate-limiting step of cholesterol
biosynthesis (Chen et al., 2013; Marcil et al., 2015). Several in vivo and in vitro studies were
conducted to investigate the role of propionate in the cholesterol lowering mechanism, only few
of which with humans and mostly with inconsistent results, therefore more human studies
would be required to explore this further (Todesco et al., 1991; Wolever et al., 1991). However,
even if propionate would be confirmed to be associated to the cholesterol-lowering properties
of soluble dietary fiber it would be likely to be confined only to a secondary role. Soluble fiber
fermentation in the colon, in fact, occurs irrespectively of the Mw of the carbohydrates, whereas
it is well known that the cholesterol-lowering effects are mainly due to viscosity, hence strictly
connected with molecular size (Gunness and Gidley, 2010).
To summarize, it can be concluded that the attenuation of cholesterol observed after a meal rich
in soluble dietary fibers is probably due to a combination of all the aforementioned effects, with
the excess bile salt secretion playing the leading role.

35
1.3.3 Additional health implications
Binding of metals. Cereals such as oat and barley are rich in nutritionally important minerals
but, at the same time, they also contain high amounts of soluble dietary fiber (e.g. β-glucan)
and phytic acid, which are known to reduce the bioavailability of iron and zinc (Camire and
Clydesdale, 1981; Persson et al., 1991; Torre et al., 1991). In an in vitro study by Persson et al.
(1991) the soluble dietary fiber fraction (mainly composed of β-glucan) of barley resulted to be
more active in complexing metals than oat, and a difference was found between different metals
(copper, zinc and cadmium). However, they showed that treatment with phytase to remove
phytic acid significantly reduced the binding capacity of the fiber extracts therefore potentially
excluding a direct interaction between fiber and metals. In a study by Platt and Clydesdale
(1984) cellulose, lignin, sodium phytate and cereal β-glucan, alone and in combination, were
evaluated with regard to their effect on iron profiles in model systems containing FeSO4·7H2O
under simulated gastrointestinal pH conditions. They observed that lowering the pH to gastric
conditions (pH 2) triggered the solubilisation of a portion of the iron bound to lignin, cellulose
and β-glucan. Bringing back the pH to neutral conditions (pH 6.8) led again to increased
portions of complexed iron and reduced portions of soluble iron. They hypothesised binding
strengths in the order lignin/ phytic acid > β-glucan > cellulose, but concluded that the binding
components in cereals interact, which critically affects iron solubility and thus bioavailability.
Faure et al. (2015) investigated the iron binding property of highly pure (>97%) commercial
oat and barley β-glucan by observing the kinetics of the Fenton reaction between Fe2+ and H2O2
in 0.6% oat and barley β-glucan solutions at pH values between 2.7 and 4.7. At pH 2.7 β-glucan
form both sources showed no influence on the rate of the Fenton reaction, while at pH 4.7 barley
β-glucan and, even more, oat β-glucan reduced the reaction rate, suggesting a binding with iron.
The difference in iron binding power was hypothesized to be due to the difference of DP3:DP4
between β-glucan from the two different cereal sources, with oat showing a stronger binding
than barley. On the other hand, in a paper by Boulos and Nyström (2016), no significant
difference was found in the kinetics of the Fenton-induced oxidation of three constitutionally
isomeric β-glucan tetrasaccharides used as model compounds, with no or one β-(1→3)-linkage.
This suggests that for the commercial β-glucan, the pure carbohydrates are not the main

36
responsible for the difference in behaviour between oat and barley, but rather a contaminant
present in different amounts depending on the cereal source.
Figure 6. Chemical structure of phytic acid.
Wang et al. (2017, a&b) unequivocally explained the differences in iron-binding behaviour of
commercial β-glucan materials from different cereal sources with the varying amounts of phytic
acid contamination, since removal of phytic acid with ion-exchange resins led to same
degradation rates and iron binding abilities of all β-glucan materials regardless of their origin.
In addition to the bioavailability of metals in the gastrointestinal tract, it is important to consider
that binding transition metals like iron and copper will also have an effect on β-glucan oxidation
(as clarified in section 2.1), as the metals are involved in the Fenton reaction and hence in the
reactions forming e.g. hydroxyl radicals.
Other health effects.As reviewed by Lazaridou et al. (2007) diets enriched with oat or barley
β-glucan have shown a reduction in feed intake and weight gain in several animal studies, but
in human studies there was no evidence of such an effect. However, some in vivo studies on
humans showed that oat β-glucan can reduce intestinal transit time and increase fecal weight,
which can decrease the opportunity for both nutrients and fecal mutagens to interact with the
intestinal epithelium (Jones, 2002). It has also been suggested that diets high in dietary fiber
such as β-glucans may have a protective effect against colon cancer (Lazaridou et al., 2007).
Furthermore, a few studies show that cereal β-glucan have a prebiotic effect and can therefore
contribute to maintain a healthy intestinal environment (Jaskari et al., 1998; Dongowski et al.,
2002; Bengmark, 2003).

37
2. β-glucan oxidation
Radical mediated oxidation of β-glucan during storage or food processing can cause chain
cleavage or oxidation of any of the hydroxyl groups of the glucose monomers, leading to the
formation of carbonyl or carboxyl groups or even to a cross-ring cleavage (Figure 7) (Boulos
& Nyström., 2016).
Figure 7. Potential oxidative modifications on the β-glucan backbone during processing and storage: (a) carboxyl formation at C6, (b) carbonyl formation at C6, (c) dicarbonyl formation with ring opening, (d) radical mediated carbonyl formation on ring, and (e) chain cleavage and carboxyl formation at C1.
All these changes can significantly affect the health and technological properties of cereal
β-glucan. In order to investigate the properties of oxidised β-glucan, or to obtain β-glucan with
specific characteristics, the new functional groups can also be engineered using targeted
chemical oxidation methods, such as TEMPO-mediated oxidation or sodium periodate
oxidation.
2.1 Unselective Fenton oxidation
During food processing and storage, cereal β-glucan can undergo enzymatic or chemical
degradation with consequent reduction of Mw and viscosity, which can impair their health
benefits (Tosh et al., 2010; Wolever et al., 2010). The chemically-induced oxidative
degradation of β-glucan has been shown to take place when traces of transition metals such as
iron or copper and a reducing agent such as ascorbic acid (AH2), which are frequently found in
food, come in contact with atmospheric oxygen (Schuchmann and von Sonntag, 1978; Kivelä

38
et al., 2009). Faure et al. (2013) identified the reactive oxygen species responsible for the loss
in viscosity and Mw of β-glucan by means of indirect spin trapping and electron spin resonance
(ESR) spectroscopy as the hydroxyl radical (•OH). It has been suggested that the catalytic cycle
of •OH production might be induced by the pro-oxidant AH2, which reduces intrinsic iron and
dissolved O2 to produce Fe2+ and hydrogen peroxide (H2O2) (Michels and Frei, 2013), which
are the two substrates for the so-called Fenton reaction (3) (Fenton, 1894).
Cu+/Fe2+ + O2 ⇄ Cu2+/Fe3+ + O2•- (1)
2O2•- + 2H+ → H2O2 + O2 (2)
Cu+/Fe2+ + H2O2 → Cu2+/Fe3+ + OH- + •OH Fenton (3)
AH2 + 2 Cu2+/Fe3+ → A + 2H+ + 2 Cu+/Fe2+ (4)
AH2 + O2 → A + H2O2 (5)
Hydroxyl radicals are highly reactive species that can oxidize the polysaccharides in their
vicinity at diffusion-controlled rates in a non-selective way, initiating a chain reaction that may
lead to several degradation products. β-Glucan have been reported to often be associated with
metals (Platt and Clydesdale, 1984), which can make these polysaccharides very susceptible to
oxidative degradation, since the metal catalyst is located in the proximity of the target molecule
(Kivelä et al., 2009). Evidence suggests that the hydroxyl-induced degradation takes place in a
random manner and is not influenced by the conformation of the glycosidic bond of the
polysaccharide, (Christensen et al., 1996; Hjerde et al., 1998).
Since glucose is the repeating unit forming the β-glucan polymeric chain, its degradation
products, which have been extensively studied by Schuchmann and von Sonntag (1977), are
expected to be similar to the degradation products of β-glucan. The authors, who studied the
degradation of glucose by hydroxyl radicals produced by ϒ-irradiation in N2O/O2 saturated
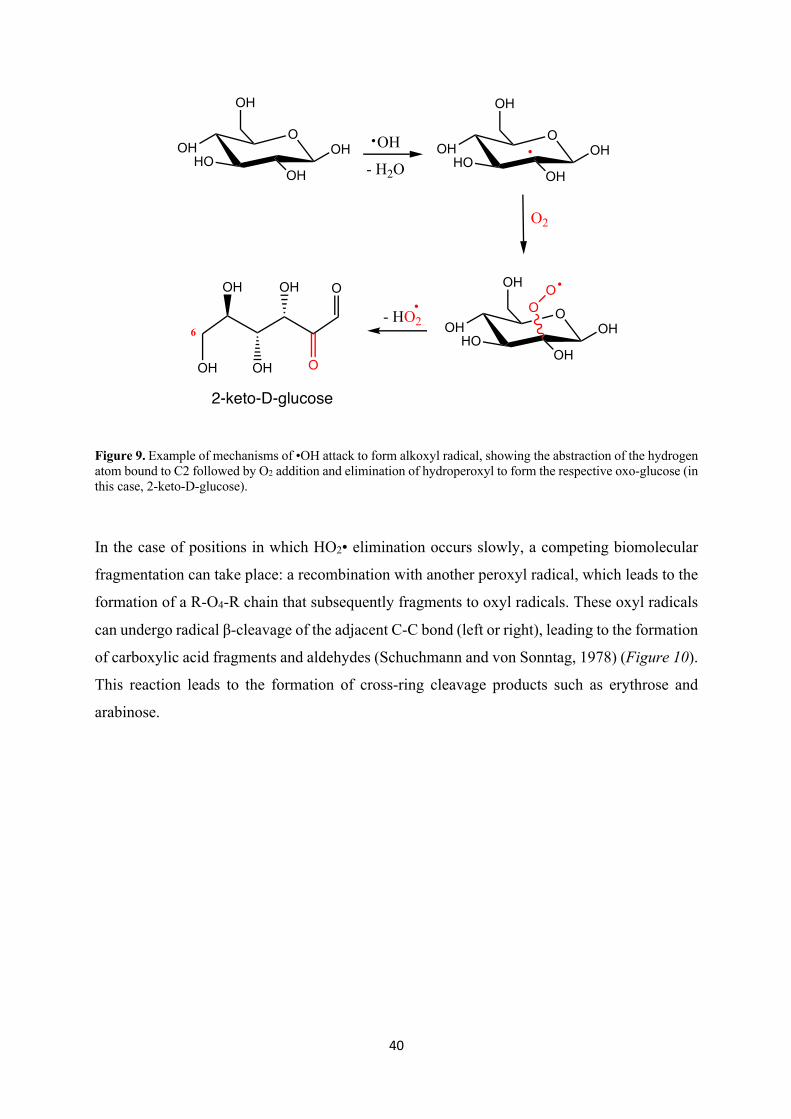
solutions, showed that these radicals can easily abstract a carbon bound hydrogen leading to the
formation of a carbon centered alkoxyl radical (6) (Schuchmann and von Sonntag 1977).
H-C-OH + •OH → •C-OH + H2O (6)
39
The newly formed alkoxyl radical can react with atmospheric O2 to produce a peroxyl radical
(7) and form a carbonyl group by elimination of hydroperoxyl radical(HO2•)(8).
•C-OH + O2 → •O-O-C-OH (7)
•O-O-C-OH → HO2• + C=O (8)
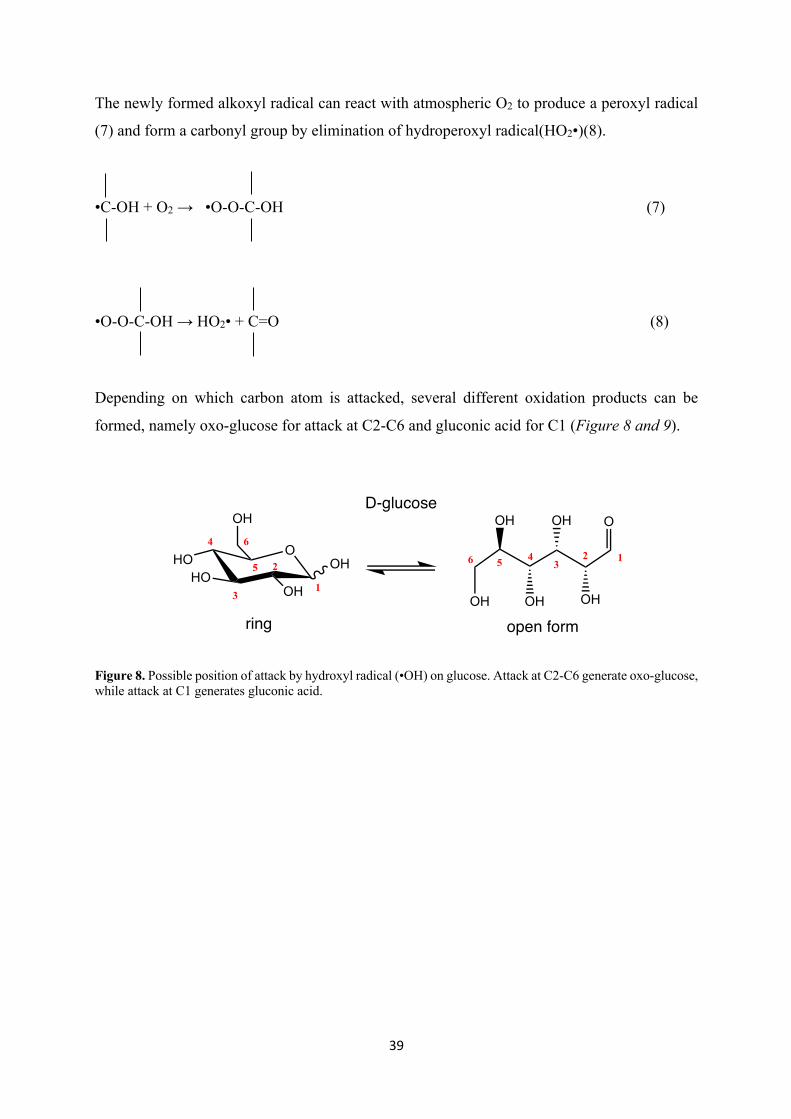
Depending on which carbon atom is attacked, several different oxidation products can be
formed, namely oxo-glucose for attack at C2-C6 and gluconic acid for C1 (Figure 8 and 9).
Figure 8. Possible position of attack by hydroxyl radical (•OH) on glucose. Attack at C2-C6 generate oxo-glucose, while attack at C1 generates gluconic acid.
OOH
OHHO
HO
OH OH
OH OH
OH
OH
O
123
46 5
1
2
3
4
5
6
D-glucose
ring open form
40
Figure 9. Example of mechanisms of •OH attack to form alkoxyl radical, showing the abstraction of the hydrogen atom bound to C2 followed by O2 addition and elimination of hydroperoxyl to form the respective oxo-glucose (in this case, 2-keto-D-glucose).
In the case of positions in which HO2• elimination occurs slowly, a competing biomolecular
fragmentation can take place: a recombination with another peroxyl radical, which leads to the
formation of a R-O4-R chain that subsequently fragments to oxyl radicals. These oxyl radicals
can undergo radical β-cleavage of the adjacent C-C bond (left or right), leading to the formation
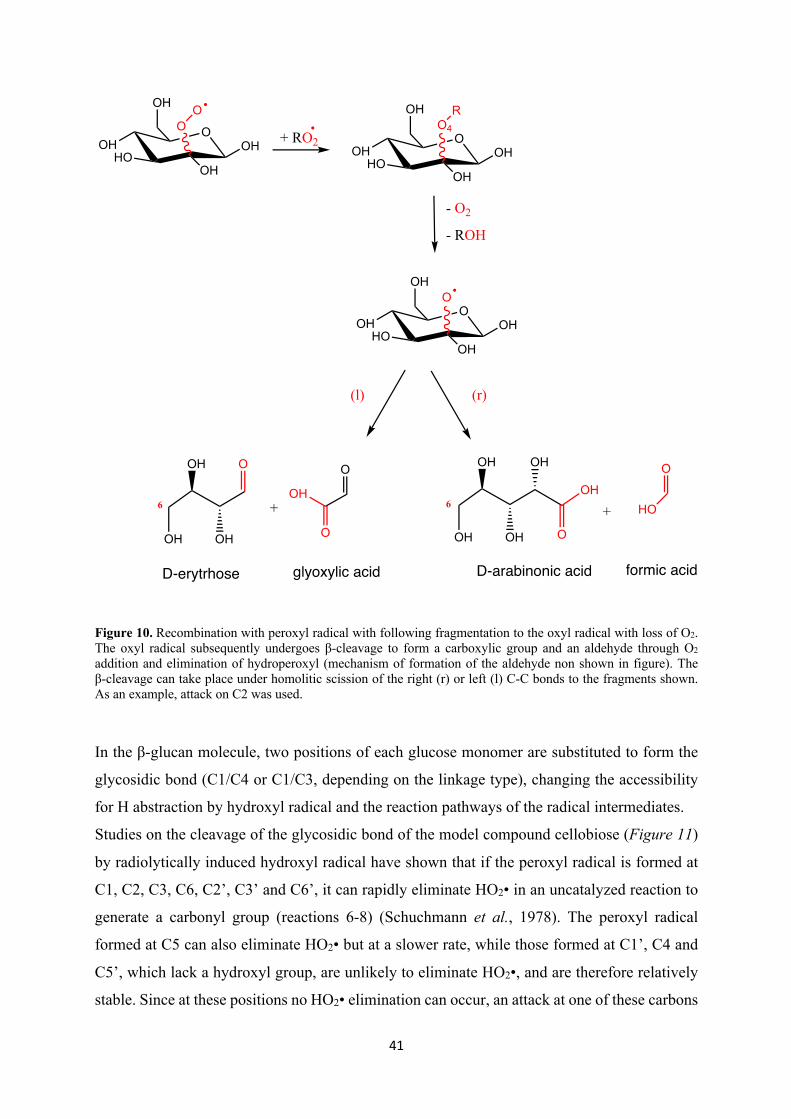
of carboxylic acid fragments and aldehydes (Schuchmann and von Sonntag, 1978) (Figure 10).
This reaction leads to the formation of cross-ring cleavage products such as erythrose and
arabinose.
O
OHHO
OH
OH
OH OH
- H2O
O
OHHO
OH
OH
OH
O2
O
OHHO
OH
OH
OHOO
- HO2
OH
OH OH
OH O
O
6
2-keto-D-glucose
41
Figure 10. Recombination with peroxyl radical with following fragmentation to the oxyl radical with loss of O2. The oxyl radical subsequently undergoes β-cleavage to form a carboxylic group and an aldehyde through O2 addition and elimination of hydroperoxyl (mechanism of formation of the aldehyde non shown in figure). The β-cleavage can take place under homolitic scission of the right (r) or left (l) C-C bonds to the fragments shown. As an example, attack on C2 was used.
In the β-glucan molecule, two positions of each glucose monomer are substituted to form the
glycosidic bond (C1/C4 or C1/C3, depending on the linkage type), changing the accessibility
for H abstraction by hydroxyl radical and the reaction pathways of the radical intermediates.
Studies on the cleavage of the glycosidic bond of the model compound cellobiose (Figure 11)
by radiolytically induced hydroxyl radical have shown that if the peroxyl radical is formed at
C1, C2, C3, C6, C2’, C3’ and C6’, it can rapidly eliminate HO2• in an uncatalyzed reaction to
generate a carbonyl group (reactions 6-8) (Schuchmann et al., 1978). The peroxyl radical
formed at C5 can also eliminate HO2• but at a slower rate, while those formed at C1’, C4 and
C5’, which lack a hydroxyl group, are unlikely to eliminate HO2•, and are therefore relatively
stable. Since at these positions no HO2• elimination can occur, an attack at one of these carbons
O
OHHO
OH
OH
OHOO
+ RO2 O
OHHO
OH
OH
OH
O4R
- O2
- ROH
O
OHHO
OH
OH
OH
O
(l) (r)
OH
OH
OH OH
OH
O
+
O
HO
OH
OH OH
O
+
O
O
OH
D-erytrhose glyoxylic acid D-arabinonic acid formic acid
6 6

42
leads to chain scission by biomolecular fragmentation, in which one oxidized unit or fragment
is formed while the other unit is assumed to stay unchanged.
Figure 11. Cellobiose structure.
Therefore, in the β-glucan molecule, an attack at any of the glycosidic acetal carbons, i.e. C1,
C3 or C5 (in case of β-(1→3) linkage) or C1, C4, C5 (in case of β-(1→4) linkage), leads to
breakage of the glycosidic bond and depolymerization of β-glucan with consequent loss of
viscosity. Attack at any other carbon, on the other hand, never leads to direct chain scission.
It has been proposed that chain scission might also take place through formation of an oxonium
ion resulting from hydroxyl radical attack, followed by O2 addition and elimination of HO2•,
even though no free hydroxyl groups are present on the attacked carbon (Lindsay and Fry,
2007). In reality, different mechanisms are possible for the glycosidic bond cleavage and the
prevailing one depends on several parameters, such as availability and concentration of
dissolved O2, Fe and H2O2 (Boulos and Nyström, 2016).
In any case, all these changes dramatically affect the physicochemical characteristics of
β-glucan and can potentially have a positive or negative impact on the functionality of this fiber
depending on the application.
2.2 Selective TEMPO oxidation
TEMPO-mediated oxidation, which is used to selectively oxidize the primary hydroxyl groups
(C6) of polysaccharides into carboxylic groups, has been widely used in carbohydrate chemistry
over the last two decades. The interest for new polyuronides is due to their valuable properties,
such as antiflocculation, adhesion, gelation, thickening, complexation, as well as a high number
of biological activities (Pierre et al., 2017). TEMPO is a nitroxyl radical (2,2,6,6-
tetramethylpiperidine-1-oxyl radical) in which an unpaired electron is delocalized between the
N and O atoms (Figure 12). Generally, a catalytic amount of TEMPO and NaBr are added in
polysaccharide solutions at pH 10-11, and the oxidation is initiated by the addition of NaClO
as primary oxidant (Isogai et al., 2011). Consequently, the primary hydroxyl groups (C6) of the

43
polysaccharides are first converted into aldehydes and, as the reaction progresses, the aldehydes
are converted into carboxylates.
TEMPO-mediated oxidation can also be used to increase the solubility of polysaccharides, as
demonstrated by Chang and Robyt (1996), who showed that after TEMPO oxidation both
soluble and insoluble polysaccharides had increased water solubility compared to the native
ones. Specifically, water-insoluble polysaccharides such as cellulose and chitin became water
soluble (9.4 and 7.9% w/v, respectively) while water-soluble polysaccharides had their water
solubility doubled or tripled. This increased solubility is especially useful in the case of curdlan,
a β-(1→3)-glucan of bacterial origin, whose interesting biological activities such as anti-HIV
agent, antitumor efficacy and anticoagulant are limited in their applicability by its water-
insolubility. A study showed that after 100 min in the presence of TEMPO/NaBr/NaClO at pH
10, all the C6 primary hydroxyl groups of curdlan were converted into carboxylates and the
oxidized products were water-soluble. Oxidation, however, led to a strong depolymerization of
the curdlan whose degree of polymerization decreased from 6790 to 86 (Tamura et al., 2009).
As shown in this study, despite the high effectiveness in selectively converting the C6 primary
hydroxyl groups of polysaccharides into carboxylic groups, this method can also cause
extensive depolymerization, possibly due to a β-elimination at the C6 aldehyde groups formed
as intermediates during oxidation or cleavage of the glycosidic bond by hydroxyl radicals
formed as by-products during oxidation (Hirota et al., 2009). For this reason, new methods
based on TEMPO/NaClO/NaClO2 under neutral or acidic conditions have been developed,
which only cause a limited depolymerization of the polysaccharide (Hirota et al., 2009; Tamura
et al., 2010).
44
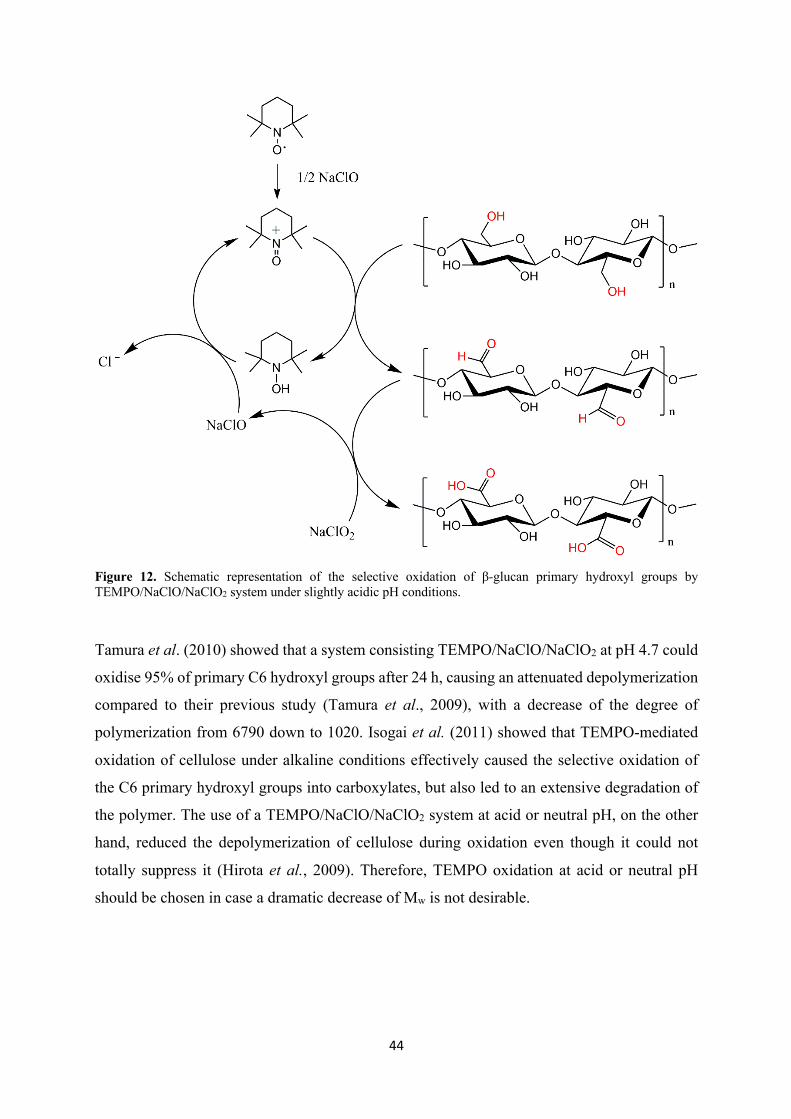
Figure 12. Schematic representation of the selective oxidation of β-glucan primary hydroxyl groups by TEMPO/NaClO/NaClO2 system under slightly acidic pH conditions.
Tamura et al. (2010) showed that a system consisting TEMPO/NaClO/NaClO2 at pH 4.7 could
oxidise 95% of primary C6 hydroxyl groups after 24 h, causing an attenuated depolymerization
compared to their previous study (Tamura et al., 2009), with a decrease of the degree of
polymerization from 6790 down to 1020. Isogai et al. (2011) showed that TEMPO-mediated
oxidation of cellulose under alkaline conditions effectively caused the selective oxidation of
the C6 primary hydroxyl groups into carboxylates, but also led to an extensive degradation of
the polymer. The use of a TEMPO/NaClO/NaClO2 system at acid or neutral pH, on the other
hand, reduced the depolymerization of cellulose during oxidation even though it could not
totally suppress it (Hirota et al., 2009). Therefore, TEMPO oxidation at acid or neutral pH
should be chosen in case a dramatic decrease of Mw is not desirable.

45
2.3 Selective sodium periodate oxidation
The chemical modification of polysaccharides is one of the most important tools to design new
materials with new structures and properties (Coseri et al., 2015). It has been shown that a
variety of oxidation mechanisms have different effects on the polysaccharides and that the
oxidized polymers bear altered health and technological properties compared to the native ones.
For example, the oxidation of the polysaccharide sodium alginate renders it more efficient in
crosslinking proteins and partial oxidation of polysaccharides may be used to introduce highly
flexible links into the otherwise stiff structures (Balakrishnan et al., 2005; Kristiansen et al.,
2010). Also, as already mentioned, radical induced oxidation of polysaccharides can occur
during food processing and storage, introducing new functional groups on the polymeric
backbone and causing chain cleavage (Kivelä et al., 2009). In order to study these structural
modifications methods for targeted oxidation, such as TEMPO oxidation (section 2.2), can be
used. Sodium periodate (NaIO4) oxidation can also be applied for this purpose, in particular to
investigate the effects of cross-ring cleavage of β-glucan caused by oxidation during food
processing. As shown by Boulos and Nyström (2017), in fact, cross-ring cleaved units were
formed in oat and barley β-glucan solutions in the presence of FeSO4 and H2O2 and clearly
detected by UPLC/MS with the help of fluorescent labelling. NaIO4 can oxidize selectively the
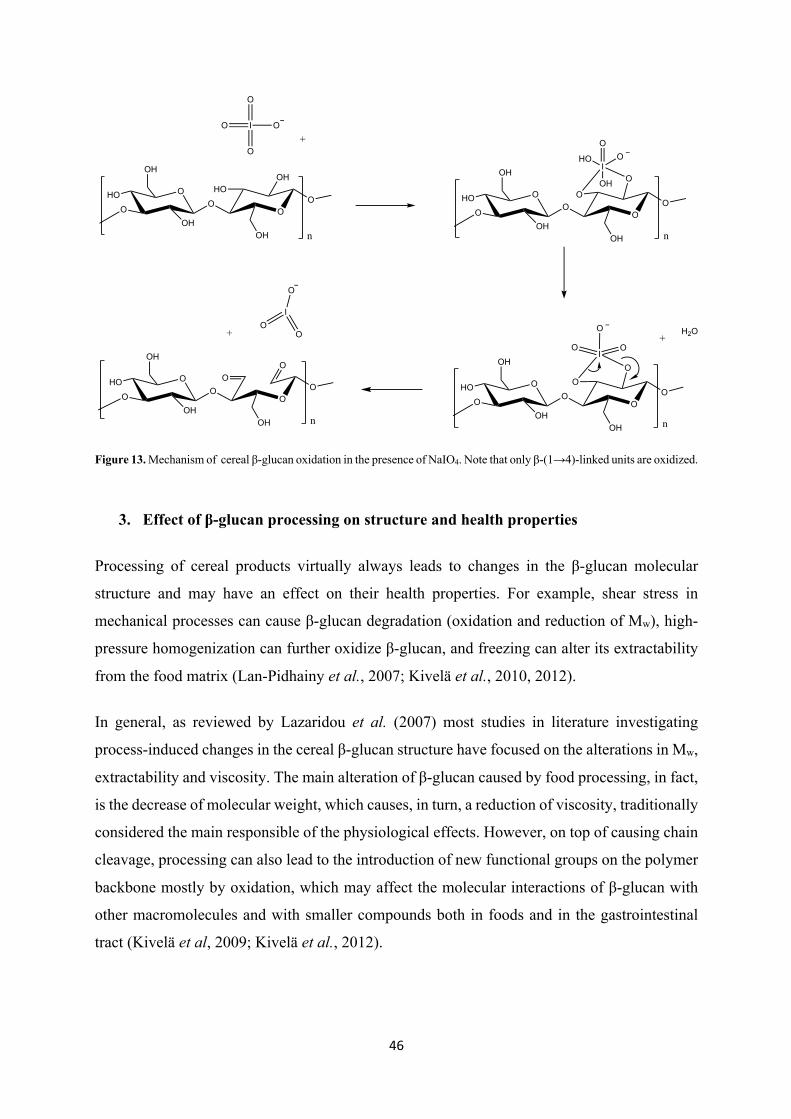
two secondary, vicinal hydroxyl groups of the glucose monomers in carbohydrates such as
cellulose. This leads to the oxidative cleavage of the C2-C3 bond, resulting in the opening of
the glucopyranose ring and the formation of two aldehyde groups (Figure 13). It must be kept
in mind, however, that NaIO4 oxidation strongly influences the hydrolytic stability of
polysaccharides and the newly formed dialdehyde derivatives are generally known to be highly
susceptible to alkaline β-elimination, which can cause significant depolymerization.
46
Figure 13. Mechanism of cereal β-glucan oxidation in the presence of NaIO4. Note that only β-(1→4)-linked units are oxidized.
3. Effect of β-glucan processing on structure and health properties
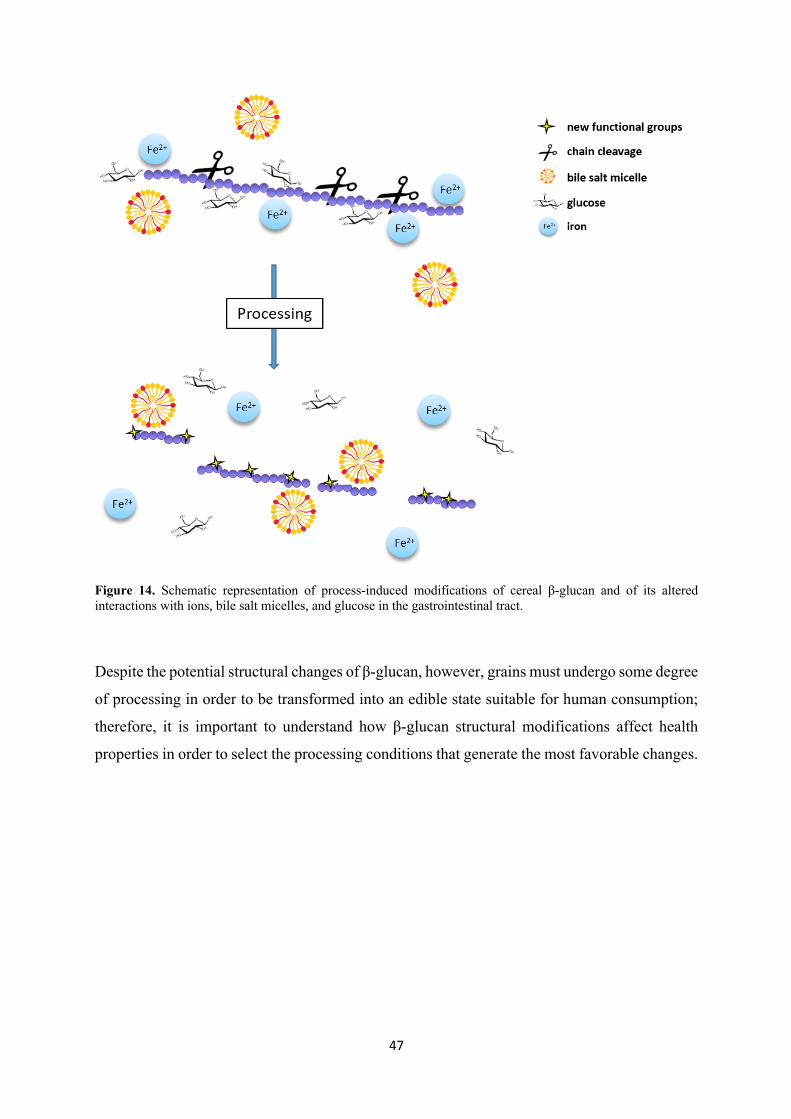
Processing of cereal products virtually always leads to changes in the β-glucan molecular
structure and may have an effect on their health properties. For example, shear stress in
mechanical processes can cause β-glucan degradation (oxidation and reduction of Mw), high-
pressure homogenization can further oxidize β-glucan, and freezing can alter its extractability
from the food matrix (Lan-Pidhainy et al., 2007; Kivelä et al., 2010, 2012).
In general, as reviewed by Lazaridou et al. (2007) most studies in literature investigating
process-induced changes in the cereal β-glucan structure have focused on the alterations in Mw,
extractability and viscosity. The main alteration of β-glucan caused by food processing, in fact,
is the decrease of molecular weight, which causes, in turn, a reduction of viscosity, traditionally
considered the main responsible of the physiological effects. However, on top of causing chain
cleavage, processing can also lead to the introduction of new functional groups on the polymer
backbone mostly by oxidation, which may affect the molecular interactions of β-glucan with
other macromolecules and with smaller compounds both in foods and in the gastrointestinal
tract (Kivelä et al, 2009; Kivelä et al., 2012).
O
OH
OH
OO
HOOH
OH
O
O
OH
OH
OO
OO
OH
OO
O
n
HO
n
HO
O
OH
OH
OO
O
O
OH
OO
n
HO
I
O
O
O O+
I
OOHO
OH
O
OH
OH
OO
OO
OH
OO
n
HO
IO
O H2O
O+
I
OO
O
+
47
Figure 14. Schematic representation of process-induced modifications of cereal β-glucan and of its altered interactions with ions, bile salt micelles, and glucose in the gastrointestinal tract.
Despite the potential structural changes of β-glucan, however, grains must undergo some degree
of processing in order to be transformed into an edible state suitable for human consumption;
therefore, it is important to understand how β-glucan structural modifications affect health
properties in order to select the processing conditions that generate the most favorable changes.

48
3.1 Chemical and enzymatic modification of β-glucan
3.1.1 Oxidation
It is well-known that β-glucan can undergo degradation (i.e. Mw reduction) with consequent
loss of viscosity in food products such as beverages or bakery products, and this modification
has traditionally been attributed to enzymatic or acidic hydrolysis, or to the use of high shear
force (Beer et al., 1997; Åman et al., 2004). Kivelä et al. (2009) were the first to demonstrate
that β-glucan in aqueous systems can undergo a type of degradation which cannot be explained
by enzymatic processed or acidic hydrolysis, the latter of which requires low pH (1-2) and high
temperatures. They showed that pure β-glucan solutions, in the presence of ascorbic acid (10
mM) and Fe2+, underwent a drastic viscosity loss, and hence Mw reduction, up to 50% in 1 h.
They suggested that this degradation process could be the result of an oxidative cleavage of the
β-glucan chain caused by hydroxyl radicals, a specific type of reactive oxygen species (ROS)
which is known to cause scission in polysaccharides and that can be produced through the
Fenton reaction (Fry, 1998; Kivelä et al., 2009). With this study, they underlined the importance
of understanding this mechanism because of its crucial role in potentially reducing the
beneficial β-glucan’s health properties, which are closely related to viscosity, but also in
undermining the stability of β-glucan in solution. The synergistic effect of ascorbic acid and
Fe2+ was also confirmed by another study by Kivelä et al.(2009), in which the addition of
ascorbic acid to an unpurified oat β-glucan extract caused the immediate decrease of viscosity
and Mw, with an even stronger reduction with the additional presence of metal ions such as Fe2+
and Cu2+. Even the sole addition of Fe2+ to the oat extract caused viscosity loss, confirming the
ability of Fe2+ to promote the degradation of β-glucan. It is important to notice that in this study
ascorbic acid induced a faster viscosity loss in the crude oat β-glucan extract compared to the
purified one, suggesting the occurrence of endogenous reactions catalysed by the free Fe2+ ions
present in the untreated extracts.
The radical mediated oxidation seems to play a key role also in the case of thermal treatments
of β-glucan solutions. A study investigated the degradation of oat β-glucan in highly purified
solutions or in crude extracts after thermal treatments at 95°C and 120 °C (Kivelä et al., 2011).
Both viscosity and Mw decreased because of the thermal treatments, with a more dramatic effect
in the case of the crude extract, containing also co-extracted phytic acid, proteins, minerals and
other components. Fenton reaction was deemed responsible for the degradation, since the
addition of Fe2+ ions accelerated the thermal degradation, strongly oxidative free radicals were

49
detected and oxygen was consumed while the β-glucan backbone was being broken down
during the thermal treatments. Additionally, when ascorbic acid was added to the solutions, the
viscosity loss was even more significant than only in the presence of Fe2+, confirming once
more the hypothesis of the Fenton reaction controlling the degradation.
On top of causing chain cleavage in the β-glucan backbone, radical mediated oxidation can also
lead to the oxidation of any of the hydroxyl groups of the glucose monomers, and to the
consequent formation of carbonyl or carboxyl groups or even to ring opening (Faure et al.,
2013). In a study by de Moura et al. (2011) the oxidation of β-glucan from oat bran was
promoted with H2O2 at different concentrations and for different reaction times at 40°C. De
Moura and her colleagues showed that the viscosity of the solutions went down with increasing
reaction times and amounts of H2O2 and also confirmed the formation of carbonyl and carboxyl
functional groups, which followed a similar pattern. Kivelä et al. (2012) demonstrated that
thermal treatments of oat β-glucan (95 and 120 °C for 30 min) led to the formation of carbonyl
groups simultaneously with the chain cleavage of cereal β-glucan, as a result of oxidation.
More recently, Faure et al. (2013) investigated the effect of temperature on the degradation of
cereal β-glucan induced by Fe2+. They showed that a higher temperature of storage (100°C) of
a commercial barley β-glucan solution in the presence of Fe2+, led to an enhanced formation of
hydroxyl radical and as a consequence a more intense depolymerization of β-glucan. At lower
temperatures (25 and 50°C), the radical catalyzed reactions were significantly slower, but an
addition of ascorbic acid or H2O2 enhanced the degradation of β-glucan. In a second study they
showed that the Mw of a 1% β-glucan solution, after 2 h of storage at 85°C, decreased from 538
kDa to molecules smaller than the detection limit of the light scattering analysis when 100 mM
H2O2 was added to the solution. Without the addition of H2O2, on the other hand, the Mw was
still 513 kDa after 2 h of storage at 85°C and 328 kDa after one week at the same temperature
(Faure et al., 2014).
3.1.2 Acidic and enzymatic hydrolysis
Major changes of the β-glucan structure are observed in processes that allow the action of
endogenous β-glucanases, or which use extreme conditions (heat, low pH, pressure). For
example, depolymerization can take place during the extraction step, caused by endogenous or
microbial β-glucanases from contaminating microorganisms (Lazaridou et al., 2007). Åman et
al. (2004) analyzed the β-glucan extracted from different oat products, finding large differences

50
in the average Mw and Mw distributions, which are likely to have an impact on the nutritional
properties of these foods. Oats, rolled oats, oat bran and oat bran concentrates all had
monomodal distributions and very high Mw (2060 to 2300 kDa), while the oat-enriched
experimental food products had lower Mw, ranging from 450 to 1920 kDa. Among the latter,
extruded flakes, macaroni and muffins had higher Mw while pasteurized apple juice, fresh pasta
and tea cakes were shown to contain significantly degraded β-glucan. Also, different types of
oat bran-based breads (normal, prefermented, and with sourdough) had average Mw ranging
from 240 to 1670 kDa, with large particle size of the oat bran and short fermentation times
limiting the degradation of β-glucan in these breads. The polymodal distribution of β-glucan in
these breads seems to point towards a degradation of enzymatic nature, probably due to
β-glucanases present in the flour mixture. In the case of the commercial foods studied, porridge
made of rolled oats, the breakfast cereal, and the extruded oat product all showed monomodal
distributions and high average Mw (1930 to 2010 kDa). The crisp bread and the yogurt-like
product contained medium degraded β-glucan (950 and 830 kDa, respectively) and bread loaf,
fried pancakes, pancake batter, and fermented oat soup had highly degraded β-glucans (1900 to
630 × 103). In general, processing including a fermentation step (e.g. baking) and production
of fresh pasta, fermented soup or pancake batter all seem to lead to a dramatic decrease in
β-glucan Mw. Heat treatment, on the other hand, resulted to be protecting β-glucan against
degradation, which could be due to the in activation of β-glucanases. In addition to this, studies
have also shown that when oat flour is mixed with wheat or rye flour for baking purposes or for
the preparation of pasta, a reduction of Mw and extractability are recorded, which is most likely
due to the β‐glucanases in wheat and rye (Degutyte-Fomins et al., 2002; Johansson et al., 2007).
This is probably why evidence exists that baking has a detrimental effect on the cholesterol
lowering properties of β‐glucan. For example, an oat β-glucan preparation showed reduced
cholesterol lowering effect when incorporated into a cookie or bread formula compared to the
same oat preparation consumed raw (Kerckhoffs et al., 2003). Gamel et al. (2015) showed that
different bread making protocols can affect the Mw of β‐glucan processing. In particular, they
showed that a shorter contact time between β‐glucan and the β‐glucanases in the wheat flower,
combined with a lower pH, translated into a reduced depolymerization of β‐glucan.
A study by Vaikousi et al. (2004) investigated the variations in the viscosity of acidic media
containing barley β-glucan during thermal processing. The pH used for this study was that
typically found in acidic food products such as fruit and tomato juice (roughly 2.7 to 4.3) and
the processing conditions those typically used in industry for the pasteurization of such products
(temperature ~ 70 to 95°C and time ~ 4 to 75 min). The effects of β-glucan degradation, caused

51
by acidic hydrolysis, were dependent on pH, temperature and time and were more dramatic for
a high Mw (250 kDa) isolate than for a low Mw one (140 kDa), suggesting that liquid products
containing β-glucan of different Mw can have different flow behavior during thermal
processing. The glycosidic linkages of cereal β-glucan have been proven to be stable at low pH
(1.5) and temperatures between 25 and 37°C (Bhatty, 1992; Johansen et al., 1993). Johansson
et al. (2006) reported that no degradation of oat β-glucan could be observed at 37°C with 0.1
M HCl over a 12 h period. At 120°C and with 3M HCl, on the other hand, total hydrolysis to
glucose monomers occurred within 1 h. Hydrolysis at 120°C with 0.1M HCl produced several
different products, most of them with very low Mw (DP 1-7) (Johansson et al., 2006).
3.2 Thermal and mechanical processing
Thermal or thermomechanical modifications of dietary fibres such as β-glucan often occur
during food production (e.g. baking or extrusion). Both very high and very low temperatures
(e.g. baking and freezing) can cause a reduction of viscosity, Mw and extractability of β-glucan
(Beer et al., 1997; Degutyte-Fomins et al., 2002). Beer et al. (1997), through an in vitro
digestion system, showed that the extractability and Mw of β-glucan was lower in baked muffins
compared to that of the oat bran used to make them and also found different extractabilities
depending on the muffin recipe. They also showed that the extractability of β-glucan was
reduced after 8 weeks of storage in the freezer, even if no change in Mw was detected. In another
study, investigating the impact of freezing on the physicochemical properties of β‐glucan and
its effectiveness in lowering postprandial blood glucose response, oat bran muffins were
subjected to several freeze-thaw cycles. It was found that the β‐glucan solubility decreased with
an increasing number of freeze‐thaw cycles, while its Mw distribution decreased slightly (Lan-
Pidhainy et al., 2007). Izydorczyk et al.(2003) also reported that hydrothermal treatments (e.g.
autoclaving and steaming) of barley had no significant effect on extractability of β-glucan, but
actually prevented enzymatic hydrolysis hence improving its ability to from viscous slurries.
Hydrothermal treatments of barley grains were therefore suggested as a potential pretreatment
before incorporation of the grain into food systems.
Most of the oats used in the food industry are processed into groats, flakes, flour or bran before
being incorporated in other food products or being cooked by the end consumer. The first step
in this process is kilning, which is a treatment that combines high temperature with moisture,
and it is used to inactivate lipases that can cause fat rancidity, significantly reducing shelf life.

52
During this process, also β-glucanases are inactivated, as confirmed in a study by Ames et al.
(2015). In this study, the Mw and viscosity of β-glucan extracted from raw oats resulted to be
significantly lower than those of β-glucan extracted from kilned oat material, which confirms
the inactivation of β-glucanases during this process and which suggests kilning as a key step to
preserve the physicochemical properties of β-glucan.
In terms of mechanical processing, the impact of milling on β‐glucan structure and function
was investigated in several papers, as reviewed by Lazaridou et al. (2007). Jaskari et al. (1995)
for example, extracted the β‐glucan from oat bran for 2h at 37°C and found that the soluble
fraction was higher in dry milled than in wet milled preparations (34% and 24%, respectively).
Heating significantly increased the amount of soluble β-glucan, on average, from 29 to 84%.
Also, the Mw of the extracted soluble fiber was influenced by the milling technique, since oat
bran obtained by dry milling and fiber-concentrated oat bran obtained by wet milling led to
extracted β-glucan averaging 840 kDa and 600 kDa, respectively, and remained unchanged
during hydrothermal treatment (up to 95°C). In another study, the effect of freeze-milling on
the properties of β-glucan from oat bran was investigated (Harasym et al., 2015). The results
show that after freezing and milling in a hammer mill, a significant reduction of particle size
was achieved, since 89% of the material was between 80 and 50 μm in comparison with the
initial material in which 79% was between 250 and 200 μm. Also, following freeze-milling of
the oat fiber, the Mw of the the β-glucan preparation was reduced to 697 kDa form 2180 kDa of
the control β-glucan preparation. Furthermore, an enhanced purity of the β-glucan preparations
was achieved, from 74% of the control to 84% of the frozen-milled samples, which is probably
connected to the lower particle size. More recently, Grundy et al. (2017) showed that the milling
of oat flakes to produce oat flour of smaller particle size increased the rate and extent of release
and dissolution of β-glucan from the cell walls of the oat tissue.
Homogenization techniques are often used in aqueous food applications to enhance product
properties such as structure and stability, in particular for polysaccharides. Kivelä et al. (2010)
investigated the influence of two high pressure homogenizers and a rotor/stator homogenizer
(colloid mill) on the flow and molecular properties of oat β-glucan in semi-dilute and
concentrated solutions. An irreversible decrease in viscosity and loss of shear thinning behavior
was recorded after each homogenization treatment for both solutions. Also, the Mw decreased
together with viscosity after the high pressure homogenization and they both decreased in
parallel with the mechanical energy input. The colloid mill treatments did not result in a
reduction of Mw and the observed reduction in viscosity was probably due to the incorporation

53
of air in the system. Interestingly, as a consequence of the Mw decrease, the shape of β-glucan
became more spherical and dense. Previous studies have shown that the alterations in the
solution properties of polysaccharides caused by high pressure homogenization are normally
linked to the breakage of glycosidic bonds in their backbone (Lagoueyte and Paquin, 1998;
Floury et al., 2002). However, in the study by Kivelä it was not possible to differentiate between
scission of glycosidic bond and simple disassociation of aggregates of β-glucan (Kivelä et al.,
2010).
To examine these structural changes further, Kivelä et al. (2012) performed homogeneization
of highly purified (OBG) and extracted native (NBG) oat β‐glucan solutions with a
microfluidizer at the lowest and highest pressures possible (300 and 1000 bar) in the continuous
mode for 5 min. The formation of carbonyl groups along the chain and the chain cleavage during
the high pressure treatments were investigated by labelling the treated solutions for fluorescent
detection with CCOA (carbazole-9-carboxylic acid [2-(2-aminooxyethoxy)ethoxy]amide)
followed by HPSEC analysis. Homogenisation significantly reduced the Mw in both OBG
(from 260 to 100 kDa) and NBG (from 1600 to 200 kDa) after 5 min of continuous exposure
to 1000 bar, similarly to what reported previously for β-glucan as a result of high pressure
homogenization (Kivelä et al., 2010). The concentration of carbonyl groups was low (5 and 10
µmol/g for NBG and OBG, respectively), but they occurred throughout the molar mass
distribution in NBG, confirming the involvement of oxidative reactions in the homogenization
process. The degradation during the microfluidizer treatment, however, was mainly caused by
the turbulent and elongational forces to which the polysaccharides are subjected under
hydrodynamic pressure and high velocity, respectively. Oxidation, therefore, cannot be
considered the predominant degradation mechanism during high pressure homogenization,
since the mechanical energy input and capillary effect play the central role. In any case,
oxidation reaction might still contribute to the final properties of the homogenized β-glucan.
Extrusion is another common industrial process used for cereals, which involves the
simultaneous application of heat, pressure and shear force. It can be used to create a variety of
products, such as snacks and breakfast cereal, using a big range of conditions. In general, the
Mw of β-glucan contained in extruded commercial oat products, despite the high temperatures
and shear rates used in the extruders, remains high (Åman et al., 2004; Beck et al., 2009; Tosh
et al., 2010). Tosh et al. (2010), however, prepared a series of extruded oat bran cereals with a
difference range of Mw (1930–251 kDa) and modified solubility by varying the temperatures
(181–237 °C) and water contents (7.0–18.7%). The reduced Mw breakfast cereals (251–950

54
kDa) were produced with higher temperature and pressure in the extruder than those used to
produce 1930 kDa cereals. The reduction of Mw from 1930 to 251 kDa caused, as expected, a
dramatic decrease of viscosity, from 2900 to 131 mPa∙s at 30 s-1. Interestingly, when the
extrusion conditions were made more severe, disruption of the cell walls followed and β‐glucan
dispersed throughout the cereal.
Zhang et al. (2009) compared the effect of steam heating, extrusion and superfine grinding on
the solubilisation of oat bran soluble dietary fiber (OSDF). The yield of OSDF increased after
processing and went from 5.9% for the native to 7.2% for the steam heated, 12.3% for the
extruded and 16.6% for the super fine-ground oat bran samples. The solubility of the OSDF
varied greatly among the different treatments, from 47.2% after extrusion to 87.8% after super-
fine grinding. Also, the portion of OSDF with a Mw higher than 500 kDa was 31% in the
untreated, 30 % in the steam heated, 95% in the extruded and 37% in the super-fine ground oat
bran. According to these results, the super-fine grinding and extrusion processes resulted to be
the most effective in enabling the extraction of high Mw OSDF. In another paper, Zhang et al.
(2011) showed that soluble dietary fiber (SDF) extracted from extruded oat bran had higher
yield, mean particle diameter, peak temperature, solubility, swelling capacity, solvent retention
capacity, foam ability, apparent viscosity and consistency coefficient, and lower flow behavior
index compared to the SDF extracted from native oat bran, indicating that extrusion can
improve some properties of oat bran.
3.3 Impact of processing on β-glucan health promoting properties
The physiological benefits of soluble dietary fibers such as β-glucan are normally attributed to
their physico-chemical properties, for example water holding capacity, swelling, the ability to
hinder diffusion through viscosity enhancement and the formation of gels, binding properties,
and susceptibility or resistance to bacterial degradation and fermentation (Lazaridou and
Biliaderis, 2007). The exact mechanisms by which these soluble fibers exert their serum
cholesterol and glucose lowering effects are still under debate but the most common hypothesis
is based on their ability to increase the viscosity of the intestinal lumen (Bell et al., 1999;
Battilana et al., 2001; Wood, 2007; Naumann et al., 2018). Wood et al. (2000) showed that the
viscosity of β-glucan solutions is controlled by its concentration and Mw and therefore food
processing that either lowers solubility or causes depolymerisation of the β-glucan might reduce
physiological effectiveness and must be carefully considered. Lan-Pidhainy et al. (2007)
investigated the impact of freezing of oat bran muffins on the physicochemical and

55
physiological effects of β-glucan. β-Glucan solubility decreased with the increasing number of
freeze-thaw cycles but the Mw only decreased slightly. The peak blood glucose rise (PBGR) of
11 healthy subjects measured after fresh muffins consumption was significantly lower than
after consumption of muffins treated with four freeze-thaw cycles (1.84 ± 0.2 vs. 2.31 ±
0.1 mmol/L). Also, compared with a control of whole wheat muffins, the reduction in the area
under the glucose (AUG) response curve after consuming fresh muffins was nearly double
than that of muffins subjected to 4 freeze-thaw cycles. An inverse linear relationship was found
between the log (concentration) of extractable β-glucan and PBGR and AUC showing that a
reduced β-glucan solubility can attenuate its effectiveness in lowering postprandial glycemia.
It has been shown, however, that certain types of food processing such as cooking or extrusion
can actually improve the physiological activity of β-glucan by increasing its availability (Van
Der Sluijs et al., 1999; Lazaridou and Biliaderis, 2007). Moreover, even though processing can
cause a partial reduction of Mw of the polymer through enzymatic hydrolysis, milling, stirring,
etc., β-glucan can still be able to improve plasma cholesterol and glucose and insulin responses
(Sundberg et al., 1995; Izydorczyk et al., 2000; Yokoyama et al., 2006). For instance, animal
studies have demonstrated that even after reduction of Mw by one order of magnitude (e.g. from
1000 to 200 kDa) β-glucan was still able to attenuate plasma cholesterol (Yokoyama et al.,
2002; Wilson et al., 2004). A human study investigated the attenuation of glucose and insulin
responses following the consumption of glucose solutions containing oat gums in the range of
1.3-10.5 g of pure β-glucan (Wood, Braaten, et al., 1994). An inverse linear relationship
between log(viscosity) and postprandial glucose and insulin responses was found, and
viscosities of the effective oat gum solutions were in the range of 20–8000 mPa∙s (at 30 s-1).
Interestingly, regression analysis of their data showed that even viscosities as low as 10 mPa∙s
resulted in an average 12-13% reduction in the peak plasma glucose.
As for the cholesterol reduction, studies have shown that cereal β-glucan with low or medium
Mw had a better bile acid binding capacity than the high Mw ones. Kim and White (2010)
evaluated the in vitro bile acid binding of oat β-glucan of low (156 kDa) medium (371 kDa)
and high (687 kDa) Mw. With the positive control cholestyramine set at 100% binding, high
Mw β-glucan bound 27%, medium Mw β-glucan 24 % and low Mw β-glucan 21%, indicating
that the lower Mw β-glucan could be more effective in attenuating serum cholesterol. Sayar et
al. (2011) produced low, medium and high Mw oat β-glucan (157, 277, 560 kDa respectively)
through lichenase treatment and added them to plain muffins at a 0.52% final concentration.

56
Also in this case, the bile acid binding capacity of the low and medium Mw β-glucan enriched
muffins was greater than that of the high Mw ones.
Some studies also found a significant difference between the bile acid binding properties of oat
and barley β-glucans, which was suggested to be due to their fine structure (DP3:DP4). In their
paper, Kahlon and Woodruff (2003) investigated the bile acid binding by rice bran, oat bran,
dehulled barley, and β-glucan enriched barley mixing them in vitro with a cocktail of bile acids
at pH of 6.3 and centrifuging the mixture, measuring the unbound bile acids in the supernatant.
Using cholestyramine al positive control and setting it at 100% binding, they showed that on an
equal total dietary fiber and insoluble dietary fiber (IDF) basis rice bran was binding 45 and
49%, oat bran 23 and 30%, dehulled barley 33 and 57%, and β-glucan enriched barley 20 and
40%, respectively. Given the results, they proposed that for oat bran, the main mechanism of
cholesterol lowering was not due to the bile acid binding by its soluble fiber. In general, bile
acid binding was not found to be proportional to the soluble fiber content of the cereal brans
tested but appeared to be relate to the IDF content, with the exception of β-glucan enriched
barley. Wolever et al. (2010), on the other hand, showed in a human study that 3 g of high Mw
(2210 kDa) and medium-Mw (530 kDa) β-glucan had similar effect in lowering LDL
cholesterol, but the effectiveness was decreased by 50% when using low Mw (210 kDa)
β-glucan in extruded breakfast cereals.
Data available suggest that also process induced oxidation may enhance both β-glucan health
benefits and physical properties of foods (De Moura et al., 2011; Lee et al., 2011). The new
functional groups introduced to the β-glucan chain by oxidation increase the solubility of
β-glucan and enhance its interactions with other molecules by van der Waals forces, hydrogen
bonding and ionic interactions. Park et al. (2009) reported that oxidation can increase water
extractability by solubilizing the non-water soluble β-glucan and oxidised β-glucan has also
been shown to inhibit the growth of tumour cells with increased efficacy linked to higher level
of oxidation (Bae et al., 2011). Several studies have focused in particular on the
hypocholesterolaemic properties of oxidised β-glucan. Park et al. (2009), for example,
investigated the effect of a TEMPO-mediated oxidation of β-glucan on the physicochemical
and cholesterol lowering properties of these polysaccharides. The C6-oxidized β-glucan
produced though a TEMPO/NaBr/NaOCl system at pH 10.8 resulted to be more effective in
improving the serum lipid profile of hypercholesterolemic rats compared to the native β-glucan.
This effect was linked to the enhanced bile acid binding by the β-glucan, due to the increased
water solubility of this polysaccharide after oxidation. The solubility in fact, went from 37% in

57
the native β-glucan to 88% with 100% oxidised C6 and was suggested to be a consequence of
the newly formed carboxylic groups on the β-glucan chain. Furthermore, unselective oxidation
induced by a Fenton system (0.9% H2O2 and Fe2+) was demonstrated to increase the bile acid
binding capability of cereal β-glucan from 11% to 16% after a 30 min oxidative treatment,
indicating an increased efficacy in blood cholesterol reduction (De Moura et al., 2011). Also
this oxidation, similarly to the TEMPO induced one, led to the formation of carbonyl and
carboxyl groups along the β-glucan chain (Park et al., 2009; De Moura et al., 2011). Wang et
al. (2011) investigated the physicochemical and health properties of the β-glucan extracted from
the fungus Poria cocos, which is an insoluble β-(1→3)-glucan with little physiological activity.
After a TEMPO-mediated oxidation 95.7% of this insoluble β-glucan became water-soluble.
And exhibited an enhanced in vitro bile acid binding capacity (15.25 µM/mg, dry matter)
compared to the native product (9.31 µM/mg, dry matter).
It is important to clarify that the studies that revealed an increased bile acid binding by oxidized
or low Mw β-glucan are based on centrifugation methods (Park et al., 2009; Kim and White,
2010; De Moura et al., 2011), in which the sample is centrifuged to separate the solid phase
with bound bile acids from the non-bound, dissolved bile acids. These methods, however, while
suitable for the evaluation of bile acid retention by insoluble dietary fiber, are not indicated for
the soluble fiber (Gunness et al., 2012). Since reduction of Mw or oxidation translates into an
increased solubility, in fact, even though the time and temperature of incubation might not lead
to a complete solubilization of β-glucan, they allow for gelation, and as a consequence for the
bile acid to be trapped in a viscous gelled layer of oxidized β-glucan. Therefore, since native
and modified extracts possess very different properties in this respect, the extent of
solubilization of the fiber will vary dramatically, rendering the samples difficult to compare.
Dialysis based methods have been used in recent years to study the impact of β-glucan of
different Mw or oxidation level on bile acid binding and, being more accurate and more similar
to the in vivo mechanisms, have allowed to obtain more reliable results. (Han et al., 2009;
Zacherl et al., 2011; Gunness et al., 2012; Naumann et al., 2018). All of these studies have
shown that viscosity is the key factor in bile acid binding by cereal β-glucan, therefore higher
Mw and/or higher concentrations are extremely important for the soluble fiber to exert its
hypocholesterolemic effect.

58
References
ADDIN Mendeley Bibliography CSL_BIBLIOGRAPHYXÅman, P., Rimsten, L. and Andersson, R. (2004) ‘Molecular Weight Distribution of β-Glucan in Oat-Based Foods’, Cereal Chemistry. doi: 10.1094/CCHEM.2004.81.3.356.
Ames, N., Storsley, J. and Tosh, S. (2015) ‘Effects of Processing on Physicochemical Properties and Efficacy of β-Glucan from Oat and Barley’, Cereal Foods World, 60(1), pp. 4–8. doi: 10.1094/CFW-60-1-0004.
Anderson, J. W., Spencer, D. B., Hamilton, C. C., Smith, S. F., Tietyen, J., Bryant, C. A. and Oeltgen, P. (1990) ‘Oat-bran cereal lowers serum total and LDL cholesterol in hypercholesterolemic men’, The American Journal of Clinical Nutrition, 52(3), pp. 495–499. doi: 10.1093/ajcn/52.3.495.
Anderson, J. W., Story, L., Sieling, B., Chen, W. J. L., Petro, M. S. and Story, J. (1984) ‘Hypocholesterolemic effects of oat-bran or bean intake for hypercholesterolemic men’, The American Journal of Clinical Nutrition, 40(6), pp. 1146–1155. doi: 10.1093/ajcn/40.6.1146.
Armand, M. (2007) ‘Lipases and lipolysis in the human digestive tract: where do we stand?’, Current Opinion in Clinical Nutrition and Metabolic Care, 10(2), pp. 156–164. doi: 10.1097/MCO.0b013e3280177687.
Bae, I. Y., Kim, H. Y., Lee, S. and Lee, H. G. (2011) ‘Effect of the degree of oxidation on the physicochemical and biological properties of Grifola frondosa polysaccharides’, Carbohydrate Polymers. Elsevier Ltd., 83(3), pp. 1298–1302. doi: 10.1016/j.carbpol.2010.09.037.
Balakrishnan, B., Lesieur, S., Labarre, D. and Jayakrishnan, A. (2005) ‘Periodate oxidation of sodium alginate in water and in ethanol-water mixture: A comparative study’, Carbohydrate Research, 340(7), pp. 1425–1429. doi: 10.1016/j.carres.2005.02.028.
Barone Lumaga, R., Azzali, D., Fogliano, V., Scalfi, L. and Vitaglione, P. (2012) ‘Sugar and dietary fibre composition influence, by different hormonal response, the satiating capacity of a fruit-based and a β-glucan-enriched beverage’, Food and Function, 3(1), pp. 67–75. doi: 10.1039/c1fo10065c.
Battilana, P., Ornstein, K., Minehira, K., Schwarz, J. M., Acheson, K., Schneiter, P., Burri, J., Jéquier, E. and Tappy, L. (2001) ‘Mechanisms of action of beta-glucan in postprandial glucose metabolism in healthy men.’, European journal of clinical nutrition, 55(5), pp. 327–333. doi: 10.1038/sj.ejcn.1601160.
Beer, M. U., Arrigoni, E. and Amadò, R. (1995) Effect of oat gum on blood cholesterol levels in healthy young men, European journal of clinical nutrition.
Beer, M. U., Wood, P. J. and Weisz, J. (1997) ‘Molecular Weight Distribution and (1→3)(1→4)-β-Glucan Content of Consecutive Extracts of Various Oat and Barley Cultivars’, Cereal Chemistry Journal, 74(4), pp. 476–480. doi: 10.1094/CCHEM.1997.74.4.476.
Beer, M. U., Wood, P. J., Weisz, J. and Fillion, N. (1997) ‘Effect of cooking and storage on the amount and molecular weight of (1→3)(1→4)-β-D-glucan extracted from oat products by an in vitro digestion system’, Cereal Chemistry. doi: 10.1094/CCHEM.1997.74.6.705.
Bell, S., Goldman, V. M., Bistrian, B. R., Arnold, A. H., Ostroff, G. and Forse, R. A. (1999) ‘Effect of β-Glucan from Oats and Yeast on Serum Lipids’, Critical Reviews in Food Science and Nutrition, 39(2), pp. 189–202. doi: 10.1080/10408399908500493.
Bengmark, S. (2003) ‘Use of some pre-, pro- and synbiotics in critically ill patients’, Bailliere’s Best Practice and Research in Clinical Gastroenterology. doi: 10.1016/S1521-6918(03)00073-8.
Bhatty, R. S. (1992) ‘Total and extractable β-glucan contents of oats and their relationship to viscosity’, Journal of Cereal Science. doi: 10.1016/S0733-5210(09)80070-2.
Biliaderis, C. G. and Izydorczyk, M. S. (2006) Functional food carbohydrates. CRC Press.
Böhm, N. and Kulicke, W.-M. (1999) ‘Rheological studies of barley (1→3)(1→4)-β-glucan in

59
concentrated solution: mechanistic and kinetic investigation of the gel formation’, Carbohydrate Research, 315(3–4), pp. 302–311. doi: 10.1016/S0008-6215(99)00036-1.
Boulos, S. and Nyström, L. (2016) ‘UPLC-MS/MS investigation of β-glucan oligosaccharide oxidation’, Analyst. Royal Society of Chemistry, 141(24), pp. 6533–6548. doi: 10.1039/c6an01125j.
Boulos, S. and Nyström, L. (2017) ‘Complementary Sample Preparation Strategies for Analysis of Cereal β-Glucan Oxidation Products by UPLC-MS/MS’, Frontiers in Chemistry, 5, pp. 157–208. doi: 10.3389/fchem.2017.00090.
Bourdon, I., Yokoyama, W., Davis, P., Hudson, C., Backus, R., Richter, D., Knuckles, B. and Schneeman, B. O. (1999) ‘Postprandial lipid, glucose, insulin, and cholecystokinin responses in men fed barley pasta enriched with β-glucan’, The American Journal of Clinical Nutrition, 69(1), pp. 55–63. doi: 10.1093/ajcn/69.1.55.
Braaten, J. T., Wood, P. J., Scott, F. W., Wolynetz, M. S., Lowe, M. K., Bradley-White, P. and Collins, M. W. (1994) ‘Oat beta-glucan reduces blood cholesterol concentration in hypercholesterolemic subjects’, European journal of clinical nutrition, 48(7), p. 465—474. Available at: http://europepmc.org/abstract/MED/7956987.
Burkus, Z. and Temelli, F. (1998) ‘Effect of Extraction Conditions on Yield , Composition , and Viscosity Stability of Barley β -Glucan Gum’, 75(6), pp. 805–809. doi: 10.1094/CCHEM.1998.75.6.805.
CAMIRE, A. L. and CLYDESDALE, F. M. (1981) ‘Effect of pH and Heat Treatment on the Binding of Calcium, Magnesium, Zinc, and Iron to Wheat Bran and Fractions of Dietary Fiber’, Journal of Food Science, 46(2), pp. 548–551. doi: 10.1111/j.1365-2621.1981.tb04907.x.
Cavallero, A., Empilli, S., Brighenti, F. and Stanca, A. M. (2002) ‘High (1→3,1→4)-β-Glucan Barley Fractions in Bread Making and their Effects on Human Glycemic Response’, 36, pp. 59–66. doi: 10.1006/jcrs.2002.0454.
Chang, P. S. and Robyt, J. F. (1996) ‘Oxidation of primary alcohol groups of naturally occurring polysaccharides with 2,2,6,6-tetramethyl-1-piperidine oxoammonium ion’, Journal of Carbohydrate Chemistry. doi: 10.1080/07328309608005694.
Chen, W.-J. L., Anderson, J. W. and Jennings, D. (2013) ‘Propionate May Mediate the Hypocholesterolemic Effects of Certain Soluble Plant Fibers in Cholesterol-Fed Rats’, Experimental Biology and Medicine. doi: 10.3181/00379727-175-41791.
Choromanska, A., Kulbacka, J., Rembialkowska, N., Pilat, J., Oledzki, R., Harasym, J. and Saczko, J. (2015) ‘Anticancer properties of low molecular weight oat beta-glucan - An in vitro study’, International Journal of Biological Macromolecules. Elsevier B.V., 80, pp. 23–28. doi: 10.1016/j.ijbiomac.2015.05.035.
Coseri, S., Biliuta, G., Zemljič, L. F., Srndovic, J. S., Larsson, P. T., Strnad, S., Kreže, T., Naderi, A. and Lindström, T. (2015) ‘One-shot carboxylation of microcrystalline cellulose in the presence of nitroxyl radicals and sodium periodate’, RSC Advances, 5(104), pp. 85889–85897. doi: 10.1039/C5RA16183E.
Cui, S. W. and Wang, Q. (2009) ‘Cell wall polysaccharides in cereals: chemical structures and functional properties’, Structural Chemistry, 20(2), pp. 291–297. doi: 10.1007/s11224-009-9441-0.
Daou, C. and Zhang, H. (2012) ‘Oat Beta-Glucan: Its Role in Health Promotion and Prevention of Diseases’, Comprehensive Reviews in Food Science and Food Safety, 11(4), pp. 355–365. doi: 10.1111/j.1541-4337.2012.00189.x.
Davy, B. M., Davy, K. P., Ho, R. C., Beske, S. D., Davrath, L. R. and Melby, C. L. (2002) ‘High-fiber oat cereal compared with wheat cereal consumption favorably alters LDL-cholesterol subclass and particle numbers in middle-aged and older men 1 – 3’, pp. 1–8.
Degutyte-Fomins, L., Sontag-Strohm, T. and Salovaara, H. (2002) ‘Oat Bran Fermentation by Rye Sourdough’, Cereal Chemistry Journal, 79(3), pp. 345–348. doi: 10.1094/CCHEM.2002.79.3.345.

60
Dongowski, G. (2007) ‘Interactions between dietary fibre-rich preparations and glycoconjugated bile acids in vitro’, Food Chemistry, 104(1), pp. 390–397. doi: 10.1016/j.foodchem.2006.11.053.
Dongowski, G., Huth, M., Gebhardt, E. and Flamme, W. (2002) ‘Dietary fiber-rich barley products beneficially affect the intestinal tract of rats.’, The Journal of nutrition. doi: 10.1093/jn/132.12.3704.
Ellegård, L. and Andersson, H. (2007) ‘Oat bran rapidly increases bile acid excretion and bile acid synthesis: an ileostomy study’, European Journal of Clinical Nutrition, 61(8), pp. 938–945. doi: 10.1038/sj.ejcn.1602607.
Estrada, A., Yun, C. H., Van Kessel, A., Li, B., Hauta, S. and Laarveld, B. (1997) ‘Immunomodulatory activities of oat β-glucan in vitro and in vivo’, Microbiology and Immunology, 41(12), pp. 991–998. doi: 10.1111/j.1348-0421.1997.tb01959.x.
Faure, A. M., Knüsel, R. and Nyström, L. (2013) ‘Effect of the temperature on the degradation of β-glucan promoted by iron(II)’, Bioactive Carbohydrates and Dietary Fibre. doi: 10.1016/j.bcdf.2013.09.003.
Faure, A. M., Koppenol, W. H. and Nyström, L. (2015) ‘Iron(II) binding by cereal beta-glucan.’, Carbohydrate polymers, 115, pp. 739–43. doi: 10.1016/j.carbpol.2014.07.038.
Faure, A. M., Sánchez-Ferrer, A., Zabara, A., Andersen, M. L. and Nyström, L. (2014) ‘Modulating the structural properties of β-D-glucan degradation products by alternative reaction pathways.’, Carbohydrate polymers, 99, pp. 679–86. doi: 10.1016/j.carbpol.2013.08.022.
Faure, A. M., Werder, J. and Nyström, L. (2013) ‘Reactive oxygen species responsible for beta-glucan degradation’, Food Chemistry. Elsevier Ltd, 141(1), pp. 589–596. doi: 10.1016/j.foodchem.2013.02.096.
FDA (1997) ‘Food Labeling: Health Claims; Oats and Coronary Heart Disease’, Federal register, 62(15), pp. 202–205.
Fenton, H. J. H. (1894) ‘LXXIII.—Oxidation of tartaric acid in presence of iron’, J. Chem. Soc., Trans., 65, pp. 899–910. doi: 10.1039/CT8946500899.
Fincher, G. B. and Stone, B. A. (1986) ‘Cell walls and their components in cereal grain technology.’, in Advances in Cereal Science and Technology, Vol. VIII.
Floury, J., Desrumaux, A., Axelos, M. A. V. and Legrand, J. (2002) ‘Degradation of methylcellulose during ultra-high pressure homogenisation’, Food Hydrocolloids. doi: 10.1016/S0268-005X(01)00039-X.
Fry, S. C. (1998) ‘Oxidative scission of plant cell wall polysaccharides by ascorbate-induced hydroxyl radicals’, Biochemical Journal, 332(2), pp. 507–515. doi: 10.1042/bj3320507.
Grundy, M. M. L., Quint, J., Rieder, A., Ballance, S., Dreiss, C. A., Butterworth, P. J. and Ellis, P. R. (2017) ‘Impact of hydrothermal and mechanical processing on dissolution kinetics and rheology of oat β-glucan’, Carbohydrate Polymers. doi: 10.1016/j.carbpol.2017.02.077.
Gunness, P., Flanagan, B. M., Shelat, K., Gilbert, R. G. and Gidley, M. J. (2012) ‘Kinetic analysis of bile salt passage across a dialysis membrane in the presence of cereal soluble dietary fibre polymers’, Food Chemistry. Elsevier Ltd, 134(4), pp. 2007–2013. doi: 10.1016/j.foodchem.2012.03.131.
Gunness, P. and Gidley, M. J. (2010) ‘Mechanisms underlying the cholesterol-lowering properties of soluble dietary fibre polysaccharides’, Food & Function, 1(2), p. 149. doi: 10.1039/c0fo00080a.
Hallfrisch, J. and Behall, K. M. (2000) ‘Mechanisms of the Effects of Grains on Insulin and Glucose Responses’, 19(3), pp. 320–325.
Hallfrisch, J., Scholfield, D. J. and Behall, K. M. (1995) ‘Diets containing soluble oat extracts improve glucose and insulin responses of moderately hypercholesterolemic men and women’, American Journal of Clinical Nutrition, 61(2), pp. 379–384.
Han, S.-H., Lee, S.-W. and Rhee, C. (2009) ‘Effect of heat treatment of digestion-resistant fraction from soybean on retarding of bile acid transport in vitro’, Nutrition Research and Practice, 3(2), p.

61
149. doi: 10.4162/nrp.2009.3.2.149.
Harasym, J., Suchecka, D. and Gromadzka-Ostrowska, J. (2015) ‘Effect of size reduction by freeze-milling on processing properties of beta-glucan oat bran’, Journal of Cereal Science, 61, pp. 119–125. doi: 10.1016/j.jcs.2014.10.010.
Hirota, M., Tamura, N., Saito, T. and Isogai, A. (2009) ‘Oxidation of regenerated cellulose with NaClO2 catalyzed by TEMPO and NaClO under acid-neutral conditions’, Carbohydrate Polymers. doi: 10.1016/j.carbpol.2009.04.012.
Hofmann, A. F. (1999) ‘Bile Acids : The Good , the Bad , and the Ugly’, News in physiologiacal sciences, 14, pp. 24–29. doi: 10.1152/physiologyonline.1999.14.1.24.
International, A. A. of C. C. (2001) The Definition of Dietary Fiber.
Isogai, A., Saito, T. and Fukuzumi, H. (2011) ‘TEMPO-oxidized cellulose nanofibers’, Nanoscale, 3(1), pp. 71–85. doi: 10.1039/C0NR00583E.
Izydorczyk, M S and Biliaderis, C. G. (2000) ‘Structural and functional aspects of cereal arabinoxylans and b-glucans’, Novel Macromolecules in Food Systems, pp. 361–384.
Izydorczyk, M. S., Jacobs, M. and Dexter, J. E. (2003) ‘Distribution and Structural Variation of Nonstarch Polysaccharides in Milling Fractions of Hull-less Barley with Variable Amylose Content’, Cereal Chemistry. doi: 10.1094/CCHEM.2003.80.6.645.
Izydorczyk, M. S., Macri, L. J. and Macgregor, A. W. (1998) ‘Structure and physicochemical properties of barley extractable P-glucans and arabinoxylans ’’, 8617(97), pp. 259–269.
Izydorczyk, M. S., Storsley, J., Labossiere, D., MacGregor, A. W. and Rossnagel, B. G. (2000) ‘Variation in total and soluble β-glucan content in hulless barley: Effects of thermal, physical, and enzymic treatments’, Journal of Agricultural and Food Chemistry. doi: 10.1021/jf991102f.
Jaskari, J., Henriksson, K., Nieminen, A., Suortti, T., Salovaara, H. and Poutanen, K. (1995) ‘Effect of Hydrothermal and Enzymatic Treatments on the Viscous Behavior of Dry-Milled and Wet-Milled Oat Brans’, Cereal Chemistry, 72(6), pp. 625–631.
Jaskari, J., Kontula, P., Siitonen, A., Jousimies-Somer, H., Mattila-Sandholm, T. and Poutanen, K. (1998) ‘Oat β-glucan and xylan hydrolysates as selective substrates for Bifidobacterium and Lactobacillus strains’, Applied Microbiology and Biotechnology. doi: 10.1007/s002530051155.
Jenkins, D. J. A., Kendall, C. W. C., Augustin, L. S. A., Franceschi, S., Hamidi, M. and Marchie, A. (2002) ‘Glycemic index : overview of implications in health and disease 1 – 4’, 76, pp. 266–273.
Johansen, H. N., Wood, P. J. and Knudsen, K. E. B. (1993) ‘Molecular weight changes in the (1.fwdarw.3)(1.fwdarw.4)-.beta.-D-glucan of oats incurred by the digestive processes in the upper gastrointestinal tract of pigs’, Journal of Agricultural and Food Chemistry, 41(12), pp. 2347–2352. doi: 10.1021/jf00036a025.
Johansson, L., Virkki, L., Anttila, H., Esselström, H., Tuomainen, P. and Sontag-Strohm, T. (2006) ‘Hydrolysis of β-glucan’, Food Chemistry, 97(1), pp. 71–79. doi: 10.1016/j.foodchem.2005.03.031.
Jones, P. J. (2002) ‘Clinical nutrition: 7. Functional foods — more than just nutrition’, 166(12), pp. 1555–1563.
Judd, P. A. and Ellis, P. R. (2005) Traditional Medicines for Modern Times, Traditional Medicines for Modern Times: Antidiabetic Plants. Edited by A. Soumyanath. CRC Press. doi: 10.1201/9781420019001.
Kahlon, T. S. and Woodruff, C. L. (2003) ‘In vitro binding of bile acids by rice bran, oat bran, barley and β-glucan enriched barley’, Cereal Chemistry. doi: 10.1094/CCHEM.2003.80.3.260.
Kim, H. J. and White, P. J. (2010) ‘In Vitro Bile-Acid Binding and Fermentation of High, Medium, and Low Molecular Weight β-Glucan’, Journal of Agricultural and Food Chemistry, 58(1), pp. 628–634. doi: 10.1021/jf902508t.
Kim, H., Stote, K. S., Behall, K. M., Spears, K. and Vinyard, B. (2009) ‘Glucose and insulin responses

62
to whole grain breakfasts varying in soluble fiber , b -glucan’, pp. 170–175. doi: 10.1007/s00394-009-0778-3.
Kivelä, Reetta, Gates, F. and Sontag-Strohm, T. (2009) ‘Degradation of cereal beta-glucan by ascorbic acid induced oxygen radicals’, Journal of Cereal Science, 49(1), pp. 1–3. doi: 10.1016/j.jcs.2008.09.003.
Kivelä, R., Henniges, U., Sontag-Strohm, T. and Potthast, A. (2012) ‘Oxidation of oat β-glucan in aqueous solutions during processing’, Carbohydrate Polymers, 87(1), pp. 589–597. doi: 10.1016/j.carbpol.2011.08.028.
Kivelä, R., Nyström, L., Salovaara, H. and Sontag-Strohm, T. (2009) ‘Role of oxidative cleavage and acid hydrolysis of oat beta-glucan in modelled beverage conditions’, Journal of Cereal Science, 50(2), pp. 190–197. doi: 10.1016/j.jcs.2009.04.012.
Kivelä, R., Pitkänen, L., Laine, P., Aseyev, V. and Sontag-Strohm, T. (2010a) ‘Influence of homogenisation on the solution properties of oat β-glucan’, Food Hydrocolloids, 24(6–7), pp. 611–618. doi: 10.1016/j.foodhyd.2010.02.008.
Kivelä, R., Sontag-Strohm, T., Loponen, J., Tuomainen, P. and Nyström, L. (2011) ‘Oxidative and radical mediated cleavage of β-glucan in thermal treatments’, Carbohydrate Polymers, 85(3), pp. 645–652. doi: 10.1016/j.carbpol.2011.03.026.
Kristiansen, K. A., Potthast, A. and Christensen, B. E. (2010) ‘Periodate oxidation of polysaccharides for modification of chemical and physical properties’, Carbohydrate Research. Elsevier Ltd, 345(10), pp. 1264–1271. doi: 10.1016/j.carres.2010.02.011.
Kwong, M. G. Y., Wolever, T. M. S., Brummer, Y. and Tosh, S. M. (2013) ‘Attenuation of glycemic responses by oat β-glucan solutions and viscoelastic gels is dependent on molecular weight distribution’, Food & function, 4, pp. 401–408. doi: 10.1039/c2fo30202k.
Lagoueyte, N. and Paquin, P. (1998) ‘Effects of microfluidization on the functional properties of xanthan gum’, Food Hydrocolloids. doi: 10.1016/S0268-005X(98)00004-6.
Lakshmanan, M. R., Nepokroeff, C. M., Ness, G. C., Dugan, R. E. and Porter, J. W. (1973) ‘Stimulation by insulin of rat liver β-hydroxy-β-methylglutaryl coenzyme A reductase and cholesterol-synthesizing activities’, Biochemical and Biophysical Research Communications, 50(3), pp. 704–710. doi: 10.1016/0006-291X(73)91301-6.
Lan-Pidhainy, X., Brummer, Y., Tosh, S. M., Wolever, T. M. and Wood, P. J. (2007a) ‘Reducing beta-glucan solubility in oat bran muffins by freeze-thaw treatment attenuates its hypoglycemic effect’, Cereal Chemistry, 84(5), pp. 512–517. doi: 10.1094/CCHEM-84-5-0512.
Lan-Pidhainy, X., Brummer, Y., Tosh, S. M., Wolever, T. M. and Wood, P. J. (2007b) ‘Reducing beta-glucan solubility in oat bran muffins by freeze-thaw treatment attenuates its hypoglycemic effect’, Cereal Chemistry. doi: 10.1094/CCHEM-84-5-0512.
Lazaridou, A., Biliaderis, C. ., Micha-Screttas, M. and Steele, B. . (2004) ‘A comparative study on structure–function relations of mixed-linkage (1→3), (1→4) linear β-d-glucans’, Food Hydrocolloids, 18(5), pp. 837–855. doi: 10.1016/j.foodhyd.2004.01.002.
Lazaridou, A. and Biliaderis, C. G. (2007) ‘Molecular aspects of cereal β-glucan functionality: Physical properties, technological applications and physiological effects’, Journal of Cereal Science, 46(2), pp. 101–118. doi: 10.1016/j.jcs.2007.05.003.
Lazaridou, A., Biliaderis, C. G. and Izydorczyk, M. S. (2003) ‘Molecular size effects on rheological properties of oat β-glucans in solution and gels’, Food Hydrocolloids, 17(5), pp. 693–712. doi: 10.1016/S0268-005X(03)00036-5.
Lazaridou, A., Biliaderis, C. G. and Izydorczyk, M. S. (2007) ‘Cereal β-Glucans: Structures, Physical Properties, and Physiological Functions’, in Functional food carbohydrates, pp. 1–72.
Lee, K. Y., Park, S. Y. and Lee, H. G. (2011) ‘Effect of oat β-glucan and its oxidised derivative on the quality characteristics of sponge cake’, International Journal of Food Science & Technology, 46(12),

63
pp. 2663–2668. doi: 10.1111/j.1365-2621.2011.02798.x.
Lindsay, S. E. and Fry, S. C. (2007) ‘Redox and wall-restructuring’, Plant Cell Monographs. doi: 10.1007/7089_2006_075.
Marcil, V., Delvin, E., Seidman, E., Poitras, L., Zoltowska, M., Garofalo, C. and Levy, E. (2015) ‘Modulation of lipid synthesis, apolipoprotein biogenesis, and lipoprotein assembly by butyrate’, American Journal of Physiology-Gastrointestinal and Liver Physiology. doi: 10.1152/ajpgi.00440.2001.
Marlett, J. A. and Fischer, M. H. (2002) ‘Nutrient Metabolism A Poorly Fermented Gel from Psyllium Seed Husk Increases Excreta Moisture and Bile Acid Excretion in Rats’, The Journal of nutrition, 132(9), pp. 2638–2643.
McIntosh, G. H., Whyte, J., McArthur, R. and Nestel, P. J. (1991) ‘Barley and wheat foods: influence on plasma cholesterol concentrations in hypercholesterolemic men’, The American Journal of Clinical Nutrition, 53(5), pp. 1205–1209. doi: 10.1093/ajcn/53.5.1205.
Michels, A. J. and Frei, B. (2013) ‘Myths, artifacts, and fatal flaws: Identifying limitations and opportunities in vitamin C research’, Nutrients. doi: 10.3390/nu5125161.
De Moura, F. A., Pereira, J. M., Da Silva, D. O., Zavareze, E. D. R., Da Silveira Moreira, A., Helbig, E. and Dias, A. R. G. (2011) ‘Effects of oxidative treatment on the physicochemical, rheological and functional properties of oat β-glucan’, Food Chemistry. Elsevier Ltd, 128(4), pp. 982–987. doi: 10.1016/j.foodchem.2011.04.003.
Naumann, S., Schweiggert-Weisz, U., Bader-Mittermaier, S., Haller, D. and Eisner, P. (2018) ‘Differentiation of adsorptive and viscous effects of dietary fibres on bile acid release by means of in vitro digestion and dialysis’, International Journal of Molecular Sciences, 19(8). doi: 10.3390/ijms19082193.
Östman, E., Rossi, E., Larsson, H., Brighenti, F. and Björck, I. (2006) ‘Glucose and insulin responses in healthy men to barley bread with different levels of (1→3;1→4)-β-glucans; predictions using fluidity measurements of in vitro enzyme digests’, Journal of Cereal Science, 43(2), pp. 230–235. doi: 10.1016/j.jcs.2005.11.001.
Park, S. Y., Bae, I. Y., Lee, S. and Lee, H. G. (2009) ‘Physicochemical and Hypocholesterolemic Characterization of Oxidized Oat β-Glucan’, Journal of Agricultural and Food Chemistry, 57(2), pp. 439–443. doi: 10.1021/jf802811b.
Pentikäinen, S., Karhunen, L., Flander, L., Katina, K., Meynier, A., Aymard, P., Vinoy, S. and Poutanen, K. (2014) ‘Enrichment of biscuits and juice with oat β-glucan enhances postprandial satiety’, Appetite, 75, pp. 150–156. doi: 10.1016/j.appet.2014.01.002.
Persson, H., Nyman, M., Liljeberg, H., Önning, G. and Frølich, W. (1991) ‘Binding of mineral elements by dietary fibre components in cereals—In vitro (III)’, Food Chemistry, 40(2), pp. 169–183. doi: 10.1016/0308-8146(91)90100-3.
Pierre, G., Punta, C., Delattre, C., Melone, L., Dubessay, P., Fiorati, A., Pastori, N., Galante, Y. M. and Michaud, P. (2017) ‘TEMPO-mediated oxidation of polysaccharides: An ongoing story’, Carbohydrate Polymers. doi: 10.1016/j.carbpol.2017.02.028.
Platt, S. R. and Clydesdale, F. M. (1984) ‘Binding of Iron by Cellulose, Lignin, Sodium Phytate and Beta-Glucan, Alone and in Combination, Under Simulated Gastrointestinal pH Conditions’, Journal of Food Science. Blackwell Publishing Ltd, 49(2), pp. 531–535. doi: 10.1111/j.1365-2621.1984.tb12460.x.
Potter, J. G., Coffman, K. P., Reid, R. L., Krall, J. M. and Albrink, M. J. (1981) ‘Effect of test meals of varying dietary fiber content on plasma insulin and glucose response’, The American Journal of Clinical Nutrition, 34(3), pp. 328–334. doi: 10.1093/ajcn/34.3.328.
Radulian, G., Rusu, E., Dragomir, A. and Posea, M. (2009) ‘Metabolic effects of low glycaemic index diets’, 8, pp. 1–8. doi: 10.1186/1475-2891-8-5.

64
Ryu, J., Lee, S., You, S., Shim, J. and Yoo, S. (2012) ‘Effects of barley and oat  -glucan structures on their rheological and thermal characteristics’, Carbohydrate Polymers. Elsevier Ltd., 89(4), pp. 1238–1243. doi: 10.1016/j.carbpol.2012.04.025.
Sayar, S., Jannink, J.-L. and White, P. J. (2011) ‘Textural and Bile Acid-Binding Properties of Muffins Impacted by Oat β-Glucan with Different Molecular Weights’, Cereal Chemistry Journal, 88(6), pp. 564–569. doi: 10.1094/CCHEM-02-11-0014.
Schuchmann, M. N. and von Sonntag, C. (1978) ‘The Effect of Oxygen on the OH-radical-induced Scission of the Glycosidic Linkage of Cellobiose’, International Journal of Radiation Biology and Related Studies in Physics, Chemistry and Medicine, 34(4), pp. 397–400. doi: 10.1080/09553007814551051.
Schuchmann, M. N. and Von Sonntag, C. (1977) ‘Radiation chemistry of carbohydrates. Part 14. Hydroxyl radical induced oxidation of D-glucose in oxygenated aqueous solution’, Journal of the Chemical Society, Perkin Transactions 2. doi: 10.1039/P29770001958.
Skendi, A., Biliaderis, C. G., Lazaridou, A. and Izydorczyk, M. S. (2003) ‘Structure and rheological properties of water soluble β-glucans from oat cultivars of Avena sativa and Avena bysantina’, Journal of Cereal Science. doi: 10.1016/S0733-5210(02)00137-6.
Van Der Sluijs, A. M. C., Behall, K. M., Douglass, L., Prather, E., Scholfield, D. J. and Hallfrisch, J. (1999) ‘Effect of cooking on the beneficial soluble β-glucans in Oatrim’, Cereal Foods World, 44(4), pp. 194–198.
Smith, K. N., Queenan, K. M., Thomas, W., Fulcher, R. G. and Slavin, J. L. (2008) ‘Physiological Effects of Concentrated Barley β-Glucan in Mildly Hypercholesterolemic Adults’, Journal of the American College of Nutrition, 27(3), pp. 434–440. doi: 10.1080/07315724.2008.10719722.
Sundberg, B., Pettersson, D. and Åman, P. (1995) ‘Nutritional properties of fibre‐rich barley products fed to broiler chickens’, Journal of the Science of Food and Agriculture. doi: 10.1002/jsfa.2740670408.
Tamura, N., Hirota, M., Saito, T. and Isogai, A. (2010) ‘Oxidation of curdlan and other polysaccharides by 4-acetamide-TEMPO/NaClO/NaClO2 under acid conditions’, Carbohydrate Polymers. Elsevier Ltd., 81(3), pp. 592–598. doi: 10.1016/j.carbpol.2010.03.016.
Tamura, N., Wada, M. and Isogai, A. (2009) ‘TEMPO-mediated oxidation of (1→3)-β-d-glucans’, Carbohydrate Polymers. Elsevier Ltd, 77(2), pp. 300–305. doi: 10.1016/j.carbpol.2008.12.040.
Tappy, L., Gugolz, E. and Wursch, P. (1996) ‘Effects of Breakfast Cereals Containing Various Amounts of -Glucan Fibers on Plasma Glucose and Insulin Responses in NIDDM Subjects’, Diabetes Care, 19(8), pp. 831–834. doi: 10.2337/diacare.19.8.831.
Theuwissen, E. and Mensink, R. P. (2008) ‘Water-soluble dietary fibers and cardiovascular disease’, Physiology & Behavior, 94(2), pp. 285–292. doi: 10.1016/j.physbeh.2008.01.001.
Todesco, T., Rao, A. V, Bosello, O. and Jenkins, D. J. (1991) ‘Propionate lowers blood glucose and alters lipid metabolism in healthy subjects’, The American Journal of Clinical Nutrition, 54(5), pp. 860–865. doi: 10.1093/ajcn/54.5.860.
Torre, M., Rodriguez, A. R. and Saura-Calixto, F. (1991) ‘Effects of Dietary Fiber and Phytic Acid on Mineral Availability’, Critical Reviews in Food Science and Nutrition, 30(1), pp. 1–22. doi: 10.1080/10408399109527539.
Tosh, S. M., Brummer, Y., Miller, S. S., Regand, A., Defelice, C., Duss, R., Wolever, T. M. S. and Wood, P. J. (2010) ‘Processing Affects the Physicochemical Properties of β-Glucan in Oat Bran Cereal’, Journal of Agricultural and Food Chemistry, 58(13), pp. 7723–7730. doi: 10.1021/jf904553u.
Tosh, S. M., Brummer, Y., Wood, P. J., Wang, Q. and Weisz, J. (2004) ‘Evaluation of structure in the formation of gels by structurally diverse (1→3)(1→4)-β-d-glucans from four cereal and one lichen species’, Carbohydrate Polymers, 57(3), pp. 249–259. doi: http://dx.doi.org/10.1016/j.carbpol.2004.05.009.

65
Vaikousi, H., Biliaderis, C. G. and Izydorczyk, M. S. (2004) ‘Solution flow behavior and gelling properties of water-soluble barley (1 → 3,1 → 4)-β-glucans varying in molecular size’, Journal of Cereal Science, 39(1), pp. 119–137. doi: 10.1016/j.jcs.2003.09.001.
Wang, Q. and Ellis, P. R. (2014) ‘Oat β-glucan: physico-chemical characteristics in relation to its blood-glucose and cholesterol-lowering properties’, British Journal of Nutrition. doi: 10.1017/s0007114514002256.
Wang, Y. J., Maina, N. H., Ekholm, P., Lampi, A. M. and Sontag-Strohm, T. (2017) ‘Retardation of oxidation by residual phytate in purified cereal β-glucans’, Food Hydrocolloids, 66, pp. 161–167. doi: 10.1016/j.foodhyd.2016.11.019.
Wang, Y. J., Zhan, R., Sontag-Strohm, T. and Maina, N. H. (2017) ‘The protective role of phytate in the oxidative degradation of cereal beta-glucans’, Carbohydrate Polymers. Elsevier Ltd., 169, pp. 220–226. doi: 10.1016/j.carbpol.2017.04.016.
Wang, Y., Liu, S., Yang, Z., Zhu, Y., Wu, Y., Huang, J. and Mao, J. (2011) ‘Oxidation of β-glucan extracted from Poria Cocos and its physiological activities’, Carbohydrate Polymers. doi: 10.1016/j.carbpol.2011.03.052.
Wilson, T. A., Nicolosi, R. J., Delaney, B., Chadwell, K., Moolchandani, V., Kotyla, T., Ponduru, S., Zheng, G.-H., Hess, R., Knutson, N., et al. (2004) ‘Reduced and High Molecular Weight Barley β-Glucans Decrease Plasma Total and Non-HDL-Cholesterol in Hypercholesterolemic Syrian Golden Hamsters’, The Journal of Nutrition, 134(10), pp. 2617–2622. doi: 10.1093/jn/134.10.2617.
Wolever, T. M. S., Jenkins, D. J., Jenkins, A. L. and Josse, R. G. (1991) ‘The glycemic index: methodology and clinical implications’, The American Journal of Clinical Nutrition, 54(5), pp. 846–854. doi: 10.1093/ajcn/54.5.846.
Wolever, T. M. S., Spadafora, P. and Eshuis, H. (1991) ‘Interaction between colonic acetate and propionate in humans’, The American Journal of Clinical Nutrition, 53(3), pp. 681–687. doi: 10.1093/ajcn/53.3.681.
Wolever, T. M. S., Tosh, S. M., Gibbs, A. L., Brand-Miller, J., Duncan, A. M., Hart, V., Lamarche, B., Thomson, B. A., Duss, R. and Wood, P. J. (2010) ‘Physicochemical properties of oat β-glucan influence its ability to reduce serum LDL cholesterol in humans: a randomized clinical trial’, The American Journal of Clinical Nutrition, 92(4), pp. 723–732. doi: 10.3945/ajcn.2010.29174.
Wood, P. J. (2007) ‘Cereal β-glucans in diet and health’, Journal of Cereal Science, 46(3), pp. 230–238. doi: 10.1016/j.jcs.2007.06.012.
Wood, P. J. (2010) ‘REVIEW: Oat and rye -glucan: Properties and function.’, Cereal Chem., 87(4), pp. 315–330.
Wood, P. J., Beer, M. U. and Butler, G. (2000) ‘Evaluation of role of concentration and molecular weight of oat beta-glucan in determining effect of viscosity on plasma glucose and insulin following an oral glucose load’, The British Journal of Nutrition, 84(1), pp. 19–23. doi: S0007114500001185 [pii].
Wood, P. J., Braaten, J. T., Scott, F. W., Riedel, K. D., Wolynetz, M. S. and Collins, M. W. (1994) ‘Effect of dose and modification of viscous properties of oat gum on plasma glucose and insulin following an oral glucose load.’, The British Journal of Nutrition, 72(5), pp. 731–743. doi: 10.1079/BJN19940075.
Wood, P. J., Weisz, J. and Blackwell, B. a. (1991) ‘Molecular characterization of cereal B-D-glucans. Structural analysis of oat B-D-glucan and rapid structural evaluation of B-D-glucans from different sources by high-performance liquid chromatography of oligosaccharides released by lichenase.’, Cereal Chemistry, pp. 31–39.
Wood, P. J., Weisz, J. and Blackwell, B. A. (1994) ‘Structural Studies of (1-3)(1-4)-beta-D-Glucans by 13C-Nuclear Magnetic Resonance Spectroscopy and by Rapid Analysis of Cellulose- Like Regions Using High-Performance Anion- Exchange Chromatography of Oligosaccharides Released by Lichenase.’, Cereal Chem, p. 301.

66
Wood, P. J., Weisz, J., Fedec, P. and Burrows, B. D. (1989) ‘Large-Scale Preparation and Properties of Oat Fractions Enriched in (1-3)(1-4)-Beta-D- Glucan.’, Cereal Chemistry, 66, pp. 97–103.
YOKOYAMA, W. H., KNUCKLES, B. E., STAFFORD, A. and INGLETT, G. (2006) ‘Raw and Processed Oat Ingredients Lower Plasma Cholesterol in the Hamster’, Journal of Food Science, 63(4), pp. 713–715. doi: 10.1111/j.1365-2621.1998.tb15820.x.
Yokoyama, W. H., Knuckles, B. E., Wood, D. and Inglett, G. E. (2002) ‘Food Processing Reduces Size of Soluble Cereal β-Glucan Polymers without Loss of Cholesterol-Reducing Properties’, in, pp. 105–116. doi: 10.1021/bk-2002-0816.ch008.
Zacherl, C., Eisner, P. and Engel, K.-H. (2011) ‘In vitro model to correlate viscosity and bile acid-binding capacity of digested water-soluble and insoluble dietary fibres’, Food Chemistry. Elsevier Ltd, 126(2), pp. 423–428. doi: 10.1016/j.foodchem.2010.10.113.
Zhang, M., Bai, X. and Zhang, Z. (2011) ‘Extrusion process improves the functionality of soluble dietary fiber in oat bran’, Journal of Cereal Science. Elsevier Ltd, 54(1), pp. 98–103. doi: 10.1016/j.jcs.2011.04.001.
Zhang, M., Liang, Y., Pei, Y., Gao, W. and Zhang, Z. (2009) ‘Effect of process on physicochemical properties of oat bran soluble dietary fiber’, Journal of Food Science. doi: 10.1111/j.1750-3841.2009.01324.x.

67
4. Aims of the study
Process-induced changes to the β-glucan structure can deeply affect the technological (such as
viscoelasticity and firmness/chewiness in baked products) and health properties (such as the
attenuation of serum cholesterol and of postprandial blood glucose response, the increase of
satiety and the immunomodulatory activity) of this fiber. The main objectives of this thesis
were the characterization of the modifications of the β-glucan structure as a consequence of
processing and the investigation of the impact of these modifications on some of this fiber’s
health effects. More in detail:
1. Characterization of the modifications of cereal β-glucan structure obtained by food
processes, specifically thermal treatments and extrusion. In particular, investigation of
the oxidation products formed as a consequence of hydroxyl radical induced
degradation by means of carbonyl labelling, followed by analysis with UPLC-MS
coupled with fluorescent detection.
2. Elucidation of the molecular interactions between processed cereal β-glucan and Fe2+
and investigation of the role of phytic acid in this mechanism through an in vitro
digestion model.
3. Investigation of the role of cereal β-glucan in the cholesterol reduction mechanism,
through the study of the interaction of this fiber with bile acids.

68

69
PART 2: RESEARCH PAPERS
Manuscript 1
Elena Marasca, Samy Boulos and Laura Nyström – “Bile acid binding by native and modified
oat and barley β-glucan” – SUBMITTED
Manuscript 2
Elena Marasca, Olivia Wyss, Samy Boulos and Laura Nyström – “In vitro estimation of iron
binding by modified cereal β-glucan extracts”
Manuscript 2
Elena Marasca, Samy Boulos and Laura Nyström – “Investigation of cereal β-glucan
degradation following thermal treatments and extrusion”

70

71
I. Bile acid binding by native and modified oat and barley
β-glucan
Elena Marasca, Samy Boulos and Laura Nyström – SUBMITTED
ABSTRACT
Foods rich in cereal β-glucan are among the most efficient dietary tools to help in controlling
several food related diseases. In particular, it has been shown that a diet rich in soluble dietary
fibers (SDF) such as β-glucan can help reducing serum cholesterol levels, leading to a decreased
risk of cardiovascular disease (CVD). However, β-glucan undergoes different reactions and
modifications during food processing, which alter its viscous properties but also its molecular
interactions with minor food components as well as with the mucosal layer of the
gastrointestinal tract. It has been proposed in the literature that oxidized and lower molecular
weight β-glucans have a higher bile acid binding activity than native β-glucan, being therefore
more effective in lowering serum cholesterol. In this study, the kinetics of passage of a bile salt
mix across a dialysis membrane, in the presence or absence of native and modified oat and
barley β-glucan extracts, were studied to investigate the mechanism of diffusion through the
unstirred water layer of the small intestine. As modifications, TEMPO oxidation, sodium
periodate (NaIO4) oxidation and acid hydrolysis were performed. The experimental data were
fitted using first order kinetics as the simplest model for diffusion under dialysis, in order to
derive rate coefficients (K) that quantify the effectiveness of the native and differently modified
β-glucan in binding the bile acids. Native oat and barley β-glucan extracts, i.e. the most viscous
ones, had the strongest bile acid binding power (smallest rate coefficient), with oat β glucan
retaining bile acids more effectively than barley. TEMPO oxidized and acid hydrolyzed oat and
barley β-glucan extracts were also quite effective in retaining the bile acids, despite their
significantly lower viscosity. Opposite of what was suggested in the literature, oxidation and
reduction of molecular weight do not seem to increase the bile acid binding capability of the
fibers.

72
1. Introduction
Cereal β-glucan (mixed linkage (1→3, 1→4)-β-D-glucan) is a soluble dietary fiber found
predominantly in the cell walls of the endosperm and aleurone layer of oat and barley grains
and in smaller amounts in rye and wheat. β-Glucan is a homopolysaccharide of
D-glucopyranose arranged as blocks of consecutive (1→4)-linked β-D-glucose residues
separated by single (1→3)-linkages. Although most of the cellulose segments are trimers (DP3)
and tetramers (DP4), longer units are also present in the polymeric chains (Wood, 1991;
Izydorczyk et al., 1998; Cui et al., 2000; Lazaridou et al., 2004). β-Glucans have several
positive health effects, in particular blood cholesterol lowering properties, recognized by the
EFSA (2011) as well as the FDA (2009), and blood glucose regulation, as evaluated by EFSA
(2011). In vivo (Wolever et al., 2010) and ex vivo (Carr et al., 2003) experiments have shown
that these properties are due to β-glucan’s ability to form a highly viscous intestinal lumen,
slowing down gastric emptying and absorption of glucose as well as inhibiting the uptake of
food lipids and bile acids (Wood, 2004; Cui et al., 2009). Since the viscosity depends on
molecular weight (Mw) and concentration, the hypothesis is that the higher the Mw, the better
the health properties (Cui et al., 2000). However, the relationships between β-glucan structure
and its resulting biological functions have not yet been fully established. This lack of
information on β-glucan structure-function relationship is still a major limitation for effectively
utilizing β-glucan in functional foods. All food processing causes some degree of damage to
the β-glucan structure, above all chain scission, which leads to a decrease in Mw and, as a
consequence, loss of viscosity (Lazaridou and Biliaderis, 2007; Wood, 2007). Processing can
also result in the oxidation of any of the hydroxyl groups of the glucose monomers, leading to
the formation of carbonyl or carboxyl groups, or even to a ring opening (Kristiansen et al.,
2010; Kivelä et al., 2012). However, some of the data available in the literature suggests that
oxidation may, in fact, enhance β-glucan’s health benefits as well as the physical properties of
β-glucan enriched foods (de Moura et al., 2011; Lee et al., 2011). For example, oxidised
β-glucan from the mushroom Grifola frondosa has been shown to inhibit the growth of tumour
cells with increased efficacy linked to higher level of oxidation (Bae et al., 2011). Therefore,
processing conditions that make favourable changes to the β-glucan structure should be
preferred.
One of the main health effects of cereal β-glucan, shared with other soluble dietary fibres, is its
ability to lower serum cholesterol, leading to a decreased risk of cardiovascular disease. The
main mechanism of action is thought to be the interaction of β-glucan with bile acids in the

73
small intestine, which prevents their re-absorption with a subsequent excess excretion in the
feces (Wood, 2004; Gunness and Gidley, 2010). Bile salts are amphipathic bio-surfactants
synthesized from cholesterol in the liver and stored in the gall bladder as conjugated salts of
glycine or taurine. Above their critical micelle concentration (typically >10 mM) they can easily
agglomerate in bile to form micelles. These micelles solubilize biliary and dietary cholesterol
and form mixed micelles with phosphatidylcholine, which are then excreted from the gall
bladder into the duodenum after meals containing lipids (Gunness et al., 2012). Three different
mechanisms have been proposed to explain the interaction of soluble dietary fibers and bile
acids that lead to a reduction of cholesterol: (i) the fibers may increase the barrier properties of
the unstirred water layer in the small intestine, hindering the absorption of the micelles by the
enterocytes; (ii) fibers and micelles are associated at a molecular level; (iii) the fibers may form
a local matrix that entraps the micelles (Gunness and Gidley, 2010).
Several researchers investigated these mechanisms, trying to clarify how β-glucan interacts
with bile acids and how processing (oxidation/degradation) affects this interaction, obtaining
mixed results. Some studies, based on heterogeneous centrifugation methods, show that
β-glucan oxidation enhances bile acid binding (Park et al., 2009; de Moura et al., 2011) and
that β-glucans with low or medium Mw have a better bile acid binding capacity than the high
Mw ones (Hyun and White, 2010). This seems to exclude viscosity as the main property
regulating bile acid binding by β-glucan, since both oxidised and low Mw β-glucan have lower
viscosity compared to the native and high Mw ones. Other studies, based on dynamic dialysis
methods, show that viscosity is indeed the main factor that affects bile acid binding (Zacherl,
et al., 2011; Gunness et al., 2012). The latter are in line with in vivo results, showing that high
viscosity (high Mw) β-glucan have a greater capacity of lowering LDL-cholesterol than low
viscosity (low Mw) β-glucan preparations (Wolever et al., 2010). However, the effects of
oxidation and degradation of cereal β-glucan on viscosity and bile acid binding have never been
studied with a dynamic, dialysis-based method.
In the present study, the kinetics of passage of a model bile salt mixture across a dialysis
membrane, in the presence and absence of modified and unmodified β-glucan from oat and
barley, were studied as a model for diffusion across the unstirred water layer that lines the small
intestine, in order to evaluate the influence of processing on viscosity and bile acid binding
capacity.

74
Figure 1. Bile salts used for the experiments: a) sodium cholate, b) sodium glycocholate, c) sodium deoxycholate, and d) sodium taurocholate.
5. Materials and methods
5.1 Chemicals and materials
Oat flour (OatWell® 28) and barley flour (Beta®Barley dehulled and untreated) were supplied
by DSM (Kaiseraugst, Switzerland) and Dieckmann Cereals GmbH (Rinteln, Germany),
respectively.
Calcium chloride, glycerol, hydrochloric acid, potassium hydroxide, sodium acetate anhydrous,
sodium azide, sodium chlorites, sodium hypochlorite (available chlorine 10-15%), sodium
nitrate, sodium (meta) periodate, 2,2,6,6-tetramethyl-piperidine 1-oxyl (TEMPO; 98%),
xylanase, pancreatin from porcine pancreas, α-amylase from Bacillus licheniformis (Termamyl
300®L), sodium taurocholate hydrate, sodium deoxycholate, sodium cholate hydrate, sodium
glycocholate were purchased from Sigma Aldrich (Switzerland). Absolute ethanol was
purchased from Fluka (Buchs, Switzerland). β-Glucan assay kit (mixed linkage) was purchased
a) b)
c) d)

75
from Megazyme, Ireland. Ultrapure water (Milli-Q) was used for all the experiments (Merck
Millipore, Merck KGaA, Darmstadt, Germany).
5.2 Oat and barley β-glucan extraction
β-Glucan was extracted and purified from oat and barley flour according to the methods
described by Rieder et al. (2015) and Lazaridou et al. (2004) with some adaptations. Firstly,
25 g of oat flour or 50 g of barey flour were dispersed in 0.5 L of 70% v/v ethanol and incubated
at 80°C for 2 h with shaking every 30 min to extract low-molar mass ethanol-soluble
carbohydrates and to inactivate the majority of endogenous enzymes. The mixture was left to
cool to room temperature and filtered using a sintered filter. The filtrate was discarded and the
residue was re-suspended in 1 L Milli-Q water and stirred overnight at room temperature.
Subsequently, 2.5 mL Termamyl (≥300 U/g) and 50 mg CaCl2 were added and the mixture was
incubated for 1 h at 96°C to hydrolyze starch. After cooling to room temperature, the mixture
was centrifuged (Centrifuge 5810 R, Eppendorf, Germany) (4000 rpm, 15 min, 4 °C), the
supernatant was collected, 10 mL sodium acetate buffer (2.5 M, pH 4.5) was added and the pH
was adjusted to 4.5 with 1 M NaOH. Furthermore, 10 mg xylanase (2500 U/g) was added and
the solution was incubated for 3 h at 40°C to degrade co-extracted arabinoxylans. The pH was
then adjusted to 7 with 1 M NaOH and, following the addition of 50 mg pancreatin, the solution
was incubated at 45 min for 3 h to digest proteins. To inactivate the enzymes, the solution was
heated to 80°C for 45 min. After the mixture was cooled, it was centrifuged (4000 rpm, 15 min,
4 °C) to remove the debris, the supernatant was collected and β-glucan was precipitated with
two volumes of ethanol overnight at 4°C. After filtration using a sintered filter, β-glucan was
collected, frozen, freeze dried (Lyolab BII, LSL Secfroid, Switzerland) for 48 h and milled with
a ball mill (Pulverisette 23, Fritsch, Germany).
5.3 Characterization of the extracts
5.3.1 Total β-glucan content
The total β-glucan content of the oat and barley extracts was determined with the β-glucan
Assay Kit (Mixed Linkage) from Megazyme, Ireland.

76
5.3.2 Protein content
The nitrogen content was determined in triplicates by a TruSpec Micro, composed of a
combination of flow-through methods carrier gas and individual, highly selective infrared (IR)
and thermal conductivity detectors (TruSpec Micro, LECO Corporation, USA). The
approximate protein content was then calculated using the nitrogen conversion factors
determined by Mossé (1990) of 5.36 for the oat and 5.50 for the barley extracts.
5.3.3 Ash content
To measure ash content, 500 mg of oat or barley β-glucan extracts were transferred into
pre-weighed crucibles and burnt into a muffle furnace (model L 51/S, Nabertherm, Germany)
at 600°C for 24 h. The ash content (dry weight basis) was calculated based on the weight
difference of the remaining ash over sample weight x 100%.
5.3.4 β-Glucan DP3 to DP4 ratio
To investigate the fine structure of oat and barley β-glucan, the native extracts were hydrolysed
in triplicates by lichenase and the concentration of the resulting DP3 and DP4 oligomers
measured with high-performance anion exchange chromatography with pulsed amperometric
detection (HPAEC-PAD). To do this, 10 mg of oat or barley β-glucan extract were dispersed in
0.2 mL 50% ethanol, then 4 mL of 20 mM NaH2PO4 buffer (pH 6.5) was added and the samples
were boiled for 10 minutes to help dissolution. After cooling to RT, 0.2 mL lichenase solution
(20 U/mL) was added and the samples were incubated for 2 hours at 60°C. Following
inactivation of lichenase by boiling the tubes for 15 minutes, 500 μL of each sample was
transferred to a fresh tube and diluted with 4.5 mL Milli-Q water. Finally, 0.3 mL of the diluted
sample solution was mixed with 0.3 mL water and 0.4 mL 0.06 mM maltotriose (internal
standard). The samples, standard solutions for calibration, water as blank, and maltotriose as
reference were filtered (0.45 μm) in HPLC vials and measured. Calibration curves were created
in the range of 10 – 100 µM for DP3 and 5 – 50 µM for DP4, relative to the area of the internal
standard.
The instrument consisted of a gradient pump (GS50), auto sampler (AS50), thermal
compartment (AS50), pneumatic controller (PC10) and electrochemical detector (ED50), all
from Dionex Bio LC, USA. The column was an Analytical Dionex CarboPac PA1 (2 mm ×

77
250 mm) with a CarboPac PA1 guard column (2 mm × 50 mm). Elution buffers consisted of
150 mM NaOH solution (A) and 5 mM NaOAc with 150 mM NaOH solution (B). The
following elution gradient was used: 90% (A) and 10% (B) from 0 to 2 min, a linear increase
of (B) from 10 to 100% between 2 and 17 min, 100% (B) between 17 and 23 min, and 90% (A)
and 10% (B) from 23 to 37 min. The flow rate was 0.25 mL/min, except between 23 and 36
min, when it was increased to 0.75 mL/min. The resulting peaks were analyzed using
Chromeleon, version 6.8, also by Dionex Bio LC, USA.
5.3.5 Molecular weight determination
The weight average molecular weight (Mw) of the oat and barley β-glucan extracts, native and
modified, was determined in triplicates by means of size exclusion chromatography (SEC). The
extracts were dissolved in eluent (0.1 M sodium nitrate and 0.02% (w/w) sodium azide) to a
final concentration of 0.1% (w/v), then filtered (0.45 μm) in HPLC vials.
Two different systems were used for analysis: the first one, suitable for Mw up to 1’000 kDa,
was an OMNISEC Resolve (pump, degasser, autosampler and column oven) coupled with an
OMNISEC Reveal (integrated multi-detector module incorporating refractive index, UV/Vis
absorbance, light scattering and intrinsic viscosity detectors), both by Malvern (Malvern
Instruments, U.K.). The column system was made of a pre-column (Viscotek AGuard Col. 50
x 6.0 mm, Malvern Instruments Ltd, UK) connected to two A6000 columns in series (Viscotek,
300 x 8 mm, Malvern Instruments Ltd, UK). The temperature of the column compartment was
kept at 30°C, the flow rate was 0.7 mL/min and the injection volume 50 μL. PEO (Mw = 24’063
g/mol) and Dextran (Mw = 70’026 g/mol) were used as standards for calibration. The average
Mw of the samples were calculated using the OMNISEC software (version 5.1, Malvern
Instruments Ltd, UK).
The other system, used for samples with Mw above 1’000 kDa, consisted of a HPLC equipped
with a binary pump, degasser, thermostated column compartment and auto sampler, all from
HP (Series 1100, Hewlett Packard, USA). The column system was made of a pre-column
(Viscotek AGuard Col. 50 x 6.0 mm, Malvern Instruments Ltd, UK) connected to a A5000
column (Viscotek, 300 x 7.8 mm, Malvern Instruments Ltd, UK) and a Suprema 30000 column
(10 μm, 8 x 300 mm, PSS Polymer Standards Service GmbH, Germany). The temperature of
the column compartment was kept at 35°C, the flow rate was 1 mL/min and the injection
volume 50 μL. Samples were analyzed with a refractive index detector (Series 1200, Agilent

78
Technologies, AG, Switzerland). For the calibration curve, 0.1% (w/v) solutions of β-glucan
standards with reported Mp of 33’600, 67’100, 187’100, 247’000, 375’000, and 667’000 g/mol
(Megazyme, Ireland) were prepared and filtered (0.45 μm) in HPLC vials. The average Mw
were calculated based on the measured standard curve with the ChemStation software
(ChemStation for LC 3D systems) and the add-on Cirrus GPC/SEC software (version 3.4.1)
from Agilent.
5.3.6 Rheological measurements
The viscosities of the mixtures used for the bile acid binding experiment (1.33% accounting for
the purity of each β-glucan extract) were measured using an AR-2000 rheometer (TA
Instruments, New Castle, DE). A cone and plate geometry was used with a plate radius of
40 mm and a cone angle of 2°. The gap between the cone and plate geometry was set at 59 µm.
The flow curves were obtained over a shear rate range of 20–2000 s-1, at 37°C.
5.3.7 FT-IR measurements
Fourier-transform infrared (FTIR) spectra of freeze-dried samples were recorded at room
temperature with a Varian 640 FTIR spectrometer (Agilent Technologies AG, Basel,
Switzerland) and a MKII Golden Gate single Attenuated Total Reflection (ATR) system.
5.4 β-Glucan structural modifications
5.4.1 Acid hydrolysis
The pH of 1% (w/v) oat or barley β-glucan solutions was adjusted to 1.0 with 1 M HCl. The
samples were then incubated at 50°C and, after 24 hours, cooled to RT and neutralized with 1M
NaOH. The partially hydrolyzed β-glucan was precipitated with two volumes of ethanol then
stored at 4°C for 48 hours to help the precipitation. Finally, the samples were centrifuged (10
minutes, 4000 rpm, 4 °C), the solids freeze dried and milled by ball mill.
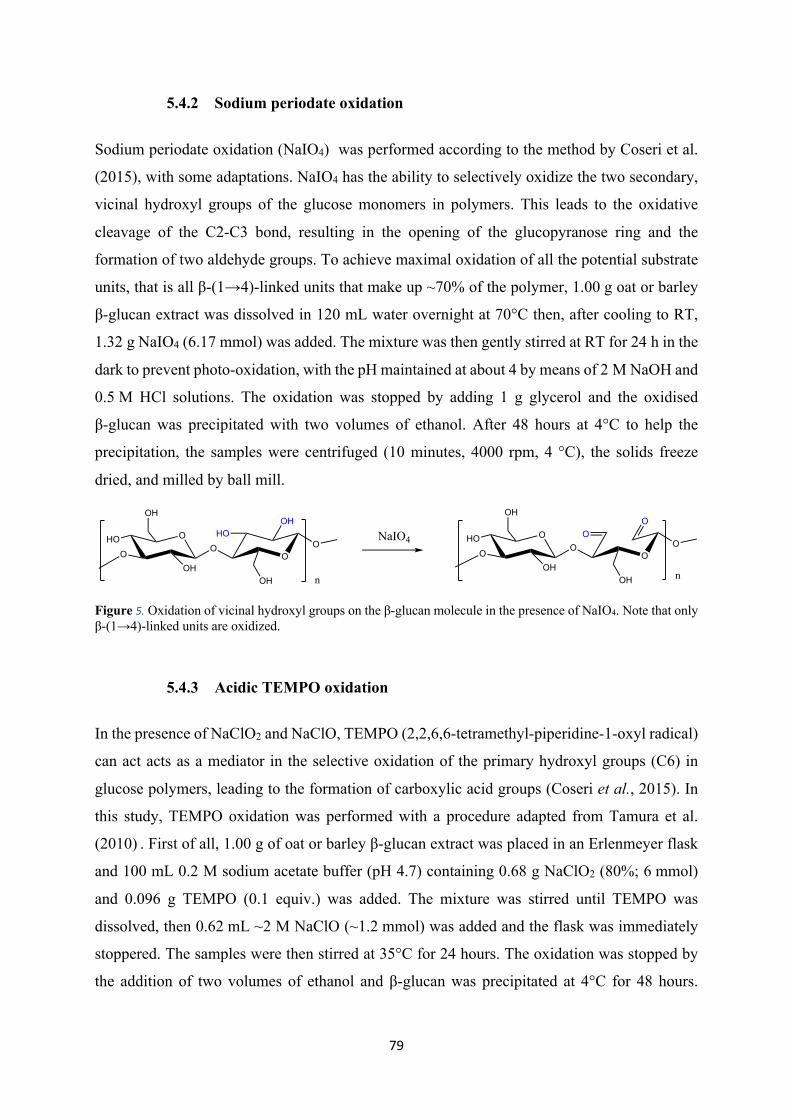
79
5.4.2 Sodium periodate oxidation
Sodium periodate oxidation (NaIO4) was performed according to the method by Coseri et al.
(2015), with some adaptations. NaIO4 has the ability to selectively oxidize the two secondary,
vicinal hydroxyl groups of the glucose monomers in polymers. This leads to the oxidative
cleavage of the C2-C3 bond, resulting in the opening of the glucopyranose ring and the
formation of two aldehyde groups. To achieve maximal oxidation of all the potential substrate
units, that is all β-(1→4)-linked units that make up ~70% of the polymer, 1.00 g oat or barley
β-glucan extract was dissolved in 120 mL water overnight at 70°C then, after cooling to RT,
1.32 g NaIO4 (6.17 mmol) was added. The mixture was then gently stirred at RT for 24 h in the
dark to prevent photo-oxidation, with the pH maintained at about 4 by means of 2 M NaOH and
0.5 M HCl solutions. The oxidation was stopped by adding 1 g glycerol and the oxidised
β-glucan was precipitated with two volumes of ethanol. After 48 hours at 4°C to help the
precipitation, the samples were centrifuged (10 minutes, 4000 rpm, 4 °C), the solids freeze
dried, and milled by ball mill.
Figure 5. Oxidation of vicinal hydroxyl groups on the β-glucan molecule in the presence of NaIO4. Note that only β-(1→4)-linked units are oxidized.
5.4.3 Acidic TEMPO oxidation
In the presence of NaClO2 and NaClO, TEMPO (2,2,6,6-tetramethyl-piperidine-1-oxyl radical)
can act acts as a mediator in the selective oxidation of the primary hydroxyl groups (C6) in
glucose polymers, leading to the formation of carboxylic acid groups (Coseri et al., 2015). In
this study, TEMPO oxidation was performed with a procedure adapted from Tamura et al.
(2010) . First of all, 1.00 g of oat or barley β-glucan extract was placed in an Erlenmeyer flask
and 100 mL 0.2 M sodium acetate buffer (pH 4.7) containing 0.68 g NaClO2 (80%; 6 mmol)
and 0.096 g TEMPO (0.1 equiv.) was added. The mixture was stirred until TEMPO was
dissolved, then 0.62 mL ~2 M NaClO (~1.2 mmol) was added and the flask was immediately
stoppered. The samples were then stirred at 35°C for 24 hours. The oxidation was stopped by
the addition of two volumes of ethanol and β-glucan was precipitated at 4°C for 48 hours.
O
OH
OH
OO
HOOH
OH
ONaIO4 O
OH
OH
OO
OO
OH
OOO
n
HO
n
HO

80
Finally, the samples were centrifuged (10 minutes, 4000 rpm, 4 °C), the solids freeze dried and
milled by ball mill.
Figure 3. Oxidation of primary hydroxyl groups of β-glucan in the presence of nitroxyl radicals (TEMPO).
5.5 Determination of bile acid-binding capacity of native and modified β-glucan
The bile acid binding capability of β-glucan was determined according to the method by
Gunness et al.(2012), with slight adaptations. For this purpose, a 40 mM bile salt mixture was
prepared, consisting of 10 mM sodium cholate, 10 mM sodium deoxycholate, 10 mM sodium
glycocholate, and 10 mM sodium taurocholate dissolved in phosphate buffer (0.1 M, pH 6.9).
β-Glucan from oat or barley, in its native or modified form, was dissolved in quadruplicates at
70°C in the phosphate buffer, to a final β-glucan concentration of 1.33% (prepared accounting
for the purity of each extract). To mimic the barrier properties of the gastrointestinal tract,
centrifuge tubes equipped with a dialysis unit (slide-A-Lyzer MINI Dialysis Device, 3.5K
MWCO, 2mL; Thermo Scientific Pierce Protein Research Products, Rockford, Illinois, United
States) were used. The β-glucan solutions and the bile salt mixture were transferred into the
dialysis unit to reach a final concentration of 1% and 10 mM respectively. The unit was then
placed back into the centrifuge tube, which was previously filled with 44.5 mL buffer, and the
tube was sealed with a screw-top cap (Figure 4). The control was prepared, in triplicates, by
replacing β-glucan solutions with the phosphate buffer. The samples and controls were
incubated with shaking at 200 rpm and 37°C. After 2, 4, 6, 9, 12, 24, and 48 hours of incubation,
50 µL of each dialysate was sampled into an Eppendorf vial, diluted with 100 µL buffer and
stirred. Subsequently, 10 µL of this mixture was transferred into a polystyrene cuvette (1.5 mL)
and analyzed with an enzymatic kit (Total bile acids assay; Diazyme Europe GmbH, Dresden,
Germany) for total bile acid concentration using a UV-spectrophotometer (Cary 100 UV-Vis,
Agilent Technologies, Santa Clara, California). The experimental data were analyzed with
Origin and were fitted using first order kinetics (Box Lucas model, Eq. (1)) as the simplest
model for diffusion under dialysis (Gunness et al., 2012),
NaClO2, NaOCl
N
OO
OH
OH
O OO
OH
OH
OO
COOH
OH
O OO
OH
COOH
OHO HO
HO HO
n n

81
Ct = Cf (1 – e[-Kt]) (1)
where Ct is the concentration of bile salts in dialysate at time t (µM), Cf is the concentration of
bile salts after equilibrium (µM); K is the apparent permeability rate constant (h-1); and t is time
(h).
Figure 4. Experimental setup for the study of the kinetics of diffusion of bile acids, based on a centrifuge tube equipped with a dialysis unit (3.5 kDa MWCO).
5.6 Statistical analysis
Statistical means and standard deviations (SD) were calculated with Microsoft Excel 2016
(Microsoft Cooperation, USA). T-test was used to compare the bile acid binding behaviour of
oat and barley β-glucan extracts, both in the native and modified form. Additionally, univariate
analyses of variance (ANOVA) with post hoc testing including Tukey’s HSD were conducted
to find out more about the effect of the different modifications on bile acid binding. These tests
were performed with SPSS (IBM Statistical Package for Social Science, Chicago, USA). The
threshold for indicating a significant difference was set at the probability level of p < 0.05,
unless otherwise stated.
82
6. Results and discussion
6.1 Extract composition and DP3:DP4 ratio
Table 1 Oat and barley β-glucan extract composition.
The total β-glucan content resulted to be 68.6 ± 2.3% for the oat extract and 54.6 ± 2.2% for
the barley extract, lower than the purities obtained by Rieder et al. (2015) (94% dwb for barley)
and Lazaridou et al. (2004) (>93% dwb for oat). However, in the present study, the extraction
method was simplified and several purification steps (mainly the extensive dialysis and some
precipitation and re-dissolution steps) were left out, which explains the difference in purity.
Protein and ash contents (Table 1) were in line with the results found in the literature for similar
extraction procedures (Bhatty, 1995; Lazaridou et al., 2004; Limberger-Bayer et al., 2014).
Ashes are higher for barley than oat (8.78% vs 5.25%), while proteins are higher for oat than
barley (3.99± 0.08% vs 3.53 ± 0.21%).
The fine structure (DP3:DP4) of the extracted β-glucans was also analysed, by means of
HPAEC-PAD. The fine structure is a crucial characteristic of these polysaccharides, since it
affects their physical properties, such as the water solubility, viscosity, and gelation properties,
but also their behaviour in the gastro-intestinal tract (Lazaridou et al., 2004). As already
mentioned, β-glucans from different cereal sources differ in their DP3:DP4 ratio, following the
order of wheat (3.0–4.5), barley (1.8–3.5), rye (1.9–3.0) and oats (1.5–2.3) (Wood, 2004;
Lazaridou and Biliaderis, 2007). The results for DP3:DP4 (Table 1) are in accordance with the
literature, with barley having a higher DP3:DP4 than oat (2.2 vs 1.5) (Lazaridou et al., 2004;
Lazaridou and Biliaderis, 2007; Izydorczyk and Dexter, 2008).
a means of triplicate measurements ± standard deviation.
Total
β-glucan (% dwb)a
Protein (% dwb)a
Ash (% dwb) DP3:DP4
Oat β-glucan 68.6 ± 2.3 3.99 ± 0.09 5.25 1.5 Barley β-glucan 54.6 ± 2.2 3.53 ± 0.21 8.78 2.2

83
6.2 Molecular size distribution and viscosity of the native and modified samples
Table 2 Weight average molecular mass (Mw) and remaining viscosity (Visc [%]) of native and modified β-glucan. For the determination of the remaining viscosity, a concentration of 1.33% β-glucan (taking the purity of the extracts into account) was used at a shear rate of 20 s-1 (absolute viscosities for the native samples in parenthesis).
The Mw of oat and barley β-glucan samples before and after the modifications was determined
in triplicates using size exclusion chromatography (Table 2). Literature values for the Mw of
cereal β-glucans are very variable because of differences in extraction and measuring methods
(Wood, 2004). However, as confirmed by t-test, there is a significant difference between
unmodified oat and barley β-glucan (p<0.05) with oat β-glucan being slightly larger (about
1600 kDa) than barley β-glucan (1300 kDa), which is in accordance to the literature (Beer,
Wood and Weisz, 1997; Lazaridou et al., 2004; Brennan and Cleary, 2005). As for the modified
samples, the results show that TEMPO treatment reduces the Mw roughly by a factor of six for
oat and seven for barley. TEMPO oxidation was chosen because it is a mild modification that
allows to investigate the effect of oxidation on the kinetics of the bile salts passage without
dramatically reducing the molecular size. Acid hydrolysis led to a stronger degradation and the
molecular size was reduced roughly by nine times. This treatment was used to study the effect
of the reduction of Mw, and hence of the viscosity, on the kinetics of the passage of bile salts,
without any other alteration of the structure through oxidation. With sodium periodate treatment
the molecular size was drastically reduced. According to Kristiansen et al. (2010) periodate
oxidation strongly influences the hydrolytic stability of polysaccharides. The dialdehyde
derivatives formed by oxidation, in fact, are highly susceptible to alkaline β-elimination, which
can lead to the breakage of β-glucan’s backbone structure and, as a consequence, to the low Mw
observed in this study.
The difference in Mw between native oat and barley β-glucan is reflected in the viscosity of
their solutions, with oat being more viscous than barley (around 4.5 and 2.4 Pa·s, respectively).
Mw [kg/mol] Remaining visc. [%] OBG 1584 ± 14 100% (4.47±0.09 Pa·s) BBG 1300 ± 7 100% (2.63±0.09 Pa·s) OBG TEMPO 275 ± 5 3% BBG TEMPO 192 ± 2 3% OBG NaIO4 6 ± 0.2 0.1% BBG NaIO4 6 ± 0.9 0.2% OBG HCl 173 ± 14 1% BBG HCl 136 ± 6 3%

84
As expected, with decreasing Mw of the polysaccharide, there is a decrease in viscosity (Table
2) (Lazaridou, et al., 2003; Lazaridou et al., 2004; Wood, 2007). Despite the fact that the Mw
of TEMPO treated samples is still roughly one sixth of the untreated ones, the viscosity
decreases dramatically: a 97% decrease, in fact, is registered for both oat and barley after
oxidation. As expected, the viscosity is even lower for hydrolysed oat β-glucan, due to the
slightly smaller Mw (99% reduction). However, in the case of barley, there is no significant
difference in viscosity between TEMPO oxidised and hydrolysed samples, despite the lower
Mw of the latter. This could be due to a different relationship of their intrinsic viscosity as a
function of salt concentration, which is known to have little effect on native, neutral
polysaccharides, but to drastically reduce the viscosity for negatively charged uronans
(Spatareanu et al., 2014). The great viscosity decrease in the oxidised and hydrolysed samples
can be explained with the fact that β-glucan is a high Mw polysaccharide with high intrinsic
viscosity which leads to polymer coil overlap and entanglement at low concentrations (0.2–
0.3% w/v), above which flow viscosities increase exponentially with concentration and Mw, to
about the fourth power (Wood, 2004). Due to this exponential relationship, even a small change
in Mw can result in a significant change in viscosity. As expected, sodium periodate oxidised
β-glucans, both from oat and barley, have the lowest viscosity (~ 3 and 4 mPa·s, respectively),
comparable to that of the control (~2 mPa·s), due to their low Mw (>99.5% reduction) compared
to the native β-glucan.
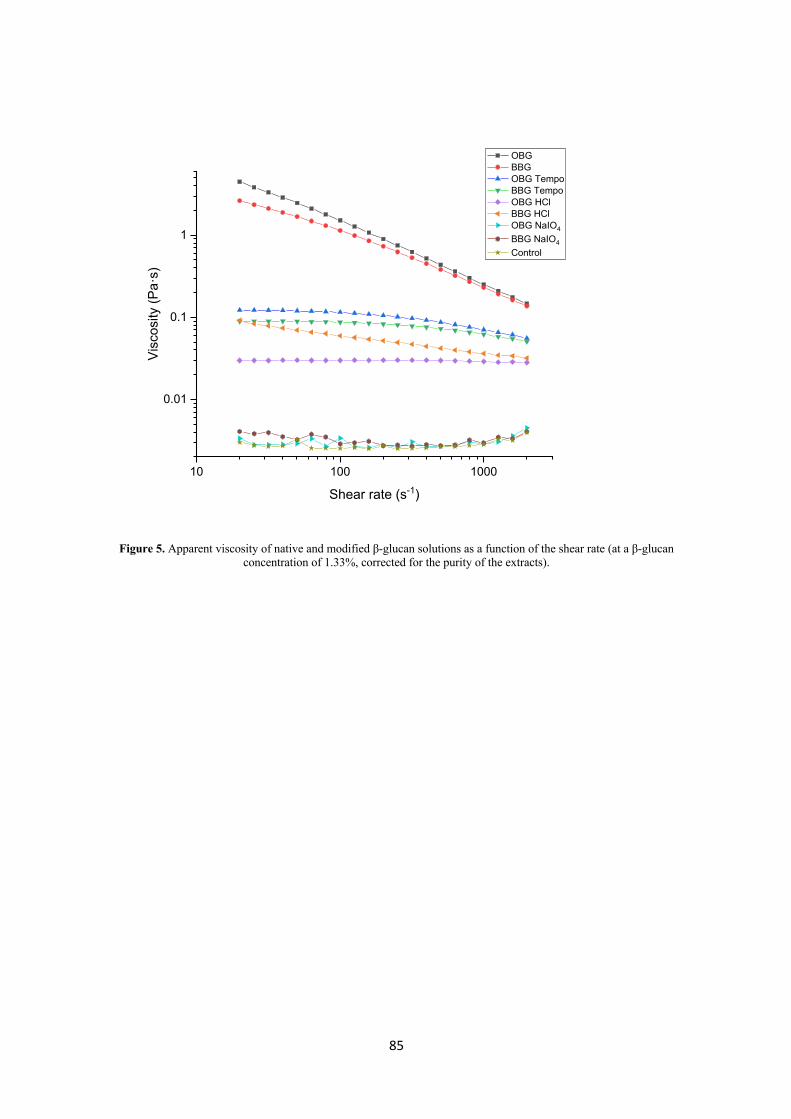
85
Figure 5. Apparent viscosity of native and modified β-glucan solutions as a function of the shear rate (at a β-glucan concentration of 1.33%, corrected for the purity of the extracts).
10 100 1000
0.01
0.1
1
OBG BBG OBG Tempo BBG Tempo OBG HCl BBG HCl OBG NaIO4
BBG NaIO4
Control
Vis
cosi
ty (P
a·s)
Shear rate (s-1)
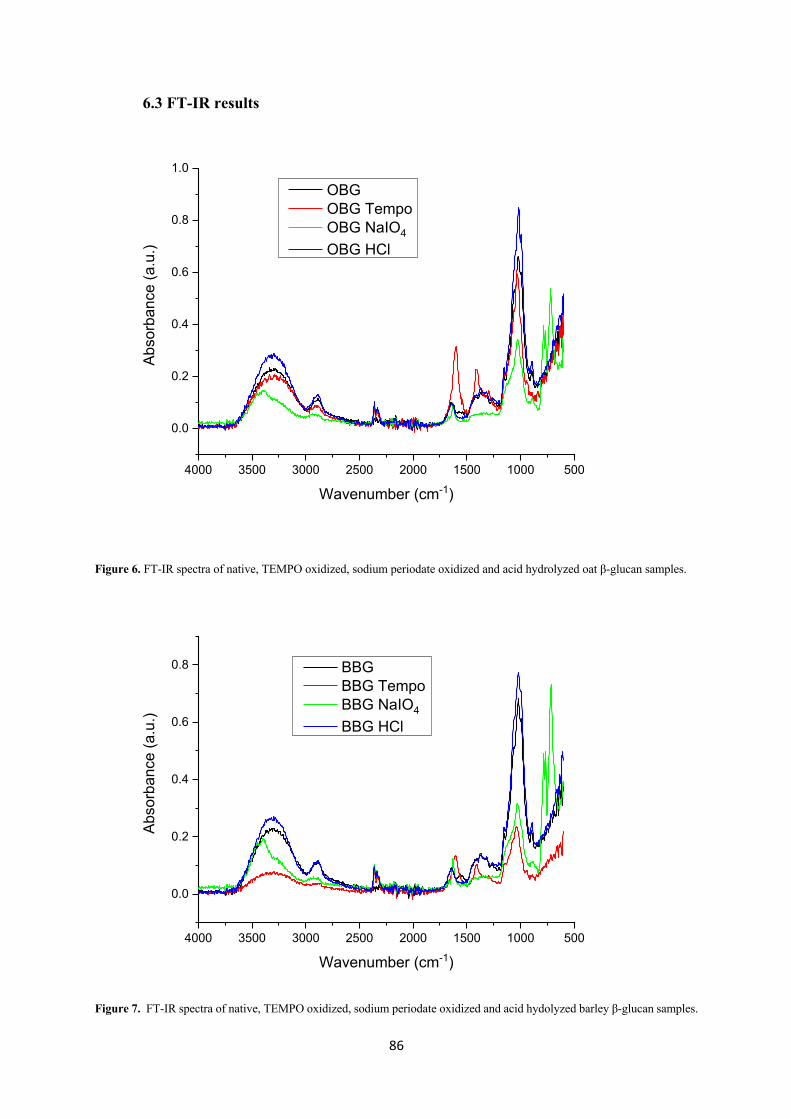
86
6.3 FT-IR results
Figure 6. FT-IR spectra of native, TEMPO oxidized, sodium periodate oxidized and acid hydrolyzed oat β-glucan samples.
Figure 7. FT-IR spectra of native, TEMPO oxidized, sodium periodate oxidized and acid hydolyzed barley β-glucan samples.
4000 3500 3000 2500 2000 1500 1000 500
0.0
0.2
0.4
0.6
0.8
1.0Ab
sorb
ance
(a.u
.)
Wavenumber (cm-1)
OBG OBG Tempo OBG NaIO4
OBG HCl
4000 3500 3000 2500 2000 1500 1000 500
0.0
0.2
0.4
0.6
0.8
Abso
rban
ce (a
.u.)
Wavenumber (cm-1)
BBG BBG Tempo BBG NaIO4
BBG HCl

87
In order to gain a deeper understanding of the changes to the molecular structure after the
modifications, in particular after oxidation, FT-IR analysis was performed on freeze-dried
samples, and Figure 6 and Figure 7 show the IR spectrum of oat and barley β-glucan extracts
before and after the modifications. Two of the most characteristic peaks of glucose polymers,
present in every sample, are the peak around 3400 cm-1 due to hydrogen bonded O–H stretching
and the peak at 2900 cm-1 due to sp3 hybridized C–H stretching. After TEMPO oxidation, a new
peak appears at around 1700 cm-1 in both oat and barley samples, suggesting the alteration in
the chemical structure of the glucose backbone units. (Coseri et al., 2015) This peak is assigned
to the C=O stretching, usually observed between 1740 cm−1 and 1650 cm−1, and it is due to the
oxidation of primary hydroxyl groups forming carboxylic acids. For the sodium periodate
oxidized samples, the formation of aldehyde groups by oxidation of vicinal secondary hydroxyl
groups is difficult to monitor with FT-IR. The absorption peak assigned to the C=O stretching
vibration of aldehyde groups are expected to be localized between 1740 – 1690 cm-1; however,
since these groups are often partially or totally hydrated, the resulting geminal diol or
hemiacetal structures do not show the classical C=O absorption (Fan, et al., 2001; Coseri et al.,
2015). Other characteristic absorption peaks of native polymeric glucose, such as the –OH in-
plane bending at around 1200 cm-1, the C-H deformation stretching vibration at around
1100 cm-1, and the C–O–C stretching vibration of the pyranose ring skeleton at roughly
1050 cm-1 become weaker after oxidation (Coseri et al., 2015). This effect, particularly evident
for oat β-glucan oxidized samples, especially for periodate oxidation, tends to indicate a partial
decomposition of the polymer after the modification (Cai et al., 2017). After sodium periodate
treatment, in fact, two C-H bonds on the ring are substituted by acetal C-H bonds.
As expected, acid hydrolysis, a well-known and commonly used method to degrade
polysaccharides, did not play a major role on the molecular structure of the glucose units. The
spectra (Figure 6 and Figure 7) clearly show that there are no major changes occurring within
the chemical structure of the hydrolysed β-glucan, for both oat and barley.
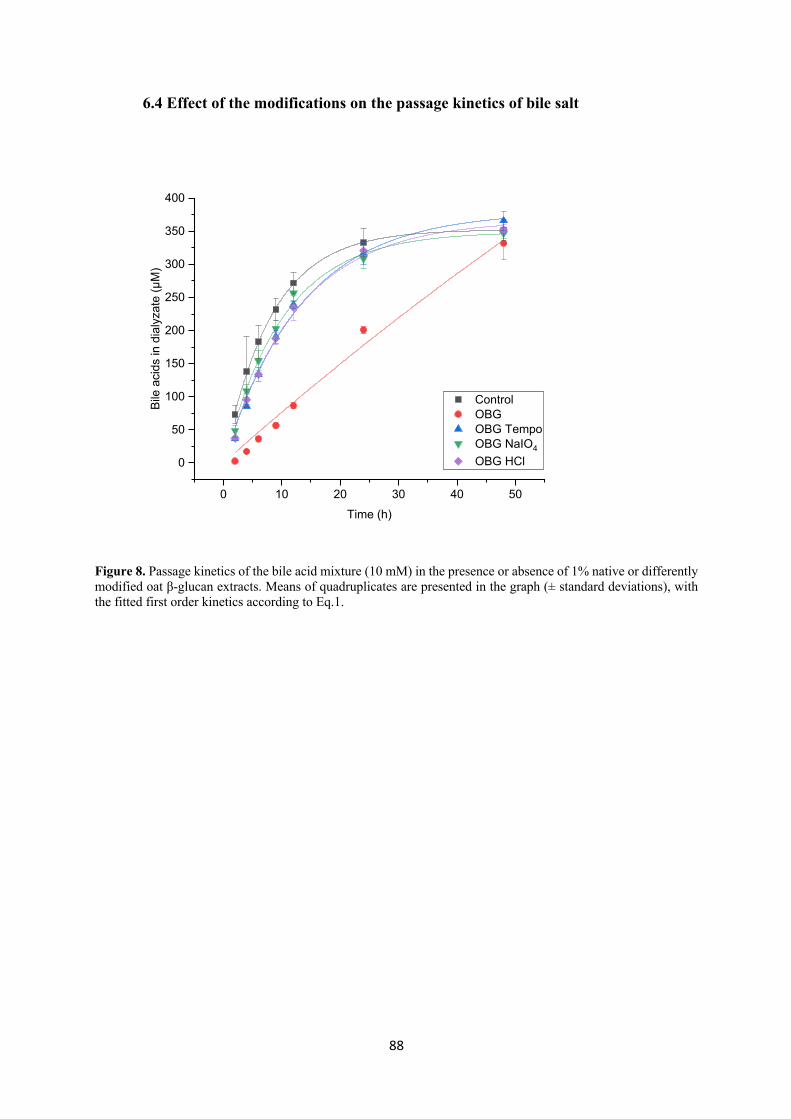
88
6.4 Effect of the modifications on the passage kinetics of bile salt
Figure 8. Passage kinetics of the bile acid mixture (10 mM) in the presence or absence of 1% native or differently modified oat β-glucan extracts. Means of quadruplicates are presented in the graph (± standard deviations), with the fitted first order kinetics according to Eq.1.
0 10 20 30 40 50
0
50
100
150
200
250
300
350
400
Control OBG OBG Tempo OBG NaIO4
OBG HCl
Bile
aci
ds in
dia
lyza
te (μ
M)
Time (h)
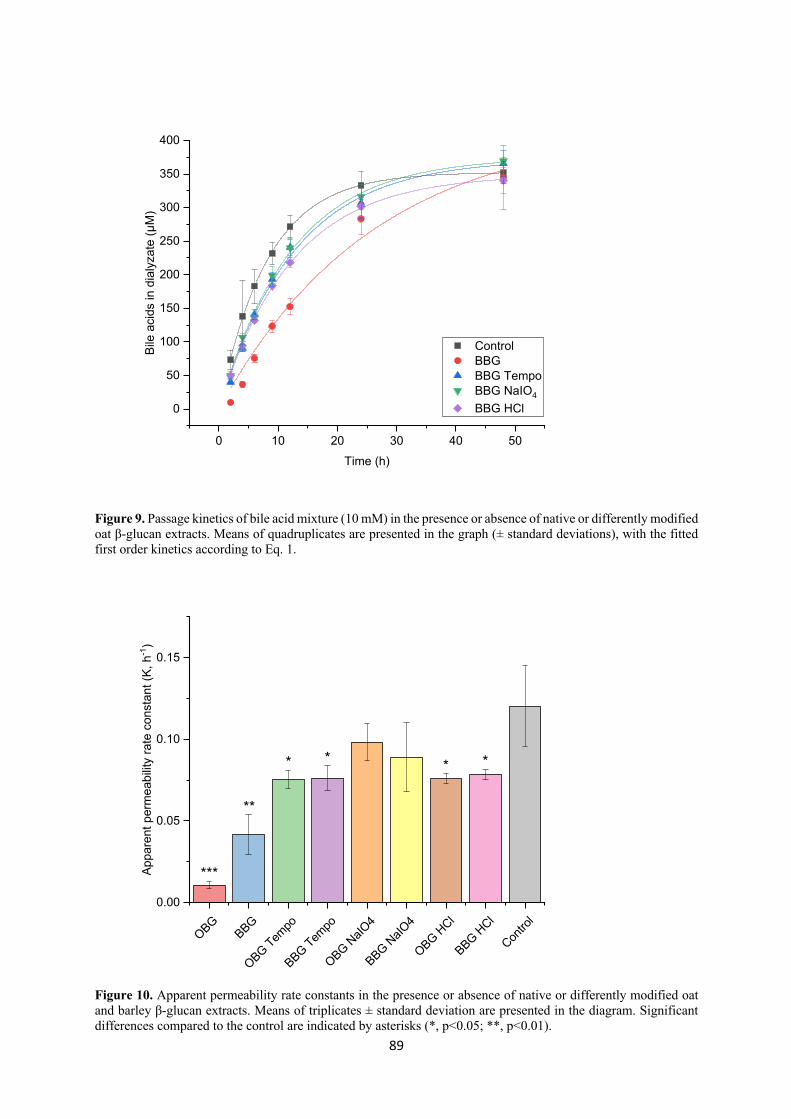
89
Figure 9. Passage kinetics of bile acid mixture (10 mM) in the presence or absence of native or differently modified oat β-glucan extracts. Means of quadruplicates are presented in the graph (± standard deviations), with the fitted first order kinetics according to Eq. 1.
Figure 10. Apparent permeability rate constants in the presence or absence of native or differently modified oat and barley β-glucan extracts. Means of triplicates ± standard deviation are presented in the diagram. Significant differences compared to the control are indicated by asterisks (*, p<0.05; **, p<0.01).
0 10 20 30 40 50
0
50
100
150
200
250
300
350
400
Control BBG BBG Tempo BBG NaIO4
BBG HCl
Bile
aci
ds in
dia
lyza
te (μ
M)
Time (h)
OBGBBG
OBG Tempo
BBG Tempo
OBG NaIO
4
BBG NaIO
4
OBG HCl
BBG HCl
Contro
l0.00
0.05
0.10
0.15
Appa
rent
per
mea
bilit
y ra
te c
onst
ant (
K, h
-1)
***
**
* * **

90
In this study, the kinetics of diffusion of a bile acid mixture across a dialysis membrane, in the
presence or absence of native or modified oat or barley β-glucan extracts, was studied as a
simplified in vitro model of the absorption across the unstirred water layer of the small intestine.
The experimental data were fitted using first-order kinetics (all R2 values were with ≥0.99
excellent) (Eq.1). The rate of diffusion of bile salts through the dialysis membrane was
characterized by the apparent permeability rate constant (K, [h-1]), which is higher the faster
the diffusion. The results obtained for the control show that the permeation of bile salts through
the dialysis membrane increases with dialysis time, reaching a plateau after 48 hours. In the
presence of the native oat and barley β-glucan extracts, the diffusion of the bile salts through
the membrane follows the same first order kinetics of the control, but with a lower diffusion
constant (Figures 8, 9 and 10). The apparent permeability rate constants of OBG (0.011 ±
0.002 h-1) and BBG (0.042 ± 0.012 h-1), in fact, are both significantly different from the constant
of the control (0.120 ± 0.025 h-1) (p<0.01). Also, OBG was more effective than BBG in slowing
down the bile acid release (p<0.01), which correlates with the viscosity of the two types of
extract (about 4.5 Pa·s for OBG and 2.5 Pa·s for BBG): the higher the viscosity, the higher the
retention of bile acids.
TEMPO OBG and TEMPO BBG were also effective in slowing down the bile acid diffusion,
since their K (0.075 ± 0.006 h-1 and 0.076 ± 0.008 h-1, respectively) is significantly different
from the control (p<0.05). Their effect, however, was not as dramatic as that of the native fibers,
possibly due to the much lower viscosity of these samples (97% lower for both oat and barley
after TEMPO oxidation). In this case, though, no significant difference was found between oat
and barley extracts, which also possess very similar viscosities. The same is valid for the
hydrolyzed samples, which also possess similar viscosities and which both significantly slowed
down the bile acid diffusion but not significantly differently from each other (K(OBG HCl)=
0.076 ± 0.003 h-1; K(BBG HCl) = 0.078 ± 0.003 h-1). TEMPO oxidized and hydrolyzed
samples, both from oat and barley, are all not significantly different from each other but all have
an impact on the passage of bile acids through the dialysis membrane. The apparent
permeability rate constants of the periodate oxidized samples (OBG NaIO4: 0.098 ± 0.011 h-1;
BBG NaIO4: 0.089 ± 0.021 h-1), on the other hand, are not significantly different from the
control. Interestingly, despite the fact that the viscosity of the extracts drops dramatically after
the modifications, going from roughly 3.5 Pa·s for the native fibers to below 0.2 Pa·s for the
processed ones (Table 2), both TEMPO oxidized and acid hydrolyzed samples significantly
reduce the diffusion of bile acids through the membrane. These viscosity values, however, are
still significantly higher than of the control and this difference is sufficient to affect the bile
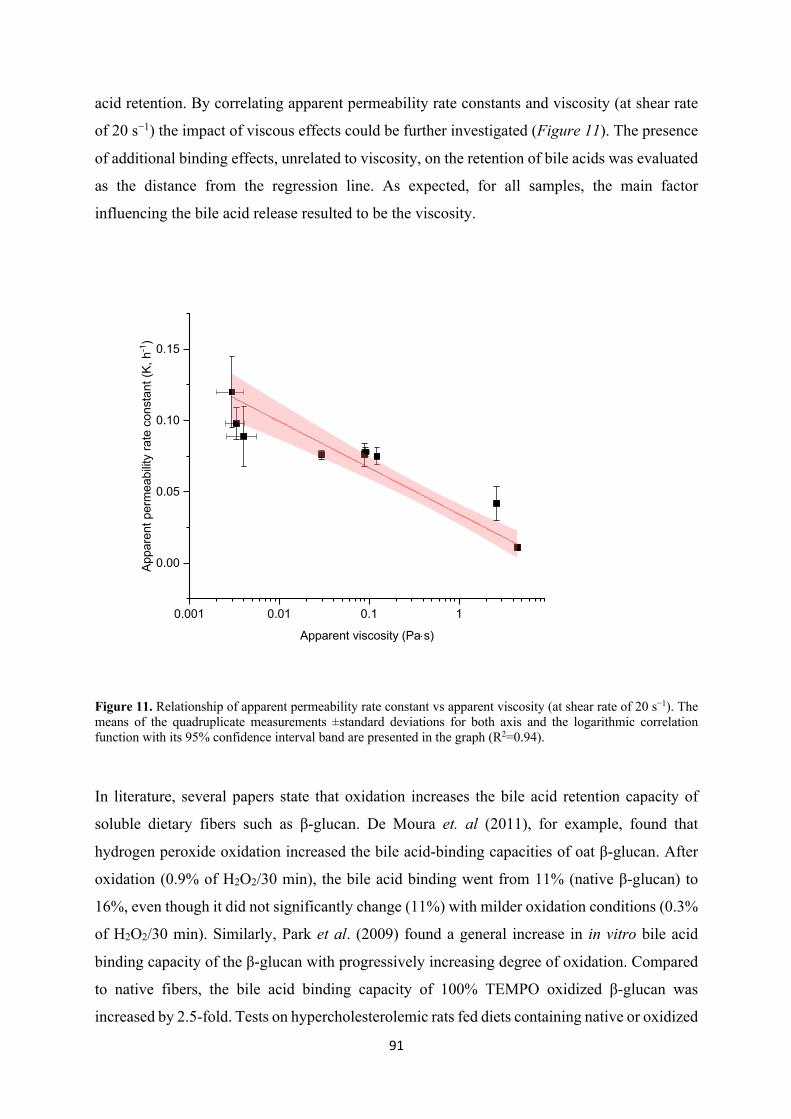
91
acid retention. By correlating apparent permeability rate constants and viscosity (at shear rate
of 20 s−1) the impact of viscous effects could be further investigated (Figure 11). The presence
of additional binding effects, unrelated to viscosity, on the retention of bile acids was evaluated
as the distance from the regression line. As expected, for all samples, the main factor
influencing the bile acid release resulted to be the viscosity.
Figure 11. Relationship of apparent permeability rate constant vs apparent viscosity (at shear rate of 20 s−1). The means of the quadruplicate measurements ±standard deviations for both axis and the logarithmic correlation function with its 95% confidence interval band are presented in the graph (R2=0.94).
In literature, several papers state that oxidation increases the bile acid retention capacity of
soluble dietary fibers such as β-glucan. De Moura et. al (2011), for example, found that
hydrogen peroxide oxidation increased the bile acid-binding capacities of oat β-glucan. After
oxidation (0.9% of H2O2/30 min), the bile acid binding went from 11% (native β-glucan) to
16%, even though it did not significantly change (11%) with milder oxidation conditions (0.3%
of H2O2/30 min). Similarly, Park et al. (2009) found a general increase in in vitro bile acid
binding capacity of the β-glucan with progressively increasing degree of oxidation. Compared
to native fibers, the bile acid binding capacity of 100% TEMPO oxidized β-glucan was
increased by 2.5-fold. Tests on hypercholesterolemic rats fed diets containing native or oxidized
0.001 0.01 0.1 1
0.00
0.05
0.10
0.15
Appa
rent
per
mea
bilit
y ra
te c
onst
ant (
K, h
-1)
Apparent viscosity (Pa×s)

92
β-glucan seemed to confirm that the diets containing the oxidized fibers were more effective at
improving the serum lipid profile of the rats, compared to those containing the native β-glucan.
In another study, the bile acid binding capacities of oat flour and of β-glucan extracts with
different molecular weights (and therefore viscosity) were compared, with cholestyramine as
positive control and cellulose as negative control. The low and medium Mw β-glucans bound
more bile acids than the high Mw ones (high Mw β-glucan: 2.12±0.04; medium and low Mw β-
glucan 2.41±0.12 (μmol/100 mg sample, dwb)). Oat flour, with the lowest β-glucan
concentration, had the greatest bile acid binding ability per amount of β-glucan, which was
hypothesized to indicate that either the oat flour contains other compounds contributing to bile-
acid binding, or that β-glucan can more efficiently bind bile acids in more dilute surroundings
(Kim et al., 2010).
It must be specified, however, that all of the studies that found oxidized or low Mw β-glucan to
have a stronger bile acid binding effect are based on heterogeneous centrifugation methods,
where the sample is centrifuged to separate the solid phase with bound bile acids from the non-
bound, dissolved bile acids. While these methods may be adequate for the evaluation of
insoluble dietary fibers they are, however, not suitable for the measurement of bile acid binding
properties of soluble dietary fibers (Gunness et al., 2012). A prerequisite for accurate
centrifugation measurements is, in fact, that no significant amount of fibers go into solution, or
that at the very least, a constant ratio between fiber supernatants and solids is ensured across all
samples (Naumann et al., 2018). The solubility of low Mw or oxidized β-glucan, however, is
greatly increased compared to the unmodified fibers, and even though the time and temperature
of incubation might not suffice to completely solubilize β-glucan, they allow for gelation, and
hence for the bile acid to be trapped in a viscous gelled layer of oxidized β-glucan. Due to the
different properties of native and modified extracts, the extent of solubilization of β-glucan will
vary significantly, violating the prerequisite and hence rendering the samples difficult to
compare.
On top of this, many of the centrifugation studies have been carried out with bile salts
concentrations far below the CMC, which greatly affects the results due to the impaired
formation of micelles. For all these reasons and for closer similarity to the in vivo mechanisms,
dialysis-based methods allow to obtain more accurate results and have been used in several
studies in recent years (Han et al., 2009; Zacherl et al., 2011; Gunness et al., 2012; Naumann
et al., 2018). Naumann et al. (2018) evaluated the bile acid binding capabilities of dietary fiber-
rich materials from different sources, comparing centrifugation and dialysis methods and using

93
cellulose and cholestyramine as negative and positive control. As confirmed by their findings,
centrifugation methods, due to the experimental setting, can only investigate the adsorptive
properties of the materials, not the viscosity effects, which greatly impairs the comparability
with in vivo studies. They show that in the case of a high fiber barley product the retention of
bile acids is mainly due to viscosity, which is confirmed by the perfect correlation with the
apparent permeability rate constant. These results are also in line with NMR studies indicating
that barley β-glucan does not bind tauroconjugated (Gunness et al., 2010) or glycoconjugated
(Bowles, 1996) bile acids. The additional absorptive effects observed are most probably due to
components of the material other than soluble dietary fiber, such as insoluble fiber or proteins.
This is also supported by the study of Sayar et al. (2011) which described that partial lichenase
hydrolysis did not affect bile acid binding of β-glucan-rich oat fractions using an in vitro
centrifugation method. In our study, no significant difference in the Cf of the different samples,
and therefore no adsorptive effects, were observed, which can be explained with the absence of
insoluble fibers in the extracts.
The results of the present work are also in line with the findings of Gunness et al. (2012), who
investigated the kinetics of diffusion of a model bile salt mix and complete porcine bile across
a dialysis membrane, in the presence or absence of β-glucan and arabinoxylans. In their study,
they found that the transport of bile salts across the dialysis membrane is delayed with
increasing concentration (and viscosity) of the soluble dietary fibers. In fact, they noticed that
the rate coefficient for passage of the bile salts micelles decreased with increasing
concentrations of the fibers, and that the viscosity of the samples increased with increasing
fibers concentrations. In particular, they noticed that viscosity, more than concentration, might
be the key property regulating the rate of dialysis: for example, a 1.67% w/v β-glucan solution
had a lower permeability rate constant K and a higher viscosity than a more concentrated 2.08%
w/v arabinoxylan system. This is also in line with the results of in vivo experiments that showed
that high viscosity (high Mw) β-glucan have a greater capacity of lowering LDL-cholesterol
compared to low viscosity (low Mw) β-glucan preparations (Wolever et al., 2010).
According to the results of the present study, oxidation and hydrolysis do not positively affect
the retention of bile acids, which is consistent with the findings of Zacherl et al. (2011), who
set up a model to investigate the correlation between the bile acid binding capacities of different
fibers, including β-glucan, with their viscosities after in vitro digestion. According to their
study, the bile acid binding capacity of the soluble dietary fibers is mainly due to the high
viscosity of the digested chime, which limits the diffusion of the bile salts through the

94
membrane in the employed model. However, additional experiments involving harsh thermal
processing to reduce the viscosity of the dietary fibers showed that bile acid binding is not solely
dependent on high viscosity and that other factors may contribute to the binding of bile acids
(Zacherl et al., 2011).
When interpreting the results obtained in our study, several aspects should be taken into
account. First of all, it is a simplified in vitro model of the very complex human gastrointestinal
system. The cholesterol lowering action exerted by the soluble dietary fibers in the gut consists
of many phenomena, such as the binding capacity of soluble fibers on neutral sterols, which
may concur to the final result. (Zacherl et al., 2011) Also, studies have shown that soluble
dietary fibers cause an increase in short chain fatty acids (SCFA) such as acetate, propionate
and butyrate, which are all products of their bacterial colonic fermentation. These SCFA,
particularly propionate, may also indirectly cause a decrease in blood cholesterol through
inhibition of hepatic cholesterol synthesis and this mechanism is not considered by the approach
developed in the present study (Gunness and Gidley, 2010).
Nevertheless, this simplified model allowed to evaluate the influence of process induced
degradation effects (e.g. oxidation and hydrolysis) of β-glucan on viscosity and bile acid-
binding capacity. This enabled to develop diffusion kinetics for different processing conditions
of the fibers, thus helping to understand how food processing can affect the health properties of
these polysaccharides.
7. Conclusions
Oat and barley β-glucan extracts were modified (TEMPO oxidation, sodium periodate
oxidation and acid hydrolysis) and characterized. The native and modified fibers were then used
to study the impact of structural changes on the bile acid binding capability of β-glucan, linked
to the reduction of serum cholesterol. The results obtained, based on a dynamic dialysis model
aimed at mimicking the absorption through the unstirred water layer of the small intestine, led
us to reject the hypothesis that oxidation improves the bile acid binding by cereal β-glucan.
High viscosity seems to be the most important property affecting the binding in vitro, as
confirmed by published in vivo studies, even though other factors concurring to the final
cholesterol reduction cannot be ruled out. The native, most viscous samples had the highest bile
acid binding effect, but also TEMPO oxidized and hydrolysed β-glucan also showed significant
bile acid binding behavior despite the lower viscosity. The correlation between apparent

95
permeability rate constant and viscosity indicated no additional adsorptive effects, confirming
that viscosity is the key property affecting bile acid binding by charged or neutral soluble
dietary fiber. The extreme modifications performed in this study were meant to investigate the
maximal effect that any processing (e.g. food production) may have on the health properties of
the fibers. Since no change in the bile acid retention mechanism was recorded, it can be
expected that no change would take place for processed fiber in foods either. Additional studies,
both in vivo and in vitro will be required to investigate this further, in order to find the best
processing conditions of β-glucan to ensure optimal health effects.
8. Acknowledgements
The study was carried out within the framework of the Healthy Nutrition and Sustainable Food
Production project, (National Research Programme NRP 69), funded by the Swiss National
Science Foundation. We thank Dr. Attila Bagdi for the training on the dialysis method, Prof.
Peter Fischer (Laboratory of Food Process Engineering, ETH Zurich) for kindly allowing us to
use the rheometer and Prof. Raffaele Mezzenga (Laboratory of Food and Soft Materials) for
usage of the FT-IR.
9. References
ADDIN Mendeley Bibliography CSL_BIBLIOGRAPHYXBae, I. Y., Kim, H. Y., Lee, S. and Lee, H. G. (2011) ‘Effect of the degree of oxidation on the physicochemical and biological properties of Grifola frondosa polysaccharides’, Carbohydrate Polymers. Elsevier Ltd., 83(3), pp. 1298–1302. doi: 10.1016/j.carbpol.2010.09.037.
Beer, M. U., Wood, P. J. and Weisz, J. (1997) ‘Molecular Weight Distribution and (1→3)(1→4)-β-Glucan Content of Consecutive Extracts of Various Oat and Barley Cultivars’, Cereal Chemistry Journal, 74(4), pp. 476–480. doi: 10.1094/CCHEM.1997.74.4.476.
Bhatty, R. . (1995) ‘Laboratory and pilot plant extraction and purification of β-glucans from hull-less barley and oat brans’, Journal of Cereal Science, 22(2), pp. 163–170. doi: 10.1016/0733-5210(95)90046-2.
Bowles, R. (1996) ‘13C CP/MAS NMR study of the interaction of bile acids with barley β-D-glucan’, Carbohydrate Polymers, 29(1), pp. 7–10. doi: 10.1016/0144-8617(95)00138-7.
Brennan, C. S. and Cleary, L. J. (2005) ‘The potential use of cereal (1→3,1→4)-β-d-glucans as functional food ingredients’, Journal of Cereal Science, 42(1), pp. 1–13. doi: 10.1016/j.jcs.2005.01.002.
Cai, H., Yang, X., Cai, Q., Ren, B., Qiu, H. and Yao, Z. (2017) ‘Lycium barbarum L. Polysaccharide (LBP) Reduces Glucose Uptake via Down-Regulation of SGLT-1 in Caco2 Cell’, Molecules, 22(2), p.

96
341. doi: 10.3390/molecules22020341.
Carr, T. P., Wood, K. J., Hassel, C. A., Bahl, R. and Gallaher, D. D. (2003) ‘Raising intestinal contents viscosity leads to greater excretion of neutral steroids but not bile acids in hamsters and rats’, Nutrition Research, 23(1), pp. 91–102. doi: 10.1016/S0271-5317(02)00476-1.
Coseri, S., Biliuta, G., Zemljič, L. F., Srndovic, J. S., Larsson, P. T., Strnad, S., Kreže, T., Naderi, A. and Lindström, T. (2015) ‘One-shot carboxylation of microcrystalline cellulose in the presence of nitroxyl radicals and sodium periodate’, RSC Advances, 5(104), pp. 85889–85897. doi: 10.1039/C5RA16183E.
Cui, S. W. and Wang, Q. (2009) ‘Cell wall polysaccharides in cereals: chemical structures and functional properties’, Structural Chemistry, 20(2), pp. 291–297. doi: 10.1007/s11224-009-9441-0.
Cui, W., Wood, P. J., Blackwell, B. and Nikiforuk, J. (2000) ‘Physicochemical properties and structural characterization by two-dimensional NMR spectroscopy of wheat β-D-glucan—comparison with other cereal β-D-glucans’, Carbohydrate Polymers, 41(3), pp. 249–258. doi: 10.1016/S0144-8617(99)00143-5.
Efsa (2011) ‘Scientific Opinion on the substantiation of health claims related to beta-glucans from oats and barley and maintenance of normal blood LDL-cholesterol concentrations ( ID 1236 , 1299 ), increase in satiety leading to a reduction in energy intake ( ID 851’, EFSA Journal, 9(6), pp. 1–21. doi: 10.2903/j.efsa.2011.2207.
Fan, Q. G., Lewis, D. M. and Tapley, K. N. (2001) ‘Characterization of Cellulose Aldehyde Using Fourier Transform Infrared Spectroscopy’.
(FDA) F.a.D.A. 21 CFR 101.81 (2009) ‘Health claims: Soluble fiber from certain foods and risk of coronary heart disease (CHD)’, in Regulatory Information, pp. 143–148.
Gunness, P., Flanagan, B. M. and Gidley, M. J. (2010) ‘Molecular interactions between cereal soluble dietary fibre polymers and a model bile salt deduced from 13C NMR titration’, Journal of Cereal Science. Elsevier Ltd, 52(3), pp. 444–449. doi: 10.1016/j.jcs.2010.07.009.
Gunness, P., Flanagan, B. M., Shelat, K., Gilbert, R. G. and Gidley, M. J. (2012) ‘Kinetic analysis of bile salt passage across a dialysis membrane in the presence of cereal soluble dietary fibre polymers’, Food Chemistry. Elsevier Ltd, 134(4), pp. 2007–2013. doi: 10.1016/j.foodchem.2012.03.131.
Gunness, P. and Gidley, M. J. (2010) ‘Mechanisms underlying the cholesterol-lowering properties of soluble dietary fibre polysaccharides’, Food & Function, 1(2), p. 149. doi: 10.1039/c0fo00080a.
Han, S.-H., Lee, S.-W. and Rhee, C. (2009) ‘Effect of heat treatment of digestion-resistant fraction from soybean on retarding of bile acid transport in vitro’, Nutrition Research and Practice, 3(2), p. 149. doi: 10.4162/nrp.2009.3.2.149.
Izydorczyk, M. S. and Dexter, J. E. (2008) ‘Barley β-glucans and arabinoxylans: Molecular structure, physicochemical properties, and uses in food products–a Review’, Food Research International. Elsevier Ltd, 41(9), pp. 850–868. doi: 10.1016/j.foodres.2008.04.001.
Izydorczyk, M. S., Macri, L. J. and Macgregor, A. W. (1998) ‘Structure and physicochemical properties of barley extractable P-glucans and arabinoxylans ’’, 8617(97), pp. 259–269.
Kim, H. J. and White, P. J. (2010) ‘In Vitro Bile-Acid Binding and Fermentation of High, Medium, and Low Molecular Weight β-Glucan’, Journal of Agricultural and Food Chemistry, 58(1), pp. 628–634. doi: 10.1021/jf902508t.
Kivelä, R., Henniges, U., Sontag-Strohm, T. and Potthast, A. (2012) ‘Oxidation of oat β-glucan in aqueous solutions during processing’, Carbohydrate Polymers, 87(1), pp. 589–597. doi: 10.1016/j.carbpol.2011.08.028.
Kristiansen, K. A., Potthast, A. and Christensen, B. E. (2010) ‘Periodate oxidation of polysaccharides

97
for modification of chemical and physical properties’, Carbohydrate Research. Elsevier Ltd, 345(10), pp. 1264–1271. doi: 10.1016/j.carres.2010.02.011.
Lazaridou, A., Biliaderis, C. ., Micha-Screttas, M. and Steele, B. . (2004) ‘A comparative study on structure–function relations of mixed-linkage (1→3), (1→4) linear β-d-glucans’, Food Hydrocolloids, 18(5), pp. 837–855. doi: 10.1016/j.foodhyd.2004.01.002.
Lazaridou, A. and Biliaderis, C. G. (2007) ‘Molecular aspects of cereal β-glucan functionality: Physical properties, technological applications and physiological effects’, Journal of Cereal Science, 46(2), pp. 101–118. doi: 10.1016/j.jcs.2007.05.003.
Lazaridou, A., Biliaderis, C. G. and Izydorczyk, M. S. (2003) ‘Molecular size effects on rheological properties of oat β-glucans in solution and gels’, Food Hydrocolloids, 17(5), pp. 693–712. doi: 10.1016/S0268-005X(03)00036-5.
Lee, K. Y., Park, S. Y. and Lee, H. G. (2011) ‘Effect of oat β-glucan and its oxidised derivative on the quality characteristics of sponge cake’, International Journal of Food Science & Technology, 46(12), pp. 2663–2668. doi: 10.1111/j.1365-2621.2011.02798.x.
Limberger-Bayer, V. M., de Francisco, A., Chan, A., Oro, T., Ogliari, P. J. and Barreto, P. L. M. (2014) ‘Barley β-glucans extraction and partial characterization’, Food Chemistry, 154, pp. 84–89. doi: 10.1016/j.foodchem.2013.12.104.
Mossé, J. (1990) ‘Nitrogen to Protein Conversion Factor for Ten Cereals and Six Legumes or Oilseeds. A Reappraisal of Its Definition and Determination. Variation According to Species and to Seed Protein Content Jacques’, Journal of Agricultural and Food Chemistry, (38), pp. 18–24.
de Moura, F. A., Pereira, J. M., da Silva, D. O., Zavareze, E. da R., da Silveira Moreira, A., Helbig, E. and Dias, A. R. G. (2011) ‘Effects of oxidative treatment on the physicochemical, rheological and functional properties of oat β-glucan’, Food Chemistry, 128(4), pp. 982–987. doi: 10.1016/j.foodchem.2011.04.003.
Naumann, S., Schweiggert-Weisz, U., Bader-Mittermaier, S., Haller, D. and Eisner, P. (2018) ‘Differentiation of adsorptive and viscous effects of dietary fibres on bile acid release by means of in vitro digestion and dialysis’, International Journal of Molecular Sciences, 19(8). doi: 10.3390/ijms19082193.
Park, S. Y., Bae, I. Y., Lee, S. and Lee, H. G. (2009) ‘Physicochemical and Hypocholesterolemic Characterization of Oxidized Oat β-Glucan’, Journal of Agricultural and Food Chemistry, 57(2), pp. 439–443. doi: 10.1021/jf802811b.
Rieder, A., Knutsen, S. H., Ulset, A. T., Christensen, B. E., Andersson, R., Mikkelson, A., Tuomainen, P., Maina, N. and Ballance, S. (2015) ‘Inter-laboratory evaluation of SEC-post-column calcofluor for determination of the weight-average molar mass of cereal β-glucan’, Carbohydrate Polymers. Elsevier Ltd., 124, pp. 254–264. doi: 10.1016/j.carbpol.2015.02.019.
Sayar, S., Jannink, J.-L. and White, P. J. (2011) ‘Textural and Bile Acid-Binding Properties of Muffins Impacted by Oat β-Glucan with Different Molecular Weights’, Cereal Chemistry Journal, 88(6), pp. 564–569. doi: 10.1094/CCHEM-02-11-0014.
Spatareanu, A., Bercea, M., Budtova, T., Harabagiu, V., Sacarescu, L. and Coseri, S. (2014) ‘Synthesis, characterization and solution behaviour of oxidized pullulan , Members of European Polysaccharides Network of Excellence (EPNOE)’, Carbohydrate Polymers. Elsevier Ltd., 111, pp. 63–71. doi: 10.1016/j.carbpol.2014.04.060.
Tamura, N., Hirota, M., Saito, T. and Isogai, A. (2010) ‘Oxidation of curdlan and other polysaccharides by 4-acetamide-TEMPO/NaClO/NaClO2 under acid conditions’, Carbohydrate Polymers. Elsevier Ltd., 81(3), pp. 592–598. doi: 10.1016/j.carbpol.2010.03.016.
Wolever, T. M., Tosh, S. M., Gibbs, A. L., Brand-Miller, J., Duncan, A. M., Hart, V., Lamarche, B., Thomson, B. A., Duss, R. and Wood, P. J. (2010) ‘Physicochemical properties of oat β-glucan

98
influence its ability to reduce serum LDL cholesterol in humans: a randomized clinical trial’, The American Journal of Clinical Nutrition, 92(4), pp. 723–732. doi: 10.3945/ajcn.2010.29174.
Wood, P. J. (1991) ‘Oat β-glucan-physicochemical properties and physiological effects’, Trends in Food Science & Technology, 2(December), pp. 311–314. doi: 10.1016/0924-2244(91)90733-Y.
Wood, P. J. (2004) ‘Relationships between solution properties of cereal β-glucans and physiological effects — a review’, Trends in Food Science & Technology, 15(6), pp. 313–320. doi: 10.1016/j.tifs.2003.03.001.
Wood, P. J. (2007) ‘Cereal β-glucans in diet and health’, Journal of Cereal Science, 46(3), pp. 230–238. doi: 10.1016/j.jcs.2007.06.012.
Zacherl, C., Eisner, P. and Engel, K.-H. (2011) ‘In vitro model to correlate viscosity and bile acid-binding capacity of digested water-soluble and insoluble dietary fibres’, Food Chemistry. Elsevier Ltd, 126(2), pp. 423–428. doi: 10.1016/j.foodchem.2010.10.113.
ADDIN Mendeley Bibliography CSL_BIBLIOGRAPHYX

99
II. In vitro estimation of iron binding by modified cereal
β-glucan extracts
Elena Marasca, Olivia Wyss, Samy Boulos and Laura Nyström
ABSTRACT
The reduction of iron bioavailability in the presence of dietary fibers is a well known fact, but
the mechanism behind this phenomenon has not been fully clarified yet. Recent literature has
shown that the main responsible factor for iron reduction in the case of native cereal β-glucan
is phytic acid, which is often found in association with dietary fibers and forms a strong complex
with iron rendering it unavailable for absorption. During food processing, however, the
molecular structure of β-glucan can undergo modifications which can affect the delicate
equilibrium between phytic acid, fiber and minerals such as iron. In the present study, an in
vitro method was used to elucidate the relationship between β-glucan structure and its iron-
binding capability and to clarify the role of phytic acid in this mechanism. TEMPO oxidation,
sodium periodate oxidation and acid hydrolysis were used as modifications of the β-glucan
fibers, which were subsequently characterized and digested in vitro in the presence of iron, with
or without prior enzymatic dephytinization. The results obtained in this study confirm that no
direct binding between native β-glucan and iron exists, and underlines the major impact of
phytic acid as an inhibitor of iron absorption in cereal foods.
1. Introduction
For living organisms, iron is an essential trace element, crucial for biological functions such as
respiration, energy production, DNA synthesis and cell proliferation (Abbaspour et al., 2015).
Despite its geologic abundance and large-scale intervention programs, more than two billion
people in developing as well as high-income countries are affected by iron deficiency, which
remains the most widely prevalent nutritional problem worldwide. Besides insufficient dietary
intake and pathologic conditions, one of the most likely causes of this nutritional problem is the
poor bioavailability of dietary iron, which is influenced by its chemical form, the type of dietary
iron (heme and non-heme), the iron status of the individual and the presence of other food
components (Hemalatha et al., 2007; Hurrell and Egli, 2010; Camaschella, 2015).
Generally, cereals can be a good source of minerals such as iron and zinc, but dietary fibers can
have a negative effect on their bioavailability (Kennefick and Cashman, 2000). Also, there is

100
considerable evidence that phytate, which is associated with fibers in many foods, reduces the
bioavailability of iron (Hallberg et al., 1987; Hurrell et al., 1992; Hurrell et al., 2003; Nielsen
et al., 2013). In particular, recent studies have confirmed phytic acid as the main responsible
for iron binding by commercial oat and barley β-glucan (Wang et al., 2017 a&b). However, to
the best of our knowledge, no literature investigating the impact of β-glucan structural
modifications on iron bioavailability exists. During food processing, in fact, β-glucan can
undergo degradation, such as chain scission and oxidation (Kivelä et al., 2011, 2012; Mäkelä
et al., 2015), which can alter its behaviour in the gastro-intestinal tract, and which can, in turn,
affect the equilibrium between phytic acid and iron.
Accurate measurement of iron bioavailability can be challenging, but in vivo human studies
using iron isotopes have been proven to be very successful (Hurrell, 1997). However, since in
vivo human studies are expensive and time consuming, and animal models have certain
limitations, such as the differences in mineral requirements, metabolism, digestive capacity,
and sensitivity to dietary factors compared to humans (Reddy and Cook, 1991), in vitro methods
for the determination of iron availability were developed as an alternative in the 1970s and
1980s and are still in use today (Miller et al., 1981; Larsson et al., 1997; Sandberg, 2005). These
methods are not as accurate as in vivo studies to determine real bioavailability, since the
interactions between the digestive system and the food ingested cannot be measured, and factors
such as transit time, site of absorption, composition of digestive secretions responding to the
meal and intestinal flora may all affect the mineral availability (Sandberg, 2005). Despite these
limitations, however, the in vitro studies are still very useful to predict trends or compare iron
absorption between different samples (Hurrell, 1997).
While there are several studies concerning iron availability in relation to wheat bran fibers and
phytic acid (Simpson et al., 1981; Hallberg et al., 1987; Kennefick et al., 2000), not many
studies investigate the impact of other fibers, in particular β-glucan, on the bioavailability of
iron. (1→3)(1→4)-β-D-Glucan (cereal β-glucan) is a partly soluble dietary fiber found
predominantly in the cell walls of the endosperm and aleurone layer of oat and barley grains
and in smaller amounts in rye and wheat. β-Glucan has several positive health effects, in
particular blood cholesterol lowering properties, recognized by EFSA as well as the FDA (FDA,
2009; EFSA, 2011) , and of beneficial blood glucose regulation, as evaluated by EFSA (EFSA,
2010).
In a study by Platt and Clydesdale (1984), cellulose, lignin, sodium phytate and cereal β-glucan,
alone and in combination, were evaluated with regard to their effect on iron profiles in model
systems containing FeSO4·7H2O under simulated gastrointestinal pH conditions. They

101
observed that lowering the pH to gastric conditions (pH 2) triggered the solubilisation of a
portion of the iron bound to lignin, cellulose and β-glucan. Bringing back the pH to neutral
conditions (pH 6.8) led again to increased portions of complexed iron and reduced portions of
soluble iron. They hypothesised binding strengths in the order lignin/ phytic acid > β-glucan >
cellulose, but concluded that the binding components in cereals interact, which critically affects
iron solubility and thus bioavailability.
Differences in iron binding properties of commercial high-viscosity oat and barley β-glucan of
high purity (>97%) were assessed also in our earlier study (Faure et al., 2015). The kinetics of
the Fenton reaction between iron(II) and hydrogen peroxide in 0.6% oat and barley β-glucan
solutions at pH values 2.7 and 4.7 were measured. At low pH, β-glucan did not affect the rate
of the Fenton reaction, which implies that all iron was available to react with hydrogen
peroxide. At pH 4.7, on the other hand, barley β-glucan and even more so oat β-glucan reduced
the reaction rate suggesting that both bind iron, with the commercial oat β-glucan material
showing a stronger binding power than barley β-glucan. The variation in iron binding was
hypothesized to be caused by the different ratio of β-(1→3)-linked cellotriosyl to cellotetraosyl
units (DP3:DP4) between β-glucan from different cereal sources, with oat β-glucan having
more β-(1→4)-linkages than barley. On the other hand, Fenton-induced oxidation of three
constitutionally isomeric β-glucan tetrasaccharides as model compounds with no or one β-
(1→3)-linkage exhibited no significant differences in degradation kinetics, suggesting that for
the commercial β-glucan polymer materials, other constituents than the pure carbohydrates are
responsible for the difference in behaviour between oat and barley β-glucan (Boulos and
Nyström, 2016). Wang et al. (2017 a&b) attributed the differences in iron-binding behaviour
of commercial β-glucan materials to the varying levels of phytic acid contamination, as removal
of phytic acid by treatment with ion-exchange resins led to same degradation rates and iron
binding capacities of all β-glucan materials irrespective of their origin.
Thanks to its health promoting properties and technological functions, β-glucan is added as
functional ingredient in different food products such as pasta, soups, bread and low-fat dairy
products (Brennan et al., 2005; Lazaridou and Biliaderis, 2007). However, it has been shown
that during production and in food systems β-glucan may be subjected to degradation processes
such as oxidation and acid hydrolysis, which alter its viscous properties (reduction of Mw), but
also its interactions with minor food components (Brennan et al., 2005; Johansson et al., 2006;
Kivelä et al., 2012). Recent literature shows that processing or modification does not
necessarily reduce β-glucan’s health benefits or its technological properties, but in certain cases
can actually improve both (Park et al., 2009; De Moura et al., 2011; Lee et al., 2011). However,

102
to be best of our knowledge, no literature investigating the impact of β-glucan structural
modifications on iron bioavailability exists. In order to get more insight on how processing
affects iron binding, β-glucan extracts from oat bran and barley flour were modified with
different targeted chemical treatments and the behaviour of the different samples was compared
in an in vitro digestion experiment. Also, the role of phytic acid was studied by means of prior
phytase treatment of the native extracts.
2. Materials
Oat flour (OatWell® 28) and barley flour (Beta®Barley dehulled and untreated) were supplied
by DSM (Herleen, Netherlands) and Dieckmann Cereals GmbH (Rinteln, Germany),
respectively. Calcium chloride, Chelex® 100 sodium form (50-100 mesh), glycerol, bile extract
porcine, hydrochloric acid, iron(II) sulfate heptahydrate, maltotriose, potassium hydroxide,
sodium acetate, sodium azide, sodium bicarbonate, sodium chlorite (80%), sodium hypochlorite
(available chlorine 10-15%), sodium nitrate, sodium (meta) periodate, 2,2,6,6-Tetramethyl-
piperidine 1-oxyl (TEMPO, 98%), α-amylase from Bacillus licheniformis (Termamyl 300®L),
pancreatin from porcine pancreas, and pepsin from porcine gastric mucosa (≥250 units/mg
solid) were purchased from Sigma-Aldrich, Switzerland. Ethanol absolute was supplied by
Alcosuisse, Switzerland. Acetic acid glacial and sodium hydroxide pellets were purchased from
Fisher Chemical, United Kingdom. EDTA, sodium hydroxide solution (50–52% in water),
sulphuric acid and sodium phosphate were purchased from Fluka, Germany. Titrisol® Iron
standard (1000 mg in 1000 mL 3.5% HCl) was purchased from Merck, Germany. Ronozyme™
P-(M) (Peniophora lycii wt phytase, 10 U/mg), was provided by Novozymes, Denmark.
Ultrapure (Milli-Q) water was used for all experiments (Merck Millipore, Merck KGaA,
Darmstadt, Germany). All glassware used for the experiments was washed in a laboratory
dishwasher, soaked overnight in 1M HCl and then rinsed with Milli-Q water.
3. Methods
3.1 β-Glucan extraction
β-Glucan was extracted and purified from oat and barley flour according to the procedure by
Bhatty (1995), with slight adaptations. Firstly, the flours were stirred in 1 M NaOH (2% w/w)

103
for 24 hours, followed by removal of debris by centrifugation for 10 min at 4000 rpm and 4°C
(Centrifuge 5810 R, Eppendorf, Germany). The supernatant was collected and the pH adjusted
to 6.5 with conc. HCl followed by the addition of 2 mL Termamyl (≥300 U/g) and 70 mg CaCl2
per liter of extraction mixture. The mixture was then incubated for 1 h at 96°C to hydrolyze
starch. After cooling to RT, the pH was adjusted to 4.5 with conc. HCl to denaturate proteins,
which were removed as precipitate by centrifugation for 15 min at 4000 rpm and 4°C. The
supernatant was collected and the β-glucan precipitated with an equal volume of ethanol. The
β-glucan precipitate was collected by centrifugation (10 min, 4000 rpm, 4°C) and decantation
of the supernatant, freeze-dried for 48 h (Lyolab BII, LSL Secfroid, Switzerland), and
homogenized with a ball mill (Pulverisette 23, Fritsch, Germany).
3.2 Iron removal
To remove intrinsic iron from the extracts, a treatment with Chelex (a chelating ion exchange
resin) was carried out. Chelex consists of styrene divinylbenzene copolymers containing paired
iminodiacetate ions, with a high preference for polyvalent transition metals such as iron. The
quantity of ions exchanged is very low at pH below 2, increases sharply from pH 2 to 4, and
reaches a maximum above pH 4 (Bio-Rad). Multiple rounds of optimisation led to the
following protocol: a 0.75% oat or barley β-glucan solution was prepared by dissolving the
extract in a shaking water bath overnight at 70°C, and subsequently treated three times with
Chelex. For each treatment, 50 g Chelex beads were added per liter of β-glucan solution and
the suspension stirred for 2 hours, then vacuum-filtered through a glass filter (250 mL, pore
size 2). After each treatment the Chelex was regenerated with aqueous HCl and NaOH washings
as described in the manufacturer’s instructions. In the first and second treatment, the pH of the
solution was lowered to 2.5 with 1 M and 0.1 M sulfuric acid to favour the solubilisation of
iron. For the third treatment, the pH was not adjusted after stirring the solution with Chelex, but
left at around pH 10, in order to enhance the chelating power of the resin. After this cycle of
treatments, two volumes of ethanol were added to the solutions, which were then stored for 48
hours at 4°C to help the precipitation of iron-free β-glucan. The samples were then centrifuged
(10 min, 4000 rpm, 4 °C), the pellet freeze-dried for 48 h and milled by ball mill.

104
3.3 Composition of the extracts
3.3.1 Total β-glucan content
The total β-glucan content of the oat and barley β-glucan extracts before and after Chelex
treatment was determined with the β-glucan Assay Kit (Mixed Linkage) from Megazyme,
Ireland.
3.3.2 Protein content
To estimate the protein content of the oat and barley β-glucan extracts before and after Chelex
treatment, samples were dried for 24 hours at 120 °C and sent to the service for microelemental
analyses at ETH (Elementaranalysen LOC, ETH Zurich). The nitrogen content was determined
in duplicates by a TruSpec Micro, composed of a combination of flow-through carrier gas and
individual, highly selective infrared (IR) and thermal conductivity detectors (TruSpec Micro,
LECO Corporation, USA). The approximate protein content was then calculated using the
nitrogen conversion factors determined by Mosse (1990) of 5.36 for the oat extracts and 5.50
for the barley extracts.
3.3.3 Phytic acid content
The phytic acid content of the oat and barley β-glucan extracts before and after Chelex treatment
was determined in triplicates with the Phytic Acid (Phytate)/Total Phosphorus Kit obtained
from Megazyme, Ireland.
3.3.4 Iron content
Iron concentration was determined by means of a graphite furnace AAS (Agilent Technologies
AG, Switzerland, VARIAN GTA 120). The wavelength for iron (248.3 nm) was selected, with
a slit width of 0.2 nm and a lamp current of 5.0 mA with background correction on. The heating
program was divided in several steps to evaporate the water (85–140°C), burn the organic
residues (800°C), and atomize the contained iron for the absorption to be measured (2300°C).
The calibration curve was automatically constructed by the instrument from a standard solution
(10 ng/mL) prepared by dilution of the standard stock solution of Fe Titrisol® (1000 mg/L).
After dissolving the β-glucan extracts, samples were diluted with aqueous HCl to reach a final

105
concentration of 0.1 M HCl and an intrinsic iron concentration below 10 µg/L to lie within the
calibration range of the atomic absorption spectrometry (AAS). Each sample or standard was
measured in triplicate. The samples were stored at 4 °C until measurement.
3.4 Structural characterisation of the extracts
3.4.1 DP3 to DP4 ratio
The DP3 to DP4 ratio of the oat and barley β-glucan was determined before and after the Chelex
treatment, as well as after the modifications. The polymer was hydrolysed by lichenase and the
concentration of the resulting main β-gluco-oligomers DP3 and DP4 measured in triplicates
with high-performance anion exchange chromatography and pulsed amperometric detection
(HPAEC-PAD).
First of all, 70 mg of the different oat and barley β-glucan extracts (untreated, Chelex- treated,
hydrolysed, TEMPO- and NaIO4-oxidised) was dispersed in 0.2 mL 50% aqueous EtOH and 4
mL 20 mM NaH2PO4/ NaHPO4 buffer (pH 6.5), and boiled for 10 min. After cooling, 0.2 mL
lichenase solution (20 U/mL) was added and the samples were incubated for 2 hours at 60°C.
Lichenase was inactivated by boiling the tubes for 15 min, then 500 μL of each sample was
transferred to a fresh tube and diluted with 4.5 mL water. An aliquot of 0.3 mL was mixed with
0.3 mL water and 0.4 mL 60 µM maltotriose (internal standard I.S.). Calibration curves were
constructed in the range of 10–100 µM for DP3 and 5–50 μM for DP4, relative to the area of
the I.S.. The samples, standard solutions for calibration, and water as blank were filtered (0.45
μm) into HPLC vials and analysed by HPAEC-PAD.
The instrument consisted of a gradient pump (GS50), auto sampler (AS50), thermal
compartment (AS50), pneumatic controller (PC10) and the electrochemical detector (ED50),
all from Dionex Bio LC, USA. The column was an Analytical Dionex CarboPac PA1 (2 mm ×
250 mm) with a CarboPac PA1 guard column (2 mm × 50 mm). The signal was measured with
pulsed amperometric detection using waveform A from Dionex technical note 21. Eluents were
(A) 150 mM NaOH solution and (B) 500 mM NaOAc with 150 mM NaOH. The following
eluent gradient was applied: 90% eluent A and 10% eluent B from 0 to 2 min, a linear increase
of eluent B from 10 to 100% between 2 and 17 min, 100% eluent B between 17 and 23 min,
and 90% eluent A and 10% eluent B from 23 min to 37 min. The flow rate was 0.25 mL/min,
except between 23 and 36 min it was increased to 0.75 mL/min. The resulting peaks were
analysed using the software Chromeleon® version 6.8, also by Dionex Bio LC, USA.

106
3.4.2 Molecular weight
The molecular weight (Mw) of the oat and barley β-glucan extracts before and after the Chelex
treatment, as well as after the modifications, was determined in triplicates by means of high
performance size exclusion chromatography (HPSEC). The extracts were dissolved in eluent
(0.1 M sodium nitrate and 0.02% (w/v) sodium azide) to a final concentration of 0.1% (w/v),
and filtered (0.45 μm) into HPLC vials. For the calibration curve, 3 mg of β-glucan standards
with reported peak molecular weight (Mp) of 33’600, 67’100, 187’100, 247’000, 375’000, and
667’000 g/mol (Megazyme, Ireland) were boiled in 3 mL eluent until dissolved and filtered
(0.45 μm) into HPLC vials.
The HPLC consisted of a binary pump, degasser, thermostated column compartment,
and auto sampler, all from Agilent Technologies (Series 1100, Hewlett Packard, USA). A pre-
column (Viscotek AGuard Col. 50 x 6.0 mm, Malvern Instruments Ltd, United Kingdom) was
used together with an A5000 column (Viscotek, 300 x 7.8 mm, Malvern Instruments Ltd, UK)
and a suprema 30000 column (10 μm, 8 x 300 mm, PSS Polymer Standards Service GmbH,
Germany). The temperature of the columns was kept at 35 °C with a flow rate of 1 mL/min and
an injection volume of 50 µL. The elution was recorded using a refractive index detector (Series
1200, Agilent Technologies (Schweiz) AG, Switzerland). The weight average molecular
weights were calculated based on the measured standard curve with the ChemStation software
(ChemStation for LC 3D systems, Rev B.04.02 SP1) and the add-on Cirrus GPC/SEC software
(version 3.4.1) from Agilent.
3.5 Structural modifications
3.5.1 Acid hydrolysis
The pH of 1% (w/v) Chelex-treated oat or barley β-glucan solutions was adjusted to 1 with 1 M
HCl. The samples were left to hydrolyse for 24 hours at 50°C and then neutralised with 1 M
NaOH. The extracted β-glucan was precipitated by adding two volumes of ethanol and then
stored at 4°C for 48 hours to help the precipitation. Finally, the samples were centrifuged
(10 min, 4000 rpm, 4 °C), the collected solids freeze dried and milled by ball mill.
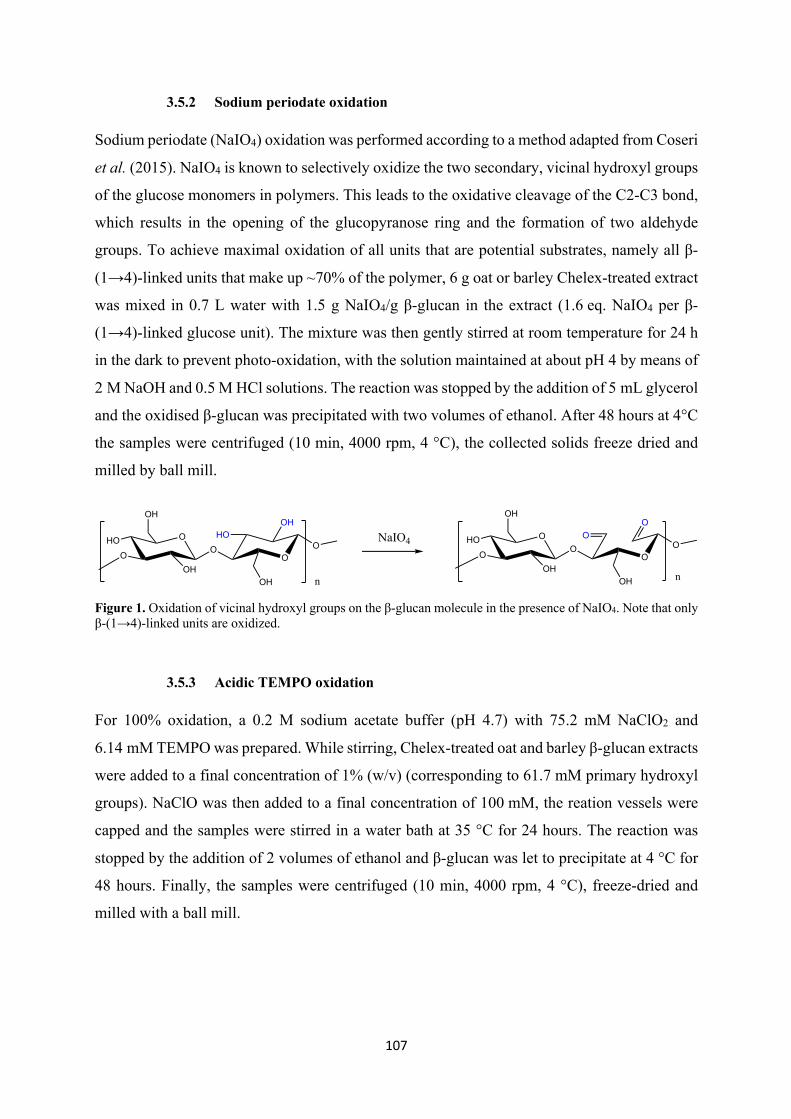
107
3.5.2 Sodium periodate oxidation
Sodium periodate (NaIO4) oxidation was performed according to a method adapted from Coseri
et al. (2015). NaIO4 is known to selectively oxidize the two secondary, vicinal hydroxyl groups
of the glucose monomers in polymers. This leads to the oxidative cleavage of the C2-C3 bond,
which results in the opening of the glucopyranose ring and the formation of two aldehyde
groups. To achieve maximal oxidation of all units that are potential substrates, namely all β-
(1→4)-linked units that make up ~70% of the polymer, 6 g oat or barley Chelex-treated extract
was mixed in 0.7 L water with 1.5 g NaIO4/g β-glucan in the extract (1.6 eq. NaIO4 per β-
(1→4)-linked glucose unit). The mixture was then gently stirred at room temperature for 24 h
in the dark to prevent photo-oxidation, with the solution maintained at about pH 4 by means of
2 M NaOH and 0.5 M HCl solutions. The reaction was stopped by the addition of 5 mL glycerol
and the oxidised β-glucan was precipitated with two volumes of ethanol. After 48 hours at 4°C
the samples were centrifuged (10 min, 4000 rpm, 4 °C), the collected solids freeze dried and
milled by ball mill.
Figure 1. Oxidation of vicinal hydroxyl groups on the β-glucan molecule in the presence of NaIO4. Note that only β-(1→4)-linked units are oxidized.
3.5.3 Acidic TEMPO oxidation
For 100% oxidation, a 0.2 M sodium acetate buffer (pH 4.7) with 75.2 mM NaClO2 and
6.14 mM TEMPO was prepared. While stirring, Chelex-treated oat and barley β-glucan extracts
were added to a final concentration of 1% (w/v) (corresponding to 61.7 mM primary hydroxyl
groups). NaClO was then added to a final concentration of 100 mM, the reation vessels were
capped and the samples were stirred in a water bath at 35 °C for 24 hours. The reaction was
stopped by the addition of 2 volumes of ethanol and β-glucan was let to precipitate at 4 °C for
48 hours. Finally, the samples were centrifuged (10 min, 4000 rpm, 4 °C), freeze-dried and
milled with a ball mill.
O
OH
OH
OO
HOOH
OH
ONaIO4 O
OH
OH
OO
OO
OH
OOO
n
HO
n
HO

108
Figure 2. Oxidation of β-glucan in the presence of nitroxyl radicals (TEMPO).
3.6 In vitro digestion and determination of dialysable iron
The procedure used for the in vitro digestion was originally developed by Miller et al. (1981),
adapted by Hurrell et al. (1988). Additional slight adaptations were made in the present study
to fit the experimental design. All β-glucan extracts, native and modified, were digested in vitro
in triplicates. The digestion of the untreated and Chelex-treated samples was performed with
and without a prior degradation of phytic acid by means of phytase. The following enzyme
solutions were prepared freshly for each digestion: phytase solution (3 mg/mL = 30 U/mL), 3.2
g pepsin + 16.8 g 0.1 M HCl (resulting in 16% (w/w) pepsin), and both 0.4% (w/v) pancreatin
and 2.5% (w/v) bile extract together in 0.1 M NaHCO3
First of all, a β-glucan solution was prepared by mixing each extract with water (1% (w/w)
effective β-glucan concentration), stirring the mixture for 30 min at RT and then shaking it
vigorously in a dry incubator at 65°C for two hours. For untreated oat, untreated barley, Chelex-
treated oat and Chelex-treated barley, this corresponded to an extract concentration of 1.7%,
2.2%, 1.5% and 2.2%, respectively.
After pH adjustment to 4–4.5 with 6 M HCl, either 1 mL water or 1 mL phytase solution (30
U) was added to the sample, which was then incubated overnight in a shaking water bath at
40°C and 160 rpm. After cooling to RT, the pH was adjusted to 2.0 using 6 M HCl, in order to
mimic the gastric conditions. The sample was then divided into quadruplicate 50 g aliquots,
which were transferred to 100 mL Schott bottles. To each of these, 25 μL 0.1 M FeSO4 in 0.1
M HCl (50 μM final concentration in samples) and 1.3 mL 16% pepsin were added, followed
by incubation for 2 h in a shaking water bath at 37°C and 160 rpm. In the simulated transition
from the stomach to the intestine, a slow increase of pH is required. This is crucial in any
attempt to simulate gastrointestinal digestion of food iron due to formation of ferrous and ferric
hydroxide aggregates at elevated pH that eventually precipitate, lowering iron availability
(Miller et al., 1981). The amount of base required to increase the pH of the different samples
was determined by titration. Titratable acidity was determined on one of the 50 g replicates by
NaClO2, NaOCl
N
OO
OH
OH
O OO
OH
OH
OO
COOH
OH
O OO
OH
COOH
OHO HO
HO HO
n n

109
adding 5 mL of the pancreatin solution followed by titration to pH 7.5 with 0.5 M KOH (roughly
1-3 mL of KOH solution required, depending on the sample). Titratable acidity was defined as
the number of equivalents of KOH required to titrate the solution to pH 7.5. A segment of
dialysis tubing 30 cm long (Spectra/Por Dialysis Membrane, MWCO 6-8 kD, 32 mm flat width,
Spectrum Labs, United States) containing 25 mL water and an amount of NaHCO3 equivalent
to the titratable acidity measured previously was placed in each of the remaining three
replicates, which were then incubated at 37°C in a shaking water bath (160 rpm) until pH
reached about 5, after approximately 40–70 min, depending on the sample. After that, 5 mL of
pancreatin solution was added and the samples were incubated for an additional 2 hours at 37
°C in the shaking water bath (160 rpm). Finally, a 20 mL aliquot of the dialysate (sample inside
the tube) was diluted two times and acidified with HCl (0.1 M final concentration) to keep the
iron in solution. The samples were then further diluted according to their expected iron content
to be below 10 µg/L and hence in the calibration range for analysis by atomic absorption
spectroscopy and the iron content measured as described in section 3.3.4.
In addition, a reference and a control were also analysed: the reference was prepared and treated
like the samples, but the β-glucan solution was replaced by water. In the control, the β-glucan
solution was also replaced by water but no FeSO4 was added, in order to check possible
contaminations of iron from the reagents and enzymes used for digestion.
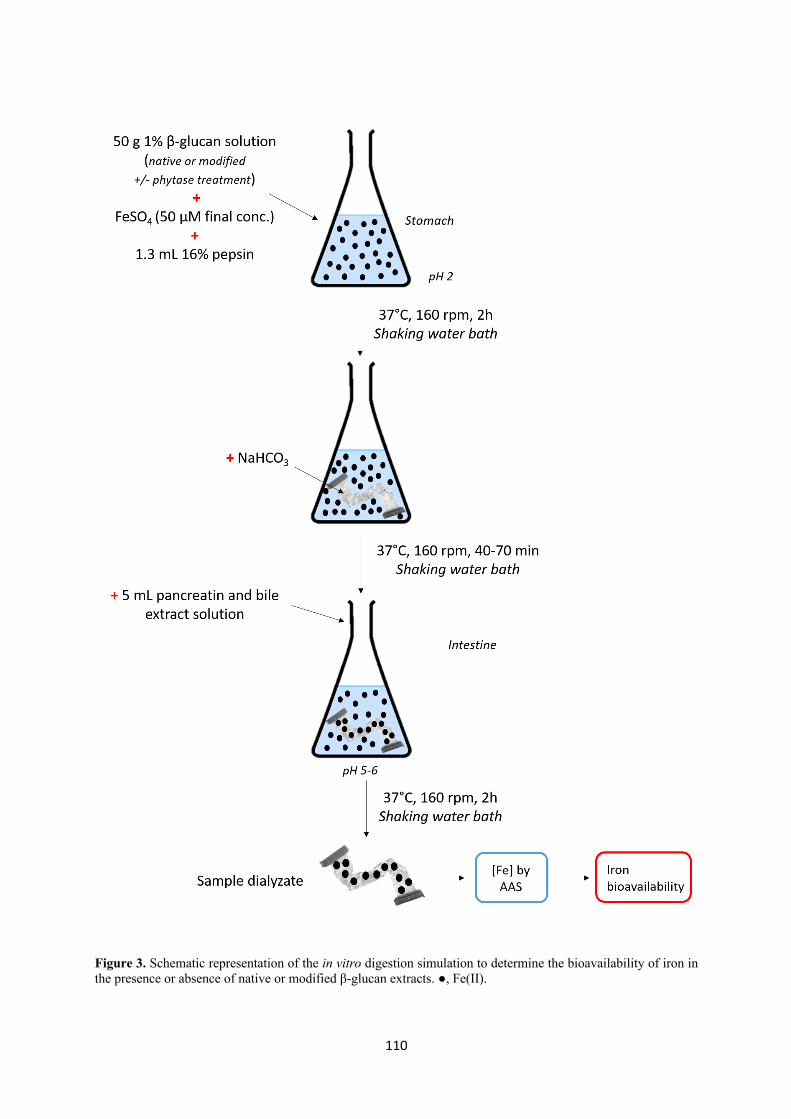
110
Figure 3. Schematic representation of the in vitro digestion simulation to determine the bioavailability of iron in the presence or absence of native or modified β-glucan extracts. ●, Fe(II).
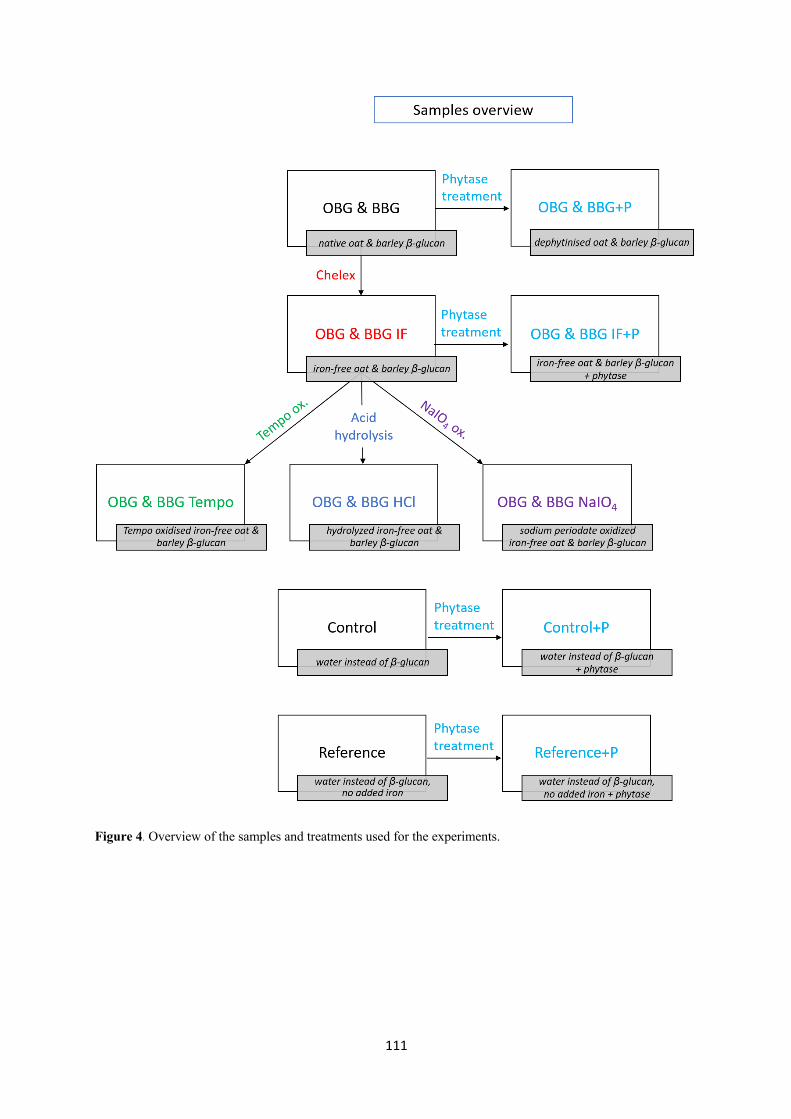
111
Figure 4. Overview of the samples and treatments used for the experiments.
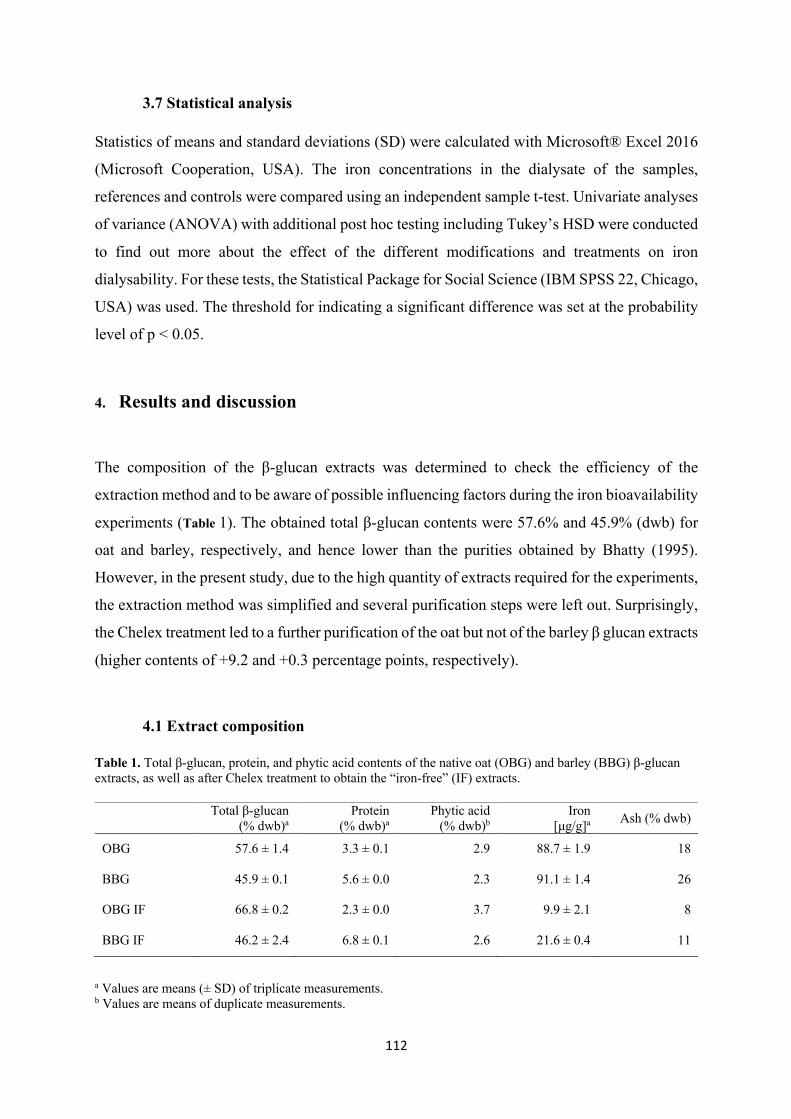
112
3.7 Statistical analysis
Statistics of means and standard deviations (SD) were calculated with Microsoft® Excel 2016
(Microsoft Cooperation, USA). The iron concentrations in the dialysate of the samples,
references and controls were compared using an independent sample t-test. Univariate analyses
of variance (ANOVA) with additional post hoc testing including Tukey’s HSD were conducted
to find out more about the effect of the different modifications and treatments on iron
dialysability. For these tests, the Statistical Package for Social Science (IBM SPSS 22, Chicago,
USA) was used. The threshold for indicating a significant difference was set at the probability
level of p < 0.05.
4. Results and discussion
The composition of the β-glucan extracts was determined to check the efficiency of the
extraction method and to be aware of possible influencing factors during the iron bioavailability
experiments (Table 1). The obtained total β-glucan contents were 57.6% and 45.9% (dwb) for
oat and barley, respectively, and hence lower than the purities obtained by Bhatty (1995).
However, in the present study, due to the high quantity of extracts required for the experiments,
the extraction method was simplified and several purification steps were left out. Surprisingly,
the Chelex treatment led to a further purification of the oat but not of the barley β glucan extracts
(higher contents of +9.2 and +0.3 percentage points, respectively).
4.1 Extract composition
Table 1. Total β-glucan, protein, and phytic acid contents of the native oat (OBG) and barley (BBG) β-glucan extracts, as well as after Chelex treatment to obtain the “iron-free” (IF) extracts.
a Values are means (± SD) of triplicate measurements. b Values are means of duplicate measurements.
Total β-glucan (% dwb)a
Protein (% dwb)a
Phytic acid (% dwb)b
Iron [μg/g]a Ash (% dwb)
OBG 57.6 ± 1.4 3.3 ± 0.1 2.9 88.7 ± 1.9 18
BBG 45.9 ± 0.1 5.6 ± 0.0 2.3 91.1 ± 1.4 26
OBG IF 66.8 ± 0.2 2.3 ± 0.0 3.7 9.9 ± 2.1 8
BBG IF 46.2 ± 2.4 6.8 ± 0.1 2.6 21.6 ± 0.4 11

113
The protein content of the barley β-glucan extract (5.6%) was higher than that of the oat extract
(3.3%), which is in line with the results by Bhatty (1995). Interestingly, the Chelex treatment
led to a reduced protein content in the oat β-glucan extract (2.3%), but to an increased
proportion of protein in the dry matter in the case of barley β-glucan (6.8%).
The phytic acid contents were not significantly different between the two extracts, with a trend
towards higher concentrations in the oat than in the barley β-glucan extract (2.9± 0.3% and 2.3
± 0.1%, respectively). This trend is in line with the used sources of whole grain flour for barley
and bran (where phytic acid is primarily located) for oat. (Haros et al., 2001; Schlemmer et al.,
2009) Comparing the results with literature values, phytic acid in the extracts is three to four
times higher than in oat or barley flour (Schlemmer et al., 2009). The Chelex treatment led to
a higher amount of phytic acid in the extracts, in particular for oat (3.7%), which might indicate
a correlation between β-glucan and phytic acid content. This is in sharp contrast to a recent
study by Wang et al. (2017) where an ion-exchange treatment of commercial oat β-glucan led
to the concomitant removal of both iron and phytic acid.
The iron content of the untreated β-glucan extracts was similar for oat (88.7 ± 1.9 μg/g) and
barley (91.1 ± 1.4 μg/g). The removal of the intrinsic iron using Chelex was similar for oat and
barley, with a reduction of 90 and 76%, respectively.
4.2 Structural characteristics
To characterise the β-glucan and investigate the effect of the modifications on the fibres, Mw
and DP3/DP4 were determined before and after the different treatments by HPSEC and
HPAEC-PAD, respectively (Table 2).
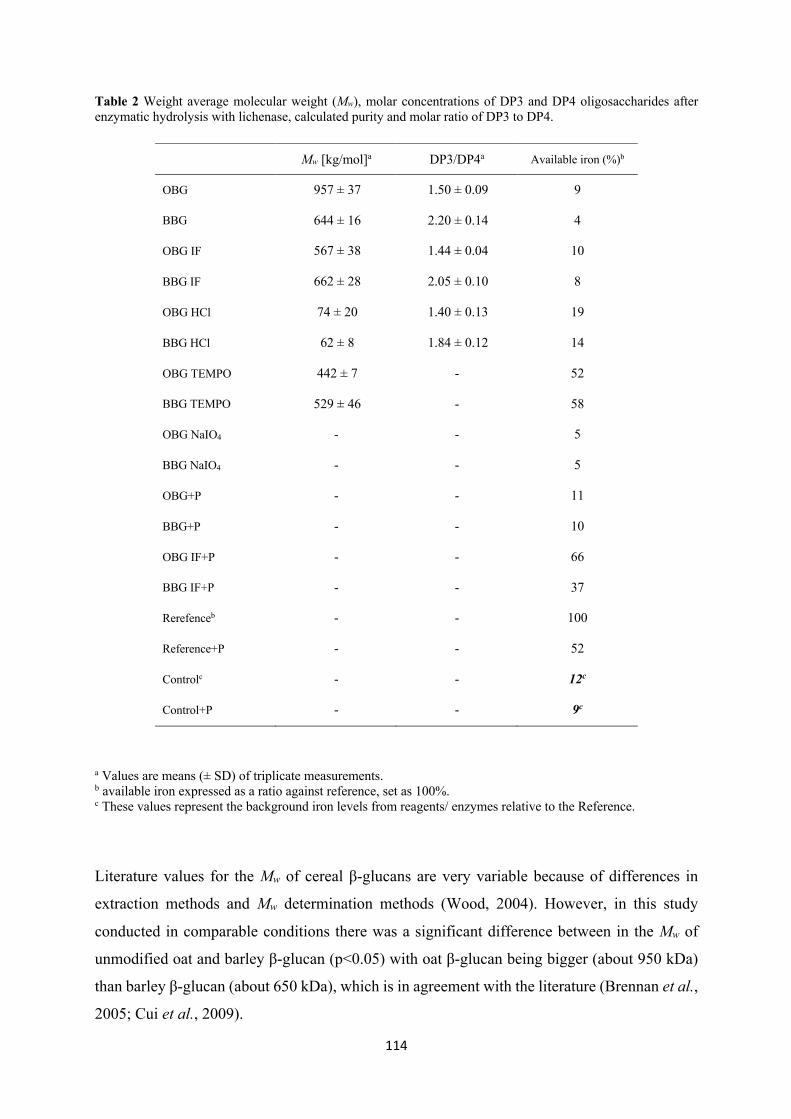
114
Table 2 Weight average molecular weight (Mw), molar concentrations of DP3 and DP4 oligosaccharides after enzymatic hydrolysis with lichenase, calculated purity and molar ratio of DP3 to DP4.
a Values are means (± SD) of triplicate measurements. b available iron expressed as a ratio against reference, set as 100%. c These values represent the background iron levels from reagents/ enzymes relative to the Reference.
Literature values for the Mw of cereal β-glucans are very variable because of differences in
extraction methods and Mw determination methods (Wood, 2004). However, in this study
conducted in comparable conditions there was a significant difference between in the Mw of
unmodified oat and barley β-glucan (p<0.05) with oat β-glucan being bigger (about 950 kDa)
than barley β-glucan (about 650 kDa), which is in agreement with the literature (Brennan et al.,
2005; Cui et al., 2009).
Mw [kg/mol]a DP3/DP4a Available iron (%)b
OBG 957 ± 37 1.50 ± 0.09 9
BBG 644 ± 16 2.20 ± 0.14 4
OBG IF 567 ± 38 1.44 ± 0.04 10
BBG IF 662 ± 28 2.05 ± 0.10 8
OBG HCl 74 ± 20 1.40 ± 0.13 19
BBG HCl 62 ± 8 1.84 ± 0.12 14
OBG TEMPO 442 ± 7 - 52
BBG TEMPO 529 ± 46 - 58
OBG NaIO4 - - 5
BBG NaIO4 - - 5
OBG+P - - 11
BBG+P - - 10
OBG IF+P - - 66
BBG IF+P - - 37
Rerefenceb - - 100
Reference+P - - 52
Controlc - - 12c
Control+P - - 9c

115
Results show that Chelex treatment reduced the Mw of oat β-glucan (-41%) but not that of barley
β-glucan. This could be due to slight differences in the preparation of the many batches of
Chelex treated β-glucan, but also to the slightly different composition of the two extracts.
Acid hydrolysis, on the other hand, led to a relatively high degree of chain scission. As
expected, the molecular size was significantly reduced, roughly by 13 times for oat and 10 times
for barley. This treatment was used to study the effect of the reduction of Mw on the iron binding
by cereal β-glucan without any other alteration of the structure.
TEMPO treatment reduced the Mw by roughly half for oat and only 20% for barley. TEMPO
oxidation was chosen because it is a reasonably mild modification that allows to investigate the
effect of oxidation on iron binding by β-glucan without dramatically reducing the Mw of the
fiber, while introducing potential new binding sites for iron, namely carboxylate groups.
There were no analyzable SEC-peaks for the NaIO4 oxidised β-glucan. In principle, sodium
periodate oxidation is not expected to directly lead to extensive chain scission. However,
extensive degradation of alginate upon oxidation with sodium periodate has been reported,
especially when complete oxidation was attempted (Balakrishnan et al., 2005). The sharp Mw
decrease after NaIO4 treatment might be due to the fact that the C=O groups introduced by
oxidation render the molecule more susceptible to β-elimination, causing breakage of the β-
glucan structure. Also, considering the crosslinking property of periodate oxidised
polysaccharides, the formation of protein-iron-β-glucan complexes can be hypothesised.
Crosslinking happens due to the formation of Schiff’s base between the amino groups of lysine
and the aldehyde groups of the polymer (Balakrishnan et al., 2005). According to Kristiansen
et al. (2010) oxidised polysaccharides show hydrolytic lability and the capacity of
macromolecular compaction as additional features which would support the hypothesis of the
formation of precipitating complexes. In the present study, since some degree of precipitation
was observed in the periodate treated samples, but not comparable to the amount of β-glucan
originally dissolved to prepare the solutions, probably both phenomena took place, so some of
the oxidized β-glucan is insoluble but also an extensive degradation of the polymer took place.
The measured molar ratio of DP3 to DP4 for the untreated oat and barley β-glucan extracts is
in the range of reported literature values. (Lazaridou et al., 2004; Lazaridou and Biliaderis,
2007; Izydorczyk et al., 2008) As for the modifications, Chelex treatment did not significantly
change the DP3 to DP4 ratio. Acid hydrolysis seemed to affect barley β-glucan in a different
way than oat β-glucan. The DP3:DP4 of the oat samples in fact, did not change significantly
after hydrolysis, while it decreased for the barley samples. The missing data for the oxidised
samples can be explained with the structural changes of the molecules, in particular the new

116
functional groups (carboxyls and aldehydes) introduced upon oxidation. First of all, they could
sterically hinder the lichenase from cleaving the (1→4)-glycosidic linkage of the glucose
residues. Secondly, there are probably no more native DP3 or DP4 left, since they got altered
during oxidation and hence would elute differently (or degrade under the highly basic HPAEC-
PAD conditions).
The introduction of these new functional groups is also expected to have an effect on the water
solubility of the oxidised β-glucan. Coseri et al. (2015) reported from their oxidation
experiments with cellulose that when the conversion of the hydroxyl to carboxylic groups
reached a certain level, the fully oxidised molecules tended to become water soluble. Indeed,
the solubilisation of the TEMPO oxidised β-glucan samples in the experiments was
unproblematic. In contrast, the sodium periodate oxidised β-glucan could not be completely
dissolved in water. Small, loose particles were always visible and if the dispersion was not
stirred or shaken, the particles would precipitate and sediment.
4.3 In vitro digestion and dialysis
After the in vitro digestion, the iron concentrations in the dialysis tubes were determined with
AAS. The iron concentration in the dialysates of all unmodified samples (OBG, BBG, OBG IF,
BBG IF) was less than 10% of iron concentration in the reference-dialysate, which means some
compounds in these samples are binding iron (p<0.001) (Figure 5 and Figure 6). Interestingly,
a comparison of the two references with and without phytase (Reference and Reference+P)
shows that the dialysate of Reference+P contains only half of the amount of iron compared to
the untreated Reference (p<0.001). These results suggest that phytase or a component of the
commercial product is able to bind iron, but, to the best of our knowledge, no literature
explaining this phenomenon is available. However, even proteins such as bovine serum albumin
(BSA) are known to bind iron in the presence of hydrogen carbonate ions (Ueno et al., 2014),
therefore, iron binding by the commercial phytase material is a reasonable explanation (Hurrell
et al.,1992).
The dialysates of OBG+P and BBG+P both contained >5 times less iron than the Reference+P,
indicating that, despite of the phytase treatment that should hydrolyse phytic acid, something is
still binding iron (p<0.001). If a Chelex treatment is applied before the phytase, however, iron
binding is as expected diminished, with the dialysate of OBG IF+P exhibiting the highest iron
concentration of all samples and significantly higher than Reference+P (p<0.05). The iron
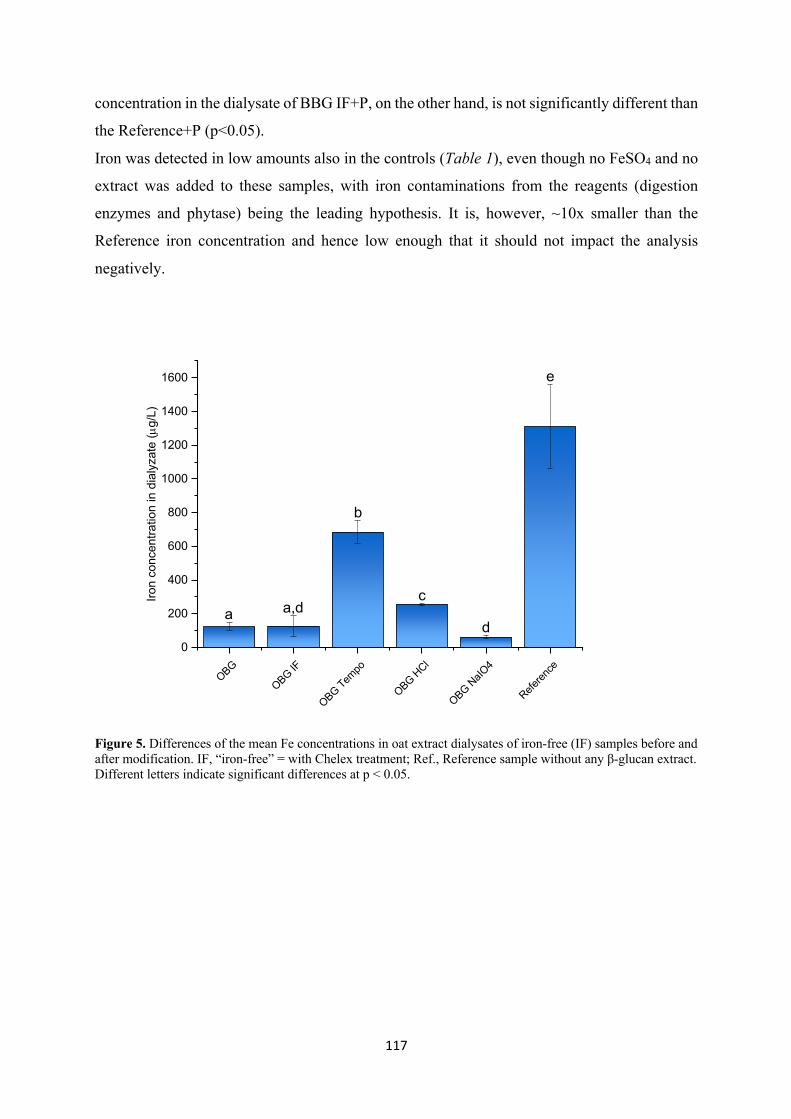
117
concentration in the dialysate of BBG IF+P, on the other hand, is not significantly different than
the Reference+P (p<0.05).
Iron was detected in low amounts also in the controls (Table 1), even though no FeSO4 and no
extract was added to these samples, with iron contaminations from the reagents (digestion
enzymes and phytase) being the leading hypothesis. It is, however, ~10x smaller than the
Reference iron concentration and hence low enough that it should not impact the analysis
negatively.
Figure 5. Differences of the mean Fe concentrations in oat extract dialysates of iron-free (IF) samples before and after modification. IF, “iron-free” = with Chelex treatment; Ref., Reference sample without any β-glucan extract. Different letters indicate significant differences at p < 0.05.
OBG
OBG IF
OBG Tempo
OBG HCl
OBG NaIO
4
Referen
ce
0
200
400
600
800
1000
1200
1400
1600 e
c
b
a,d
Iron
conc
entra
tion
in d
ialy
zate
(µg/
L)
ad
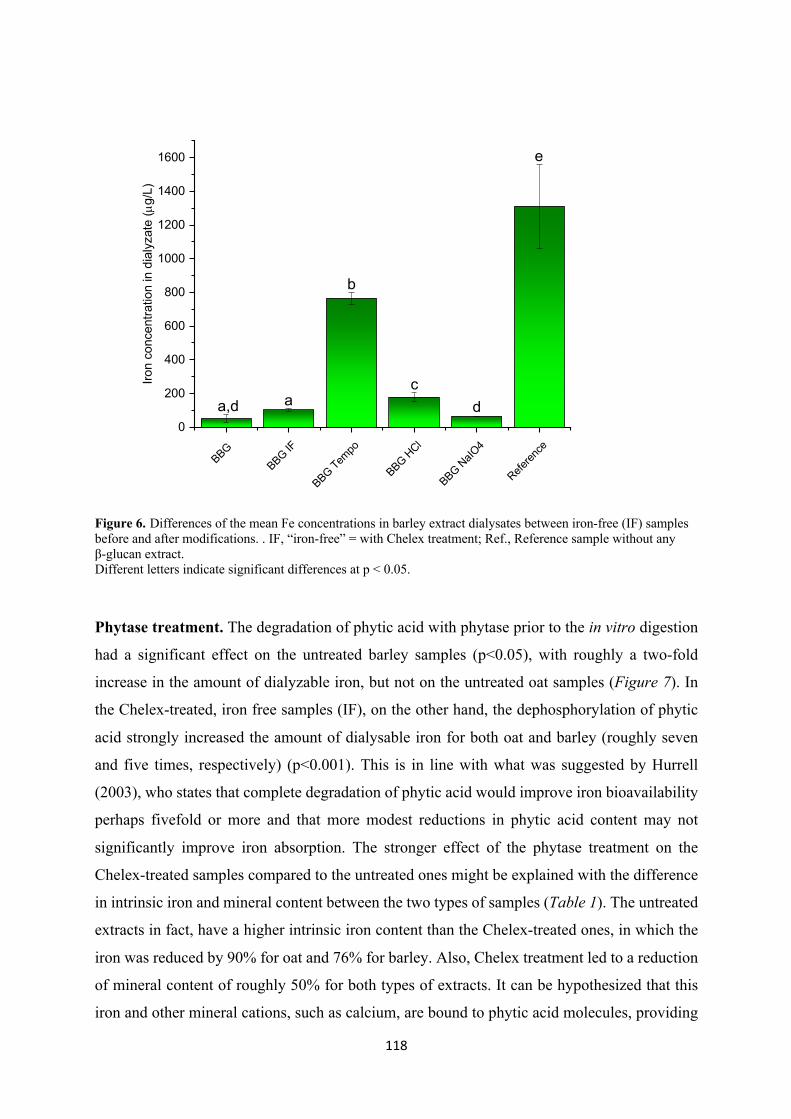
118
Figure 6. Differences of the mean Fe concentrations in barley extract dialysates between iron-free (IF) samples before and after modifications. . IF, “iron-free” = with Chelex treatment; Ref., Reference sample without any β-glucan extract. Different letters indicate significant differences at p < 0.05.
Phytase treatment. The degradation of phytic acid with phytase prior to the in vitro digestion
had a significant effect on the untreated barley samples (p<0.05), with roughly a two-fold
increase in the amount of dialyzable iron, but not on the untreated oat samples (Figure 7). In
the Chelex-treated, iron free samples (IF), on the other hand, the dephosphorylation of phytic
acid strongly increased the amount of dialysable iron for both oat and barley (roughly seven
and five times, respectively) (p<0.001). This is in line with what was suggested by Hurrell
(2003), who states that complete degradation of phytic acid would improve iron bioavailability
perhaps fivefold or more and that more modest reductions in phytic acid content may not
significantly improve iron absorption. The stronger effect of the phytase treatment on the
Chelex-treated samples compared to the untreated ones might be explained with the difference
in intrinsic iron and mineral content between the two types of samples (Table 1). The untreated
extracts in fact, have a higher intrinsic iron content than the Chelex-treated ones, in which the
iron was reduced by 90% for oat and 76% for barley. Also, Chelex treatment led to a reduction
of mineral content of roughly 50% for both types of extracts. It can be hypothesized that this
iron and other mineral cations, such as calcium, are bound to phytic acid molecules, providing
BBG
BBG IF
BBG Tempo
BBG HCl
BBG NaIO
4
Referen
ce
0
200
400
600
800
1000
1200
1400
1600 e
dc
a
Iron
conc
entra
tion
in d
ialy
zate
(µg/
L)
a,d
b
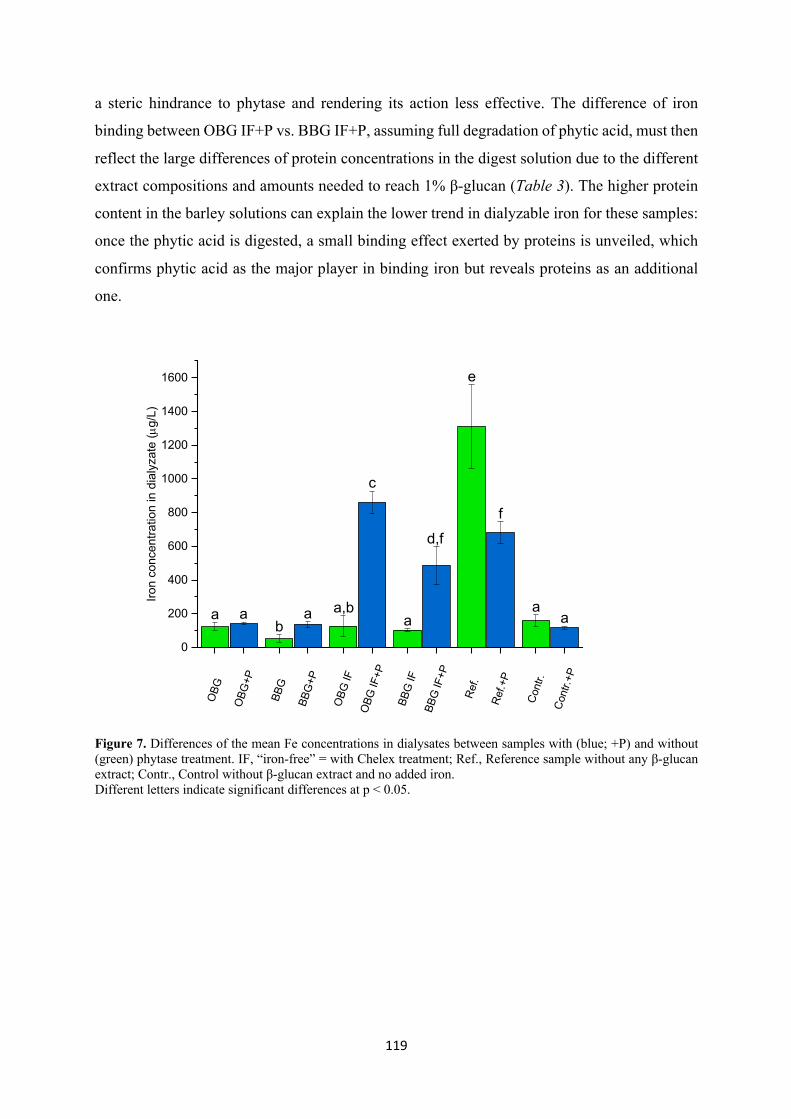
119
a steric hindrance to phytase and rendering its action less effective. The difference of iron
binding between OBG IF+P vs. BBG IF+P, assuming full degradation of phytic acid, must then
reflect the large differences of protein concentrations in the digest solution due to the different
extract compositions and amounts needed to reach 1% β-glucan (Table 3). The higher protein
content in the barley solutions can explain the lower trend in dialyzable iron for these samples:
once the phytic acid is digested, a small binding effect exerted by proteins is unveiled, which
confirms phytic acid as the major player in binding iron but reveals proteins as an additional
one.
Figure 7. Differences of the mean Fe concentrations in dialysates between samples with (blue; +P) and without (green) phytase treatment. IF, “iron-free” = with Chelex treatment; Ref., Reference sample without any β-glucan extract; Contr., Control without β-glucan extract and no added iron. Different letters indicate significant differences at p < 0.05.
OBG
+P
BBG
+P
OBG
IF+P
BBG
IF+P
Ref.+
P
Cont
r.+P
OBG
BBG
OBG
IF
BBG
IF
Ref.
Cont
r.
0
200
400
600
800
1000
1200
1400
1600
f
aa
e
d,f
a
c
ba,baa
Iron
conc
entra
tion
in d
ialy
zate
(µg/
L)
a
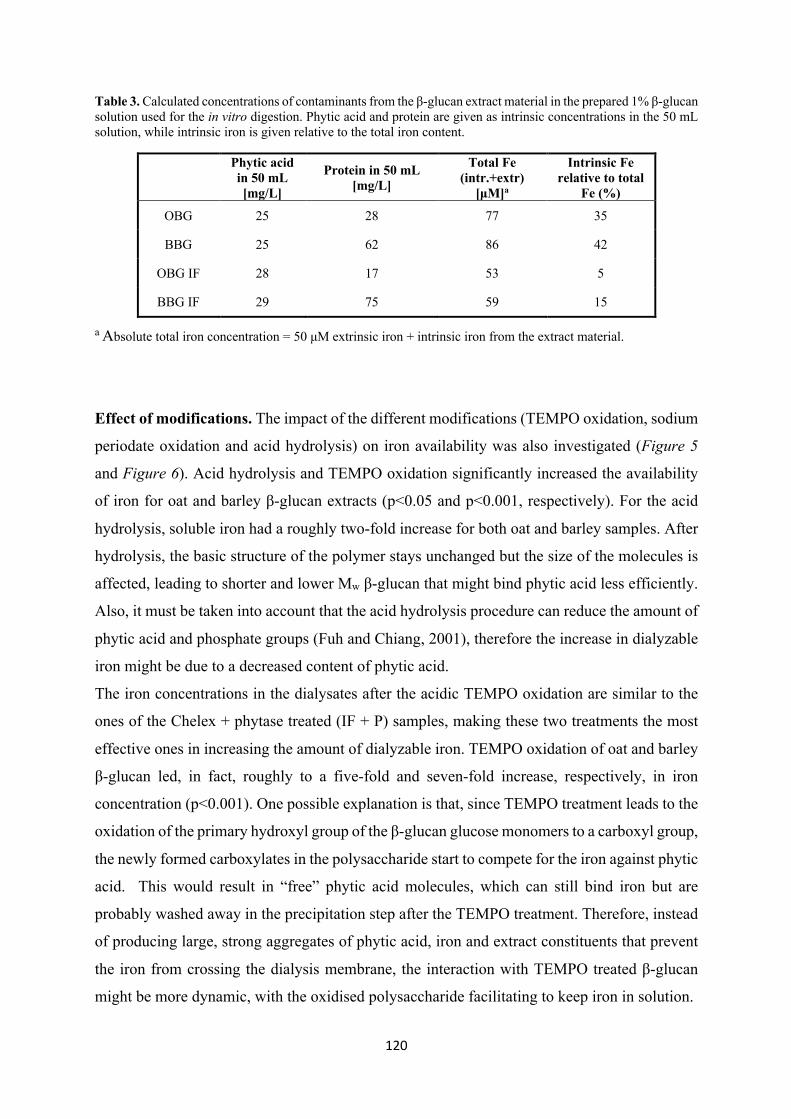
120
Table 3. Calculated concentrations of contaminants from the β-glucan extract material in the prepared 1% β-glucan solution used for the in vitro digestion. Phytic acid and protein are given as intrinsic concentrations in the 50 mL solution, while intrinsic iron is given relative to the total iron content.
Phytic acid in 50 mL [mg/L]
Protein in 50 mL [mg/L]
Total Fe (intr.+extr)
[μM]a
Intrinsic Fe relative to total
Fe (%)
OBG 25 28 77 35
BBG 25 62 86 42
OBG IF 28 17 53 5
BBG IF 29 75 59 15
a Absolute total iron concentration = 50 μM extrinsic iron + intrinsic iron from the extract material.
Effect of modifications. The impact of the different modifications (TEMPO oxidation, sodium
periodate oxidation and acid hydrolysis) on iron availability was also investigated (Figure 5
and Figure 6). Acid hydrolysis and TEMPO oxidation significantly increased the availability
of iron for oat and barley β-glucan extracts (p<0.05 and p<0.001, respectively). For the acid
hydrolysis, soluble iron had a roughly two-fold increase for both oat and barley samples. After
hydrolysis, the basic structure of the polymer stays unchanged but the size of the molecules is
affected, leading to shorter and lower Mw β-glucan that might bind phytic acid less efficiently.
Also, it must be taken into account that the acid hydrolysis procedure can reduce the amount of
phytic acid and phosphate groups (Fuh and Chiang, 2001), therefore the increase in dialyzable
iron might be due to a decreased content of phytic acid.
The iron concentrations in the dialysates after the acidic TEMPO oxidation are similar to the
ones of the Chelex + phytase treated (IF + P) samples, making these two treatments the most
effective ones in increasing the amount of dialyzable iron. TEMPO oxidation of oat and barley
β-glucan led, in fact, roughly to a five-fold and seven-fold increase, respectively, in iron
concentration (p<0.001). One possible explanation is that, since TEMPO treatment leads to the
oxidation of the primary hydroxyl group of the β-glucan glucose monomers to a carboxyl group,
the newly formed carboxylates in the polysaccharide start to compete for the iron against phytic
acid. This would result in “free” phytic acid molecules, which can still bind iron but are
probably washed away in the precipitation step after the TEMPO treatment. Therefore, instead
of producing large, strong aggregates of phytic acid, iron and extract constituents that prevent
the iron from crossing the dialysis membrane, the interaction with TEMPO treated β-glucan
might be more dynamic, with the oxidised polysaccharide facilitating to keep iron in solution.

121
In addition to this, another possible explanation for the higher dialyzable iron content is that the
reagents used for TEMPO oxidation (TEMPO, sodium hypochlorite) could also have an effect
on the phytic acid molecule. The slightly acidic conditions used during the treatment might
destabilise the phytic acid-iron complex, favouring the dephosphorylation of the inositols
similarly as during natural digestion processes. In lower phosphorylated inositol phosphates,
the free hydroxyl groups can be oxidised, which promotes further dephosphorylation of the
molecule by β-elimination due to an acidifed α-C-H bond. The phosphate groups as well as iron
can therefore be released during the TEMPO treatment and washed away in the precipitation
step to isolate the modified extract.
The iron concentration in the dialysates of the sodium periodate samples (NaIO4) was
statistically significantly lower than that of the Chelex-treated samples (IF) in the case of barley,
but the difference was so small that it is not relevant. No significant difference was found for
oat (p<0.05).
Differences between oat and barley β-glucan. T-tests showed that barley β-glucan extract has
stronger iron binding properties than oat β-glucan extract for most of the samples (untreated,
Chelex + phytase treated, and acid hydrolysed samples) (p<0.05) which is in discordance with
previous studies (Faure et al., 2015). To explain this, the purities of the oat and barley β-glucan
extracts used in the present study must be considered, since two sets of samples with differing
prerequisites are being compared. First of all, it must be clarified that the final β-glucan
concentration was 1% in every sample, but, since oat and barley β-glucan extracts had different
purities, the amount of extract used to prepare the samples was not the same. Barley β-glucan
extract is less pure and thus a higher extract concentration was required to prepare the samples,
causing the compounds other than β-glucan binding iron to have a stronger effect. In particular,
while the phytic acid content did not vary too dramatically between the different samples, the
protein content was much higher in the barley than in the oat β-glucan solution (Table 3) and it
is known that iron can be bound by insoluble peptides containing serine phosphate or high
proportions of carboxylic acid groups. (Hurrell et al., 1992) Likewise, the intrinsic content of
iron was higher in barley than in oat, which also remained higher after Chelex treatment.
Although it cannot be excluded, the lack of any correlation between intrinsic iron content of the
extract vs. dialyzable iron concentration after the in vitro digestion suggests that most of the
intrinsic iron was not released from the extract matrix, or at the very least, did not significantly
interfere with the analysis. The difference in fine structure between oat and barley β-glucan
might have only secondary and probably negligible effects. Due to its DP3:DP4, in fact, barley

122
is more prone to gelation and the formation of these networks might cause a physical
entrapment of extract costituents such as phytic acid that can hinder the access to enzymes. To
summarize, based on the results of the present study and on recent data found in literature
(Wang, 2017 a&b), the combination of phytic acid content plus an additional contribution from
proteins dictates the iron binding effects in the presence of cereal β-glucan.
Precipitation issues. After storage of the samples in the fridge after in vitro digestion, before
an aliquot was taken and HCl added to it to make a 0.1M solution for AAS analysis, visible
precipitates had formed in the falcon tubes. The formation of precipitates in β-glucan solutions
has been reported before and compositional analysis showed they consisted of protein, phytic
acid and minerals (R. Kivelä et al., 2009). Presumably, the precipitates in our samples contained
iron, because rough estimations of the total iron in the system after the digestion did not add up
to the expected total amount (data not shown). The formation of precipitates in the digest does
not seem to be problematic since this might similarly be happening in vivo, where it its known
that iron can precipitate either due to insoluble complex or iron hydroxides formation, lowering
iron absorption. Therefore, in order to estimate iron bioavailability correctly, these precipitates
should not be taken into account and this is achieved thanks to the dialysis method, which only
allows solubilized iron to pass into the membrane.
Although there is an abundance of literature about iron bioavailability in foods, only few studies
investigate or discuss the role of the partly soluble dietary fibre such as β-glucan (Platt et al.,
1984; Torre et al., 1991; Faure et al., 2014, 2015; Wang et al., 2017; Wang, Zhan, et al., 2017).
However, it is nowadays an established fact that phytic acid plays a major role in the observed
reduced bioavailability of the iron present in cereals and other vegetable food, as recently
confirmed by Wang et al. (2017 a&b). As previously discussed, both oat and barley β-glucan
extracts contain high levels of phytic acid (Table 1) and it is well known that even small
amounts of phytic acid can bind considerable amounts of iron (Schlemmer et al., 2009; Nielsen
et al., 2013). Therefore, the key role of phytic acid in iron binding is once more confirmed by
the present study. It has to be clarified that the results discussed here do not represent the true
iron bioavailability, which is a complex combination of factors. They offer, however, a relative
comparison, which can help to understand how processing of oat and barley β-glucan affects
iron binding, and, as a consequence, iron bioavailability.

123
5. Conclusions
Oat and barley β-glucan extracts, native or modified (TEMPO oxidation, sodium periodate
oxidation and acid hydrolysis), were studied to investigate the impact of molecular structure
and phytic acid content on iron bioavailability. To do this, an in vitro digestion featuring a
reverse dialysis system was performed to mimic the behaviour of the fiber extracts in the human
gastro-intestinal tract in the presence of iron. The results confirm that no direct binding between
β-glucan and iron exists. On the other hand, they confirm the major impact phytic acid has on
the inhibition of iron absorption and suggest that protein also play a part in iron binding.
TEMPO oxidation and dephytinization of Chelex-treated extracts were the most effective
treatments for improving iron bioavailability, both for oat and barley samples. Extracts from
barley showed in all cases at the very least an equal or a more significant reduction of iron
bioavailability than oat, which can be either due to the different compositions of the β-glucan
extracts or an indirect effect due to the different fine structures of β-glucan from the two
different cereal sources. Phytase treatment was only successful in liberating iron when the
contents of minerals that complex phytic acid were reduced first, which has major implications
when attempting to increase iron bioavailability through phytase treatments in β-glucan
containing food products. Additional in vivo and in vitro experiments studying the interactions
between β-glucan, phytic acid and iron are required to further investigate these mechanisms and
ensure optimal iron bioavailability in foods.
6. Acknowledgements
The study was carried out within the framework of the Healthy Nutrition and Sustainable Food
Production project (National Research Programme NRP 69), funded by the Swiss National
Science Foundation. We thank Prof. Peter Fischer (Laboratory of Food Process Engineering,
ETH Zurich) for kindly allowing us to use the rheometer. We also thank Prof. Zimmermann
(Laboratory of Human Nutrition, ETH Zurich) for letting us use the AAS and Mr. Christophe
Zeder for helping us with the measurements.
References
ADDIN Mendeley Bibliography CSL_BIBLIOGRAPHYXAbbaspour, N., Hurrell, R. and Kelishadi, R. (2015) ‘Review on iron and its importance for human health Review on iron and its importance for human health’, Journal of Research in Medical Sciences, 19(FEBRUARY 2014), pp. 1–9.

124
Balakrishnan, B., Lesieur, S., Labarre, D. and Jayakrishnan, A. (2005) ‘Periodate oxidation of sodium alginate in water and in ethanol-water mixture: A comparative study’, Carbohydrate Research, 340(7), pp. 1425–1429. doi: 10.1016/j.carres.2005.02.028.
Bhatty, R. . (1995) ‘Laboratory and pilot plant extraction and purification of β-glucans from hull-less barley and oat brans’, Journal of Cereal Science, 22(2), pp. 163–170. doi: 10.1016/0733-5210(95)90046-2.
Bio-Rad (no date) ‘Chelex® 100 and Chelex 20 Chelating Ion Exchange Resin Instruction Manual’, Bio-Rad Laboratories, pp. 1–24.
Boulos, S. and Nyström, L. (2016) ‘UPLC-MS/MS investigation of β-glucan oligosaccharide oxidation’, Analyst. Royal Society of Chemistry, 141(24), pp. 6533–6548. doi: 10.1039/c6an01125j.
Brennan, C. S. and Cleary, L. J. (2005) ‘The potential use of cereal (1→3,1→4)-β-d-glucans as functional food ingredients’, Journal of Cereal Science, 42(1), pp. 1–13. doi: 10.1016/j.jcs.2005.01.002.
Camaschella, C. (2015) ‘Iron-Deficiency Anemia’, New England Journal of Medicine. Massachusetts Medical Society, 372(19), pp. 1832–1843. doi: 10.1056/NEJMra1401038.
Coseri, S., Biliuta, G., Zemljič, L. F., Srndovic, J. S., Larsson, P. T., Strnad, S., Kreže, T., Naderi, A. and Lindström, T. (2015) ‘One-shot carboxylation of microcrystalline cellulose in the presence of nitroxyl radicals and sodium periodate’, RSC Advances, 5(104), pp. 85889–85897. doi: 10.1039/C5RA16183E.
Cui, S. W. and Wang, Q. (2009) ‘Cell wall polysaccharides in cereals: chemical structures and functional properties’, Structural Chemistry, 20(2), pp. 291–297. doi: 10.1007/s11224-009-9441-0.
EFSA Panel on Dietetic Products Nutrition and Allergies (NDA) (2010) ‘Scientific Opinion on the substantiation of health claims related to beta-glucans and maintenance or achievement of normal blood glucose concentrations ( ID 756 , 802 , 2935 ) pursuant to Article 13 ( 1 ) of’, EFSA Journal, 8(1924), pp. 1–14. doi: 10.2903/j.efsa.2010.1482.
European Food Safety Authority (EFSA) (2011) ‘Scientific Opinion on the substantiation of a health claim related to barley beta-glucans and lowering of blood cholesterol and reduced risk of (coronary) heart disease pursuant to Article 14 of Regulation (EC) No 1924/2006’, EFSA Journal, 9(12), p. 2470. doi: 10.2903/j.efsa.2011.2470.
Faure, A. M., Koppenol, W. H. and Nyström, L. (2015) ‘Iron(II) binding by cereal beta-glucan.’, Carbohydrate polymers, 115, pp. 739–43. doi: 10.1016/j.carbpol.2014.07.038.
Faure, A. M., Sánchez-Ferrer, A., Zabara, A., Andersen, M. L. and Nyström, L. (2014) ‘Modulating the structural properties of β-D-glucan degradation products by alternative reaction pathways.’, Carbohydrate polymers, 99, pp. 679–86. doi: 10.1016/j.carbpol.2013.08.022.
(FDA) F.a.D.A. 21 CFR 101.81 (2009) ‘Health claims: Soluble fiber from certain foods and risk of coronary heart disease (CHD)’, in Regulatory Information, pp. 143–148.
Fuh, W. S. and Chiang, B. H. (2001) ‘Dephytinisation of rice bran and manufacturing a new food ingredient’, Journal of the Science of Food and Agriculture, 81(15), pp. 1419–1425. doi: 10.1002/jsfa.962.
Hallberg, L., Rossander, L. and Skaanberg, A. B. (1987) ‘Phytates and the inhibitory effect of bran on iron absorption in man’, American Journal of Clinical Nutrition, 45(5), pp. 988–996. doi: 10.1093/ajcn/45.5.988.
Haros, M., Rosell, C. M. and Benedito, C. (2001) ‘Fungal phytase as a potential breadmaking additive’, European Food Research and Technology, 213(4–5), pp. 317–322. doi: 10.1007/s002170100396.

125
Hemalatha, S., Platel, K. and Srinivasan, K. (2007) ‘Zinc and iron contents and their bioaccessibility in cereals and pulses consumed in India’, Food Chemistry, 102(4), pp. 1328–1336. doi: 10.1016/j.foodchem.2006.07.015.
Hurrell, R. and Egli, I. (2010) ‘Iron bioavailability and dietary reference values.’, The American journal of clinical nutrition, 91(5), pp. 1461S-1467S. doi: 10.3945/ajcn.2010.28674F.Am.
Hurrell, R. F. (1997) ‘Bioavailability of iron.’, European journal of clinical nutrition, 51 Suppl 1(1), pp. S4–S8.
Hurrell, R. F., Reddy, M. B., Juillerat, M. A. and Cook, J. D. (2003) ‘Degradation of phytic acid in cereal porridges improves iron absorption by human subjects’, American Journal of Clinical Nutrition, 77(5), pp. 1213–1219. doi: 10.1093/ajcn/77.5.1213.
Hurrell RF, Juillerat MA, Reddy MB, Lynch SR, Dassenko SA, C. J. (1992) ‘Soy protein, phytate, and iron absorption in humans.’, Am J. Clin. Nutr, 56(3), pp. 573–8.
Izydorczyk, M. S. and Dexter, J. E. (2008) ‘Barley β-glucans and arabinoxylans: Molecular structure, physicochemical properties, and uses in food products–a Review’, Food Research International. Elsevier Ltd, 41(9), pp. 850–868. doi: 10.1016/j.foodres.2008.04.001.
Johansson, L., Virkki, L., Anttila, H., Esselström, H., Tuomainen, P. and Sontag-Strohm, T. (2006) ‘Hydrolysis of β-glucan’, Food Chemistry, 97(1), pp. 71–79. doi: 10.1016/j.foodchem.2005.03.031.
Kennefick, S. and Cashman, K. D. (2000) ‘Inhibitory effect of wheat fibre extract on calcium absorption in Caco-2 cells: Evidence for a role of associated phytate rather than fibre per se’, European Journal of Nutrition, 39(1), pp. 12–17. doi: 10.1007/s003940050071.
Kivelä, R., Henniges, U., Sontag-Strohm, T. and Potthast, A. (2012) ‘Oxidation of oat β-glucan in aqueous solutions during processing’, Carbohydrate Polymers, 87(1), pp. 589–597. doi: 10.1016/j.carbpol.2011.08.028.
Kivelä, R., Nyström, L., Salovaara, H. and Sontag-Strohm, T. (2009) ‘Role of oxidative cleavage and acid hydrolysis of oat beta-glucan in modelled beverage conditions’, Journal of Cereal Science, 50(2), pp. 190–197. doi: 10.1016/j.jcs.2009.04.012.
Kivelä, R., Sontag-Strohm, T., Loponen, J., Tuomainen, P. and Nyström, L. (2011) ‘Oxidative and radical mediated cleavage of β-glucan in thermal treatments’, Carbohydrate Polymers, 85(3), pp. 645–652. doi: 10.1016/j.carbpol.2011.03.026.
Larsson, M., Minekus, M. and Havenaar, R. (1997) ‘Estimation of the bioavailability of iron and phosphorus in cereals using a dynamic in vitro gastrointestinal model’, Journal of the Science of Food and Agriculture, 74(1), pp. 99–106. doi: 10.1002/(SICI)1097-0010(199705)74:1<99::AID-JSFA775>3.0.CO;2-G.
Lazaridou, A., Biliaderis, C. ., Micha-Screttas, M. and Steele, B. . (2004) ‘A comparative study on structure–function relations of mixed-linkage (1→3), (1→4) linear β-d-glucans’, Food Hydrocolloids, 18(5), pp. 837–855. doi: 10.1016/j.foodhyd.2004.01.002.
Lazaridou, A. and Biliaderis, C. G. (2007) ‘Molecular aspects of cereal β-glucan functionality: Physical properties, technological applications and physiological effects’, Journal of Cereal Science, 46(2), pp. 101–118. doi: 10.1016/j.jcs.2007.05.003.
Lee, K. Y., Park, S. Y. and Lee, H. G. (2011) ‘Effect of oat β-glucan and its oxidised derivative on the quality characteristics of sponge cake’, International Journal of Food Science & Technology, 46(12), pp. 2663–2668. doi: 10.1111/j.1365-2621.2011.02798.x.
Mäkelä, N., Sontag-Strohm, T. and Maina, N. H. (2015) ‘The oxidative degradation of barley β-glucan in the presence of ascorbic acid or hydrogen peroxide’, Carbohydrate Polymers. Elsevier Ltd., 123, pp. 390–395. doi: 10.1016/j.carbpol.2015.01.037.

126
Miller, D., Schricker, R., Ph, D. and Rasmussen, R. (1981) ‘An in vitro availability method for estimation iron availability from meals’, The American Journal of Clinical Nutrition, 43, pp. 2248–2256.
De Moura, F. A., Pereira, J. M., Da Silva, D. O., Zavareze, E. D. R., Da Silveira Moreira, A., Helbig, E. and Dias, A. R. G. (2011) ‘Effects of oxidative treatment on the physicochemical, rheological and functional properties of oat β-glucan’, Food Chemistry. Elsevier Ltd, 128(4), pp. 982–987. doi: 10.1016/j.foodchem.2011.04.003.
Nielsen, A. V. F., Tetens, I. and Meyer, A. S. (2013) ‘Potential of phytase-mediated iron release from cereal-based foods: A quantitative view’, Nutrients, 5(8), pp. 3074–3098. doi: 10.3390/nu5083074.
Park, S. Y., Bae, I. Y., Lee, S. and Lee, H. G. (2009) ‘Physicochemical and Hypocholesterolemic Characterization of Oxidized Oat β-Glucan’, Journal of Agricultural and Food Chemistry, 57(2), pp. 439–443. doi: 10.1021/jf802811b.
Platt, S. R. and Clydesdale, F. M. (1984) ‘Binding of Iron by Cellulose, Lignin, Sodium Phytate and Beta-Glucan, Alone and in Combination, Under Simulated Gastrointestinal pH Conditions’, Journal of Food Science. Blackwell Publishing Ltd, 49(2), pp. 531–535. doi: 10.1111/j.1365-2621.1984.tb12460.x.
Reddy, M. B. and Cook, J. D. (1991) ‘Assessment of dietary determinants of nonheme-iron absorption in humans and rats’, American Journal of Clinical Nutrition, 54(4), pp. 723–728. doi: 10.1093/ajcn/54.4.723.
Sandberg, A. S. (2005) ‘Methods and options for in vitro dialyzability; benefits and limitations’, International Journal for Vitamin and Nutrition Research, 75(6), pp. 395–404. doi: 10.1024/0300-9831.75.6.395.
Schlemmer, U., Frølich, W., Prieto, R. M. and Grases, F. (2009) ‘Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis’, Molecular Nutrition and Food Research, 53(SUPPL. 2), pp. 330–375. doi: 10.1002/mnfr.200900099.
Simpson, K. M., Morris, E. R. and Cook, J. D. (1981) ‘The inhibitory effect of bran on iron absorption in man’, The American Journal of Clinical Nutrition, pp. 1469–1478.
Torre, M., Rodriguez, A. R. and Saura-Calixto, F. (1991) ‘Effects of Dietary Fiber and Phytic Acid on Mineral Availability’, Critical Reviews in Food Science and Nutrition, 30(1), pp. 1–22. doi: 10.1080/10408399109527539.
Ueno, H. M., Urazono, H. and Kobayashi, T. (2014) ‘Serum albumin forms a lactoferrin-like soluble iron-binding complex in presence of hydrogen carbonate ions’, Food Chemistry. Elsevier Ltd, 145, pp. 90–94. doi: 10.1016/j.foodchem.2013.07.143.
Wang, Y. J., Maina, N. H., Ekholm, P., Lampi, A. M. and Sontag-Strohm, T. (2017) ‘Retardation of oxidation by residual phytate in purified cereal β-glucans’, Food Hydrocolloids, 66, pp. 161–167. doi: 10.1016/j.foodhyd.2016.11.019.
Wang, Y. J., Zhan, R., Sontag-Strohm, T. and Maina, N. H. (2017) ‘The protective role of phytate in the oxidative degradation of cereal beta-glucans’, Carbohydrate Polymers. Elsevier Ltd., 169, pp. 220–226. doi: 10.1016/j.carbpol.2017.04.016.
Wood, P. J. (2004) ‘Relationships between solution properties of cereal β-glucans and physiological effects — a review’, Trends in Food Science & Technology, 15(6), pp. 313–320. doi: 10.1016/j.tifs.2003.03.001.

127
III. Investigation of cereal β-glucan degradation following
thermal treatments and extrusion
Elena Marasca, Samy Boulos and Laura Nyström
ABSTRACT
Cereal β-glucan is a soluble dietary fiber with well-established health promoting benefits, which
have been shown to depend mainly on its high molecular weight and on its ability to form highly
viscous solutions. During processing, β-glucan can undergo many alterations, such as chain
scission and introduction of new functional groups, which can affect its technological and health
properties. This study investigated the effects of thermal treatments (125°C and 175°C) and
extrusion cooking (120°C and 150°C, 25% moisture content) on the molecular structure of
β-glucan extracted from oat and barley flour. Both viscosity and molecular weight decreased
during the thermal treatments, and the degradation was more intense with increasing
temperatures. For the extruded samples, no significant structural change took place for oat
β-glucan, while viscosity had a dramatic increase in the case of barley, in particular for the
samples extruded at 120°C. Oxidative reactions linked to Fenton chemistry most probably took
place during thermal treatments, and the oxidation products were investigated through carbonyl
labelling by reductive amination, followed by enzymatic digestion with lichenase and
β-glucosidase, purification/fractionation by SPE and analysis by UPLC-MS with fluorescent
detection. Several lytic as well as non-lytic oxidation products could be detected in particular
in the case of the samples treated at 175°C, together with a marked increase in the reducing end
unit concentration, confirming this treatment as the one causing the highest degree of oxidation
and depolymerisation, in line with the reduction of Mw and viscosity. No conclusive results
were found in the case of the extruded samples, therefore further investigation will be required
to clarify this process.
1. Introduction
Cereal β-glucan, a soluble dietary fiber found mainly in oat and barley, has drawn the attention
of the scientific community in recent years due to its beneficial health properties, such as the
cholesterol lowering effect and blood glucose regulation, confirmed by governmental health

128
and food agencies (FDA, 2009; EFSA, 2010). These beneficial health effects have often been
linked to the ability of this polysaccharide to form highly viscous solutions in the
gastrointestinal tract, preventing or slowing down the absorption of compounds associated with
diseases such as coronary heart disease and diabetes (Wood, 2010). β-Glucan is a
homopolysaccharide of D-glucopyranose arranged as blocks of consecutive (1→4)-linked β-D-
glucose residues separated by single (1→3)-linkages. The flexible (1→3)-linkages interrupt the
rigid cellulose-like sections formed by (1→4)-linked glucose units, thus increasing the
solubility of this polymer in water and its ability to form highly viscous solutions (Lazaridou et
al., 2007). The viscosity of the solutions depends on the molecular weight (Mw), which in the
native form can go up to 1000-3000 kDa, on the solubility and on the extractability of β-glucan
(Lazaridou and Biliaderis, 2007). However, since during food production cereals are subjected
to a high degree of processing (milling, baking, extrusion, etc.), these characteristics can be
altered, which leads to a modification of the health and technological properties of the fiber
(Lazaridou et al., 2007; Wood, 2007).
The degradation of β-glucan in foods, with consequent loss of viscosity and reduction of Mw,
has been demonstrated to take place already during storage, and this has been traditionally
considered to originate from enzymatic or acid hydrolysis (Lazaridou and Biliaderis, 2007).
Kivelä et al. (2009), however, proved that while the addition of citric and malic acids in an
unpurified oat β-glucan extract only caused a moderate effect and only with the simultaneous
presence of heat, the addition of ascorbic acid triggered an immediate decrease in viscosity of
the solution and of the Mw of β-glucan. This led to reject the hypothesis of acid hydrolysis,
which anyway requires very low pH and high temperature to be effective (Johansson et al.,
2006), being the main culprit of β-glucan molecular degradation, and to propose instead that
the depolymerization of β-glucan in the ascorbic acid treated extract was induced by metal-
catalysed hydroxyl radicals (Kivelä et al., 2009). Macromolecules are known to be susceptible
to degradation induced by reactive oxygen species (ROS). Among these, the hydroxyl radical
(•OH) has been established to be able to scission polysaccharides (Fry, 1998) and has recently
been confirmed, by means of indirect spin trapping and electron spin resonance (ESR)
spectroscopy, as the main responsible for the loss in viscosity and Mw of cereal β-glucan (Faure
et al., 2013).
The production of •OH radicals is believed to take place through the so-called Fenton reaction
(Fenton, 1894) (1), in which catalytic amounts of reduced metals such as Fe2+, Cu+ and Zn+ can
generate aggressive •OH from the modestly reactive H2O2.

129
Fe2+ + H2O2 → Fe3+ + OH− + •OH (1)
In order to initiate the Fenton reaction, a reducing agent such as AH2 is required to reduce the
catalysing metal to the active state and the dissolved oxygen to H2O2. All these reagents are
normally found in the aqueous phases of cereal foods, which also contain the soluble fibers
(Slavin et al., 1999).
Kivelä et al. (2011) showed that oxidative degradation can take place also during thermal
treatment (95°C and 120°C) of oat β-glucan solutions, with consequent reduction of viscosity
and Mw. Fenton chemistry was confirmed to play an important role in the degradation, since
the addition of Fe2+, a known catalyst for the Fenton reaction, accelerated the thermal
degradation. Also, oxidative free radicals and hydrogen peroxide were formed during heating,
corroborating the hypothesis of extensive oxidation taking place. In a following study, Kivelä
et al. (2012) demonstrated the formation of carbonyl groups along the β-glucan chain
simultaneously with chain cleavage during thermal treatment (120°C). To do this, they labelled
the carbonyl groups with carbazole-9-carboxylic acid [2-(2-aminooxyethoxy)ethoxy]amide
(COOA) and subsequently determined their concentration by size exclusion chromatography.
They reported an increase in the concentration of carbonyl groups of an extract of native oat
β-glucan from 1 to 12µmol/g, that is 11µmol/g or 0.2 carbonyls per 100 repeating units after 30
min at 120°C. Both studies showed that the strength of thermal degradation in solution is
affected by co-extracted compounds such as phytates, proteins and minerals (Kivelä et al.,
2011, 2012).
High pressure treatments such as homogenization have also been shown to cause
depolymerization of β-glucan, with consequent loss of viscosity (Kivelä et al., 2010, 2012).
During homogenization, also a certain degree of oxidation takes place, even though the main
degradation mechanism is the hydrolysis caused by mechanical energy (Kivelä et al., 2012).
Varying results have been published regarding extrusion, another important industrial
processing technique that involves the simultaneous application of high temperature, pressure
and sheer force. Some studies have shown that harsh extrusion conditions can cause a severe
depolymerisation of β-glucan (Åman et al., 2004; Tosh et al., 2010), while others have
demonstrated that extrusion at conditions commonly used for commercial products can lead to
a higher Mw and viscosity, which result in improved physiological effects of β-glucan (Zhang
et al., 2009; Tosh et al., 2010; Zhang et al., 2011).

130
All the aforementioned studies have shown that in standard food processes dramatic changes in
β-glucan structure and functionality can take place, ranging from a reduction of viscosity and
Mw to the formation of new functional groups along the polymer chain. However, there is still
a lack of detailed information about β-glucan structure-function relationships, which is a major
limitation for effectively utilizing the full potential of β-glucan in functional foods. The aim of
this study was to explore the mechanisms of oat and barley β-glucan degradation following
thermal treatment and extrusion. In particular, an in depth analysis of the oxidation products by
means of UPLC-MS was carried out.
2. Materials
Oat bran concentrate (OatWell® 28) was obtained from DSM (Kaiseraugst, Switzerland) and
barley flour (Beta®Barley dehulled and untreated) was supplied by Dieckmann Cereals GmbH
(Rinteln, Germany). Calcium chloride, hydrochloric acid, sodium acetate, sodium nitrate,
sodium phosphate mono- and di-basic (NaH2CO3 and Na2HCO3, respectively), sodium azide,
methanol (LC-MS grade), 2-propanol, acetonitrile (LC-MS grade), 25% aqueous ammonia and
formic acid (both LC-MS grade), 2-aminobenzamide (2-AB), 2-methylpyridine borane
complex 95% (2-picoline borane), β-gentiobiose (6-O-β-D-glucopyranosyl-D-glucose), α-
amylase (from Bacillus licheniformis, Termamyl 300®L), xylanase (2500 U/g), pancreatin
from porcine pancreas and pepsin from porcine gastric mucosa (≥250 units/mg solid) were
purchased from Sigma-Aldrich, Switzerland. Lichenase (from Bacillus subtilis; EC 3.2.1.73;
GH family 16) and β-glucosidase (from Aspergillus niger; EC 3.2.1.21; GH family 3) were
supplied by Megazyme (Ireland).
Ethanol absolute was supplied by Alcosuisse, Switzerland. Acetic acid glacial and sodium
hydroxide pellets were purchased from Fisher Chemical, United Kingdom. Ultrapure (Milli-Q)
water was used for all of the experiments (Merck Millipore, Merck KGaA, Darmstadt,
Germany).
3. Methods
6.1 β-glucan extraction
The extraction of β-glucan from oat and barley flour (native or extruded) was performed
according to the methods by Rieder et al. (2015) and Lazaridou et al. (2004) with some
adaptations. Oat or barley flour (25 g and 50 g, respectively) was suspended in 0.5 L 70% v/v

131
ethanol and incubated at 80°C for 2 h to extract low-molar mass ethanol soluble carbohydrates
and to inactivate endogenous enzymes. The mixture was cooled to RT and filtered through a
sintered filter, the filtrate was then discarded and the residue re-suspended in 1 L Milli-Q water
and stirred overnight at RT. Next, 2.5 mL Termamyl (≥300 U/g) and 50 mg CaCl2 were added
and the mixture was incubated for 1 h at 96°C to hydrolyze starch. The mixture was then cooled
to RT and, after centrifugation (Centrifuge 5810 R, Eppendorf, Germany) (4000 rpm, 15 min,
4 °C), the supernatant was collected, 10 mL sodium acetate buffer (2.5 M, pH 4.5) was added
and the pH was adjusted to 4.5 with 1 M NaOH. Subsequently, in order to degrade co-extracted
arabinoxylans, 10 mg xylanase (2500 U/g) was added and the solution was incubated for 3 h at
40°C. The pH was then adjusted to 7 with 1 M NaOH and, following the addition of 50 mg
pancreatin, the solution was incubated at 45 min for 3 h to digest proteins. To inactivate the
enzymes, the solution was heated to 80°C for 45 min. After cooling to RT, the mixture was then
centrifuged to remove the debris (4000 rpm, 15 min, 4 °C), the supernatant was collected and
β-glucan was precipitated with two volumes of ethanol overnight at 4°C. After filtration using
a sintered filter, β-glucan was collected, frozen, freeze dried (Lyolab BII, LSL Secfroid,
Switzerland) for 48 h and milled with a ball mill (Pulverisette 23, Fritsch, Germany).
6.2 Characterization of the extracts
6.2.1 Total β-glucan content
The total β-glucan content of the oat and barley β-glucan extracts was determined with the
β-glucan Assay Kit (Mixed Linkage) from Megazyme, Ireland.
6.2.2 Protein content
To evaluate the protein content of the β-glucan extracts from oat and barley, samples were dried
for 24 hours at 110 °C and sent to the service for microelemental analyses at ETH
(Elementaranalysen LOC, ETH Zurich). The nitrogen (N) content was determined in triplicates
by a TruSpec Micro, composed of a combination of flow-through carrier gas and individual,
highly selective infrared (IR) and thermal conductivity detectors (TruSpec Micro, LECO
Corporation, USA). The approximate protein content was calculated using the conversion
factors determined by Mossé (1990) of 5.36 for the oat extracts and 5.50 for the barley extracts.

132
6.2.3 Phytic acid and iron contents
Phosphorus (P) and iron (Fe) content were determined by ICP-MS. First of all, 0.5 ml nitric
acid 65% (sub-boiled, in house production) was added to 0.5 ml sample (0.6% oat and barley
β-glucan solution, adjusted for the extract purity), then the samples were digested overnight at
80°C in an oven (Binder, Germany). The mineralised samples were filled up to 10 ml with
MilliQ water. For the analysis of P, the digested samples were further diluted ten times. The
determination of Fe and P was done on an iCAP RQ ICP-MS (Thermo Fisher Scientific, USA),
equipped with a cyclonic spray chamber and a SC-2 DX FAST autosampler (Elemental
Scientific, USA). Measurements (56Fe and 31P) were done in KED (kinetic energy
discrimination) mode, with He as a collision gas. Calibration ranges were 0-500 ng/ml for Fe,
and 0-1000 ng/ml for P. Calibrators were prepared from 100 µg/ml single element standard
solutions (Inorganic Ventures, USA). Indium (20 ng/ml) was used as internal standard.
Instrumental limits of detection were 0.02 ng/ml for Fe, and 1.4 ng/ml for P.
The phytic acid content was determined as follows:
%𝑃ℎ𝑦𝑡𝑖𝑐𝑎𝑐𝑖𝑑(𝑑𝑤𝑏) = 𝑃𝑐𝑜𝑛𝑡𝑒𝑛𝑡(𝑔/100𝑔)
0.282 ∙100%
were 0.282 is the factor used to convert the measured phosphorus content to phytic acid content
since phytic acid comprises 28.2% phosphorus (McKie and McCleary, 2016).
6.3 Treatments
Oat and barley extracts were dissolved overnight in water in a shaking water bath at 70°C to
prepare 0.6% β-glucan solutions, adjusted for the purity of each extract.
High temperature (HT):
15 ml of oat or barley extract solution was incubated in triplicates at 125°C or 175°C in the
oven for 2 h in triplicates in 25 mL PTFE lined hydrothermal reactors (Great Wall Instruments,
China).

133
High pressure (HP):
High pressure treatment was performed in a industrial-scale, high pressure device (Wave 55,
N.C Hyperbaric, Burgos, Spain). Samples were triple packed in PE bags and water was used as
the pressure transmitting medium. Treatment was performed at 600 MPa for a holding time of
5 min, not including the pressure build-up and release time. The treatment was repeated 6 times
and the temperature in the high-pressure chamber was kept at 10°C. After the high-pressure
treatment, the viscosity of the samples was immediately measured.
Extrusion:
Oat and barley flours were adjusted at 25% moisture content and extruded in a Brabender
Do-Corder C3 twin screw extruder, at a feed rate of 1 kg/h and with a diameter of 4 mm at
120°C or 150°C. Extruded samples were dried at 70°C for 6 hours then homogenized with a
ball mill.
Figure 1. Schematic representation of sample preparation and analysis.
6.4 Molecular weight determination
The weight average molecular weight (Mw) of the oat and barley β-glucan extracts, native and
processed, was determined in triplicates using size exclusion chromatography (SEC). Two
different systems were used for analysis and in both cases the extracts were dissolved in eluent

134
(0.1 M sodium nitrate and 0.02% (w/w) sodium azide) to a final concentration of 0.1% (w/v),
then filtered (0.45 μm) in HPLC vials.
The first system, more suitable for molecular weights up to 1’000 kDa, was an OMNISEC
Resolve (pump, degasser, autosampler and column oven) coupled with an OMNISEC Reveal
(integrated multi-detector module equipped with refractive index, UV/Vis absorbance, light
scattering and intrinsic viscosity detectors), both by Malvern (Malvern Instruments, U.K.). The
column system consisted of a pre-column (Viscotek AGuard Col. 50 x 6.0 mm, Malvern
Instruments Ltd, UK) connected to two A6000 columns in series (Viscotek, 300 x 8 mm,
Malvern Instruments Ltd, UK). The temperature of the column compartment was set at 30°C,
the flow rate was 0.7 mL/min and the injection volume 50 μL. The standard used for calibration
were PEO (polyethyleneoxide, Mw = 24’063 g/mol) and dextran (Mw = 70’026 g/mol). To
calculate the Mw of each sample the OMNISEC software (version 5.1, Malvern Instruments
Ltd, UK) was used.
The second system, used for samples with higher Mw (above 1’000 kDa), consisted of a HPLC
equipped with a binary pump, degasser, thermostated column compartment and auto sampler,
all from HP (Series 1100, Hewlett Packard, USA). The column system was comprised of a pre-
column (Viscotek AGuard Col. 50 x 6.0 mm, Malvern Instruments Ltd, UK) linked to a A5000
column (Viscotek, 300 x 7.8 mm, Malvern Instruments Ltd, UK) and a suprema 30000 column
(10 μm, 8 x 300 mm, PSS Polymer Standards Service GmbH, Germany). The column
compartment was kept at 35°C, the flow rate was set at 1 mL/min and the injection volume was
50 μL. Samples were analyzed with a refractive index (RI) detector (Series 1200, Agilent
Technologies, AG, Switzerland). For the calibration curve, solutions of β-glucan standards
(0.1% (w/v)) with reported Mp of 33’600, 67’100, 187’100, 247’000, 375’000, and 667’000
g/mol (Megazyme, Ireland) were prepared in eluent and filtered (0.45 μm) in HPLC vials. The
average Mw were calculated based on the measured standard curve with the ChemStation
software (ChemStation for LC 3D systems) and the add-on Cirrus GPC/SEC software (version
3.4.1) from Agilent
6.5 Rheology measurements
The viscosities of the native and treated samples (0.6% β-glucan, accounting for the purity of
each extract) were measured using an AR-2000 rheometer (TA Instruments, New Castle, DE).
A cone and plate geometry was used with a plate radius of 40 mm and a cone angle of 2°. The

135
gap between the cone and plate geometry was set at 59 µm. The flow curves were obtained over
a shear rate range of 20–2000 s-1, at 20°C.
6.6 Labelling by reductive amination
In order to investigate the oxidative changes in the β-glucan structure after thermal and high
pressure treatments, newly formed carbonyl groups and reducing ends were labelled with the
fluorescent 2-aminobenzamide (2-AB) by reductive amination according to the method by
Boulos and Nyström (2017), with some adaptations (Figure 2).
Labelling. First of all, 300 mg 0.6% native or treated oat or barley β-glucan solution
(adjusted for the purity of each extract) was accurately weighed in triplicates into a Falcon
tubes. Subsequently, acetic acid (285 μL), water (190 μL) and 2-AB (170 μL of a 0.8 M solution
in 50% methanol) were added, the tubes vortexed and then placed in a water bath at 40°C for
30 min. Afterwards, picoline borane (50 μL of a freshly prepared 0.4 M solution in methanol)
was added, the tubes vortexed and heated in a water bath at 40°C for 30 min. The samples where
then stored at RT in the dark overnight.
Removal of excess reagents. Two volumes of isopropanol followed by four volumes
of acetonitrile were added to each tube to precipitate the labelled β-glucan, the samples were
thoroughly mixed, then centrifuged (Centrifuge 5810 R, Eppendorf, Germany) (4000 rpm,
15 min, 4 °C). The supernatant was discarded, the pellets resuspended in 10 mL 95:5
ACN/H2O, mixed well and centrifuged again (4000 rpm, 15 min, 4 °C). The supernatant was
discarded and the process was repeated twice more with 10 mL methanol. After the last
centrifugation, the supernatant was discarded and the pellet suspended in 2 mL water.
Enzymatic treatment. First of all, 200 μL lichenase (20 U/mL in 20 mM phosphate
buffer) and 40 μL phosphate buffer (1M) were pipetted to each suspended pellet, then the
samples were incubated in a water bath at 37°C for 3 h under shaking. Afterwards, 50 μL
β-glucosidase (20 U/mL in 50 mM sodium acetate buffer) together with 250 μL sodium acetate
buffer (0.5 M) were added and the samples incubated overnight at RT. After the incubation
time, the enzymes were deactivated by heating the tubes at 90°C for 30 minutes in a waterbath.
Finally, 250 μL of internal standard were added to each sample before the next steps.
Solid phase extraction (SPE). The SPE cartridges (Supelclean ENVI-Carb 3mL, 0.25
g) were preconditioned with 3 mL ACN followed by 3 mL water. After loading the samples,
the cartridges were washed once with 3 mL water followed by two washings with 1.5 mL water.

136
After this, the samples were eluted with 2.5 mL 1:3 ACN/H2O + 0.1% formic acid in order to
collect fraction 1. Fraction 2 was collected by elution with 2.5 mL 1:1 ACN/H2O + 0.1% formic
acid. All tubes containing fraction 1 and fraction 2 were evaporated to dryness using a nitrogen
evaporator, then the samples were reconstituted with 200 μL and transferred to HPLC vials with
glass inserts and stored frozed at -20°C prior analysis.
Internal standard preparation. 3 ml 0.05mM gentiobiose solution were mixed with
acetic acid (2850 μL), H2O (1900 μL), and 2-AB (1700 μL of a 0.8 M solution in 50%
methanol), the tubes vortexed and then placed in a water bath at 40°C for 30 min. Afterwards,
picoline borane (500 μL of a freshly prepared 0.4 M solution in methanol) was added, the tubes
vortexed and heated in a water bath at 40°C for 30 min. The samples where then stored at RT
in the dark overnight. After the complete incubation period, the reaction mixture was diluted
with 5 mL H2O and run through a SPE cartridge (Supelclean ENVI-Carb 6 mL, 0.5 g)
preconditioned with 6 mL ACN followed by 6 mL water. After loading the samples, the
cartridges were washed once with 3 mL water followed by two washings with 2 mL water. In
this case, fraction 1 and 2 (namely 4.5 mL 1:3 ACN/H2O + 0.1% formic acid and 4.5 mL 1:1
ACN/H2O + 0.1% formic acid) were collected in the same 20mL volumetric flask. After this,
the samples were eluted with to 20 mL with water and the exact concentration of this purified
internal standard solution was measured by constructing a calibration curve with the unpurified
standard prepared before SPE.
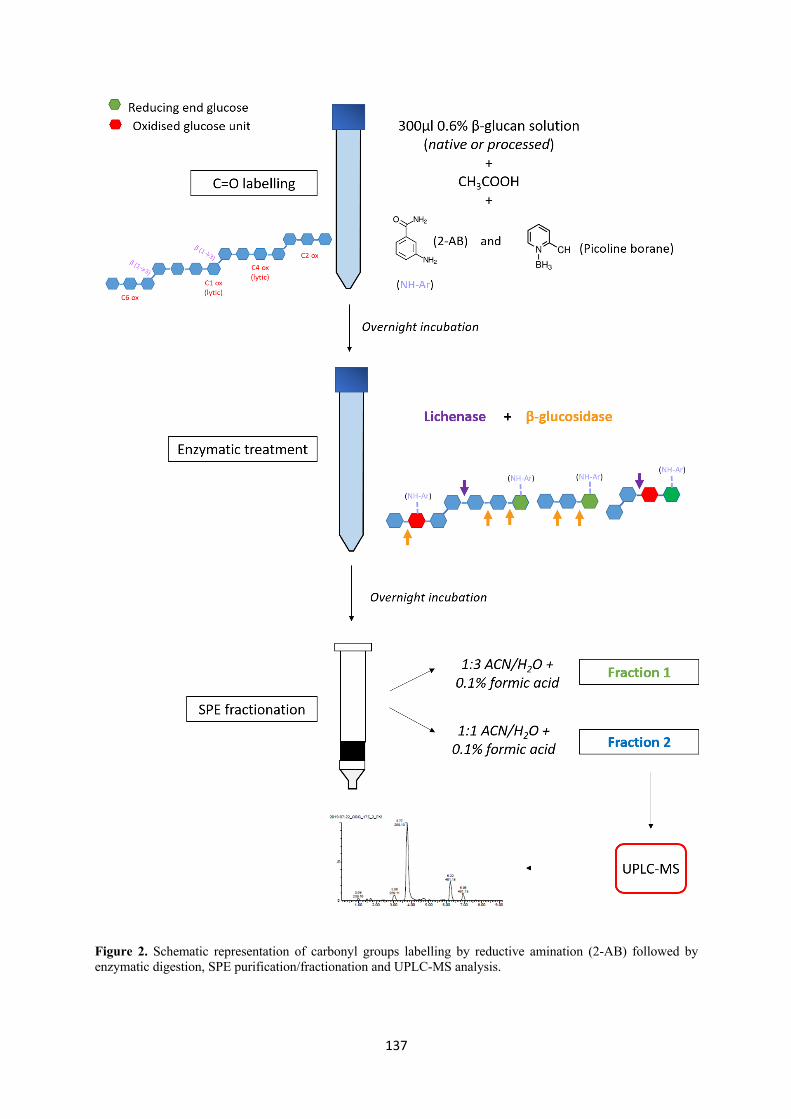
137
Figure 2. Schematic representation of carbonyl groups labelling by reductive amination (2-AB) followed by enzymatic digestion, SPE purification/fractionation and UPLC-MS analysis.

138
6.7 UPLC-MS analysis
The analysis of the labelled products (2.5 µl from each fraction injected) was performed with a
Waters Acquity UPLC system equipped with an Acquity UPLC BEH Amide column (2.1 ×
150mm, 1.7μm) and a VanGuard BEH Amide precolumn (5 mm) coupled to a Synapt G2 MS
system composed of an electrospray ionization (ESI) source and a quadrupole time-of-flight
(qToF) analyzer (Waters Corp., Milford, MA, USA). The MS was calibrated with a sodium
formate solution with leucine-enkephalin as the lock mass (m/z 554.2615), and run in negative
ion mode. The temperature of the column was kept at 35°C and the samples at 10°C. A 8:2 and
a 2:8 mixture of ACN/H2O were used as weak and strong needle wash, respectively. As eluents,
ACN with 0.1% ammonia and H2O with 0.1% ammonia were used, starting the gradient with
90% ACN and 10% H2O. The gradient increased linearly to 60% ACN and 40% H2O after 12
min, was kept constant for 2 min, then in 0.75 min the gradient returned to initial conditions,
followed by 2.25 min of re-equilibration. The flow rate was set to 0.34 ml/min.
The Synapt G2 was operated in the resolution mode. The voltages for the capillary, the sample
cone, and the extraction cone were set to 2500, 25, and 4 V, respectively, while the cone and
the desolvation gas flow rates were set to 20 l/h and 850 l/h. The temperature of the electrospray
desolvation heater was set to 350 °C and the source temperature was 120 °C. At a scan rate of
1 Hz, full scan mass spectra were recorded from m/z 50 to 1200 in centroid mode. The data
collected were analysed with MassLynx software, version 4.1 (Waters Corp., Milford, MA,
USA). The samples were also detected with a Fluorescent Detector (FLD), for which the
excitation and emission wavelengths were set to 360 nm and 424 nm, respectively. The peaks
of the Fluorescent Detector Analogue Signal (FLD signal) were integrated using Apex Track
Peak Integration.
6.8 Statistical analysis
Means and standard deviations (SD) were calculated with Microsoft® Excel 2016 (Microsoft
Cooperation, USA). The molecular weight, viscosity and FLD peak areas generated by UPLC-
MS analysis were compared using independent sample t-tests. For these tests, the Statistical
Package for Social Science (IBM SPSS 22, Chicago, USA) was used. The threshold for
indicating a significant difference was set at the probability level of p < 0.05.
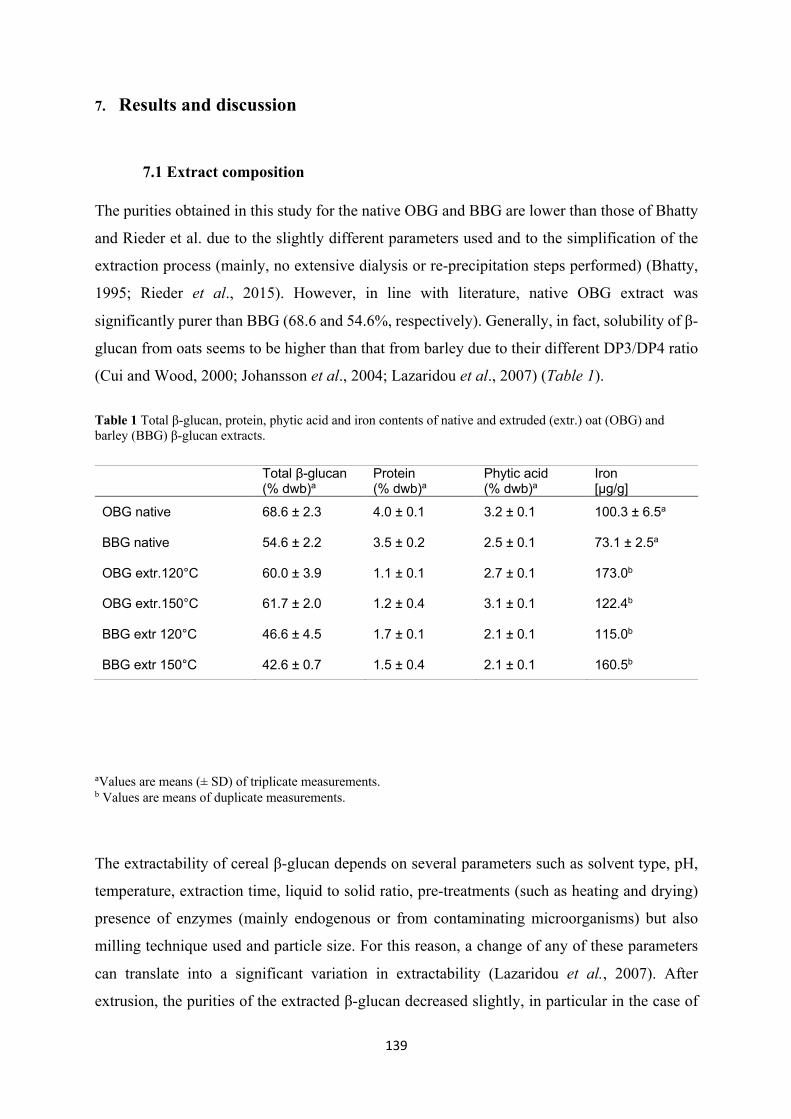
139
7. Results and discussion
7.1 Extract composition
The purities obtained in this study for the native OBG and BBG are lower than those of Bhatty
and Rieder et al. due to the slightly different parameters used and to the simplification of the
extraction process (mainly, no extensive dialysis or re-precipitation steps performed) (Bhatty,
1995; Rieder et al., 2015). However, in line with literature, native OBG extract was
significantly purer than BBG (68.6 and 54.6%, respectively). Generally, in fact, solubility of β-
glucan from oats seems to be higher than that from barley due to their different DP3/DP4 ratio
(Cui and Wood, 2000; Johansson et al., 2004; Lazaridou et al., 2007) (Table 1).
Table 1 Total β-glucan, protein, phytic acid and iron contents of native and extruded (extr.) oat (OBG) and barley (BBG) β-glucan extracts.
aValues are means (± SD) of triplicate measurements. b Values are means of duplicate measurements.
The extractability of cereal β-glucan depends on several parameters such as solvent type, pH,
temperature, extraction time, liquid to solid ratio, pre-treatments (such as heating and drying)
presence of enzymes (mainly endogenous or from contaminating microorganisms) but also
milling technique used and particle size. For this reason, a change of any of these parameters
can translate into a significant variation in extractability (Lazaridou et al., 2007). After
extrusion, the purities of the extracted β-glucan decreased slightly, in particular in the case of
Total β-glucan (% dwb)a
Protein (% dwb)a
Phytic acid (% dwb)a
Iron [μg/g]
OBG native 68.6 ± 2.3 4.0 ± 0.1 3.2 ± 0.1 100.3 ± 6.5a
BBG native 54.6 ± 2.2 3.5 ± 0.2 2.5 ± 0.1 73.1 ± 2.5a
OBG extr.120°C 60.0 ± 3.9 1.1 ± 0.1 2.7 ± 0.1 173.0b
OBG extr.150°C 61.7 ± 2.0 1.2 ± 0.4 3.1 ± 0.1 122.4b
BBG extr 120°C 46.6 ± 4.5 1.7 ± 0.1 2.1 ± 0.1 115.0b
BBG extr 150°C 42.6 ± 0.7 1.5 ± 0.4 2.1 ± 0.1 160.5b

140
barley, which might be explained with the higher particle size of the flours after extrusion and
milling. The milling process of the extruded samples, in fact, proved to be quite difficult,
particularly for barley, and a particle size as small as the original flours could not be obtained.
It has been shown that the larger the particle size the lower the extractability, which explains
these results (Grundy et al., 2017; Johansson et al., 2018).
In the present study, the iron content of the extracts was higher for oat (100.3 ± 6.5 μg/g) than
barley (73.1 ± 2.5 μg/g), which reflects the literature values of oat and barley flour (USDA,
2019). Interestingly, after extrusion, an increase of iron content in the extracts was recorded.
Iron, phytic acid and protein contents are crucial for the oxidative degradation under
examination here. The Fenton reaction, in fact, which has been shown to be responsible for the
β-glucan degradation during thermal treatments (Kivelä et al., 2011, 2012), is catalysed by iron,
which is present in significant amounts in cereals such as oat and barley (Ragaee et.al, 2006).
Hydroxyl radicals have very short half-life (~1 ns), which means they are only able to attack
the molecules in their vicinity (Pryor, 1986). The localization of the metals, therefore, is one of
the key factors in the Fenton oxidation of macromolecules. Since β-glucan is often found in
association with metals (Platt and Clydesdale, 1984), this polysaccharide is a specific target for
hydroxyl radical induced degradation, as the short living radicals are produced in immediate
vicinity of β-glucan molecules (Chevion, 1988). It has been shown that the formation of
complexes with chelating agents (e.g. phytic acid) and proteins heightens the reduction power
of iron (Welch et al., 2002), therefore these co-extracted compounds can contribute to an
enhanced oxidative degradation of β-glucan (Kivelä et al., 2012).
The phytic acid content was higher in the oat than in the barley β-glucan extract (3.2 and 2.5%,
respectively), which is in line with the used sources of whole grain flour for barley and bran
flour for oat, where phytic acid is primarily located (Haros et al., 2001; Schlemmer et al., 2009).
Phytic acid in the extracts was three to four times higher compared to the literature values for
oat or barley flour (Schlemmer et al., 2009). After extrusion, a slight reduction in phytic acid
content was recorded.
The protein content of the barley β-glucan extract (3.5%) was slightly lower than that of the oat
extract (4%), which is in line with the literature (Bhatty, 1995; Kahlon and Woodruff, 2003;
Lazaridou et al., 2007). As for the samples extracted after extrusion, the nitrogen content was
very close to the limit of detection of the instrument, therefore the results cannot be considered
completely reliable. However, a significant decrease in the protein amount was recorded after
extrusion, which is in line with literature. It has been shown, in fact, that extrusion can cause
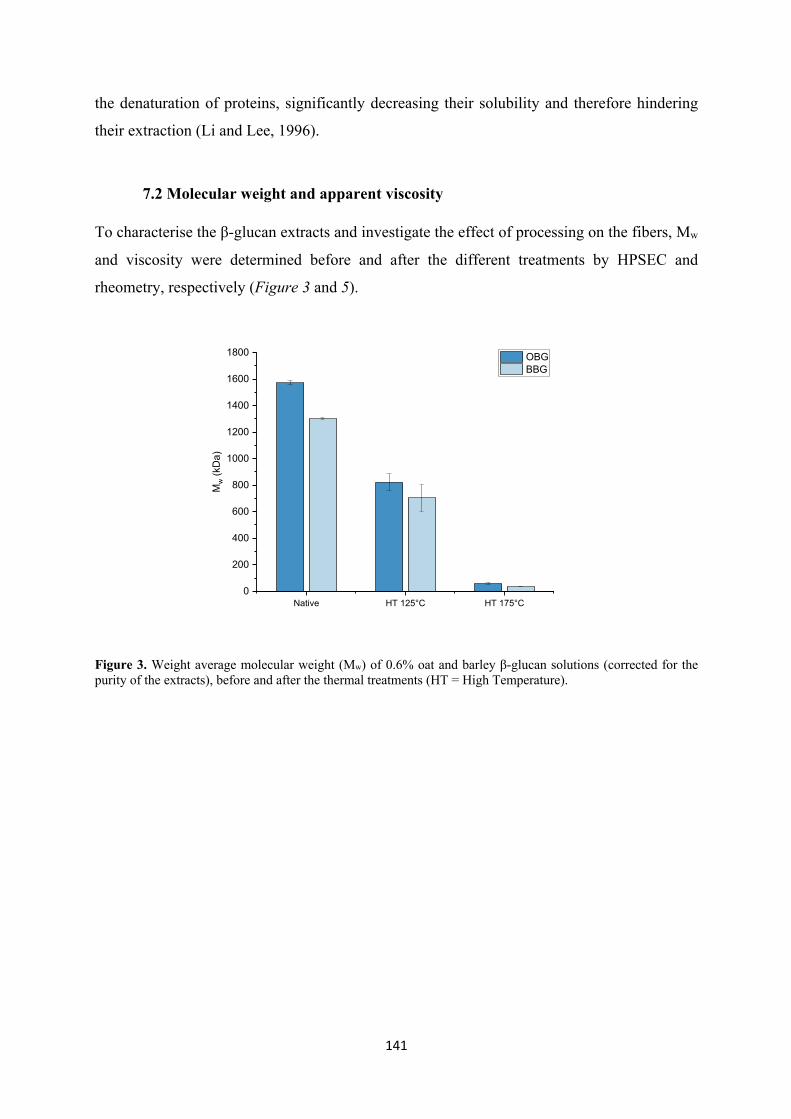
141
the denaturation of proteins, significantly decreasing their solubility and therefore hindering
their extraction (Li and Lee, 1996).
7.2 Molecular weight and apparent viscosity
To characterise the β-glucan extracts and investigate the effect of processing on the fibers, Mw
and viscosity were determined before and after the different treatments by HPSEC and
rheometry, respectively (Figure 3 and 5).
Figure 3. Weight average molecular weight (Mw) of 0.6% oat and barley β-glucan solutions (corrected for the purity of the extracts), before and after the thermal treatments (HT = High Temperature).
Native HT 125°C HT 175°C0
200
400
600
800
1000
1200
1400
1600
1800
Mw (k
Da)
OBG BBG
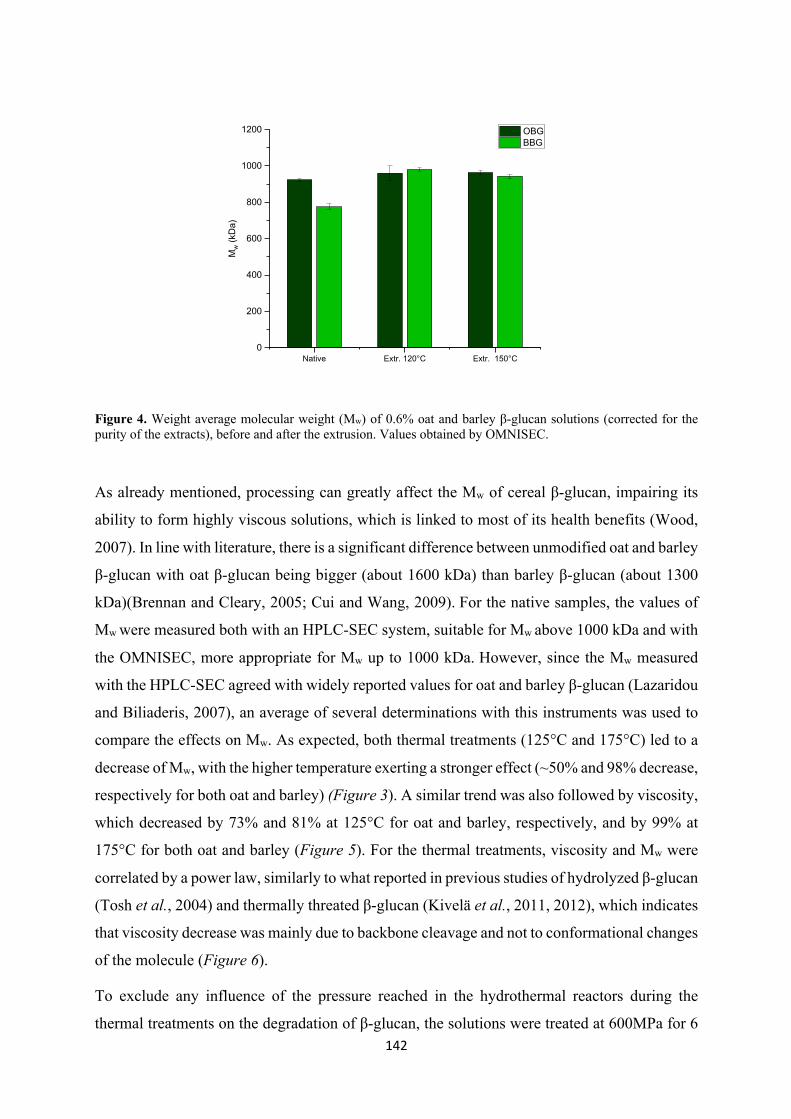
142
Figure 4. Weight average molecular weight (Mw) of 0.6% oat and barley β-glucan solutions (corrected for the purity of the extracts), before and after the extrusion. Values obtained by OMNISEC.
As already mentioned, processing can greatly affect the Mw of cereal β-glucan, impairing its
ability to form highly viscous solutions, which is linked to most of its health benefits (Wood,
2007). In line with literature, there is a significant difference between unmodified oat and barley
β-glucan with oat β-glucan being bigger (about 1600 kDa) than barley β-glucan (about 1300
kDa)(Brennan and Cleary, 2005; Cui and Wang, 2009). For the native samples, the values of
Mw were measured both with an HPLC-SEC system, suitable for Mw above 1000 kDa and with
the OMNISEC, more appropriate for Mw up to 1000 kDa. However, since the Mw measured
with the HPLC-SEC agreed with widely reported values for oat and barley β-glucan (Lazaridou
and Biliaderis, 2007), an average of several determinations with this instruments was used to
compare the effects on Mw. As expected, both thermal treatments (125°C and 175°C) led to a
decrease of Mw, with the higher temperature exerting a stronger effect (~50% and 98% decrease,
respectively for both oat and barley) (Figure 3). A similar trend was also followed by viscosity,
which decreased by 73% and 81% at 125°C for oat and barley, respectively, and by 99% at
175°C for both oat and barley (Figure 5). For the thermal treatments, viscosity and Mw were
correlated by a power law, similarly to what reported in previous studies of hydrolyzed β-glucan
(Tosh et al., 2004) and thermally threated β-glucan (Kivelä et al., 2011, 2012), which indicates
that viscosity decrease was mainly due to backbone cleavage and not to conformational changes
of the molecule (Figure 6).
To exclude any influence of the pressure reached in the hydrothermal reactors during the
thermal treatments on the degradation of β-glucan, the solutions were treated at 600MPa for 6
Native Extr. 120°C Extr. 150°C0
200
400
600
800
1000
1200
Mw (k
Da)
OBG BBG
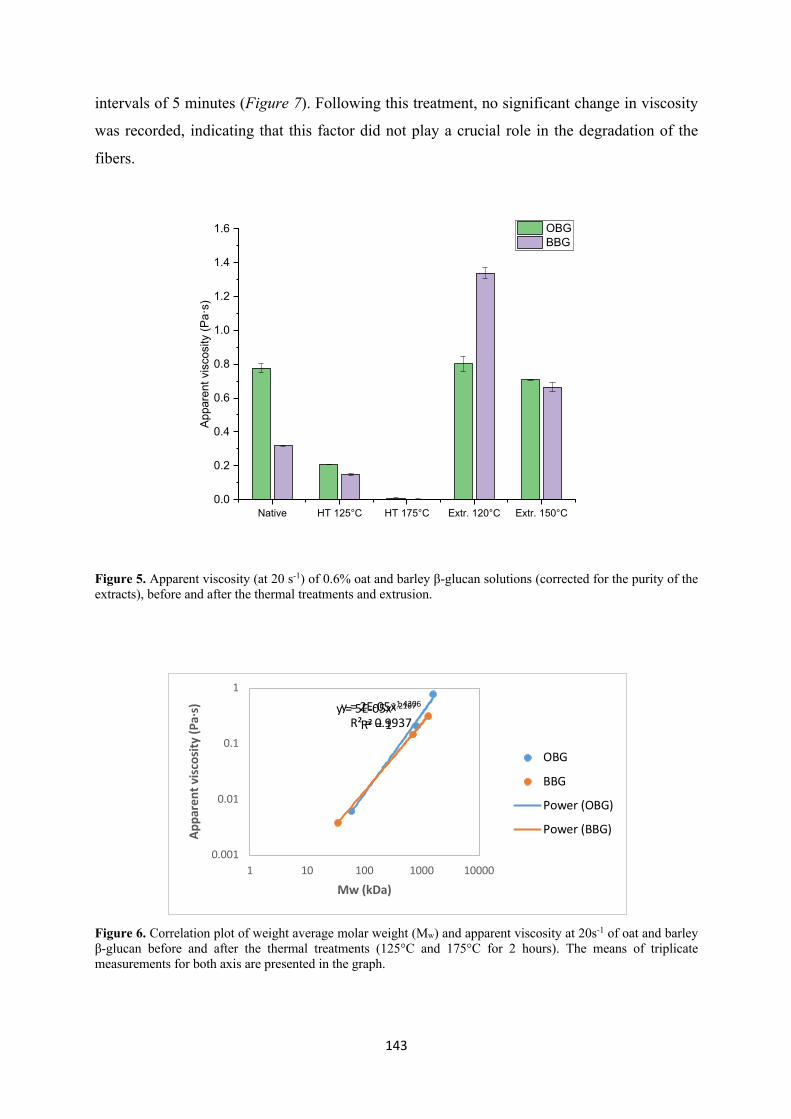
143
intervals of 5 minutes (Figure 7). Following this treatment, no significant change in viscosity
was recorded, indicating that this factor did not play a crucial role in the degradation of the
fibers.
Figure 5. Apparent viscosity (at 20 s-1) of 0.6% oat and barley β-glucan solutions (corrected for the purity of the extracts), before and after the thermal treatments and extrusion.
Figure 6. Correlation plot of weight average molar weight (Mw) and apparent viscosity at 20s-1 of oat and barley β-glucan before and after the thermal treatments (125°C and 175°C for 2 hours). The means of triplicate measurements for both axis are presented in the graph.
Native HT 125°C HT 175°C Extr. 120°C Extr. 150°C0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
Appa
rent
vis
cosi
ty (P
a·s)
OBG BBG
y = 2E-05x1.4306
R² = 0.9937y = 5E-05x1.2167
R² = 1
0.001
0.01
0.1
1
1 10 100 1000 10000
Appa
rent
vis
cosi
ty (P
a·s)
Mw (kDa)
OBG
BBG
Power (OBG)
Power (BBG)
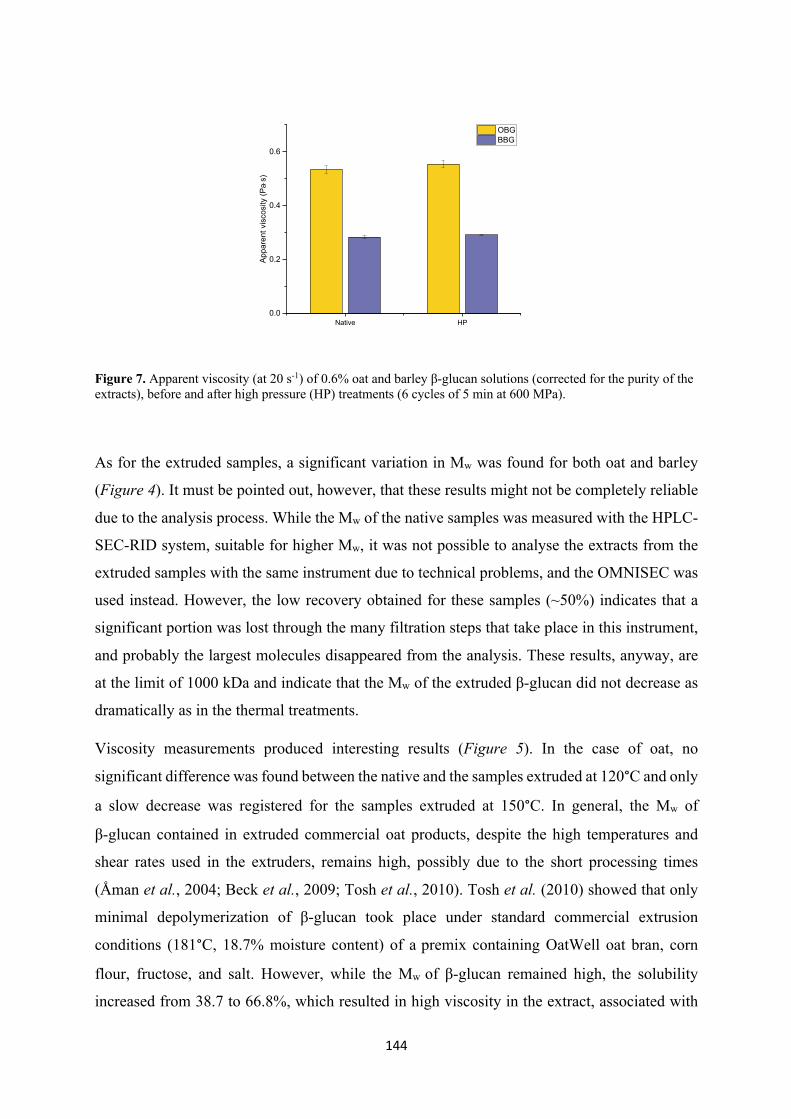
144
Figure 7. Apparent viscosity (at 20 s-1) of 0.6% oat and barley β-glucan solutions (corrected for the purity of the extracts), before and after high pressure (HP) treatments (6 cycles of 5 min at 600 MPa).
As for the extruded samples, a significant variation in Mw was found for both oat and barley
(Figure 4). It must be pointed out, however, that these results might not be completely reliable
due to the analysis process. While the Mw of the native samples was measured with the HPLC-
SEC-RID system, suitable for higher Mw, it was not possible to analyse the extracts from the
extruded samples with the same instrument due to technical problems, and the OMNISEC was
used instead. However, the low recovery obtained for these samples (~50%) indicates that a
significant portion was lost through the many filtration steps that take place in this instrument,
and probably the largest molecules disappeared from the analysis. These results, anyway, are
at the limit of 1000 kDa and indicate that the Mw of the extruded β-glucan did not decrease as
dramatically as in the thermal treatments.
Viscosity measurements produced interesting results (Figure 5). In the case of oat, no
significant difference was found between the native and the samples extruded at 120°C and only
a slow decrease was registered for the samples extruded at 150°C. In general, the Mw of
β-glucan contained in extruded commercial oat products, despite the high temperatures and
shear rates used in the extruders, remains high, possibly due to the short processing times
(Åman et al., 2004; Beck et al., 2009; Tosh et al., 2010). Tosh et al. (2010) showed that only
minimal depolymerization of β-glucan took place under standard commercial extrusion
conditions (181°C, 18.7% moisture content) of a premix containing OatWell oat bran, corn
flour, fructose, and salt. However, while the Mw of β-glucan remained high, the solubility
increased from 38.7 to 66.8%, which resulted in high viscosity in the extract, associated with
Native HP0.0
0.2
0.4
0.6
Appa
rent
vis
cosi
ty (P
a×s)
OBG BBG

145
high physiological effectiveness. On the other hand, increasing the cooking temperature and
reducing the water content of the mixture, resulted in a harsher depolymerization of the fiber.
Further increasing the temperature and standard mechanical energy dramatically enhanced the
β-glucan solubility at physiological conditions from 66.8% to 100%. This, however, took place
along a reduction of viscosity of the extracts, due to the simultaneous decrease in β-glucan Mw
(Tosh et al., 2010). In the present study, no dramatic change was noticed in the viscosity (and,
possibly, Mw) of the extruded oat flours, probably due to the high moisture contents and mild
cooking temperatures. In the case of barley, however, a significant increase in viscosity was
obtained with the extrusion at 150°C, and an even more marked effect with the extrusion at
120°C. This was also observed by Zhang et al. (2011), who noticed an intense increase in the
viscosity of solutions of soluble dietary fiber extracted from extruded oat bran compared to
those from the native bran.
7.3 UPLC-MS analysis of native and processed extracts.
The oxidation products generated during thermal treatments and, possibly, extrusion of cereal
β-glucan were examined by UPLC-MS, in order to further clarify the degradation mechanisms
of this fiber.
The investigation of the oxidation products of polysaccharides by MS techniques is hindered
by several difficulties, such as the high Mw of these macromolecules, the low concentration of
oxidized units and the complexity of the products in terms of position and type of oxidation
(Boulos and Nyström, 2016). Several strategies have been used to solve these difficulties, such
as the use of low Mw model oligosaccharides, enzymatic treatment to produce selectively
hydrolysed parts of the polysaccharide or functional group labelling for higher MS sensitivity
(Lamari et al., 2003). Carbonyl and reducing end labelling by reductive amination with 2-AB
followed by enzymatic treatment, SPE purification/fractionation and UPLC-MS analysis, has
been shown to allow the selective investigation of β-glucan oxidation products formed during
mild oxidation conditions, such as those caused by Fenton reaction in foodstuff (Boulos et al.,
2017).
For laboratory scale applications, sodium cyanoborohydride (NaBH3CN) is the most commonly
used reagent for reductive amination, since it is highly selective, soluble in many solvents, and
stable in acidic mediums (Lane, 1975). The major drawback of NaBH3CN, however, is that

146
upon hydrolysis it generates the highly toxic and volatile hydrogen cyanide (HCN). Ruhaak et
al. (2010) demonstrated that 2-picoline-borane can be applied as an equally efficient but non-
toxic alternative to NaBH3CN in labelling of oligosaccharides with 2-aminobenzamide (2-AB)
by reductive amination, therefore this reagent was selected for our study.
Boulos & Nyström (2017) investigated commercial β-glucan oxidation products by UPLC-
MS/MS after four sample preparation strategies: SPE, enzymatic digestion though lichenase
and β-glucosidase and two functional group labelling techniques. They were able to detect all
lytic (OH attack on C1, C3/C4, C5) and non-lytic (OH attack on C2, C4/C3, C6) oxidation
products with a combination of the four sample preparation strategies (Figure 8). In particular,
they were able to demonstrate the occurrence of 5-oxo-glucose as a direct result of lytic C5-
oxidation under Fenton conditions for the first time, which seemed to represent a clear marker
of β-glucan oxidation. On top of this, neutral reducing ends were also detected as labeled
glucose and cross-ring cleaved units (arabinose and erythrose). Determination of the reducing
end concentration is particularly useful to follow the hydrolysis of polysaccharides, since this
concentration increases simultaneously to the depolymerization of the molecule.
Figure 8. Possible position of attack by hydroxyl radical (•OH) on D-glucose, shown in the equilibrium between ring and open form. Attack at at C2-C6 generate oxo-glucose, while attack at C1 generates gluconic acid.
Several of these products were detected in the current study, in particular for the oat and barley
solutions treated at 175°C.
7.3.1 Main products in Fraction 2
The main signals for all the β-glucan samples (native and treated) were detected in fraction 2
(Figure 9) namely labelled Glc (Glc-2AB) and labelled GlcGlc (GlcGlc-2AB), with m/z of
OOH
OHHO
HO
OH OH
OH OH
OH
OH
O
123
46 5
1
2
3
4
5
6
D-glucose
ring open form

147
299.12 and 461.18, respectively. GlcGlc consisted of two peaks, the first one being intrinsic of
the sample and the second one being the internal standard used for analysis (labelled
gentiobiose). Also, a peak with m/z 623.23, that is exactly 162 Da mass (anhydrous glucose
unit) difference from m/z 461.18 (labelled GlcGlc), was found in the native and treated samples,
and this was recognized as labelled Glc3 (GlcGlcGlc-2AB), a product of incomplete hydrolysis
of β-glucan during sample preparation. Additionally, Glc4-2AB with m/z 785.28, Glc5-2AB
with m/z 947.34 and Glc6-2AB with m/z 1109.39 were found in some of the fractions. In
particular, Glc4-2AB was detected in the thermally threated BBG (at both 125°C and 175°C),
BBG extruded at 120°C and both OBG and BBG extruded at 150°C. Glc5-AB and Glc6-AB
were detected in the case of thermally threated BBG (at both 125°C and 175), and in quite
significant amounts in all OBG and BBG extruded samples.
Figure 9. Example of UPLC-MS base peak ion chromatograms (BPI) of samples of OBG Fraction 2: A) native, B) thermally treated at 125°C, C) thermally treated at 175°C, D) extruded at 120°C, E) extruded at 150°C.
Other relevant products found in fraction 2 were cross-ring cleavage products such as labelled
arabinose (Ara-AB) with m/z 269.11 and labelled erytrose (Ery-2AB) with m/z 239.1, but also
GlcAra-AB with m/z 431.17 and GlcEry-2AB with m/z 401.16. In particular, Ery-2AB and
A
B
C
D
E
148
GlcEry-2AB were only found in the 175°C threated samples, both from OBG and BBG, and
therefore represent a clear marker of β-glucan oxidation (Figure 10).
Figure 10. MS spectra of BBG samples treated at 175°, showing the m/z 401.16, corresponding to GlcEry-2AB and the m/z 239.10, corresponding to Ery-2AB
In theory, after labelling, enzymatic treatment and SPE purification and fractionation, only
labelled monosaccharides are expected to be generated from reducing end sugars, such as
labelled Glc, Ara, Ery and 5oxoGlc together with labelled oxo-Glcn oligosaccharides (non-
reducing end/mid-chain oxo-products). Boulos and Nyström (2017) also observed that the used
β-glucosidase was not able to hydrolyse β-(1→3)-linked modified reducing end units (i.e.
labelled), leading to Glc-2AB and GlcGlc-2AB as products of intact reducing ends. By means
of MS/MS analysis they were able to demonstrate that the labelled disaccharides, such as
GlcGlc-2AB, Glc5oxoGlc-2AB, and GlcAra-2AB exhibited a β-(1→3)-linkage. On the basis
of this, the authors hypothesized that probably β-glucosidase cannot cleave off the non-reducing
Glc-unit, if it is β-(1→3)-linked and linked to a labelled monosaccharide (Glc-2AB, Ara-2AB,
5oxoGlc-2AB). On the other hand, β-(1→4)-linkages were fully hydrolyzed irrespective of
labelling (with the exception of Glc-Ery-2AB, which can only be β-(1→4)-linked, and was also
sometimes detected).
7.3.2 Main products in Fraction 1
The main products found in fraction 2, in particular Glc-2AB, GlcGlc-2AB, Ara-2AB and
GlcAra-2AB were also found in fraction 1 of both native and processed samples, even if in
A
B

149
smaller amounts compared to fraction 2. Ery-2AB and GlcEry-2AB, on the other hand were
exclusive to fraction 2.
In fraction 1 of oxidized β-glucan, Boulos and Nyström (2017) could detect a peak with m/z
281.11, corresponding to a lytic C5 oxidation product, namely 5oxoGlc, a clear marker of
β-glucan oxidation, which could not be found in the present study. On the other hand, small
amounts of other labelled lytic non–reducing end/non lytic mid-chain oxo-Glcn, with n equal to
2, 3 and 4 (m/z 459.16, 621.21 and 783.27, respectively) were detected in the MS, even though
the mixture was very complex and the FLD signal did not allow their individual identification.
Despite this, these products can be considered clear markers for β-glucan oxidation.
Figure 11. Example of UPLC-MS base peak ion chromatograms (BPI) of samples of OBG Fraction 1: A) native, B) thermally treated at 125°C, C) thermally treated at 175°C, D) extruded at 120°C, E) extruded at 150°C.
A
B
C
D
E
150
Table 3 Main products found in fraction 1 and fraction 2 with corresponding m/z.
Product (monosaccharide) m/z where Product (disaccharide) m/z where
299.12 Fr. 2
461.16 Fr. 2
269.11 Fr. 2
431.17 Fr. 2
239.10 Fr. 2
401.16 Fr. 2
7.3.3 FLD peak area (relative to I.S.) comparison
For analysis and comparison of the products before and after the treatments, the FLD peak areas
(relative to I.S.) of the corresponding MS peaks in fraction 1 and 2 were summed up (Figure
12).
Comparing the FLD peak areas of the intact reducing end products Glc-2AB and GlcGlc-2AB
of the heat treated β-glucan samples with the native, a marked increase was found, in particular
for the 175°C treated samples, when the peak areas of both fractions were added. For OBG,
H
HN
H
Ar
OHOH
OH OHOH
Glc-2AB
H
HN
H
Ar
OOH
OH OHOH
(Glc)
GlcGlc-2AB
OHOH
OHOH
H
H
HN
Ara-2ABAr
OOH
OHOH
H
H
HN
GlcAra-2ABAr
(Glc)
H
HN
H
ArOH
OHOH
Ery-2AB
H
HN
H
ArOH
OOH
GlcEry-2AB
(Glc)
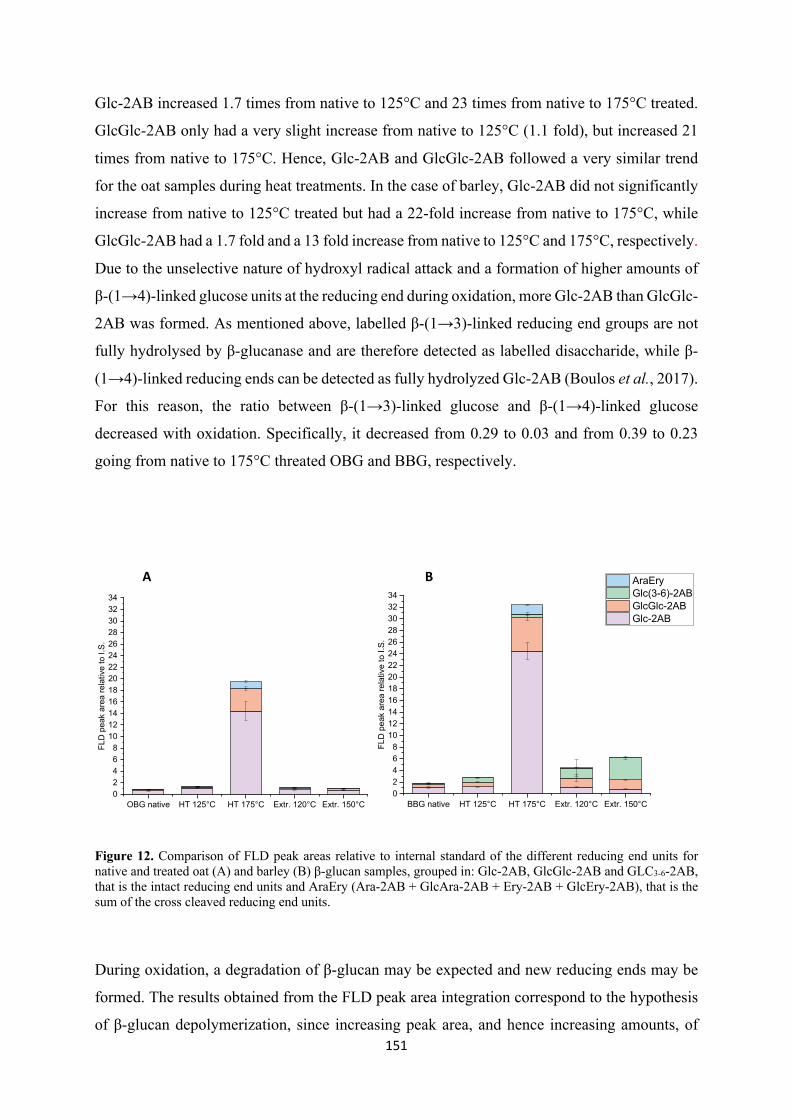
151
Glc-2AB increased 1.7 times from native to 125°C and 23 times from native to 175°C treated.
GlcGlc-2AB only had a very slight increase from native to 125°C (1.1 fold), but increased 21
times from native to 175°C. Hence, Glc-2AB and GlcGlc-2AB followed a very similar trend
for the oat samples during heat treatments. In the case of barley, Glc-2AB did not significantly
increase from native to 125°C treated but had a 22-fold increase from native to 175°C, while
GlcGlc-2AB had a 1.7 fold and a 13 fold increase from native to 125°C and 175°C, respectively.
Due to the unselective nature of hydroxyl radical attack and a formation of higher amounts of
β-(1→4)-linked glucose units at the reducing end during oxidation, more Glc-2AB than GlcGlc-
2AB was formed. As mentioned above, labelled β-(1→3)-linked reducing end groups are not
fully hydrolysed by β-glucanase and are therefore detected as labelled disaccharide, while β-
(1→4)-linked reducing ends can be detected as fully hydrolyzed Glc-2AB (Boulos et al., 2017).
For this reason, the ratio between β-(1→3)-linked glucose and β-(1→4)-linked glucose
decreased with oxidation. Specifically, it decreased from 0.29 to 0.03 and from 0.39 to 0.23
going from native to 175°C threated OBG and BBG, respectively.
Figure 12. Comparison of FLD peak areas relative to internal standard of the different reducing end units for native and treated oat (A) and barley (B) β-glucan samples, grouped in: Glc-2AB, GlcGlc-2AB and GLC3-6-2AB, that is the intact reducing end units and AraEry (Ara-2AB + GlcAra-2AB + Ery-2AB + GlcEry-2AB), that is the sum of the cross cleaved reducing end units.
During oxidation, a degradation of β-glucan may be expected and new reducing ends may be
formed. The results obtained from the FLD peak area integration correspond to the hypothesis
of β-glucan depolymerization, since increasing peak area, and hence increasing amounts, of
OBG native HT 125°C HT 175°C Extr. 120°C Extr. 150°C02468
10121416182022242628303234
FLD
pea
k ar
ea re
lativ
e to
I.S.
BBG native HT 125°C HT 175°C Extr. 120°C Extr. 150°C02468
10121416182022242628303234
FLD
pea
k ar
ea re
lativ
e to
I.S.
AraEry Glc(3-6)-2AB GlcGlc-2AB Glc-2AB
A B
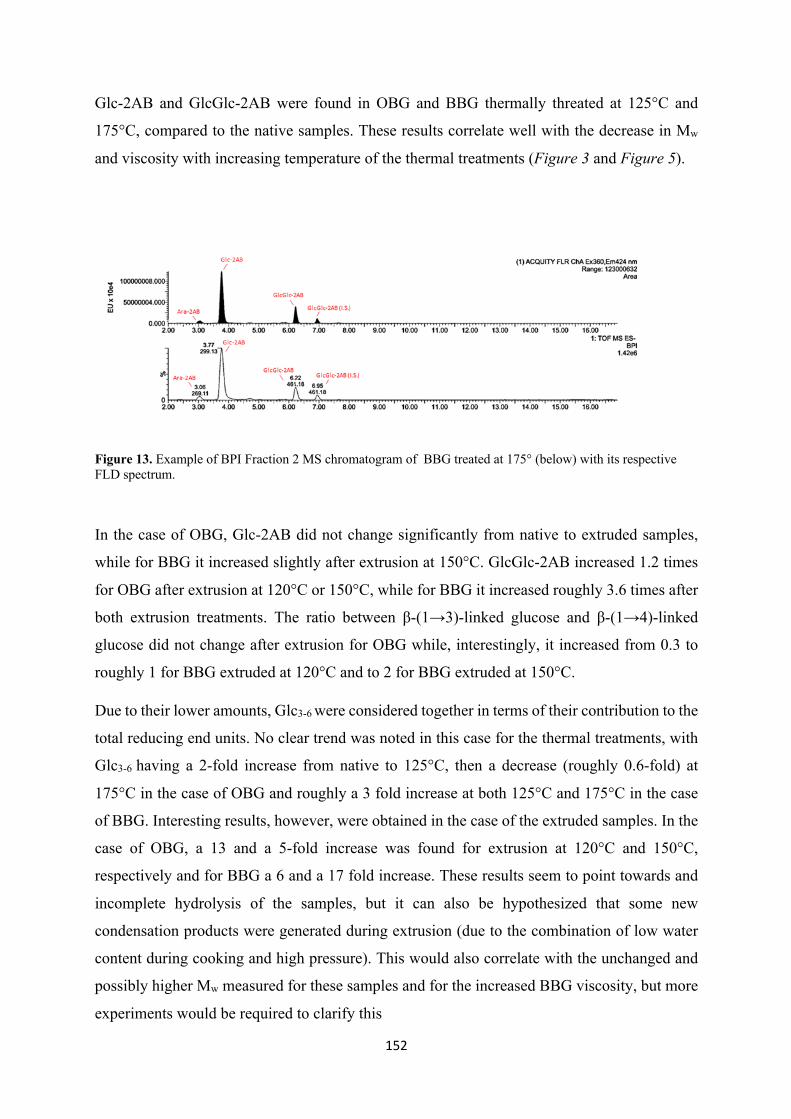
152
Glc-2AB and GlcGlc-2AB were found in OBG and BBG thermally threated at 125°C and
175°C, compared to the native samples. These results correlate well with the decrease in Mw
and viscosity with increasing temperature of the thermal treatments (Figure 3 and Figure 5).
Figure 13. Example of BPI Fraction 2 MS chromatogram of BBG treated at 175° (below) with its respective FLD spectrum.
In the case of OBG, Glc-2AB did not change significantly from native to extruded samples,
while for BBG it increased slightly after extrusion at 150°C. GlcGlc-2AB increased 1.2 times
for OBG after extrusion at 120°C or 150°C, while for BBG it increased roughly 3.6 times after
both extrusion treatments. The ratio between β-(1→3)-linked glucose and β-(1→4)-linked
glucose did not change after extrusion for OBG while, interestingly, it increased from 0.3 to
roughly 1 for BBG extruded at 120°C and to 2 for BBG extruded at 150°C.
Due to their lower amounts, Glc3-6 were considered together in terms of their contribution to the
total reducing end units. No clear trend was noted in this case for the thermal treatments, with
Glc3-6 having a 2-fold increase from native to 125°C, then a decrease (roughly 0.6-fold) at
175°C in the case of OBG and roughly a 3 fold increase at both 125°C and 175°C in the case
of BBG. Interesting results, however, were obtained in the case of the extruded samples. In the
case of OBG, a 13 and a 5-fold increase was found for extrusion at 120°C and 150°C,
respectively and for BBG a 6 and a 17 fold increase. These results seem to point towards and
incomplete hydrolysis of the samples, but it can also be hypothesized that some new
condensation products were generated during extrusion (due to the combination of low water
content during cooking and high pressure). This would also correlate with the unchanged and
possibly higher Mw measured for these samples and for the increased BBG viscosity, but more
experiments would be required to clarify this
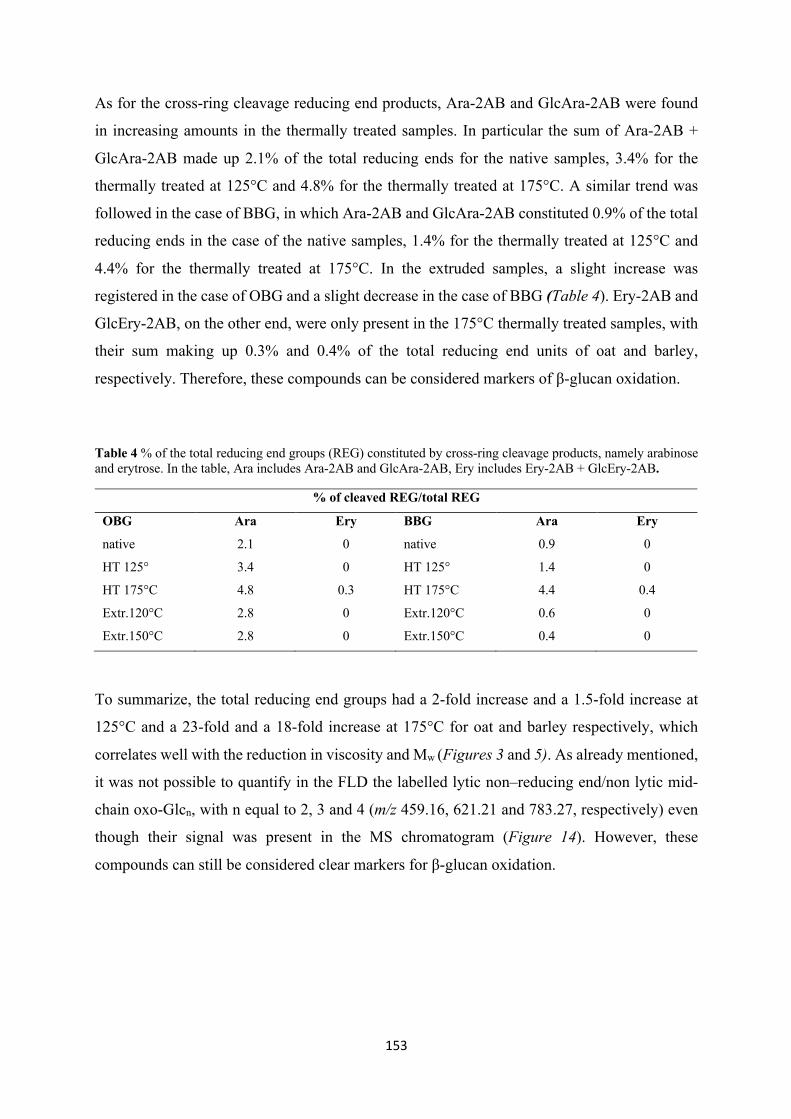
153
As for the cross-ring cleavage reducing end products, Ara-2AB and GlcAra-2AB were found
in increasing amounts in the thermally treated samples. In particular the sum of Ara-2AB +
GlcAra-2AB made up 2.1% of the total reducing ends for the native samples, 3.4% for the
thermally treated at 125°C and 4.8% for the thermally treated at 175°C. A similar trend was
followed in the case of BBG, in which Ara-2AB and GlcAra-2AB constituted 0.9% of the total
reducing ends in the case of the native samples, 1.4% for the thermally treated at 125°C and
4.4% for the thermally treated at 175°C. In the extruded samples, a slight increase was
registered in the case of OBG and a slight decrease in the case of BBG (Table 4). Ery-2AB and
GlcEry-2AB, on the other end, were only present in the 175°C thermally treated samples, with
their sum making up 0.3% and 0.4% of the total reducing end units of oat and barley,
respectively. Therefore, these compounds can be considered markers of β-glucan oxidation.
Table 4 % of the total reducing end groups (REG) constituted by cross-ring cleavage products, namely arabinose and erytrose. In the table, Ara includes Ara-2AB and GlcAra-2AB, Ery includes Ery-2AB + GlcEry-2AB.
% of cleaved REG/total REG
OBG Ara Ery BBG Ara Ery
native 2.1 0 native 0.9 0
HT 125° 3.4 0 HT 125° 1.4 0
HT 175°C 4.8 0.3 HT 175°C 4.4 0.4
Extr.120°C 2.8 0 Extr.120°C 0.6 0
Extr.150°C 2.8 0 Extr.150°C 0.4 0
To summarize, the total reducing end groups had a 2-fold increase and a 1.5-fold increase at
125°C and a 23-fold and a 18-fold increase at 175°C for oat and barley respectively, which
correlates well with the reduction in viscosity and Mw (Figures 3 and 5). As already mentioned,
it was not possible to quantify in the FLD the labelled lytic non–reducing end/non lytic mid-
chain oxo-Glcn, with n equal to 2, 3 and 4 (m/z 459.16, 621.21 and 783.27, respectively) even
though their signal was present in the MS chromatogram (Figure 14). However, these
compounds can still be considered clear markers for β-glucan oxidation.
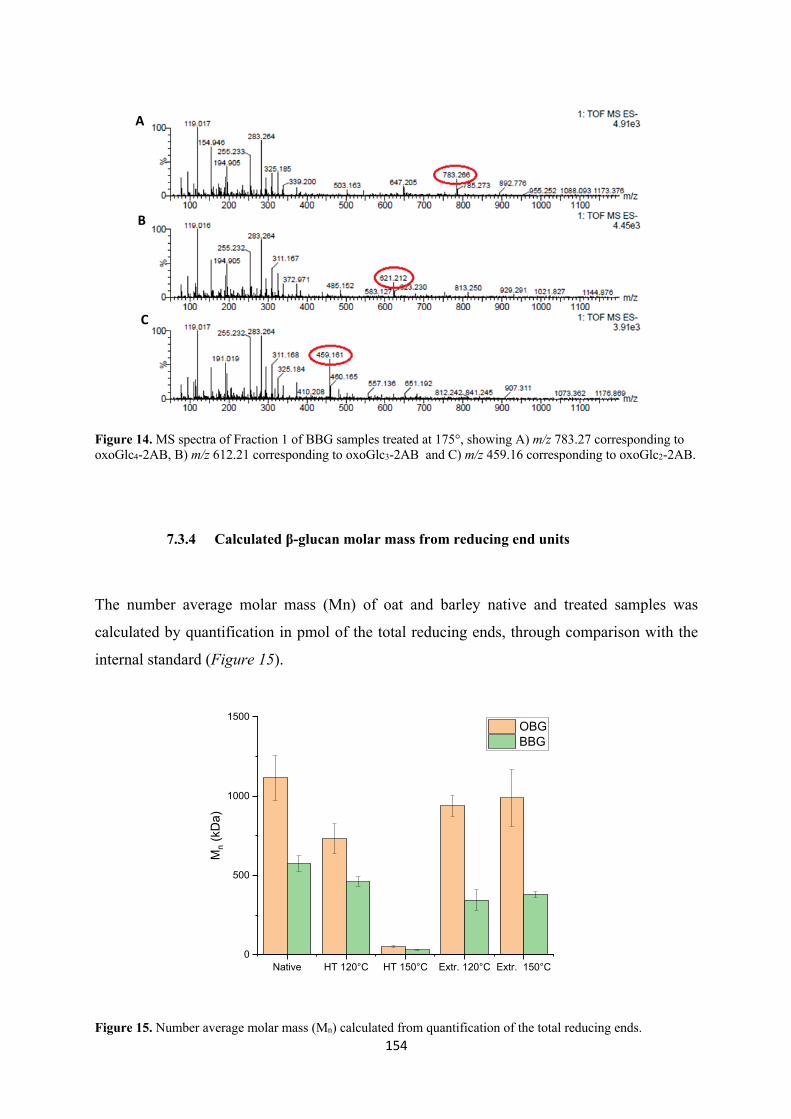
154
Figure 14. MS spectra of Fraction 1 of BBG samples treated at 175°, showing A) m/z 783.27 corresponding to oxoGlc4-2AB, B) m/z 612.21 corresponding to oxoGlc3-2AB and C) m/z 459.16 corresponding to oxoGlc2-2AB.
7.3.4 Calculated β-glucan molar mass from reducing end units
The number average molar mass (Mn) of oat and barley native and treated samples was
calculated by quantification in pmol of the total reducing ends, through comparison with the
internal standard (Figure 15).
Figure 15. Number average molar mass (Mn) calculated from quantification of the total reducing ends.
Native HT 120°C HT 150°C Extr. 120°C Extr. 150°C0
500
1000
1500
Mn (
kDa)
OBG BBG
A
B
C

155
Confirming the degradation following the thermal treatments, the calculated Mn of both oat and
barley β-glucan decreased at 125°C and, even more, at 175°, in agreement with the data shown
so far. As a consequence of extrusion, no significant difference was found in the case of oat, at
both the extrusion conditions. For barley, a significant but small decrease took place after
extrusion, but no significant difference as found between the two extrusion temperatures. While
the data collected from the thermal treatments can be fully explained and matches previous
studies (Kivelä et al., 2011, 2012), data from extrusion is more uncertain. The comparison of
Mp and Mw confirms that the molecular size of β-glucan does not dramatically decrease with
extrusion, given the uncertainties of Mw measurement. For oat, this correlates well with the
viscosity, which did not change after extrusion at 120°C and only decreased very slightly at
150°C. In the case of barley, on the other end, the reducing ends are increasing slightly, and
therefore the Mp decreasing, while the viscosity dramatically increased, in particular in the case
of the samples extruded at 120°C, which might indicate a higher solubility of β-glucan. It can,
however, also be hypothesized that condensation reactions might take place, due to the high
pressure and low moisture in the extruder, since the decrease in Mw is too small to justify the
dramatic increase in viscosity. In any case, it must be kept in mind that the flours and the
extracts used for the experiments contain a myriad of other compound that can interact with
β-glucan and can play a part in this.
OBG vs BBG
Several studies in literature have reported different degradation rates of oat vs barley, which
mainly differ in their ratio of β-(1→3)-linked cellotriosyl to cellotetraosyl units (DP3:DP4),
with oat β-glucan having more β-(1→4)-linkages than barley (Faure et al., 2012, 2015). Boulos
and Nyström (2016), however, explored the degradation kinetics of the Fenton-induced
oxidation of three constitutionally isomeric β-glucan tetrasaccharides as model compounds with
no or one β-(1→3)-linkage. No significant differences in degradation kinetics could be found,
suggesting that for the commercial β-glucan used in the studies aforementioned, other
constituents than the pure carbohydrates were responsible for the difference in behavior
between oat and barely β-glucan (Boulos and Nyström, 2016). This lack of difference between
linkage type can be attributed to the aggressive nature of the hydroxyl radical and to its lack of
selectivity. Hence, this difference is probably due to the different amounts of iron, phytic acid

156
and proteins present in the extracts. It has been shown, in fact, that iron can form complexes
with chelating agents such as phytic acid but also with proteins, which enhances the reducing
power of iron (Welch et al., 2002). In the present study, the native oat extract contained a higher
amount of iron but also of phytic acid and proteins, compared to barley (Table 1). Both cereal
β-glucans had a similar trend for the thermal degradation, with oat having a slightly stronger
degradation than barley, as shown by the UPLC-MS (reducing end groups had a 2 fold and a
1.5 fold increase at 125°C and at 175°C and a 23 fold and 18 fold increase for oat and barley
respectively), viscosity and Mw data. This might be due to the higher amount of iron in oat than
barley, which catalyzes the Fenton reaction, and the simultaneous higher contents of phytic acid
and protein which, through complexation, can stabilize this metal. As shown by Welch et al.
(2002), in fact, in neutral water solutions, the redox potential of Fe3+/Fe2+ (110mV) is not
actually sufficient to reduce the dissolved oxygen with redox potential of 150mV. However,
they suggested that iron may complex with chelates or other compounds such as proteins and
act as a strong oxidants via Fenton chemistry.
As for the extrusion treatments, these will require further investigation. Also, the contents of
iron, phytic acid and proteins in the flours were unfortunately not determined, therefore their
influence on the process is unknown. However, in particular for the barley samples, Glc(3-6)
increased significantly following extrusion, indicating and incomplete enzymatic hydrolysis
during sample preparation or even some other type of reaction taking place during processing
which needs to be addressed further.
8. Conclusions
The influence of thermal (125°C and 175°C) and extrusion treatments on the molecular
structure of cereal β-glucan were investigated. A strong reduction of molecular weight and
viscosity took place during the thermal treatments, in particular at the higher temperature,
indicating a strong degradation taking place. For the extruded samples, oat β-glucan did not
suffer significant changes in terms of viscosity and molecular weight, while viscosity had a
dramatic increase in the case of barley, in particular for the samples extruded at 120°C. In order
to investigate the degradation and the oxidation products formed, a combination of carbonyl
labelling by reductive amination, enzymatic digestion with lichenase and β-glucosidase,
purification/fractionation by SPE and analysis by UPLC-MS with fluorescent detection was
used. A significant increase in the reducing end groups concentration was found for the
thermally treated samples for both oat and barley, in particular for those incubated at 175°, in

157
line with the reduction of molecular weight and viscosity. Oat had a slightly higher degradation
rate compared to barley, which can be due to its higher content in phytic acid, iron and proteins,
which can stabilize iron in its calalytic role in the Fenton reaction. Several lytic and non-lytic
oxidation products could be detected in particular in the case of the samples treated at 175°C,
even though quantification through FLD detection was possible for all of them. However, these
products still represent a good marker of β-glucan oxidation, confirming this method sensitive
enough to investigate β-glucan oxidation even in complex food systems. Further adjustments
will be required to achieve higher sensitivity and additional investigation will be needed for the
extruded samples, since no conclusive results could be reached with this study.
9. References
(FDA) F.a.D.A. 21 CFR 101.81 (2009) ‘Health claims: Soluble fiber from certain foods and risk of coronary heart disease (CHD)’, in Regulatory Information, pp. 143–148.
Åman, P., Rimsten, L. and Andersson, R. (2004) ‘Molecular Weight Distribution of β-Glucan in Oat-Based Foods’, Cereal Chemistry. doi: 10.1094/CCHEM.2004.81.3.356.
Beck, E. J. et al. (2009) ‘Oat β-glucan increases postprandial cholecystokinin levels, decreases insulin response and extends subjective satiety in overweight subjects’, Molecular Nutrition & Food Research, 53(10), pp. 1343–1351. doi: 10.1002/mnfr.200800343.
Bhatty, R. . (1995) ‘Laboratory and pilot plant extraction and purification of β-glucans from hull-less barley and oat brans’, Journal of Cereal Science, 22(2), pp. 163–170. doi: 10.1016/0733-5210(95)90046-2.
Boulos, S. and Nyström, L. (2016) ‘UPLC-MS/MS investigation of β-glucan oligosaccharide oxidation’, Analyst. Royal Society of Chemistry, 141(24), pp. 6533–6548. doi: 10.1039/c6an01125j.
Boulos, S. and Nyström, L. (2017) ‘Complementary Sample Preparation Strategies for Analysis of Cereal β-Glucan Oxidation Products by UPLC-MS/MS’, Frontiers in Chemistry, 5, pp. 157–208. doi: 10.3389/fchem.2017.00090.
Brennan, C. S. and Cleary, L. J. (2005) ‘The potential use of cereal (1→3,1→4)-β-d-glucans as functional food ingredients’, Journal of Cereal Science, 42(1), pp. 1–13. doi: 10.1016/j.jcs.2005.01.002.
Cui, S. W. and Wang, Q. (2009) ‘Cell wall polysaccharides in cereals: chemical structures and functional properties’, Structural Chemistry, 20(2), pp. 291–297. doi: 10.1007/s11224-009-9441-0.
Cui, W. and Wood, P. J. (2000) ‘Relationships between structural features, molecular weight and rheological properties of cereal β-D-glucans’, in Hydrocolloids. Elsevier, pp. 159–168. doi: 10.1016/B978-044450178-3/50019-6.
EFSA Panel on Dietetic Products Nutrition and Allergies (NDA) (2010) ‘Scientific Opinion on the substantiation of health claims related to beta-glucans and maintenance or achievement of normal blood glucose concentrations ( ID 756 , 802 , 2935 ) pursuant to Article 13 ( 1 ) of’, EFSA Journal, 8(1924), pp. 1–14. doi: 10.2903/j.efsa.2010.1482.
Faure, A. M., Andersen, M. L. and Nyström, L. (2012) ‘Ascorbic acid induced degradation of beta-glucan: Hydroxyl radicals as intermediates studied by spin trapping and electron spin resonance spectroscopy’, Carbohydrate Polymers. Elsevier Ltd., 87(3), pp. 2160–2168. doi:

158
10.1016/j.carbpol.2011.10.045.
Faure, A. M., Koppenol, W. H. and Nyström, L. (2015) ‘Iron(II) binding by cereal beta-glucan.’, Carbohydrate polymers, 115, pp. 739–43. doi: 10.1016/j.carbpol.2014.07.038.
Faure, A. M., Werder, J. and Nyström, L. (2013) ‘Reactive oxygen species responsible for beta-glucan degradation’, Food Chemistry. Elsevier Ltd, 141(1), pp. 589–596. doi: 10.1016/j.foodchem.2013.02.096.
Fenton, H. J. H. (1894) ‘LXXIII.—Oxidation of tartaric acid in presence of iron’, J. Chem. Soc., Trans., 65, pp. 899–910. doi: 10.1039/CT8946500899.
Fry, S. C. (1998) ‘Oxidative scission of plant cell wall polysaccharides by ascorbate-induced hydroxyl radicals’, Biochemical Journal, 332(2), pp. 507–515. doi: 10.1042/bj3320507.
Grundy, M. M. L. et al. (2017) ‘Impact of hydrothermal and mechanical processing on dissolution kinetics and rheology of oat β-glucan’, Carbohydrate Polymers. doi: 10.1016/j.carbpol.2017.02.077.
Haros, M., Rosell, C. M. and Benedito, C. (2001) ‘Fungal phytase as a potential breadmaking additive’, European Food Research and Technology, 213(4–5), pp. 317–322. doi: 10.1007/s002170100396.
Johansson, D. P. et al. (2018) ‘Larger particle size of oat bran inhibits degradation and lowers extractability of β-glucan in sourdough bread – Potential implications for cholesterol-lowering properties in vivo’, Food Hydrocolloids. doi: 10.1016/j.foodhyd.2017.09.016.
Johansson, L. et al. (2004) ‘Structural analysis of water-soluble and -insoluble β-glucans of whole-grain oats and barley’, Carbohydrate Polymers, 58(3), pp. 267–274. doi: 10.1016/j.carbpol.2004.06.041.
Johansson, L. et al. (2006) ‘Hydrolysis of β-glucan’, Food Chemistry, 97(1), pp. 71–79. doi: 10.1016/j.foodchem.2005.03.031.
Kahlon, T. S. and Woodruff, C. L. (2003) ‘In vitro binding of bile acids by rice bran, oat bran, barley and β-glucan enriched barley’, Cereal Chemistry. doi: 10.1094/CCHEM.2003.80.3.260.
Kivelä, R. et al. (2010) ‘Influence of homogenisation on the solution properties of oat β-glucan’, Food Hydrocolloids, 24(6–7), pp. 611–618. doi: 10.1016/j.foodhyd.2010.02.008.
Kivelä, R. et al. (2011) ‘Oxidative and radical mediated cleavage of β-glucan in thermal treatments’, Carbohydrate Polymers, 85(3), pp. 645–652. doi: 10.1016/j.carbpol.2011.03.026.
Kivelä, R. et al. (2012) ‘Oxidation of oat β-glucan in aqueous solutions during processing’, Carbohydrate Polymers, 87(1), pp. 589–597. doi: 10.1016/j.carbpol.2011.08.028.
Kivelä, R., Gates, F. and Sontag-Strohm, T. (2009) ‘Degradation of cereal beta-glucan by ascorbic acid induced oxygen radicals’, Journal of Cereal Science, 49(1), pp. 1–3. doi: 10.1016/j.jcs.2008.09.003.
Lamari, F. N., Kuhn, R. and Karamanos, N. K. (2003) ‘Derivatization of carbohydrates for chromatographic, electrophoretic and mass spectrometric structure analysis’, Journal of Chromatography B, 793(1), pp. 15–36. doi: 10.1016/S1570-0232(03)00362-3.
Lane, C. F. (1975) ‘Sodium Cyanoborohydride - A Highly Selective Reducing Agent for Organic Functional Groups’, Synthesis, 1975(03), pp. 135–146. doi: 10.1055/s-1975-23685.
Lazaridou, A. et al. (2004) ‘A comparative study on structure-function relations of mixed-linkage (1→3), (1→4) linear β-D-glucans’, Food Hydrocolloids, 18(5), pp. 837–855. doi: 10.1016/j.foodhyd.2004.01.002.
Lazaridou, A. and Biliaderis, C. G. (2007) ‘Molecular aspects of cereal β-glucan functionality: Physical properties, technological applications and physiological effects’, Journal of Cereal Science, 46(2), pp. 101–118. doi: 10.1016/j.jcs.2007.05.003.
Lazaridou, A., Biliaderis, C. G. and Izydorczyk, M. S. (2007) ‘Cereal β -Glucans: Structures, Physical Properties, and Physiological Functions’, in Functional food carbohydrates, pp. 1–72.

159
Li, M. and Lee, T. C. (1996) ‘Effect of extrusion temperature on solubility and molecular weight distribution of wheat flour proteins’, Journal of Agricultural and Food Chemistry. doi: 10.1021/jf950582h.
McKie, V. A. and McCleary, B. V. (2016) ‘A novel and rapid colorimetric method for measuring total phosphorus and phytic acid in foods and animal feeds’, Journal of AOAC International, 99(3), pp. 738–743. doi: 10.5740/jaoacint.16-0029.
Mossé, J. (1990) ‘Nitrogen to Protein Conversion Factor for Ten Cereals and Six Legumes or Oilseeds. A Reappraisal of Its Definition and Determination. Variation According to Species and to Seed Protein Content Jacques’, Journal of Agricultural and Food Chemistry, (38), pp. 18–24.
Ragaee, S., Abdel-Aal, E. S. M. and Noaman, M. (2006) ‘Antioxidant activity and nutrient composition of selected cereals for food use’, Food Chemistry. doi: 10.1016/j.foodchem.2005.04.039.
Rieder, A. et al. (2015) ‘Inter-laboratory evaluation of SEC-post-column calcofluor for determination of the weight-average molar mass of cereal β-glucan’, Carbohydrate Polymers. Elsevier Ltd., 124, pp. 254–264. doi: 10.1016/j.carbpol.2015.02.019.
Ruhaak, L. R. et al. (2010) ‘2-Picoline-borane: A non-toxic reducing agent for oligosaccharide labeling by reductive amination’, Proteomics. doi: 10.1002/pmic.200900804.
Schlemmer, U. et al. (2009) ‘Phytate in foods and significance for humans: Food sources, intake, processing, bioavailability, protective role and analysis’, Molecular Nutrition and Food Research, 53(SUPPL. 2), pp. 330–375. doi: 10.1002/mnfr.200900099.
Slavin, J. L. et al. (1999) ‘Plausible mechanisms for the protectiveness of whole grains’, The American Journal of Clinical Nutrition, 70(3), pp. 459s-463s. doi: 10.1093/ajcn/70.3.459s.
Tosh, S. M. et al. (2004) ‘Structural characteristics and rheological properties of partially hydrolyzed oat β-glucan: The effects of molecular weight and hydrolysis method’, Carbohydrate Polymers. doi: 10.1016/j.carbpol.2003.11.004.
Tosh, S. M. et al. (2010) ‘Processing Affects the Physicochemical Properties of β-Glucan in Oat Bran Cereal’, Journal of Agricultural and Food Chemistry, 58(13), pp. 7723–7730. doi: 10.1021/jf904553u.
Welch, K. D., Davis, T. Z. and Aust, S. D. (2002) ‘Iron Autoxidation and Free Radical Generation: Effects of Buffers, Ligands, and Chelators’, Archives of Biochemistry and Biophysics, 397(2), pp. 360–369. doi: 10.1006/abbi.2001.2694.
Wood, P. J. (2007) ‘Cereal β-glucans in diet and health’, Journal of Cereal Science, 46(3), pp. 230–238. doi: 10.1016/j.jcs.2007.06.012.
Wood, P. J. (2010) ‘REVIEW: Oat and rye -glucan: Properties and function.’, Cereal Chem., 87(4), pp. 315–330.
Zhang, M. et al. (2009) ‘Effect of process on physicochemical properties of oat bran soluble dietary fiber’, Journal of Food Science. doi: 10.1111/j.1750-3841.2009.01324.x.
Zhang, M., Bai, X. and Zhang, Z. (2011) ‘Extrusion process improves the functionality of soluble dietary fiber in oat bran’, Journal of Cereal Science. doi: 10.1016/j.jcs.2011.04.001.

160

161
APPENDIX
Figure S1. MS chromatograms for the Fraction 1 of OBG samples, native and treated, with their respective FLD signals. Starting from above: native OBG, treated at 125°C, treated at 150°C, extruded at 120°C, extruded at 150°C.
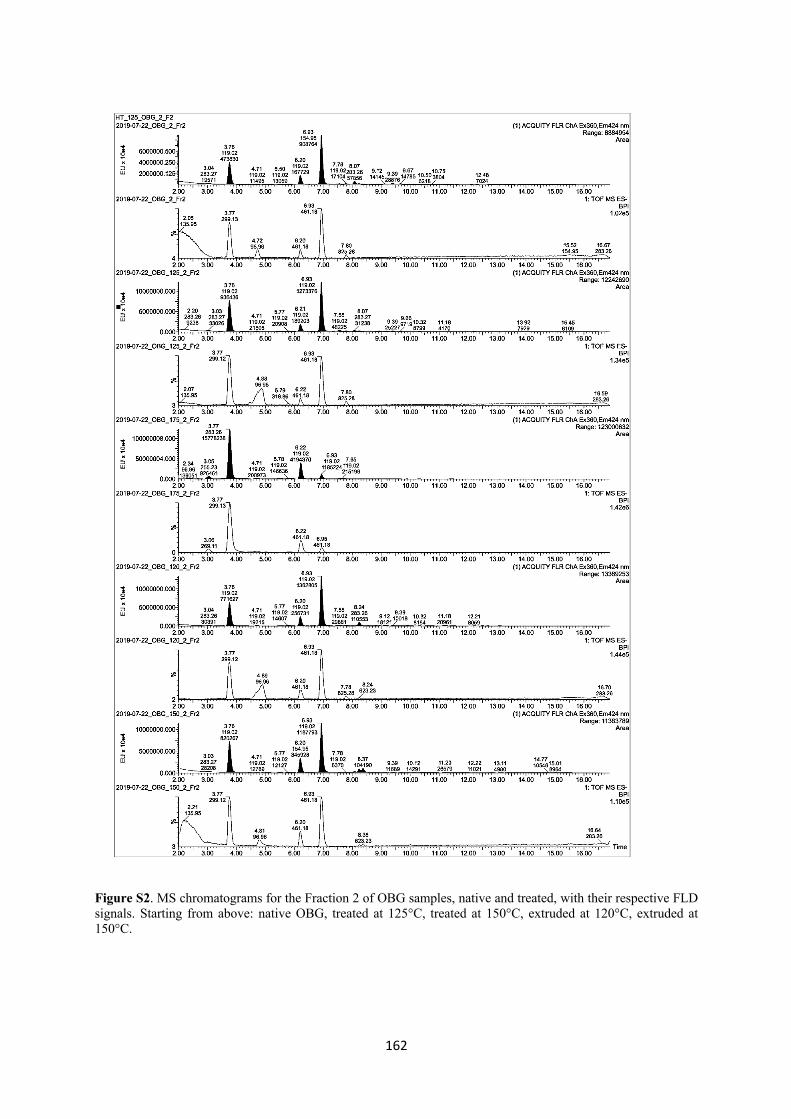
162
Figure S2. MS chromatograms for the Fraction 2 of OBG samples, native and treated, with their respective FLD signals. Starting from above: native OBG, treated at 125°C, treated at 150°C, extruded at 120°C, extruded at 150°C.

163
Figure S3. MS chromatograms for the Fraction 1 of BBG samples, native and treated, with their respective FLD signals. Starting from above: native OBG, treated at 125°C, treated at 150°C, extruded at 120°C, extruded at 150°C.
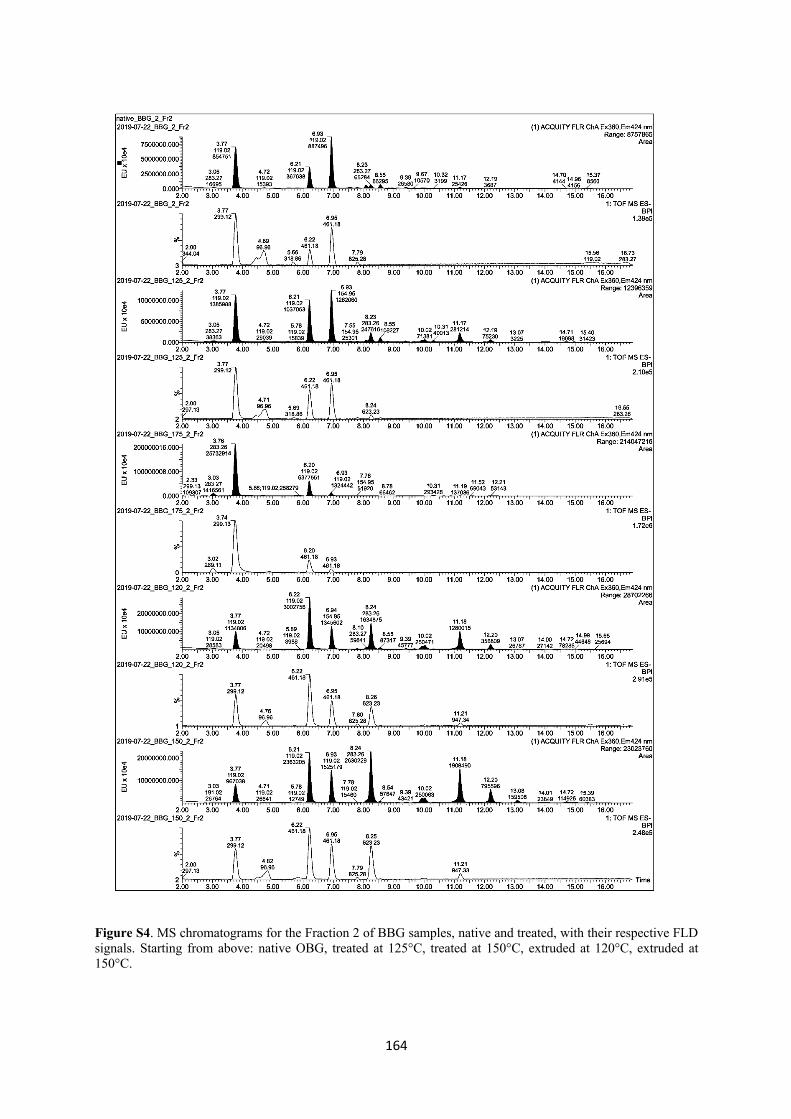
164
Figure S4. MS chromatograms for the Fraction 2 of BBG samples, native and treated, with their respective FLD signals. Starting from above: native OBG, treated at 125°C, treated at 150°C, extruded at 120°C, extruded at 150°C.

165
PART 3: CONCLUSIONS & OUTLOOK
The present work investigated the structural modifications of cereal β-glucan extracted from
oat and barley flours and the impact of these alterations on some of the fiber’s health aspects.
In particular, it was shown that, in contrast of what was reported previously in the literature,
oxidation or reduction of molecular weight do not improve in any way the bile acid retention
capacity of cereal β-glucan (Park et al., 2009; Kim et al., 2010; de Moura et al., 2011). This
property, in fact, only depends on the ability of β-glucan to form highly viscous solutions.
However, in the case of TEMPO oxidized and hydrolysed samples, despite the lower Mw, the
viscosity of the solutions was still sufficient to exert the bile acid “binding” effect, showing that
a certain degree of processing does not impair completely the health property of β-glucan, in
particular regarding the attenuation of serum cholesterol, which is in line with other reports
(Yokoyama et al., 2002). Also, given the good correlation with viscosity, other kinds of binding
were excluded, in particular a direct interaction between β-glucan and bile acids. However,
additional factors contributing to the overall cholesterol lowering effect cannot be ruled out and
further studies are required to clarify this.
Also, a direct binding of iron by β-glucan was excluded, in contrast with some reports (Faure
et al., 2015), confirming instead other studies that deem phytic acid responsible for iron binding
(Wang et al., 2017 a&b) and highlighting the necessity of keeping a close eye on this
contaminant when in need to ensure maximum iron availability. Additional in vivo and in vitro
experiments studying the interactions between β-glucan, phytic acid and iron would be required
to further investigate these mechanisms in order to ensure optimal iron bioavailability in foods.
The investigation of the oxidative degradation of cereal β-glucan induced by hydroxyl radical
generated through Fenton reaction, after high temperature treatments, gave quite clear and
consistent results, with a reduction of viscosity and Mw more intense the higher the temperature,
in line with the literature (Kivelä et al., 2012; Kivelä et al., 2011). The simultaneous increase
of the reducing end units and of the cross-ring cleavage oxidation products (such as arabinose
and erythrose) further confirmed this and shed some light on the modification of the β-glucan
structure during thermal degradation. However, unclear results were obtained in the case of
extrusion, where Mw and viscosity appeared to have different trends, in particular in the case of
barley. This, associated with unexpected products in the MS spectra, possibly resulting from an
incomplete enzymatic degradation, highlights the necessity to investigate this process further,

166
in particular the potential formation of condensation products due to the high pressure and low
moisture content during the extrusion cooking.
Overall, this thesis helped to add a few more pieces to the complicated jigsaw that is the
investigation of the structural properties of cereal β-glucan and how these are affected by
different types of processing and consequent structural changes. As Einstein said, “if we knew
what it was we were doing, it would not be called research, would it?”. Therefore, more studies
would be needed to understand further all these complex mechanism, in vitro and in vivo, to be
able to completely unravel the links between β-glucan structure and its biological value, to
ensure its beneficial health effects are maximized.
References
Faure, A. M., Koppenol, W. H. and Nyström, L. (2015) ‘Iron(II) binding by cereal beta-glucan.’, Carbohydrate polymers, 115, pp. 739–43. doi: 10.1016/j.carbpol.2014.07.038.
Kim, H. J. and White, P. J. (2010) ‘In Vitro Bile-Acid Binding and Fermentation of High, Medium, and Low Molecular Weight β-Glucan’, Journal of Agricultural and Food Chemistry, 58(1), pp. 628–634. doi: 10.1021/jf902508t.
Kivelä, R., Henniges, U., Sontag-Strohm, T. and Potthast, A. (2012) ‘Oxidation of oat β-glucan in aqueous solutions during processing’, Carbohydrate Polymers, 87(1), pp. 589–597. doi: 10.1016/j.carbpol.2011.08.028.
Kivelä, R., Sontag-Strohm, T., Loponen, J., Tuomainen, P. and Nyström, L. (2011) ‘Oxidative and radical mediated cleavage of β-glucan in thermal treatments’, Carbohydrate Polymers, 85(3), pp. 645–652. doi: 10.1016/j.carbpol.2011.03.026.
de Moura, F. A., Pereira, J. M., da Silva, D. O., Zavareze, E. da R., da Silveira Moreira, A., Helbig, E. and Dias, A. R. G. (2011) ‘Effects of oxidative treatment on the physicochemical, rheological and functional properties of oat β-glucan’, Food Chemistry, 128(4), pp. 982–987. doi: 10.1016/j.foodchem.2011.04.003.
Park, S. Y., Bae, I. Y., Lee, S. and Lee, H. G. (2009) ‘Physicochemical and Hypocholesterolemic Characterization of Oxidized Oat β-Glucan’, Journal of Agricultural and Food Chemistry, 57(2), pp. 439–443. doi: 10.1021/jf802811b.
Wang, Y. J., Maina, N. H., Ekholm, P., Lampi, A. M. and Sontag-Strohm, T. (2017) ‘Retardation of oxidation by residual phytate in purified cereal β-glucans’, Food Hydrocolloids, 66, pp. 161–167. doi: 10.1016/j.foodhyd.2016.11.019.
Wang, Y. J., Zhan, R., Sontag-Strohm, T. and Maina, N. H. (2017) ‘The protective role of phytate in the oxidative degradation of cereal beta-glucans’, Carbohydrate Polymers. Elsevier Ltd., 169, pp. 220–226. doi: 10.1016/j.carbpol.2017.04.016.
Yokoyama, W. H., Knuckles, B. E., Wood, D. and Inglett, G. E. (2002) ‘Food Processing Reduces Size of Soluble Cereal β-Glucan Polymers without Loss of Cholesterol-Reducing Properties’, in, pp. 105–116. doi: 10.1021/bk-2002-0816.ch008.





























